
Expression of G Protein-Coupled Receptor 30 in the Hamster Ovary: Differential Regulation by...
12
Expression of G Protein-Coupled Receptor 30 in the Hamster Ovary: Differential Regulation by Gonadotropins and Steroid Hormones Cheng Wang, Eric R. Prossnitz, and Shyamal K. Roy Departments of Obstetrics and Gynecology (C.W., S.K.R.) and Cellular and Integrative Physiology (S.K.R.), Durham Research Center, University of Nebraska Medical Center, Omaha, Nebraska 68198; and Department of Cell Biology and Physiology (E.R.P.), University of New Mexico Health Sciences Center, Albuquerque, New Mexico 87131 The nongenomic actions of estradiol-17 are mediated by transmembrane estrogen receptors. Recently, G protein-cou- pled receptor 30 (GPR30) has been suggested to be a trans- membrane estrogen receptor that can mediate rapid and tran- scription-independent estradiol-17 signaling in different cell types. However, the expression, regulation, or biological rel- evance of GPR30 in the ovary remains unknown. We examined the expression and hormonal regulation of GPR30 mRNA and protein in hamster ovarian cells during the estrous cycle and after hypophysectomy and hormone replacement. GPR30 pro- tein expression was high in the theca, appreciable in the gran- ulosa, but low in luteal cells. GPR30 protein levels in granu- losa and theca cells increased steadily with the development of preantral and antral follicles, respectively. GPR30 mRNA and protein levels increased significantly on diestrous (d 3 of the estrous cycle), but decreased on d 4 at 1600 h after the LH surge. GPR30 mRNA levels increased significantly after hy- pophysectomy. Although steroid treatment failed to alter ovarian GPR30 mRNA levels, either FSH or LH effectively reduced the levels. Interestingly, the decrease in GPR30 mRNA corresponded to a marked increase in the receptor protein levels. FSH treatment, either alone or together with LH, resulted in a marked increase in GPR30 immunostaining in granulosa cells. LH alone significantly increased immuno- staining in theca cells. These results suggest that GPR30 is expressed in the membrane of hamster granulosa and theca cells, and the expression is regulated by gonadotropins. The unique pattern of GPR30 expression suggests that gonado- tropin-regulated follicular cell functions may involve GPR30 activity. (Endocrinology 148: 4853– 4864, 2007) E STROGEN PLAYS AN important role in the mammalian ovary (1– 6) that includes the formation of gap junc- tions between granulosa cells and enhancement of FSH-stim- ulated gene expression in granulosa cells (7, 8). Immunohis- tochemical, biochemical, and genetic studies have demonstrated the presence of classical estrogen receptor (ESR), such as ESR1 (also known as ER) and ESR2 (also known as ER), and their role in regulating ovarian follicular development in many species (1, 2, 9 –14). Evidence has ac- cumulated to indicate that estrogen also affects reproductive target tissues via nongenomic mechanisms that do not in- volve the classic ESR (15–17). However, the identity of the membrane ESR (mESR) remains controversial. It has been suggested that the mESR is a variant of the classical ESR, such as ESR1 and ESR2 that mediate estrogen action in the target cells, such as endothelial, neuronal, and pituitary cells (18 – 21). On the other hand, evidence has also accumulated for the presence of novel G protein-coupled mESRs, which are un- related to nuclear ESR but mediate the nongenomic action of estrogen in many cell types (22–25). Recent studies have confirmed that the G protein-coupled receptor 30 (GPR30) functions as a novel transmembrane ESR in many cancer cell lines (26 –28). Thomas et al. (27) have demonstrated that in SKBR3 breast cancer cells, which lack nuclear ESR, GPR30 functions as a high-affinity, saturable, displaceable, and sin- gle-binding-site-specific receptor for estrogens. Further- more, progesterone-induced increases and RNA interfer- ence-induced decreases in GPR30 expression in SKBR3 cells coincide with altered estradiol-17 (E2) binding. Revankar et al. (28) have demonstrated that GPR30 specifically binds E2 and fluorescent E2 derivatives and initiates intracellular cal- cium mobilization and synthesis of phosphatidylinositol 3,4,5-trisphosphate. All these lines of evidence clearly estab- lish that GPR30 is an alternative form of mESR, which exists and functions independent of the nuclear ESR. Although the presence of classical ESRs in ovarian fol- licles has been documented (2), evidence for the presence of mESRs in follicular cells is lacking. Preliminary studies in our laboratory suggest that E2 can rapidly phosphor- ylate MAPK in hamster granulosa cells (Roy, S. K., un- published observation); therefore, a functional mESR may exist in follicular cells. Although the membrane versions of the ESR1 and ESR2 can mediate the E2 action in hamster follicular cells, the involvement of an independent mESR, such as GPR30, cannot be ruled out. Therefore, the objec- tives of the present studies were to determine whether hamster ovarian cells would express GPR30 mRNA and protein and whether gonadotropins and ovarian steroid hormones would regulate the expression as the first step toward understanding the role of GPR30 in follicular development. First Published Online July 19, 2007 Abbreviations: E2, Estradiol-17; ECL, enhanced chemilumines- cence; ESR, estrogen receptor; GPCR, G protein-coupled receptor; GPR30, G protein-coupled receptor 30; mESR, membrane ESR. Endocrinology is published monthly by The Endocrine Society (http:// www.endo-society.org), the foremost professional society serving the endocrine community. 0013-7227/07/$15.00/0 Endocrinology 148(10):4853– 4864 Printed in U.S.A. Copyright © 2007 by The Endocrine Society doi: 10.1210/en.2007-0727 4853
Transcript of Expression of G Protein-Coupled Receptor 30 in the Hamster Ovary: Differential Regulation by...

Expression of G Protein-Coupled Receptor 30 in theHamster Ovary: Differential Regulation byGonadotropins and Steroid Hormones
Cheng Wang, Eric R. Prossnitz, and Shyamal K. Roy
Departments of Obstetrics and Gynecology (C.W., S.K.R.) and Cellular and Integrative Physiology (S.K.R.), DurhamResearch Center, University of Nebraska Medical Center, Omaha, Nebraska 68198; and Department of Cell Biology andPhysiology (E.R.P.), University of New Mexico Health Sciences Center, Albuquerque, New Mexico 87131
The nongenomic actions of estradiol-17� are mediated bytransmembrane estrogen receptors. Recently, G protein-cou-pled receptor 30 (GPR30) has been suggested to be a trans-membrane estrogen receptor that can mediate rapid and tran-scription-independent estradiol-17� signaling in different celltypes. However, the expression, regulation, or biological rel-evance of GPR30 in the ovary remains unknown. We examinedthe expression and hormonal regulation of GPR30 mRNA andprotein in hamster ovarian cells during the estrous cycle andafter hypophysectomy and hormone replacement. GPR30 pro-tein expression was high in the theca, appreciable in the gran-ulosa, but low in luteal cells. GPR30 protein levels in granu-losa and theca cells increased steadily with the developmentof preantral and antral follicles, respectively. GPR30 mRNAand protein levels increased significantly on diestrous (d 3 of
the estrous cycle), but decreased on d 4 at 1600 h after the LHsurge. GPR30 mRNA levels increased significantly after hy-pophysectomy. Although steroid treatment failed to alterovarian GPR30 mRNA levels, either FSH or LH effectivelyreduced the levels. Interestingly, the decrease in GPR30mRNA corresponded to a marked increase in the receptorprotein levels. FSH treatment, either alone or together withLH, resulted in a marked increase in GPR30 immunostainingin granulosa cells. LH alone significantly increased immuno-staining in theca cells. These results suggest that GPR30 isexpressed in the membrane of hamster granulosa and thecacells, and the expression is regulated by gonadotropins. Theunique pattern of GPR30 expression suggests that gonado-tropin-regulated follicular cell functions may involve GPR30activity. (Endocrinology 148: 4853–4864, 2007)
ESTROGEN PLAYS AN important role in the mammalianovary (1–6) that includes the formation of gap junc-
tions between granulosa cells and enhancement of FSH-stim-ulated gene expression in granulosa cells (7, 8). Immunohis-tochemical, biochemical, and genetic studies havedemonstrated the presence of classical estrogen receptor(ESR), such as ESR1 (also known as ER�) and ESR2 (alsoknown as ER�), and their role in regulating ovarian folliculardevelopment in many species (1, 2, 9–14). Evidence has ac-cumulated to indicate that estrogen also affects reproductivetarget tissues via nongenomic mechanisms that do not in-volve the classic ESR (15–17). However, the identity of themembrane ESR (mESR) remains controversial. It has beensuggested that the mESR is a variant of the classical ESR, suchas ESR1 and ESR2 that mediate estrogen action in the targetcells, such as endothelial, neuronal, and pituitary cells (18–21). On the other hand, evidence has also accumulated for thepresence of novel G protein-coupled mESRs, which are un-related to nuclear ESR but mediate the nongenomic action ofestrogen in many cell types (22–25). Recent studies haveconfirmed that the G protein-coupled receptor 30 (GPR30)functions as a novel transmembrane ESR in many cancer cell
lines (26–28). Thomas et al. (27) have demonstrated that inSKBR3 breast cancer cells, which lack nuclear ESR, GPR30functions as a high-affinity, saturable, displaceable, and sin-gle-binding-site-specific receptor for estrogens. Further-more, progesterone-induced increases and RNA interfer-ence-induced decreases in GPR30 expression in SKBR3 cellscoincide with altered estradiol-17� (E2) binding. Revankar etal. (28) have demonstrated that GPR30 specifically binds E2and fluorescent E2 derivatives and initiates intracellular cal-cium mobilization and synthesis of phosphatidylinositol3,4,5-trisphosphate. All these lines of evidence clearly estab-lish that GPR30 is an alternative form of mESR, which existsand functions independent of the nuclear ESR.
Although the presence of classical ESRs in ovarian fol-licles has been documented (2), evidence for the presenceof mESRs in follicular cells is lacking. Preliminary studiesin our laboratory suggest that E2 can rapidly phosphor-ylate MAPK in hamster granulosa cells (Roy, S. K., un-published observation); therefore, a functional mESR mayexist in follicular cells. Although the membrane versionsof the ESR1 and ESR2 can mediate the E2 action in hamsterfollicular cells, the involvement of an independent mESR,such as GPR30, cannot be ruled out. Therefore, the objec-tives of the present studies were to determine whetherhamster ovarian cells would express GPR30 mRNA andprotein and whether gonadotropins and ovarian steroidhormones would regulate the expression as the first steptoward understanding the role of GPR30 in folliculardevelopment.
First Published Online July 19, 2007Abbreviations: E2, Estradiol-17�; ECL, enhanced chemilumines-
cence; ESR, estrogen receptor; GPCR, G protein-coupled receptor;GPR30, G protein-coupled receptor 30; mESR, membrane ESR.Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving theendocrine community.
0013-7227/07/$15.00/0 Endocrinology 148(10):4853–4864Printed in U.S.A. Copyright © 2007 by The Endocrine Society
doi: 10.1210/en.2007-0727
4853

Materials and MethodsChemicals
The antibody directed against a C-terminal GPR30 peptide was de-veloped and characterized as previously described (28). Ribogreen RNAquantification kit and Alexa-conjugated second antibodies were fromMolecular Probes, Inc. (Eugene, OR); AmpliScribe T7-Flash and T3-FlashTranscription Kits were from Epicenter (Madison, WI); peroxidase-con-jugated second antibodies for Western blotting were from Jackson Im-munoresearch Laboratories, Inc. (West Grove, PA); chemiluminescencedetection kit was from GE Healthcare (Piscataway, NJ); Optitran transfermembranes were from Schleicher & Schuell Bioscience (Dassel, Ger-many); PCR chemicals were from Roche Molecular Biochemicals (Indi-anapolis, IN), Amersham Pharmacia Biotech Boehringer (Piscataway,NJ), and Promega Corp. (Madison, WI); riboprobe synthesis kit was fromPromega Corp.; TOPO4 PCR cloning kit was from Invitrogen (Carlsbad,CA); and [32P]UTP (specific activity, 800 Ci/mmol) was from MP Bio-chemicals (Costa Mesa, CA). All other molecular-grade chemicals werepurchased from Sigma Chemical Co. (St. Louis, MO), Fisher ScientificCorp. (Pittsburgh, PA), or United States Biochemical (Cleveland, OH).Ovine-FSH-20 and ovine-LH-26 were purchased from the National Pi-tuitary Hormone Program [National Institute of Diabetes and Digestiveand Kidney Diseases, National Institutes of Health (NIH), Bethesda,MD] and provided by Dr. A. F. Parlow. E2-cipionate and progesteronewere from Pharmacia-Upjohn Co. (Kalamazoo, MI) and Steraloids (Wil-ton, NH), respectively.
Animals and treatments
Female golden hamsters (90–100 g; Charles River Laboratories, Inc.,Wilmington, MA) were housed in climate-controlled conditions with a14-h dark, 10-h light cycle and free access to food and water accordingto the U.S. Department of Agriculture and Institutional Animal Care andUse Committee (IACUC) guidelines. The use of hamsters in this studywas approved by the IACUC. Ovaries were obtained from hamsters withthree consecutive estrous cycles at 0900 h of each day of the estrous cycleand also at 1600 h on proestrous (d 4). There were three hamsters for eachday of the cycle, and the experiment was repeated twice. To obtainovaries from hypophysectomized hamsters, females were hypophysec-tomized on d 1 at 0900 h (estrous) as previously described (29, 30). Tendays after HX, hamsters were injected sc with 1) 0.5% BSA in saline(vehicle control), 2) 10 �g ovine-FSH-20 twice daily for 2 d, 3) 10 �govine-LH-26 twice daily for 2 d, or 4) a combination of FSH and LH in0.5% BSA in saline, and ovaries were collected 48 h after the first in-jection. The second group of hypophysectomized hamsters was injectedsc with a single dose of 1) 100 �g E2-cipionate, 2) 500 �g progesterone,or 3) a combination of E2 and progesterone, all in U.S. pharmacopoeia-grade sesame oil. Control animals received an equal volume of sesameoil vehicle. We never observed any estrogenic effect of sesame oil in thehamster (10). There were three hamsters per group, and the entireexperiment was repeated three times for reproducibility. Ovaries werecollected 24 h after the injection. Ovaries from all animals were eitherembedded in OCT Tissue-TEK medium for frozen sections or frozen inliquid N2 for RNA and protein extraction and stored at �80 C until use.
Cloning of hamster GPR30 cDNA
RT-PCR was used for obtaining a partial hamster GPR30 cDNA clone.cDNA was amplified from total ovarian RNA according to the protocoldescribed previously (31). The forward primer, 5�-ACATCCTCATCCT-GGTGGTGAAC-3�, and the reverse primer, 5�-TGGAATGACGGGC-CTTGAGCG-3�, corresponding to 230-1083 bases of mouse GPR30cDNA (accession no. NM_029771) were designed using Amplify (Dr. BillEngels, Genetics Department, University of Wisconsin) and Vector NTIprimer design software version 10 (Invitrogen), and synthesized in TheUniversity of Nebraska DNA synthesis core (Eppley Institute). The PCRwas done for 30 cycles with an annealing temperature of 55 C, and thecDNA was cloned into a PCR 4-TOPO plasmid vector, sequenced in theDNA sequencing core facility (University of Nebraska at Lincoln, Lin-coln, NE), and blast-searched in the NIH GenBank. After confirmation,hamster GPR30 sequence was compared with the rat, mouse, and humanGPR30 sequences using a Vector NTI gene analysis software version 10
[Molecular Analysis Core, University of Nebraska Medical Center(UNMC)].
Real-time RT-PCR quantification of GPR30 mRNA levels inthe hamster ovary
RNA was isolated from frozen ovaries using Trizol (Invitrogen) fol-lowed by the RNeasy mini kit (QIAGEN Inc., Valencia, CA) accordingto the manufacturers’ instructions and previously used protocols (10,31). The amount of RNA was quantified using the Ribogreen kit (Mo-lecular Probes) according to the manufacturer’s protocol. For real-timequantification, mRNA standards for GPR30 and actin were used inparallel with the sample RNA to obtain true quantitative values. PCRprimers and probes were designed using Primer Express software ver-sion 2.0 (Applied Biosystems, Inc., Foster City, CA) from hamster GPR30and actin nucleic acid sequences, which were partially cloned in thisstudy and previously (10), respectively. The forward and reverse prim-ers and probe for GPR30 were 5�-TTCCGCACCAAGCACCAT-3�, 5�-AGCCACTGCACCTCTCTGACA-3�, and 6-FAM-5�-CGTGCACCT-GCGGCACACTG-3�-BLACKHOLE, respectively, and for actin theywere 5�-TGACCGAGCGTGGCTACAG-3�, 5�-CTTCTCTTTGATGT-CACGCACAAT-3�, and 6-FAM-5�-TCACCACCACAGCCGAGAGGGA-3�-BLACKHOLE, respectively. Hamster GPR30 and actin mRNA weresynthesized using specific cDNA and the AmpliScribe T7-Flash transcrip-tion kit and quantified using Ribogreen, and the authenticity was verifiedby RT-PCR using the forward and reverse primers designed specifically forreal-time RT-PCR. A series of eight standards ranging from 0.1 fg to 1 ng(in 10-fold increments) of specific mRNA were reverse transcribed in du-plicates along with ovarian RNA samples and used for real-time quanti-fication of GPR30 and actin using the Opticon thermocycler (Bio-Rad,Hercules, CA). PCR amplification was carried out for 40 cycles after aninitial denaturation at 95 C for 15 min to activate the HotStart Taq poly-merase (QIAGEN). Each cycle of PCR consisted of 30 sec of denaturationat 94 C and 30 sec annealing at 55 C, followed by a plate reading with a finalextension at the end of 40 cycles for 10 min at 72 C. The authenticity of thePCR signal was verified using reactions without RNA or RNA without RT.The values were presented as femtograms of mRNA per microgram of totalRNA. The levels of actin mRNA for each sample were also presented toprove the specificity of GPR30 gene expression.
Northern hybridization detection of hamster GPR30 inthe ovary
Northern hybridization of hamster GPR30 mRNA was done essen-tially as described (31) to verify a critical finding of the real-time RT-PCR.The membrane was exposed to a storage phosphor screen and the signaldigitized in a Cyclone phosphorimager (PerkinElmer, Norwalk, CT).The approximate size of the GPR30 transcript was calculated from therelative positions of RNA size markers (Invitrogen).
Localization of GPR30 protein by fluorescenceimmunohistochemistry
Although the GPR30 antibody was thoroughly characterized usinghuman cell lines (28), the specificity of detection in hamster ovarian cellswas verified by neutralizing the antibody with 10-fold molar excess ofthe antigen peptide and using the neutralized antibody in immunoflu-orescence and Western blot detection of GPR30 protein. Additionally, asingle band of the expected molecular weight was observed by Westernblotting using this antibody.
GPR30 protein was localized in 6-�m-thick frozen sections fixed infreshly prepared ice-cold 4% paraformaldehyde in PBS (pH 7.4) asdescribed previously (10, 32) using 1:25,000 dilution of a rabbit poly-clonal anti-GPR30 antibody. The GPR30 signal was developed using adonkey antirabbit-IgG-Alexa 488, and nuclei were stained with 4�,6-diamidino-2-phenylindole. The images were captured with a Leica DMRmicroscope (North Central Instruments, Inc., Plymouth, MN) fitted witha Retiga EX1394 digital camera (QImaging Corp., Surrey, British Co-lumbia, Canada) and Openlab image analysis software (Improvision,Lexington, MA). The exposure time of the camera was set to subtractbackground fluorescence that was present in sections incubated with thenonimmune IgG of the host species. The 4�,6-diamidino-2-phenylindole
4854 Endocrinology, October 2007, 148(10):4853–4864 Wang et al. • Expression of GPR30 in the Ovary

signal was converted to red for contrast, and the green GPR30-specificfluorescence signal was merged with the nuclear signal to determine thecellular site of protein expression. GPR30 fluorescence intensity wasmeasured using the NIH ImageJ Image analysis software. Signal inten-sity (OD/pixel) was recorded from the granulosa and thecal compart-ments of five to 10 follicles per section from three animals, whereas theintensity for the interstitial cell compartment was recorded from threeovary sections from three animals. At least 10 areas of interest in eachcell compartment were digitized to obtain the average for the compart-ment, which was considered as one observation (n � 1). The totalnumber of follicles per section provided the mean for the granulosa andthecal cell compartments for that ovary. The mean and the sem repre-sented three ovaries (n � 3), and was calculated using the InStat sta-tistical software (GraphPad Software, San Diego, CA). A d-1 ovarysection was examined by a Zeiss confocal microscope (UNMC ImagingCore) at �630 for determining the cellular distribution of GPR30 protein.
Immunoblot detection of GPR30 protein in the ovary
The first experiment focused on determining the cellular sites ofGPR30 expression in the hamster ovary. Ovaries from hamsters indiestrous (d 3 at 0900 h) were homogenized in a 50 mm Tris-HCl buffer(pH 7.0) containing 150 mm NaCl, 1 mm EDTA, 1 mm EGTA, 1 mm NaF,20 mm Na-pyruvate, 2 mm Na-orthovanadate, 10% glycerol, and 10% ofa protease inhibitor cocktail (Sigma) on ice using a Dounce homogenizer.The homogenate was centrifuged at 1000 � g at 4 C, and the pellet wassonicated on ice in 30 �l epidermal growth factor receptor-lysis buffer(10 mm Tris-HCl, pH 7.4, 100 mm NaCl, 1 mm EDTA, 1 mm EGTA, 1 mmNaF, 20 mm Na4P2O7, 2 mm Na3VO4) containing 1% Triton X-100 andthe protease inhibitor cocktail and centrifuged at 26,000 � g for 20 minat 4 C. The supernatant was used as the crude nuclear fraction. The1000 � g supernatant was centrifuged at 100,000 � g for 1 h at 4 C, andthe supernatant was used as the cytosolic fraction, whereas the pelletwas sonicated in 50 �l epidermal growth factor receptor-lysis buffer withTriton X-100 and protease inhibitors on ice, kept on ice for 20 min, andcentrifuged at 26,000 � g at 4 C for 20 min, and the supernatant was usedas the crude membrane protein. The protein concentration was deter-mined by the micro-BCA kit (Pierce, Rockford, IL), and 5 �g membraneprotein and 20 �g cytosolic and nuclear protein were fractionated in a10% polyacrylamide gel, transferred to an Optitran nitrocellulose mem-brane, and probed with the same anti-GPR30 antibody that was used forimmunofluorescence detection or antibodies directed against Na�-K�-ATPase 2� (membrane marker), GATA4 (nuclear marker), or glyceral-dehyde-3-phosphate dehydrogenase (cytosol marker). Secondary anti-body binding was detected with the enhanced chemiluminescence(ECL) Advance Western blotting detection kit (GE Healthcare) andrecorded by a UVP gel documentation system (UVP, Upland, CA). Theexperiment was repeated three times to confirm reproducibility.
Based on the results of subcellular localization, crude membranepreparations of ovaries from cyclic hamsters and hypophysectomizedhamsters with or without hormonal replacement were used for GPR30protein expression determination by Western immunoblotting as de-scribed for experiment 1, except the intensity of the ECL signal wasquantified. Because a housekeeping membrane protein was difficult toselect, the proteins on the nitrocellulose membrane were stained withSyproRuby stain (Invitrogen) according to the manufacturer’s instruc-tion and digitized using an ethidium bromide filter and the UVP geldocumentation system. The OD corresponding to the total immunoflu-orescence signal in each lane was used for normalizing the ECL signalfor GPR30 protein.
To determine GPR30 expression during ovarian follicular develop-ment, follicles at primary (stage 1) through large antral (stage 10) wereisolated from diestrous (d 3, 0900 h) hamsters as described previously(33). The description of developmental stages is as follows: stages 1–4,follicles with one to four layers of granulosa cells without theca; stage5, follicles with five to six layers of granulosa cells with a few thecal cells;stage 6, follicles with seven to eight layers of granulosa cells and adeveloped theca; stage 7, follicles with incipient antral cavities; andstages 8–10, follicles with increasing antrum (33). Follicles were rup-tured to extract granulosa cells, and thecal shells (whenever present)were cleaned of granulosa cells manually as best as possible without theuse of any enzyme to avoid any damage to the receptor protein. Noattempt was made to collect the oocytes. Therefore, granulosa cell prep-
arations represented a pure population, whereas theca preparation con-tained a few attached granulosa cells. Cells were washed thoroughly toremove BSA, and the crude plasma membrane was prepared as de-scribed earlier. After measuring the protein concentration, 5 �g totalprotein was used for Western blot detection of GPR30 as describedearlier. Each group had three replicates representing three animals.
Results
Cloning of the hamster GPR30 cDNA and comparison ofthe cDNA and deduced amino acid sequences with itsmouse, rat, and human counterparts
To determine nucleic acid and protein informationabout hamster GPR30, an 854-bp GPR30 cDNA, which was231 bp shorter than the full-length rat or mouse GPR30cDNA (NM_029771, NM_133573) was cloned, and the se-quence was submitted to the GenBank (accession no.DQ237895). Comparison of the nucleic acid sequence re-vealed that hamster GPR30 cDNA was 94.7, 95.1, and 95%similar to the corresponding segments of rat, mouse, andhuman GPR30 cDNA, respectively (Fig. 1). Similarly, theamino acid sequence of hamster GPR30 was 99.3, 98.2, and98.1% similar to that of rat, mouse, and human, respec-tively (Fig. 2). The partial amino acid sequence of hamsterGPR30 protein contained six of the seven transmembrane-spanning domains (Transmembrane Predictor: http://www.ch.embnet.org/cgi-bim/TMPTED_for_parser). Fur-thermore, analysis of the amino acid sequence revealedthat it contained a conserved cystine residue in each of thefirst two extracellular loops (Cys130, Cys205 and 207) thatare believed to be necessary for the formation of intramo-lecular disulfide bridges to maintain the receptor struc-ture. The hamster GPR30 protein also contained a D-R-Ytriplet, an exceptionally conserved sequence among manyG protein-coupled receptors (GPCRs) and believed to playa role in signal transduction (34), located in the secondintracytoplasmic loop after the third transmembranedomain. NetPhos (http://www.cbs.dtu.dk/services/NetPhos/) analysis revealed the presence of four putativeserine and threonine and two tyrosine phosphorylationsites. Prosite (http://www.expasy.ch/prosite/) analysisindicated the presence of a putative N-glycosylation siteand four possible protein kinase C phosphorylation sites(Fig. 2) that matched closely with those in mouse GPR30protein. The presence of a Cys352 in the last hydrophilicregion might suggest a putative palmitoylation site thatanchors the carboxy terminus to the inside of the cellmembrane. Subcellular distribution analysis revealed thatGPR30 was expressed exclusively in the membrane frac-tion of the hamster ovary (Fig. 2).
GPR30 mRNA and protein expression in the hamster ovarythroughout the estrous cycle
The objectives were to determine whether GPR30 ex-pression in ovarian cells would alter with correspondingchanges in endogenous gonadotropin levels during theestrous cycle. GPR30 mRNA levels remained steady untild 2 (metestrous) followed by a significant increase on themorning of d 3 (diestrous) (Fig. 3A). GPR30 mRNA levelsdeclined to d 1 level by d 4 (proestrous) morning with afurther decline 2 h after the preovulatory gonadotropin
Wang et al. • Expression of GPR30 in the Ovary Endocrinology, October 2007, 148(10):4853–4864 4855

surge (Fig. 3A). No change in the levels of actin mRNA wasevident throughout the estrous cycle (Fig. 3A), suggestingthe specificity of GPR30 mRNA expression. The purposeof the Northern blot analysis was to verify the increase inGPR30 mRNA levels observed on d 3 morning by real-timeRT-PCR and also to gather information about GPR30 tran-scripts in the hamster ovary. Four transcripts were visiblein the blot (Fig. 3C) regardless of the cycle day. The ap-proximate size of the most prominent transcript was 1.4 kb(Fig. 3C). Furthermore, the levels of the 1.4-kb transcriptincreased 2-fold on d 3 compared with d 1 (Fig. 3, C andD), whereas the expression levels of other three transcriptsdid not change significantly (Fig. 3C). These results notonly confirmed the real-time data but also corroboratedthe protein data and implicated the 1.4-kb transcript to bebiologically relevant in ovarian functions. Based on theinformation, no attempt was made to repeat the Northernanalysis for all days of the estrous cycle.
Western blot analysis of ovarian protein revealed a 38-kDaband, and the expression levels closely followed the GPR30mRNA levels (Fig. 3E). However, in contrast to the mRNAlevels, GPR30 protein levels increased moderately but sig-
nificantly on d 2 morning, indicating the presence of addi-tional posttranscriptional regulation.
Based on the results of the Western immunoblotting, wewanted to examine whether there was any cell-type-spe-cific expression of GPR30 protein in the hamster ovary.Although immunofluorescence localization corroboratedthe immunoblot results, an estrous cycle-specific expres-sion of the receptor protein was evident (Fig. 4, A–F).Appreciable GPR30 immunosignal was present mainly inthe granulosa cells of preantral follicles on d1 at 0900 h, butmodest immunosignal was also present in the theca in-terna cells (Fig. 4, A and E). Confocal microscopic exam-ination confirmed the epifluorescence microscopic find-ings but also revealed that GPR30 immunostaining wasnot uniformly distributed in the granulosa cells. Further-more, punctate immunostaining was primarily intracel-lular (Fig. 4B). Low but discernible expression was evidentin cells of the newly formed corpora lutea (Fig. 4A), butinterstitial cells showed modest expression (Fig. 4, A andE). GPR30 expression increased significantly only in thethecal cells on d 2 at 0900 h (Fig. 4E); however, receptorexpression increased markedly in all cell types on d 3 at
HamsterHumanMouse
Rat
HamsterHumanMouseRat
HamsterHumanMouseRat
HamsterHumanMouseRat
HamsterHumanMouseRat
HamsterHumanMouseRat
HamsterHumanMouseRat
HamsterHumanMouseRat
Mouse
230 329ACATCCTCATCCTGGTGGTGAACATCAGCTTCCGGGAGAAGATGACAATCCCAGACCTGTACTTCATCAACCTGGCGGCGGCCGACCTCATCCTGGTGGCACATCCTGATCCTGGTGGTGAACATCAGCTTCCGCGAGAAGATGACCATCCCCGACCTGTACTTCATCAACCTGGCGGTGGCGGACCTCATCCTGGTGGCACATCCTCATCCTGGTGGTGAACATCAGCTTCCGGGAGAAGATGACCATCCCAGACCTGTACTTCATCAACCTGGCGGCGGCCGACCTCATCCTGGTGGCACATCCTCATCTTGGTGGTGAACATCAGCTTCCGGGAGAAGATGACTATCCCAGACCTGTACTTCATCAACCTGGCAGCGGCTGACCTCATCCTGGTGGC
330 429CGACTCCCTGATCGAGGTATTCAACCTGGATGAGCAGTACTACGACATTGCTGTGCTCTGCACCTTCATGTCCCTCTTCCTGCAGATCAACATGTACAGCCGACTCCCTCATTGAGGTGTTCAACCTGCACGAGCGGTACTACGACATCGCCGTCCTGTGCACCTTCATGTCGCTCTTCCTGCAGGTCAACATGTACAGCTGACTCCCTGATTGAGGTGTTCAACCTGGACGAGCAGTACTACGACATCGCAGTGCTCTGCACCTTCATGTCCCTCTTCCTGCAGATCAACATGTACAGCCGACTCCCTGATCGAGGTGTTCAACCTGGACGAGCAGTATTACGATATCGCCGTGCTCTGCACCTTCATGTCCCTCTTCCTGCAGATCAACATGTACAGC430 529AGTGTCTTCTTCCTCACCTGGATGAGCTTCGACAGGTACCTGGCGCTGGCCAAGGCAATGCGCTGCAGCCTCTTCCGCACCAAGCACCATGCCCGGCTCAAGCGTCTTCTTCCTCACCTGGATGAGCTTCGACCGCTACATCGCCCTGGCCAGGGCCATGCGCTGCAGCCTGTTCCGCACCAAGCACCACGCCCGGCTGAAGCGTCTTCTTCCTCACCTGGATGAGCTTCGACAGGTACCTAGCGCTGGCCAAGGCCATGCGCTGTGGCCTCTTCCGCACCAAGCACCACGCACGGCTCAAGCGTCTTCTTCCTCACCTGGATGAGCTTCGACAGGTACCTGGCGCTGGCCAAAGCCATGCGCTGTGGCCTCTTCCGCACCAAGCACCACGCGCGGCTCA
HamsterHuman
Rat
530 629GCTGTGGCCTCATCTGGATGGCCTCAGTGTCGGCCACGCTGGTACCCTTCACGGCCGTGCACCTGCGGCACACTGAAGAGGCCTGCTTTTGCTTTGCTGAGCTGTGGCCTCATCTGGATGGCATCCGTGTCAGCCACGCTGGTGCCCTTCACCGCCGTGCACCTGCAGCACACCGACGAGGCCTGCTTCTGTTTCGCGGAGCTGTGGCCTCATCTGGATGGCCTCAGTGTCCGCCACGCTGGTGCCCTTCACAGCGGTGCACCTGCGGCACACGGAGGAGGCCTGCTTCTGCTTTGCTGAGCTGTGGCCTCATCTGGATGGCCTCAGTGTCCGCCACGCTGGTGCCCTTCACGGCCGTGCATCTGCGGCACACCGAGGAGGCCTGCTTCTGCTTTGCCGA630 729TGTCAGAGAGGTGCAGTGGCTGGAGGTCACGCTGGGCTTCATTGTGCCCTTCGCCATCATTGGCCTCTGCTACTCCCCCATCGTGCGGGCCCTCATCCGGTGTCCGGGAGGTGCAGTGGCTCGAGGTCACGCTGGGCTTCATCGTGCCCTTCGCCATCATCGGCCTGTGCTACTCCCTCATTGTCCGGGTGCTGGTCAGGTGTCAGGGAGGTGCAGTGGCTGGAGGTCACACTGGGCTTCATCATGCCCTTCGCCATCATTGGCCTCTGCTACTCCCTCATCGTGCGAGCCCTCATCCGGTGTCAGGGAGGTGCAGTGGCTGGAGGTCACGCTGGGCTTCATTGTGCCCTTCGCCATCATCGGCCTGTGCTATTCCCTCATCGTGCGGGCCCTCATCCGG730 829GCCCACAGGCACCGTGGGCTGCGCCCGCGCAGGCAGAAAGCCCTGAGGATGATCTTTGCCGTGGTCCTCGTCTTCTTCATCTGCTGGCTGCCGGAGAATGGCGCACCGGCACCGTGGGCTGCGGCCCCGGCGGCAGAAGGCGCTCCGCATGATCCTCGCGGTGGTGCTGGTCTTCTTCGTCTGCTGGCTGCCGGAGAACGGCCCACAGGCACCGCGGCCTGCGCCCACGCAGGCAGAAAGCCCTGAGGATGATCTTCGCAGTGGTCCTTGTTTTCTTCATCTGCTGGCTGCCGGAGAACGGCCCACAGGCATCGTGGCCTGCGCCCACGCAGGCAGAAAGCCCTGAGGATGATCTTCGCAGTGGTCCTTGTCTTCTTCATCTGCTGGCTGCCGGAGAACG830 929TGTTCATCAGTGTCCATCTCCTGCAATGGACGCAGTCAGGGGACACTCCCTGCAAGCAGTCTTTCCGTCATGCTTACCCCTTGACAGGCCACATTGTCAATCTTCATCAGCGTGCACCTCCTGCAGCGGACGCAGCCTGGGGCCGCTCCCTGCAAGCAGTCTTTCCGCCATGCCCACCCCCTCACGGGCCACATTGTCAATCTTCATCAGTGTCCACCTACTGCAGTGGACGCAGCCAGGGGACACTCCCTGCAAGCAGTCTTTCCGTCACGCCTACCCCTTGACAGGCCACATAGTCAATCTTCATCAGCGTCCACCTACTGCAGTGGGCGCAGCCAGGGGACACTCCCTGCAAGCAGTCTTTCCGTCATGCCTACCCCTTGACAGGCCACATAGTCAA930 1029CCTTGCAGCCTTCTCCAACAGCTGCCTGAACCCCCTCATCTACAGCTTCCTTGGAGAGACCTTCAGGGACAAGCTCAGGCTGTACGTGGAGCAGAAGACGCCTCGCCGCCTTCTCCAACAGCTGCCTAAACCCCCTCATCTACAGCTTTCTCGGGGAGACCTTCAGGGACAAGCTGAGGCTGTACATTGAGCAGAAAACACCTTGCAGCCATCTCCAACAGCTGCCTGAATCCCCTCATCTACAGCTTCCTGGGAGAGACCTTCAGGGACAAGCTCAGGCTCTATGTGGAGCAGAAGACGCCTGGCAGCCTTCTCCAACAGCTGCCTGAGTCCCCTCATCTATAGCTTCCTGGGAGAGACCTTCAGGGACAAGCTCAGGCTGTATGTGGCGCAGAAGACG
1030 1083AGCCTGCCAGCTCTGAACCGCTTCTGCCACGCCACGCTCAAGGCCGTCATTCCAAATTTGCCGGCCCTGAACCGCTTCTGTCACGCTGCCCTGAAGGCCGTCATTCCAAGCCTGCCGGCTCTGAACCGCTTCTGCCATGCCACGCTCAAGGCCGTCATTCCAAGCCTGCCAGCTCTCAACCGCTTCTGCCATGCCACGCTCAAGGCAGTCATACCA
FIG. 1. Comparison of the partial cDNA sequence of the hamster GPR30 with the corresponding sequences of rat, mouse, and human GPR30.Differences in the nucleic acids are indicated in bold.
4856 Endocrinology, October 2007, 148(10):4853–4864 Wang et al. • Expression of GPR30 in the Ovary

0900 h with maximal expression in the theca (Fig. 4, C andE). Focal increased expression in the granulosa cell com-partment and considerable immunostaining in the oocyteswas also evident (Fig. 4C). GPR30 expression declinedconsiderably by the morning of d 4 primarily in the gran-ulosa and theca cells (Fig. 4E) and then further declined inall cell types by d 4 at 1600 h (Fig. 4, D and E). NoGPR30-specific immunostaining was observed when sec-
tions of a d-3 ovary were exposed to the antibody pre-neutralized with GPR30 peptide (Fig. 4F).
To determine the pattern of GPR30 protein expression indifferent follicular cell types during follicular developmentin vivo, proteins from follicles in each stage of developmenton d 3 at 0900 h were examined by immunoblotting. Whereasa modest level of GPR30 was detected in the granulosa cellsof primary follicles, the expression levels increased markedly
1 50Hamster .......... .......... .......... .......... ..........Human MDVTSQARGV GLEMYPGTAQ PAAPNTTSPE LNLSHPLLGT ALANGTGELSMouse MDATTPAQTV GVEIYLGPVW PAPSNSTPLA LNLSLALRED APGNLTGDLSRat MAATTPAQDV GVEIYLGPVW PAPSNSTPLA LNLSLALRED APGNLTGDLS
51 100Hamster .......... .......... .......ILI LVVNISFREK MTIPDLYFIN
Human EHQQYVIGLF LSCLYTIFLF PIGFVGNILI LVVNISFREK MTIPDLYFINMouse EHQQYVIALF LSCLYTIFLF PIGFVGNILI LVVNISFREK MTIPDLYFINRat EHQQYVIALF LSCLYTIFLF PIGFVGNILI LVVNISFREK MTIPDLYFIN
101 150Hamster LAAADLILVA DSLIEVFNLD EQYYDIAVLC TFMSLFLQIN MYSSVFFLTWHuman LAVADLILVA DSLIEVFNLH ERYYDIAVLC TFMSLFLQVN MYSSVFFLTWMouse LAAADLILVA DSLIEVFNLD EQYYDIAVLC TFMSLFLQIN MYSSVFFLTWRat LAAADLILVA DSLIEVFNLD EQYYDIAVLC TFMSLFLQIN MYSSVFFLTW
151 200Hamster MSFDRYLALA KAMRCSLFRT KHHARLSCGL IWMASVSATL VPFTAVHLRHHuman MSFDRYIALA RAMRCSLFRT KHHARLSCGL IWMASVSATL VPFTAVHLQHMouse MSFDRYLALA KAMRCGLFRT KHHARLSCGL IWMASVSATL VPFTAVHLRHRat MSFDRYLALA KAMRCGLFRT KHHARLSCGL IWMASVSATL VPFTAVHLRH
201 250Hamster TEEACFCFAD VREVQWLEVT LGFIVPFAII GLCYSPIVRA LIRAHRHRGLHuman TDEACFCFAD VREVQWLEVT LGFIVPFAII GLCYSLIVRV LVRAHRHRGLMouse TEEACFCFAD VREVQWLEVT LGFIMPFAII GLCYSLIVRA LIRAHRHRGLRat TEEACFCFAD VREVQWLEVT LGFIVPFAII GLCYSLIVRA LIRAHRHRGL
251 300
Hamster RPRRQKALRM IFAVVLVFFI CWLPENVFIS VHLLQWTQSG DTPCKQSFRHHuman RPRRQKALRM ILAVVLVFFV CWLPENVFIS VHLLQRTQPG AAPCKQSFRHMouse RPRRQKALRM IFAVVLVFFI CWLPENVFIS VHLLQWTQPG DTPCKQSFRHRat RPRRQKALRM IFAVVLVFFI CWLPENVFIS VHLLQWAQPG DTPCKQSFRH
301 350Hamster AYPLTGHIVN LAAFSNSCLN PLIYSFLGET FRDKLRLYVE QKTSLPALNR
Human AHPLTGHIVN LAAFSNSCLN PLIYSFLGET FRDKLRLYIE QKTNLPALNRMouse AYPLTGHIVN LAAISNSCLN PLIYSFLGET FRDKLRLYVE QKTSLPALNRRat AYPLTGHIVN LAAFSNSCLS PLIYSFLGET FRDKLRLYVA QKTSLPALNR
351 375Hamster FCHATLKAVI P......... .....
Human FCHAALKAVI PDSTEQSDVR FSSAVMouse FCHATLKAVI PDSTEQSEVR FSSAVRat FCHATLKAVI PDSTEQSDVK FSSAV
A
B
Na+-k+-ATPase-2α
GPR30
GAPDH
W C N M
GATA4
FIG. 2. A, Comparison of the deducedpartial amino acid sequence of ham-ster GPR30 with the correspondingsequence of that of rat, mouse, andhuman. Differences in the amino acidresidues are presented in bold, puta-tive serine phosphorylation sites arehighlighted with a long overline, thre-onine phosphorylation sites are high-lighted with a dashed underline, twotyrosine phosphorylation sites areboxed, protein kinase C phosphoryla-tion sites are highlighted with a shortoverline, and a glycosylation motif isin bold italics. Sequences noncompa-rable to that of the hamster GPR30are in italics. B, Subcellular distribu-tion of GPR30 in hamster ovariancells. C, Cytosol; M, crude membrane;N, nuclear; W, whole ovarian homog-enate.
Wang et al. • Expression of GPR30 in the Ovary Endocrinology, October 2007, 148(10):4853–4864 4857

in the granulosa cells of follicles at stages 2–10 (Fig. 5). GPR30protein levels in the granulosa cells dropped significantlywhen preantral follicles transited from stage 5 to stage 6 andwhen theca layer started to appear (33) and then increasedagain with the growth of the antral follicles (Fig. 5). Thecalcells had a relatively higher level of GPR30 protein expres-sion from the outset, and the levels increased steadily withthe growth of antral follicles (Fig. 5).
Effect of gonadotropins and steroid hormones on GPR30mRNA and protein expression in the hamster ovary
The objectives were to examine the selective effects ofgonadotropins or ovarian steroid hormones on ovarianGPR30 mRNA and protein expression and to determine
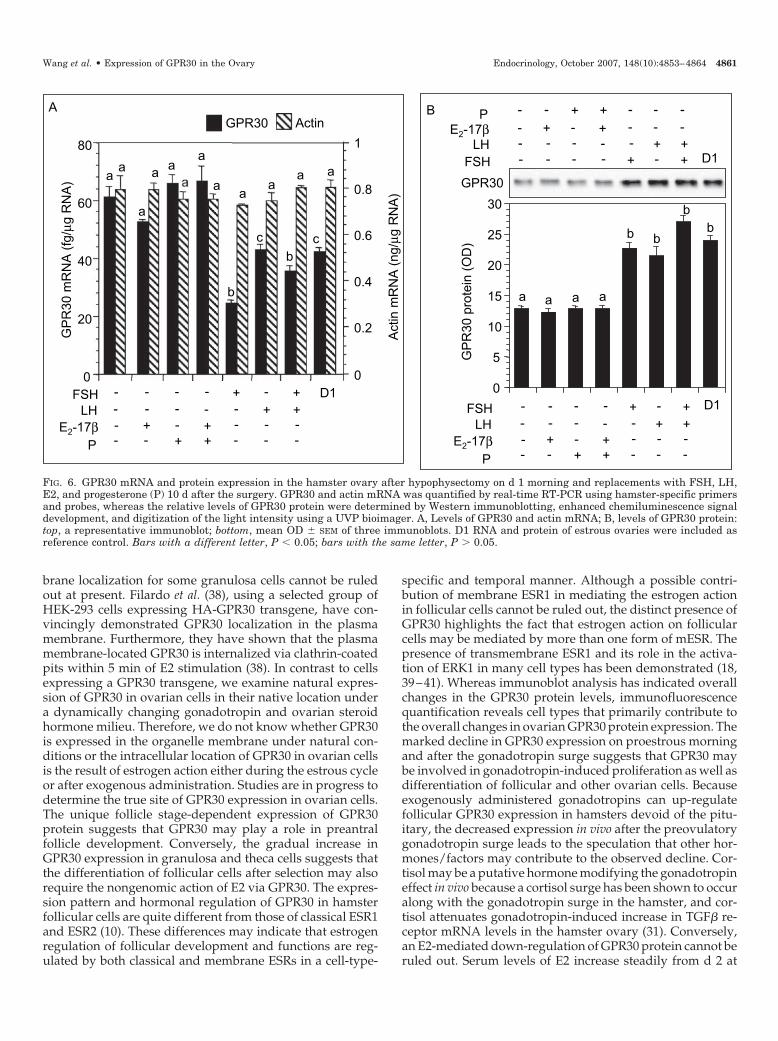
whether the effects of endogenous gonadotropins could bemimicked by exogenously administered hormones. Hy-pophysectomy resulted in a significant increase in thesteady-state levels of GPR30 mRNA compared with pituitaryintact hamsters on d 1 (Fig. 6A), which corresponded with adecrease in the levels of GPR30 protein (Fig. 6B). Neither themRNA nor the protein levels were affected by E2 or pro-gesterone treatment, either alone or in combination (Fig. 6, Aand B). In contrast, treatment with FSH, LH, or both resultedin a marked decrease in GPR30 mRNA levels (Fig. 6A) thatcorresponded with increases in the levels of GPR30 protein(Fig. 6B). Interestingly, FSH had the strongest effect onGPR30 mRNA levels that was somewhat blunted by LH (Fig.6B). No change in the levels of ovarian actin mRNA was
1-09h 2-09h 3-09h 4-09h 4-16h
Days and time of the Estrous cycle
aa
b b
c
1-09h 2-09h 3-09h 4-09h 4-16h
Days and time of the Estrous cycle
GP
R30
pro
tein
(OD
)
GPR30
0
10
20
30
40
50
60
E
D1 D3
1.4
7.5
2.4
4.4
kb
CB
D1 D3
1.4
7.5
2.4
4.4
kb
28S
18S
D1 D30
1
2
3
4G
PR
30 m
RN
A (
DLU
x 1
03 ) b
a
D
A
1-09h 2-09h 3-09h 4-09h 4-16hDays and time of the Estrous cycle
GP
R30
mR
NA
(fg/
µg R
NA)
01020304050607080
0
0.2
0.4
0.6
0.8
1
GPR30 Actin
Act
inm
RN
A (n
g/µg
RN
A)
FIG. 3. GPR30 mRNA and protein expression in the hamster ovary throughout the estrous cycle. A, Levels of GPR30 and actin mRNA in thehamster ovary on each day of the estrous cycle and after the preovulatory gonadotropin surge. B, Methylene blue-stained estrous (d 1) anddiestrous (d 3) ovarian RNA. RNA ladder shows the approximate size of the mRNA species. C, Northern hybridization of RNA depicted in Bshowing GPR30 mRNA transcripts. The most abundant transcript of approximately 1.4 kb has been indicated by an arrow. D, quantificationof the digital light unit (DLU) associated with the 1.4-kb GPR30 mRNA shown in C. E, Western immunoblotting analysis of the levels of GPR30protein in the hamster ovary during the estrous cycle: top, a representative immunoblot; bottom, mean OD � SEM of three immunoblots. Barswith a different letter, P � 0.05; bars with the same letter, P � 0.05.
4858 Endocrinology, October 2007, 148(10):4853–4864 Wang et al. • Expression of GPR30 in the Ovary

GC
Th
IC
GC
F
Th
A
GC
IC
CL
GCD
GC
Th
GC
IC
antrum
C
GCGC
IC Th
IC
O
1-09h 2-09h 3-09h 4-09h 4-16hDays and time of the Estrous cycle
GP
R30
IF (O
D/p
ixel
)
0
500
1000
1500E
GC
Th
B
a
c
b
a
ThecaGC
IC
aa
b
c
b
aa
a
b
b
a
FIG. 4. ImmunofluorescencelocalizationofGPR30proteininhamsterovarysections.GreenrepresentsGPR30,whereasredrepresentsthenucleus.A,Imagefrom an ovary section on d 1 at 0900 h; B, confocal image of a portion of a follicle on d 1 at 0900 h showing intracellular localization of GPR30 in granulosacells, with a few cells exhibiting possible plasma membrane staining (arrowheads); C, image from an ovary section on d 3 at 0900 h; D, image from an ovarysection on d 4 at 1600 h, 2 h after an ovary section on the preovulatory gonadotropin surge; E, intensity (OD/pixel) of the GPR30 immunosignal presentin the granulosa, theca, and interstitial cells; F, a d-3, 0900-h ovary section stained with GPR30 antibody that was preneutralized with the antigen peptide.Each bar represents a mean OD � SEM of three ovaries (see text for detail). Comparisons are for granulosa, theca, or interstitial cells across the cycle days:bars with a different letter, P � 0.05; bars with the same letter, P � 0.05. CL, Corpus luteum; GC, granulosa cells; IC, interstitial cells; Th, theca; O, oocyte.Bar, 10 �m.
Wang et al. • Expression of GPR30 in the Ovary Endocrinology, October 2007, 148(10):4853–4864 4859

evident (Fig. 6A), indicating the specificity of the hormonaleffect.
To determine the cell types contributing to the changes inGPR30 protein expression in hypophysectomized hamsters,GPR30 protein expression was also examined by immuno-fluorescence localization and quantification of the immu-nosignal. Hypophysectomy resulted in ovaries with smallpreantral follicles, which were associated with a modest levelof GPR30 expression in all three cell types (Fig. 7, A and E).Treatment with FSH alone resulted in the formation of antralfollicles with increased GPR30 expression exclusively in thegranulosa cells (Fig. 7, B and E). In contrast, whereas treat-ment with LH alone resulted in increased GPR30 expressiononly in the theca cells (Fig. 7, C and E), the level of expressionin the granulosa cells increased remarkably without anychange in the theca when LH was administered along withFSH (Fig. 7, D and E). It was also noteworthy that the in-creased expression was localized in the antral granulosacells.
Discussion
The results of the present study demonstrate for the firsttime the presence of a novel transmembrane ESR, GPR30, inhamster ovarian cells and provide evidence that reproduc-tive hormones affect GPR30 expression. The results also pro-vide convincing evidence that both granulosa and theca cellsexpress GPR30. Although the present study does not addressE2 and GPR30 interaction, accumulating evidence identifiesGPR30 as one of the receptors that are responsible for rapidestrogen-elicited responses in many cell lines (26–28, 35).
The deduced amino acid sequence from the cloned ham-ster GPR30 cDNA represents a significant portion of theGPR30 protein (from 78 to 361 of 375 total amino acids). Thissequence has almost all of the structural features of the GPCRsuperfamily. The conserved cystine residues in the extracel-lular loops have been suggested to provide structural integ-
rity by forming intramolecular disulfide bridges (34). TheD-R-Y triplet present in the hamster GPR30 protein is anexceptionally conserved sequence among many GPCRs andbelieved to play a role in signal transduction (34). The pres-ence of a Cys352 in the last hydrophilic region has beensuggested to be a putative palmitoylation site that anchorsthe carboxy terminus to the inside of the cell membrane.However, because of the length limitation of the presentsequence, we cannot detect N-glycosylation sites in the ex-tracellular N terminus of the receptor, a feature common tomany GPCRs (34). Although we cannot categorically identifythe transcript variant primarily responsible for generatingGPR30 protein in ovarian cells, the appreciably higher levelof expression of the 1.4-kb transcript compared with theother three variants and its increased expression on d 3corresponding to GPR30 protein suggest that the 1.4-kb tran-script may be the physiologically relevant form in the ovary.The reported size of the mouse and rat GPR30 mRNA is 2.5and 1.3 kb, respectively, and both encode an identical GPR30protein. Four GPR30 transcript variants have been identifiedin the human (36, 37); however, such information aboutrodent GPR30 is not available in the GenBank. HumanGPR30 transcript variants differ significantly at the 5�-un-translated region, primarily at the 5�-site, but are 100% iden-tical at the reading frame and 99% identical at the 3�-un-translated region. Furthermore, all four transcripts encode anidentical GPR30 protein.
Previous studies have indicated that GPR30 is expressedin the organelle membrane of cultured cells (28). Unlikecultured cells, granulosa cells within follicles present scantycytosolic compartment; hence, a clear distinction betweenintracellular and plasma membrane localization of GPR30presents a challenge. Nevertheless, the distinct punctate in-tracellular localization of GPR30 protein in granulosa andtheca cells in the confocal image suggests a possible associ-ation with the organelle membrane, although a plasma mem-
GPR
30 p
rote
in (O
D)
S1 S2 S3 S4 S5 S6 S7 S8 S9 S100
5
10
15
20
25
30
35
40
Granulosa cells
Stages of follicles
Theca cells
S1 S2 S3 S4 S5 S6 S7 S8 S9 S10
Stages of follicles
b
a
c
d d
cc
e
f f
a
b
a
c c
FIG. 5. Western immunoblotting offollicle stage-dependent expression ofGPR30 protein in theca (hatched bars)and granulosa (solid bars) cells. Folli-cles at different stages (S1–4 indicateone through four layers of granulosacells and no theca; S5 indicates five tosix layers of granulosa cells and a fewthecal cells; S6 indicates large prean-tral follicles with seven to eight layersof granulosa cells and well developedtheca; S7 indicates follicles with incip-ient antrum; and S8-S10 indicatesantral follicles with increasing size)of development were isolated fromdiestrous (d 3, 0900 h) ovaries as de-scribed in the text followed by separa-tion of the granulosa and theca cells.Top, A representative immunoblot ofthecal expression of GPR30; middle, arepresentative immunoblot of granu-losa cell expression of GPR30; bottom,mean OD � SEM of three immunoblotsof each cell compartment. Bars with adifferent letter, P � 0.05; bars with thesame letter, P � 0.05.
4860 Endocrinology, October 2007, 148(10):4853–4864 Wang et al. • Expression of GPR30 in the Ovary
brane localization for some granulosa cells cannot be ruledout at present. Filardo et al. (38), using a selected group ofHEK-293 cells expressing HA-GPR30 transgene, have con-vincingly demonstrated GPR30 localization in the plasmamembrane. Furthermore, they have shown that the plasmamembrane-located GPR30 is internalized via clathrin-coatedpits within 5 min of E2 stimulation (38). In contrast to cellsexpressing a GPR30 transgene, we examine natural expres-sion of GPR30 in ovarian cells in their native location undera dynamically changing gonadotropin and ovarian steroidhormone milieu. Therefore, we do not know whether GPR30is expressed in the organelle membrane under natural con-ditions or the intracellular location of GPR30 in ovarian cellsis the result of estrogen action either during the estrous cycleor after exogenous administration. Studies are in progress todetermine the true site of GPR30 expression in ovarian cells.The unique follicle stage-dependent expression of GPR30protein suggests that GPR30 may play a role in preantralfollicle development. Conversely, the gradual increase inGPR30 expression in granulosa and theca cells suggests thatthe differentiation of follicular cells after selection may alsorequire the nongenomic action of E2 via GPR30. The expres-sion pattern and hormonal regulation of GPR30 in hamsterfollicular cells are quite different from those of classical ESR1and ESR2 (10). These differences may indicate that estrogenregulation of follicular development and functions are reg-ulated by both classical and membrane ESRs in a cell-type-
specific and temporal manner. Although a possible contri-bution of membrane ESR1 in mediating the estrogen actionin follicular cells cannot be ruled out, the distinct presence ofGPR30 highlights the fact that estrogen action on follicularcells may be mediated by more than one form of mESR. Thepresence of transmembrane ESR1 and its role in the activa-tion of ERK1 in many cell types has been demonstrated (18,39–41). Whereas immunoblot analysis has indicated overallchanges in the GPR30 protein levels, immunofluorescencequantification reveals cell types that primarily contribute tothe overall changes in ovarian GPR30 protein expression. Themarked decline in GPR30 expression on proestrous morningand after the gonadotropin surge suggests that GPR30 maybe involved in gonadotropin-induced proliferation as well asdifferentiation of follicular and other ovarian cells. Becauseexogenously administered gonadotropins can up-regulatefollicular GPR30 expression in hamsters devoid of the pitu-itary, the decreased expression in vivo after the preovulatorygonadotropin surge leads to the speculation that other hor-mones/factors may contribute to the observed decline. Cor-tisol may be a putative hormone modifying the gonadotropineffect in vivo because a cortisol surge has been shown to occuralong with the gonadotropin surge in the hamster, and cor-tisol attenuates gonadotropin-induced increase in TGF� re-ceptor mRNA levels in the hamster ovary (31). Conversely,an E2-mediated down-regulation of GPR30 protein cannot beruled out. Serum levels of E2 increase steadily from d 2 at
B
0
5
10
15
20
25
30
b
a a a a
b b
b
GPR30
GP
R30
pro
tein
(OD
)
FSHLH
E2-17βP
D1- - - - + +-- - - - - ++- + - + - --- - + + - --
FSH D1- - - - + +-LH - - - - - ++
E2-17β - + - + - --P - - + + - --A
GP
R30
mR
NA
(fg/
µg R
NA)
FSHLH
E2-17βP
D1----
--+-
---+
--++
+---
++--
-+--
0
20
40
60
80
aa
b
a
cb
c
a
0
0.2
0.4
0.6
0.8
1
a aa a
aa a
Actin
mR
NA
(ng/
µgR
NA)
GPR30 Actin
FIG. 6. GPR30 mRNA and protein expression in the hamster ovary after hypophysectomy on d 1 morning and replacements with FSH, LH,E2, and progesterone (P) 10 d after the surgery. GPR30 and actin mRNA was quantified by real-time RT-PCR using hamster-specific primersand probes, whereas the relative levels of GPR30 protein were determined by Western immunoblotting, enhanced chemiluminescence signaldevelopment, and digitization of the light intensity using a UVP bioimager. A, Levels of GPR30 and actin mRNA; B, levels of GPR30 protein:top, a representative immunoblot; bottom, mean OD � SEM of three immunoblots. D1 RNA and protein of estrous ovaries were included asreference control. Bars with a different letter, P � 0.05; bars with the same letter, P � 0.05.
Wang et al. • Expression of GPR30 in the Ovary Endocrinology, October 2007, 148(10):4853–4864 4861

A
GC
Th
IC
C
GCTh
IC
GC
Th
IC
B
D
GC
Th
IC
Theca cells
Granulosa cells
Interstial cells
GPR
30 im
mun
oflu
ores
cenc
e (O
D/ p
ixel
)
0
200
400
600
FSHLH
--
+ +-- ++
a a a aa
aa
b
c
b
aa
E
FIG. 7. Immunofluorescence localization of GPR30 protein in hypophysectomized and hormone-replaced ovary sections. Green represents GPR30, whereasred represents the nucleus. A, Image of an ovary section from a vehicle-treated hamster 10 d after hypophysectomy; B, image of an ovary section from ahamster treated with 10 �g ovine-FSH-20 twice daily for 48 h; C, image of an ovary section from a hamster treated with 10 �g ovine-LH-25 twice daily for48 h; D, image of an ovary section from a hamster treated with FSH plus LH; E, intensity (OD/pixel) of the GPR30 immunostaining present in the granulosa,theca, and interstitial cells in ovaries represented in A–D. Each bar represents a mean OD � SEM of three ovaries (see text for detail). Comparisons are forgranulosa, theca, or interstitial cells across treatment groups and granulosa vs. theca cells within each treatment group: bars with a different letter, P �0.05; bars with the same letter, P � 0.05. Bar, 10 �m. GC, Granulosa cells; IC, interstitial cells; Th, theca.
4862 Endocrinology, October 2007, 148(10):4853–4864 Wang et al. • Expression of GPR30 in the Ovary

0900 h through d 4 at 1500 h followed by a decline (42). Theunique expression pattern of GPR30 mRNA and protein inresponse to steroid hormones or gonadotropins leads to thespeculation that a hormone-and cell-type-specific posttran-scriptional regulation may exist in hamster ovarian cells.Ovarian GPR30 mRNA and protein levels during the estrouscycle reflect the end effect of dynamically changing serumlevels of ovarian steroids and pituitary gonadotropins. Se-rum progesterone and FSH levels are high on d 1, and thelevels drop by d 2 through d 3, when serum levels of E2increase (42, 43). FSH causes significant growth of the gran-ulosa cell compartment that is increased further in the pres-ence of LH (44). LH acts primarily on the theca and cannotinduce preantral follicular growth by itself in hypophysec-tomized hamsters. Therefore, GPR30 expression in LH-treated hypophysectomized ovary remains only in the theca.Because thecal development corresponds to follicular devel-opment, steroid hormones alone in hypophysectomized con-dition do not cause significant thecal growth, which mayexplain low levels of GPR30 protein despite high levels ofreceptor mRNA expression. In contrast, progesterone hasbeen shown to increase GPR30 mRNA levels in MCF7 breastcancer cells (45, 46), and the increase coincides with a pro-portionate decrease in cell proliferation (45). Whether pro-gesterone plays a similar role in hypophysectomized ham-sters is not known at present. Conversely, it is equallypossible that GPR30 mRNA destabilization is directly pro-portional to protein translation, and steroid hormones andgonadotropins differentially influence the stability of GPR30mRNA in ovarian cells. It has been demonstrated for pro-teins, such as histone, c-Myc, etc. that a block in translationresults in mRNA stabilization (47). A 2- to 4-fold fluctuationin mRNA half-life is quite common in mammalian cells, andsuch changes may result in a greater than 1000-fold differ-ence in the steady-state levels of mRNA, which ultimatelydetermine the corresponding levels of protein (47). Further-more, estrogen has been shown to regulate mRNA stabilityin many cell types (47). Apparently, a translational blockexists when reproductive hormones are withdrawn by hy-pophysectomy, resulting in mRNA stabilization; however,steroid hormones alone cannot overcome the block. Gonad-otropins, on the other hand, can overcome the block, leadingto GPR30 mRNA destabilization. Because gonadotropintreatment results in follicular development and stimulationof ovarian steroidogenesis, it is possible that steroid hormoneregulation of ovarian GPR30 expression requires prior actionof gonadotropins. Such a mechanism will play an importantrole in the GPR30-mediated estrogen effect on follicular cellsthroughout the estrous cycle when marked changes in en-dogenous hormone levels are expected.
In summary, the results of the present study provide thefirst evidence that GPR30, a transmembrane ESR, is ex-pressed in the granulosa and theca cells of growing folliclesin the hamster ovary. Furthermore, the expression level ofGPR30 mRNA and protein is follicle cell type specific and isaffected by reproductive hormones. The results help us tospeculate that GPR30 may be involved, at least in part, inmediating estrogen action on follicular cells.
Acknowledgments
Received June 1, 2007. Accepted July 10, 2007.Address all correspondence and requests for reprints to: Shyamal K.
Roy, Departments of Obstetrics and Gynecology and Cellular and In-tegrative Physiology, DRC 5013, University of Nebraska Medical Center,984515 Nebraska Medical Center, Omaha, Nebraska 68198-4515. E-mail:[email protected].
This work was supported by grants (R01-HD38468) from the NationalInstitute of Child Health and Human Development, National Institutesof Health (NIH); the Leland and Dorothy Olson Foundation of theDepartment of Obstetrics and Gynecology, University of Nebraska Med-ical Center to S.K.R.; and a grant (R01-CA116662) from the NationalCancer Institute, NIH, to E.R.P. C.W. was a recipient of the Lalor Foun-dation postdoctoral fellowship.
Disclosure Statement: The authors have nothing to disclose.
References
1. Britt KL, Saunders PK, McPherson SJ, Misso ML, Simpson ER, Findlay JK2004 Estrogen actions on follicle formation and early follicle development. BiolReprod 71:1712–1723
2. Palter SF, Tavares AB, Hourvitz A, Veldhuis JD, Adashi EY 2001 Are estro-gens of import to primate/human ovarian folliculogenesis? Endocr Rev 22:389–424
3. Daniel SA, Armstrong DT 1983 Involvement of estrogens in the regulation ofgranulosa cell aromatase activity. Can J Physiol Pharmacol 61:507–511
4. Drummond AE, Findla JK 1999 The role of estrogen in folliculogenesis. MolCell Endocrinol 151:57–64
5. Rosenfeld CS, Roberts RM, Lubahn DB 2001 Estrogen receptor- and aro-matase-deficient mice provide insight into the roles of estrogen within theovary and uterus. Mol Reprod Dev 59:336–346
6. Britt KL, Findlay JK 2002 Estrogen actions in the ovary revisited. J Endocrinol175:269–276
7. Hsueh AJ, Billig H, Tsafriri A 1994 Ovarian follicle atresia: a hormonallycontrolled apoptotic process. Endocr Rev 15:707–724
8. Richards JS 1994 Hormonal control of gene expression in the ovary. EndocrRev 15:725–751
9. Couse JF, Lindzey J, Grandien K, Gustafsson JA, Korach KS 1997 Tissuedistribution and quantitative analysis of estrogen receptor-� (ER�) and estro-gen receptor-beta (ER�) messenger ribonucleic acid in the wild-type andER�-knockout mouse. Endocrinology 138:4613–4621
10. Yang P, Kriatchko A, Roy SK 2002 Expression of ER� and ER� in the hamsterovary: differential regulation by gonadotropins and ovarian steroid hormones.Endocrinology 143:2385–2398
11. Pepe GJ, Billiar RB, Leavitt MG, Zachos NC, Gustafsson JA, Albrecht ED2002 Expression of estrogen receptors � and � in the baboon fetal ovary. BiolReprod 66:1054–1060
12. Kuiper GG, Carlsson B, Grandien K, Enmark E, Haggblad J, Nilsson S,Gustafsson JA 1997 Comparison of the ligand binding specificity and tran-script tissue distribution of estrogen receptors � and �. Endocrinology 138:863–870
13. Zachos NC, Billiar RB, Albrecht ED, Pepe GJ 2004 Regulation of oocytemicrovilli development in the baboon fetal ovary by estrogen. Endocrinology145:959–966
14. Taniguchi F, Couse JF, Rodriguez KF, Emmen JM, Poirier D, Korach KS 2007Estrogen receptor-� mediates an intraovarian negative feedback loop on thecalcell steroidogenesis via modulation of Cyp17a1 (cytochrome P450, steroid17�-hydroxylase/17,20 lyase) expression. FASEB J 21:586–595
15. Ho KJ, Liao JK 2002 Nonnuclear actions of estrogen. Arterioscler Thromb VascBiol 22:1952–1961
16. Levin ER 1999 Cellular functions of the plasma membrane estrogen receptor.Trends Endocrinol Metab 10:374–377
17. Nadal A, Ropero AB, Fuentes E, Soria B 2001 The plasma membrane estrogenreceptor: nuclear or unclear? Trends Pharmacol Sci 22:597–599
18. Pedram A, Razandi M, Levin ER 2006 Nature of functional estrogen receptorsat the plasma membrane. Mol Endocrinol 20:1996–2009
19. Watson CS, Norfleet AM, Pappas TC, Gametchu B 1999 Rapid actions ofestrogens in GH3/B6 pituitary tumor cells via a plasma membrane version ofestrogen receptor-�? Steroids 64:5–13
20. Chen Z, Yuhanna IS, Galcheva-Gargova Z, Karas RH, Mendelsohn ME,Shaul PW 1999 Estrogen receptor � mediates the nongenomic activation ofendothelial nitric oxide synthase by estrogen. J Clin Invest 103:401–406
21. Monje P, Boland R 1999 Characterization of membrane estrogen bindingproteins from rabbit uterus. Mol Cell Endocrinol 147:75–84
22. Qiu J, Bosch MA, Tobias SC, Grandy DK, Scanlan TS, Ronnekleiv OK, KellyMJ 2003 Rapid signaling of estrogen in hypothalamic neurons involves a novelG-protein-coupled estrogen receptor that activates protein kinase C. J Neurosci23:9529–9540
Wang et al. • Expression of GPR30 in the Ovary Endocrinology, October 2007, 148(10):4853–4864 4863

23. Nadal A, Ropero AB, Laribi O, Maillet M, Fuentes E, Soria B 2000 Non-genomic actions of estrogens and xenoestrogens by binding at a plasma mem-brane receptor unrelated to estrogen receptor � and estrogen receptor �. ProcNatl Acad Sci USA 97:11603–11608
24. Sylvia VL, Boyan BD, Dean DD, Schwartz Z 2000 The membrane effects of17�-estradiol on chondrocyte phenotypic expression are mediated by activa-tion of protein kinase C through phospholipase C and G proteins. J SteroidBiochem Mol Biol 73:211–224
25. Filardo EJ, Quinn JA, Frackelton R, Bland KI 2002 Estrogen action via the Gprotein-coupled receptor, GPR30: stimulation of adenyl cyclase and cAMP-mediated attenuation of the epidermal growth factor receptor-to-MAPL sig-naling axis. Mol Endocrinol 16:70–84
26. Vivacqua A, Bonofiglio D, Recchia AG, Musti AM, Picard D, Ando S,Maggiolini M 2006 The G protein-coupled receptor GPR30 mediates the pro-liferative effects induced by 17�-estradiol and hydroxytamoxifen in endome-trial cancer cells. Mol Endocrinol 20:631–646
27. Thomas P, Pang Y, Filardo EJ, Dong J 2005 Identity of an estrogen membranereceptor coupled to a G protein in human breast cancer cells. Endocrinology146:624–632
28. Revankar CM, Cimino DF, Sklar LA, Arterburn JB, Prossnitz ER 2005 Atransmembrane intracellular estrogen receptor mediates rapid cell signaling.Science 307:1625–1630
29. Roy SK, Greenwald GS 1986 Effects of FSH and LH on incorporation of[3H]thymidine into follicular DNA. J Reprod Fertil 78:201–209
30. Roy SK, Kole AR 1995 Transforming growth factor-� receptor type II expres-sion in the hamster ovary: cellular site(s), biochemical properties, and hor-monal regulation. Endocrinology 136:4610–4620
31. Roy SK 2000 Regulation of transforming growth factor-�-receptor type I andtype II messenger ribonucleic acid expression in the hamster ovary by go-nadotropin and steroid hormone. Biol Reprod 62:1858–1865
32. Wang C, Roy SK 2006 Expression of growth differentiation factor 9 in theoocytes is essential for the development of primordial follicles in the hamsterovary. Endocrinology 147:1725–1734
33. Roy SK, Greenwald GS 1985 An enzymatic method for dissociation of intactfollicles from the hamster ovary: histological and quantitative aspects. BiolReprod 32:203–215
34. Strader CD, Fong TM, Tota MR, Underwood D, Dixon RA 1994 Structure andfunction of G protein-coupled receptors. Annu Rev Biochem 63:101–132
35. Filardo EJ, Thomas P 2005 GPR30: a seven-transmembrane-spanning estrogenreceptor that triggers EGF release. Trends Endocrinol Metab 16:362–367
36. Bologa CG, Revankar CM, Young SM, Edwards BS, Arterburn JB, KiselyovAS, Parker MA, Tkachenko SE, Savchuck NP, Sklar LA, Oprea TI, ProssnitzER 2006 Virtual and biomolecular screening converge on a selective agonist forGPR30. Nat Chem Biol 2:207–212
37. Carmeci C, Thompson DA, Ring HZ, Francke U, Weigel RJ 1997 Identifica-tion of a gene (GPR30) with homology to the G-protein-coupled receptorsuperfamily associated with estrogen receptor expression in breast cancer.Genomics 45:607–617
38. Filardo E, Quinn J, Pang Y, Graeber C, Shaw S, Dong J, Thomas P 2007Activation of the novel estrogen receptor G protein-coupled receptor 30(GPR30) at the plasma membrane. Endocrinology 148:3236–3245
39. Razandi M, Pedram A, Merchenthaler I, Greene GL, Levin ER 2004 Plasmamembrane estrogen receptors exist and functions as dimers. Mol Endocrinol18:2854–2865
40. Levin ER 2003 Bidirectional signaling between the estrogen receptor and theepidermal growth factor receptor. Mol Endocrinol 17:309–317
41. Evinger 3rd AJ, Levin ER 2005 Requirements for estrogen receptor � mem-brane localization and function. Steroids 70:361–363
42. Saidapur S, Greenwald GS 1978 Peripheral blood and ovarian levels of sexsteroids in the cyclic hamster. Biol Reprod 18:401–408
43. Bast JD, Greenwald GS 1974 Serum profiles of follicle-stimulating hormone,luteinizing hormone, and prolactin during the estrous cycle of the hamster.Endocrinology 94:1295–1299
44. Greenwald GS, Roy SK 1994 Follicular development and its control. In: KnobilE, Neill JD, eds. The physiology of reproduction. 2nd ed. New York: RavenPress; 629–724
45. Ahola TM, Purmonen S, Pennanen P, Zhuang YH, Tuohimaa P, Ylikomi T2002 Progestin upregulates G-protein-coupled receptor 30 in breast cancercells. Eur J Biochem 269:2485–2490
46. Ahola TM, Manninen T, Alkio N, Ylikomi T 2002 G protein-coupled receptor30 is critical for a progestin-induced growth inhibition in MCF-7 breast cancercells. Endocrinology 143:3376–3384
47. Ross J 1995 mRNA stability in mammalian cells. Microbiol Rev 59:423–450
Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving theendocrine community.
4864 Endocrinology, October 2007, 148(10):4853–4864 Wang et al. • Expression of GPR30 in the Ovary




















