
Gi-coupled P2Y-ADP receptor mediates GSK-3 phosphorylation and β-catenin nuclear translocation in...
12
Gi-coupled P2Y-ADP receptor mediates GSK-3 phosphorylation and b-catenin nuclear translocation in granule neurons Felipe Ortega, Raquel Pe ´rez-Sen and MªTeresa Miras-Portugal Department of Biochemistry, Veterinary Faculty, Complutense University of Madrid, Madrid, Spain Glycogen synthase kinase-3 (GSK-3) is a versatile signaling molecule that regulates development and cell fate at the CNS (Grimes and Jope 2001; Jope and Johnson 2004). The function of GSK-3 as a modulator of synaptic remodeling is based on its action on cytoskeletal re-arrangement (Hall et al. 2002; Sayas et al. 2002). In addition, it is well established that GSK-3 is involved in the survival signaling pathway of growth factors and neurotrophins in many neuronal cells (Pap and Cooper 1998; Hetman et al. 2000; Cross et al. 2001). In this respect, the impairment of GSK-3 regulation has been related to neurodegeneration (A ´ lvarez et al. 1999; Lucas et al. 2001; Bhat et al. 2004; Engel et al. 2006). Moreover GSK-3 actions can be even more diverse consid- ering it can modulate the expression of a large number of transcription factors (Grimes and Jope 2001). The best-known inputs targeting GSK-3 are growth and developmental factors. It is well characterized that insulin and other growth factors are coupled to GSK-3 catalytic activity inhibition by involving its phosphorylation at residues Ser21/9 (for a and b isoforms, respectively), through a phosphatidyl inositol-3 kinase (PI3-K)/Akt-depen- dent pathway. In a different way, Wnt developmental factors inhibit GSK-3 activity through its dissociation from a multiprotein complex. This inactivation abolished GSK-3 phosphorylation of one of its better known substrates, b-catenin, which triggers its degradation by the proteasome pathway. The lack of inhibition over b-catenin stabilizes it in the cytosol and allows its translocation to the nucleus, where it interacts with the lymphoid enhancer factor/T-cell factor Received May 8, 2007; revised manuscript received July 13, 2007; accepted August 7, 2007. Address correspondence and reprint requests to MªTeresa Miras- Portugal, Chairman of the Biochemistry Department, Veterinary Faculty, Complutense University of Madrid, Madrid, Spain. E-mail: [email protected] Abbreviations used: 2MeSADP, 2-methyl-thio-ADP; 2MeSAMP, 2- methyl-thio-AMP; BSA, bovine serum albumin; DIV, days in vitro; ERK, extracellular signal-regulated kinase; GSK-3, glycogen synthase kinase-3; IGF-I, insulin-like growth factor-I; Lef/Tcf, lymphoid enhancer factor/T-cell factor; MRS-2179, 2¢-deoxy-N 6 -methyl-adenosine 3¢,5¢- diphosphate; PBS, phosphate-buffered saline; PI3-K, phosphatidyl ino- sitol-3 kinase; PTX, pertussis toxin; SDS, sodium dodecyl sulfate. Abstract Glycogen synthase kinase-3 (GSK-3) is a multifaceted en- zyme involved in development, neurogenesis, and survival at the CNS. We investigated nucleotides signaling to GSK-3 in cerebellar granule neurons and found that the metabotropic agonist 2-methyl-thio-ADP (2MeSADP) was able to induce GSK-3 phosphorylation and inhibition of its catalytic activity. 2MeSADP could be acting through several P2Y-ADP recep- tors expressed in granule neurons, as RT-PCR expression was found for P2Y 1 , P2Y 12 , and P2Y 13 receptors, but the pharmacological data fitted well with a Gi-coupled P2Y 13 receptor: the effect was sensitive to pertussis toxin, was unaffected by specific antagonists of P2Y 1 and P2Y 12 recep- tors, such as 2¢-deoxy-N 6 -methyl-adenosine 3¢,5¢-diphosphate and 2-methyl-thio-AMP, respectively, and the EC 50 values for 2MeSADP and ADP were in the same low nanomolar range. 2MeSADP was able to phosphorylate and activate extracel- lular signal-regulated kinase (ERK)-1,2 and Akt proteins, but its effect on GSK-3 phosphorylation was primarily dependent on the phosphatidyl inositol-3 kinase (PI3-K)/Akt pathway, as it was abolished by the PI3-K inhibitor wortmannin. GSK-3 inactivation by 2MeSADP in granule neurons resulted in nu- clear translocation of its substrate b-catenin, which functions as a transcriptional regulator, this effect being lost with wortmaninn. The present study first describes the coupling of a Gi-coupled P2Y 13 -like receptor to GSK-3 and b-catenin through PI3-K/Akt signaling. Keywords: 2-methyl-thio-ADP, glycogen synthase kinase-3, granule neurons, insulin-like growth factor-I, nucleotide receptors, phosphatidyl inositol-3 kinase/Akt, b-catenin. J. Neurochem. (2008) 104, 62–73. d JOURNAL OF NEUROCHEMISTRY | 2008 | 104 | 62–73 doi: 10.1111/j.1471-4159.2007.05021.x 62 Journal Compilation ȑ 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 62–73 ȑ 2007 The Authors
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Gi-coupled P2Y-ADP receptor mediates GSK-3 phosphorylation and β-catenin nuclear translocation in...

Gi-coupled P2Y-ADP receptor mediates GSK-3 phosphorylationand b-catenin nuclear translocation in granule neurons
Felipe Ortega, Raquel Perez-Sen and MªTeresa Miras-Portugal
Department of Biochemistry, Veterinary Faculty, Complutense University of Madrid, Madrid, Spain
Glycogen synthase kinase-3 (GSK-3) is a versatile signalingmolecule that regulates development and cell fate at the CNS(Grimes and Jope 2001; Jope and Johnson 2004). Thefunction of GSK-3 as a modulator of synaptic remodeling isbased on its action on cytoskeletal re-arrangement (Hall et al.2002; Sayas et al. 2002). In addition, it is well establishedthat GSK-3 is involved in the survival signaling pathway ofgrowth factors and neurotrophins in many neuronal cells(Pap and Cooper 1998; Hetman et al. 2000; Cross et al.2001). In this respect, the impairment of GSK-3 regulationhas been related to neurodegeneration (Alvarez et al. 1999;Lucas et al. 2001; Bhat et al. 2004; Engel et al. 2006).Moreover GSK-3 actions can be even more diverse consid-ering it can modulate the expression of a large number oftranscription factors (Grimes and Jope 2001).
The best-known inputs targeting GSK-3 are growth anddevelopmental factors. It is well characterized that insulinand other growth factors are coupled to GSK-3 catalyticactivity inhibition by involving its phosphorylation atresidues Ser21/9 (for a and b isoforms, respectively),through a phosphatidyl inositol-3 kinase (PI3-K)/Akt-depen-
dent pathway. In a different way, Wnt developmental factorsinhibit GSK-3 activity through its dissociation from amultiprotein complex. This inactivation abolished GSK-3phosphorylation of one of its better known substrates,b-catenin, which triggers its degradation by the proteasomepathway. The lack of inhibition over b-catenin stabilizes it inthe cytosol and allows its translocation to the nucleus, whereit interacts with the lymphoid enhancer factor/T-cell factor
Received May 8, 2007; revised manuscript received July 13, 2007;accepted August 7, 2007.Address correspondence and reprint requests to MªTeresa Miras-
Portugal, Chairman of the Biochemistry Department, Veterinary Faculty,Complutense University of Madrid, Madrid, Spain.E-mail: [email protected] used: 2MeSADP, 2-methyl-thio-ADP; 2MeSAMP, 2-
methyl-thio-AMP; BSA, bovine serum albumin; DIV, days in vitro;ERK, extracellular signal-regulated kinase; GSK-3, glycogen synthasekinase-3; IGF-I, insulin-like growth factor-I; Lef/Tcf, lymphoid enhancerfactor/T-cell factor; MRS-2179, 2¢-deoxy-N6-methyl-adenosine 3¢,5¢-diphosphate; PBS, phosphate-buffered saline; PI3-K, phosphatidyl ino-sitol-3 kinase; PTX, pertussis toxin; SDS, sodium dodecyl sulfate.
Abstract
Glycogen synthase kinase-3 (GSK-3) is a multifaceted en-
zyme involved in development, neurogenesis, and survival at
the CNS. We investigated nucleotides signaling to GSK-3 in
cerebellar granule neurons and found that the metabotropic
agonist 2-methyl-thio-ADP (2MeSADP) was able to induce
GSK-3 phosphorylation and inhibition of its catalytic activity.
2MeSADP could be acting through several P2Y-ADP recep-
tors expressed in granule neurons, as RT-PCR expression
was found for P2Y1, P2Y12, and P2Y13 receptors, but the
pharmacological data fitted well with a Gi-coupled P2Y13
receptor: the effect was sensitive to pertussis toxin, was
unaffected by specific antagonists of P2Y1 and P2Y12 recep-
tors, such as 2¢-deoxy-N6-methyl-adenosine 3¢,5¢-diphosphate
and 2-methyl-thio-AMP, respectively, and the EC50 values for
2MeSADP and ADP were in the same low nanomolar range.
2MeSADP was able to phosphorylate and activate extracel-
lular signal-regulated kinase (ERK)-1,2 and Akt proteins, but
its effect on GSK-3 phosphorylation was primarily dependent
on the phosphatidyl inositol-3 kinase (PI3-K)/Akt pathway, as
it was abolished by the PI3-K inhibitor wortmannin. GSK-3
inactivation by 2MeSADP in granule neurons resulted in nu-
clear translocation of its substrate b-catenin, which functions
as a transcriptional regulator, this effect being lost with
wortmaninn. The present study first describes the coupling of
a Gi-coupled P2Y13-like receptor to GSK-3 and b-catenin
through PI3-K/Akt signaling.
Keywords: 2-methyl-thio-ADP, glycogen synthase kinase-3,
granule neurons, insulin-like growth factor-I, nucleotide
receptors, phosphatidyl inositol-3 kinase/Akt, b-catenin.
J. Neurochem. (2008) 104, 62–73.
d JOURNAL OF NEUROCHEMISTRY | 2008 | 104 | 62–73 doi: 10.1111/j.1471-4159.2007.05021.x
62 Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 62–73� 2007 The Authors

(Lef/Tcf) family of DNA-binding proteins and activatetranscription of genes containing Lef/Tcf binding sites (Dinget al. 2000; Domınguez and Green 2001; Grimes and Jope2001).
We raise the question whether this enzyme could also bemodulated by nucleotides, as they act as relevant signals atthe CNS. Nucleotides exhibit neuroprotective actionsbecause of the modulation of neurotransmitter release, anddisplay survival responses after brain injury or in neurode-generative disorders (Burnstock 2005, 2006; Neary andKang 2005). They cooperate with growth factors to triggerproliferation and differentiation in glial cells (Neary et al.1999) and mediate specific actions, such as inflammatoryresponses (DiVirgilio 2005; Kucher and Neary 2005), painsignaling and transmission regulation (Gerevich and Illes2004; Inoue et al. 2005) and the neural and endothelialcontrol of vascular tone (Erlinge 1998). The versatility ofnucleotide actions is explained by the diversity of nucle-otide receptors expressed at the nervous system. Theionotropic nucleotide receptors, P2X, are ligand-operatedchannels coupled to extracellular calcium entry and areformed by the assembly of several subunits (P2X1–7) (North2002). The metabotropic receptors, P2Y, are G protein-coupled receptors mainly associated to phospholipase Cactivation and intracellular calcium mobilization. A sub-family is composed by the most recently cloned, P2Y12,P2Y13, and P2Y14, which respond to ADP and UDP-glucose, respectively, and are coupled to adenylate cyclaseinhibition (Ralevic and Burnstock 1998; Communi et al.2001; Hollopeter et al. 2001).
Cerebellar granule neurons provide a good neuronalmodel in which to approach the function of nucleotidereceptors, as they express a great variety of bothionotropic (P2X1, P2X2, P2X4, and P2X7) and metabo-tropic (P2Y1, P2Y4, P2Y6, and P2Y12) receptors (Hervaset al. 2003). In addition, the agents and signaling cascadesinvolved in survival and differentiation processes are wellcharacterized in these cells (Contestabile 2002). Amongthese routes, we have focused on GSK-3 and investigatedif this enzyme is a target for nucleotides signaling ingranule neurons. In the present study, we have demon-strated that the metabotropic nucleotide agonist 2-methyl-thio-ADP (2MeSADP) is coupled to GSK-3 signaling ingranule neurons through a PI3-K/Akt-dependent pathway.This action is mediated by a Gi-coupled receptor, whichexhibits pharmacological characteristics similar to thatdescribed for a P2Y13-like receptor (Communi et al.2001; Fumagalli et al. 2004). While coupling of nucleo-tides to GSK-3 has already been reported by Neary andKang (2006) in rat cortical astrocytes, the novel contribu-tion of the present study is the link of 2MeSADP signalingto b-catenin nuclear translocation, implying transcriptionalregulation mediated by 2MeSADP in cerebellar granuleneurons.
Materials and methods
MaterialsMaterial from cell cultures was purchased from Worthington (Lake
Wood, NJ, USA) (papain, ovomucoid, and Dnase), GIBCO BRL
(Paisley, Renfrewshire, UK) (b-mercaptoethanol, B-27, and Neuro-
basal-A medium), and Biochrom (AG, Berlin, Germany) (poly-L-
lysine). Antibodies for phospho-GSK-3, phospho-Akt, and Akt
were from Cell Signalling Technology (Beverly, MA, USA),
antibody for total GSK-3 was from Biosource (Nivelles, Belgium),
antibodies for phospho-ERK-1,2 and total ERK-1,2 were from Santa
Cruz Biotechnologies Inc. (Santa Cruz, CA, USA), antibodies for b-catenin were from Transduction Laboratories (Lexington, KY,
USA), and antibodies for Neu-N were from Chemikon International
(Temecula, CA, USA). Secondary antibody donkey anti-mouse IgG
fluorescein-conjugated was from Jackson ImmunoResearch Labo-
ratories (West Grove, PA, USA). Amersham Biosciences Europe
GmbH (Barcelona, Spain) provided polyvinylidene difluoride
membranes and ECL method (kit Super Signal substrate western
blotting) used in western blot experiments and anti-rabbit horserad-
ish peroxidase-conjugated antibody. Anti-mouse horseradish perox-
idase-conjugated antibodies were from Santa Cruz Biotechnologies
Inc. Prolong Anti-Fade used in immunocytochemistry was from
Molecular Probes (Eugene, OR, USA). Micro-Bradford assay and
molecular-weight protein standards were from Bio-Rad (Munich,
Germany). RNA isolation was made employing the RNAeasy
system Midi-kit from Qiagen (Chatsworth, CA, USA), Dnase
digestion by employing Dnase free from Ambion (Austin, TX,
USA), and RNA quantification by the RiboGreen RNA Quantitation
Reagent (Molecular Probes). Applied Biosystems (Branchburg, NJ,
USA) provided MultiScribe Taq ManR Reverse Transcription
Reagents and AmpliTaq Gold PCR Master Mix. SYBR gold probe
for DNA detection was from Molecular Probes.
Nucleotide agonists and antagonists, insulin-like growth factor-I
(IGF-I) and transducing inhibitors were routinely purchased from
Sigma Aldrich (St Louis, MO, USA) [2MeSADP, ADP, 2-methyl-
thio-AMP (2MeSAMP), and IGF-I], Tocris Bioscience (Essex, UK)
(2¢-deoxy-N6-methyl-adenosine 3¢,5¢-diphosphate; MRS-2179), and
Calbiochem Co. (San Diego, CA, USA) [pertussis toxin (PTX),
wortmaninn, U-0126, and PP-2]. All other reagents not specified
were routinely supplied by Sigma Aldrich, Merck (Darmstadt,
Germany), or Roche Diagnostics SL (Barcelona, Spain).
Culture of cerebellar granule neuronsAll experiments carried out at the Universidad Complutense de
Madrid followed the guidelines of the International Council for
Laboratory Animal Science. Cerebellar cultures were performed
according to the procedure described by Meyer-Franke et al. (1995)and modified by Pons et al. (2001). Cerebella from Wistar rat pups
(P7) were aseptically removed and submitted to digestion with papain
100 U/mL (previously activated in Earle’s balanced salt solution
buffer containing of 5 mmol/L L-Cys, 2 mmol/L EDTA, and
0.067 mmol/L b-mercaptoethanol), in the presence of 100 U/mL of
Dnase, 1 mmol/L CaCl2, and 1 mmol/L MgCl2. The obtained cells
were re-suspended in neurobasal medium supplemented with B-27
and containing 21 mmol/L KCl, 2 mmol/L glutamine, and
antibiotics, 100 U/mL penicillin, 0.1 mg/mL streptomycin and
0.25 lg/mL amphotericin B, and plated in glass coverslips or plastic
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 62–73
ADP signals to GSK-3 and b-catenin in granule neurons | 63

Petri dishes (100 mm) pre-coated with 0.1 mg/mL poly-L-lysine at a
density of 200 000 cells/cm2. They were maintained in a humidified
incubator at 37�C in 5% CO2. Culture medium was routinely
replaced every 4 days. AraC (10 lmol/L) was added to avoid the
proliferation of glial cells.
Cultured cerebellar granule neurons were used at 10–14 DIV
(days in vitro). Prior to the stimulation of cells with nucleotide
agonists or other agents, cells were routinely washed for about 2 h
at 37�C in a low potassium medium [Locke solution, in mmol/L:
NaCl 140; KCl 4.7; CaCl2 2.5; KH2PO4 1.2; MgSO4 1.2; glucose
5.5; and HEPES (acid) 10; pH 7.4] in order to lower the basal
levels of GSK-3, Akt, and ERK phosphorylation, normally being
elevated as a consequence of the culture in high potassium
medium. The specified treatments were also applied in the same
Locke medium. For longer incubation times, as those required in
b-catenin experiments, the nucleotide agonists were applied in the
culture medium.
Western blot experimentsCells were stimulated in the presence of the nucleotide agonists
and IGF-I with different treatments for the required times.
Stimulation was stopped by addition of lysis buffer [20 mmol/L
MOPS, 50 mmol/L NaF, 40 mmol/L b-glycerophosphate,1 mmol/L sodium o-vanadate, 5 mmol/L EDTA, 2 mmol/L EGTA,
0.5% Triton X-100, pH = 7.2, 1 mmol/L phenylmethylsulfonyl
fluoride, and protease inhibitor cocktail (Complete; Roche)].
Protein determination of the cell extracts was carried out and then
mixed with sample buffer 4x [50% glycerol, 125 mmol/L Tris
pH = 6.8, 4% sodium dodecyl sulfate (SDS), 1% bromophenol
blue, 5% b-mercaptoetanol, and 4.5% H2O]. The samples were
heated at 99�C and aliquots (25 lg protein for GSK-3 and Akt
determinations, 15 lg protein for ERK and Neu-N, and 8 lg for
b-catenin) were subjected to SDS gel electrophoresis (25 mmol/L
Tris, 200 mmol/L glycine, and 0.1% SDS, pH = 8.3) using 12%
acrylamide gels. Immunotransference was performed in polyviny-
lidene difluoride membranes (25 mmol/L Tris, 192 mmol/L gly-
cine, and 20% methanol), according to that previously described
(Jimenez et al. 1999). The Tris buffered saline buffer (100 mmol/L
NaCl and 10 mmol/L Tris–HCl, pH = 7.5) containing 1% (v/v)
Tween-20 and 5% bovine serum albumin (BSA) was employed as
blocking medium and in subsequent incubations with the antibod-
ies. Incubation with the antibodies was performed at the following
dilutions: 1 : 750 for phospho-GSK-3 a/b (Ser21/9), 1 : 1000 for
total GSK-3, phospho-ERK-1,2 (Thr202 and Tyr204), total ERK,
phospho-Akt (Thr308), total Akt, and b-catenin; and 1 : 500 for
the neuron-specific nuclear protein antibody Neu-N. Primary
antibodies were detected with horseradish peroxidase-conjugated
antibodies (1 : 4000 anti-mouse and 1 : 4000 anti-rabbit) and
visualized by the ECL method. The quimioluminscence images
were quantified by densitometry employing the Fluo-S Imager of
Bio-Rad.
Measurement of GSK-3 activity in vitroGlycogen synthase kinase-3 activity was measured by the method
described by Engel et al. (2006). After the stimulation of granule
cells with the nucleotide agonists, cell extracts were obtained by
adding the lysis buffer [20 mmol/L HEPES, pH = 7.4, 100 mmol/L
NaCl, 10 mmol/L NaF, 1 mmol/L sodium o-vanadate, 1% Triton
X-100, 1 mmol/L EDTA, 1 mmol/L EGTA, and protease inhibitor
cocktail (Complete; Roche)]. After 30 min lysis on ice, cells were
centrifuged at 13 000 g for 5 min at 4�C. Reactions were performed
by incubating 25 lL of the protein extract in a buffer containing
25 mmol/L Tris pH = 7.5, 1 mmol/L dithiothreitol, and 10 mmol/L
MgCl2, together with a GSK-3 specific peptide GS-1 (final
concentration 30 lmol/L) and ATP (50 lmol/L plus 2 lCi of
[c32P]ATP) at 30�C for 10 min. GS-1 is a synthetic peptide
(YRRAAVPPSPSLSRHSSPHQS*EDEE) derived from glycogen
synthase sequence, containing a Ser (21S*) residue in phospho-
rylated form and three consensus sites for GSK-3 specific
phosphorylation (Stambolic and Woodgett 1994). This reaction
was run in parallel in the presence of 20 mmol/L LiCl, an inhibitor
which exhibits a high degree of selectivity towards GSK-3 (Davies
et al. 2000). Thereafter, aliquots were spotted onto p81 phospho-
cellulose paper (Merck), washed twice with 75 mmol/L phosphoric
acid, and measured by scintillation counting. The difference
between the kinase activity in the presence or absence of LiCl
was considered to represent the specific GSK-3 activity.
Immunocytochemistry experimentsCells plated onto glass coverslips were stimulated with the
specific effectors for the required times and then fixed with
paraformaldehyde (4%, w/v) for 15 min and permeabilized with
Triton X-100 (0.1%, v/v) in phosphate-buffered saline (PBS)/BSA
(3%, w/v) containing 5% of donkey serum for 1 h at 37�C. Theprimary antibody, mouse anti-b-catenin (1 : 200), was diluted in
PBS/BSA and applied for 1 h at 37�C. The secondary antibody,
donkey anti-mouse IgG fluorescein-conjugated (1 : 200), was also
incubated in PBS/BSA for 1 h at 37�C. Finally, cells were
washed three times and mounted following standard procedures
(Prolong Anti-Fade). Controls were prepared according to the
same protocol by replacing primary antibodies with the same
volume of PBS/BSA solution, and followed by the incubation
with the secondary antibody.
The coverslips were viewed with a Nikon TE-200 microscope
(Nikon Instruments Europe B. V., Badhoevedorp, The Netherlands)
and a Kappa DX2 camera controlled by kappa Image Base Control
software (Kappa opto-electronics GmbH, Gleichen, Germany).
Densitometric analysis of the photographs was performed using
Lucida 4.0 analyze software (Kinetic Imaging Ltd, Liverpool, UK).
RT-PCR experimentsTotal RNAwas isolated from granule cell cultures maintained at 10–
14 DIV and from whole P7 rat. First strand cDNA synthesis was
performed according to manufacturer’s protocol and RT reactions
were performed in parallel in the absence of the reverse transcriptase
enzyme, in order to evaluate the level of contaminating genomic
DNA. PCR reactions were carried out using 1–5 lL of the RT
product and specific oligonucleotide primers (300 nmol/L each) for
P2Y receptor: P2Y1, forward primer (AAGTCTCTGGGCAGGCT-
CAAGAAG) and reverse primer (CCGAATCCCAGTGCCAGA-
GTAGAA); P2Y12, forward primer (AACTTCCAGCCCCAGCAA-
TCTTTT) and reverse primer (CGTTATGTCCTTATCTTTTGGC-
CTCC); and P2Y13, forward primer: (CGTCGTACCGTTTAGG-
AAAACTTTTGTGA) and reverse primer (TTCACGGTTGATG-
CCGTTGCCTCTT). The amplification was run for 40 cycles (94�C30 s, 60�C 30 s, and 72�C 60 s). Control reactions (H2O control)
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 62–73� 2007 The Authors
64 | F. Ortega et al.

were carried out in the absence of RT product (template) to avoid
cross contamination.
Nuclear extracts preparationTo isolate nuclei, cells stimulated with the nucleotide agonists for
the specified times were washed and scraped in cold-ice PBS buffer.
After a brief centrifugation for 1 min at 500 g, the alive cells in
the pellet were then re-suspended in 300 lL lysis buffer [2/3 PBS,
1% (v/v) Nonidet P-40, 1 mmol/L Na3VO4, 50 mmol/L NaF,
0.5 mmol/L EDTA, 10% (v/v) glycerol, 1 mmol/L dithiothreitol,
1 mmol/L phenylmethylsulfonyl fluoride, and protease inhibitor
cocktail (Complete, Roche)] and kept on ice for 15 min. Following
incubation, extracts were centrifuged at 4�C for 1 min at 4000 g,and the supernatants recovered as the cytosolic fractions. The
remaining nuclear pellet was then re-suspended in 300 lL lysis
buffer containing 0.1% (v/v) SDS. Nuclear extracts were then mixed
with sample buffer 4x and the samples heated at 99�C to be ready
for western blot experiments.
Statistical analysisData are represented as mean ± SEM of at least three independent
experiments obtained from different cultures. Comparison between
sample and controls were carried out using the Dunnett test, and
comparisons between different samples were carried out by applying
the Tukey test.
Results
2MeSADP induces GSK-3 phosphorylation in cerebellargranule neuronsCultured cerebellar granule neurons were used in the secondweek (10–14 DIV), because at this stage granule neuronsexhibited the most significant responses to nucleotides. Astrophic actions of nucleotides are mostly associated to thestimulation of metabotropic P2Y receptors, we first analyzedif metabotropic nucleotide agonists were linked to GSK-3signaling in granule neurons. Among them 2MeSADP turnedout to be able to induce significant levels of GSK-3phosphorylation increasing about twofold from the basallevel. This effect was transient and early observed at the first5 min, reaching a maximum around 10–15 min after stim-ulation and rapidly declining to basal values in a 60 minperiod (Fig. 1). For subsequent experiments, a stimulationtime of 10 min period with 2MeSADP was chosen to obtainsignificant levels of GSK-3 phosphorylation.
Analysis of the receptor involved in the responses of2MeSADP on GSK-3 phosphorylation in granule neurons2MeSADP could be activating several types of nucleotidicADP receptors in granule neurons, such as P2Y1 and P2Y12
receptors, previously shown to be expressed in this cellularmodel by RT-PCR, and also in functional studies, at least forP2Y1 (Hervas et al. 2003; Leon et al. 2006). To analyze thereceptor involved in GSK-3 phosphorylation, specific antag-onists for these receptors were tested, such as MRS-2179 and
2MeSAMP, for P2Y1 and P2Y12 receptors, respectively(Boyer et al. 1998; Hollopeter et al. 2001). As shown inFig. 2a, none of these antagonists abolished the effect of2MeSADP on GSK-3 phosphorylation. The same behaviorwas observed when the incubations with the nucleotideagonists were performed in the continuous presence ofadenosine deaminase (1 U/mL), confirming that 2MeSADP-mediated effect was independent on extracellular nucleotidehydrolysis and excluding the participation of adenosinereceptors (results not shown). These results suggested thatanother receptor different from P2Y1 and P2Y12 andresponding to ADP nucleotides could be present in granuleneurons. The expression of P2Y13 receptors by RT-PCR(Fig. 2b) in these cells suggests that this receptor could beresponsible for the effect of 2MeSADP on GSK-3 phos-phorylation.
Dose–response experiments were then performed with2MeSADP and the endogenous agonist ADP, as relativeaffinities of both compounds can be used to distinguishbetween P2Y12 and the closely related P2Y13 receptor. Asshown in Fig. 2c, the calculated EC50 values were9.12 ± 2.2 nmol/L for 2MeSADP, and slightly lower,
C 1 5 15 30 60 120
400
GSK-3
pGSK-3
300
GS
K-3
ph
osp
ho
ryla
tio
n,
% in
crea
se o
ver
bas
al
200
100
0
1 µmol/L 2MeSADP
1 µmol/L 2MeSADP
min
C
**
**
*
1 5 15 30 60 120 min
Fig. 1 Effect of 2MeSADP on GSK-3 phosphorylation as a function of
time in cerebellar granule neurons. Cultured granule cells were stim-
ulated with 1 lmol/L 2MeSADP concentration at different stimulation
times. Then cells were harvested and phosphorylation of GSK-3 was
analyzed by immunoblotting, as described in Materials and methods.
Histograms represent the percentage increase with respect to non-
stimulated cells (100% control value) and were obtained by normali-
zation of densitometric values of phospho-GSK-3 with respect to total
GSK-3. Values are the mean ± SD of three experiments performed
from different cultures. The blot corresponds to a representative
experiment. Data were analyzed by Dunnett test and were statistically
significant at *p < 0.05 and **p < 0.01.
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 62–73
ADP signals to GSK-3 and b-catenin in granule neurons | 65

3.58 ± 1.3 nmol/L, for ADP, which fitted well with theestimated values for the activation of nucleotide ADPreceptors (Communi et al. 2001; Sellers et al. 2001).These data suggest the possibility that 2MeSADP-induced
GSK-3 phosphorylation could be mediated by a P2Y13
receptor, for which similar potency of ADP and 2MeSADPhas been described (Communi et al. 2001; Fumagalli et al.2004).
2MeSADP
(2MeSADP) (nmol/L) (ADP) (nmol/L)
pGSK-3
GSK-3
P2Y12
P2Y13
P2Y1
H2O Gn -RT Cb M
C 1 3 10 30 100 1000 C 1 3 10 30 100 1000
2MeSADP
Cont
pGSK-3
(a)
(c)
(b)
GSK-3
250
120
100
80
60
40
20
0
120
100
80
60
40
20
0
–10 –9 –8–Log (2MeSADP) (nmol/L) –Log (ADP) (nmol/L)
GS
K-3
ph
osp
ho
ryla
tio
n(%
max
imu
m)
–7 –6 –10 –9 –8 –7 –6
GS
K-3
ph
osp
ho
ryla
tio
n%
incr
reas
e ov
er b
asal
200
150
50
0
100
2MeSADP
2MeSAMP
MRS–2179
Cont
** ** **
2MeSADP
2MeSAMP
MRS–2179
Fig. 2 Analysis of the receptor mediating responses of 2MeSADP on
GSK-3 phosphorylation in cerebellar granule neurons. (a) Effect of
P2Y-ADP receptor antagonists. Cultured granule cells were pre-
incubated for 5 min with 10 lmol/L MRS-2179 and 100 lmol/L
2MeAMP before being stimulated with 1 lmol/L 2MeADP for 10 min.
Controls in the presence of these antagonists did not significantly
change basal levels of phosphorylation in non-stimulated cells. Then
cells were harvested and phosphorylation of GSK-3 was analyzed by
immunoblotting, as described in Materials and methods. Histograms
represent the percentage increase with respect to non-stimulated cells
(100% control value) and were obtained by normalization of densito-
metric values of phospho-GSK-3 with respect to total GSK-3. Values
are the mean ± SD of three experiments performed from different
cultures. A representative experiment is shown in the top-left panel.
Data were analyzed by Dunnett and Tukey tests and were statistically
significant at *p < 0.05 and **p < 0.01 when phosphorylation values
obtained from non-stimulated cells (control, empty histogram) were
taken as references. (b) RT-PCR expression of P2Y-ADP receptors.
Amplified PCR bands obtained in RT-PCR experiments as described
in Materials and methods were electrophoresed on a 2% agarose gel
and visualized by SYBR Gold staining. The band for P2Y receptors
were around 150 bp and were amplified both from cultured cerebellar
granule neurons (GN) and P7 whole rat cerebellum RNA extracts (Cb).
No amplification products were observed in parallel assays carried out
without reverse transcriptase ()RT) or template (H2O). M: 100-bp
DNA ladder. (c) Dose–response curves for P2Y-ADP receptors.
Concentration–dependence curves for 2MeSADP and ADP were
obtained by stimulating cells for 10 min in the presence of different
concentrations of both agonists. Data were obtained by normalization
of densitometric values of phospho-GSK-3 with respect to total GSK-3.
Results were normalized with respect to the maximal response elicited
by 1 lmol/L 2MeSADP and ADP (100%), and the curves fitted by
Boltzmann analysis. Values are the mean ± SD of three experiments
performed from different cultures. The blots correspond to represen-
tative experiments. Data were analyzed by Dunnett test and were
statistically significant at *p < 0.05 and **p < 0.01.
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 62–73� 2007 The Authors
66 | F. Ortega et al.

2MeSADP-mediated increase on GSK-3 phosphorylationoccurs through a PI3-K-Akt-dependent pathwayNext studies were carried out in order to go deeply intothe mechanism of action of 2MeSADP on GSK-3phosphorylation. In this respect two intracellular interme-diate routes were analyzed which could converge on GSK-3. We looked into the PI3-K/Akt and extracellular signal-regulated kinase (ERK) pathways to investigate whetherthese were upstream cascades of GSK-3. In these studies,IGF-I, an important survival factor in granule neurons,was taken as positive control of the PI3-K/Akt/GSK-3pathway.
In the same way to that observed for GSK-3 phosphor-ylation, 2MeSADP was able to induce the phosphorylationand subsequent activation of both Akt (Thr308) and ERK-1,2(Thr202 and Tyr204). These effects were maintained in thepresence of the P2Y1 antagonist MRS-2179 (Fig. 3). There-fore both signaling pathways were possible candidates asintermediate upstream routes leading to GSK-3. IGF-I, at theconcentration used, produced around twice as much phos-phorylation levels of GSK-3 and four times higher phos-phorylation of Akt, than those obtained with 2MeSADP,while inducing similar levels of ERK-1,2 phosphorylation tothat of the nucleotidic agonist.
Further analysis of the signaling pathway mediating2MeSADP effect on GSK-3 phosphorylation was carriedout with specific transducing inhibitors. First, the effect ofPTX was investigated. The experiments performed in thepresence of PTX showed that the toxin completely abolished2MeSADP-mediated effect, confirming the requirementof a Gi-coupled P2Y receptor specific for ADP involved inGSK-3 phosphorylation in granule neurons (Fig. 4).
In addition, the PI3-K inhibitor wortmannin, at 1 lmol/Lconcentration, which inhibits PI3-K signaling in this cellularmodel, completely abolished 2MeSADP-mediated effects onGSK-3 phosphorylation. The ERK-activating kinase-1 inhib-itor, U-0126, which prevents ERK-phosphorylation, keepsthe same pattern response of control experiments (Fig. 4).These data suggests that phosphorylation of GSK-3 by2MeSADP stimulation is mediated primarily via the PI3-K-Akt-dependent pathway, although some crosstalk with themitogen-activated protein kinase pathway cannot be ruledout. Finally, the treatment of granule neurons with theinhibitor of src-tyrosine kinases PP-2, produced a significantdecrease in the basal phosphorylation levels, although it didnot abolish the effect of 2MeSADP on GSK-3 phosphory-lation (Fig. 4). Thus src-tyrosine kinases did not appeared tobe an intermediate step in the PI3-K to GSK-3 pathway
C 2MeS 2MeS+ MRS
IGF-I
** **
***
p-GSK3/GSK-3 tot
1000
900
GS
K-3
ph
osp
ho
ryla
tio
nin
crea
se o
ver
bas
al
600
500
400
300
200
100
0C 2MeS 2MeS
+ MRSIGF-I C 2MeS 2MeS
+ MRSIGF-I
****
**
###
###
****
*
Cont2M
eS
+MRS
IGF-I
Cont2M
eS
+MRS
IGF-I
Cont2M
eS
+MRS
IGF-I
p-Akt/Akt tot p-ERK1,2/ERK tot
Fig. 3 2MeSADP induced Akt and ERK-1,2 phosphorylation in cul-
tured cerebellar granule neurons. Akt and ERK activation by 2MeSADP
in cerebellar granule neurons were analyzed by using specific anti-
bodies that recognize the phosphorylated forms of these two proteins,
Akt at Thr308, and ERK-1,2 at Thr202 and Tyr204. Cells were stimu-
lated with 2MeSADP, in the presence or absence of 10 lmol/L MRS-
2179, and IGF-I (50 ng/mL) for 10 min. Cell extracts were obtained and
these proteins were analyzed by immunoblotting, as described in
Materials and methods. GSK-3 phosphorylation was analyzed in
the same extracts as described before. Histograms represent the
percentage increase with respect to non-stimulated cells (100% control
value) and were obtained by normalization of densitometric values of
the phosphoproteins with respect to the total forms of GSK-3, Akt, and
ERK. Values are the mean ± SD of three experiments performed from
different cultures. The blots correspond to representative experiments.
Data were analyzed by Dunnett and Tukey tests and were statistically
significant at *p < 0.05 and **p < 0.01, when the phosphorylation
values of non-stimulated cells (empty bars) were taken as reference for
each phosphoprotein separately, and at ###p < 0.001 with respect to
treatment with 2MeSADP ± MRS-2179.
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 62–73
ADP signals to GSK-3 and b-catenin in granule neurons | 67
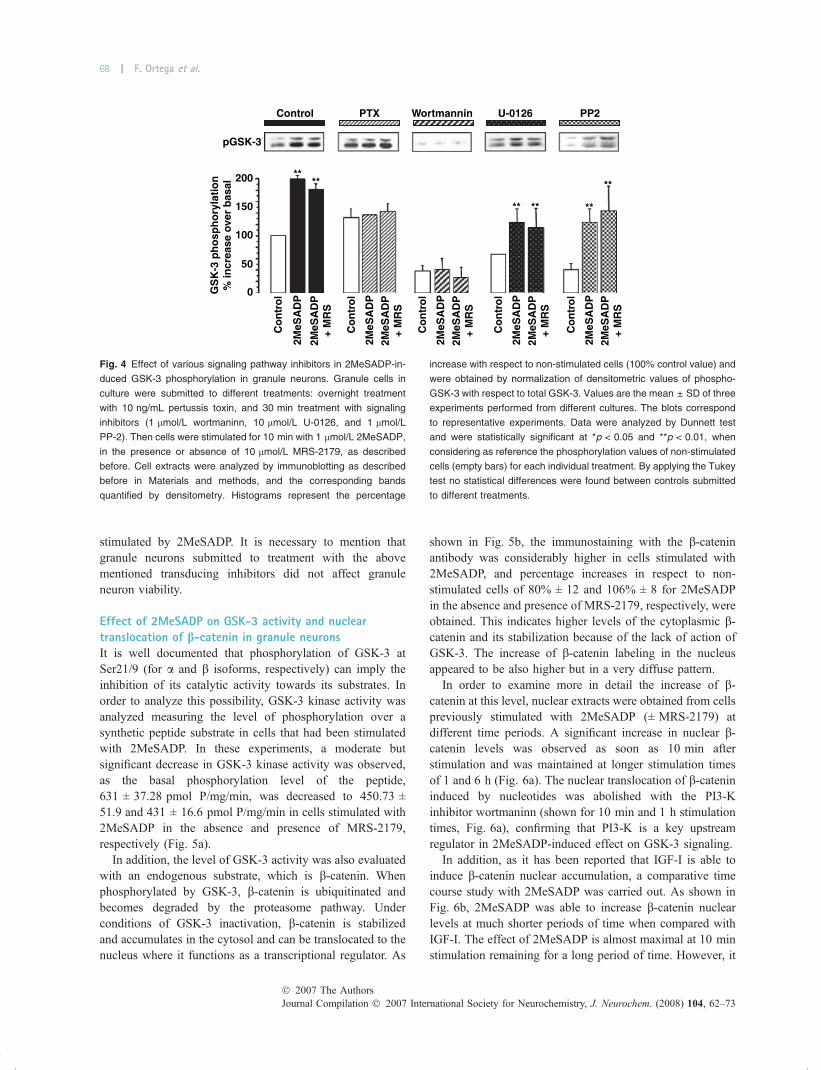
stimulated by 2MeSADP. It is necessary to mention thatgranule neurons submitted to treatment with the abovementioned transducing inhibitors did not affect granuleneuron viability.
Effect of 2MeSADP on GSK-3 activity and nucleartranslocation of b-catenin in granule neuronsIt is well documented that phosphorylation of GSK-3 atSer21/9 (for a and b isoforms, respectively) can imply theinhibition of its catalytic activity towards its substrates. Inorder to analyze this possibility, GSK-3 kinase activity wasanalyzed measuring the level of phosphorylation over asynthetic peptide substrate in cells that had been stimulatedwith 2MeSADP. In these experiments, a moderate butsignificant decrease in GSK-3 kinase activity was observed,as the basal phosphorylation level of the peptide,631 ± 37.28 pmol P/mg/min, was decreased to 450.73 ±51.9 and 431 ± 16.6 pmol P/mg/min in cells stimulated with2MeSADP in the absence and presence of MRS-2179,respectively (Fig. 5a).
In addition, the level of GSK-3 activity was also evaluatedwith an endogenous substrate, which is b-catenin. Whenphosphorylated by GSK-3, b-catenin is ubiquitinated andbecomes degraded by the proteasome pathway. Underconditions of GSK-3 inactivation, b-catenin is stabilizedand accumulates in the cytosol and can be translocated to thenucleus where it functions as a transcriptional regulator. As
shown in Fig. 5b, the immunostaining with the b-cateninantibody was considerably higher in cells stimulated with2MeSADP, and percentage increases in respect to non-stimulated cells of 80% ± 12 and 106% ± 8 for 2MeSADPin the absence and presence of MRS-2179, respectively, wereobtained. This indicates higher levels of the cytoplasmic b-catenin and its stabilization because of the lack of action ofGSK-3. The increase of b-catenin labeling in the nucleusappeared to be also higher but in a very diffuse pattern.
In order to examine more in detail the increase of b-catenin at this level, nuclear extracts were obtained from cellspreviously stimulated with 2MeSADP (± MRS-2179) atdifferent time periods. A significant increase in nuclear b-catenin levels was observed as soon as 10 min afterstimulation and was maintained at longer stimulation timesof 1 and 6 h (Fig. 6a). The nuclear translocation of b-catenininduced by nucleotides was abolished with the PI3-Kinhibitor wortmaninn (shown for 10 min and 1 h stimulationtimes, Fig. 6a), confirming that PI3-K is a key upstreamregulator in 2MeSADP-induced effect on GSK-3 signaling.
In addition, as it has been reported that IGF-I is able toinduce b-catenin nuclear accumulation, a comparative timecourse study with 2MeSADP was carried out. As shown inFig. 6b, 2MeSADP was able to increase b-catenin nuclearlevels at much shorter periods of time when compared withIGF-I. The effect of 2MeSADP is almost maximal at 10 minstimulation remaining for a long period of time. However, it
Control PTX
pGSK-3
200 ****
** ** **
**
100
150G
SK
-3 p
ho
sph
ory
lati
on
% in
crea
se o
ver
bas
al
50
0
Co
ntr
ol
Co
ntr
ol
2MeS
AD
P
2MeS
AD
P
2MeS
AD
P+
MR
S
2MeS
AD
P+
MR
S
Co
ntr
ol
2MeS
AD
P2M
eSA
DP
+ M
RS
Co
ntr
ol
2MeS
AD
P2M
eSA
DP
+ M
RS
Co
ntr
ol
2MeS
AD
P2M
eSA
DP
+ M
RS
Wortmannin U-0126 PP2
Fig. 4 Effect of various signaling pathway inhibitors in 2MeSADP-in-
duced GSK-3 phosphorylation in granule neurons. Granule cells in
culture were submitted to different treatments: overnight treatment
with 10 ng/mL pertussis toxin, and 30 min treatment with signaling
inhibitors (1 lmol/L wortmaninn, 10 lmol/L U-0126, and 1 lmol/L
PP-2). Then cells were stimulated for 10 min with 1 lmol/L 2MeSADP,
in the presence or absence of 10 lmol/L MRS-2179, as described
before. Cell extracts were analyzed by immunoblotting as described
before in Materials and methods, and the corresponding bands
quantified by densitometry. Histograms represent the percentage
increase with respect to non-stimulated cells (100% control value) and
were obtained by normalization of densitometric values of phospho-
GSK-3 with respect to total GSK-3. Values are the mean ± SD of three
experiments performed from different cultures. The blots correspond
to representative experiments. Data were analyzed by Dunnett test
and were statistically significant at *p < 0.05 and **p < 0.01, when
considering as reference the phosphorylation values of non-stimulated
cells (empty bars) for each individual treatment. By applying the Tukey
test no statistical differences were found between controls submitted
to different treatments.
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 62–73� 2007 The Authors
68 | F. Ortega et al.
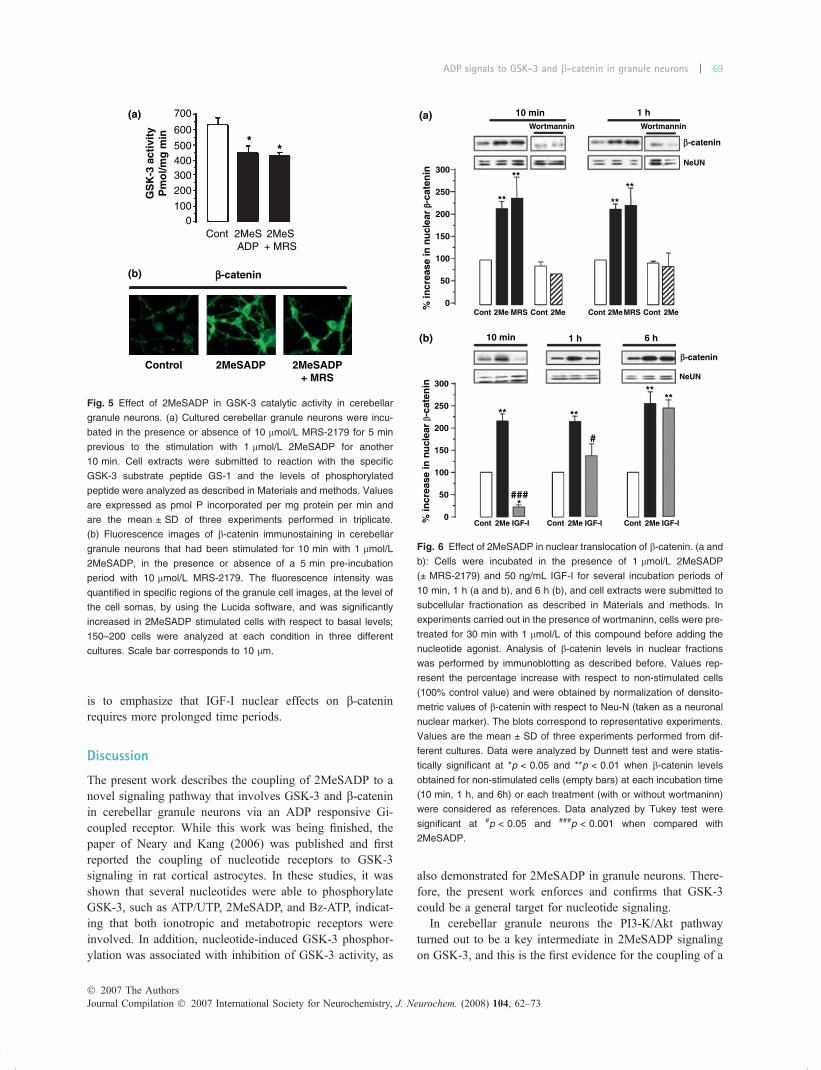
is to emphasize that IGF-I nuclear effects on b-cateninrequires more prolonged time periods.
Discussion
The present work describes the coupling of 2MeSADP to anovel signaling pathway that involves GSK-3 and b-cateninin cerebellar granule neurons via an ADP responsive Gi-coupled receptor. While this work was being finished, thepaper of Neary and Kang (2006) was published and firstreported the coupling of nucleotide receptors to GSK-3signaling in rat cortical astrocytes. In these studies, it wasshown that several nucleotides were able to phosphorylateGSK-3, such as ATP/UTP, 2MeSADP, and Bz-ATP, indicat-ing that both ionotropic and metabotropic receptors wereinvolved. In addition, nucleotide-induced GSK-3 phosphor-ylation was associated with inhibition of GSK-3 activity, as
also demonstrated for 2MeSADP in granule neurons. There-fore, the present work enforces and confirms that GSK-3could be a general target for nucleotide signaling.
In cerebellar granule neurons the PI3-K/Akt pathwayturned out to be a key intermediate in 2MeSADP signalingon GSK-3, and this is the first evidence for the coupling of a
(a)
(b)
700
**
GS
K-3
act
ivit
yP
mo
l/mg
min 600
500400300200
1000
Cont 2MeSADP
bb-catenin
Control 2MeSADP 2MeSADP+ MRS
2MeS+ MRS
Fig. 5 Effect of 2MeSADP in GSK-3 catalytic activity in cerebellar
granule neurons. (a) Cultured cerebellar granule neurons were incu-
bated in the presence or absence of 10 lmol/L MRS-2179 for 5 min
previous to the stimulation with 1 lmol/L 2MeSADP for another
10 min. Cell extracts were submitted to reaction with the specific
GSK-3 substrate peptide GS-1 and the levels of phosphorylated
peptide were analyzed as described in Materials and methods. Values
are expressed as pmol P incorporated per mg protein per min and
are the mean ± SD of three experiments performed in triplicate.
(b) Fluorescence images of b-catenin immunostaining in cerebellar
granule neurons that had been stimulated for 10 min with 1 lmol/L
2MeSADP, in the presence or absence of a 5 min pre-incubation
period with 10 lmol/L MRS-2179. The fluorescence intensity was
quantified in specific regions of the granule cell images, at the level of
the cell somas, by using the Lucida software, and was significantly
increased in 2MeSADP stimulated cells with respect to basal levels;
150–200 cells were analyzed at each condition in three different
cultures. Scale bar corresponds to 10 lm.
(a)
(b)
10 min
10 min
1 h
1 h 6 h
Wortmannin
300
250
200
% in
crea
se in
nu
clea
r bb-
cate
nin
% in
crea
se in
nu
clea
r b-
cate
nin
150
100
0Cont
Cont 2Me IGF-I Cont 2Me IGF-I Cont 2Me IGF-I
Cont Cont Cont2Me 2Me 2Me 2MeMRS MRS
50
300
250
200
150
100
0
50
Wortmannin
NeUN
**
****
****
*
#
###
**
**
**
b-catenin
NeUN
b-catenin
Fig. 6 Effect of 2MeSADP in nuclear translocation of b-catenin. (a and
b): Cells were incubated in the presence of 1 lmol/L 2MeSADP
(± MRS-2179) and 50 ng/mL IGF-I for several incubation periods of
10 min, 1 h (a and b), and 6 h (b), and cell extracts were submitted to
subcellular fractionation as described in Materials and methods. In
experiments carried out in the presence of wortmaninn, cells were pre-
treated for 30 min with 1 lmol/L of this compound before adding the
nucleotide agonist. Analysis of b-catenin levels in nuclear fractions
was performed by immunoblotting as described before. Values rep-
resent the percentage increase with respect to non-stimulated cells
(100% control value) and were obtained by normalization of densito-
metric values of b-catenin with respect to Neu-N (taken as a neuronal
nuclear marker). The blots correspond to representative experiments.
Values are the mean ± SD of three experiments performed from dif-
ferent cultures. Data were analyzed by Dunnett test and were statis-
tically significant at *p < 0.05 and **p < 0.01 when b-catenin levels
obtained for non-stimulated cells (empty bars) at each incubation time
(10 min, 1 h, and 6h) or each treatment (with or without wortmaninn)
were considered as references. Data analyzed by Tukey test were
significant at #p < 0.05 and ###p < 0.001 when compared with
2MeSADP.
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 62–73
ADP signals to GSK-3 and b-catenin in granule neurons | 69

P2Y13 receptor to this signal. There are many examples in theliterature involving this route in signaling displayed bynucleotides acting through other ADP receptors, such asP2Y1 receptors in adventitial fibroblasts and astrocytomacells, and P2Y12 receptors in C6 glioma, in which they aremediating proliferative actions (Sellers et al. 2001; Gerasi-movskaya et al. 2005; Van Kolen et al. 2006). However, inrat cortical astrocytes ATP-induced GSK-3 phosphorylationis only slightly regulated by PI3-K/Akt and is mainlysubmitted to protein kinase C-dependent regulation (Nearyand Kang 2006).
The receptor mediating the 2MeSADP-induced increase inGSK-3 phosphorylation fits to a P2Y13-like receptor in
granule neurons
The target of 2MeSADP-described effect on GSK-3 phos-phorylation in granule neurons fits better to the pharmaco-logical profile of a P2Y13 receptor, rather than other ADPreceptors expressed, such as P2Y1 and P2Y12. These cellsexpress a significant level of P2Y1 receptor and it has beendescribed to be coupled to an increase in the intracellularcalcium concentration ([Ca2+]i) and calcium calmodulin-dependent kinase II activation (Hervas et al. 2003; Leonet al. 2006). However, 2MeSADP action on GSK-3 phos-phorylation did not appear to be mediated by P2Y1, becausethe selective antagonist MRS-2179 did not inhibit this effect.In addition, the effect was sensitive to PTX suggesting theinvolvement of a Gi-coupled receptor, such as P2Y12 andP2Y13 receptor subtypes that have been described to bemainly expressed in blood cells and brain (Communi et al.2001; Hollopeter et al. 2001; Laitinen et al. 2001). TheP2Y12 receptor was discarded on the basis of the lack ofeffect of the specific antagonist 2MeSAMP. In addition, thesimilar relative potencies of 2MeSADP and ADP, whichallows the discrimination between P2Y12 and P2Y13 sub-types, agrees with the presence of P2Y13, specially for thatdescribed for mouse and rat orthologs, in which the potencyof ADP may be similar or even higher than 2MeSADP(Zhang et al. 2002; Marteau et al. 2003; Fumagalli et al.2004). Besides, it is well documented for P2Y12 receptor,that 2MeSADP is much more potent than ADP, around 1order of magnitude (Hollopeter et al. 2001; Zhang et al.2001). Therefore, all these considerations favor the possibil-ity of a P2Y13 over a P2Y12 receptor mediating 2MeSADPeffects on granule neurons.
In this respect, P2Y13 expression in cultured granule cellshas been confirmed by RT-PCR in cerebellar granuleneurons, which also showed a clear expression of functionalP2Y1 receptors. This agrees with the observed co-expressionof several types of ADP receptors in the same cellular modelthat seems to be a common feature in mammalian cells. Indendritic cells P2Y13 co-exists with another not yet charac-terized ADP receptor, they are both coupled to ERKactivation and exhibit different anti-inflammatory effects
(Marteau et al. 2004). Similarly, in cerebellar astrocytesP2Y1 and P2Y13 receptors are co-expressed and mediateERK phosphorylation and intracellular calcium mobilization(Carrasquero et al. 2005, 2006). In addition, it is generallyobserved that the co-expression of several ADP receptorsresults in crosstalk and reciprocal interaction between them.A good example for such interaction is the platelet model, inwhich P2Y1 and P2Y12 receptors interact reciprocally. P2Y12
receptor potentiates the P2Y1-induced calcium response viaPI3-K and adenylate cyclase inhibition, whereas P2Y1
inhibits the P2Y12 signal through src-kinase activation(Hardy et al. 2004). In this respect, an interaction betweenP2Y13 and other P2Y receptors mediating the effect of2MeSADP cannot be excluded in granule neurons.
Physiological meaning of 2MeSADP signaling throughGSK-3 and b-catenin in granule neuronsTaking into consideration the multiple regulatory actions ofGSK-3, the physiological meaning of its inactivation bynucleotides implies a wide range of actions, which woulddepend on the cellular model, the differentiation state of thecell, the environmental factors, the specific targets orsubstrates, and the various pools located in different cellularmicrodomains in which a particular signal could be acting. Incortical astrocytes, when exposed to stress conditions ofmechanical strain, GSK-3 is inhibited, and this effect ismediated by the newly released nucleotides. Therefore,nucleotides are mediating a protective role during processesof brain injury through GSK-3 signaling (Neary and Kang2006).
In this respect, GSK-3 has been considered a key mediatorin processes of death and survival, especially in neuronalmodels. In fact, growth and survival factors trigger differentsignaling cascades that converge in GSK-3 (Hetman et al.2000). This has been extensively studied in cerebellargranule neurons, which are considered a cell cultureparadigm to understand the molecular mechanism underlyingapoptosis. In this cellular model, some factors are able torescue from apoptotic-stimuli: depolarizing K+ levels, neu-rotrophic factors (IGF-I), cAMP, and Li+ (D’Mello et al.1993, 1994; Miller et al. 1997; Subramaniam et al. 2003).Interestingly all these survival factors trigger a cascade ofintracellular events that have in common GSK-3 phosphor-ylation and inhibition through the PI3-K/Akt pathway, asshown for IGF-I in the literature and in the present study (Papand Cooper 1998; Chin et al. 2005). Considering the lowerlevels of GSK-3 phosphorylation obtained for 2MeSADP inrespect to that obtained for IGF-I, it is tempting to assumethat 2MeSADP could mimic survival actions of growthfactors, acting in combination with them or potentiating theirneuroprotective actions under certain conditions. In thisrespect, previous work reported protective actions of ADPand adenosine from low-potassium-induced apoptosis,which clearly involved a nucleotide receptor, although the
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 62–73� 2007 The Authors
70 | F. Ortega et al.

underlying mechanisms were not fully elucidated (Vitoloet al. 1998).
The modest decrease in GSK-3 activity observed withnucleotides in the present study is only slightly lower thanthat obtained for the most representative signals coupled toGSK-3 inhibition, such as growth factors, for which no morethan 40–50% decrease in GSK-3 activity has been reported.This seems to be sufficient to trigger significant downstreamresponses (Ding et al. 2000; Neary and Kang 2006), and inthe case of nucleotides these events lead to b-catenin. Thesignaling of 2MeSADP to b-catenin described in the presentwork has not been previously reported for nucleotides. b-Catenin is a multifunctional protein whose stability is mainlyregulated by GSK-3. The cytosolic accumulation of b-catenin followed by its translocation to the nucleus, allows itsinteraction with Lef/Tcf family of transcription factors(Domınguez and Green 2001; Grimes and Jope 2001).Recent evidence confirms the b-catenin regulation by diversesignaling stimuli. For example, b-catenin-induced Tcf/Lef-dependent transcriptional activity has been described forIGF-I in hepatoma cells (Desbois-Mouthon et al. 2001),hepatocyte growth factor in mammary epithelial cells(Papkoff and Aikawa 1998), and fibroblast growth factor-IIin human umbilical vein endothelial cells (Holnthoner et al.2002). This activity was demonstrated to be directly linked toGSK-3 phosphorylation, as it was lost after over-expressionof an unhibitable GSK-3 mutant. It is to be emphasized thatthe IGF-I effect in hepatoma cells (Desbois-Mouthon et al.2001) and fibroblast growth factor-induced b-catenin stabil-ization in cortical neurons (Hashimoto et al. 2002), involvesthe PI3-K/Akt pathway, as we report in this work for2MeSADP in granule neurons. In other systems, growth anddevelopmental factors are coupled to b-catenin stabilizationand transcriptional activity, but this does not correlate withPI3-K/Akt-dependent regulation of GSK-3 activity (Dinget al. 2000; Playford et al. 2000; Chen et al. 2005;McManus et al. 2005). Nevertheless, this PI3-K/Akt path-way seems to be important for the maintenance of growthfactor-mediated b-catenin cascade in other cellular models(Naito et al. 2005; Kim et al. 2007). In addition to growthfactors, b-catenin-dependent transcriptional activity has beenrecently reported for some G protein-coupled receptors(Shevtsov et al. 2006), such as those of PGE2 and lyso-phosphatidic acid in colon cancer cells, and has turned out tobe, at least partially, dependent on GSK-3 inactivation bySer21/9 phosphorylation (Castellone et al. 2005; Shao et al.2005; Yang et al. 2005).
Different genes have been reported to be regulated byb-catenin, such as those involved in cell cycle regulation,cell adhesion, migration, and survival (Novak and Dedhar1999; Harris and Peifer 2005). Besides, two genes related topurinergic metabolism were recently described to containLef-1 binding sites in their promoter region, the 5¢-nucle-otidase (CD 73), a nucleotide hydrolytic ectoenzyme, and
adenosine deaminase, which deaminates adenosine intoinosine (Spychala and Kitajewski 2004). In relation to thegenes that 2MeSADP could be regulating in granuleneurons, it is interesting to notice the rapid time course ofb-catenin nuclear accumulation found for 2MeSADP in thepresent study. For other signals, such as hepatocyte growthfactor and lysophosphatidic acid, b-catenin has beenreported to accumulate at the nucleus in less than 30 min(Papkoff and Aikawa 1998; Yang et al. 2005). These effectsare consistent with b-catenin being a short-lived protein thatis present at very low cytosolic levels in the absence ofstimuli, because of its continuous degradation by theubiquitin-proteasome pathway (Papkoff 1996; Aberle et al.1997). The rapid effect seen with 2MeSADP in comparisonwith IGF-I in granule neurons suggests that this nucleotide,which is a more diffusible signal, can be responding faster tochanges in granule cell viability and being able to regulateearly inducible genes. Further work is required to elucidatethe mechanism underlying b-catenin stabilization andnuclear translocation by ADP in granule neurons in orderto clarify the functional meaning of b-catenin being aphysiological target of the ADP signaling in this cellularmodel.
Acknowledgements
We are in debt with Dr Felix Hernandez (CBMSO, Madrid, Spain)
for the in vitro determination of GSK-3 activity, and we thank Dr
Esmerilda G. Delicado for her useful comments and help in the
preparation of the manuscript. We thank Elena Fariza for English
corrections of the manuscript. FO is a fellowship of MECD. This
works was supported by research grants from the Spanish Ministry
of Education and Science BFU2005-0207079, the Fundacion La
Caixa nº BM05-114-0, and The Fundacion Marcelino Botın.
References
Aberle H., Bauer A., Stappert J., Kispet A. and Kemler R. (1997)b-\Catenin is a target for the ubiquitin-proteasome pathway. EMBOJ. 16, 3797–3804.
Alvarez G., Munoz-Montano J. R., Satrustegui J., Avila J., Bogonez E.and Dıaz-Nido J. (1999) Lithium protects cultured neurons againstb-amyloid-induced neurodegeneration. FEBBS Lett. 453, 260–264.
Bhat R. V., Budd Haeberlein S. L. B. and Avila J. (2004) Glycogensynthase kinase-3: a drug target for CNS therapies. J. Neurochem.89, 1313–1317.
Boyer J. L., Mohanram A., Camaioni E., Jacobson K. A. and HardenT. K. (1998) Competitive and selective antagonism of P2Y1receptors by N6-methyl 2¢-deoxyadenosine 3¢,5¢-bisphosphate. Br.J. Pharmacol. 124, 1–3.
Burnstock G. (2005) Purinergic signalling: therapeutic potential. Anal.Real. Acad. Nac. Farm. 71, 283–319.
Burnstock G. (2006) Historical review: ATP as a neurotransmitter.Trends Pharmacol. Sci. 27, 166–176.
Carrasquero L. M. G., Delicado E. G., Jimenez A. I., Perez-Sen R. andMiras-Portugal M. T. (2005) Cerebellar astrocytes co-expressseveral ADP receptors. Presence of functional P2Y13-like recep-tors. Purinergic Signal. 1, 153–159.
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 62–73
ADP signals to GSK-3 and b-catenin in granule neurons | 71

Carrasquero L. M. G., Delicado E. G. and Miras-Portugal M. T. (2006)Which is the receptor mediating ADP signaling in rat cerebellarastrocytes. Purinergic Signalling, Vol. 2 Abstract. 8th InternationalSymposium on Adenosine and Adenine Nucleotides, Ferrara, Italy,pp. 323.
Castellone M. D., Teramoto H., Williams B. O., Druey K. M. andGutkind S. (2005) Protaglandin E2 promotes colon cancer cellgrowth through a Gs-axin-b-catenin signalling axis. Science 310,1504–1510.
Chen J., Wu A., Sun H., Drakas R., Garofalo C., Cascio S., Surmacz E.and Baserga R. (2005) Functional significance of type 1 insulin-like growth factor-mediated nuclear translocation of the insulinreceptor substrate-1 and b-catenin. J. Biol. Chem. 280, 29912–29920.
Chin P. C., Majdzadeh N. M. and D’Mello S. R. (2005) Inhibition ofGSK-3 b is a common event in neuroprotection by different sur-vival factors. Mol. Brain Res. 137, 193–201.
Communi D., Gonzalez N. D., Detheux M., Brezillon S., Lannoy V.,Parmentier M. and Boeynaems J. M. (2001) Identification of anovel human ADP receptor coupled to G(i). J. Biol. Chem. 276,41479–41485.
Contestabile A. (2002) Cerebellar granule cells as a model to studymechanisms of neuronal apoptosis or survival in vivo and in vitro.Cerebellum 1, 41–55.
Cross D. A. E., Culbert A. A., Chalmers K. A., Facci L., Skaper S. D.and Reith A. D. (2001) Selective small-molecule inhibitors ofglycogen synthase kinase-3 activity protect primary neurons fromdeath. J. Neurochem. 77, 94–102.
D’Mello S. R., Galli C., Ciotti T. and Calissano P. (1993) Induction ofapoptosis in cerebellar granule neurons by low potassium: inhibi-tion of death by insulin-like growth factor-I and cAMP. Proc. NatlAcad. Sci. USA 90, 10989–10993.
D’Mello S. R., Anelli R. and Calissano P. (1994) Lithium inducesapoptosis in immature cerebellar granule neurons but promotessurvival of mature neurons. Exp. Cell Res. 211, 332–338.
Davies S. P., Reddy H., Caivano M. and Cohen P. (2000) Specificity andmechanism of action of some commonly used protein kinaseinhibitors. Biochem. J. 351, 95–105.
Desbois-Mouthon C., Cadoret A., Blivet-Van Eggelpoel M-J., BertrandF., Cherqui G., Perret C. and Capeau J. (2001) Insulin and IGF-1stimulate the b-catenin pathway through two signalling cascadesinvolving GSK-3b inhibition and Ras activation. Oncogene 20,252–259.
Ding V. W., Chen R.-H. and McCormick F. (2000) Differential regula-tion of glycogen synthase kinase 3b by insulin and Wnt signalling.J. Biol. Chem. 275, 32475–32481.
DiVirgilio F. (2005) Purinergic mechanism in the immune system: asignal of danger for dendritic cells. Purinergic Signal. 1, 205–209.
Domınguez I. and Green J. B. A. (2001) Missing links in GSK3 regu-lation. Dev. Biol. 235, 303–313.
Engel T., Hernandez F., Avila J. and Lucas J. J. (2006) Full reversal ofAlzheimer’s disease-like phenotype in a mouse model with con-ditional overexpression of glycogen synthase kinase-3. J. Neurosci.26, 5083–5090.
Erlinge D. (1998) Extracellular ATP: a growth factor for vascular smoothmuscle cells. Gen. Pharmacol. 31, 1–8.
Fumagalli M., Trincavelli L., Lecca D., Martini C., Ciana P. and Ab-bracchio M. P. (2004) Cloning, pharmacological characterizationand distribution of the rat G-protein-coupled P2Y13 receptor. Bio-chem. Pharmacol. 68, 113–124.
Gerasimovskaya E. V., Tucker D. A., Weiser-Evans M., Wenzlau J. M.,Klemm D. J., Banks M. and Stenmark K. R. (2005) ExtracellularATP-induced proliferation of adventitial fibroblasts requiresphosphoinositide 3-kinase, Akt, mammalian target of rapamycin
and p70 S6 kinase signaling pathways. J. Biol. Chem. 280, 1838–1848.
Gerevich Z. and Illes P. (2004) P2Y receptors and pain transmission.Purinergic Signal. 1, 3–10.
Grimes C. A. and Jope R. S. (2001) The multifaceted roles of glycogensynthase kinase 3b in cellular signalling. Prog. Neurobiol. 65, 391–426.
Hall A. C., Brennan A., Goold R. G., Cleverley K., Lucas F. R., Gordon-Weeks P. R. and Salinas P. C. (2002) Valproate regulates GSK-3-mediated axonal remodelling and synapsin I clustering in devel-oping neurons. Mol. Cell. Neurosci. 20, 257–270.
Hardy A. R., Jones M. L., Mundell S. J. and Poole A. W. (2004) Re-ciprocal cross-talk between P2Y1 and P2Y12 receptors at the levelof calcium signalling in human platelets. Blood 104, 1745–1752.
Harris T. J. C. and Peifer M. (2005) Decisions: b-catenin chooses be-tween adhesion and transcription. Trends Cell Biol. 15, 234–237.
Hashimoto M., Sagara Y., Langford D., Everall I. P., Mallory M.,Everson A., Digicaylioglu M. and Masliah E. (2002) Fibroblastgrowth factor 1 regulates signalling via the glycogen synthase ki-nase-3b pathway. J. Biol. Chem. 277, 32985–32991.
Hervas C., Perez-Sen R. and Miras-Portugal M. T. (2003) Coexpressionof functional P2X and P2Y nucleotide receptors in single cerebellargranule cells. J. Neurosci. Res. 73, 384–399.
Hetman M., Cavanaugh J. E., Kimelman D. and Xia Z. (2000) Role ofglycogen synthase kinase-3b in neuronal apoptosis induced bytrophic withdrawal. J. Neurosci. 20, 2567–2574.
Hollopeter G., Jantzen H.-M., Vincent D. et al. (2001) Identification ofplatelet ADP receptor targeted by antithrombotic drugs. Nature409, 202–207.
Holnthoner W., Pillinger M., Groger M., Wolff K., Ashton A. W.,Albanese C., Neumeister P., Pestell R. G. and Petzelbauer P. (2002)Fibroblast growth factor-2 induces Lef/Tcf-dependent transcriptionin human endothelial cells. J. Biol. Chem. 277, 45847–45853.
Inoue K., Tsuda M. and Koizumi S. (2005) ATP receptors in painsensation: involvement of spinal microglia and P2X4 receptors.Purinergic Signal. 1, 95–100.
Jimenez A. I., Castro E., Mirabet M., Franco R., Delicado E. G. andMiras-Portugal M. T. (1999) Potentiation of ATP calcium re-sponses by A2B receptors stimulation and other signals coupled toGs proteins in type-1 cerebellar astrocytes. Glia 26, 119–128.
Jope R. S. and Johnson G. V. W. (2004) The glamour and gloom ofglycogen synthase kinase-3. Trends Biochem. Sci. 29, 95–102.
Kim S-E., Lee W-J. and Choi K-Y. (2007) The PI3 kinase-Akt pathwaymediates Wnt3a-induced proliferation. Cell. Signal. 19, 511–518.
Kucher B. M. and Neary J. T. (2005) Bi-functional effects of ATP/P2receptor activation on tumour necrosis factor-a release inlipopolysaccharide-stimulated astrocytes. J. Neurochem. 82, 525–535.
Laitinen J. T., Uri A., Raidaru G. and Miettinen R. (2001) [35S]GTPcSautoradiography reveals a wide distribution of Gi/o-linked ADPreceptors in the nervous system: close similarities with the plateletP2YADP receptor. J. Neurochem. 77, 505–518.
Leon D., Hervas C. and Miras-Portugal M. T. (2006) Activation of P2Y1
and P2X7 receptors induce calcium/calmodulin-dependent proteinkinase II phosphorylation in cerebellar granule neurons. Eur. J.Neurosci. 23, 2999–3013.
Lucas J. J., Hernandez F., Gomez-Ramos P., Moran M. A., Hen R. andAvila J. (2001) Decreased nuclear beta-catenin, tau hyperphorph-orylation and neurodegeneration in GSK-3beta conditional trans-genic mice. EMBO J. 20, 27–39.
McManus E. J., Sakamoto K., Armit L. J., Ronaldson L., Shpiro N.,Marquez R. and Alessi D. R. (2005) Role that phosphorylation ofGSK-3 plays in insulin and Wnt signalling defined by knockinganalysis. EMBO J. 24, 1571–1583.
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 62–73� 2007 The Authors
72 | F. Ortega et al.

Marteau F., Le Poul E., Communi D., Communi D., Labouret C., Savi P.,Boeynaems J.-M. and Suarez-Gonzalez N. (2003) Pharmacologicalcharacterization of the human P2Y13.Mol. Pharmacol. 64, 104–112.
Marteau F., Communi D., Boeynaems J.-M. and Suarez-Gonzalez N.(2004) Involvement of multiple P2Y receptors and signalling path-ways in the action of adenine nucleotides diphosphates on humanmonocyte-derived dendritic cells. J. Leukoc. Biol. 76, 796–803.
Meyer-Franke A., Kaplan M. R., Pfrieger F. W. and Barres B. A. (1995)Characterisation of the signalling interactions that promote thesurvival and growth of developing retinal ganglion cells in culture.Neuron 15, 805–819.
Miller T. M., Tansey M. G., Johnson E. M. and Creedon D. J. (1997)Inhibition of phosphatidylinositol 3-kinase activity blocks depo-larization- and insulin-like growth factor I-mediated survival ofcerebellar granule neurons. J. Biol. Chem. 272, 9847–9853.
Naito A. T., Akazawa H., Takano H., Minamino T., Nagai T., AburataniH. and Komuro I. (2005) Phosphatidylinositol 3-kinase-Akt path-way plays a critical role in early cardiomyogenesis by regulatingcanonical Wnt signaling. Circ. Res. 97, 144–151.
Neary J. T. and Kang Y. (2005) Signaling from P2 nucleotide receptorsto protein kinase cascades induced by CNS injury. Mol. Neurobiol.31, 95–103.
Neary J. T. and Kang Y. (2006) P2 purinergic receptors signal to gly-cogen synthase kinase-3beta in astrocytes. J. Neurosci. Res. 84,515–524.
Neary J. T., McCarthy M., Cornel-Bell A. and Kang Y. (1999) Trophicsignalling pathways activated by purinergic receptors in rat andhuman astroglia. Prog. Brain Res. 120, 323–332.
North R. A. (2002) Molecular physiology of P2X receptors. Physiol.Rev. 82, 1013–1067.
Novak A. and Dedhar S. (1999) Signaling through b-catenin and Lef/Tcf. Cell. Mol. Life Sci. 56, 532–537.
Pap M. and Cooper G. M. (1998) Role of glycogen synthase kinase-3 inthe phosphotidylinositol 3-kinase/Akt cell survival pathway.J. Biol. Chem. 273, 19929–19932.
Papkoff J. (1996) Regulation of complexed and free catenin pools bydistinct mechanisms. J. Biol. Chem. 272, 4536–4543.
Papkoff J. and Aikawa M. (1998) Wnt and HGF regulate GSK3 betaactivity and beta-catenin signalling in mammary epithelial cells.Biochem. Biophys. Res Comm. 247, 851–858.
Playford M. P., Bicknell D., Bodmer W. F. and Macaulay V. M. (2000)Insulin-like growth factor 1 regulates the location, stability, andtranscriptional activity of b-catenin. Proc. Natl Acad. Sci. 97,12103–12108.
Pons S., Trejo J. L., Martınez-Morales J. R. and Martı E. (2001) Vi-tronectin regulates sonic hedgehog activity during cerebellum
development through CREB phosphorylation. Development 128,1481–1492.
Ralevic V. and Burnstock G. (1998) Receptors for purines and pyrimi-dines. Pharmacol. Rev. 50, 413–492.
Sayas C. L., Avila J. and Wandosell F. (2002) Regulation of neuronalcytoskeleton by lysophosphatidic acid: role of GSK-3. Biochim.Biophys. Acta 1582, 144–153.
Sellers L. A., Simon J., Lundahl T. S., Cousens D. J., Humphrey P. P. A.and Barnard E. (2001) Adenosine nucleotides acting at the humanP2Y1 receptor stimulate mitogen-activated protein kinases andinduce apoptosis. J. Biol. Chem. 276, 16379–16390.
Shao J., Jung C., Liu C. and Sheng H. (2005) Prostaglandin E2 stimu-lates the b-catenin/T cell factor-dependent transcription in coloncancer. J. Biol. Chem. 280, 26565–26572.
Shevtsov S. P., Haq S. and Force T. (2006) Activation of b-cateninsignalling pathways by classical G-protein-coupled receptors. CellCycle 5, 2295–2300.
Spychala J. and Kitajewski J. (2004) Wnt and b-catenin signalling targetthe expression of ecto-5¢-nucleotidase and increase extracellularadenosine generation. Exp. Cell Res. 296, 99–108.
Stambolic V. and Woodgett J. R. (1994) Mitogen inactivation of gly-cogen synthase kinase-3b in intact cells via serine 9 phosphory-lation. Biochem. J. 303, 701–704.
Subramaniam S., Strelau J. and Unsicker K. (2003) Growth differenti-ation factor-15 prevents low potassium-induced cell death of cer-ebellar granule neurons by differential regulation of Akt and ERKpathways. J. Biol. Chem. 278, 8904–8912.
Van Kolen K., Gilany K., Moens L., Esmans E. and Slegers H. (2006)P2Y12 receptor signalling towards PKB proceeds through IGF-Ireceptor cross-talk and requires activation of Src, Pyk2 and Rap1.Cell. Signal. 18, 1169–1181.
Vitolo O. V., Ciotti M. T., Galli C., Borsello T. and Calissano P. (1998)Adenosine and ADP prevent apoptosis in cultured rat cerebellargranule cells. Brain Res. 809, 297–301.
Yang M., Zhong W. W., Srivastava N., Slavin A., Yang J., Hoey T. andAn S. (2005) G protein-coupled lysophosphatidic acid receptorsstimulate proliferation of colon cancer cells through the b-cateninpathway. Proc. Natl Acad. Sci. 102, 6027–6032.
Zhang F. L., Luo L., Gustafson E. et al. (2001) ADP is the cognateligand for the orphan G-protein-coupled receptor SP1999. J. Biol.Chem. 276, 8608–8615.
Zhang F. L., Luo L., Gustafson E., Palmer K., Qiao X., Fan X., Yang S.,Laz T. Z., Bayne M. and Monsma F. J. Jr (2002) P2Y13: iden-tification and characterization of a novel Gai-coupled ADPreceptor from human and mouse. J. Pharmacol. Exp. Ther. 301,705–713.
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 62–73
ADP signals to GSK-3 and b-catenin in granule neurons | 73




















