
Nicotinic Receptor Function in Schizophrenia
15
VOL22.NO. 3, 1996 Nicotinic Receptor Function in Schizophrenia 431 by Sherry Leonard, Catherine Adams, Charles R. Breese, Lawrence E. Adler, Paula Blckford, William Byerley, Hilary Coon, Jay M. Griffith, Christine Miller, Marina Myles- Worsley, Herbert T. Nagamoto, Yvonne Rollins, Karen E. Stevens, Merilyne Waldo, and Robert Freedman Abstract Schizophrenia can be partially char- acterized by deficits in sensory proc- essing. Biochemical, molecular, and genetic studies of one such endo- phenotype, the P50 auditory-evoked potential gating deficit, suggest that one of the neuronal nicotinic recep- tors, the a7 nicotinic receptor, may function in an inhibitory neuronal pathway involved in this pheno- type. The P50 deficit is normalized in nongating subjects by nicotine. Although most schizophrenia patients are heavy smokers, the effects of nicotine may be transient, as a7 receptors are known to desen- sitize rapidly. In an animal model of the P50 gating deficit, antagonists of the a7 nicotinic receptor block nor- mal gating of the second of paired auditory stimuli. Regional localiza- tion of receptor expression includes areas known to function in sensory filtering. An inhibitory mechanism, in the hippocampus, may involve nicotinic stimulation of -y-amino- butyric acid (GABA)ergic inter- neurons, resulting in decreased response to repetitive stimuli. Expression of the a7 receptor is decreased in hippocampal brain tis- sue, dissected postmortem, from schizophrenia subjects. The P50 deficit is inherited in schizophrenia pedigrees, but it is not sufficient for disease development and thus rep- resents a predisposition factor. Linkage analysis between the P50 deficit in multiplex schizophrenia pedigrees and deoxyribonucleic acid (DNA) markers throughout the genome yielded positive lod scores to DNA markers mapping to a region of chromosome 15 containing the oc7 nicotinic receptor gene. Elucidation of possible interactions of the P50 with other factors, known to be important in the etiology of the disease, is important in deter- mining an overall pathobiology of schizophrenia. Schizophrenia Bulletin, 22(3): 431-445,1996. Schizophrenia is a complex disease that has been difficult to characterize by either biochemical or classical genetic methods. Although a genetic component for the disease has been demonstrated from family and twin studies (Gottesman and Shields 1982; McGue and Gottesman 1989), there is not a Mendelian pattern of inheri- tance, and linkage of schizophrenia with a specific chromosomal locus has yet to be definitively established (Cloninger 1994). Biochemical mea- surements, candidate gene analysis, and environmental factor investiga- tion have failed to find a single deter- minant that makes a major contribu- tion to this mental illness (Sarkar et al. 1991; Catalano et al. 1992). In a broad sense, however, schizophrenia might be viewed as a disease consist- ing of a number of interactive pheno- types, each biochemically and geneti- cally independent, which together contribute to clinical manifestation of the disease. In the early 1960s, P. E. Meehl suggested that a useful strat- egy would be to search for a neuro- physiologica] "schizotaxic" factor, described by a single neuronal abnor- mality that would be inherited but would not necessarily be sufficient to cause schizophrenia by itself (Meehl 1962). Specific focus on such an endophenotypic factor might afford the investigator a single neuronal pathway for investigation and per- Reprint requests should be sent to Dr. S. Leonard, Dept. of Pharmacology, University of Colorado Health Sciences Center, Box C-268-71,4200 E. 9th Ave., Denver, CO 80262. by guest on July 14, 2011 schizophreniabulletin.oxfordjournals.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Nicotinic Receptor Function in Schizophrenia

VOL22.NO. 3, 1996 Nicotinic Receptor Functionin Schizophrenia
431
by Sherry Leonard, CatherineAdams, Charles R. Breese,Lawrence E. Adler, PaulaBlckford, William Byerley,Hilary Coon, Jay M. Griffith,Christine Miller, Marina Myles-Worsley, Herbert T. Nagamoto,Yvonne Rollins, Karen E.Stevens, Merilyne Waldo, andRobert Freedman
Abstract
Schizophrenia can be partially char-acterized by deficits in sensory proc-essing. Biochemical, molecular, andgenetic studies of one such endo-phenotype, the P50 auditory-evokedpotential gating deficit, suggest thatone of the neuronal nicotinic recep-tors, the a7 nicotinic receptor, mayfunction in an inhibitory neuronalpathway involved in this pheno-type. The P50 deficit is normalizedin nongating subjects by nicotine.Although most schizophreniapatients are heavy smokers, theeffects of nicotine may be transient,as a7 receptors are known to desen-sitize rapidly. In an animal model ofthe P50 gating deficit, antagonists ofthe a7 nicotinic receptor block nor-mal gating of the second of pairedauditory stimuli. Regional localiza-tion of receptor expression includesareas known to function in sensoryfiltering. An inhibitory mechanism,in the hippocampus, may involvenicotinic stimulation of -y-amino-butyric acid (GABA)ergic inter-neurons, resulting in decreasedresponse to repetitive stimuli.Expression of the a7 receptor isdecreased in hippocampal brain tis-sue, dissected postmortem, fromschizophrenia subjects. The P50deficit is inherited in schizophreniapedigrees, but it is not sufficient fordisease development and thus rep-resents a predisposition factor.Linkage analysis between the P50deficit in multiplex schizophreniapedigrees and deoxyribonucleicacid (DNA) markers throughout thegenome yielded positive lod scoresto DNA markers mapping to aregion of chromosome 15 containingthe oc7 nicotinic receptor gene.Elucidation of possible interactionsof the P50 with other factors, knownto be important in the etiology of
the disease, is important in deter-mining an overall pathobiology ofschizophrenia.
Schizophrenia Bulletin, 22(3):431-445,1996.
Schizophrenia is a complex diseasethat has been difficult to characterizeby either biochemical or classicalgenetic methods. Although a geneticcomponent for the disease has beendemonstrated from family and twinstudies (Gottesman and Shields 1982;McGue and Gottesman 1989), there isnot a Mendelian pattern of inheri-tance, and linkage of schizophreniawith a specific chromosomal locushas yet to be definitively established(Cloninger 1994). Biochemical mea-surements, candidate gene analysis,and environmental factor investiga-tion have failed to find a single deter-minant that makes a major contribu-tion to this mental illness (Sarkar etal. 1991; Catalano et al. 1992). In abroad sense, however, schizophreniamight be viewed as a disease consist-ing of a number of interactive pheno-types, each biochemically and geneti-cally independent, which togethercontribute to clinical manifestation ofthe disease. In the early 1960s, P. E.Meehl suggested that a useful strat-egy would be to search for a neuro-physiologica] "schizotaxic" factor,described by a single neuronal abnor-mality that would be inherited butwould not necessarily be sufficient tocause schizophrenia by itself (Meehl1962). Specific focus on such anendophenotypic factor might affordthe investigator a single neuronalpathway for investigation and per-
Reprint requests should be sent toDr. S. Leonard, Dept. of Pharmacology,University of Colorado Health SciencesCenter, Box C-268-71,4200 E. 9th Ave.,Denver, CO 80262.
by guest on July 14, 2011schizophreniabulletin.oxfordjournals.org
Dow
nloaded from

432 SCHIZOPHRENIA BULLETIN
haps simplify the genetic componentas well.
Schizophrenia patients suffer fromseveral types of sensory deficiencies,some of which can be measuredexperimentally (Adler et al. 1982;Braff and Saccuzzo 1985; Braff et al.1992). Patients have problems withthe interpretation of both sound andsight, resulting in the commonlyoccurring auditory and visual hal-lucinations (Andreasen 1985;McGlashan and Fenton 1992; Tsuang1993). It is as though the informationis reaching sensory nuclei in thebrain, but normal processing of thatinput and comparison with priorknowledge of a similar event do nottake place. This article reviews theprogress on the use of an auditory-evoked potential deficit as a modelendophenotype in schizophrenia forbiochemical, molecular, and geneticresearch.
Measurement of SensoryProcessing Deficits InSchizophrenia
The schizophrenia patient can bedescribed as hypervigilant; sensoryinput that a normal individual canfilter out continues to stimulate thepatient (Venables 1964). Repetitivesounds in the environment, such astraffic or construction sounds, areheard but ignored by normal sub-jects; the schizophrenia patient con-tinues to respond, often interpretingthe noise as threatening or directive.Auditory stimulation, such as soundsfrom a refrigerator or a furnace, oftenbecome overwhelmingly bother-some.
Several approaches to measuringthis deficit in human subjects havebeen successful (Braff and Saccuzzo1985; Freedman et al. 1987; Braff et al.1992). All involve the delivery of a
conditioning stimulus, followed aftera set period of time by a test stimu-lus. The normal subject has thecapacity to filter out or gate theresponse to the second input; theschizophrenia subject does not. Inone paradigm, a soft sound precedesa louder test sound by 100 ms; nor-mally there is a diminution ofresponse to the second and loudersound. However, this diminution ofresponse, referred to as prepulse inhi-bition, is less robust in the schizo-phrenia subject (Braff et al. 1992).
In the visual backward maskingassay of visual processing, two differ-ent objects are presented within 100ms, and the subject is asked to recallthe first. In schizophrenia subjects,identification of the first object isobscured or masked by visual inputfrom the second object. Normal sub-jects can easily identify both objects(Braff and Saccuzzo 1985). Thesetypes of analyses suggest that schizo-phrenia subjects cannot control orinhibit their responses to sensorystimuli.
Our laboratory uses yet anotherparadigm to study a neuronal path-way in the brain that regulates theprocessing of auditory stimuli. It waschosen not only because the pheno-type can be studied by recordingauditory-evoked potentials inpatients and normal subjects, but alsobecause it can be assayed in a labora-tory animal in which invasive elec-troencephalogram (EEG) and phar-macological experiments are possi-ble. The procedure, derived from aphenomenon originally reported byDavis et al. (1966), has been adaptedhere to measure inhibitory neuronalprocesses, or gating, of sensory audi-tory stimuli in the brain (Adler et al.1982; Boutros et al. 1991; Freedman etal. 1991). An initial auditory stimulusactivates or conditions an inhibitoryresponse, and a second auditory
stimulus then tests the extent of inhi-bition (Eccles 1969). The conditioningresponse is stimulatory becauseinhibitory mechanisms have not beenactivated; the test response normallyis decreased by inhibitory circuitsactivated by the first stimulus. Al-though several waves can be mea-sured, as a result of evoked potentialsfrom the delivery of auditory stimuli(Simpson and Knight 1993), we havefocused on the P50, a positive waveoccurring 50 ms after the stimulus,for analysis. The response at 50 ms isdecreased for several seconds follow-ing the test stimulus, defining theinhibition of extraneous auditoryinput, but the P50 does not exhibitlonger-term habituation, and manypairs of stimuli can thus be examinedin a given testing session (Adler et al.1982; Freedman et al. 1983; Boutros etal. 1991; Judd et al. 1992).
This inhibitory gating effect isreported as the ratio of the amplitudeof the test (T) P50 wave over theamplitude of the conditioning (C)wave or P50 ratio. In a study of 37schizophrenia subjects and 43 normalsubjects, 94 percent of normal sub-jects had P50 ratios s 0.50; 91 percentof the schizophrenia subjects hadratios a 0.50 (x2 = 55.5, df=l,p<0.001) (Waldo et al. 1994). The meanfor normal subjects was 0.18 ± 0.17and for schizophrenia subjects, 1.0 ±0.32. The schizophrenia subject isthus unable to filter or gate out repet-itive auditory input—a measurablephenotype that may result in abnor-mal interpretation of extraneoussounds.
Gating of auditory-evoked poten-tials in control subjects can be over-come by directing attention to thetest stimulus, which can be demon-strated by requiring an observationabout the test stimulus, such ascounting odd tones. Gating of the testresponse is then lost (Guterman et al.
by guest on July 14, 2011schizophreniabulletin.oxfordjournals.org
Dow
nloaded from

VOL 22, NO. 3, 1996 433
1992). Gating is also lost in normalsubjects by increased catecholaminerelease, introduced either by discom-fort (Johnson and Adler 1993) or bypharmacological intervention (Adleret al. 1994). This capacity to overridethe inhibitory response when neces-sary, by increasing attention to allstimuli in the environment, allows usto respond when required, even if thestimulus would normally be ignored.Effects of selective attention and sub-ject comfort, however, are importantparameters in the comparison ofauditory gating between schizophre-nia subjects and controls. These andother procedural considerations af-fect the reliability and repeatability ofthe assay (Boutros et al. 1991; Jergeret al. 1992; Waldo et al. 1992).
Interestingly, the gating deficit isalso found in about 50 percent of theschizophrenia patient's first-degreerelatives, who do not have the illness.The deficit therefore probably has agenetic component, and indeed, itappears to be inherited from the sideof the family carrying the schizophre-nia phenotype (Siegel et al. 1984;Waldo et al. 1991). Other sensoryendophenotypes, such as deficits insmooth-pursuit eye movements(Holzman et al. 1988) and in reactiontime (De Amicis et al. 1986), may beinherited in a similar manner. Mostsuch deficits, including the P50deficit, have an apparent autosomaldominant transmission pattern, thatis, the deficit is found in one parent,in the proband, in half the siblings,and in half of the siblings' andproband's descendents. The P50deficit thus represents an endopheno-type that may predispose the personwho expresses it to schizophrenia.
Pharmacology of AuditoryGating In SchizophreniaSubjects and Controls
Neuroleptic Medication. Clinical
drug treatments and the measure-ment of auditory gating deficits inother psychiatric illnesses such asmania suggest that the deficits seenin schizophrenia are not directlyrelated to aberrant dopaminergicneurotransmission. Typical neurolep-tic drugs such as haloperidol do notnormalize the P50 deficit (Freedmanet al. 1983; Adler et al. 1990). Al-though unmedicated schizophreniasubjects have smaller P50 waves thando normal controls and the size ofthe waves is normalized by neurolep-tics, both the conditioning and thetest waves are affected equally, re-sulting in retention of the abnormal77 C ratio (Straumanis et al. 1982;Freedman et al. 1983; Adler et al.1990). In patients who respond clini-cally to the atypical neurolepticclozapine, P50 ratios are normalized(Nagamoto et al., in press). Manicpatients exhibit abnormal P50 gatingratios, but only during their manicphase; the abnormal P50 ratios nor-malize when the patients areeuthymic. The response duringmania is probably related to in-creased adrenergic activity, as theP50 ratio is correlated with levels ofthe norepinephrine metabolite 3-methoxy,4-hydroxyphenylglycol(Adler et al. 1990). Increased adrener-gic response also has been implicatedin altered P50 ratios in controls,resulting either from discomfort orfrom exposure to potentially danger-ous situations (Waldo et al. 1992;Johnson and Adler 1993). However,in schizophrenia subjects, the P50ratio remains abnormal in conditionsof discomfort and following adminis-tration of conventional neurolepticmedication, suggesting that their P50ratios are controlled by neurotrans-mitter systems other than dopamine.
Regulation of the Auditory GatingDeficit by Nicotine. Nicotine is aself-administered drug that has
received only limited interest in thefield, probably due to the relativefailure of cholinergjc agonists andacetylcholinesterase inhibitors to sig-nificantly ameliorate the symptomsof schizophrenia (Pfeiffer and Jenny1957; Rosenthal and Bigelow 1973;Berger et al. 1979); muscarinic anti-cholinergics, however, have exacer-bated some symptoms of schizophre-nia (Tandon and Greden 1989). Theidea that a deficiency in nicotiniccholinergic neurotransmitter systemsmight underlie sensory deficits inschizophrenia patients and theirfirst-degree relatives has onlyrecently been considered (Hughes etal. 1986; Adler et al. 1992,1993; Goffet al. 1992; Karson et al. 1993). Yetschizophrenia subjects may have dis-covered this long ago; it is clinicallywell recognized that smoking inci-dence in the mentally ill is muchhigher than normal and is inordi-nately increased in the populationwith schizophrenia (Greeman andMcClellan 1991; Menza et al. 1991;Goff et al. 1992). Smoking in schizo-phrenia subjects can be pathologic;patients may smoke or chew tobaccoto the point of nausea and havedeveloped water intoxication fromdecreased secretion of antidiuretichormone, a side effect of nicotine(Kirch et al. 1985).
Investigation of smoking effects onthe auditory-evoked potential deficit,however, could be complicated bychronic exposure to nicotine. In ini-tial experiments to investigate theeffects of nicotine on the P50 deficit,we chose six nonschizophrenia, first-degree relatives of schizophreniasubjects who exhibited the P50 gat-ing deficit but who were nonsmokersand who, because they had no psy-chiatric illness, were neuroleptic-free. In a double-blind, placebo-con-trolled crossover study, nicotine wasadministered in the form of nicotine-containing chewing gum (total of 6
by guest on July 14, 2011schizophreniabulletin.oxfordjournals.org
Dow
nloaded from
434 SCHIZOPHRENIA BULLETIN
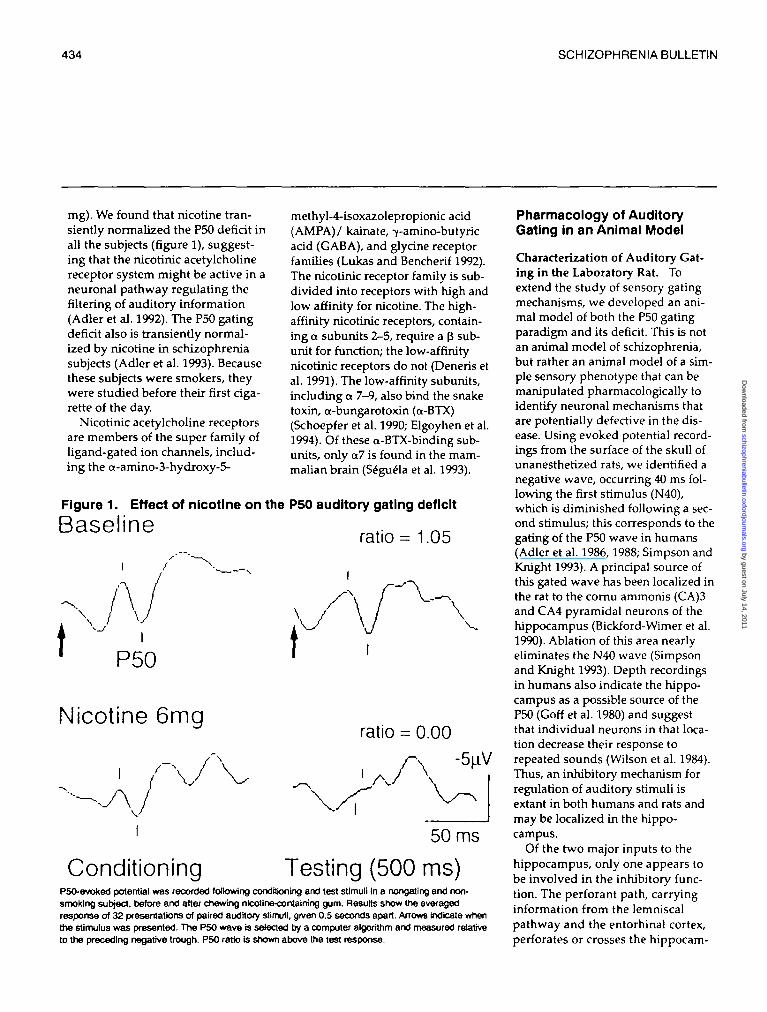
mg). We found that nicotine tran-siently normalized the P50 deficit inall the subjects (figure 1), suggest-ing that the nicotinic acetylcholinereceptor system might be active in aneuronal pathway regulating thefiltering of auditory information(Adler et al. 1992). The P50 gatingdeficit also is transiently normal-ized by nicotine in schizophreniasubjects (Adler et al. 1993). Becausethese subjects were smokers, theywere studied before their first ciga-rette of the day.
Nicotinic acetylcholine receptorsare members of the super family ofligand-gated ion channels, includ-ing the a-amino-3-hydroxy-5-
methyl-4-isoxazolepropionic acid(AMPA)/ kainate, •y-amino-butyricacid (GABA), and glycine receptorfamilies (Lukas and Bencherif 1992).The nicotinic receptor family is sub-divided into receptors with high andlow affinity for nicotine. The high-affinity nicotinic receptors, contain-ing a subunits 2-5, require a B sub-unit for function; the low-affinitynicotinic receptors do not (Deneris etal. 1991). The low-affinity subunits,including a 7-9, also bind the snaketoxin, a-bungarotoxin (a-BTX)(Schoepfer et al. 1990; Elgoyhen et al.1994). Of these ct-BTX-binding sub-units, only a7 is found in the mam-malian brain (S£gu£la et al. 1993).
Figure 1. Effect of nicotine on the P50 auditory gating deficit
Baseline ,. , __ratio = 1.05
P50
Nicotine 6mgratio = 0.00
\J V -5[iV/ \
\
50 ms
Conditioning Testing (500 ms)P50-evoked potential was recorded following conditioning and test stimuli In a nongating and non-smoking subject, before and after chewing nicotine-cortfaining gum. Results show the averagedresponse of 32 presentations of paired auditory stimuli, given 0.5 seconds apart. Arrows Indicate whenthe stimulus was presented. The P50 wave is selected by a computer algorithm and measured relativeto the preceding negative trough. P5O ratio is shown above the test response.
Pharmacology of AuditoryGating in an Animal Model
Characterization of Auditory Gat-ing in the Laboratory Rat. Toextend the study of sensory gatingmechanisms, we developed an ani-mal model of both the P50 gatingparadigm and its deficit. This is notan animal model of schizophrenia,but rather an animal model of a sim-ple sensory phenotype that can bemanipulated pharmacologically toidentify neuronal mechanisms thatare potentially defective in the dis-ease. Using evoked potential record-ings from the surface of the skull ofunanesthetized rats, we identified anegative wave, occurring 40 ms fol-lowing the first stimulus (N40),which is diminished following a sec-ond stimulus; this corresponds to thegating of the P50 wave in humans(Adler et al. 1986,1988; Simpson andKnight 1993). A principal source ofthis gated wave has been localized inthe rat to the cornu ammonis (CA)3and CA4 pyramidal neurons of thehippocampus (Bickford-Wimer et al.1990). Ablation of this area nearlyeliminates the N40 wave (Simpsonand Knight 1993). Depth recordingsin humans also indicate the hippo-campus as a possible source of theP50 (Goff et al. 1980) and suggestthat individual neurons in that loca-tion decrease their response torepeated sounds (Wilson et al. 1984).Thus, an inhibitory mechanism forregulation of auditory stimuli isextant in both humans and rats andmay be localized in the hippo-campus.
Of the two major inputs to thehippocampus, only one appears tobe involved in the inhibitory func-tion. The perforant path, carryinginformation from the lemniscalpathway and the entorhinal cortex,perforates or crosses the hippocam-
by guest on July 14, 2011schizophreniabulletin.oxfordjournals.org
Dow
nloaded from
VOL. 22, NO. 3, 1996 435
pal fissure and enters CA3-CA4.This pathway also synapses ongranule cells of the dentate gyrus,whose mossy fibers project to theCA3-CA4. Neurons of the entorhi-nal cortex do not show a decreasedresponse to auditory stimulation, asseen in the hippocampus (Stafekhinaand Vinogradova 1975). The secondmajor input to the hippocampus isthrough the fimbria-fornix, whichseparates the two lateral ventriclesand carries afferent fibers from theseptum and brainstem into most
areas of the hippocampal formation.Lesion of the fimbria-fornix fibersresults in a diminished response tosensory stimuli in neurons in thisregion of the hippocampus (Vino-gradova 1975); administration ofnicotine to the fimbria-fornix-lesioned animal normalized audi-tory gating (Bickford and Wear1995). Cholinergic neurons of theseptal region, in a response similarto that of neurons in the hippo-campus, also exhibit gating of audi-tory stimuli (Miller and Freedman
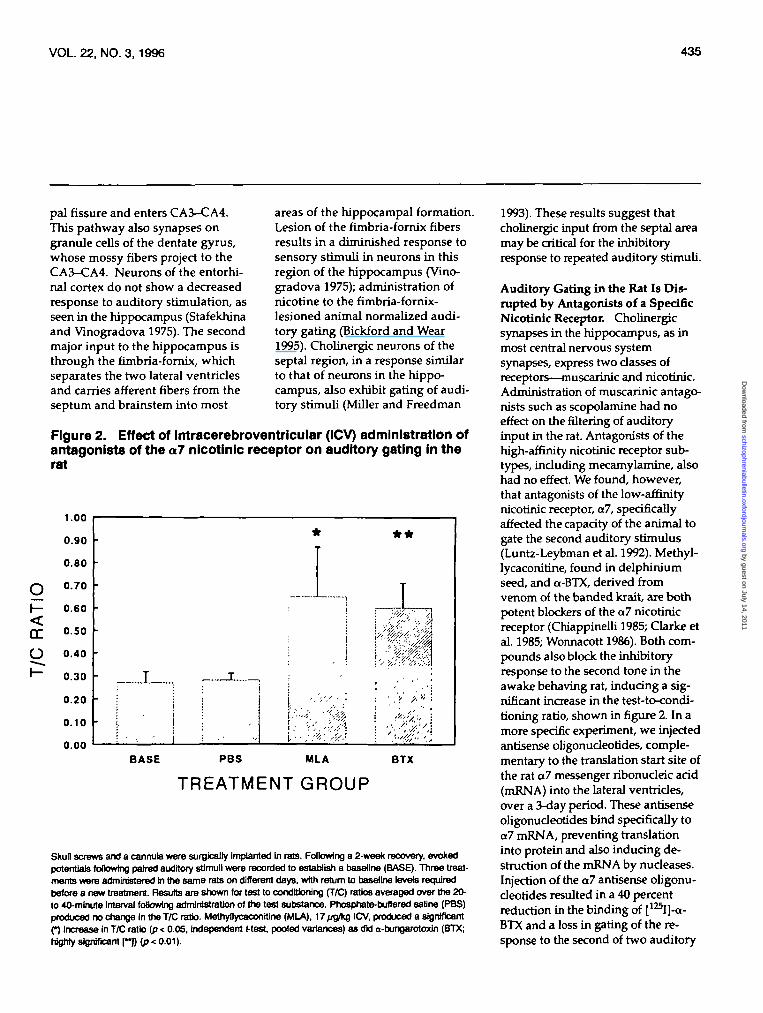
Figure 2. Effect of Intracerebroventricular (ICV) administration ofantagonists of the al nicotinic receptor on auditory gating in therat
o
RA
TI
O
1.00
0.90
0.80
0.70
0.60
0.50
0.40
0.30
0.20
0.10
0.00
-
-
; i ',
: • i :i. 1 :: . ' •
t * *
- !
<>••::'*'•.'//.
I
•'••'•','/•''X
BASE PBS MLA
TREATMENT GROUPBTX
Skull screws and a cannula were surgically Implanted In rats. Following a 2-week recovery, evokedpotentials following paired auditory stimuli were recorded to establish a baseline (BASE). Three treat-ments were administered in the same rats on different days, with return to baseline levels requiredbefore a new treatment. Results are shown for test to conditioning (T/C) ratios averaged over the 20-to 40-minute Interval foBowing administration of the test substance. Phosphate-buffered saflne (PBS)produced no change In the T/C ratio. Methyflycaconitine (MLA), 17/jg/kglCV, produced a significant(*) Increase in T/C ratio (p < 0.05, independent J-test, pooled variances) as did a-bungarotoxin (BTX;Wghty significant [**D (P < 001).
1993). These results suggest thatcholinergic input from the septal areamay be critical for the inhibitoryresponse to repeated auditory stimuli.
Auditory Gating in the Rat Is Dis-rupted by Antagonists of a SpecificNicotinic Receptor. Cholinergicsynapses in the hippocampus, as inmost central nervous systemsynapses, express two classes ofreceptors—muscarinic and nicotinic.Administration of muscarinic antago-nists such as scopolamine had noeffect on the filtering of auditoryinput in the rat. Antagonists of thehigh-affinity nicotinic receptor sub-types, including mecamylamine, alsohad no effect. We found, however,that antagonists of the low-affinitynicotinic receptor, al, specificallyaffected the capacity of the animal togate the second auditory stimulus(Luntz-Leybman et al. 1992). Methyl-lycaconitine, found in delphiniumseed, and a-BTX, derived fromvenom of the banded krait, are bothpotent blockers of the a7 nicotinicreceptor (Chiappinelli 1985; Clarke etal. 1985; Wonnacott 1986). Both com-pounds also block the inhibitoryresponse to the second tone in theawake behaving rat, inducing a sig-nificant increase in the test-to-condi-tioning ratio, shown in figure 2. In amore specific experiment, we injectedantisense oligonudeotides, comple-mentary to the translation start site ofthe rat al messenger ribonucleic acid(mRNA) into the lateral ventricles,over a 3-day period. These antisenseoligonudeotides bind specifically toal mRNA, preventing translationinto protein and also inducing de-struction of the mRNA by nudeases.Injection of the al antisense oligonu-deotides resulted in a 40 percentreduction in the binding of [125I]-a-BTX and a loss in gating of the re-sponse to the second of two auditory
by guest on July 14, 2011schizophreniabulletin.oxfordjournals.org
Dow
nloaded from

436 SCHIZOPHRENIA BULLETIN
stimuli (Rollins et al. 1993).More recently, nicotine has been
found to normalize an amphetamine-induced P50 gating impairment infreely moving rats (Stevens et al., inpress), and mice expressing low lev-els of a-BTX-binding nicotinic recep-tors have deficits in P50 auditory gat-ing (Stevens et al. 1995).
In both humans and laboratoryanimals, normalization of gatingdeficits by nicotine, and introduction
of the gating deficit in rats by specifica! antagonists suggests that thisnicotinic receptor may play an im-portant role in the inhibition of extra-neous noise in our environment.
Expression of the oil NicotinicAcetylchollne Receptor inHuman Brain
Presence of the a.7 receptor in hip-pocampal neurons has been shown
by receptor autoradiography, using["^J-a-BTX, at sites near the dentate
Figure 3. Binding of [1sl]-a-bungarotoxin In rat brain
:3&In rat brain, receptor autoradlography using [1I5t]-a-bungarotoxin shows binding on large nonprfndpalcefls and processes lying tn and near the dentate gyrus. Section was counterstained with cresyl violetThe targe labeled neuron with processes is 40 fjm In size.
gyrus and the CA3-CA4 region(Freedman et al. 1993). In a compara-tive study of a-BTX binding in thehippocampus of schizophrenia andcontrol subjects, done by quantitativereceptor autoradiography on humanpostmortem brain samples, we foundthat the level of binding in the schiz-ophrenia samples was significantlydecreased in both dentate gyrus andCA3 regions of the hippocampus ascompared to matched controls. Themean reduction in expression of thereceptor was 40 percent (Freedman etal. 1995).
We have recently completed aregional localization of the receptorprotein in both rat and human brain,using [125!]-a-BTX autoradiography.The c*7 receptor was found in manyareas of the brain, including areasknown to be involved in sensoryprocessing, such as hippocampus,medial and lateral geniculate, amyg-dala, and brainstem. In manyregions, binding is present on boththe cell bodies and on cellularprocesses, as shown on a cell locatedin the hilar region of the rat hip-pocampus (figure 3). Examples ofbinding in several regions of humanpostmortem brain are shown in fig-ure 4. In the hippocampus (figure4B), a cell with labeled processesextending into both the hilar regionand molecular layers is shown. Notall the cells in the hippocampus ex-hibited labeled processes. In the diag-onal band of Broca, near the septalnucleus (figure 4D), binding is alsoseen only on the cell bodies. Bindingon cellular processes of these neuronscannot be ruled out, but it might beexpressed at some distance from thecell; none were observed in the planeof the tissue section examined.
Of particular interest was the pat-tern of [125I]-a-BTX binding in the
by guest on July 14, 2011schizophreniabulletin.oxfordjournals.org
Dow
nloaded from

VOL 22, NO. 3, 1996 437
Figure 4. Binding of [ial]-a-bungarotoxin in human postmortembrain
A
t •» »
Guillery 1993). Anatomically, RTN isa capsular structure that surroundsthe dorsal and ventral thalamus.Neurons of the RTN are known to beuniformly GABAergic (Lozsadi1994), and nerve-growth-factor recep-tor is highly expressed in this nucleus(Chen and Bentivoglio 1993), as it isin the hippocampus. The heavy con-centration of a.7 receptor expressionin the RTN and its presence inGABAergic interneurons of the hip-pocampus (Freedman et al. 1993)suggest that a.7 may also be localizedon inhibitory neurons in this thalamicnucleus and might participate in theprocessing or filtering of sensoryinformation in thalamocortical cir-cuits as well as in the hippocampus.
LRt
?-?;&<;• •
pUv'' : - • ; ; • • *
•• k.
(A) Reticular thalamic nucleus showing binding to both cell bodies and a network of processes. LRt =lateral reticular thalamus. (B) A large ceB, similar to those seen In rat hippocampus, positioned Just out-side the dentate gyrus with processes extending into the hilar region and molecular layers, dg ° den-tate gyrus; h = Wlus. (C) Lateral geniculate showing [""ll-a-bungarotodn binding on cell bodies only.(D) Diagonal band of Broca with labeling on elongate cetl bodies. Scale ban (A) 200 ̂ m; (B.C.D) 75
reticular thalamic nucleus (figure4A). Neuxonal cell bodies are heavilylabeled by the ligand, as are proc-esses. The latter form dense andintertwined bundles of fibers that runthroughout this nucleus. The reticu-
iaj thalamic nucleus (RTN) is thoughtto be a pacemaker in corticothalamicand thalamocortical connections(Crick 1984), since it provides theprincipal inhibitory input into thedorsal thalamus (Mitrofanis and
A Neuronal Pathway forProcessing of AuditoryInformation in the Brain
The animal model of auditory gatinghas been useful for elucidating a pos-sible neuronal pathway in the brainfor the processing of auditory input.A model pathway, based on our ex-perimental evidence and on that ofothers, illustrates how such a mecha-nism might function in the hippo-campus (figure 5). The major inputfor excitatory information comesfrom the perforant path to the den-tate gyrus and CA3 pyramidal neu-rons. These neurons relay informa-tion through the Schaffer collateralsto CA1. The output from CA1 ismainly to the frontal lobe via thesubiculum. A mechanism for regula-tion of the information received bythe CA1 might be the inhibitory neu-rons of the CA3-CA4 regions. Theseneurons bear modulatory a7 nico-tinic receptors, but they are activatedprimarily by glutamitergic synapsesfrom the mossy fibers of the dentategyrus and from the perforant path.Stimulation by auditory input would
by guest on July 14, 2011schizophreniabulletin.oxfordjournals.org
Dow
nloaded from

438 SCHIZOPHRENIA BULLETIN
Figure 5. Neuronal model for gating of auditory stimuli in thehippocampus
PerforantPath
C A 3 A GABAPyramidalNeuron
Inter-neuron
SchafferCollaterals
Fimbria-Fornix
Auditory information reaches the hippocampus through the perforant path. Mossy fibers synapse onCA3 pyramidal cells and on CA3 irrterneurons as well. The Intemeuron Inhibits the pyramidal cell byGABAergic and other inhibitory synapses. Additionally, the pyramidal cefl and the Intemeuron are stim-ulated by cholinergic synapses coming from the septum through the fomix Blockade of this septalchofinergic Input removes the effect of the Intemeuron and permits the pyramidal cell to fire Inresponse to the second stimulus. In this model, choBnerglc stimulation is necessary for inhibition.Ach = acetytcholine; Otu = glutamate; GABA = -y-amino-butyrtc add.
cause the intemeuron to releaseGABA and other inhibitory neuro-transmitters onto the pyramidal neu-ron, resulting in cell discharge (Millerand Freedman 1995).
There are two types of GABAreceptors on pyramidal cells of thehippocampus: GABAAand GABAB.The combined effect is an inhibitioncapable of lasting 300 ms (Dutar andNicoll 1988). Additionally, GABAB
and other G, protein-linked receptorsare found presynaptically (Manzoneet al. 1994) and can depress gluta-mate release. Although the inhibitory
neurotransmitters released by theintemeuron mainly contact the cellbody receptors of the CA3 neurons,diffusion can occur through thesynaptic cleft to activate presynapticreceptors (Isaacson et al. 1993). Thiswould result in inhibition of gluta-mate release from perforant path andmossy fiber terminals. Indeed,GABAB antagonists block auditorygating in the rat (Hershman et al.1995). Because interneurons are acti-vated for longer than 250 ms after thefirst tone is delivered, the combinationof these events could result in the
long-term inhibition seen when thetest auditory stimulus is given 500 msafter the conditioning stimulus.
Input onto the intemeuron fromthe perforant path and input fromthe pyramidal neurons probably arenot sufficient to cause the intemeu-rons to discharge the necessary burstof action potentials. The intemeuron,however, also receives cholinergicinput from the medial septal nucleus.Our data, showing that antagonistsof the a.7 neuronal nicotinic receptorblock inhibition of the evoked re-sponse to paired auditory stimuli,suggest that this receptor may playan important role in the filtering ofauditory information passingthrough the hippocampus. A defectin the expression or function of thea7 receptor could thus result in aphenotype of failed inhibitory re-sponses to sensory input, which par-tially characterizes schizophrenia.
Cholinergic modulation of in-hibitory responses may also functionelsewhere in the brain. Acerylcholinemodulates an inhibitory response toauditory stimuli in the dentate gyrus(Foster and Deadwyler 1992), andfibers extending from the CA3 pyra-midal neurons back through thefornix excite the lateral septal nu-cleus. The lateral septum has a weakinhibitory input into the medial sep-tal nucleus, and inhibitory fibers alsoproject from the CA3 interneurons tothe medial septum (Alonso andKohler 1982). The principal input tothe medial septal nucleus, however,comes from the brainstem reticularformation, which also exhibits adecreased response to the second ofpaired stimuli (Bickford et al. 1993).Reticular neurons express nicotinicreceptors (Stevens et al. 1993) and arelabeled by [123I]-a-BTX, suggestingthat they may also express the a.7receptor. Auditory informationpasses through the nucleus of the lat-
by guest on July 14, 2011schizophreniabulletin.oxfordjournals.org
Dow
nloaded from

VOL22.NO. 3, 1996 439
eral lemniscus into a group of neu-rons of the reticular formation thatexpress auditory-induced startle, andit has been proposed that the hip-pocampus is responsible for the pre-pulse inhibition of this startle re-sponse (Swerdlow et al. 1994). Sev-eral of the inhibitory phenotypesmeasured in schizophrenia subjectsmay thus have common functionalelements in their neuronal circuitry.
Processing of AuditoryInformation In the Human Brain
Although the animal model of audi-tory gating has been helpful in thepharmacological analysis of this phe-notype, the neuronal pathway in thehuman brain is likely to be morecomplex because of increased devel-opment in the forebrain areas. Thesuperior temporal gyms is a majorsource of auditory input to thehuman limbic system; a dipole forthe P50 response has been localizedto this gyrus by magnetoencephalog-raphy (Reite et al. 1988). The superiortemporal gyrus receives a cholinergicinnervation from the nucleus basalis,which regulates an auditory responsein the neocortex (Metherate and Ashe1993), probably by a mechanism sim-ilar to the one we have found in thehippocampus. Abnormal P300 re-sponses to auditory-evoked poten-tials have been recorded from surfaceelectrodes over the superior temporalgyrus in schizophrenia subjects(McCarley et al. 1993), although it isnot known whether this aberrancy isbiochemical or is related to structuralchanges in the superior temporalgyms.
In addition to the cholinergic neu-rotransmitter system, other inputs toinhibitory neurons are likely to beimportant in proper function. Nor-
adrenergic and serotonergic inputsfrom the brainstem are known todecrease interneuron activity (Madi-son and Nicoll 1988; Freund 1992).This effect could be aberrant inmania, where remission of symptomsand normal P50 gating is seen whennorepinephrine metabolite levelsreturn to baseline levels (Adler et al.1990). Qozapine, a serotonergicantagonist, may be effective in someschizophrenia subjects by blockingserotonin's normal inhibitory inputto the interneurons (Schorte et al.1993). This would have the effect ofincreasing the interneurons' capacityto gate sensory response and wouldaccount for the normalization of theP50 deficit seen in schizophreniapatients on dozapine therapy (Nag-amoto et al., in press). Typical neu-roleptics do not normalize auditorygating in schizophrenia patients, sug-gesting that these drugs may nothave a direct effect on inhibitoryresponses in the hippocampus butinstead may decrease the synapticresponses in the forebrain nuclei tosensory activity coming through theperforant pathway, resulting in ageneral decrease in excitability of thehippocampal region (Johnson et al.1983).
Other neurotransmitter systemscould interact with cholinergic inner-vation to enhance or inhibit nicotine-induced responses. Nicotinic recep-tors are found on many catechol-aminergic neurons, and stimulationof these high-affinity receptors bynicotine is known to induce release ofnorepinephrine. These receptors areblocked by mecamylamine in thehuman, and the normalizing effect ofnicotine on the abnormal P50 isenhanced by mecamylamine, per-haps because the decrease in inter-neuronal activity caused by norepi-nephrine has been removed.
The P50 Deficit as a RiskFactor for Schizophrenia
The P50 Deficit Is Inherited. TheP50 deficit is one of several measur-able phenotypes, including decreasedP300 amplitude (Blackwood et al.1991) and aberrant smooth-pursuiteye movement (Holzman et al. 1988),that are found in approximately 50percent of the first-degree relatives ofschizophrenia subjects, who do nothave the disease. Usually, the deficitis observed in one parent and in halfof the siblings, describing an autoso-mal dominant type of inheritance.Nine families with multiple-genera-tional schizophrenia were studied forcodistribution of schizophrenia andthe P50 gating deficit. A total of 120subjects were recorded, of whom 34were diagnosed with schizophrenia.Subjects with P50 quotients below0.40 were classified as normal, thosewith 77 C ratios of 0.50 or more wereclassified as abnormal, and thosewith ratios in between were regardedas unknown. Based on these criteria,32 of the 34 schizophrenia subjectshad abnormal ratios and 2 wereunknown. Schizophrenia was alwaystransmitted through a parent with anabnormal P50 ratio, suggesting thephenotype came from only one sideof the family. A mathematical modelfitted to the pedigrees supports thehypothesis that presence of an abnor-mal P50 is directly related to theinheritance of risk for schizophrenia(Waldo et al. 1991).
A Secondary Factor Is Required forExpression of the SchizophreniaSyndrome. Presence of the gatingdeficit in nonschizophrenia relativesthat have an abnormal P50 suggeststhat this deficit is not sufficient fordevelopment of the schizophreniadisease state. A study of siblings dis-
by guest on July 14, 2011schizophreniabulletin.oxfordjournals.org
Dow
nloaded from

440 SCHIZOPHRENIA BULLETIN
cordant for the P50 deficit and schiz-ophrenia was undertaken. Magneticresonance imaging was used todetermine the hippocampa] volumein three groups: siblings with schizo-phrenia and abnormal P50 ratios,nonschizophrenia siblings withabnormal P50 ratios, and normal sib-lings with normal ratios. We foundthat a reduction in volume of thehippocampus was related to thedevelopment of schizophrenia in sib-lings with abnormal P50 ratios(Waldo et al. 1994). Although therewas variability in hippocampal sizein normal subjects, the nonschizo-phrenia siblings with abnormal P50shad normal to large hippocampi; theschizophrenia subjects had bothabnormal P50s and significantlysmaller hippocampal volumes.Expressed alone, neither of thesedeficits was sufficient for diseasedevelopment, as many of the siblingswith normal P50 ratios had reducedhippocampal volumes. Interestingly,although the nonschizophrenia sib-lings with abnormal P50 gatingdeficits had no history of chronicpsychiatric disease, the siblings withnormal P50 ratios often were diag-nosed with disorders including alco-holism and minor depression, sug-gesting that a second factor may beinvolved (Waldo et al. 1994).
Hippocampal pathology is welldocumented in schizophrenia Qesteand Lohr 1989; Suddath et al. 1990).However, it is not dear whether itscause is genetic, developmental, orenvironmental. Correct temporalexpression of growth factors andneurotransmitter systems both havebeen shown to be necessary for com-petent maturation of neuronal cir-cuits in the central nervous system(Snider and Johnson 1989; Knusel etal. 1990; McDonald and Johnstone1990; Crowley et al. 1994). The inves-tigation of interactive biochemical
systems such as the P50 deficit inestablished endophenotypes found inschizophrenia thus represents a logi-cal approach to a disease of remark-able complexity.
Possible Linkage of the P50Deficit to SpecificChromosomal Regions
Since the P50 deficit appeared to beinherited in the nine pedigrees exam-ined, these families were used in apreliminary linkage study with morethan 300 mapped markers, coveringthe entire genome. Linkage analysisrepresents an independent approachfor analysis of a disease or phenotypebecause it makes no assumptionabout the identification of the genesinvolved. Linkage studies in schizo-phrenia, however, are difficultbecause of the low rates of reproduc-tion in many patients and the possi-bility that multiple genes may beinvolved. As an exploratoryapproach, a 95-percent confidencelevel for lod (log to the base 10 of theodds ratio) scores was determinedfor markers that were not linked tothe P50 or schizophrenia. The resultsshowed positive lod scores (p < 0.05)for the P50 deficit, which were alsononnegative for schizophrenia at foursites in the genome (Coon et al. 1993).One of these regions, 15ql4, on thelong arm of chromosome 15, has sub-sequently been shown to be the locusof the ot7 nicotinic receptor gene(Chini et al. 1994).
Although the study was prelimi-nary and only suggests further explo-ration of a rather large region of chro-mosome 15, it is important that threeindependent lines of investigation—gene expression, pharmacologicalstudies, and genetic-linkage analy-sis—have all suggested involvementof the a7 nicotinic receptor in gating
of sensory stimuli and schizophrenia.Work is in progress to determinewhether abnormalities in the a7 geneitself are linked to the P50 deficit andschizophrenia.
Two other disorders with psychoticelements are caused by geneticdefects with loci near 15ql4. Prader-Willi syndrome, usually the resujt ofa deletion involving 15qll-14, has ahigh association with schizophrenia(Clarke 1993). Marian's syndrome,linked to 15ql5, also has beenobserved to cosegregate with schizo-phrenia (Sirota et al. 1990). It is possi-ble that genes localized in the interimregion between 15qll and 15q22 areassociated with schizophrenia and, insome families, are inherited withthese disease alleles.
Conclusion
Concentrated study of endopheno-types in diseases of complex originprovides several advantages. Unlikethe disease state, which is difficult toquanritate, endophenotypes such asthe P50 deficit and aberrant smooth-pursuit eye movement can be exam-ined in humans and limited pharma-cological research can be accom-plished. This information can then beused for linkage analysis, since a sin-gle endophenotype is more likely tohave a simple, Mendelian form ofinheritance. Additionally, several ofthe traits associated with schizophre-nia can be examined in laboratoryanimals where specific neuroanatom-ical areas, such as the hippocampus,can be studied using pharmacologi-cal and electrophysiologjcal methods.We have characterized one suchtrait—the auditory-evoked potentialdeficit—found in nearly all schizo-phrenia subjects and about half oftheir clinically unaffected first-degreerelatives. The deficit is inherited, pos-
by guest on July 14, 2011schizophreniabulletin.oxfordjournals.org
Dow
nloaded from

VOL 22, NO. 3, 1996 441
sibly as a necessary but not sufficientcondition for schizophrenia, and thusrepresents a predisposition factor.Interaction with other biochemicaland/or developmental defects orenvironmental factors may be neces-sary for full presentation of the ill-ness. Although several neurorrans-mitter systems probably function inthe neuronal pathway regulating gat-ing of the P50, and have beendemonstrated with state-dependentchanges in P50 gating, we haveexamined the role of the neuronalnicotinic acetylcholine receptor fam-ily in this pathway.
Biological and pharmacologicalevidence suggests that a subtype ofthis receptor family—the a7 nicotinicreceptor—is specifically involved intrait-related P50 gating deficits. Lev-els of oil expression are decreased inschizophrenia subjects, and thegenomic locus for this gene lies in aregion of chromosome 15, identifiedin a genetic-linkage study of P50 as aregion of interest. The a.7 receptor isexpressed in many sensory-filteringnuclei in the brain, including the hip-pocampus and RTN. The synapticcharacteristics of auditory gating,which we have measured in the hip-pocampus, may also be extant inthese other nuclei. It will be impor-tant to determine the expression andfunction of the receptor, wherever itis found, to understand possiblepathogenic effects of dysfunction.Although evidence for involvement ofthe a7 receptor as a candidate gene ispromising, interaction with otherknown pathologies in the diseasemust be investigated before an un-derlying model for the pathogenesisof schizophrenia can be determined.
References
Adler, L.E.; Gerhardt, G.A.; Franks,R.; Baker, N.; Nagamoto, H.; Drebing,
C; and Freedman, R. Sensory physi-ology and catecholamines in schizo-phrenia and mania. PsychiatryResearch, 31:297-309,1990.
Adler, L.E.; Hoffer, L; Griffith, J.M.;Waldo, M.C.; and Freedman, R. Nor-malization by nicotine of deficientauditory sensory gating in the rela-tives of schizophrenics. Biological Psy-chiatry, 32:607-616,1992.
Adler, L.E.; Hoffer, L.; Nagamoto,H.T.; Waldo, M.C.; Kisley, M.A.; andGriffith, J.M. Yohimbine impairs P50auditory sensory gating in normalsubjects. Neuropsychopharmacology,10:249-257,1994.
Adler, L.E.; Hoffer, L; Wiser, A.; andFreedman, R. Normalization of audi-tory physiology by cigarette smokingin schizophrenic patients. Americanjournal of Psychiatry, 150:1856-1861,1993.Adler, L.E.; Pachtman, E.; Franks, R.;Pecevich, M.; Waldo, M.C.; andFreedman, R. Neurophysiologicalevidence for a defect in neuronalmechanisms involved in sensory gat-ing in schizophrenia. Biological Psy-chiatry, 17:639-654,1982.
Adler, L.E.; Rose, G.M.; and Freed-man, R. Neurophysiological studiesof sensory gating in rats: Effects ofamphetamine, phencyclidine, andhaloperidol. Biological Psychiatry,21:787-798,1986.
Adler, L.E.; Rose, G.M.; Pang,}.; andGerhardt, G. Modulation of the gat-ing of auditory evoked potentials bynorepinephrine and dopamine: Phar-macological evidence obtained usinga selective neurotoxin. Biological Psy-chiatry, 24:179-190,1988.
Alonso, A., and Kohler, C. Evidencefor separate projections of hippocam-pal and non-pyramidal neurons todifferent parts of the septum in ratbrain. Neuroscience Letters, 21:209-214,1982.
Andreasen, N.C. Positive vs. negative
schizophrenia: A critical evaluation.Schizophrenia Bulletin, ll(3):380-389,1985.Berger, P.A.; Davis, K.L.; and Hollis-ter, L.E. Pharmacological investiga-tions of cholinergic mechanisms ofpsychosis in schizophrenia andmanic psychosis. In: Davis, K.L., andBerger, P.A., eds. Brain Acetylcholineand Neuropsychiatric Disease. NewYork, NY: Plenum Press, 1979.pp. 15-32.
Bickford, PC; Luntz-Leybman, V.;and Freedman, R. Auditory sensorygating in the rat hippocampus: Mod-ulation by brainstem activity. BrainResearch, 607:33-38,1993.Bickford, P.C., and Wear, K. Fimbriafornix lesions disrupt auditory sen-sory gating in the rat hippocampus.Brain Research, 705:235-240,1995.
Bickford-Wimer, PC; Nagamoto, H.;Johnson, R.; Adler, L; Egan, M.; Rose,G.M.; and Freedman, R. Auditorysensory gating in hippocampal neu-rons: A model system in the rat. Bio-logical Psychiatry, 27:183-192,1990.Blackwood, D.H.; St. Clair, D.M.;Muir, W.J.; and Duffy, J.C. AuditoryP300 and eye tracking dysfunction inschizophrenic pedigrees. Archives ofGeneral Psychiatry, 48:899-909,1991.Boutros, N.N.; Overall, J.; and Zouri-dakis, G. Test-retest reliability of theP50 mid-latency auditory evokedresponse. Psychiatry Research, 39:181-192,1991.
Braff, D.L.; Grillon, C; and Geyer,M. A. Gating and habituation of thestartle reflex in schizophrenicpatients. Archiites of GeneralPsychiatry, 49:206-215,1992.
Braff, D.L., and Saccuzzo, D.P. Thetime course of information-process-ing deficits in schizophrenia. Ameri-can Journal of Psychiatry, 142:170-174,1985.Catalano, M.; Nobile, M.; Novelli, E.;and Smeraldi, E. Use of polymerase
by guest on July 14, 2011schizophreniabulletin.oxfordjournals.org
Dow
nloaded from

442 SCHIZOPHRENIA BULLETIN
chain reaction DNA denaturing gra-dient gel electrophoresis to identifypolymorphisms in three exons ofdopamine D2 receptor gene in schiz-ophrenic and delusional patients. Bio-logical Psychiatry, 26:1-3,1992.
Chen, S., and Bentivoglio, M. Nervegrowth factor receptor-containingcholinergic neurons of the basal fore-brain project to the thalamic reticularnucleus in the rat. Brain Research,606:207-212,1993.Chiappinelli, V. Actions of snakevenom toxins on neuronal nicotinicreceptors and other neuronal recep-tors. Pharmacology and Therapeutics,31:1-32,1985.
Chini, B.; Raimond, E.; Elgoyhen,A.B.; Moralli, D.; Balzaretti, M.; andHeinemann, S. Molecular cloningand chromosomal localization of thehuman 7-nicotinic receptor subunitgene (CHRNA7). Genomics, 19:379-381,1994.
Clarke, D.J. Prader-Willi syndromeand psychoses. British Journal of Psy-chiatry, 163:680-684,1993.
Clarke, P.B.; Schwartz, R.D.; Paul,S.M.; Pert, C.B.; and Pert, A. Nicotinicbinding in rat brain: Autoradio-graphic comparison of [3H]-acetyl-choline, [3H]-nicotine, and [125I]-a-bungarotoxin. Journal of Neuroscience,5:1307-1315,1985.
Cloninger, C.R. Turning point in thedesign of linkage studies of schizo-phrenia. America Journal of MedicalGenetics: Neuropsychiatric Genetics,54:83-92,1994.
Coon, H.; Plaetke, R.; Holik, J.; Hoff,M.; Myles-Worsley, M.; Waldo, M.;Freedman, R.; and Byerley, W. Use ofa neurophysiological trait in linkageanalysis of schizophrenia. BiologicalPsychiatry, 34:277-289,1993.
Crick, F. Function of the thalamicreticular complex: The searchlighthypothesis. Proceedings of the National
Academy of Sciences of the United Statesof America, 81:4586-1590,1984.Crowley, C; Spencer, S.D.; Nishimura,M.C.; Chem, K.S.; Pitts-Meek, S.;Armanini, M.P.; ling, L.H.; McMahon,S.B.; Shelton, D.L.; Levinson, A.D.;and Phillips, H.S. Mice lacking nervegrowth factor display perinatal loss ofsensory and sympathetic neurons yetdevelop basal forebrain cholinergicneurons. Cell, 76:1001-1011,1994.Davis, H.; Mast, T.; Yoshie, N.; andZerlin, S. The slow response of thehuman cortex to auditory stimuli:Recovery cycle process. Electro-encephalography and Clinical Neuro-physiology, 21:105-113,1966.
De Amicis, L.E.; Wagstaff, D.A.; andCromwell, R.L. Reaction time cross-over as a marker of schizophreniaand of higher functioning. Journal ofNervous and Mental Disease,174:177-179,1986.
Deneris, E.S.; Connolly, J.; Rogers,S.W.; and Duvoisin, R. Pharmaco-logical and functional diversity ofneuronal nicotinic acetylcholinereceptors. Trends in PharmacologicalSciences, 12:34-40,1991.
Dutar, P., and Nicoll, R.A. Pre- andpostsynaptic GABAB receptors in thehippocampus have different pharma-cological properties. Neuron, 1:585-591,1988.Ecdes, J.C. The Inhibitory Pathways ofthe Central Nervous System. Spring-field, IL: Charles C Thomas, 1969.Elgoyhen, A.B.; Johnson, D.S.; Boul-ter, J.; Vetter, D.E.; and Heinemann, S.Alpha 9: An acetylcholine receptorwith novel pharmacological proper-ties expressed in rat cochlear haircells. Cell, 79:705-715,1994.Foster, T.C., and Deadwyler, S.A.Acetylcholine modulates averagedsensory evoked response and per-forant path evoked field potentials inthe rat dentate gyrus. Brain Research,587:95-101,1992.
Freedman, R.; Adler, L.E.; Baker, N.;Waldo, M.; and Mizner, G. Candidatefor inherited neurobiological dys-function in schizophrenia. SomaticCell and Molecular Genetics,13:479-484,1987.
Freedman, R.; Adler, L.E.; Waldo,M.C.; Pachtman, E.; and Franks, R.D.Neurophysiological evidence for adefect in inhibitory pathways inschizophrenia: Comparison of med-icated and drug-free patients. Biologi-cal Psychiatry, 18:537-551,1983.
Freedman, R.; Hall, M.; Adler, L.E.;and Leonard, S. Evidence in post-mortem brain tissue for decreasednumbers of hippocampal nicotinicreceptors in schizophrenia. BiologicalPsychiatry, 38:22-33,1995.Freedman, R.; Waldo, M.; Bickford-Wimer, P.; and Nagamoto, H. Ele-mentary neuronal dysfunctions inschizophrenia. Schizophrenia Research,4:233-243,1991.Freedman, R.; Wetmore, C; Stein-berg, I.; Leonard, S.; and Olson, L.ot-Bungarotoxin binding to hippo-campal interneurons: Immunocyto-chemical characterization and effectson growth factor expression. Journalof Neuroscience, 13:1965-1975,1993.
Freund, R.F. GABAergjc septal andserotonergic median raphe afferentspreferentially innervate inhibitoryinterneurons in the hippocampusand dentate gyrus. Epilepsy Research,7(Suppl.):79-91,1992.
Goff, D.C.; Henderson, D.C.; andAmico, E. Cigarette smoking in schiz-ophrenia: Relationship to psycho-pathology and medication sideeffects. American Journal of Psychiatry,149:1189-1194,1992.
Goff, W.R.; Williamson, P.D.;VanGilder, J.C; Allison, R.; andFisher, T.C. Neural origins of longlatency evoked potentials recordedfrom the depth and from the cortical
by guest on July 14, 2011schizophreniabulletin.oxfordjournals.org
Dow
nloaded from

VOL.22, NO. 3, 1996 443
surface of the brain in man. Progressin Clinicol Neurophysiology, 7:126-145,1980.Gottesman, I.I., and Shields, J. Schizo-phrenia: The Epigenetic Puzzle. Cam-bridge, England: Cambridge Univer-sity Press, 1982.
Greeman, M., and McClellan, T.A.Negative effects of a smoking ban onan inpatient psychiatry service. Hos-pital and Community Psychiatry,42:408-412,1991.
Guterman, Y; Josiassen, R.C.; andBashore, T.R., Jr. Attentional influ-ence on the P50 component of theauditory event-related brain poten-tial. International Journal of Psycho-physiology, 12:197-209,1992.
Hershman, K.M.; Freedman, R.; andBickford, P.C. GABAB antagonistsdiminish the inhibitory gating ofauditory response in the rat hip-pocampus. Neuroscience Letters,190:133-136,1995.
Holzman, P.S.; Kringlen, E.;Matthysse, S.; Flanagan, S.D.; lipton,R.B.; Cramer, G.; Levin, S.; Lange, K.;and Levy, D.L. A single dominantgene can account for eye trackingdysfunctions and schizophrenia inoffspring of discordant twins.Archives of General Psychiatry,45:641-647,1988.
Hughes, J.R.; Hatsukami, D.K.;Mitchell, J.E.; and Dahlgren, I.A.Prevalence of smoking among psy-chiatric outpatients. American Journalof Psychiatry, 143:993-997,1986.
Isaacson, J.S.; Solis, J.M.; and Nicoll,R.A. Local and diffuse synapticactions of GABA in the hippocam-pus. Neuron, 10:165-175,1993.
Jerger, K.; Biggins, C; and Fein, G.P50 suppression is not affected byattentional manipulations. BiologicalPsychiatry, 31:365-377,1992.
Jeste, D.V., and Lohr, J.B. Hippo-
campal pathologic findings in schizo-phrenia. Archives of General Psy-chiatry, 46:1019-1024,1989.Johnson, M.R., and Adler, L.E. Tran-sient impairment in P50 auditorysensory gating induced by a coldpressor test. Biological Psychiatry,33:380-387,1993.
Johnson, S.W.; Palmer, M.R.; andFreedman, R. Effects of dopamine onspontaneous and evoked activity ofcaudate neurons. Neuropharmacology,22:843-851,1983.
Judd, L.L.; McAdams, L.; Budnick, B.;and Braff, D.L. Sensory gating def-icits in schizophrenia: New results.American Journal of Psychiatry,149:488-493,1992.
Karson, C.N.; Casanova, M.F.; Klein-man, J.E.; and Griffin, W.S. Cholineacetyltransferase in schizophrenia.American Journal of Psychiatry,150:454-459,1993.Kirch, D.G.; Bigelow, L.B.; Wein-berger, D.R.; Lawson, W.B.; andWyatt, R.J. Polydipsia and chronichyponatremia in schizophrenic inpa-tients. Journal of Clinical Psychiatry,46:179-181,1985.
Knusel, B.; Michel, P.P.; Schwaber,J.S.; and Hefti, F. Selective and non-selective stimulation of centralcholinergic and dopaminergjc devel-opment in vitro by nerve growth fac-tor, basic fibroblast growth factor,epidermal growth factor, insulin andthe insulin-like growth factors I andII. Journal of Neuroscience, 10:558-570,1990.
Lozsadi, D. A. Organization of corti-cal afferents to the rostral, limbic sec-tor of the rat thalamic reticularnucleus. Journal of Comparative Neu-rology, 341:520-533,1994.
Lukas, R.J., and Bencherif, M. Hetero-geneity and regulation of nicotinicacetylcholine receptors. International
Review of Neurobiology, 34:25-131,1992.
Luntz-Leybman, V.; Bickford, P.C;and Freedman, R. Cholinergic gatingof response to auditory stimuli in rathippocampus. Brain Research,587:130-136,1992.
Madison, D.V., and Nicoll, R.A. Nor-epinephrine decreases synaptic inhi-bition in the rat hippocampus. BrainResearch, 442:131-138,1988.
Manzone, O.J.; Manabe, T.; andNicoll, R. Release of adenosine byactivation of NMD A receptors in thehippocampus. Science, 265:2098-2101,1994.
McCarley, R.W.; Shenton, M.E.;O'Donnell, B.F.; and Nestor, P.G.Uniting Kraepelin and Bleuler: Thepsychology of schizophrenia and thebiology of temporal lobe abnormali-ties. Harvard Review of Psychiatry,1:36-52,1993.
McDonald, J.W., and Johnstone, M.V.Physiological and pathophysiologicalroles of excitatory amino acids dur-ing central nervous system develop-ment. Brain Research Reviews, 15:41-70,1990.
McGlashan, T.H., and Fenton, W.S.The positive-negative distinction inschizophrenia: Review of natural his-tory validators. Archives of GeneralPsychiatry, 49:63-72,1992.
McGue, M., and Gottesman, I.I.Genetic linkage in schizophrenia:Perspectives from genetic epidemiol-ogy. Schizophrenia Bulletin, 15(3):453-464,1989.
Meehl, P.E. Schizotaxia, schizotypy,schizophrenia. American Psychologist,17:827-838,1962.
Menza, M.A.; Grossman, N.; VanHorn, C; Cody, R.; and Forman, N.Smoking and movement disorders inpsychiatric patients. Biological Psychi-atry, 30:109-115,1991.
by guest on July 14, 2011schizophreniabulletin.oxfordjournals.org
Dow
nloaded from

444 SCHIZOPHRENIA BULLETIN
Metherate, R., and Ashe, J.H.Nucleus basalis stimulation facili-tates thalamocortical synaptic trans-mission in the rat auditory cortex.Synapse, 14:132-143,1993.
Miller, C, and Freedman, R. Medialseptal neuron activity in relation toan auditory sensory gating para-digm. Neuroscience, 55:373-380, 1993.
Miller, C, and Freedman, R. Theactivity of hippocampal intemeuronsand pyramidal cells during theresponse of the hippocampus torepeated auditory stimuli. Neuro-science, 69:371-381,1995.
Mitrofanis, J., and Guillery, R.W. Newviews of the thalamic reticularnucleus in the adult and the develop-ing brain. Trends in Neurosciences,16:240-245,1993.
Nagamoto, H.T.; Adler, L.E.; Hea,R.A.; Griffith, J.M.; McRae, K.A.; andFreedman, R. Gating of auditory P50in schizophrenics: Unique effects ofclozapine. Biological Psychiatry, inpress.
Pfeiffer, C.C., and Jenny, E.H. Theinhibition of the conditioned re-sponse and the counteraction ofschizophrenia by muscarinic stimula-tion of the brain. Annals of the NewYork Academy of Sciences, 66:753-764,1957.
Reite, M.; Teals, P.; Zimmerman, J.E.;Davis, K.; Whalen, J.; and Edrich, J.Source origin of a 50-msec latencyauditory evoked field component inyoung schizophrenic men. BiologicalPsychiatry, 24:495-506,1988.
Rollins, YD.; Stevens, K.E.; Harris,K.R.; Hall, M.E.; Rose, G.M.; andLeonard, S. Reduction in auditorygating following inrracerebroventric-ular application of a-bungarotoxinbinding site ligands and a7 antisenseoligonudeotides. Society of Neuro-science Abstracts, 19:837,1993.
Rosenthal, R., and Bigelow, L.G. The
effects of physostigmine in pheno-thiazine resistant chronic schizo-phrenic patients: Preliminary obser-vations. Comprehensive Psychiatry,14:489-494,1973.
Sarkar, G.; Kapelner, S.; Grandy, D.K.;Marchionni, M.; Civelli, O.; Sobell, J.;Heston, L.; and Sommer, S.S. Directsequencing of the dopamine D2receptor (DRD2) in schizophrenicsreveals three polymorphisms but nostructural change in the receptor.Genomics, 11:8-14,1991.
Schoepfer, R.; Conroy, W.G.; Whiting,P.; Gore, M.; and Lindstrom, J. Brain-bungarotoxin binding protein cDNAsand MAbs reveal subtypes of thisbranch of the ligand-gated ion chan-nel gene superfamily. Neuron,5:35-^8,1990.Schorte, A.; Janssen, PR; Megens,A. A.; and Leysen, J.E. Occupancy ofcentral neurotransmirter receptors byrisperidone, clozapine, and haloperi-dol, measured ex vivo by quantita-tive autoradiography. Brain Research,631:191-202,1993.
S<§gu<§la, P.; Wadiche, J.; Dineley-MiUer, K.; Dani, J.A.; and Patrick, J.W.Molecular cloning, functional proper-ties, and distribution of rat brain a7:A nicotinic cation channel highly per-meable to calcium. Journal of Neuro-science, 13:596-604,1993.
Siegel, C; Waldo, M.; Mizner, G.;Adler, L.E.; and Freedman, R. Deficitsin sensory gating in schizophrenicpatients and their relatives. Archives ofGeneral Psychiatry, 41:607-612,1984.
Simpson, G.V., and Knight, R.T. Mul-tiple brain systems generating the ratauditory evoked potential: II. Disso-ciation of auditory cortex and non-lemniscal generator systems. BrainResearch, 602:251-263,1993.
Sirota, P.; Frydman, M.; and Sirota, L.Schizophrenia and Marfan syn-drome. British Journal of Psychiatry,157:433-436,1990.
Snider, W.D., and Johnson, E.M., Jr.Neurorrophic molecules. Annals ofNeurology, 26:489-506,1989.
Stafekhina, V.S., and Vinogradova,O.S. Sensory characteristics of thecortical input to the hippocampus:The entorhinal cortex. Zhurnal VyssheiNervnoi Deiatelnosti Imeni I. P. Pavlova,25:119-127,1975.
Stevens, D.R., Birnstiel, S.; Gerber, U.;McCarley, R.W.; and Greene, R.W.Nicotine depolarizations of ratmedial pontine reticular formationneurons studied in vitro. Neuro-science, 57:419-424,1993.
Stevens, K.E.; Freedman, R.; Collins,A.C.; Hall, M.; Leonard, S.; Marks,M.J.; and Rose, G.M. Genetic correla-tion of inhibitory gating of hippo-campal auditory evoked responseand a-bungarotoxin-binding nico-tinic cholinergic receptors in inbredmouse strains. Neuropsychopharma-cology, in press.Stevens, K.E.; Meltzer, J.; and Rose,G.M. Nicotinic cholinergic normal-ization of amphetamine-induced lossof auditory gating in freely-movingrats. Psychopharmacology, 119:163-170,1995.Straumanis, J.J.; Shagass, C.; and Roe-mer, R.A. Influence of antipsychoticand antidepressant drugs on evokedpotential correlates of psychosis. Bio-logical Psychiatry, 17:1101-1122,1982.
Suddath, R.L.; Christison, G.W.; Tor-rey, E.F.; Casanova, M.F.; and Wein-berger, D.R. Anatomical abnormali-ties in the brains of monozygotictwins discordant for schizophrenia.New England Journal of Medicine,322:789-794,1990.
Swerdlow, N.R.; Braff, D.L.; Taaid,N.; and Geyer, M.A. Assessing thevalidity of an animal model of defi-cient sensorimotor gating in schizo-phrenic patients. Archives of GeneralPsychiatry, 51:139-154, 1994.
by guest on July 14, 2011schizophreniabulletin.oxfordjournals.org
Dow
nloaded from

VOL. 22, NO. 3, 1996 445
Tandon, R., and Greden, J.F. Cholin-ergic hyperactivity and negativeschizophrenic symptoms. Archiixs ofGeneral Psychiatry, 46:745-753,1989.
Tsuang, M.T. Genotypes, phenotypes,and the brain: A search for connec-tions in schizophrenia. British Journalof Psychiatry, 163:299-307,1993.
Venables, P. Input dysfunction inschizophrenia. In: Maher, B.A., ed.Progress in Experimental PersonalityResearch. New York, NY: AcademicPress, 1964. pp. 1-47.Vinogradova, O. Functional organi-zation of the limbic system in theprocess of registration of information.In: Issacson, R.L., and Pribram, K.H.,eds. The Hippocampus: Neurophysiol-ogy and Behavior. Vol. 1. New York,NY: Plenum Press, 1975. pp. 3-69.
Waldo, M.C.; Carey, G.; Myles-Wors-ley, M.; Cawthra, E.; Adler, L.E.; Nag-amoto, H.T.; Wender, P.; Byerley, W.;Plaetke, R.; and Freedman, R. Codis-tribution of a sensory gating deficitand schizophrenia in multi-affectedfamilies. Psychiatry Research,39:257-268,1991.
Waldo, M.C.; Cawthra, E.; Adler,L.E.; Dubester, S.; Staunton, M.; Nag-amoto, H.T.; Baker, N.; Madison, A.;Simon, J.; Scherzinger, A.; Drebing,C; Gerhardt, G.; and Freedman, R.Auditory sensory gating, hippocam-pal volume, and catecholaminemetabolism and schizophrenics andtheir siblings. Schizophrenia Research,12:93-106,1994.
Waldo, M.; Gerhardt, G.; Baker, N.;Drebing, C; Adler, L.E.; and Freed-
man, R. Auditory sensory gating andcatecholamine metabolism in schizo-phrenic and normal subjects. Psychia-try Research, 44:21-32,1992.Wilson, C.L.; Babb, T.L.; Halgren, E.;Wang, M.L.; and Crandall, PH.Habituation of human limbic neu-ronal response to sensory stimula-tion. Experimental Neurology, 84:74-97,1984.
Wonnacott, S. a-Bungarotoxin bindsto low-affinity nicotine binding sitesin rat brain. Journal of Neurochemistry,47:1706-1712,1986.
Acknowledgments
This research was supported byUSPHS grant MH-44212 from theNational Institute of Mental Health,DA-09457 from the National Instituteon Dmg Abuse, and by the VeteransAdministration Medical ResearchService.
The Authors
Sherry Leonard, Ph.D., is AssistantProfessor, Department of Pharma-cology, University of ColoradoHealth Sciences Center and DenverVeterans Affairs Medical Center, Den-ver, CO. Catherine Adams, Ph.D.,and Charles R. Breese, Ph.D., areInstructors, Department of Psychia-try, University of Colorado HealthSciences Center, Denver, CO.Lawrence E. Adler, M.D., is AssociateProfessor, Department of Psychiatry,
and Paula Bickford, Ph.D., is Assis-tant Professor, Department of Phar-macology, University of ColoradoHealth Sciences Center and DenverVeterans Affairs Medical Center, Den-ver, CO. William Byerley, Ph.D., isAssociate Professor and Hilary Coon,Ph.D., is Assistant Professor, Depart-ment of Psychiatry, University ofUtah Medical Center, Salt Lake City,UT. Jay M. Griffith, M.D., is AssistantProfessor, Department of Psychiatry,Southwestern University School ofMedicine and Dallas Veterans AffairsMedical Center, Dallas, TX. ChristineMiller, Ph.D., is a Graduate Student,Department of Psychiatry, Universityof Colorado Health Sciences Center,Denver, CO. Marina Myles-Worsley,Ph.D., is Assistant Professor, Depart-ment of Psychiatry, University ofUtah M«dical Center, Salt Lake City,UT. Herbert T. Nagamoto, M.D., isAssistant Professor, Department ofPsychiatry, University of ColoradoHealth Sciences Center and DenverVeterans Affairs Medical Center, Den-ver, CO. Yvonne Rollins is a GraduateStudent, Department of Pharmacol-ogy, and Karen E. Stevens, Ph.D., isAssistant Professor, Department ofPsychiatry, University of ColoradoHealth Sciences Center, Denver, CO.Merilyne Waldo, Ph.D., is AssistantProfessor, Department of Psychiatry,and Robert Freedman, M.D., is Pro-fessor, Departments of Psychiatryand Pharmacology, University ofColorado Health Sciences Center andDenver Veterans Affairs MedicalCenter, Denver, CO.
by guest on July 14, 2011schizophreniabulletin.oxfordjournals.org
Dow
nloaded from



















