
Toluene inhalation modulates dentate gyrus granule cell output in vivo

NRC
STa
FMMb
Aoe(pufefwrh�mOsissaftwnm�vrgeP
Kf
*EACDsltw[5
Neuroscience 127 (2004) 53–61
0d
ICOTINIC CHOLINERGIC STIMULATION PROMOTES SURVIVAL ANDEDUCES MOTILITY OF CULTURED RAT CEREBELLAR GRANULE
ELLSNatntrM2pvreht(1msebmFssn2
ironiifTpeKrieeedtp
. FUCILE,a* M. RENZI,a C. LAURO,a C. LIMATOLA,a
. CIOTTIb AND F. EUSEBIa
Istituto Pasteur Fondazione Cenci-Bolognetti and Dipartimento diisiologia Umana e Farmacologia, Centro di Eccellenza Biologia eedicina Molecolare, Universita di Roma “La Sapienza,” P. le Aldooro 5, I-00185 Roma, Italy
Istituto di Neurobiologia, CNR, Viale Marx 43-15, I-00137 Roma, Italy
bstract—Despite many studies on the functional expressionf neuronal nicotinic acetylcholine receptors (nAChRs), anxhaustive description of the long-term effects of nicotineNic) stimulation in cerebellar granules is still far to be com-leted. For this reason, we addressed the experiments stim-lating cultured cerebellar granule neurons (CGN) with Nic,ocusing on the effects on cell motility and survival. Usinglectrophysiological and Ca2�-fluorescence techniques, weound a subset of rat CGN that responded to Nic by inwardhole cell currents and by short-delay Ca2� transients. These
esponses were mediated through both homomeric andeteromeric nAChRs, as assessed by their sensitivity to-bungarotoxin (�-BTX), dihydro-�-erythroidine (DH�E),ethyllicaconitine (MLA) and 5-hydroxyindole (5OH-indole).nce established the expression of �-BTX-sensitive and in-ensitive nAChRs and their ability to trigger Ca2� responses
n CGN, we aimed at investigating their possible role on cellurvival and motility. We demonstrate that Nic stimulationignificantly increases the survival of CGN exposed to thepoptosis-promoting low K� medium. This anti-apoptotic ef-ect is likely mediated through �7* nAChRs since we foundhat it was mimicked by choline, was insensitive to DH�E andas fully inhibited by �-BTX. Furthermore, we report that Nicegatively modulates CGN motility, reducing the basal cellovement through a pored membrane by the activation of-BTX-insensitive nAChRs. We conclude that CGN expressarious types of nAChRs, which are differently involved inegulating Nic-mediated modulation of cell survival and mi-ration, and we suggest potential regulatory roles for cholin-rgic receptors during cerebellar development. © 2004 IBRO.ublished by Elsevier Ltd. All rights reserved.
ey words: neuroprotection, migration, 5-hydroxyindole,ura-2, homomeric nAChR, heteromeric nAChR.
Corresponding author: Tel: �39-06-4991-0971; fax: �39-06-4991-0851.-mail address: [email protected] (S. Fucile).bbreviations: BME, basal Eagle’s medium; �-BTX, �-bungarotoxin;GN, cerebellar granule neurons; DH�E, dihydro-�-erythroidine;MEM, Dulbecco’s Modified Eagle’s medium; FBS, fetal bovineerum; MLA, methyllicaconitine; nAChR, neuronal nicotinic acetylcho-ine receptor; Nic, nicotine; P/S, penicillin/streptomycin; R, ratio be-ween fluorescence images obtained at 340 nm and 380 nm excitationavelengths; �R, difference between peak and basal R values;
Ca2�]i, intracellular free Ca2� concentration; TTX, tetrodotoxin;
mOH-indole, 5-hydroxyindole.306-4522/04$30.00�0.00 © 2004 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2004.04.017
53
euronal nicotinic acetylcholine receptors (nAChRs)re pentameric receptor-channels widely expressed
hroughout the vertebrate CNS, where they modulateeurotransmitter release and mediate fast synaptic
ransmission by acting at pre- and post-synaptic sites,espectively (reviewed by McGehee and Role, 1996;acDermott et al., 1999; Vizi and Lendvai 1999; Dani,001). Many studies focused on the expression and thehysiological role of nAChRs in the cerebellum (for re-iew see Jaarsma et al., 1997). Cerebellar granule neu-ons (CGN), the most represented neurons in brain,xpress many nAChR subunits, including those formingeteromeric receptors like �3, �4, �5, �2 and �4, andhe �7, that mainly constitutes homomeric receptorsDidier et al., 1995; Zoli et al., 1995; Caruncho et al.,997; Graham et al., 2002). nAChR subunit expressionay change throughout cerebellar development: for in-
tance, cultured CGN obtained from 7-day-old (p7) ratsxpressed the �7 subunit both at neurites and cellodies (Didier et al., 1995); while CGN from adult ani-als lack somatic �7 subunits (Caruncho et al., 1997).urthermore, nAChRs are localized at pre- and post-ynaptic sites of granule cells at early developmentaltages (p5–p14), modulating both synaptogenesis andeurogenesis (Bianchi et al., 2000; De Filippi et al.,001; Kawa, 2002).
It is well established that the activation of nAChRsnduces Ca2� permeation through the ion pore (see foreview Fucile, 2004). Specifically, nicotine (Nic) stimulationf CGN promotes Ca2� influx (Didier et al., 1995), andeurotransmitter release (Bianchi et al., 2000). However, it
s not yet established whether, in addition to their functionn synaptic transmission, nAChRs may play a role on otherunctional events such as CGN survival and migration.his is an important issue, since nAChRs regulate bothroliferation and survival of CGN progenitors (Opanashukt al., 2001), similarly to GABAA receptors (Owens andriegstein, 2002), pointing to a key role of neurotransmitter
eceptors during cerebellar development. To address thisssue, we made experiments to establish the functionalxpression of nAChR subtypes, and their efficacy in influ-ncing the survival and migration of CGN. Specifically,lectrophysiological recordings and Ca2�-fluorescenceeterminations were performed upon acute Nic stimula-ion, in order to establish nAChR functional expression; inarallel experiments, the regulation of CGN survival and
igration was assayed upon long-lasting Nic stimulation.ved.

C
P((bwEosEfccgcT1
C
TiECccFww
F
Fflbifr3dtaFccflcw
E
A2ic(MMswImutmps
S
C(tk1((n2d
M
Camg1lnlArtsavCo
S
Derdrw[ac
S
Ws2w1cPUma
N
Tp(Cst
S. Fucile et al. / Neuroscience 127 (2004) 53–6154
EXPERIMENTAL PROCEDURES
GN
rimary cultures of CGN were obtained from p8 Wistar ratsCharles River, Como, Italy) according to the method of Levi et al.1989). Experiments were designed in order to minimize the num-er and the suffering of animals used. Rats were anesthetizedith Fluothane and killed by decapitation, in agreement with theuropean Directive 86/609/EEC requirements and the guidelinesf the Ethical Committee of the Medical Faculty of Rome Univer-ity La Sapienza. Cells were cultured (5% CO2, 37 °C) in basalagle’s medium (BME), supplemented with: 10% heat-inactivated
etal bovine serum (FBS); 2 mM L-glutamine; penicillin/streptomy-in (P/S); and 25 mM KCl. Cells were plated onto poly-L-lysine-oated Petri dishes at a density of 2.5�105 cells/cm2. To preventlial cell proliferation, 20 h after plating, cultures were treated withytosine-�-d-arabinofuranoside at a final concentration of 10 �M.hese cultures were used 7–8 days after plating (Levi et al.,989).
ell lines
he human retroviral packaging cell line HEK 293 stably express-ng the human �4�2 nAChRs was grown in Dulbecco’s Modifiedagle’s medium (DMEM; with Glutamax-I; �10% FBS; �P/S; 5%O2, 37 °C) supplemented with geneticin. The rat anterior pituitaryell line GH4C1 stably expressing the human �7 nAChRs wasultured in F-10 (HAM) nutrient mixture (with L-glutamine; �10%BS; �P/S; 5% CO2, 37 °C) supplemented with geneticin. Cellsere plated onto Petri dishes at a density of 104 cells/cm2 andere used for electrophysiological recording 24–72 h after plating.
luorescence measurements
luorescence determinations were made using a conventionaluorescence microscopy system driven by Axon Imaging Work-ench software (Axon Instruments, Union City, CA, USA) detailed
n a previous work (Fucile et al., 2000). The level of intracellularree Ca2� concentration ([Ca2�]i) was expressed as the time-esolved ratio, R, between fluorescence images obtained at40 nm and 380 nm excitation wavelengths, and averaged from aomain approximating the cell shape. The amplitude of [Ca2�]iransients (�R) was measured as the difference between peaknd basal R values. The experimental procedures used are as inucile et al. (2000). Briefly, cells were incubated (37 °C) with theell membrane permeant fura-2 AM (4 �M) for 45 min in theulture medium. Responsive cells, identified as those generatinguorescence transient to a brief agonist application, were whole-ell patch clamped. Fluorescence determinations were carried outhen the basal fluorescence was maintained.
lectrophysiological recordings
Ch- or Nic-induced whole-cell currents were recorded at 23–5 °C from cells voltage-clamped at �70 mV. Unless otherwise
ndicated, the Nic concentration routinely used was 50 �M. Re-ordings were performed using borosilicate glass patch pipettesHilgenberg, Malsfeld, Germany) having a tip resistance of 3–5�. Whole-cell capacitance and patch series resistances (5–15�) were estimated from slow transient compensations, with a
eries resistance compensation of 70–85%. Membrane currentsere filtered at 2 kHz on acquisition with an Axopatch 200A (Axon
nstruments) and analyzed using pClamp8 software (Axon Instru-ents). During recordings, cells were continuously superfusedsing a gravity-driven perfusion system consisting of independentubes for normal and drug-containing external solutions. The ter-inals of the tubes were positioned 50–100 �m away from theatched cell and connected, on the other side, to a fast exchanger
ystem (RSC-100; Bio-Logic, Claix, France). burvival assay
GN (5�105) were seeded on poly-L-lysine-coated plastic 24-well15.5-mm) dishes. After 6 days of culture the cells were switchedo serum-free BME containing 5 mM K� to induce apoptosis orept in 25 mM K� medium for control, as described (D’Mello et al.,993); cells switched to low K� medium were treated with Nic1 �M), in the presence or in the absence of �-bungarotoxin�-BTX; 100 nM) or dihydro-�-erythroidine (DH�E; 50 �M). Alter-atively, cells were treated with choline chloride (200 �M). After4 h, the cells were counted in a hemocytometer for viability, asescribed (Volonte et al., 1994).
otility assay
erebellar neurons were obtained from rats, as described above,nd immediately plated in DMEM and HAM’s 12 (3:1), supple-ented with insulin, putrescine, transferrin, Na�-selenite and pro-esterone (Bottenstein et al., 1979), onto polylysine-treated2 mm transwells (3 �m pore size filters, 500,000 cells/well). The
ower chambers contained vehicle (water) or the cholinergic ago-ist studied (Nic 1 �M or choline 200 �M). When necessary, the
ower chamber also contained DH�E (50 �M) or �-BTX (100 nM).fter 2 h incubation at 37 °C, cells were treated with 10% trichlo-
oacetic acid on ice for 5 min. Cells adhering to the upper side ofhe filter were scraped off, while cells on the lower side weretained with a solution containing 50% isopropanol, 1% formiccid and 0.5% (w/v) Brilliant Blue R 250. Stained cells wereisually counted in more than 20 fields with a 40� objective.hemotactic index is obtained by the ratio between the numbersf migrated cells in treated vs untreated samples.
tatistical analysis
ata are reported as meansS.E.M. Two data sets were consid-red statistically different with P0.05 Student’s t-test. Dose-esponse curves were constructed by fitting values obtained atifferent concentrations, after normalization. The non-linear fittingoutine of Sigma Plot software (Systat, Point Richmond, CA, USA)as used to fit the data to the Hill-type equation: I�1/(1�(EC50/
A])nH, where I is the normalized current amplitude induced by thegonist at concentration [A]; nH, the Hill coefficient; and EC50 theoncentration at which a half-maximum response was induced.
olutions and chemicals
hole-cell patch-clamped cells were superfused with an externalolution containing (in mM): NaCl, 140; KCl, 2.8; CaCl2, 2; MgCl2,; HEPES-NaOH, 10; glucose, 10 (pH 7.3). The patch pipettesere filled with a solution containing (in mM): 140 CsCl, 2 MgATP,0 HEPES-CsOH and 5 BAPTA (pH 7.3). Chemicals were pur-hased from Sigma (USA), except fura-2 AM from Molecularrobes (Eugene, OR, USA), DH�E from RBI/Sigma (Natick, MA,SA) and tetrodotoxin (TTX) from Tocris (Bristol, UK). Cultureedia and FBS were purchased from Gibco (USA). L-Glutaminend P/S were purchased from Euroclone (UK).
RESULTS
ic-induced inward currents and Ca2� transients
he effects of Nic on CGN were first studied using theatch-clamp technique, in the whole-cell configuration. Nic200 �M) elicited inward currents in the 30% of the testedGN (12 out 40). Nic-induced currents were divided in twoubtypes, depending on their sensitivity to methyllicaconi-ine (MLA) at the concentration of 1 nM, at which MLA
ehaves as selective antagonist of �7-containing nAChRs
(Fp(c
smtosaripTi00[ct
w(iRrstitiwnefltac
bwdltmvfsewb1vm1ont1mBnCemt
Nb
Tcc2vsentt
FWt(ibciapctroin
S. Fucile et al. / Neuroscience 127 (2004) 53–61 55
Brioni et al., 1997). The MLA-insensitive CGN (5 out 40;ig. 1A) exhibited a mean current amplitude of �5843A, while the current amplitude of the MLA-sensitive cellsfive out 40; Fig. 1B) was much lower, �71 pA. Two moreells exhibited both current types (Fig. 1C).
In order to better characterize the cholinergic re-ponses, we used the Ca2� imaging analysis, quicklyonitoring a high number of cells. Specifically, we inves-
igated the effects of nAChR activation by Nic on the levelf [Ca2�]i, from fura-2-loaded CGN, and found that a smallubset of cells responded to Nic (50 �M) application withn elevation of [Ca2�]i, localized in the cell bodies (56esponsive cells from 313 optic field representing approx-mately 0.2% of total cells; see Fig. 2A) and/or in therocesses (63 responsive cells from same optic fields).he �R values, used to estimate the amplitudes of Nic-
nduced [Ca2�]i transients, exhibited a mean value of.360.05 (range: 0.02–0.95) in CGN cell bodies, and of.180.02 in the processes. Interestingly, the kinetics ofCa2�]i rise was different between cell bodies and pro-esses: the mean rise time (i.e. the mean time from basal
2�
ig. 1. Typical inward currents elicited by Nic in cultured CGN. (A)hole-cell inward currents recorded from a single CGN (representa-
ive of five out of total 40 cells) in the absence (left) or in the presenceright) of the competitive �7-nAChR antagonist MLA (1 nM). Note MLAnefficacy, indicative of non-�7 nAChRs activation. Horizontal blackars in here and below represent Nic applications, at the indicatedoncentration. Horizontal open bars represent MLA applications, at thendicated concentration. (B) Whole-cell inward currents recorded fromnother CGN (representative of five cells) in the absence (left) or in theresence (right) of MLA. Note both the efficacy of MLA and the fasturrent desensitization, indicative of �7* nAChRs activation. (C) Mul-iphasic whole-cell inward current recorded from a different CGN (rep-esentative of two cells) in the absence (left) or in the presence (right)f MLA. Note the MLA-induced suppression of the fast component,
ndicative of both �7* and non-�7 nAChRs activation in the sameeuron.
o peak value) of the [Ca ]i in the soma was 4.040.12 s, t
hile in cell processes it was significantly shorter1.740.12 s, e.g. Fig. 2A), indicating that Ca2� flowednto the cell somata over the whole nAChR activation.epetitive applications of Nic induced a rundown of Ca2�
esponses (Fig. 2B; n�8) that could be due to many rea-ons including nAChR desensitization, Ca2� store deple-ion and dye bleaching. Treatment with TTX (1 �M) did notnhibit the Ca2� transients, indicating that the action po-ential propagation did not modulate Nic-induced Ca2�
nflow (n�6; not shown). Nic-induced Ca2� transientsere strongly reduced in Ca2�-free medium (Fig. 2C;�5), both in soma and processes, showing that [Ca2�]ilevation was mainly due to Ca2� entry from extracellularuid rather than to cytosolic Ca2� mobilization. Consideredogether, these data indicate that functional Ca2� perme-ble nAChRs are expressed in both cell body and pro-esses of CGN.
In further experiments we asked whether the low num-er of CGN exhibiting Ca2� transients (0.2%), comparedith those displaying Nic-induced currents (30%), could beue to changes in [Ca2�]i detectable only in CGN with
arge nicotinic currents. Actually, all CGN that respondedo Nic by Ca2� transients also displayed currents withean amplitude of �18238 pA (n�19; see Fig. 3), a
alue more than three-fold larger than the mean currentrom CGN randomly patched. The decay of these currents,elected through Ca2� transients, was fitted to a singlexponential, with a mean time constant of 1.770.22 s;hile the dose-response relation (see Fig. 3A; n�4) wasest fitted to a Hill equation, giving an EC50 value of1.50.1 �M, and a nH value of 2.20.1. Both these latteralues were compatible with an apparent affinity of hetero-eric rather than of homomeric receptors (Brioni et al.,997). I–V relations showed that Nic-currents exhibited anbvious inward rectification (Fig. 3B). Interestingly, the �7AChR antagonist �-BTX (100 nM) was unable to inhibithe Nic-elicited currents and, in all CGN (n�6) treated for
h with �-BTX, the current amplitude and kinetics re-ained stable compared with untreated CGN (Fig. 3A, C).y contrast, the competitive inhibitor of the heteromericAChRs, DH�E (50 �M), blocked Nic-currents (Fig. 3C).onsidered together, these findings indicate that CGNxpress �-BTX-insensitive, non-�7 containing, hetero-eric nAChRs identified by detecting Nic-induced Ca2�
ransients.
ic-induced currents and Ca2� transients modulatedy 5-hydroxyindole (5OH-indole)
he amplitude of Ca2� transients measured using fluores-ence microscopy depends on the time integral of Ca2�
urrent, i.e. Ca2� charge entering the cells (Fucile et al.,003). Since Nic-induced currents generated by the acti-ation of homomeric �7 nAChRs desensitize quickly (Fen-ter et al., 1997), it would be possible that Ca2� transientslicited by the activation of �7* nAChRs (i.e. �7-containingAChRs; for nomenclature see Lukas et al., 1999) were
oo small and too fast to be detected under our experimen-al conditions. To address this issue, we thought useful to
est higher Nic concentrations (300–500 �M) in CGN
tbbttas(dt5(apicG4b�p
I
LraS1ewbdtd(ascdtp5C�sao(1ln
(tirN
Fr(tfloet
S. Fucile et al. / Neuroscience 127 (2004) 53–6156
reated with 5OH-indole, an analog of serotonin, which haseen elsewhere shown to enhance the currents generatedy the activation of �7 nAChR, decreasing its desensitiza-ion (Zwart et al., 2002). We found that 5OH-indole poten-iated Nic-mediated Ca2� transients only in a low percent-ge of CGN cell bodies (3 cells from 54 optic fields repre-enting the 0.06% of total cells; Fig. 4A) and processestwo in the same fields). The remaining Nic-sensitive CGNisplayed a marked impairment of Ca2� transients ampli-ude upon 5OH-indole treatment (e.g. Fig. 4B). Invariably,OH-indole treatment potentiated or inhibited Nic-currentsFig. 4C, D). In order to clarify these effects, we tested thection of 5OH-indole on heteromeric �4�2 nAChRs ex-ressed in HEK293 cells and on homomeric �7 expressed
n GH4C1 cells. Interestingly, upon 5OH-indole treatment,holinergic whole-cell currents were enhanced in �7-H4C1 cells (Fig. 4E), but reduced in �4�2-HEK cells (Fig.F). These findings suggest that the currents potentiatedy 5OH-indole in CGN are originated by the activation of7* nAChRs while those inhibited by 5OH-indole are cou-led to heteromeric nAChRs.
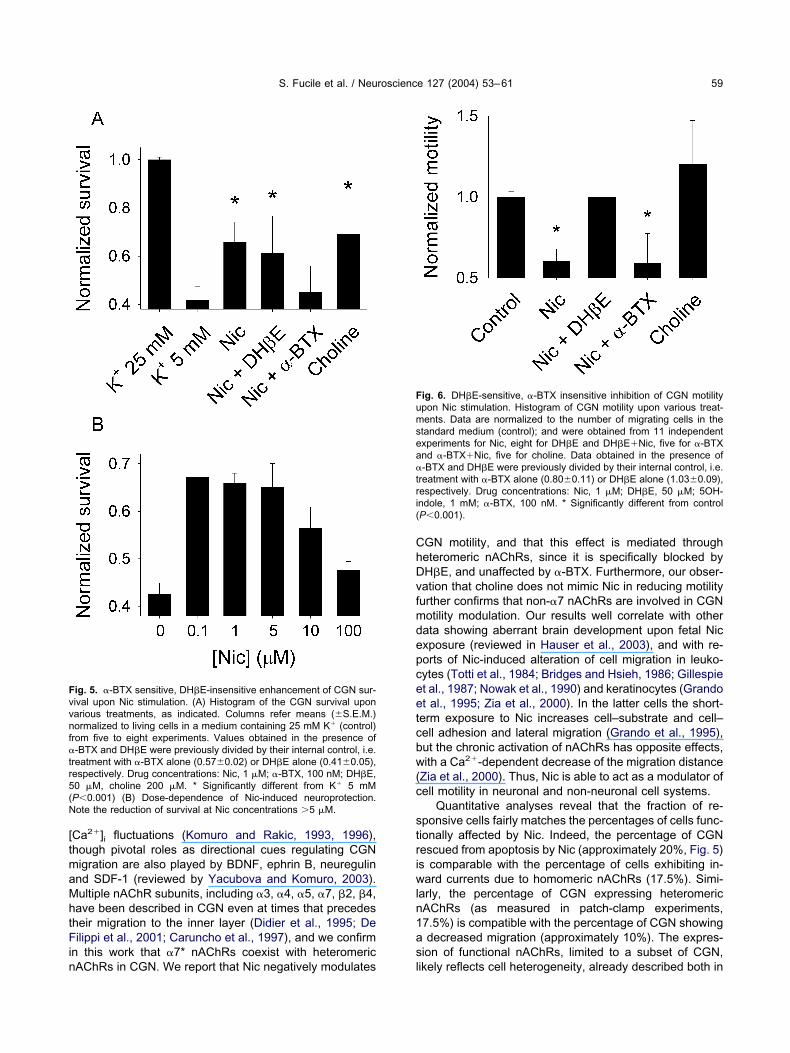
ncrease of CGN survival mediated by �7* nAChRs
ow concentrations of Nic have been proved to exert neu-oprotective activities in several experimental paradigms,nd in different neuronal systems (Messi et al., 1997;himohama et al., 1998; Kihara et al., 1997; Kaneko et al.,997; Li et al., 1999, 2000; Belluardo et al., 2000; Stevenst al., 2003). In this study we were interested to seehether Nic could protect CGN from cellular death inducedy K� deprivation, a classical model of apoptotic neuraleath (Gallo et al., 1987). As expected, when the concen-ration of K� in medium was reduced to 5 mM, a markedrop in the number of living cells was observed, after 24 hFig. 5A). When Nic at 0.1–5 �M concentrations wasdded to 5 mM K� containing medium, cell death wasignificantly inhibited (Fig. 5B), while relatively high con-entrations of Nic (100 �M) failed to protect CGN fromeath (Fig. 5B), likely due to increased receptor desensi-
ization. The neurotrophic effect of Nic was blocked by theresence of �-BTX (100 nM), and not of DH�E 50 �M (Fig.A), strongly suggesting the involvement of �7* nAChRs inGN neuroprotection. Interestingly, CGN treatment with-BTX alone induced a slight but significant increase in cellurvival (see legend of Fig. 5), suggesting that both �-BTXnd Nic effects might be due, at least in part, to receptorccupancy rather than to channel opening. Choline200 �M), an agonist of �7* nAChRs (Alkondon et al.,997), induced a significant increase in the percentage of
ive cells (Fig. 5A), supporting the involvement of �7*AChRs in Nic-induced neuroprotection.
C) Typical fluorescence transients elicited by repetitive Nic applica-ions performed in the presence or absence of extracellular Ca2�, asndicated. Nic applications, every 5 min. In eight similar experiments,esponses in Ca2�-free medium were reduced to 33% of control.
2�
ote recovery after restoring normal extracellular Ca concentration.ig. 2. [Ca2�]i transients induced by Nic in cultured CGN. (A) Fluo-escence changes (top) recorded from a CGN upon application of Nic50 �M, horizontal bar). Note the different kinetics of fluorescenceransients between cell body and process (as indicated). (B) Typicaluorescence transients elicited by repetitive Nic applications to an-ther CGN as in (A). Nic applications, every 5 min. In eight similarxperiments, the responses to the second and the third Nic applica-ions were reduced to 7214% and 4524% of control, respectively.
In
Ict1CtPciceb(msm
I�tNNtsn
eeeeBmwTCarasktdan(2ooChann
F((ic
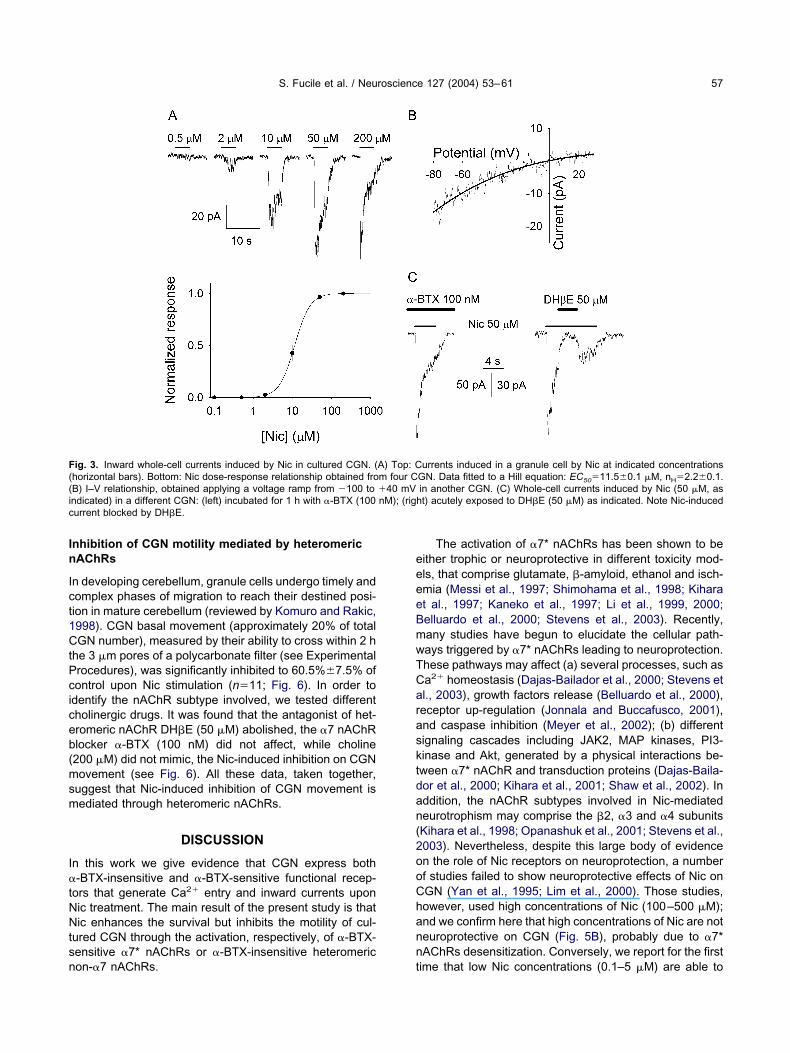
S. Fucile et al. / Neuroscience 127 (2004) 53–61 57
nhibition of CGN motility mediated by heteromericAChRs
n developing cerebellum, granule cells undergo timely andomplex phases of migration to reach their destined posi-ion in mature cerebellum (reviewed by Komuro and Rakic,998). CGN basal movement (approximately 20% of totalGN number), measured by their ability to cross within 2 h
he 3 �m pores of a polycarbonate filter (see Experimentalrocedures), was significantly inhibited to 60.5%7.5% ofontrol upon Nic stimulation (n�11; Fig. 6). In order todentify the nAChR subtype involved, we tested differentholinergic drugs. It was found that the antagonist of het-romeric nAChR DH�E (50 �M) abolished, the �7 nAChRlocker �-BTX (100 nM) did not affect, while choline200 �M) did not mimic, the Nic-induced inhibition on CGNovement (see Fig. 6). All these data, taken together,
uggest that Nic-induced inhibition of CGN movement isediated through heteromeric nAChRs.
DISCUSSION
n this work we give evidence that CGN express both-BTX-insensitive and �-BTX-sensitive functional recep-
ors that generate Ca2� entry and inward currents uponic treatment. The main result of the present study is thatic enhances the survival but inhibits the motility of cul-
ured CGN through the activation, respectively, of �-BTX-ensitive �7* nAChRs or �-BTX-insensitive heteromeric
ig. 3. Inward whole-cell currents induced by Nic in cultured CGN. (horizontal bars). Bottom: Nic dose-response relationship obtained froB) I–V relationship, obtained applying a voltage ramp from �100 tondicated) in a different CGN: (left) incubated for 1 h with �-BTX (100urrent blocked by DH�E.
on-�7 nAChRs. t
The activation of �7* nAChRs has been shown to beither trophic or neuroprotective in different toxicity mod-ls, that comprise glutamate, �-amyloid, ethanol and isch-mia (Messi et al., 1997; Shimohama et al., 1998; Kiharat al., 1997; Kaneko et al., 1997; Li et al., 1999, 2000;elluardo et al., 2000; Stevens et al., 2003). Recently,any studies have begun to elucidate the cellular path-ays triggered by �7* nAChRs leading to neuroprotection.hese pathways may affect (a) several processes, such asa2� homeostasis (Dajas-Bailador et al., 2000; Stevens etl., 2003), growth factors release (Belluardo et al., 2000),eceptor up-regulation (Jonnala and Buccafusco, 2001),nd caspase inhibition (Meyer et al., 2002); (b) differentignaling cascades including JAK2, MAP kinases, PI3-inase and Akt, generated by a physical interactions be-ween �7* nAChR and transduction proteins (Dajas-Baila-or et al., 2000; Kihara et al., 2001; Shaw et al., 2002). Inddition, the nAChR subtypes involved in Nic-mediatedeurotrophism may comprise the �2, �3 and �4 subunitsKihara et al., 1998; Opanashuk et al., 2001; Stevens et al.,003). Nevertheless, despite this large body of evidencen the role of Nic receptors on neuroprotection, a numberf studies failed to show neuroprotective effects of Nic onGN (Yan et al., 1995; Lim et al., 2000). Those studies,owever, used high concentrations of Nic (100–500 �M);nd we confirm here that high concentrations of Nic are noteuroprotective on CGN (Fig. 5B), probably due to �7*AChRs desensitization. Conversely, we report for the first
urrents induced in a granule cell by Nic at indicated concentrationsGN. Data fitted to a Hill equation: EC50�11.50.1 �M, nH�2.20.1.in another CGN. (C) Whole-cell currents induced by Nic (50 �M, as
ht) acutely exposed to DH�E (50 �M) as indicated. Note Nic-induced
A) Top: Cm four C�40 mVnM); (rig
ime that low Nic concentrations (0.1–5 �M) are able to

pt�m�padibc
tnmsp
wes(
F1i5Naie
S. Fucile et al. / Neuroscience 127 (2004) 53–6158
artially rescue CGN from apoptosis caused by exposureo low K� medium. This effect, which is eliminated by-BTX cell treatment and unaffected by DH�E, is likelyediated through �7* nAChRs, which are blocked by-BTX and insensitive to DH�E. This view is further sup-orted by cell treatment with choline, a rather selectivegonist of �7* nAChRs (Alkondon et al., 1997), that in-uced neuroprotective effect comparable to Nic. Interest-
ngly, we report also that �-BTX itself promotes a limitedut significant enhancement of CGN survival, suggesting a
ig. 4. Effects of 5OH-indole on nicotinic responses in cultured CGN.mM 5OH-indole administration (as indicated). Typical of three resp
ndicated). Typical of six responses. (C) Whole-cell current elicited bOH-indole (1 mM), as indicated (middle); and 3 min after 5OH-indole wote reversible current potentiation upon 5OH-indole treatment. (D) Wpplied as in A. (A–D) Different CGN. (E) Whole-cell current generated
n a stably transfected GH4C1 cell and enhanced by 5OH-indole, appxpressed in a HEK293 cell, and activated by Nic (2 �M, horizontal b
ontribution to neuroprotection due to receptor occupancy, m
riggering signaling cascade (Kihara et al., 2001). A similareuroprotective action obtained with low doses of Nic andediated mainly through �3* nAChRs has been elsewhere
hown for external granule layer neuroblasts, the CGNrecursors (Opanashuk et al., 2001).
During development, CGN migrate first tangentiallyithin the external granular layer and then radially from thexternal to the inner granular layer, on the basis of multipleignaling mechanisms and external guidance cuesYacubova and Komuro, 2003). The modulation of CGN
]i transients elicited by Nic (500 �M, horizontal bar) potentiated uponB) Nic-induced [Ca2�]i transients inhibited by 1 mM 5OH-indole (as00 �M, horizontal bar) in control condition (left); in the presence ofight). 5OH-indole administration started 4 s before agonist application.current elicited by Nic (50 �M, horizontal bar) inhibited by 5OH-indolectivation of �7 nAChRs with ACh (100 �M, horizontal bars), recordedA, B. (F) Whole-cell current mediated through �4�2 nAChRs, stablycurrent reduction with 5OH-indole administration.
(A) [Ca2�
onses. (y Nic (5ashout (rhole-cellby the a
lied as inar). Note
ovement is mainly due to neurotransmitter-induced
[tmaMhtFin
ChDvfmdepceetcbw(c
striwln1as
Fvvnf�tr5(N
Fumsea�tri(
S. Fucile et al. / Neuroscience 127 (2004) 53–61 59
Ca2�]i fluctuations (Komuro and Rakic, 1993, 1996),hough pivotal roles as directional cues regulating CGNigration are also played by BDNF, ephrin B, neuregulinnd SDF-1 (reviewed by Yacubova and Komuro, 2003).ultiple nAChR subunits, including �3, �4, �5, �7, �2, �4,ave been described in CGN even at times that precedesheir migration to the inner layer (Didier et al., 1995; Deilippi et al., 2001; Caruncho et al., 1997), and we confirm
n this work that �7* nAChRs coexist with heteromeric
ig. 5. �-BTX sensitive, DH�E-insensitive enhancement of CGN sur-ival upon Nic stimulation. (A) Histogram of the CGN survival uponarious treatments, as indicated. Columns refer means (S.E.M.)ormalized to living cells in a medium containing 25 mM K� (control)rom five to eight experiments. Values obtained in the presence of-BTX and DH�E were previously divided by their internal control, i.e.
reatment with �-BTX alone (0.570.02) or DH�E alone (0.410.05),espectively. Drug concentrations: Nic, 1 �M; �-BTX, 100 nM; DH�E,0 �M, choline 200 �M. * Significantly different from K� 5 mMP0.001) (B) Dose-dependence of Nic-induced neuroprotection.ote the reduction of survival at Nic concentrations �5 �M.
AChRs in CGN. We report that Nic negatively modulates l
GN motility, and that this effect is mediated througheteromeric nAChRs, since it is specifically blocked byH�E, and unaffected by �-BTX. Furthermore, our obser-ation that choline does not mimic Nic in reducing motilityurther confirms that non-�7 nAChRs are involved in CGNotility modulation. Our results well correlate with otherata showing aberrant brain development upon fetal Nicxposure (reviewed in Hauser et al., 2003), and with re-orts of Nic-induced alteration of cell migration in leuko-ytes (Totti et al., 1984; Bridges and Hsieh, 1986; Gillespiet al., 1987; Nowak et al., 1990) and keratinocytes (Grandot al., 1995; Zia et al., 2000). In the latter cells the short-erm exposure to Nic increases cell–substrate and cell–ell adhesion and lateral migration (Grando et al., 1995),ut the chronic activation of nAChRs has opposite effects,ith a Ca2�-dependent decrease of the migration distance
Zia et al., 2000). Thus, Nic is able to act as a modulator ofell motility in neuronal and non-neuronal cell systems.
Quantitative analyses reveal that the fraction of re-ponsive cells fairly matches the percentages of cells func-ionally affected by Nic. Indeed, the percentage of CGNescued from apoptosis by Nic (approximately 20%, Fig. 5)s comparable with the percentage of cells exhibiting in-ard currents due to homomeric nAChRs (17.5%). Simi-
arly, the percentage of CGN expressing heteromericAChRs (as measured in patch-clamp experiments,7.5%) is compatible with the percentage of CGN showingdecreased migration (approximately 10%). The expres-
ion of functional nAChRs, limited to a subset of CGN,
ig. 6. DH�E-sensitive, �-BTX insensitive inhibition of CGN motilitypon Nic stimulation. Histogram of CGN motility upon various treat-ents. Data are normalized to the number of migrating cells in the
tandard medium (control); and were obtained from 11 independentxperiments for Nic, eight for DH�E and DH�E�Nic, five for �-BTXnd �-BTX�Nic, five for choline. Data obtained in the presence of-BTX and DH�E were previously divided by their internal control, i.e.
reatment with �-BTX alone (0.800.11) or DH�E alone (1.030.09),espectively. Drug concentrations: Nic, 1 �M; DH�E, 50 �M; 5OH-ndole, 1 mM; �-BTX, 100 nM. * Significantly different from controlP0.001).
ikely reflects cell heterogeneity, already described both in

CeDG
nupsoL
Af
A
B
B
B
B
N
B
C
C
D
D
D
D
D
D
F
F
F
F
G
G
G
G
H
H
J
J
K
K
K
K
K
K
K
S. Fucile et al. / Neuroscience 127 (2004) 53–6160
GN cultures and slices, with physiological roles to belucidated (Didier et al., 1995; Caruncho et al., 1997;ominguez del Toro et al., 1997; De Filippi et al., 2001;raham et al., 2002; Kawa, 2002).
In conclusion, we report that CGN express both �7*AChRs and non-�7 heteromeric nAChRs: the latter mod-late neuronal motility while the first ones mediate neuro-rotective responses. Our data support the view that Nicignaling may play relevant roles during cerebellar devel-pment (Court et al., 1995; Hellstrom-Lindahl et al., 1998;ee et al., 2002).
cknowledgements—This work is supported by a co-fin grantrom MIUR and FIRB to F.E.
REFERENCES
lkondon M, Pereira EF, Cortes WS, Maelicke A, Albuquerque EX(1997) Choline is a selective agonist of �7 nicotinic acetylcholinereceptors in the rat brain neurons. Eur J Neurosci 9:2734–2742.
elluardo N, Mudo G, Blum M, Amato G, Fuxe K (2000) Neurotrophiceffects of central nicotinic receptor activation. J Neural Transm60:227–245.
ianchi C, Tomasini MC, Antonelli T, Marani L, Beani L (2000) Nico-tinic modulation of [(3)H]D-aspartate outflow from cultured cerebel-lar granule cells. Synapse 36:307–313.
ottenstein J, Hayashi I, Hutchings S, Masui H, Mather J, McClure DB,Ohasa S, Rizzino A, Sato G, Serrero G, Wolfe R, Wu R (1979) Thegrowth of cells in serum-free hormone-supplemented media. Meth-ods Enzymol 58:94–109.
ridges RB, Hsieh L (1986) Effects of cigarette smoke fractions on thechemotaxis of polymorphonuclear leukocytes. J Leukoc Biol 40:73–85.
owak D, Ruta U, Piasecka G (1990) Nicotine increases humanpolymorphonuclear leukocytes chemotactic response: a possibleadditional mechanism of lung injury in cigarette smokers. ExpPathol 39:37–43.
rioni JD, Decker MW, Sullivan JP, Arneric SP (1997) The pharma-cology of (�)-nicotine and novel cholinergic channel modulators.Adv Pharmacol 37:153–214.
aruncho HJ, Guidotti A, Lindstrom J, Costa E, Pesold C (1997)Subcellular localization of the �7 nicotinic receptor in rat cerebellargranule cell layer. Neuroreport 8:1431–1433.
ourt JA, Perry EK, Spurden D, Griffiths M, Kerwin JM, Morris CM,Johnson M, Oakley AE, Birdsall NJ, Clementi F, Perry RH (1995)The role of the cholinergic system in the development of the humancerebellum. Brain Res Dev Brain Res 90:159–167.
ajas-Bailador FA, Lima PA, Wonnacott S (2000) The �7 nicotinicacetylcholine receptor subtype mediates nicotine protection againstNMDA excitotoxicity in primary hippocampal cultures through aCa2� dependent mechanism. Neuropharmacology 39:2799–2807.
ani JA (2001) Overview of nicotinic receptors and their roles in thecentral nervous system. Biol Psychiatry 49:166–174.
e Filippi G, Baldwinson T, Sher E (2001) Evidence for nicotinicacetylcholine receptor activation in rat cerebellar slices. PharmacolBiochem Behav 70:447–455.
idier M, Berman SA, Lindstrom J, Bursztain S (1995) Characteriza-tion of nicotinic acetylcholine receptors expressed in primary cul-tures of cerebellar granule cells. Brain Res Mol Brain Res 30:17–28.
’Mello SR, Galli C, Ciotti T, Calissano P (1993) Induction of apoptosisin cerebellar granule neurons by low potassium: inhibition of deathby insulin-like growth factor I and cAMP. Proc Natl Acad Sci USA90:10989–10993.
ominguez del Toro E, Juiz JM, Smillie FI, Lindstrom J, Criado M
(1997) Expression of �7 neuronal nicotinic receptors during post- Knatal development of the rat cerebellum. Brain Res Dev Brain Res98:125–133.
enster CP, Rains MF, Noerager B, Quick MW, Lester RA (1997)Influence of subunit composition on desensitization of neuronalacetylcholine receptors at low concentrations of nicotine. J Neuro-sci 17:5747–5759.
ucile S (2004) Ca2� permeability of nicotinic acetylcholine receptors.Cell Calcium 35:1–8.
ucile S, Palma E, Mileo AM, Miledi R, Eusebi F (2000) Humanneuronal threonine-for-leucine-248 �7 mutant nicotinic acetylcho-line receptors are highly Ca2� permeable. Proc Natl Acad Sci USA97:3643–3648.
ucile S, Renzi M, Lax P, Eusebi F (2003) Fractional Ca2� currentthrough human neuronal alpha7 nicotinic acetylcholine receptors.Cell Calcium 34:205–209.
allo V, Suergiu R, Giovannini C, Levi G (1987) Glutamate receptorsubtypes in cultured cerebellar neurons: modulation of glutamateand gamma-aminobutyric acid release. J Neurochem 49:1801–1809.
illespie MN, Owasoyo JO, Kojima S, Jay M (1987) Enhanced che-motaxis and superoxide anion production by polymorphonuclearleukocytes from nicotine-treated and smoke-exposed rats. Toxicol-ogy 45:45–52.
raham A, Court JA, Martin-Ruiz CM, Jaros E, Perry R, Volsen SG,Bose S, Evans N, Ince P, Kuryatov A, Lindstrom J, Gotti C, PerryEK (2002) Immunohistochemical localisation of nicotinic acetylcho-line receptor subunits in human cerebellum. Neuroscience 113:493–507.
rando SA, Horton RM, Pereira EF, Diethelm-Okita BM, George PM,Albuquerque EX, Conti-Fine BM (1995) A nicotinic acetylcholinereceptor regulating cell adhesion and motility is expressed in hu-man keratinocytes. J Invest Dermatol 105:774–781.
auser KF, Khurdayan VK, Goody RJ, Nath A, Saria A, Pauly JR(2003) Selective vulnerability of cerebellar granule neuroblasts andtheir progeny to drugs with abuse liability. Cerebellum 2:184–195.
ellstrom-Lindahl E, Gorbounova O, Seiger A, Mousavi M, Nordberg A(1998) Regional distribution of nicotinic receptors during prenataldevelopment of human brain and spinal cord. Brain Res Dev BrainRes 108:147–160.
aarsma D, Ruigrok TJ, Caffe R, Cozzari C, Levey AI, Mugnaini E,Voogd J (1997) Cholinergic innervation and receptors in the cere-bellum. Prog Brain Res 114:67–96.
onnala RR, Buccafusco JJ (2001) Relationship between the in-creased cell surface �7 nicotinic receptor expression and neuro-protection induced by several nicotinic receptor agonists. J Neuro-sci Res 66:565–572.
aneko S, Maeda T, Kume T, Kochiyama H, Akaike A, Shimohama S,Kimura J (1997) Nicotine protects cultured cortical neurons againstglutamate-induced cytotoxicity via �7-neuronal receptors and neu-ronal CNS receptors. Brain Res 765:135–140.
awa K (2002) Acute synaptic modulation by nicotinic agonists indeveloping cerebellar Purkinje cells of the rat. J Physiol 538:87–102.
ihara T, Shimohama S, Sawada H, Kimura J, Kume T, Kochiyama H,Maeda T, Akaike A (1997) Nicotinic receptor stimulation protectsneurons against beta-amyloid toxicity. Ann Neurol 42:159–163.
ihara T, Shimohama S, Urushitani M, Sawada H, Kimura J, Kume T,Maeda T, Akaike A (1998) Stimulation of �4�2 nicotinic acetylcho-line receptors inhibits �-amyloid toxicity. Brain Res 792:331–334.
ihara T, Shimohama S, Sawada H, Honda K, Nakamizo T, ShibasakiH, Kume T, Akaike A (2001) �7 nicotinic receptor transducessignals to phosphatidylinositol 3-kinase to block A�-amyloid-in-duced neurotoxicity. J Biol Chem 276:13541–13546.
omuro H, Rakic P (1993) Modulation of neuronal migration by NMDAreceptors. Science 260:95–97.
omuro H, Rakic P (1996) Intracellular Ca2� fluctuations modulate therate of neuronal migration. Neuron 17:275–285.
omuro H, Rakic P (1998) Orchestration of neuronal migration by

L
L
L
L
L
L
M
M
M
M
O
O
S
S
S
T
V
V
Y
Y
Z
Z
Z
S. Fucile et al. / Neuroscience 127 (2004) 53–61 61
activity of ion channels, neurotransmitter receptors, and intracellu-lar Ca2� fluctuations. J Neurobiol 37:110–130.
ee M, Martin-Ruiz C, Graham A, Court J, Jaros E, Perry R, Iversen P,Bauman M, Perry E (2002) Nicotinic receptor abnormalities in thecerebellar cortex in autism. Brain 125:1483–1495.
evi G, Aloisi F, Ciotti MT, Thangnipon W, Kingsbury A, Balazs R(1989) Preparation of 98% pure cerebellar granule cell cultures. In:A dissection and tissue culture manual of the nervous system(Shahar A, de Vellis J, Vernadakis A, Haber B, Eds), pp 211–214.New York: Alan R. Liss Inc.
i Y, King MA, Meyer EM (2000) �7 nicotinic receptor-mediated pro-tection against ethanol-induced oxidative stress and cytotoxicity inPC12 cells. Brain Res 861:165–167.
i Y, Papke RL, He YJ, Millard WJ, Meyer EM (1999) Characterizationof the neuroprotective and toxic effects of �7 nicotinic receptoractivation in PC12 cells. Brain Res 830:218–225.
im DK, Park SH, Choi WJ (2000) Subacute nicotine exposure incultured cerebellar cells increased the release and uptake of glu-tamate. Arch Pharm Res 23:488–494.
ukas RJ, Changeux JP, Le Novere N, Albuquerque EX, Balfour DJK,Berg DK, Bertrand D, Chiappinelli VA, Clarke PBS, Collins AC, DaniJA, Grady SR, Kellar KJ, Lindstrom JM, Marks MJ, Quik M, TaylorPW, Wonnacott S (1999) International union of pharmacology: XX.Curent status of the nomenclature for nicotinic acetylcholine recep-tors and their subunits. Pharmacol Rev 51:397–401.
acDermott AB, Role LW, Siegelbaum SA (1999) Presynaptic iono-tropic receptors and the control of transmitter release. Annu RevNeurosci 22:443–485.
cGehee DS, Role LW (1996) Presynaptic ionotropic receptors. CurrOpin Neurobiol 6:342–349.
essi ML, Renganathan M, Grigorenko E, Delbono O (1997) Activa-tion of �7 nicotinic acetylcholine receptor promotes survival ofspinal cord motoneurons. FEBS Lett 411:32–38.
eyer EL, Gahring LC, Rogers SW (2002) Nicotine preconditioningantagonizes activity-dependent caspase proteolysis of a glutamatereceptor. J Biol Chem 277:10869–10875.
panashuk LA, Pauly JR, Hauser KF (2001) Effect of nicotine oncerebellar granule neuron development. Eur J Neurosci 13:48–56.
wens DF, Kriegstein AR (2002) Is there more to GABA than synaptic
inhibition? Nat Rev Neurosci 3:715–727.haw S, Bencherif M, Marrero MB (2002) Janus kinase 2, an earlytarget of �7 nicotinic acetylcholine receptor-mediated neuroprotec-tion against Abeta-(1–42) amyloid. J Biol Chem 277:44920–44924.
himohama S, Greenwald DL, Shafron DH, Akaika A, Maeda T,Kaneko S, Kimura J, Simpkins CE, Day AL, Meyer EM (1998)Nicotinic �7 receptors protect against glutamate neurotoxicity andneuronal ischemic damage. Brain Res 779:359–363.
tevens TR, Krueger SR, Fitzsimonds RM, Picciotto MR (2003) Neu-roprotection by nicotine in mouse primary cortical cultures involvesactivation of calcineurin and L-type calcium channel inactivation.J Neurosci 23:10093–10099.
otti N, McCusker KT, Campbell EJ, Griffin GL, Senior RM (1984)Nicotine is chemotactic for neutrophils and enhances neutrophilresponsiveness to chemotactic peptides. Science 223:169–171.
izi ES, Lendvai B (1999) Modulatory role of presynaptic nicotinicreceptors in synaptic and non-synaptic chemical communication inthe central nervous system. Brain Res Brain Res Rev 30:219–235.
olonte C, Ciotti MT, Battistini L (1994) Development of a method formeasuring cell number: application to CNS primary neuronal cul-tures. Cytometry 17:274–276.
acubova E, Komuro H (2003) Cellular and molecular mechanisms ofcerebellar granule cell migration. Cell Biochem Biophys 37:213–234.
an GM, Lin SZ, Irwin RP, Paul SM (1995) Activation of muscariniccholinergic receptors blocks apoptosis of cultured cerebellar gran-ule neurons. Mol Pharmacol 47:248–257.
ia S, Ndoye A, Lee TX, Webber RJ, Grando SA (2000) Receptor-mediated inhibition of keratinocyte migration by nicotine involvesmodulations of calcium influx and intracellular concentration.J Pharmacol Exp Ther 293:973–981.
oli M, Le Novere N, Hill JA Jr, Changeux JP (1995) Developmentalregulation of nicotinic ACh receptor subunit mRNAs in the ratcentral and peripheral nervous systems. J Neurosci 15:1912–1939.
wart R, De Filippi G, Broad LM, McPhie GI, Pearson KH, BaldwinsonT, Sher E (2002) 5-Hydroxyindole potentiates human �7 nicotinicreceptor-mediated responses and enhances acetylcholine-inducedglutamate release in cerebellar slices. Neuropharmacology43:374–384.
(Accepted 13 April 2004)
Copyright © 2022 FDOKUMEN




















