
Coincident signalling between the Gi/Go-coupled δ-opioid receptor and the Gq-coupled m3 muscarinic...
13
Journal of Neurochemistry, 2001, 76, 1688–1700 Coincident signalling between the Gi/Go-coupled d-opioid receptor and the Gq-coupled m3 muscarinic receptor at the level of intracellular free calcium in SH-SY5Y cells A. Yeo,* D. S. K. Samways,* C. E. Fowler,* F. Gunn-Moore² ,1 and G. Henderson* Departments of *Pharmacology and ²Biochemistry, University of Bristol, Bristol, UK Abstract In SH-SY5Y cells, activation of d-opioid receptors with [D-Pen 2,5 ]-enkephalin (DPDPE; 1 mM) did not alter the intracellular free Ca 21 concentration [Ca 21 ] i . However, when DPDPE was applied during concomitant Gq-coupled m3 muscarinic receptor stimulation by carbachol or oxotremorine- M, it produced an elevation of [Ca 21 ] i . The DPDPE-evoked increase in [Ca 21 ] i was abolished when the carbachol- sensitive intracellular Ca 21 store was emptied. There was a marked difference between the concentration–response relationship for the elevation of [Ca 21 ] i by carbachol (EC 50 13 mM, Hill slope 1) and the concentration–response relation- ship for carbachol’s permissive action in revealing the d-opioid receptor-mediated elevation of [Ca 21 ] (EC 50 0.7 mM; Hill slope 1.8). Sequestration of free G protein bg dimers by transient transfection of cells with a bg binding protein (residues 495– 689 of the C terminal tail of G protein-coupled receptor kinase 2) reduced the ability of d opioid receptor activation to elevate [Ca 21 ] i . However, DPDPE did not elevate either basal or oxotremorine-M-evoked inositol phosphate production indicat- ing that d-opioid receptor activation did not stimulate phospholipase C. Furthermore, d-opioid receptor activation did not result in the reversal of muscarinic receptor desensi- tization, membrane hyperpolarization or stimulation of sphin- gosine kinase. There was no coincident signalling between the d-opioid receptor and the lysophosphatidic acid receptor which couples to elevation of [Ca 21 ] i in SH-SY5Y cells by a PLC-independent mechanism. In SH-SY5Y cells the coinci- dent signalling between the endogenously expressed d-opioid and m3 muscarinic receptors appears to occur in the receptor activation-Ca 21 release signalling pathway at a step after the activation of phospholipase C. Keywords: Ca 21 mobilization, d-opioid receptors, m3 muscarinic receptors, phospholipase C, receptor cross-talk, SH-SY5Y cells. J. Neurochem. (2001) 76, 1688–1700. G protein-coupled receptor activation results in the modu- lation of a number of cell signalling pathways such as adenylyl cyclase, phospholipase C (PLC) and ion channels (Neer 1995). In addition to these well documented, linear signalling pathways there is now considerable evidence for more complex coincident signalling, or cross-talk, between G protein-coupled receptors and between G protein-coupled receptors and other types of receptor (Neer 1995; Selbie and Hill 1998). In a number of cell types including neurones and neuronal cell lines, astrocytes, smooth muscle and mammalian expression systems, Gi/Go-coupled receptor activation alone does not activate the PLC/IP 3 /intracellular Ca 21 release pathway. However, in the presence of concomitant Gq coupled receptor activation a Gi/Go-coupled receptor- mediated response was revealed (Gerwins and Fredholm 1992; Okajima et al. 1993; Dickenson and Hill 1994; Connor and Henderson 1996; Connor et al. 1996, 1997a, 1997b; Biber et al. 1997; Tomura et al. 1997; Toms and Roberts 1999; Cho et al. 2000). This Gi/Go-coupled 1688 q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1688–1700 Received July 10, 2000; revised manuscript received November 3, 2000; accepted November 8, 2000. Address correspondence and reprint requests to G. Henderson, Department of Pharmacology, Medical Sciences Building, University of Bristol, University Walk, Bristol, BS8 1TD, UK. E-mail: [email protected] 1 Present address: Department of Preclinical Veterinary Sciences, Royal (Dick) College of Veterinary Studies, University of Edinburgh, Summerhall, Edinburgh EH9 1QH, UK. Abbreviations used: BSA, bovine serum albumin; carbachol, carbamylcholine chloride; CHO-d, Chinese hamster ovary cells transfected with the mouse d-opioid receptor; DHS, dl-threo-dihydro- sphingosine; DPDPE, [d-Pen 2,5 ]-enkephalin; PCL, phospholipase C; GRK, G protein-coupled receptor kinase; GFP, green fluorescent protein; IP 1 , inositol-1-phosphate; [Ca 21 ] i , intracellular calcium; LPA, lysophosphatidic acid; oxo-M, oxotremorine-M; SEM, standard error of the mean; UTP, uridine triphosphate.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Coincident signalling between the Gi/Go-coupled δ-opioid receptor and the Gq-coupled m3 muscarinic...

Journal of Neurochemistry, 2001, 76, 1688±1700
Coincident signalling between the Gi/Go-coupled d-opioid
receptor and the Gq-coupled m3 muscarinic receptor at the
level of intracellular free calcium in SH-SY5Y cells
A. Yeo,* D. S. K. Samways,* C. E. Fowler,* F. Gunn-Moore²,1 and G. Henderson*
Departments of *Pharmacology and ²Biochemistry, University of Bristol, Bristol, UK
Abstract
In SH-SY5Y cells, activation of d-opioid receptors with
[D-Pen2,5]-enkephalin (DPDPE; 1 mM) did not alter the
intracellular free Ca21 concentration [Ca21]i. However, when
DPDPE was applied during concomitant Gq-coupled m3
muscarinic receptor stimulation by carbachol or oxotremorine-
M, it produced an elevation of [Ca21]i. The DPDPE-evoked
increase in [Ca21]i was abolished when the carbachol-
sensitive intracellular Ca21 store was emptied. There was a
marked difference between the concentration±response
relationship for the elevation of [Ca21]i by carbachol (EC50
13 mM, Hill slope 1) and the concentration±response relation-
ship for carbachol's permissive action in revealing the d-opioid
receptor-mediated elevation of [Ca21] (EC50 0.7 mM; Hill slope
1.8). Sequestration of free G protein bg dimers by transient
transfection of cells with a bg binding protein (residues 495±
689 of the C terminal tail of G protein-coupled receptor kinase
2) reduced the ability of d opioid receptor activation to elevate
[Ca21]i. However, DPDPE did not elevate either basal or
oxotremorine-M-evoked inositol phosphate production indicat-
ing that d-opioid receptor activation did not stimulate
phospholipase C. Furthermore, d-opioid receptor activation
did not result in the reversal of muscarinic receptor desensi-
tization, membrane hyperpolarization or stimulation of sphin-
gosine kinase. There was no coincident signalling between
the d-opioid receptor and the lysophosphatidic acid receptor
which couples to elevation of [Ca21]i in SH-SY5Y cells by a
PLC-independent mechanism. In SH-SY5Y cells the coinci-
dent signalling between the endogenously expressed d-opioid
and m3 muscarinic receptors appears to occur in the receptor
activation-Ca21 release signalling pathway at a step after the
activation of phospholipase C.
Keywords: Ca21 mobilization, d-opioid receptors, m3
muscarinic receptors, phospholipase C, receptor cross-talk,
SH-SY5Y cells.
J. Neurochem. (2001) 76, 1688±1700.
G protein-coupled receptor activation results in the modu-
lation of a number of cell signalling pathways such as
adenylyl cyclase, phospholipase C (PLC) and ion channels
(Neer 1995). In addition to these well documented, linear
signalling pathways there is now considerable evidence for
more complex coincident signalling, or cross-talk, between
G protein-coupled receptors and between G protein-coupled
receptors and other types of receptor (Neer 1995; Selbie and
Hill 1998).
In a number of cell types including neurones and neuronal
cell lines, astrocytes, smooth muscle and mammalian
expression systems, Gi/Go-coupled receptor activation alone
does not activate the PLC/IP3/intracellular Ca21 release
pathway. However, in the presence of concomitant Gq
coupled receptor activation a Gi/Go-coupled receptor-
mediated response was revealed (Gerwins and Fredholm
1992; Okajima et al. 1993; Dickenson and Hill 1994;
Connor and Henderson 1996; Connor et al. 1996, 1997a,
1997b; Biber et al. 1997; Tomura et al. 1997; Toms and
Roberts 1999; Cho et al. 2000). This Gi/Go-coupled
1688 q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1688±1700
Received July 10, 2000; revised manuscript received November 3,
2000; accepted November 8, 2000.
Address correspondence and reprint requests to G. Henderson,
Department of Pharmacology, Medical Sciences Building, University of
Bristol, University Walk, Bristol, BS8 1TD, UK.
E-mail: [email protected] address: Department of Preclinical Veterinary Sciences,
Royal (Dick) College of Veterinary Studies, University of Edinburgh,
Summerhall, Edinburgh EH9 1QH, UK.
Abbreviations used: BSA, bovine serum albumin; carbachol,
carbamylcholine chloride; CHO-d, Chinese hamster ovary cells
transfected with the mouse d-opioid receptor; DHS, dl-threo-dihydro-
sphingosine; DPDPE, [d-Pen2,5]-enkephalin; PCL, phospholipase C;
GRK, G protein-coupled receptor kinase; GFP, green ¯uorescent
protein; IP1, inositol-1-phosphate; [Ca21]i, intracellular calcium; LPA,
lysophosphatidic acid; oxo-M, oxotremorine-M; SEM, standard error of
the mean; UTP, uridine triphosphate.

receptor response appeared to be mediated by free G protein
bg dimers because it was reduced under conditions where
free bg dimers were sequestered by over expressed bg
binding proteins such as the bg binding domain of the G
protein-coupled receptor kinase 2 (GRK2) or the a subunit
of transducin (Chan et al. 2000; Selbie et al. 1997). A
number of different modes of action for free bg dimers
in receptor cross-talk have been proposed. These include
excess bg dimers associating with free aq dimers to
enhance the turnover of Gq (Quitterer and Lohse 1999),
synergistic activation of PLCb isozymes by aq and bg
dimers (Zhu and Birnbaumer 1996; Selbie and Hill 1998) or
sensitization of the IP3 receptor to IP3 (Neylon et al. 1998;
Xu et al. 1996; Zeng et al. 1996).
We have previously demonstrated that activation of
several types of Gi/Go-coupled receptors (d-opioid, m-opioid,
ORL1, sst2 and NPY Y2 receptors) in the SH-SY5Y human
neuroblastoma cell line required concomitant m3 muscarinic,
Gq-coupled receptor activation to reveal the Gi/Go-coupled
receptor-mediated elevation of intracellular calcium ([Ca21]i)
(Connor and Henderson 1996; Connor et al. 1996, 1997a,
1997b). The Gi/Go-coupled receptor-mediated response was
abolished by pretreatment with pertussis toxin and by deple-
tion of the [Ca21]i stores with thapsigargin. The requirement
for concomitant Gq-coupled receptor activation was absolute
in that if the Gi/Go-coupled receptor agonist was applied
simultaneously with a muscarinic receptor antagonist then
the Gi/Go-coupled receptor-mediated response was also
abolished.
In the present paper we have further characterized the
nature of the coincident signalling between the endogen-
ously expressed d-opioid and m3 muscarinic receptors. We
have sought to determine (i) if the d-opioid and m3
muscarinic receptors release Ca21 from the same intracel-
lular Ca21 store; (ii) if activation of the d-opioid receptor
reverses m3 muscarinic receptor desensitization; (iii) the
role of bg dimers from Gi/Go in the d-opioid receptor
elevation of [Ca21]I; (iv) if the coincident signalling is
mediated through concomitant activation of PLCb; and (v) if
the d-opioid receptor exhibits cross-talk with a receptor
which elevates [Ca21]i through a PLC-independent path-
way. Some of this work has appeared previously in abstract
form (Yeo et al. 1996; Yeo and Henderson 1997a,1997b).
Materials and methods
Cell culture
SH-SY5Y cells were cultured as described previously (Connor and
Henderson 1996). Chinese hamster ovary cells transfected with the
mouse d-opioid receptor (CHO-d) were cultured in Nutrient Mixture
Ham's F12 Medium supplemented with penicillin (100 i.u./mL),
streptomycin (100 mg/mL), amphotericin B (2.5 mg/mL), geneticin
(100 mg/mL) and fetal bovine serum (10%) in a humidi®ed
incubator with 5% CO2. For the monolayer studies, cells were
plated on to plastic slides and cultured in Leighton tubes (Costar),
whereas for single cell imaging experiments cells were plated at a
low density on to glass coverslips. For measurement of inositol
phosphate production, 4 � 12-well plates were seeded with cells
and the experiments performed once the cells had reached
con¯uence.
Cell transfection
SH-SY5Y cells were transiently cotransfected with a mutant green
¯uorescent protein (S65T GFP) as a marker for successful
transfection and a G protein bg dimer binding protein comprising
part of the C terminal region (amino acids 495±689) of G protein-
coupled receptor kinase 2 (GRK2). Cells were transfected at
378C by incubation for 2 h with a mixture of Tfx-50 (Promega)
and the plasmids containing the construct for S65T GFP (pCMV-
GFP) and the construct for the bg dimer binding protein
(pcDNAneo-GRK2495±689) at a Tfx : DNA ratio of 3 : 1 and a
pCMV-GFP : pcDNAneo-GRK2495±689 ratio of 1 : 4. 24 h after
transfection this protocol resulted in 30±50% of cells exhibiting
GFP ¯uorescence observed using a 490-nm excitation ®lter and a
520-nm broad-pass emission ®lter. GFP-positive cells were assumed
to be coexpressing the G protein bg dimer binding protein.
Measurement of intracellular Ca21
Intracellular free Ca21 concentration ([Ca21]i) was measured using
the ¯uorescent Ca21-sensitive dye fura-2 as described previously
(Connor and Henderson 1996). Brie¯y, cells were superfused with
a buffer containing: NaCl 140 mM, KCl 2 mM, CaCl2 2.5 mM,
MgCl2 1 mM, HEPES 10 mM, glucose 10 mM, sucrose 40 mM,
bovine serum albumin (BSA) 0.05%, pH 7.3. In those experiments
in which lysophosphatidic acid was studied, fatty acid free BSA
was used.
Monolayers
After loading fura-2 the plastic slides were ®xed in a quartz cuvette.
The cuvette was then placed in an LS-5B Perkin-Elmer spectro-
¯uorimeter and perfused with buffer (4 mL/min at 378C). Drugs
were added to the perfusion buffer in known concentrations. The
spectro¯uorimeter was controlled by a computer running a Perkin-
Elmer software package. The fura-2 loaded cells were alternately
exposed to light at 340 nm and 380 nm and the emission of the
cells at 510 nm was recorded.
Single cell imaging
After loading fura-2, the coverslips were transferred into a 1-mL
chamber (maintained at 378C), placed onto an inverted microscope
(Nikon Diaphot) and superfused with buffer at 4 mL/min. Drugs
were added to the perfusion buffer in known concentrations. Cells
were initially categorized as GFP-positive or GFP-negative (see
above) and then alternately exposed to light at 340 nm and 380 nm
and the intensity of the emission of the cells at 510 nm was
measured using a Photonic Science camera. Ionvision software was
used to collect data and also to control the position of the ®lter
wheel selecting the excitation wavelength. Fluorescence intensity
was measured at the two emission wavelengths every 5 s or every
20 s (acquisition speed being faster during drug application and
slower during periods of washout). Background ¯uorescence
intensity was taken from an area of the coverslip with no cells.
Groups of 5±20 cells were imaged in any single experiment.
Co-incident signalling between d-opioid and m3 muscarinic receptors 1689
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1688±1700

Measurement of [3H] inositol phosphate accumulation
Phosphoinositide hydrolysis was determined by monitoring the
accumulation of inositol monophosphate in the presence of LiCl as
described previously (Toms et al. 1995). SH-SY5Y and CHO-d
cells were incubated with 4 mCi/mL [3H]inositol for 20±24 h at
378C to allow incorporation of [3H] into inositol phosphates.
Prelabelled cells were washed with a buffer containing: NaCl
118 mM, KCl 4.7 mM, NaHCO3 25 mM, KHPO4 1.2 mM, CaCl21.2 mM, MgSO4 1.2 mM, HEPES 10 mM, glucose 10 mM and
then incubated with Li1 (10 mm) for 20 min. Drugs were then
added as required, the reaction was terminated by aspiration and
addition of 500 mL ice cold perchloric acid (7.5%). Total inositol
phosphates were extracted by freeze thawing, and IP1 separated
using Dowex chromatography. The amount of [3H]-IP1 was
determined by scintillation counting.
Whole-cell patch clamp
Whole-cell patch clamp experiments were performed at 19±218C
as described previously (Seward and Henderson 1990). The super-
fusing solution consisted of: NaCl 140 mM, KCl 2 mM, CaCl22.5 mM, MgCl2 1 mM, HEPES 10 mM, glucose 10 mM, sucrose
40 mM, pH 7.4; and the pipette solution: KCl 140 mM, MgCl21 mM, CaCl2 1 mM, EGTA 10 mM, Na2 ATP 5 mM, NaGTP
0.2 mM, HEPES 10 mM, pH 7.2. The cells were superfused at
2.5 mL/min. Pipette resistances were 2±5 mV.
Drugs and chemicals
N-acetylsphingosine, apyrase, atropine methylbromide, carbamyl-
choline chloride (carbachol), dl-threo-dihydrosphingosine (DHS),
[d-Pen2,5]-enkephalin (DPDPE), fura-2AM, lysophosphatidic acid
(LPA), mecamylamine hydrochloride, oxotremorine-M (oxo-M)
and uridine triphosphate (UTP) were obtained from (Sigma).
[3H] inositol was obtained from American Radiolabelled Chemicals
Ltd.
Data analysis
Data are reported as mean ^ standard error of the mean (SEM);
statistical comparisons were made using either an unpaired
Student's t-test or a Chi-square test as appropriate; p , 0.05 was
considered signi®cant.
Results
Coincident signalling between d-opioid receptors and
m3 muscarinic receptors
When applied to SH-SY5Y cells the d-opioid receptor
agonist DPDPE on its own did not elevate [Ca21]i whereas
either carbachol (1 mm, Fig. 1a) or oxo-M (1 mm), acting at
Gq-coupled muscarinic receptors in these cells, evoked a
biphasic elevation of [Ca21]i. However, in the continued
presence of carbachol or oxo-M, application of DPDPE now
evoked a rapid elevation of [Ca21]i (Fig. 1a).
It was not possible with population studies on cell
monolayers to determine whether the same cells were
responding to muscarinic and d-opioid receptor activation.
We therefore employed single cell imaging of [Ca21]i to
identify the cells that responded to Gq-and Gi/Go-coupled
receptor activation. 95% of cells studied responded to
oxo-M (1 mm) with a rise in [Ca21]i (Table 1a). In the
absence of oxo-M a few cells (5%) responded to DPDPE
(1 mm) alone but there was no correlation between a lack of
response to oxo-M and a response to DPDPE in the absence
of oxo-M. In the continued presence of oxo-M (1 mm) 60%
of the oxo-M-responding cells exhibited an elevation of
[Ca21]i in response to DPDPE (1 mm).
We next sought to compare the concentration-dependence
of the carbachol elevation of [Ca21]i with that for its
permissive role in revealing the d-opioid receptor-mediated
elevation of [Ca21]i. In monolayers carbachol alone evoked
a concentration-dependent elevation of [Ca21]i (Figs 1b
and c). The EC50 for carbachol was 13 mm (95% con®dence
limits 1±137 mm), the Hill slope was not signi®cantly
different from unity and the response reached a maximum at
1 mm. In these experiments mecamylamine (100 mm) was
present to exclude any contribution at high agonist con-
centrations of nicotinic receptors present in SH-SY5Y cells
(Forsythe et al. 1992). The maximum response to carbachol
(3 mm) in the presence of mecamylamine was 93 ^ 14%
(n � 3) of the response seen to a maximally effective
concentration of the selective muscarinic receptor agonist,
oxo-M (30 mm). We then determined the concentration-
dependence of the permissive action of carbachol to reveal
the d-opioid receptor-mediated elevation of [Ca21]i by
applying a ®xed concentration of DPDPE (30 nm) in the
presence of increasing concentrations of carbachol (Figs 1b
and c). The amplitude of the response evoked by DPDPE
(30 nm) increased as the concentration of carbachol was
increased, reaching 50% of its maximum in the presence of
720 nm carbachol (95% con®dence limits 305 nm to
1.7 mm). The concentration±response curve for carbachol
had a Hill slope of 1.8 and was maximal at 10 mm carbachol.
Thus the concentration±response curve for carbachol's
permissive action to reveal the DPDPE elevation of
[Ca21]i lies approximately 18-fold to the left of that for
carbachol alone elevating [Ca21]i.
d-Opioid receptor activation releases Ca21 from the
same intracellular stores as m3 muscarinic receptor
activation
To determine whether d-opioid receptor activation released
Ca21 from the same intracellular stores as Gq-coupled
muscarinic receptor activation we sought to empty the
muscarinic receptor-sensitive stores by exposing the cells to
a maximally effective concentration of carbachol in the
absence of extracellular Ca21. In the absence of extra-
cellular Ca21 the carbachol-sensitive Ca21 stores should be
prevented from re®lling through capacitative Ca21 entry
pathways (Grudt et al. 1996). When carbachol (1 mm) was
applied in the absence of extracellular Ca21 it evoked a
large transient rise in [Ca21]i that returned to baseline levels
even in the continued presence of the drug (Fig. 2).
Following exposure to high carbachol, the cells failed to
1690 A. Yeo et al.
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1688±1700

respond when challenged with another Gq-coupled receptor
agonist, bradykinin (1 mm) that prior to carbachol evoked a
rise in [Ca21]i, indicating that indeed the Ca21 stores had
been emptied (data not shown). In the absence of extra-
cellular Ca21 but in the presence of a low concentration of
carbachol (1 mm) (i.e. when the intracellular Ca21 stores had
not been depleted) DPDPE (30 nm) evoked an elevation in
[Ca21]i (Fig. 2a). However, when applied in the presence of
the high carbachol concentration (1 mm), in the absence of
extracellular Ca21 (i.e. when the intracellular Ca21 stores
were depleted), DPDPE (30 nm) failed to evoke a rise in
[Ca21]i (Fig. 2b). These data demonstrate that the d-opioid
receptor-mediated rise in [Ca21]i resulted from Ca21 release
from the same stores as the Gq-coupled muscarinic receptor.
d-Opioid receptor activation does not reverse m3
muscarinic receptor desensitization
One possible explanation for the coincident nature of the
d-opioid receptor and muscarinic receptor elevations of
[Ca21]i is that stimulation of the d-opioid receptors in some
way reversed the desensitization of muscarinic receptors
developing during the continuous exposure to the muscarinic
agonist. If this was the case then it would be expected that
the DPDPE-evoked rise in [Ca21]i would be smallest if
applied at the start of the exposure to carbachol and be larger
if applied at a later time point as the desensitization of the
muscarinic receptors increased. To investigate this, DPDPE
(30 nm) was applied in the continued presence of carbachol
(1 mm) at various time points (0, 1, 2 and 10 min) after the
Fig. 1 Carbachol concentration±response
relationships. (a) The trace represents a
continuous record of [Ca21]i in a monolayer
of SH-SY5Y cells. DPDPE alone did not
elevate [Ca21]i whereas carbachol pro-
duced a biphasic elevation of [Ca21]i. In the
presence of carbachol, DPDPE now also
elevated [Ca21]i. Drugs were applied for the
periods indicated by the bars. (b) The two
traces represent continuous records of
[Ca21]i in monolayers of SHSY5Y cells
taken from two halves of the same cover-
slip. Carbachol produces a concentration-
dependent elevation of [Ca21]i. At the
low carbachol concentration (300 nM) the
response to DPDPE (30 nM) was small,
whereas in a higher carbachol concentra-
tion (3 mM) the response evoked by DPDPE
(30 nM) was larger. (c) The concentration-
response curve for the peak elevation of
[Ca21]i by carbachol lies to the right of that
for carbachol revealing the DPDPE-evoked
elevation of [Ca21]i. The closed circles
represent the concentration±response
curve for the peak elevation of [Ca21]i by
carbachol. The experiment was performed
in the presence of the nicotinic antagonist
mecamylamine (100 mM). The open circles
represent the concentration-response curve
for carbachol revealing the DPDPE evoked
elevation of [Ca21]i. Monolayers of cells
were exposed to a ®xed concentration of
DPDPE (30 nM) in the presence of different
concentrations of carbachol. Each point
represents the mean data obtained from
between 5 and 9 experiments, the bars
represent the SEM.
Co-incident signalling between d-opioid and m3 muscarinic receptors 1691
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1688±1700

beginning of the carbachol application (Fig. 3). There was
no signi®cant difference in the responses evoked at the
different time points. The mechanism by which DPDPE
elevated [Ca21]i in the continued presence of carbachol
therefore does not appear to be reversal of muscarinic
receptor desensitization.
Role of G protein bg dimers in d-opioid
receptor-mediated elevation of [Ca21]i
G protein-coupled receptor kinases (GRK) 2 and 3 bind to
bg dimers using a plekstrin homology domain contained in
their C terminal regions (Koch et al. 1993). This interaction
normally facilitates the translocation of GRKs to the plasma
membrane. Use can also be made of this interaction to
examine the role of bg dimers in cellular processes because
in cells overexpressing the C terminal portion of GRK 2 or 3
this binding protein will sequester bg dimers and thus
inactivate them (Inglese et al. 1994).
SH-SY5Y cells were transiently cotransfected with two
plasmids, one containing an insert coding for GFP and the
other an insert encoding for the C terminal region (amino
acids 495±689) of GRK2. Successfully transfected cells
were identi®ed by their green ¯uorescence. Because only a
proportion of cells (30±50%) were successfully transfected
we used single cell imaging to determine if transfection with
a bg binding protein decreased the response to m3
muscarinic and d-opioid receptor activation. Even in non-
transfected cells there was considerable variability in the
amplitude of the responses to m3 muscarinic and d-opioid
receptor activation in different cells. Therefore in these
experiments we considered a cell to be responsive if the
application of an agonist raised [Ca21]i above basal levels.
In the population of successfully transfected cells the
number of cells responding to oxo-M (1 mm) with an
increase in [Ca21]i was not different from cells transfected
with the vector alone or from wild-type cells (Table 1b).
However, the proportion of cells responding to DPDPE
(1 mm) was signi®cantly lower in cells transfected with the
bg binding protein (Chi-square test, p , 0.05). These data
indicate that the d-opioid receptor-mediated elevation of
[Ca21]i was mediated through bg dimers.
Potential role of PLCb activation in d-opioid
receptor-mediated elevation of [Ca21]i
To examine whether Gi/Go-coupled receptor activation
altered PLCb activity we measured inositol-1-phosphate
Table 1 (a) Number of SH-SY5Y cells exhibiting an elevation of [Ca21]i in response to oxo-M, DPDPE and DPDPE in the presence of oxo-M
Drug treatment Number of cells responding
Oxo-M (1 mM) 112 of 118 (95%)
DPDPE (1 mM) 6 of 118 (5%)
DPDPE (1 mM) in the presence of oxo-M (1 mM) 67 of 112 (60%)
Data are from eight coverslips on each of which 11±23 cells were examined.
(b) Effect of transient transfection of SH-SY5Y cells with a bg binding protein on the elevation of [Ca21]i by oxo-M and DPDPE in the presence of
oxo-M
Number of cells responding
Non-transfected (GFP-negative) Transfected (GFP-positive)
Transfection Oxo-M (1 mM)
DPDPE (1 mM)
in the presence of oxo-M (1 mM) Oxo-M (1 mM)
DPDPE (1 mM)
in the presence of oxo-M (1 mM)
bg binding protein 216 of 236 145 of 216 36 of 43 10 of 36*
(92%) (67%) (84%) (28%)
[0.950] [0�.278] [0�.650]1 [0.000]11
Vector control 58 of 67 28 of 58 24 of 28 16 of 24
(87%) (48%) (86%) (66%)
Cells were transiently transfected with a mixture of two plasmids; one containing the insert for GFP and one containing either the insert for amino
acids 495±689 of GRK2 (bg binding protein) or no insert as control (vector control). Cells expressing GFP (30±50% of total) were assumed to also
be expressing the bg binding protein. For each drug treatment the data were obtained from 8±10 coverslips on each of which a number of cells
were examined. *Indicates a signi®cant difference (p , 0.05, Chi-square test) compared with non-transfected cells and with cells transfected with
the vector alone. Values in square brackets are the median increases in the ratio of ¯uorescence emission at 340 : 380 nm. 1 and11 indicate
signi®cant difference from non-transfected cells at the 0.01 and 0.001 levels, respectively, determined using a Mann±Whitney U-test.
1692 A. Yeo et al.
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1688±1700

(IP1) accumulation in the presence of Li1. Oxo-M (0.1±
100 mm) evoked a concentration-dependent stimulation of
IP1 accumulation (Fig. 4a). The concentration±response
curve for oxo-M stimulation of IP1 production lies slightly
to the left of that for oxo-M elevation of [Ca21]i, with both
curves having Hill slopes close to unity (Fig. 4a). We then
used a concentration of oxo-M (1 mm) that evoked a similar
rise in [Ca21]i as did a maximally effective concentration of
DPDPE (1 mm). DPDPE did not alter either the basal level
of IP1 production or the increase in IP1 production evoked
by oxo-M at 2, 5 or 20 min (Fig. 4b).
It was surprising that in SH-SY5Y cells the coincident
signalling between d-opioid receptors muscarinic receptors
did not result in enhanced PLC activation and thus an
increase in IP1 production given that other workers, using
other cell types have reported an interaction at the level of
PLC (Gerwins and Fredholm 1992; Okajima et al. 1993;
Dickenson and Hill 1994; Biber et al. 1997; Tomura et al.
1997; Selbie and Hill 1998; Chan et al. 2000). To ensure
that the lack of PLC activation in SH-SY5Y cells was not
some methodological artefact we examined whether we
could observe d-opioid receptor activation of PLC in another
cell type. In monolayers of CHO cells stably transfected
with the d-opioid receptor (CHO-d cells), DPDPE (30±
1000 nm) applied alone did not elevate [Ca21]i (Fig. 5a).
However, in the presence of UTP (1 mm), an agonist at Gq
Fig. 2 Depletion of the carbachol-releasa-
ble Ca21 pool prevented the DPDPE ele-
vation of [Ca21]i. The traces represent
continuous records of [Ca21]i in monolayers
of SH-SY5Y cells. Drugs were applied for
the periods indicated by the bars. (a) In the
absence of extracellular Ca21 both carba-
chol (1 mM) and DPDPE (in the continued
presence of carbachol) evoked a transient
increase in [Ca21]i. (b) In the absence of
extracellular Ca21 a maximally effective
concentration of carbachol (1 mM) caused a
large transient rise in [Ca21]i that declined
back to baseline levels in the continued
presence of carbachol. Thereafter, in the
continuous presence of carbachol, DPDPE
(30 nM) failed to elevate [Ca21]i. The insert
shows an enlarged portion of the trace
during the application of DPDPE.
Co-incident signalling between d-opioid and m3 muscarinic receptors 1693
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1688±1700

coupled P2Y receptors in these cells which evoked a
biphasic increase in [Ca21]i, DPDPE now caused a rapid
elevation in [Ca21]i. We then measured IP1 accumulation in
the presence of Li1 in CHO-d cells. UTP (1±1000 mm)
evoked a concentration-dependent elevation of IP1 accumu-
lation in CHO-d cells (Fig. 5b). In contrast to SH-SY5Y
cells, in CHO-d cells the concentration-response curve for
agonist stimulation of IP1 production lies slightly to the right
of that for agonist elevation of [Ca21]i, both curves having
Hill slopes close to unity (Fig. 5b).We used a concentration
of UTP (10 mm) which evoked a similar rise in [Ca21]i as a
maximally effective concentration of DPDPE (1 mm).
DPDPE (1 mm) potentiated the UTP-evoked accumulation
of IP1 at 2, 5 and 20 min but did not affect the basal
accumulation of IP1 at the same time points (Fig. 5c). Thus,
in the presence of concomitant Gq coupled receptor activa-
tion d receptor activation stimulates PLC in CHO-d cells but
not SH-SY5Y cells.
Sphingosine kinase does not mediate the d-opioid
receptor-mediated elevation of [Ca21]i
Activation of sphingosine kinase leads to the generation
of sphingosine-1-phosphate which is an alternative second
messenger for intracellular Ca21 mobilization (Beaven
Fig. 3 The amplitude of the DPDPE evoked elevation of [Ca21]i
was independent of the time after the start of the carbachol applica-
tion at which DPDPE was applied. (a) Three superimposed traces of
[Ca21]i from the same coverslip of SH-SY5Y cells. These traces
show (i) a control response to carbachol (1 mM), (ii) a response to
DPDPE (30 nM) applied 1 min after the beginning of the carbachol
exposure, and (iii) a response to DPDPE (30 nM) applied 10 min
after the beginning of the carbachol exposure. To overlay the traces
the ratios of 340/380 nm have been normalized to 1 at the start of
each trace. (b) Comparison of the rise in [Ca21]i evoked by DPDPE
(30 nM) applied at 0, 1, 2 and 10 mins after the beginning of the
carbachol exposure. Histograms represent the mean and SEM from
4 to 7 experiments and each response has been normalized to the
amplitude of the response to DPDPE applied 10 min after the start
of the carbachol exposure.
Fig. 4 Effect of oxo-M and DPDPE on IP1 production. (a) Concen-
tration±response relationships for oxo-M-evoked elevation of [Ca21]i
(B) and accumulation of IP1 (A) in SH-SY5Y cells. The data repre-
sent the mean ^SEM from 4 experiments. (b) Time course of IP1
accumulation in SH-SY5Y cells under the following conditions (i)
basal (A), (ii) in the presence of DPDPE (1 mM)(B), (iii) in the
presence of oxo-M (1 mM) (K), and (iv) in the presence of oxo-M
(1 mM) plus DPDPE (1 mM)(O). Each point represents the mean
^SEM of 3±7 experiments.
1694 A. Yeo et al.
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1688±1700

1996). Inhibitors of sphingosine kinase have been shown to
block m2 muscarinic receptor-mediated elevation of [Ca21]i
(Meyer zu Heringdorf et al. 1998). The sphingosine kinase
inhibitor, dl-threo-dihydrosphingosine (DHS, 100 mm), was
applied to SHSY5Y cells for 10 min prior to and during
agonist application. DHS did not change the amplitude of
the response to oxo-M (1 mm) and augmented, rather than
inhibited, the response to DPDPE (1 mm). After DHS
treatment the responses to oxo-M were 111 ^ 31% of con-
trol (n � 3, p . 0.05) whereas those to DPDPE were
198 ^ 44% of control (n � 3, p , 0.05). The inactive
analogue of DHS, N-acetylsphingosine (100 mm) which
does not inhibit sphingosine kinase at micromolar concen-
trations (Yatomi et al. 1996) had no effect on the responses
to oxo-M or DPDPE (data not shown).
We next sought to determine if the d-opioid receptor
exhibited cross-talk with a G protein-coupled receptor
whose activation gave rise to an elevation of [Ca21]i
independent of PLC activation and inositol phosphate
production. One such PLC-independent pathway is through
sphingosine kinase activation (Meyer zu Heringdorf et al.
1998). In SH-SY5Y cells the elevation of [Ca21]i by
lysophosphatidic acid (LPA) was independent of PLC and
was blocked by a sphingosine kinase inhibitor (Young et al.
1999). In monolayers of SH-SY5Y cells LPA (30 nm-
10 mm) evoked a transient increase in [Ca21]i that declined
to basal levels or just above basal levels (see Fig. 6a) in the
continued presence of LPA. In the continued presence of
LPA, coapplication of DPDPE did not evoke a further
elevation of [Ca21]i (Fig. 6a). Because the sustained plateau
elevation of [Ca21]i by LPA was absent in some experi-
ments and in the other experiments was much less than that
evoked by carbachol we also sought to determine if d-opioid
receptor activation could elevate the initial peak response to
LPA. When DPDPE (1 mm) was co-applied with a sub-
maximal concentration of LPA (1 mm) the amplitude of the
peak elevation of [Ca21]i was not different from that evoked
by LPA alone (Fig. 6b). For comparison, when DPDPE was
coapplied with carbachol (1 mm) the amplitude of the
elevation of [Ca21]i was signi®cantly elevated above that
evoked by carbachol alone (Fig. 6b). Therefore the d-opioid
receptor does not exhibit cross-talk with the sphingosine
kinase-coupled LPA receptor in SH-SY5Y cells.
Potassium channel activation
Opioid receptors activate several different types of potas-
sium conductance (Wimpey and Chavkin 1991; North 1993;
Vaughan et al. 1997). Although most of the elevation of
Fig. 5 Effect of UTP and DPDPE on [Ca21]i and IP1 production in
CHO-d cells. (a) The trace represents a continuous record of [Ca21]i
in a monolayer of CHO-d cells. DPDPE alone did not elevate [Ca21]i
whereas UTP produced a biphasic elevation of [Ca21]i. In the
presence of UTP, DPDPE now also elevated [Ca21]i. Drugs were
applied for the periods indicated by the bars. (b) Concentration±
response relationships for UTP-evoked elevation of [Ca21]i (open
squares) and accumulation of IP1 (closed squares) in CHO-d cells.
The data represent the mean ^SEM from 4 to 6 experiments. (c)
Time course of IP1 accumulation in CHO-d cells under the following
conditions: (i) basal (A), (ii) in the presence of DPDPE (1 mM) (B),
(iii) in the presence of UTP (10 mM) (K), and (iv) in the presence of
UTP (10 mM) plus DPDPE (1 mM) (O). Each point represents the
mean ^SEM of 3±7 experiments; * indicates a signi®cant difference
(p , 0.05) over UTP alone.
Co-incident signalling between d-opioid and m3 muscarinic receptors 1695
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1688±1700
[Ca21]i evoked by d-opioid receptor activation resulted from
Ca21 release from intracellular stores, part of the elevation
of [Ca21]i could be secondary to increased potassium con-
ductance and membrane hyperpolarization that would
increase the driving force for Ca21 entry through capacitative
Ca21 entry channels opened on store depletion during Gq-
coupled muscarinic receptor activation (Grudt et al. 1996).
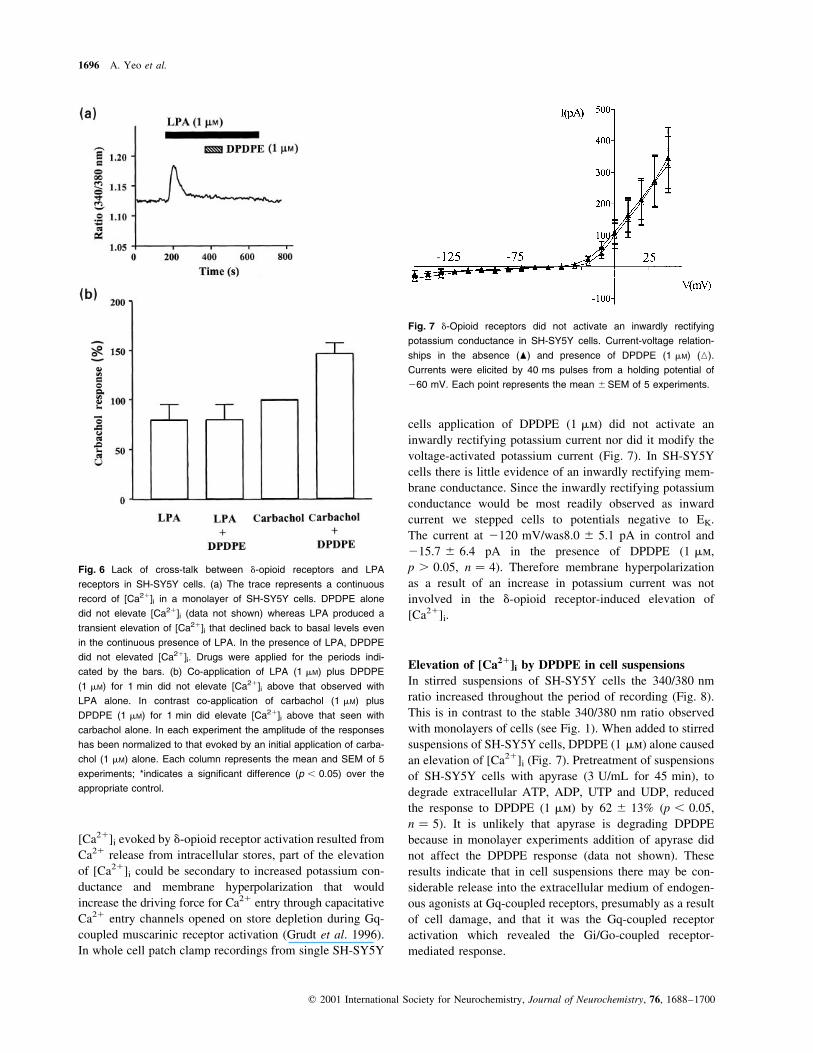
In whole cell patch clamp recordings from single SH-SY5Y
cells application of DPDPE (1 mm) did not activate an
inwardly rectifying potassium current nor did it modify the
voltage-activated potassium current (Fig. 7). In SH-SY5Y
cells there is little evidence of an inwardly rectifying mem-
brane conductance. Since the inwardly rectifying potassium
conductance would be most readily observed as inward
current we stepped cells to potentials negative to EK.
The current at 2120 mV/was8.0 ^ 5.1 pA in control and
215.7 ^ 6.4 pA in the presence of DPDPE (1 mm,
p . 0.05, n � 4). Therefore membrane hyperpolarization
as a result of an increase in potassium current was not
involved in the d-opioid receptor-induced elevation of
[Ca21]i.
Elevation of [Ca21]i by DPDPE in cell suspensions
In stirred suspensions of SH-SY5Y cells the 340/380 nm
ratio increased throughout the period of recording (Fig. 8).
This is in contrast to the stable 340/380 nm ratio observed
with monolayers of cells (see Fig. 1). When added to stirred
suspensions of SH-SY5Y cells, DPDPE (1 mm) alone caused
an elevation of [Ca21]i (Fig. 7). Pretreatment of suspensions
of SH-SY5Y cells with apyrase (3 U/mL for 45 min), to
degrade extracellular ATP, ADP, UTP and UDP, reduced
the response to DPDPE (1 mm) by 62 ^ 13% (p , 0.05,
n � 5). It is unlikely that apyrase is degrading DPDPE
because in monolayer experiments addition of apyrase did
not affect the DPDPE response (data not shown). These
results indicate that in cell suspensions there may be con-
siderable release into the extracellular medium of endogen-
ous agonists at Gq-coupled receptors, presumably as a result
of cell damage, and that it was the Gq-coupled receptor
activation which revealed the Gi/Go-coupled receptor-
mediated response.
Fig. 6 Lack of cross-talk between d-opioid receptors and LPA
receptors in SH-SY5Y cells. (a) The trace represents a continuous
record of [Ca21]i in a monolayer of SH-SY5Y cells. DPDPE alone
did not elevate [Ca21]i (data not shown) whereas LPA produced a
transient elevation of [Ca21]i that declined back to basal levels even
in the continuous presence of LPA. In the presence of LPA, DPDPE
did not elevated [Ca21]i. Drugs were applied for the periods indi-
cated by the bars. (b) Co-application of LPA (1 mM) plus DPDPE
(1 mM) for 1 min did not elevate [Ca21]i above that observed with
LPA alone. In contrast co-application of carbachol (1 mM) plus
DPDPE (1 mM) for 1 min did elevate [Ca21]i above that seen with
carbachol alone. In each experiment the amplitude of the responses
has been normalized to that evoked by an initial application of carba-
chol (1 mM) alone. Each column represents the mean and SEM of 5
experiments; *indicates a signi®cant difference (p , 0.05) over the
appropriate control.
Fig. 7 d-Opioid receptors did not activate an inwardly rectifying
potassium conductance in SH-SY5Y cells. Current-voltage relation-
ships in the absence (O) and presence of DPDPE (1 mM) (K).
Currents were elicited by 40 ms pulses from a holding potential of
260 mV. Each point represents the mean ^SEM of 5 experiments.
1696 A. Yeo et al.
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1688±1700

Discussion
In SH-SY5Y neuroblastoma cells, as in other neuronal and
non-neuronal cell types, activation of various Gi/Go-coupled
receptors can elevate [Ca21]i but only if there is concomi-
tant activation of a Gq-coupled receptor (Gerwins and
Fredholm 1992; Okajima and Kondo 1992; Okajima et al.
1993; Dickenson and Hill 1994; Connor and Henderson
1996; Connor et al. 1996, 1997a, 1997b; Biber et al. 1997;
Tomura et al. 1997; Selbie and Hill 1998; Toms and Roberts
1999; Cho et al. 2000). We have chosen in this study to
examine the interaction between the Gi/Go-coupled d-opioid
receptor and the m3 muscarinic receptor because in SH-
SY5Y cells, co-application of agonists at each of these
receptors evoked a robust, coincident elevation of [Ca21]i.
The coincident signalling required concomitant activation
of both receptor types because it was abolished when a
muscarinic receptor antagonist was applied simultaneously
with the d-opioid receptor agonist (Connor and Henderson
1996). The d-opioid receptor-mediated component was
abolished by pretreating the cells with pertussis toxin demon-
strating that activation of Gi/Go proteins was involved.
Furthermore, the coincidence was speci®c between Gi/Go-
and Gq-coupled receptors in that activation of the LPA
receptor did not reveal the d-opioid receptor-mediated
response. In SH-SY5Y cells LPA did not stimulate inositol
phosphate turnover but elevated [Ca21]i by stimulating a G
protein-coupled receptor that, by an as yet unknown mech-
anism, activated sphingosine kinase and thus elevated [Ca21]i
(Young et al. 1999). We can also rule out coupling of the Gi/
Go-coupled d-opioid receptor to sphingosine kinase to elevate
[Ca21]i in SH-SY5Y cells because the d-opioid receptor-
mediated response was not inhibited by a sphingosine kinase
inhibitor, indeed it was potentiated which may indicate that
part of the d-opioid receptor-elevation of [Ca21]i pathway is
under the negative control of sphingosine kinase.
bg dimers released from Gi/Go proteins on d-opioid
receptor activation play a key role in the elevation of [Ca21]i
because the elevation was abolished when free bg dimers
were sequestered by overexpression of the bg binding
domain of GRK2 (Koch et al. 1993). On the other hand the
m3 muscarinic, Gq-mediated elevation of [Ca21]i was
unaffected. Free bg dimers have been shown to directly
activate PLCb isozymes (Park et al. 1993; Smrcka and
Sternweiss 1993; Rhee and Bae 1997) but the isozymes
are not equally sensitive to stimulation by bg dimers. The
order of sensitivity of PLC isozymes to bg stimulation is
PLCb3 . PLCb2 . PLCb1. Free bg dimers from Gi/Go
were not suf®cient to stimulate PLC in SH-SY5Y cells
because activation of d-opioid receptors alone did not either
enhance inositol phosphate turnover or elevate [Ca21]i. This
may indicate that in SH-SY5Y cells PLCb1 is the pre-
dominant form of the enzyme expressed. We have detected
PLCb1, PLCb2 and PLCb3 in lysates of SH-SY5Y cells
using the Western blotting technique (Yeo and Henderson,
unpublished observations) but have been unable to quantify
their respective levels of expression. Elevation of [Ca21]i
was only observed with concomitant Gi/Go-and Gq-coupled
receptor activation. Therefore bg dimers from Gi/Go were
crucial to this coincident signalling but were not on their
own suf®cient to elevate [Ca21]i.
To give rise to coincident signalling bg dimers released
on Gi/Go-coupled receptor activation and aq dimers acti-
vated by Gq-coupled receptor activation may both bind to
PLC and thus produce a synergistic activation (Chan et al.
2000; Zhu and Birnbaumer 1996; Selbie et al. 1997; Tomura
et al. 1997; Selbie and Hill 1998). Furthermore, bg dimers
have been shown to decrease phosphorylation of PLCb by
PKC (Litosch 1997) which could delay the onset of negative
feedback by PKC and prolong PLCb activation. Given that
such synergy at the level of PLC should result in enhanced
inositol phosphate production as well as enhanced elevation
Fig. 8 Apyrase reduced the elevation of [Ca21]i by DPDPE alone in
stirred suspensions of SH-SY5Y or CHO-d cells. (a) The traces
represent continuous records of [Ca21]i in stirred suspensions of
SH-SY5Y cells. Addition of DPDPE (1 mM) evoked an elevation of
[Ca21]i. The response evoked by DPDPE was reduced when the cell
suspension had been treated with apyrase (3 U/mL). The traces are
representative of data obtained in 5 separate experiments. (b) The
traces represent continuous records of [Ca21]i in stirred suspensions
of CHO-d cells. Addition of DPDPE (1 mM) evoked an elevation of
[Ca21]i. The response evoked by DPDPE was reduced when the cell
suspension had been treated with apyrase (3 U/mL). The traces are
representative of data obtained in 3 separate experiments.
Co-incident signalling between d-opioid and m3 muscarinic receptors 1697
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1688±1700

of [Ca21]i this mechanism cannot explain our ®ndings in
SH-SY5Y cells where the elevation of [Ca21]i occurred in
the absence of enhanced inositol phosphate production. It
would, however, explain our results from CHO-d cells where
enhanced inositol phosphate production was observed.
Could we, for technical reasons, have missed d-opioid
receptor-mediated enhancement of inositol phosphate pro-
duction in SH-SY5Y cells? Dickenson and Hill (1998)
demonstrated for the Gi/Go-coupled 5-HT1B receptor in
CHO cells that elevation of [Ca21]i, but not inositol
phosphate production, could be observed at low levels of
receptor expression but that at higher levels of receptor
expression both an elevation of [Ca21]i and an increase in
inositol phosphate production could be observed. We feel,
however, that it was unlikely that we were missing an
enhancement of inositol phosphate production in SH-SY5Y
cells for the following reasons. First, we used concentrations
of DPDPE and oxo-M which produced similar peak
elevations of [Ca21]i and could measure enhanced inositol
phosphate production only in response to oxo-M but not
DPDPE. Second, the proportion of cells responding and the
amplitude of the elevations of [Ca21]i by DPDPE in SH-
SY5Y and CHO-d cells was similar but we could only
measure enhanced inositol phosphate production in CHO-d
cells not in SH-SY5Y cells. Given that bg and aq dimers
exert an additive stimulatory effect on PLCb3 but not
PLCb1 (Smrcka and Sternweiss 1993), an interaction at the
level of PLC to enhance inositol phosphate production
would not be observed in SH-SY5Y cells if the predominant
isozyme of PLC expressed in these cells was PLCb1.
Neuronal cells express higher amounts of Gi and Go than
Gq (Hepler and Gilman 1992) and excess bg dimers from
activated Gi/Go proteins could associate with free aq dimers
in their GDP bound state to replenish the supply of Gq. In
the continued presence of ongoing Gq-coupled receptor
activation, replenishment of the available Gq would result in
more rapid production of active aq dimers in their GTP
bound state and thus enhanced PLC activation (Quitterer and
Lohse 1999). However, this mechanism for Gi/Go-and Gq-
coupled receptor cross-talk should also be revealed as an
enhancement of inositol phosphate production and therefore
is unlikely to apply in SH-SY5Y cells.
The m3 muscarinic receptor desensitized on continuous
agonist exposure (Szekeres et al. 1998). We can discount the
possibility that Gi/Go-coupled receptor activation in some
way reversed the desensitization of the Gq-coupled m3
muscarinic receptor for two reasons. First, enhancement of
the muscarinic response through reduced desensitization
would again result in enhancement of inositol phosphate
production which we did not observe. Second, the amplitude
of the d-opioid receptor-mediated elevation in [Ca21]i was
equal in amplitude at various time points during the
muscarinic receptor-mediated response whereas desensitiza-
tion would have been expected to increase with time.
It has been suggested that activated G proteins may in
some way sensitize the IP3 receptor to IP3 (Xu et al. 1996;
Neylon et al. 1998) thus enhancing Ca21 release from
intracellular stores and that this effect was mediated by free
bg dimers (Zeng et al. 1996). With this mechanism the
cross-talk between Gi/Go-and Gq-coupled receptors would
occur at the level of the IP3 receptor, not at the level of PLC,
and would give rise to an elevation of [Ca21]i without an
enhancement of inositol phosphate production. The IP3
receptor is a tetrameric ligand-gated ion channel with four IP3
binding sites (Taylor et al. 1999). An interaction between bg
dimers and IP3 receptors could explain why the concentra-
tion±response curve for carbachol's permissive action to
reveal the d-opioid receptor response is shifted to the left.
There is some controversy surrounding the d-opioid
receptor-mediated elevation of [Ca21]i in the NG108-15
neuroblastoma � glioma hybrid cell line. We and colleagues
have demonstrated that, in that cell line, as in SH-SY5Y cells,
d-opioid receptor-mediated elevation of [Ca21]i required
concomitant Gq-coupled receptor activation (Okajima and
Kondo 1992; Okajima et al. 1993; Connor et al. 1994).
However, Thayer and colleagues have reported that in
NG108-15 cells d-opioid receptor activation alone was
suf®cient to increase [Ca21]i (Jin et al. 1994; Strassheim
et al. 1998; Yoon et al. 1999). It is possible that under
different culture conditions in different laboratories NG108-
15 cells may express different PLCb isoforms. If under their
culture conditions NG108-15 cells expressed PLCb3 then
on d-opioid receptor activation free bg dimers could directly
activate PLCb3, generate IP3 and release Ca21 from intra-
cellular stores.
In our single cell imaging experiments we observed a
rise in [Ca21]i in response to d-opioid receptor activation
in only 5% of cells, whereas 60% of cells responded to d-
opioid receptor activation during concomitant m3 muscarinic
receptor activation. Most neuronal cell lines are not entirely
homogeneous. We have evidence that SH-SY5Y cells are
somewhat heterogeneous with respect to their expression of
d-and m-opioid receptors. In single cell imaging experi-
ments, in the presence of concomitant m3 muscarinic
receptor activation, there were cells which responded to
d-opioid receptor activation, some to m-opioid receptor
activation but the majority responded to both (Yeo and
Henderson, unpublished observations). If a small population
of SH-SY5Y cells expressed predominantly PLCb3 then
these cells might exhibit a rise in [Ca21]i in response to
d-opioid receptor activation alone. In addition, we cannot
exclude the possibility that those cells responding to
d-opioid receptor activation alone were not being concomi-
tantly activated by endogenous Gq receptor agonists
released into the bathing medium from the cells themselves
or from neighbouring cells. Even though we used a fast
superfusion rate (4 mL/min) we cannot be sure that we had
rapid ¯uid exchange in the spaces between adjacent cells or
1698 A. Yeo et al.
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1688±1700

in the space between the cells and the coverslip on which
they were grown. Our observations on stirred cell suspen-
sions, where concomitant m3 muscarinic receptor activation
was not required to see a d-opioid receptor elevation of
[Ca21]i and apyrase markedly depressed the response to
d-opioid receptor activation alone, indicate that with this
method there was considerable release into the extracellular
medium of endogenous agonists at Gq-coupled receptors
presumably as a result of cell damage and that it was the
Gq-coupled receptor activation which revealed the Gi/Go-
coupled receptor-mediated response.
In neuronal and non-neuronal cells activation of Gi/Go-
coupled receptors has been reported to elevate [Ca21]i by
enhancing Ca21 entry into the cell through dihydropyridine-
sensitive, presumably l-type voltage-operated, Ca21 channels
(Jin et al. 1992; Eriksson et al. 1993; Tang et al. 1994; Smart
et al. 1995; Wandless et al. 1996). We have no experimental
evidence for any L channel component to the d-opioid
receptor-mediated elevation of [Ca21]i in SH-SY5Y cells. In
our hands the rise in [Ca21]i persisted in the absence of
extracellular Ca21 and was not reduced by nimodipine or
La31, agents which block l-type Ca21 channels (Connor
and Henderson 1996). Furthermore, m3 muscarinic receptor
activation and d-opioid receptor activation released Ca21
from the same thapsigargin-sensitive intracellular Ca21
store (Connor and Henderson 1996; this paper). Under our
culture conditions SH-SY5Y cells expressed few l-type
channels although, on prolonged depolarization, a rise in
[Ca21]i due to Ca21 entry through l-type channels could be
observed (Seward and Henderson 1990; Morton et al. 1992).
Another Ca21 entry pathway in SH-SY5Y cells is through
capacitative Ca21 entry pathways (Grudt et al. 1996).
d-opioid receptor activation did not however, modify this
Ca21 entry process (Connor et al. 1995).
In conclusion, the coincident signalling between d-opioid
and m3 muscarinic receptors in SH-SY5Y cells involves bg
subunits released from Gi/Go on d-opioid receptor activa-
tion. The site of interaction does not appear to be PLC but at
a step after PLC activation.
Acknowledgements
Single cell imaging experiments were carried out in the MRC
Cell Imaging Facility at the University of Bristol. We thank Dr
R. Lefkowitz for the gift of the plasmid containing the C
terminal region (amino acids 495±689) of GRK2, Dr L. Devi
for the gift of CHO-d cells, Prof. J. Tavare for advice and
facilities for plasmid preparation and Dr M. Connor for helpful
discussions.
References
Beaven M. A. (1996) Calcium signalling: sphigosine kinase versus
phospholipase C? Curr. Biol. 6, 798±801.
Biber K., Klotz K.-N., Berger M., Gebicke-Harter P. J. and van
Calker D. (1997) Adenosine A1 receptor-mediated activation of
phospholipase C in cultured astrocytes depends on the level of
receptor expression. J. Neurosci. 17, 4956±4964.
Chan J. S. C., Lee J. W. M., Ho M. K. C. and Wong Y. H. (2000)
Preactivation permits subsequent stimulation of phospholipase C
by Gi-coupled receptors. Mol. Pharmacol. 57, 700±708.
Cho K., Kemp N., Noel J., Aggleton J. P., Brown M. W. and Bashir Z. I.
(2000) A new form of long-term depression in the perirhinal
cortex. Nature Neurosci. 3, 150±156.
Connor M. A. and Henderson G. (1996) d- and m-opioid receptor
mobilization of intracellular calcium in SH-SY5Y human neuro-
blastoma cells. Br. J. Pharmacol. 117, 333±340.
Connor M. A., Planner A. and Henderson G. (1994) Delta opioid and
mu opioid receptor mobilisation of intracellular calcium in
neuroblastoma cells. Regul. Peptides 54, 65±66.
Connor M. A., Grudt T. J. and Henderson G. (1995) The interaction
between opioid and muscarinic receptors in the mobilization of
intracellular calcium in SH-SY5Y cells. Analgesia 1, 371±374.
Connor M. A., Yeo A. and Henderson G. (1996) The effect of
nociceptin on calcium channel current and intracellular calcium in
the SH-SY5Y human neuroblastoma cell line. Br. J. Pharmacol.
118, 205±207.
Connor M. A., Yeo A. and Henderson G. (1997a) Neuropeptide Y Y2
receptor and somatostatin sst2 receptor coupling to mobilization
of intracellular calcium in SH-SY5Y human neuroblastoma cells.
Br. J. Pharmacol. 120, 455±463.
Connor M. A., Keir M. J. and Henderson G. (1997b) d-opioid receptor
mobilization of intracellular calcium in SH-SY5Y cells: lack
of evidence for d-receptor subtypes. Neuropharmacology 36,
125±133.
Dickenson J. M. and Hill S. J. (1994) Interactions between adenosine
A1- and histamine H1-receptors. Int. J. Biochem. 26, 959±969.
Dickenson J. M. and Hill S. J. (1998) Human 5-HT1B receptor
stimulated inositol phospholipid hydrolysis in CHO cells: synergy
with Gq-coupled receptors. Eur. J. Pharmacol. 348, 279±285.
Eriksson P. S., Nilsson M., Wagberg M., Hansson E. and Ronnback L.
(1993) Kappa opioid receptors on astrocytes stimulate l-type
calcium channels. Neuroscience 54, 401±407.
Forsythe I. D., Lambert D. G., Nahorski S. R. and Linsdell P. (1992)
Elevation of cytosolic calcium by cholinoceptor agonists in SH-
SY5Y human neuroblastoma cells: estimation of the contribution
of voltage-dependent currents. Br. J. Pharmacol. 107, 207±214.
Gerwins P. and Fredholm B. B. (1992) Stimulation of adenosine A1
receptors and bradykinin receptors which act via different G
proteins, synergistically raises inositol 1,4,5-trisphophate and
intracellular free calcium in DDT MF-2 smooth muscle cells.
Proc. Natl Acad. Sci. USA 89, 7330±7334.
Grudt T. J., Usowicz M. M. and Henderson G. (1996) Calcium entry
following store depletion in SH-SY5Y neuroblastoma cells. Mol.
Brain Res. 36, 93±100.
Hepler J. R. and Gilman A. G. (1992) G proteins. Trends Biochem. Sci.
17, 383±387.
Inglese J., Luttrell L. M., Iniguez-Lluhi J. A., Touhara K., Koch W. J.
and Lefkowitz R. J. (1994) Functionally active targetting domain
of the b-adrenergic receptor kinase: an inhibitor of Gbg-mediated
stimulation of type II adenylyl cyclase. Proc. Natl Acad. Sci. USA
91, 3637±3641.
Jin W., Lee N. M., Loh H. H. and Thayer S. A. (1992) Dual excitatory
and inhibitory effects of opioids on intracellular calcium in
neuroblastoma x glioma hybrid NG108-15 cells. Mol. Pharmacol.
42, 1083±1089.
Jin W., Lee N. M., Loh H. H. and Thayer S. A. (1994) Opioids mobilize
calcium from inositol 1,4,5-trisphosphate-sensitive stores in
NG108-15 cells. J. Neurosci. 14, 1920±1929.
Koch W. J., Inglese J., Stone W. C. and Lefkowitz R. J. (1993) The
Co-incident signalling between d-opioid and m3 muscarinic receptors 1699
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1688±1700

binding site for the bg subunits of heteromeric G proteins on the
b-adrenergic receptor kinase. J. Biol. Chem. 268, 8256±8260.
Litosch I. (1997) G protein bg subunits antagonize protein kinase
C-dependent phosphorylation and inhibition of phospholipase
Cb1. Biochem. J. 326, 701±707.
Meyer zu Heringdorf D., Lass H., Alemany R., Laser K. T., Neumann
E., Zhang C., Schmidt M., Rauen U., Jakobs K. H. and van
Koppen C. J. (1998) Sphingosine kinase-mediated calcium signal-
ing by G protein coupled receptors. EMBO J. 17, 2830±2837.
Morton A. J., Hammond C., Mason W. T. and Henderson G. (1992)
Characterisation of the N- and l-type calcium channels in differ-
entiated SH-SY5Y neuroblastoma cells: calcium imaging and
single channel recording. Mol. Brain. Res. 13, 53±61.
Neer E. (1995) heteromeric G proteins: organizers of transmembrane
signals. Cell 80, 249±257.
Neylon C. B., Nickashin A., Tkachuck V. A. and Bobik A. (1998)
Heterotrimeric Gi protein is associated with the inositol 1,4,5-
trisphosphate receptor complex and modulates calcium in¯ux.
Cell Calcium 23, 281±289.
North R. A. (1993) Opioid actions on membrane ion channels, in
Opioids 1 (Herz A. ed. ), pp. 773±797. Springer Verlag, Berlin.
Okajima F. and Kondo Y. (1992) Synergism in cytosolic calcium
mobilisation between bradykinin and agonists for pertussis toxin-
sensitive G protein coupled receptors in NG108-15 cells. FEBS
Lett. 301, 223±226.
Okajima F., Tomura H. and Kondo Y. (1993) Enkephalin activates the
phospholipase C/calcium system through cross-talk between
opioid receptors and P2-purinergic or bradykinin receptors in
NG108-15 cells. Biochem. J. 290, 241±247.
Park D., Jhon D.-Y., Lee C.-W. and Rhee S. G. (1993) Activation of
phospholipase C isozymes by G protein bg subunits. J. Biol.
Chem. 268, 4573±4576.
Quitterer U. and Lohse M. J. (1999) Crosstalk between Gai- and Gaq-
coupled receptors is mediated by Gbg exchange. Proc. Natl Acad.
Sci. USA 96, 10626±10631.
Rhee S. G. and Bae Y. S. (1997) Regulation of phosphoinositide-speci®c
phospholipase C isozymes. J. Biol. Chem. 272, 15045±15048.
Selbie L. A. and Hill S. J. (1998) G protein-coupled receptor cross-talk:
the ®ne-tuning of multiple receptor-signalling pathways. Trends
Pharmacol. Sci. 19, 87±93.
Selbie L. A., Dickenson J. M. and Hill S. J. (1997) Role of G protein bg
subunits in the augmentation of P2Y2 (P2U) receptor-stimulated
responses by neuropeptide Y Y1 Gi/o-coupled receptors. Biochem.
J. 328, 153±158.
Seward E. P. and Henderson G. (1990) Characterization of two com-
ponents of the N-like, high threshold-activated calcium channel
current in differentiated SH-SY5Y cells. P.A. Eur. J. Physiol. 417,
223±230.
Smart D., Smith G. and Lambert D. G. (1995) m-opioids activate
phospholipase C in SH-SY5Y neuroblastoma cells via calcium
channel opening. Biochem. J. 305, 577±582.
Smrcka A. V. and Sternweiss P. C. (1993) Regulation of puri®ed
subtypes of phosphatidylinositol-speci®c phospholipase Cb by G
protein a and b subunits. J. Biol. Chem. 268, 9667±9674.
Strassheim D., Law P.-Y. and Loh H. H. (1998) Contribution of
phospholipase C-b3 phosphorylation to the rapid attenuation of
opioid-activated phosphoinositide response. Mol. Pharmacol. 53,
1047±1053.
Szekeres P. G., Koenig J. A. and Edwardson M. J. (1998) The
relationship between agonist intrinsic activity and the rate of
endocytosis of muscarinic receptors in a human neuroblastoma
cell line. Mol. Pharmacol. 53, 759±765.
Tang T., Kiang J. G. and Cox B. M. (1994) Opioids acting through
delta receptors elicit a transient increase in the intracellular free
calcium concentration in dorsal root ganglion neuroblastoma
hybrid ND8-47 cells. J. Pharmacol. Exp. Ther. 270, 40±46.
Taylor C. W., Genazzani A. A. and Morris S. A. (1999) Expression of
inositol trisphosphate receptors. Cell Calcium 26, 237±251.
Toms N. J. and Roberts P. J. (1999) Group 1 mGlu receptors elevate
calcium in rat cultured cortical type 2 astrocytes: synergy with
adenosine A1 receptors. Neuropharmacolgy 38, 1511±1517.
Toms N. J., Jane D. J., Tse H.-W. and Roberts P. J. (1995) Charac-
terization of metabotropic glutamate receptor-stimulated phos-
phoinositide hydrolysis in rat cultured cerebellar granule cells.
Br. J. Pharmacol. 116, 2824±2827.
Tomura H., Itoh H., Sho K., Sato K., Nagao M., Ui M., Kondo Y. and
Okajima F. (1997) bg Subunits of pertussis toxin-sensitive G
proteins mediate A1 adenosine receptor agonist-induced activa-
tion of phospholipase C in collaboration with thyrotropin. J. Biol.
Chem. 272, 23130±23137.
Vaughan C. W., Ingram S. L., Connor M. A. and Christie M. J. (1997)
How opioids inhibit GABA-mediated neurotransmission. Nature
390, 611±614.
Wandless A. L., Smart D. and Lambert D. G. (1996) Fentanyl increases
intracellular calcium concentrations in SH-SY5Y cells. Br. J.
Anaesth. 76, 461±463.
Wimpey T. L. and Chavkin C. (1991) Opioids activate both an inward
recti®er and a novel voltage-gated potassium conductance in the
hippocampal formation. Neuron 6, 281±289.
Xu X., Zeng W. and Muallem S. (1996) Regulation of the inositol
1,4,5-trisphosphate-activated calcium channel by activation of G
proteins. J. Biol. Chem. 271, 11737±11744.
Yatomi Y., Ruan F., Megdish T., Toyokuni T., Hakomori S. and
Igarashi Y. (1996) N,N-Dimethylsphingosine inhibition of sphin-
gosine kinase and sphingosine-1-phosphate activity in human
platelets. Biochemistry 35, 626±633.
Yeo A. and Henderson G. (1997a) Coincident signalling between
d-opioid and muscarinic receptors in SH-SY5Y cells. Br. J.
Pharmacol. 120, 125P.
Yeo A. and Henderson G. (1997b) Elevation of intracellular calcium by
opioids in single SH-SY5Y cells. Soc. Neurosci. 23, 78.14.
Yeo A., Kelly E. P., Roberts P. J. and Henderson G. (1996) Elevation of
intracellular calcium by carbachol and delta opioids in SH-SY5Y
cells: characterisation of the coincident signalling. Soc. Neurosci.
22, 247.5.
Yoon S. H., Lo T.-M., Loh H. H. and Thayer S. A. (1999) d-Opioid-
induced liberation of Gbg mobilizes calcium stores in NG108-15
cells. Mol. Pharmacol. 56, 902±908.
Young K. W., Challiss R. A. J., Nahorski S. R. and Mackrill J. J. (1999)
Lysophosphatidic acid-mediated Ca21 mobilization in human
SH-SY5Y neuroblastoma cells is independent of phosphoinositide
signalling, but dependent on sphingosine kinase activation.
Biochem. J. 343, 45±52.
Zeng W. K., Xu X. and Muallem S. (1996) Gbg transduces calcium
oscillations and Gq a sustained response during stimulation of
pancreatic acinar cells with calcium mobilizing agonists. J. Biol.
Chem. 271, 18520±18526.
Zhu X. and Birnbaumer L. (1996) G protein subunits and the lack of
stimulation of phospholipase C by Gs- and Gi-coupled receptors.
Proc. Natl Acad. Sci. USA 93, 2827±2831.
1700 A. Yeo et al.
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1688±1700




















