
Coincident pre-and postsynaptic activity modifies GABAergic synapses by postsynaptic changes in...
14
Neuron, Vol. 39, 807–820, August 28, 2003, Copyright 2003 by Cell Press Coincident Pre- and Postsynaptic Activity Modifies GABAergic Synapses by Postsynaptic Changes in Cl Transporter Activity At many excitatory synapses, coincident pre- and postsynaptic spiking of different temporal patterns has been shown to result in either LTP or LTD (Bi and Poo, 1998, 2001; Debanne et al., 1998; Feldman, 2000; Froemke and Dan, 2002; Magee and Johnston, 1997; Melanie A. Woodin, 1 Karunesh Ganguly, 1,2 and Mu-ming Poo* Division of Neurobiology Department of Molecular and Cell Biology University of California Berkeley, California 94720 Markram et al., 1997; Zhang et al., 1998); excitatory inputs which are repetitively activated before postsyn- aptic spiking become strengthened, while those acti- vated after postsynaptic spiking become weakened. Summary This allows for associative modification of coincident excitatory inputs, as suggested by Hebb’s postulate of Coincident pre- and postsynaptic activation is known learning (Hebb, 1949). Furthermore, this precise timing to induce long-term modification of glutamatergic syn- requirement reflects a dynamic interplay of membrane apses. We report here that, in both hippocampal cul- depolarization and Ca 2 influx through the NMDA recep- tures and acute hippocampal slices, repetitive post- tor as well as voltage-gated Ca 2 channels (Bi and Poo, synaptic spiking within 20 ms before and after the 1998; Koester and Sakmann, 1998) and argues for the activation of GABAergic synapses also led to a persis- incorporation of temporal specificity into the concept tent change in synaptic strength. This synaptic modifi- of coincidence detection at excitatory synapses (Abbott, cation required Ca 2 influx through postsynaptic 2000; Bi and Poo, 2001). Whether similar coincidence L-type Ca 2 channels and was due to a local decrease detection mechanisms also exist at inhibitory synapses in K -Cl cotransport activity, effectively reducing the is unclear. The precision with which GABAergic syn- strength of inhibition. Thus, GABAergic synapses can apses can detect coincidence will be of particular inter- detect and be modified by coincident pre- and post- est in light of findings that spike timing may be used to synaptic spiking, allowing the level of inhibition to be encode information in neural networks (Hopfield, 1995). modulated in accordance to the temporal pattern of In the present work, we have examined whether inhibi- postsynaptic excitation. tory synapses can be associatively modified through coincident pre- and postsynaptic spiking and further Introduction explored the cellular basis of coincidence detection at GABAergic synapses. Our findings suggest that coinci- Activity-induced modification of synaptic strength is be- dent pre- and postsynaptic spiking alters the activity of lieved to be a cellular mechanism underlying the refine- KCC2, a K -Cl cotransporter, resulting in changes in ment of developing connections (Katz and Shatz, 1996) the reversal potential of GABAergic synaptic currents as well as the learning and memory functions of the (E GABA ) and modulation of the strength of GABAergic inhi- nervous system (Bliss and Collingridge, 1993; Martin bition. This novel form of postsynaptic modulation of et al., 2000). Persistent activity-dependent changes in GABAergic synapses allows coordinated changes in ex- synaptic strength are often referred to as long-term po- citatory and inhibitory inputs to the postsynaptic neuron tentiation (LTP) or long-term depression (LTD), which are in response to neuronal activity. best characterized at hippocampal excitatory synapses (Bliss and Lomo, 1973; Nicoll and Malenka, 1995). In Results different brain regions and at different developmental stages, GABAergic synapses may also undergo bidirec- Coincident Pre- and Postsynaptic Activation tional long-term changes in synaptic strength following Modifies GABAergic Synapses repetitive presynaptic stimulation (Fitzsimonds et al., We first examined whether the timing of postsynaptic 1997; Komatsu and Iwakiri, 1993; Komatsu, 1996; Marty spiking relative to presynaptic stimulation affects the and Llano, 1995; McLean et al., 1996). In the cerebellum, efficacy of GABAergic synapses. Whole-cell perforated brief high-frequency stimulation of presynaptic fibers patch-clamp recordings with gramicidin (25 g/ml) were results in a rebound depolarization-induced spiking and made from pairs of interconnected neurons in low-den- LTP of inhibitory postsynaptic potentials (Kano et al., sity rat hippocampal cultures. Gramicidin forms pores 1992), whereas the same presynaptic activation with which are permeable to monovalent cations and small reduced postsynaptic spiking results in LTD (Aizenman uncharged molecules but not to Cl , permitting reliable et al., 1998). While these studies suggest that postsyn- recordings of GABAergic currents (Kyrozis and Reich- aptic activity can regulate the strength of inhibitory syn- ling, 1995; Owens et al., 1996). GABAergic postsynaptic apses, it is not known whether these synapses can de- currents (GPSCs) were identified by the time course, tect and be persistently modified by precisely coincident reversal potential (50 to 90 mV), and sensitivity to pre- and postsynaptic activity. blockade by bicuculline (10 M), a GABA A receptor an- tagonist. Current pulses were injected into the postsyn- *Correspondence: [email protected] aptic neuron (current clamped at 70 mV) to fire action 1 These authors contributed equally to this work. potentials in synchrony with repetitive presynaptic stim- 2 Present Address: School of Medicine, University of California at San Diego, La Jolla, California 92093. ulation (5 Hz, 30 s), with the peak of postsynaptic spiking
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Coincident pre-and postsynaptic activity modifies GABAergic synapses by postsynaptic changes in...

Neuron, Vol. 39, 807–820, August 28, 2003, Copyright !2003 by Cell Press
Coincident Pre- and Postsynaptic ActivityModifies GABAergic Synapses by PostsynapticChanges in Cl! Transporter Activity
At many excitatory synapses, coincident pre- andpostsynaptic spiking of different temporal patterns hasbeen shown to result in either LTP or LTD (Bi and Poo,1998, 2001; Debanne et al., 1998; Feldman, 2000;Froemke and Dan, 2002; Magee and Johnston, 1997;
Melanie A. Woodin,1 Karunesh Ganguly,1,2
and Mu-ming Poo*Division of NeurobiologyDepartment of Molecular and Cell BiologyUniversity of CaliforniaBerkeley, California 94720 Markram et al., 1997; Zhang et al., 1998); excitatory
inputs which are repetitively activated before postsyn-aptic spiking become strengthened, while those acti-vated after postsynaptic spiking become weakened.SummaryThis allows for associative modification of coincidentexcitatory inputs, as suggested by Hebb’s postulate ofCoincident pre- and postsynaptic activation is knownlearning (Hebb, 1949). Furthermore, this precise timingto induce long-term modification of glutamatergic syn-requirement reflects a dynamic interplay of membraneapses. We report here that, in both hippocampal cul-depolarization and Ca2" influx through the NMDA recep-tures and acute hippocampal slices, repetitive post-tor as well as voltage-gated Ca2" channels (Bi and Poo,synaptic spiking within 20 ms before and after the1998; Koester and Sakmann, 1998) and argues for theactivation of GABAergic synapses also led to a persis-incorporation of temporal specificity into the concepttent change in synaptic strength. This synaptic modifi-of coincidence detection at excitatory synapses (Abbott,cation required Ca2! influx through postsynaptic2000; Bi and Poo, 2001). Whether similar coincidenceL-type Ca2! channels and was due to a local decreasedetection mechanisms also exist at inhibitory synapsesin K!-Cl! cotransport activity, effectively reducing theis unclear. The precision with which GABAergic syn-strength of inhibition. Thus, GABAergic synapses canapses can detect coincidence will be of particular inter-detect and be modified by coincident pre- and post-est in light of findings that spike timing may be used tosynaptic spiking, allowing the level of inhibition to beencode information in neural networks (Hopfield, 1995).modulated in accordance to the temporal pattern of
In the present work, we have examined whether inhibi-postsynaptic excitation.tory synapses can be associatively modified throughcoincident pre- and postsynaptic spiking and furtherIntroductionexplored the cellular basis of coincidence detection atGABAergic synapses. Our findings suggest that coinci-Activity-induced modification of synaptic strength is be-dent pre- and postsynaptic spiking alters the activity oflieved to be a cellular mechanism underlying the refine-KCC2, a K"-Cl! cotransporter, resulting in changes inment of developing connections (Katz and Shatz, 1996)the reversal potential of GABAergic synaptic currentsas well as the learning and memory functions of the(EGABA) and modulation of the strength of GABAergic inhi-nervous system (Bliss and Collingridge, 1993; Martinbition. This novel form of postsynaptic modulation ofet al., 2000). Persistent activity-dependent changes inGABAergic synapses allows coordinated changes in ex-synaptic strength are often referred to as long-term po-citatory and inhibitory inputs to the postsynaptic neurontentiation (LTP) or long-term depression (LTD), which arein response to neuronal activity.best characterized at hippocampal excitatory synapses
(Bliss and Lomo, 1973; Nicoll and Malenka, 1995). InResultsdifferent brain regions and at different developmental
stages, GABAergic synapses may also undergo bidirec-Coincident Pre- and Postsynaptic Activationtional long-term changes in synaptic strength followingModifies GABAergic Synapsesrepetitive presynaptic stimulation (Fitzsimonds et al.,We first examined whether the timing of postsynaptic1997; Komatsu and Iwakiri, 1993; Komatsu, 1996; Martyspiking relative to presynaptic stimulation affects theand Llano, 1995; McLean et al., 1996). In the cerebellum,efficacy of GABAergic synapses. Whole-cell perforatedbrief high-frequency stimulation of presynaptic fiberspatch-clamp recordings with gramicidin (25 #g/ml) wereresults in a rebound depolarization-induced spiking andmade from pairs of interconnected neurons in low-den-LTP of inhibitory postsynaptic potentials (Kano et al.,sity rat hippocampal cultures. Gramicidin forms pores1992), whereas the same presynaptic activation withwhich are permeable to monovalent cations and smallreduced postsynaptic spiking results in LTD (Aizenmanuncharged molecules but not to Cl!, permitting reliableet al., 1998). While these studies suggest that postsyn-recordings of GABAergic currents (Kyrozis and Reich-aptic activity can regulate the strength of inhibitory syn-ling, 1995; Owens et al., 1996). GABAergic postsynapticapses, it is not known whether these synapses can de-currents (GPSCs) were identified by the time course,tect and be persistently modified by precisely coincidentreversal potential (!50 to !90 mV), and sensitivity topre- and postsynaptic activity.blockade by bicuculline (10 #M), a GABAA receptor an-tagonist. Current pulses were injected into the postsyn-
*Correspondence: [email protected] neuron (current clamped at !70 mV) to fire action1These authors contributed equally to this work.potentials in synchrony with repetitive presynaptic stim-2 Present Address: School of Medicine, University of California at
San Diego, La Jolla, California 92093. ulation (5 Hz, 30 s), with the peak of postsynaptic spiking

Neuron808
Figure 1. Modification of GABAergic Synapses by Coincident Pre- and Postsynaptic Activity
(A) Example recording of GPSC amplitudes before and after repetitive presynaptic stimulation paired with postsynaptic spiking (5 Hz, 30 s,marked by the arrow). Data points are GPSC amplitudes normalized to the mean value before repetitive stimulation (dotted line). Each pointis the average of all data obtained in a 1 min bin. The spike-timing interval between the onset of the GPSP and the peak of the postsynapticaction potential was "10 ms (see boxed inset for a sample trace during stimulation, scales: 100 mV, 10 ms). Traces below (left) depict samplesof GPSCs before (1) and after (2) the coincident stimulation, as well as overlapped traces of these samples after scaling (scales: 30 pA, 5 ms).(B) Summary of all experiments on the effect of coincident activity similar to (A), with the spike-timing interval within $20 ms. Data shownare the average GPSC amplitudes ($SEM), normalized for data from each experiment before averaging, in the absence (circles, amphotericin,n % 24; diamonds, gramicidin, n % 6) and the presence of D-APV (25 #m) and CNQX (15 #m) (squares, n % 13). Control experiments whereno coincident stimulation was given showed no significant change in the GPSC amplitude (triangles, p % 0.13, n % 8).(C) Summary of all experiments similar to those shown in (A), except that postsynaptic spikes were initiated more than 50 ms before or afterthe onset of GPSPs (circles, amphotericin, n % 19; diamonds, gramicidin, n % 6).(D) Summary of all experiments similar to (C), except that the postsynaptic neuron was not stimulated (circles, amphotericin, n % 11; diamonds,gramicidin, n % 5).
relative to presynaptic activation varied from 0 to $90 access resistance during long-term recordings withgramicidin, further experiments were performed withms (Figure 1A, inset). We found a persistent increase in
the amplitude of GPSCs (voltage clamped between !65 amphotericin B perforation (150–220 #g/ml). Since am-photericin B is partially permeable to Cl!, experimentsto !85 mV) when repetitive postsynaptic spiking oc-
curred within 20 ms before or after the onset of the began only when control recordings ensured a steady-state [Cl!]i was established. In the latter experiments,GABAergic postsynaptic potentials (GPSPs) (Figures 1A
and 1B). The changes in the GPSC amplitude induced by coincident pre- and postsynaptic spiking within $20 msalso produced a significant persistent increase in thecoincident activity averaged 24.2% $ 4.0%, a significant
difference compared to baseline control values (p & GPSC amplitude (26.4% $ 5.1%; p & 0.001, n % 24;Figure 1B). No significant differences were observed0.001, unpaired two-tailed Student’s t test; n % 6; Figure
1B). Experiments where no spike-timing induction pro- between results obtained using either amphotericin Bor gramicidin (p % 0.3, unpaired two-tailed Student’stocol was given showed no significant change in GPSC
amplitude over the same duration (p % 0.13, n % 8; t test). Furthermore, coincident pre- and postsynapticactivation also induced similar significant increases inFigure 1B). Due to the difficulty in obtaining stable

Coincident Spiking Modifies GABAergic Synapses809
aptic activation determines the polarity of synapticchanges (Bi and Poo, 1998; Debanne et al., 1998; Mageeand Johnston, 1997; Markram et al., 1997; Zhang et al.,1998). This suggests that different mechanisms underliethe coincidence detection at GABAergic synapses.
Changes in Postsynaptic Reversal Potentialfor GPSCsChanges in the amplitude of synaptic currents are usu-ally attributed to changes in the total synaptic conduc-tance, resulting from alterations in either transmitter re-lease or the postsynaptic responses to the transmitter.However, other mechanisms may also contribute. Forexample, changes in the intracellular Cl! concentration([Cl!]i) could alter the reversal potential for GPSCs(EGPSC), thus changing their amplitude (Ling and Benardo,1995; Thompson and Gahwiler, 1989a, 1989b). Using
Figure 2. Dependence of Synaptic Modification on Spike Timing gramicidin-perforated patch-clamp recordings, we havemeasured both the EGPSC and the slope conductancePercent change in the GPSC amplitude 10–20 min after repetitive
coincident activation (5 Hz, 30 s) is plotted against the interval be- of GPSCs at various times before and after repetitivetween the onset of GPSPs and the peak of the postsynaptic spike stimulation (Figure 3). We found that the voltage depen-(circles, amphotericin; squares, gramicidin), which varied from 0 to dence of GPSCs and the slope conductance did not$90 ms. Positive intervals: GPSP onset precedes spiking.
change during the control period. However, after coinci-dent pre- and postsynaptic activity, EGPSC shifted towardmore depolarizing values, consistent with the increaseGPSC amplitude in the presence of 6-cyano-7-nitroquin-
oxaline-2,3-dione (CNQX, 15 #M) or D-2-amino-5-phos- in the GPSC amplitude, with no obvious change in slopeconductance (Figures 3A1, 3A2, and 3C). This shift inphopentanoic acid (D-AP5, 25 #M; 25.2% $ 9.2%, p &
0.001; n % 13; Figure 1B), antagonists for the AMPA and EGPSC was statistically significant when compared to theshift in EGPSC for noncoincident, presynaptic-only, orNMDA subtype of glutamate receptors, respectively.
There was no significant difference in the amount GPSC postsynaptic-only protocols (p % 0.002, one-wayANOVA followed by Scheffe pairwise comparison).amplitudes increased between experiments performed
in the absence or presence of the glutamate antagonists Since GPSC amplitudes during the control period weremonitored with the postsynaptic neuron voltage(p % 0.3). Thus, glutamatergic transmission is not neces-
sary for the observed changes in GABAergic transmis- clamped below the EGPSC, the shift toward more positivelevels following coincident activity led to an increase insion, although glutamatergic transmission is normally
responsible for initiating the spiking of the postsynaptic the driving force for synaptic currents and, thus, anincrease in the GPSC amplitude.cell in vivo.
When repetitive postsynaptic spiking was initiated at In contrast to the above observations, we found nosignificant change in either EGPSC or the slope conduc-a time more than 50 ms before or after the onset of
GPSPs (termed “noncoincident spiking”), a small but tance when the interval between pre- and postsynapticspiking was outside the interval of $50 ms (p ' 0.05;significant reduction in the GPSC amplitude was found
(gramicidin: !13.2% $ 7.6%, p & 0.001, n % 6; ampho- Figures 3B1, 3B2, and 3C). Furthermore, repetitive spik-ing of either the pre- or postsynaptic neuron alone (attericin B: !8.1% $ 3.7%, p & 0.001, n % 19; no signifi-
cant difference between the two data sets, p % 0.2, 5 Hz, 30 s) had no significant effect on EGPSC (p ' 0.05;Figure 3C). Interestingly, repetitive presynaptic stimula-unpaired two-tailed Student’s t test; Figure 1C). In addi-
tion, repetitive stimulation of the presynaptic neuron tion alone resulted in a significant reduction of the slopeconductance when compared to coincident pre- andalone (5 Hz, 30 s) without inducing spiking of the post-
synaptic neuron (current clamped at !70 mV) resulted in postsynaptic activity (p % 0.01, Figure 3C), which couldaccount for the persistent reduction in the GPSC ampli-a persistent significant reduction of the GPSC amplitude
(gramicidin: !23.9 $ 15%, p & 0.001, n % 5; amphoteri- tude. In all experiments involving coincident activity,both the total conductance and the rise time of GPSCscin B: !22.1% $ 5.3%, p & 0.001, n % 11; no significant
difference, p % 0.6; Figure 1D). were unchanged. The 10%–90% rise time during thecontrol period (2.6 $ 0.9 ms; n % 9) and that found 10–20The dependence of synaptic modification on the rela-
tive timing of pre- and postsynaptic spiking was ana- min after stimulation were not significantly different(99.7% $ 5.1% of control values, n % 9), suggesting nolyzed by plotting the percentage change in the GPSC
amplitude against the time interval between the onset obvious alteration in the space clamp of the recording.Taken together, our results support the notion that thereof the GPSP and the peak of the postsynaptic action
potential (Figure 2). We found a clear elevation of the was a true shift in the EGPSC, triggered only by coincidentpre- and postsynaptic activity.GPSC amplitude when postsynaptic spiking occurred
within a $20 ms interval. This observation of a symmetri- To determine whether GABAergic synaptic transmis-sion, in addition to the coincident pre- and postsynapticcal window for modification of GABAergic synapses is
in sharp contrast to that observed at glutamatergic syn- activity, is required for the increase in GPSC amplitude,experiments were performed with bicuculline (10 #M)apses, where the temporal order of pre- and postsyn-

Neuron810
Figure 3. Mechanisms Underlying Changes in the GPSC Amplitude
(A1 and B1) Example recordings of changes in the GPSC amplitude induced by repetitive coincident ("5 ms, [A1]) and noncoincident ("75 ms,[B1]) activity, using gramicidin perforated patch. Traces below depict sample GPSCs before (1) and after (2) repetitive stimulation (scales: 100pA, 20 ms).(A2 and B2) GPSC amplitudes were measured at different clamping voltage and plotted against the holding voltage before (diamonds, solidline) and 10 min after (circles, dashed line) repetitive stimulation. The line represents the best linear fit; its abscissa intercept determines theEGPSC, and its slope is taken as the synaptic conductance.(C) Summary of changes in EGPSC and synaptic conductance under various stimulation conditions. Data represent average changes ($SEM)in EGPSC and slope conductance at 10–20 min after repetitive coincident activity relative to that observed during the control period. Changein EGPSC induced by coincident activity (spike-timing interval within $20 ms) is significantly different from that found for all other conditions(p % 0.002), including noncoincident activity (spike-timing interval beyond $50 ms). The change in slope conductance for presynaptic stimulationonly is significantly different from that found for coincident activity (p % 0.01). The number refers to the number of experiments.(D) Examples of recordings performed with picrotoxin (top panel; 50 #M; black bar) or bicuculline (bottom panel; 10 #M; black bar) presentduring the coincident stimulation protocol ("5 ms; arrow), using gramicidin perforated patch.

Coincident Spiking Modifies GABAergic Synapses811
or picrotoxin (50 #M), both GABAA antagonists, present induced increase in GPSC amplitudes also showed furo-semide sensitivity when tested with the drug at the endduring the coincident stimulation. No apparent increase
in the GPSC amplitude was induced by the stimulation of the experiment (n % 9; Figure 4A, bottom panel). Wenote that these experiments were performed in HCO3
!-protocol, as indicated by the measurements done afterthe antagonists were completely washed out (bicucul- free extracellular solution, thus GABAA receptor activa-
tion-dependent HCO3! efflux (Frech et al., 1999) did notline, n % 5; picrotoxin, n % 3; Figure 3D). Thus, GABAer-
gic synaptic transmission is required for the associative contribute to the increase in [Cl!]i observed.Furosemide may also inhibit the NKCC cotransportermodification of GPSCs.
To further examine potential presynaptic changes in (Cabantchik and Greger, 1992), which differs from KCC2by being insensitive to extracellular K" (Thompson andtransmitter release after coincident activation, we mea-
sured the paired-pulse ratio of GPSCs, which normally Gahwiler, 1989b). However, we found that in these hip-pocampal neurons EGPSC was sensitive to external K",reflects the release probability of the presynaptic neu-
ron. Paired-pulse ratios (PPRs) were obtained by com- and the sensitivity was blocked in the presence of furo-semide (Figure 4B), consistent with the specific effectparing the amplitude of GPSCs produced by stimulating
the presynaptic neuron with a pair of pulse stimuli. At of furosemide on KCC2. Figure 4C summarizes the ef-fects of furosemide treatment on activity-inducedan interval of 50 ms, the amplitude of the second GPSC
was found to be the same as the first GPSC, resulting changes in the GPSC amplitude after coincident andnoncoincident activity. The elevation of GPSC amplitudein an average PPR of 1.0 $ 0.06 (n % 4) during the control
period. At 10–20 min following coincident spiking, the induced by coincident activity was totally abolished bythe furosemide treatment. Taken together, these resultsPPR was 1.0 $ 0.08 (n % 4). The absence of any signifi-
cant change in PPR does not support presynaptic support the notion that changes in the GPSC amplitudeby coincident activity result from an activity-inducedchanges in transmitter release probability as an underly-
ing mechanism of synaptic modification by coincident inhibition of KCC2 and a shift of EGPSC toward more posi-tive levels.activity.
Somadendritic Differences in [Cl!]iThe Role of K!-Cl! Cotransporter ActivityTo further examine whether differential KCC2 activityAn obvious cellular mechanism mediating acute activity-in hippocampal neurons is capable of creating a localinduced changes in [Cl!]i is the modulation of variouschange in [Cl!]i, we compared the difference betweenCl! cotransporters (Jarolimek et al., 1999; Misgeld etsynaptic EGPSC and the reversal potential of GABA-al., 1986; Payne et al., 1996; Rivera et al., 1999; Rohr-induced currents (EGABA) measured at the soma. The syn-bough and Spitzer, 1996; Sun and Murali, 1999) andaptic EGPSC was determined by stimulating the presynap-channels (Smith et al., 1995) that are known to regulatetic neuron and recording the resulting current that[Cl!]i. Members of the K"/Cl! cotransport gene familypassed when the postsynaptic neuron was held at vary-expressed in hippocampal neurons include the Na"/K"/ing holding potentials (!90 to !50 mV, 5 mV incre-2Cl! (NKCC) cotransporter, which normally raises [Cl!]i
ments). The reversal potential of EGABA was determined(Sun and Murali, 1999), and KCC2, which normally low-by puffing pulses of GABA (20 ms) onto the soma, iners it (Payne et al., 1996; Rivera et al., 1999). To examinethe absence of presynaptic stimulation. The averagethe mechanism underlying the coincident activity-EGABA at the soma was !60.0 $ 2.5 mV (n % 9), which wasinduced shift of EGPSC toward more positive levels, wesignificantly more positive than that of EGPSC observed atbath applied furosemide (100 #M), a loop diuretic thatthe synapse in the same neurons (!65.3 $ 3.3 mV, p %inhibits cation chloride cotransport (Jarolimek et al.,0.0002, paired two-tailed Student’s t test). Moreover, the1999; Payne et al., 1996; Thompson and Gahwiler,difference between EGABA at the soma and EGPSC became1989b). We found that in most neurons examined (7/9)insignificant after repetitive coincident stimulationthe furosemide treatment resulted in an increase in(!61.9 $ 2.7 and !61.5 $ 3.1 mV, respectively, p %GPSC amplitude due to a shift in EGABA toward positive0.5). These results are consistent with the notion of alevels (from !66.2 $ 2.1 to !57.1 $ 4.4 mV; n % 7), andhigher KCC2 activity at the dendrite and the subsequentsubsequent coincident pre- and postsynaptic activityreduction of KCC2 activity induced by coincident activ-became ineffective in modifying the GPSC amplitudesity, which eliminated the somadendritic difference.(Figures 4A, top panel, and 4C).
Two types of hippocampal neurons have been distin-guished after 27–31 days in vitro under experimental The Role of Postsynaptic Ca2! Elevation
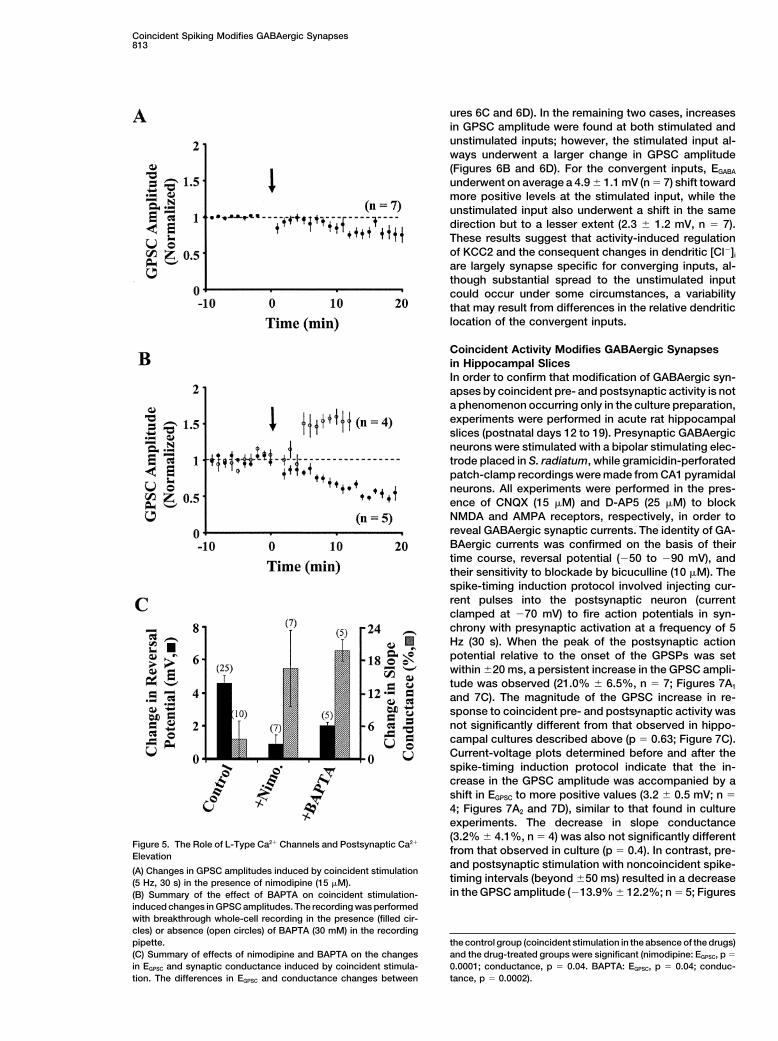
Since voltage-gated Ca2" channels are likely to be acti-[K"]o and [Cl!]i conditions: cells that show either similaror different dendritic and somatic GABA-induced cur- vated by postsynaptic spiking, we further examined the
effect of nimodipine, an inhibitor of high-threshold volt-rents, based on differences in the driving force of thesecurrents (Kelsch et al., 2001). In cells that showed differ- age-gated L-type Ca2" channels, and 1,2-bis(2-Amino-
phenoxy)ethane-N,N,N(,N(-tetraacetic acid (BAPTA), aences in dendritic versus somatic currents, furosemideabolished the differences, suggesting furosemide-sen- fast [Ca2"]i buffer, on the increase of the GPSC amplitude
resulting from coincident activation. As shown in Figuresitive KCC2 may generate a somatodendritic [Cl!]i gradi-ent. Consistent with this idea, we found that in furose- 5, in the presence of nimodipine (15 #M) or with BAPTA
(30 mM) in the postsynaptic whole-cell pipette, theremide-insensitive neurons, which were encounteredinfrequently in our cultures, coincident activity was inef- was a gradual reduction in the GPSC amplitude after
repetitive coincident stimulation within the intervalfective in altering GPSCs (n % 2; Figure 4A, middlepanel). Furthermore, neurons that exhibited an activity- of $20 ms (5 Hz, 30 s; Figures 5A and 5B). Furthermore,

Neuron812
Figure 4. The Effect of Furosemide
(A) Example recordings showing the effect of furosemide on GPSC amplitudes. (Top) Furosemide induced an increase in the GPSC amplitude,and subsequent coincident stimulation (at time % 0, arrow) failed to induce any further changes in the amplitude. Values were normalized tothe control period obtained after the addition of furosemide but before the coincident stimulation. (Middle) An example showing a GABAergicconnection unaffected by repetitive coincident stimulation that was also unresponsive to furosemide treatment. (Bottom) An example showingthe increase in the GPSC amplitude induced by repetitive coincident stimulation, followed by further increase due to furosemide.(B) Perfusion with saline containing 1 mM K" induced a shift of EGPSC to more negative values (upper trace), which was largely blocked bytreatment with furosemide (lower trace).(C) Summary of furosemide effects on changes in the GPSC amplitude observed following coincident and noncoincident pre- and postsynapticstimulation. The numbers refer to the number of experiments performed.
we observed that there was no significant change in postsynaptic activation? We address this question ofsynapse specificity using triple-perforated patch-clampEGPSC. The decrease in GPSC amplitude was accounted
for by the reduction in total synaptic conductance (Fig- recordings from neurons making divergent or converg-ing GABAergic connections. For the GABAergic presyn-ure 5C).
To exclude the possibility that the above result using aptic neuron making diverging connections with twopostsynaptic cells, we found that repetitive coincidentBAPTA was due to the increased Cl! exchange from
the pipette to the cytoplasm by the use of whole-cell stimulation of one pair of pre- and postsynaptic neuronsresulted in a significant elevation of the GPSC amplituderecordings, which are required for effective loading of
BAPTA, we examined the effects of coincident pre- and for the synaptic connection between them, without af-fecting the efficacy of the connection made on to thepostsynaptic activation on GPSCs, using whole-cell re-
cordings in the absence of BAPTA (Figure 5B). We found other postsynaptic neuron (Figure 6A; n % 5), whichwas voltage clamped at !70 mV during the period ofthat changes in the GPSC amplitude induced by coinci-
dent activity averaged 47.4% $ 19.1% (n % 4; Figure coincident stimulation. Repetitive coincident stimulationproduced a 4.2 $ 0.8 mV depolarizing shift in EGPSC at5B), which was not significantly different from those
observed using either gramicidin or amphotericin B (p % the stimulated input, while EGPSC at the unstimulated in-put also shifted in the depolarizing direction but to a0.6). Thus, postsynaptic spiking appears to activate a
nimodipine-sensitive Ca2" influx, leading to both a shift much lesser extent (0.9 $ 0.3 mV).Seven triplets with convergent GABAergic inputs ontoof EGPSC and a protection against a decrease in the Cl!
conductance due to repetitive synaptic activation. a single postsynaptic neuron were also examined. In5/7 cases, we found that the increase in GPSC amplitudeinduced by coincident pre- and postsynaptic activationSynapse Specificity in the Activity-Induced
Modification resulted in significant increase in the GPSC amplitudeat the stimulated input, while the input from the unstimu-Is the activity-induced shift in EGPSC restricted to acti-
vated synapses that are exposed to coincident pre- and lated neuron remained unchanged or depressed (Fig-
Coincident Spiking Modifies GABAergic Synapses813
ures 6C and 6D). In the remaining two cases, increasesin GPSC amplitude were found at both stimulated andunstimulated inputs; however, the stimulated input al-ways underwent a larger change in GPSC amplitude(Figures 6B and 6D). For the convergent inputs, EGABA
underwent on average a 4.9 $ 1.1 mV (n % 7) shift towardmore positive levels at the stimulated input, while theunstimulated input also underwent a shift in the samedirection but to a lesser extent (2.3 $ 1.2 mV, n % 7).These results suggest that activity-induced regulationof KCC2 and the consequent changes in dendritic [Cl!]i
are largely synapse specific for converging inputs, al-though substantial spread to the unstimulated inputcould occur under some circumstances, a variabilitythat may result from differences in the relative dendriticlocation of the convergent inputs.
Coincident Activity Modifies GABAergic Synapsesin Hippocampal SlicesIn order to confirm that modification of GABAergic syn-apses by coincident pre- and postsynaptic activity is nota phenomenon occurring only in the culture preparation,experiments were performed in acute rat hippocampalslices (postnatal days 12 to 19). Presynaptic GABAergicneurons were stimulated with a bipolar stimulating elec-trode placed in S. radiatum, while gramicidin-perforatedpatch-clamp recordings were made from CA1 pyramidalneurons. All experiments were performed in the pres-ence of CNQX (15 #M) and D-AP5 (25 #M) to blockNMDA and AMPA receptors, respectively, in order toreveal GABAergic synaptic currents. The identity of GA-BAergic currents was confirmed on the basis of theirtime course, reversal potential (!50 to !90 mV), andtheir sensitivity to blockade by bicuculline (10 #M). Thespike-timing induction protocol involved injecting cur-rent pulses into the postsynaptic neuron (currentclamped at !70 mV) to fire action potentials in syn-chrony with presynaptic activation at a frequency of 5Hz (30 s). When the peak of the postsynaptic actionpotential relative to the onset of the GPSPs was setwithin $20 ms, a persistent increase in the GPSC ampli-tude was observed (21.0% $ 6.5%, n % 7; Figures 7A1
and 7C). The magnitude of the GPSC increase in re-sponse to coincident pre- and postsynaptic activity wasnot significantly different from that observed in hippo-campal cultures described above (p % 0.63; Figure 7C).Current-voltage plots determined before and after thespike-timing induction protocol indicate that the in-crease in the GPSC amplitude was accompanied by ashift in EGPSC to more positive values (3.2 $ 0.5 mV; n %4; Figures 7A2 and 7D), similar to that found in cultureexperiments. The decrease in slope conductance(3.2% $ 4.1%, n % 4) was also not significantly different
Figure 5. The Role of L-Type Ca2" Channels and Postsynaptic Ca2"
from that observed in culture (p % 0.4). In contrast, pre-Elevationand postsynaptic stimulation with noncoincident spike-
(A) Changes in GPSC amplitudes induced by coincident stimulationtiming intervals (beyond $50 ms) resulted in a decrease(5 Hz, 30 s) in the presence of nimodipine (15 #M).in the GPSC amplitude (!13.9% $ 12.2%; n % 5; Figures(B) Summary of the effect of BAPTA on coincident stimulation-
induced changes in GPSC amplitudes. The recording was performedwith breakthrough whole-cell recording in the presence (filled cir-cles) or absence (open circles) of BAPTA (30 mM) in the recording
the control group (coincident stimulation in the absence of the drugs)pipette.(C) Summary of effects of nimodipine and BAPTA on the changes and the drug-treated groups were significant (nimodipine: EGPSC, p %
0.0001; conductance, p % 0.04. BAPTA: EGPSC, p % 0.04; conduc-in EGPSC and synaptic conductance induced by coincident stimula-tion. The differences in EGPSC and conductance changes between tance, p % 0.0002).

Neuron814
Figure 6. Synapse Specificity in the Coincident Stimulation-Induced Modification of GABAergic Synapses
(A) Example recording of a triplet with divergent connections made by a GABAergic neuron onto two postsynaptic neurons. Enhancement ofGPSC amplitude was observed at the connection exposed to coincident stimulation, but no change was observed at that made onto the(unstimulated, control) neuron.(B and C) Example recording of two triplets with convergent connections made by two GABAergic neurons onto one postsynaptic neuron.Increase in the GPSC amplitude induced by coincident stimulation resulted in either a similar increase (B) or a reduction (C) in the GPSCamplitude of the other (unstimulated, control) convergent input.(D) Summary of results on the spread of synaptic modification between divergent GABAergic outputs and converging GABAergic inputs. Datacollected from triplets similar to those shown in (A)–(C), and percentage changes in the average GPSC amplitude recorded from the sametriplet are connected by the line.
7B1 and 7C). The percentage decrease in the GPSC 32.0% $ 5.9% (n % 5) increase in GPSC amplitude,while noncoincident activity produced a slight decreaseamplitude observed in acute hippocampal slices was
not significantly different from the decrease produced (!12.8% $ 9.8%, n % 4). This was not significantlydifferent from that observed at room temperature (Figureby noncoincident activity in culture (p % 0.52; Figure
7C). Similar to the experiments performed in culture, the 7C). Similarly, we found at 35)C, there was a similar shiftin EGPSC (4.4 $ 0.5 mV, Figure 7D) following coincidentdecrease in GPSC amplitude resulting from noncoinci-
dent activity protocols did not result from a significant activity, as compared to that observed at room temper-ature.change in EGPSC (1.4 $ 0.2 mV; n % 3, p % 0.75; Figure
7D). The decrease in GPSC amplitude produced by non-coincident activity was accompanied by a 4.1% $ 3.4% Discussionchange in slope conductance, which was not signifi-cantly different from that observed in hippocampal cul- In the present study, we have demonstrated that coinci-
dent pre- and postsynaptic activity within a window oftures (p % 0.3).Neuronal excitability and the efficacy of synaptic $20 ms decreases cation chloride cotransporter activ-
ity, resulting in a shift in the EGPSC to more positive valuestransmission can be dependent upon the temperatureof the neuronal tissue (Schoepfle and Erlanger, 1941; and an increase in the GPSC amplitude when assayed
at a level below the reversal potential. Repetitive presyn-Takeya et al., 2002). To determine whether the abilityof coincident pre- and postsynaptic activity to modify aptic activity alone led to a reduction of the GPSC am-
plitude, which is mediated by a decrease in the totalGABAergic synapses was temperature dependent, ex-periments were also performed on acute hippocampal synaptic conductance. Coincident postsynaptic spiking
appears to prevent this decrease in synaptic conduc-slices heated to 35)C. At this elevated temperature, coin-cident pre- and postsynaptic activity produced a tance. Furthermore, the effects of coincident activity,

Coincident Spiking Modifies GABAergic Synapses815
Figure 7. Coincident Activity Modifies GABAergic Synapses in Acute Hippocampal Slices
(A1 and B1) Example recordings of changes in the GPSC amplitude induced by repetitive coincident ("5 ms, [A1]) and noncoincident ("75 ms,[B1]) activity, using gramicidin perforated patch. Traces below (left) depict sample GPSCs before (1) and after (2) repetitive stimulation, whichare also shown on the right after scaling. Scales: 20 pA, 20 ms.(A2 and B2) Examples of current-voltage relation of GPSCs measured during experiments shown in (A1) and (B1), before (solid lines, diamondsand squares) and 10 min after (dashed line, circles) the repetitive coincident (A2) and noncoincident (B2) stimulation.(C) Summary of percent change in the GPSC amplitude in both hippocampal cultures and slices. Data represent average changes in GPSCamplitude ($SEM) 10–20 min after the repetitive coincident and noncoincident stimulation, relative to that observed during the control period.There was no significant difference observed between slice and culture preparations for either coincident or noncoincident stimulation (p %0.6). There was also no significant difference between experiments performed on slices at room temperature or when heated to 35)C (coincident,p % 0.2; noncoincident, p % 0.9).(D) Summary of the change in EGPSC induced by coincident and noncoincident spike-timing intervals for recordings in both hippocampalcultures and slices. There was no significant difference observed between slice and culture preparations for either coincident or noncoincidentstimulation (p % 0.2), at room temperature, or at 35)C (coincident, p % 0.2; noncoincident, p % 0.6).
both the protection from the reduction in synaptic con- spiking. Thus, inhibitory inputs can be regulated bychanges in either synaptic conductance or reversal po-ductance and the positive shift in EGPSC, require postsyn-
aptic high-threshold nimodipine-sensitive Ca2" chan- tential, thereby allowing for a dynamic control of inhi-bition.nels, which are likely to be activated by postsynaptic

Neuron816
Coincidence Detection by GABAergic Synapses et al., 1996). Serine/threonine phosphorylation has beenThe existence of the $20 ms time window for the modifi- shown to regulate KCC2 activity in oocytes (Strange etcation of GABAergic synapses demonstrates that these al., 2000), while cytosolic tyrosine kinase activity regu-synapses are indeed capable of detecting and being lates K"/Cl! cotransport activity during the maturationmodified by coincident pre- and postsynaptic activity, of hippocampal neurons (Kelsch et al., 2001). Increaseda well-known property of most excitatory glutamatergic Ca2" levels associated with spike-induced L-type Ca2"
synapses. GABAergic inputs typically do not trigger channel activation, in conjunction with GABAA receptorpostsynaptic spiking, but other convergent excitatory activation, may activate intracellular signaling moleculesinputs do. The latter was simulated in the present study that downregulate KCC2 activity by dephosphorylation,by injecting depolarizing currents into the postsynaptic in a manner similar to the dephosphorylation of AMPAneuron. Thus, our findings suggest that GABAergic in- receptors following the induction of LTD (Lee et al.,puts can be modified via association with coincident 1998).excitatory inputs. Since excitatory inputs that cause re- The percent change in GPSC amplitude in responsepetitive postsynaptic spiking are likely to be potentiated to coincident pre- and postsynaptic differences, suchthemselves, the plasticity of excitatory inputs would oc- as the composition of GABAA receptor activity, variedcur in parallel with a depolarizing shift in EGPSC of all from !27.4% to 79.8% (Figure 2). While this variabilitytemporally coincident GABAergic inputs, resulting in an may result from subunits, the magnitude of the Ca2"
enhanced net excitation of the postsynaptic neuron. rise, and the level of KCC2 expression, it is also likelyThus, the associative modification of GABAergic inputs that the variability was due in part to different typesallows the level of inhibition to be modulated in accor- of presynaptic neurons. It is known that presynapticdance with the temporal pattern of postsynaptic activity, interneuron identity may influence the properties ofproviding a more active role for inhibitory inputs in the postsynaptic currents (Maccaferri et al., 2000). Becauseplasticity of neuronal networks. of the potential existence of a somadendritic gradient
The capacity for coincidence detection is believed to of [Cl!]i, dendritically projecting interneurons (Callawaybe important for learning and memory functions of the and Ross, 1995; Hoffman et al., 1997; Miles et al., 1996)nervous system. For glutamatergic synapses, the NMDA may be more likely to be susceptible to modification ofreceptor operates as a molecular coincidence detector, the GPSC amplitude via the mechanism reported here.since it allows high levels of Ca2" influx only when the Further studies that more thoroughly classify GABAergicpostsynaptic neuron is depolarized in the presence of neurons on the basis of their projections, neurochemicalpresynaptic release of glutamate (Bliss and Collingridge, content, and physiological and morphological proper-1993; Malenka and Nicoll, 1993). Furthermore, the coin- ties (Klausberger et al., 2003; McBain and Fisahn, 2001)cidence detection machinery at glutamatergic synapses are required to determine cell type dependence of activ-appears to respond differentially to the temporal order ity-induced modification.of pre- and postsynaptic activity (Bi and Poo, 1998,2001; Debanne et al., 1998; Markram et al., 1997; Zhang LTD of GABAergic Synapseset al., 1998). In contrast, the coincidence detector at The persistent reduction of synaptic conductance fol-GABAergic synapses does not appear to respond to the lowing repetitive presynaptic stimulation (without post-temporal order of pre- and postsynaptic activity. It does,
synaptic spiking) is similar to LTD of GABAergic re-however, require synaptic GABAA receptor activation insponses reported previously in these hippocampalthe presence of postsynaptic spike-induced Ca2" influx.cultures (Fitzsimonds et al., 1997) and hippocampalSpikes initiated at the axonal initial segment are knownslices (McLean et al., 1996). The decrease in inhibitoryto propagate backward to the dendritic tree and, thus,synaptic conductance may serve as a means to regulatemay activate high-threshold voltage-gated Ca2" chan-the gain of inhibition in accordance to the level andnels, such as L-type Ca2" channels at the dendritetemporal pattern of excitation, in this case, a lack of(Magee and Johnston, 1997; Stuart et al., 1997). Thesynchronous postsynaptic excitation. The mechanismrequirement of the spatial and temporal correlation ofunderlying this reduction of synaptic conductance—the spike-induced Ca2" influx with components of GA-whether it is due to a reduced transmitter release or aBAergic transmission, such as GABAA receptor activa-reduction of postsynaptic GABAA channel activity––istion or Cl! fluxes, is likely to underlie the apparent timepresently unknown. Our results suggest that when post-window for the modification of GABAergic synapses.synaptic spiking triggered by convergent excitatory in-Given the rapidity of the effect of coincident activity,puts is temporally correlated with presynaptic activa-alterations in the level of gene transcription or proteintion, there is an apparent Ca2"-dependent signal thatsynthesis are unlikely. Thus, changes in the level of ex-counteracts against the process underlying the reduc-pression or posttranslational modification of cationtion of synaptic conductance. Since the total synapticchloride cotransporters are likely to account for the de-conductance of inhibitory synapses is important forcrease in Cl! efflux. Recruitment and removal of synap-shunting inhibition, preventing the decrease in conduc-tic AMPA receptors have been reported following thetance also contributes to the reduction of inhibition forinduction of LTP (Hayashi et al., 2000) and LTD (Carroll etthose inhibitory inputs that are temporally correlatedal., 1999), respectively, at hippocampal CA1 excitatorywith the postsynaptic excitation. However, substantialsynapses. The decrease in Cl! transporter activity mayspontaneous in vivo spiking activity may influence thealso result from removal of functional KCC2 from thestability of synaptic modifications (Zhou et al., 2003), andplasma membrane. The KCC2 protein contains five con-bursting spiking activity is likely to impose additionalsensus phosphorylation sites for protein kinase C and
one tyrosine protein kinase phosphorylation site (Payne spiking history-dependent pre- and postsynaptic effects

Coincident Spiking Modifies GABAergic Synapses817
that alter the nature of synaptic modification (Froemke [Cl!]i by coincident activity. This conclusion is based onand Dan, 2002). Further studies in an in vivo preparation several lines of evidence. First, we showed that EGPSC
are required to verify the findings reported here. was sensitive to external K", a property specific toKCC2, and this sensitivity to K" was blocked by furose-
Postsynaptic Shifts in EGPSC as a Mechanism mide, consistent with a specific effect of furosemidefor GABAergic Synaptic Modification on the KCC2 activity. Second, our experiments wereOur data showed that coincident spiking led to GPSCs performed at a developmental stage when KCC2 ex-that were more depolarizing. In a few cases, initially pression is known to be upregulated, while that of NKCChyperpolarizing GPSCs were converted to depolarizing is downregulated (Ganguly et al., 2001; Kakazu et al.,GPSCs following coincident activity. Furthermore, since 1999; Lu et al., 1999; Plotkin et al., 1997), Third, KCC2EGPSC often resides near the resting membrane potential, is localized at inhibitory synapses in both mouse spinaleven small changes in EGPSC can produce marked effects cord (Hubner et al., 2001) and rat retina (Vu et al., 2000),on the amplitude of GPSCs. The change in the driving suggesting KCC2 is strategically positioned to maintainforce for GPSCs due to the shift in EGPSC after coincident local postsynaptic [Cl!]i. Finally, KCC2 is known to un-activity is sufficient to account for the change in the dergo activity-dependent regulation in cultured hippo-GPSC amplitude. For example, in the example illustrated campal neurons (Ganguly et al., 2001). Upregulation ofin Figure 7A, the 6.8 mV change in EGPSC would result in KCC2 expression during neonatal development resultsa 57% change in the amplitude of GPSCs, which is close in a hyperpolarizing shift of EGPSC, leading to the switchto that observed.
of GABAergic transmission from excitatory to inhibitoryOur finding that coincident activity induces a shift of(Ganguly et al., 2001; Rivera et al., 1999). Our resultsthe reversal potential of GABAergic responses suggestssuggest that coincident activity acts specifically to de-that there is a rapid activity-dependent and persistentcrease KCC2 activity leading to the establishment of amodulation of [Cl!]i at the synapse. Results from tripletlocalized change in [Cl!]i. Such bidirectional regulationrecordings on convergent GABAergic inputs (Figure 6)of GABA synapses via changes in the Cl! transportersuggest that such modulations of [Cl!]i can be locallyactivity supports the notion that developmental remod-restricted, only affecting synapses undergoing coinci-eling of synapses and activity-dependent synaptic plas-dent activation. The average measured change in theticity share common cellular mechanisms.reversal potential was 4.6 $ 0.8 mV (n % 11) at 10–20
min after coincident stimulation (Figure 3), which wouldpredict a change in the [Cl!]i by about 2 mM. Our ob-
Multiple Functions of GABAergic Inputsserved changes are consistent with the previous findingInhibitory inputs are usually located in regions of the(Collin et al., 1995) that following pairing of sustainedneuron where they can exert substantial influence onpostsynaptic depolarization with exogenous GABA ap-signal integration; small changes in their efficacy canplication to hippocampal CA1 pyramidal neurons re-result in large effects on neuronal output. In general, asulted in persistent modification of somatic responsesGABAergic input can be inhibitory through eitherto exogenous GABA. During the coincident pre- andshunting or hyperpolarization of the membrane (Staleypostsynaptic stimulation protocol, postsynaptic stimu-
lation may create a positive driving force for Cl! at the and Mody, 1992). Therefore, changes in either the syn-synapse, resulting in a local increase in [Cl!]i. A rough aptic conductance or EGPSC will affect the characteristicscalculation based on the average synaptic conductance of inhibition. With the shift in EGPSC toward positive levelsof !10 nS and a dendritic segment volume of 100 #m3, above the resting potential following correlated stimula-coincident postsynaptic activity of 150 spikes may in- tion, a depolarizing GABAergic input will exert a compli-duce an increase of [Cl!]i in the order of 1 mM. However, cated effect of both shunting inhibition and facilitatingsuch transient elevation of [Cl!]i can not account for excitation depending on the relative timing and spatialthe persistent changes in GPSC amplitude, unless the distribution of GABAergic and glutamatergic inputs. Foractivity of Cl! transporter is also persistently reduced. example, depolarizing GABAergic responses, while ca-Transient changes in [Cl!]i in the order of mM, e.g., that pable of shunting spatially and temporally coincidentresulted from the reduction of [K"]o, should be quickly excitatory inputs, may facilitate distant subthreshold ex-abolished after the termination of experimental manipu- citatory inputs that arrive on the decay phase of GPSCslations (see Figure 4B). We hypothesize that increased (Chen et al., 1996). In addition, depolarizing GABAergic[Ca2"]i, through its downstream effectors, may persis-
inputs may help to activate voltage-gated calcium chan-tently modify the activity of KCC2 cotransporters at thenels or even unblock NMDA receptors, thus influencingactivated GABAergic synapses. Furthermore, the resultsthe function and/or plasticity of other convergent syn-from triplet experiments suggest that coincident activityapses (Bazhenov et al., 1999). Finally, shifts of EGPSC ofmay induce local modification of Cl! transporter activityinhibitory inputs may modify the reliability of initiatingin these neurons. Local regulation of [Cl!]i has beenforward propagating action potentials (Debanne et al.,observed in retinal neurons, where a differential tar-1998) and subsequent transmitter release from the post-geting of Cl! transporter KCC2 can produce both depo-synaptic neuron following activation of inhibitory inputs.larizing and hyperpolarizing GABA responses in theThrough associative modification based on coincidencesame cell (Vardi et al., 2000).with postsynaptic spiking, these multiple functions ofGABAergic synapses can be coordinated in accordanceInvolvement of KCC2 Activitywith the activity of glutamatergic inputs to the sameOur results indicate that among the family of K"/Cl!
transporters, KCC2 is responsible for the increase of postsynaptic neuron.

Neuron818
Experimental Procedures ron was voltage clamped at !70 to !90 mV, resulting in inwardGPSCs. In a few cases when the reversal potential was around !80mV, the postsynaptic neuron was clamped at !60 mV, resulting inCell Culture
Low-density cultures of dissociated embryonic rat hippocampal outward GPSCs. Prior to the induction protocol, a minimum of 10min of control baseline recording was performed. The inductionneurons were prepared as previously described (Bi and Poo, 1998).
In brief, hippocampi were removed from E18–20 embryonic rats and protocol consisted of repetitive presynaptic stimulation (100 mVstep depolarization from Vc of !70 mV, duration 1 ms, 5 Hz for 30treated with trypsin for 20 min at 37)C, followed by gentle trituration.
The dissociated cells were plated at densities of 25,000 to 100,000 s) each paired with a postsynaptic spike induced by injection ofdepolarizing current pulses ("2 nA, 2 ms). Experiments were contin-cells/ml on poly-L-lysine coated glass coverslips in 35 mm petri
dishes. The plating medium was Dulbecco’s minimum essential me- ued only when a stable baseline GPSC amplitude (&10% variation)and reversal potential were observed before induction. The changedium (DMEM; BioWhittaker), supplemented with 10% fetal calf se-
rum (Hyclone), 10% Ham’s F12 with glutamine (BioWhittaker), and in GPSC amplitude resulting from the spike-timing induction proto-col was calculated by comparing the average size of the GPSC50 U/ml penicillin-streptomycin (Sigma). Twenty-four hours after
plating, the culture medium was changed to the above medium amplitude between 10 and 20 min after the induction protocol tothe average GPSC amplitude obtained in a 10 min baseline recordingcontaining 20 mM KCl. Both glia and neurons are present under
these culture conditions. Cells were recorded from 9 to 14 days in made prior to the induction protocol.The synaptic reversal potential of GPSCs was determined by vary-culture.
ing the holding potential of the postsynaptic cell in 5 mV increments(!90 to !30 mV) and measuring the resulting GPSC amplitude.Hippocampal SlicesTo determine the reversal potential, we used linear regression toHippocampal slices were prepared from postnatal day 12 to 19calculate a best-fit line for the voltage dependence of GPSCs. TheSprague-Dawley rats. Rats were briefly anesthetized with halothaneinterpolated intercept of this line with the abscissa was taken asand decapitated. Brains were removed and submerged in ice-coldthe reversal potential. The slope of the same line was then takensucrose-CSF: 206 mM sucrose, 2.8 mM KCl, 1 mM CaCl2, 1 mMas the respective slope conductance. Determination of EGABA at theMgCl2, 2 mM MgSO4, 1.25 mM NaH2PO4, 26 mM NaHCO3, 10 mMsoma was made in a manner similar to that described for the syn-D-glucose, 0.4 mM ascorbic acid, pH 7.4 when equilibrated withapse, except that GABA was puffed onto the cell body in 30 s95% O2-5% CO2, 295 mOsmol. Transverse hippocampal slices (400intervals (100 #M, 20 ms, 5 psi) through a micropipette (!1.5–2 M*)#M) were cut using a Vibratome 3000 sectioning system (St. Louis,with a Picospritzer II (Parker Instrumentation). In the present study,MO). Slices were transferred to a holding chamber (for a minimumthe amplitude of GPSCs was in the range of 50–400 pA. Therefore,of 1.5 hr) where they were maintained in continually perfused oxy-we estimate that per presynaptic action potential, release occurredgenated (95% O2-5% CO2) artificial CSF: 124 mM NaCl, 2.8 mM KCl,at about 8 to 60 release sites (given a conductance per single release2 mM MgSO4, 1.25 mM NaH2PO4, 2 mM CaCl2, 26 mM NaHCO3, 10site of about 0.44 $ 0.02 pS; Nusser et al., 1998). D-amino-5-phos-mM D-Glucose, 0.4 mM ascorbic acid, pH 7.4 when equilibratedphonopentanoic acid (AP5) and 6-cyano-7-nitroquinoxaline-2,with 95% O2-5% CO2, 295 mOsmol.3-dione (CNQX), and nimodipine were purchased from ResearchBiochemicals International. Amphotericin B was used as the perfo-
Electrophysiology rating agent for experiments performed with AP5 and CNQX. Furose-Hippocampal Cultures mide was purchased from Sigma. All Student’s t tests performedWhole-cell perforated patch recordings were performed on pairs or were two tailed. The use of either unpaired or paired Student’s ttriples of interconnected cultured neurons (Bi and Poo, 1998). For tests is noted in the text. One-way ANOVAs followed by Scheffeperforated patch recordings, either gramicidin or amphotericin B pairwise comparisons were used to test for significance betweenwas used for perforation. The recording pipettes were made from multiple comparisons.glass capillaries (VWR), with a resistance of !2 M*. For experiments Acute Hippocampal Slicesusing amphotericin B, the pipettes were tip filled with internal solu- A bipolar stimulating electrode placed in the S. radiatum was usedtion (K-Gluconate, 154 mM; NaCl, 9 mM; MgCl2, 1 mM; HEPES, to activate interneurons (100 #s, 5 V, 0.05 Hz). Whole-cell perforated1 mM; EGTA, 0.2 mM) and then back filled with internal solution patch recordings using gramicidin were made from CA1 pyramidalcontaining amphotericin B (150 to 220 #g/ml, Calbiochem). Experi- neurons (gramicidin and internal solution were prepared as de-ments using gramicidin were tip filled with an internal solution of scribed above). Recordings were made with Kimax-51 electrodes150 mM KCl and 10 mM HEPES, and back filled with this internal with a tip resistance of !6 M*. The external bath solution was thesolution containing gramicidin (25 #g/ml). For experiments using same as the aCSF that the slices had been maintained in, with theBAPTA loading into the postsynaptic cell, breakthrough whole-cell addition of 10 #M CNQX and 25 #M D-AP5. The bath solution wasrecordings were made. In these experiments, the internal solution oxygenated with 95% O2-5% CO2 and continually perfused at awas modified (K-Gluconate, 100 mM; BAPTA, 30 mM; KCl, 15 mM; rate of 2 ml/min and, in some experiments, heated to 35)C with anNaCl, 5 mM; EGTA, 0.5 mM; HEPES, 10 mM; Mg-ATP, 2 mM). The automatic temperature controller (Warner Instruments). The neuronsexternal bath solution was a HEPES-buffered saline (HBS: NaCl, were visualized with a CCD camera (Hamamatsu). Recordings were150 mM; KCl, 3 mM; HEPES, 10 mM; CaCl2, 3 mM; glucose, 5 mM; performed with patch clamp amplifiers (Axopatch 200B; Axon Inc.),MgCl2, 2 mM). The bath was constantly perfused with fresh recording and data were collected and analyzed in similar manner as thatmedium at a slow rate throughout the recording, and all experiments described above for hippocampal cultures.were performed at room temperature. The neurons were visualizedby phase-contrast optics (Nikon Diaphot).
AcknowledgmentsRecordings were performed with patch clamp amplifiers (Axo-patch 200B; Axon Inc.). Signals were filtered at 5 kHz using amplifier
This work was supported by grants from National Institutes of Healthcircuitry. Data was acquired and analyzed using Axoscope 8.0 (Axon(NS 36999), Alberta Heritage Foundation for Medical Research, andInc.). Recordings started after the access resistance had droppedNational Science and Engineering Research Council (to M.A.W.).to between 20 and 30 M*. The series resistance was compensated
to 80% throughout the experiment. In addition, series resistanceReceived: December 16, 2002was monitored throughout the experiments and did not change byRevised: May 30, 2003more than 5% during a typical experiment. For assaying synapticAccepted: July 17, 2003connectivity in culture, each neuron was stimulated at a low fre-Published: August 27, 2003quency (0.05 Hz) by a 1 ms step depolarization from !70 to "20
mV in voltage-clamp mode. All GPSCs were distinguished from ex-Referencescitatory postsynaptic currents by the following criteria: longer decay
times, reversal potentials between !50 and !90 mV, and in somecases, sensitivity to bicuculline methiodide (10 #M; Research Bio- Abbott, L.F. (2000). Integrating with action potentials. Neuron 26,
3–4.chemcals International). In most recordings, the postsynaptic neu-

Coincident Spiking Modifies GABAergic Synapses819
Aizenman, C.D., Manis, P.B., and Linden, D.J. (1998). Polarity of mulation and depletion in cultured rat midbrain neurons. J. Neurosci.19, 4695–4704.long-term synaptic gain change is related to postsynaptic spike
firing at a cerebellar inhibitory synapse. Neuron 21, 827–835. Kakazu, Y., Akaike, N., Komiyama, S., and Nabekura, J. (1999). Regu-lation of intracellular chloride by cotransporters in developing lateralBazhenov, M., Timofeev, I., Steriade, M., and Sejnowski, T.J. (1999).superior olive neurons. J. Neurosci. 19, 2843–2851.Self-sustained rhythmic activity in the thalamic reticular nucleus
mediated by depolarizing GABAA receptor potentials. Nat. Neurosci. Kano, M., Rexhausen, U., Dreessen, J., and Konnerth, A. (1992).2, 168–174. Synaptic excitation produces a long-lasting rebound potentiation
of inhibitory synaptic signals in cerebellar Purkinje cells. Nature 356,Bi, G.Q., and Poo, M.M. (1998). Synaptic modifications in cultured601–604.hippocampal neurons: dependence on spike timing, synaptic
strength, and postsynaptic cell type. J. Neurosci. 18, 10464–10472. Katz, L.C., and Shatz, C.J. (1996). Synaptic activity and the construc-tion of cortical circuits. Science 274, 1133–1138.Bi, G., and Poo, M. (2001). Synaptic modification by correlated activ-
ity: Hebb’s postulate revisited. Annu. Rev. Neurosci. 24, 139–166. Kelsch, W., Hormuzdi, S., Straube, E., Lewen, A., Monyer, H., andMisgeld, U. (2001). Insulin-like growth factor 1 and a cytosolic tyro-Bliss, T.V., and Collingridge, G.L. (1993). A synaptic model of mem-sine kinase activate chloride outward transport during maturationory: long-term potentiation in the hippocampus. Nature 361, 31–39.of hippocampal neurons. J. Neurosci. 21, 8339–8347.Bliss, T.V., and Lomo, T. (1973). Long-lasting potentiation of synapticKlausberger, T., Magill, P.J., Marton, L.F., Roberts, J.D., Cobden,transmission in the dentate area of the anaesthetized rabbit follow-P.M., Buzsaki, G., and Somogyi, P. (2003). Brain-state- and cell-ing stimulation of the perforant path. J. Physiol. 232, 331–356.type-specific firing of hippocampal interneurons in vivo. Nature 421,Cabantchik, Z.I., and Greger, R. (1992). Chemical probes for anion844–848.transporters of mammalian cell membranes. Am. J. Physiol. 262,Koester, H.J., and Sakmann, B. (1998). Calcium dynamics in singleC803–C827.spines during coincident pre- and postsynaptic activity depend onCallaway, J.C., and Ross, W.N. (1995). Frequency-dependent propa-relative timing of back-propagating action potentials and subthresh-gation of sodium action potentials in dendrites of hippocampal CA1old excitatory postsynaptic potentials. Proc. Natl. Acad. Sci. USApyramidal neurons. J. Neurophysiol. 74, 1395–1403.95, 9596–9601.
Carroll, R.C., Lissin, D.V., von Zastrow, M., Nicoll, R.A., and Malenka,Komatsu, Y. (1996). GABAB receptors, monoamine receptors, and
R.C. (1999). Rapid redistribution of glutamate receptors contributespostsynaptic inositol trisphosphate-induced Ca2" release are in-
to long-term depression in hippocampal cultures. Nat. Neurosci. 2,volved in the induction of long-term potentiation at visual cortical
454–460.inhibitory synapses. J. Neurosci. 16, 6342–6352.
Chen, G., Trombley, P.Q., and van den Pol, A.N. (1996). Excitatory Komatsu, Y., and Iwakiri, M. (1993). Long-term modification of inhibi-actions of GABA in developing rat hypothalamic neurones. J. Phys- tory synaptic transmission in developing visual cortex. Neuroreportiol. 494, 451–464.
4, 907–910.Collin, C., Devane, W.A., Dahl, D., Lee, C.J., Axelrod, J., and Alkon, Kyrozis, A., and Reichling, D.B. (1995). Perforated-patch recordingD.L. (1995). Long-term synaptic transformation of hippocampal CA1 with gramicidin avoids artifactual changes in intracellular chloridegamma-aminobutyric acid synapses and the effect of anandamide. concentration. J. Neurosci. Methods 57, 27–35.Proc. Natl. Acad. Sci. USA 92, 10167–10171.
Lee, H.-K., Kameyama, K., Huganir, R.L., and Bear, M.F. (1998).Debanne, D., Gahwiler, B.H., and Thompson, S.M. (1998). Long-term NMDA induces long-term synaptic depression and dephosphoryla-synaptic plasticity between pairs of individual CA3 pyramidal cells tion of the GluR1 subunit of AMPA receptors in the hippocampus.in rat hippocampal slice cultures. J. Physiol. 507, 237–247. Neuron 21, 1151–1162.Feldman, D.E. (2000). Timing-based LTP and LTD at vertical inputs Ling, D.S., and Benardo, L.S. (1995). Activity-dependent depressionto layer II/III pyramidal cells in rat barrel cortex. Neuron 27, 45–56. of monosynaptic fast IPSCs in hippocampus: contributions fromFitzsimonds, R.M., Song, H.J., and Poo, M.M. (1997). Propagation of reductions in chloride driving force and conductance. Brain Res.activity-dependent synaptic depression in simple neural networks. 670, 142–146.Nature 388, 439–448. Lu, J., Karadsheh, M., and Delpire, E. (1999). Developmental regula-Frech, M.J., Deitmer, J.W., and Backus, K.H. (1999). Intracellular tion of the neuronal-specific isoform of K-Cl cotransporter KCC2 inchloride and calcium transients evoked by gamma-aminobutyric postnatal rat brains. J. Neurobiol. 39, 558–568.acid and glycine in neurons of the rat inferior colliculus. J. Neurobiol. Maccaferri G., Roberts, J.D., Szucs, P., Cottingham, C.A., and Som-40, 386–396. ogyi, P. (2000). Cell surface domain specific postsynaptic currentsFroemke, R.C., and Dan, Y. (2002). Spike-timing-dependent synaptic evoked by identified GABAergic neurones in rat hippocampus inmodification induced by natural spike trains. Nature 416, 433–438. vitro. J. Physiol. 524, 91–116.Ganguly, K., Schinder, A.F., Wong, S.T., and Poo, M. (2001). GABA Magee, J.C., and Johnston, D. (1997). A synaptically controlled,itself promotes the developmental switch of neuronal GABAergic associative signal for Hebbian plasticity in hippocampal neurons.responses from excitation to inhibition. Cell 105, 521–532. Science 275, 209–213.
Malenka, R.C., and Nicoll, R.A. (1993). NMDA-receptor-dependentHayashi, Y., Shi, S.H., Esteban, J.A., Piccini, A., Poncer, J.C., andsynaptic plasticity: multiple forms and mechanisms. Trends Neu-Malinow, R. (2000). Driving AMPA receptors into synapses by LTProsci. 16, 521–527.and CaMKII: requirement for GluR1 and PDZ domain interaction.
Science 287, 2262–2267. Markram, H., Lubke, J., Frotscher, M., and Sakmann, B. (1997).Regulation of synaptic efficacy by coincidence of postsynaptic APsHebb, D.O. (1949). The Organization of Behavior (New York: Wileyand EPSPs. Science 275, 213–215.Interscience).Martin, S.J., Grimwood, P.D., and Morris, R.G.M. (2000). SynapticHoffman, D.A., Magee, J.C., Colbert, C.M., and Johnston, D. (1997).plasticity and memory: An evaluation of the hypothesis. Annu. Rev.K" channel regulation of signal propagation in dendrites of hippo-Neurosci. 23, 649–711.campal pyramidal neurons. Nature 387, 869–875.Marty, A., and Llano, I. (1995). Modulation of inhibitory synapses inHopfield, J.J. (1995). Pattern recognition computation using actionthe mammalian brain. Curr. Opin. Neurobiol. 5, 335–341.potential timing for stimulus representation. Nature 376, 33–36.McBain, C.J., and Fisahn, A. (2001). Interneurons unbound. Nat.Hubner, C.A., Stein, V., Hermans-Borgmeyer, I., Meyer, T., Ballanyi,Rev. Neurosci. 2, 11–23.K., and Jentsch, T.J. (2001). Disruption of KCC2 reveals an essential
role of K-Cl cotransport already in early synaptic inhibition. Neuron McLean, H.A., Caillard, O., Ben Ari, Y., and Gaiarsa, J.L. (1996).30, 515–524. Bidirectional plasticity expressed by GABAergic synapses in the
neonatal rat hippocampus. J. Physiol. 496, 471–477.Jarolimek, W., Lewen, A., and Misgeld, U. (1999). A furosemide-sensitive K"-Cl! cotransporter counteracts intracellular Cl! accu- Miles, R., Toth, K., Gulyas, A.I., Hajos, N., and Freund, T.F. (1996).

Neuron820
Differences between somatic and dendritic inhibition in the hippo- and developmental expression patterns of the neuronal K-Cl co-transporter (KCC2) in the rat retina. J. Neurosci. 20, 1414–1423.campus. Neuron 16, 815–823.
Zhang, L.I., Tao, H.W., Holt, C.E., Harris, W.A., and Poo, M. (1998). AMisgeld, U., Deisz, R.A., Dodt, H.U., and Lux, H.D. (1986). The rolecritical window for cooperation and competition among developingof chloride transport in postsynaptic inhibition of hippocampal neu-retinotectal synapses. Nature 395, 37–44.rons. Science 232, 1413–1415.Zhou, Q., Tao, H.W., and Poo, M.M. (2003). Reversal and stabilizationNicoll, R.A., and Malenka, R.C. (1995). Contrasting properties of twoof synaptic modifications in developing visual system. Science 20,forms of long-term potentiation in the hippocampus. Nature 377,1953–1957.115–118.
Nusser, Z., Hajos, N., Somogyi, P., and Mody, I. (1998). Increasednumber of synaptic GABA(A) receptors underlies potentiation athippocampal inhibitory synapses. Nature 395, 172–177.
Owens, D.F., Boyce, L.H., Davis, M.B., and Kriegstein, A.R. (1996).Excitatory GABA responses in embryonic and neonatal corticalslices demonstrated by gramicidin perforated-patch recordings andcalcium imaging. J. Neurosci. 16, 6414–6423.
Payne, J.A., Stevenson, T.J., and Donaldson, L.F. (1996). Molecularcharacterization of a putative K-Cl cotransporter in rat brain. A neu-ronal-specific isoform. J. Biol. Chem. 271, 16245–16252.
Plotkin, M.D., Snyder, E.Y., Hebert, S.C., and Delpire, E. (1997).Expression of the Na-K-2Cl cotransporter is developmentally regu-lated in postnatal rat brains: a possible mechanism underlyingGABA’s excitatory role in immature brain. J. Neurobiol. 33, 781–795.
Rivera, C., Voipio, J., Payne, J.A., Ruusuvuori, E., Lahtinen, H.,Lamsa, K., Pirvola, U., Saarma, M., and Kaila, K. (1999). The K"/Cl!co-transporter KCC2 renders GABA hyperpolarizing during neuronalmaturation. Nature 397, 251–255.
Rohrbough, J., and Spitzer, N.C. (1996). Regulation of intracellularCl! levels by Na(")-dependent Cl! cotransport distinguishes depo-larizing from hyperpolarizing GABAA receptor-mediated responsesin spinal neurons. J. Neurosci. 16, 82–91.
Schoepfle, G.M., and Erlanger, J. (1941). The action of temperatureon the excitability, spike height and configuration, and the refractoryperiod observed in the responses on single medullated nerve fibers.Am. J. Physiol. 134, 694–704.
Smith, R.L., Clayton, G.H., Wilcox, C.L., Escudero, K.W., and Staley,K.J. (1995). Differential expression of an inwardly rectifying chlorideconductance in rat brain neurons: a potential mechanism for cell-specific modulation of postsynaptic inhibition. J. Neurosci. 15, 4057–4067.
Staley, K.J., and Mody, I. (1992). Shunting of excitatory input todentate gyrus granule cells by a depolarizing GABAA receptor-medi-ated postsynaptic conductance. J. Neurophysiol. 68, 197–212.
Strange, K., Singer, T.D., Morrison, R., and Delpire, E. (2000). Depen-dence of KCC2 K-Cl cotransporter activity on a conserved carboxyterminus tyrosine residue. Am. J. Physiol. Cell Physiol. 279, C860–C867.
Stuart, G., Spruston, N., Sakmann, B., and Hausser, M. (1997). Actionpotential initiation and backpropagation in neurons of the mamma-lian CNS. Trends Neurosci. 20, 125–131.
Sun, D., and Murali, S.G. (1999). Na"-K"-2Cl! cotransporter inimmature cortical neurons: A role in intracellular Cl! regulation. J.Neurophysiol. 81, 1939–1948.
Takeya, M., Hasuo, H., and Akasu, T. (2002). Effects of temperatureincrease on the propagation of presynaptic action potentials in thepathway between the Schaffer collaterals and Hippocampal CA1neurons. Neurosci. Res. 42, 175–185.
Thompson, S.M., and Gahwiler, B.H. (1989a). Activity-dependentdisinhibition. I. Repetitive stimulation reduces IPSP driving forceand conductance in the hippocampus in vitro. J. Neurophysiol. 61,501–511.
Thompson, S.M., and Gahwiler, B.H. (1989b). Activity-dependentdisinhibition. II. Effects of extracellular potassium, furosemide, andmembrane potential on ECl- in hippocampal CA3 neurons. J. Neuro-physiol. 61, 512–523.
Vardi, N., Zhang, L.L., Payne, J.A., and Sterling, P. (2000). Evidencethat different cation chloride cotransporters in retinal neurons allowopposite responses to GABA. J. Neurosci. 20, 7657–7663.
Vu, T.Q., Payne, J.A., and Copenhagen, D.R. (2000). Localization




















