
A novel angiogenic pathway mediated by non-neuronal nicotinic acetylcholine receptors
Upload
independentCategory
view
0download
0
Journal of Neurocytology 33, 543–556 (2004)
Ablation of primary afferent terminals reducesnicotinic receptor expression and the nociceptiveresponses to nicotinic agonists in the spinal cordIMRAN M. KH AN 1,∗ , MI CHELLE WENNERHOLM 1, ERINSINGLETARY 1, KI MBER LEY P O LSTON 1, LIMIN ZHANG 1,TOM DEERIN CK 2, TO N Y L. YAKS H 1,3 and PALMER TAYLOR 1
1Department of Pharmacology University of California, San Diego, CA 92093-0636, USA; 2National Center for Microscopy and ImagingResearch University of California, San Diego, CA 92093-0636, USA; 3Department of Anesthesiology, University of California, SanDiego, CA 92093-0636, [email protected]
Received 22 June 2004; revised 31 January 2005; accepted 2 February 2005
Abstract
A variety of studies indicate that spinal nicotinic acetylcholine receptors modulate the behavioral and autonomic responseselicited by afferent stimuli. To examine the location of and role played by particular subtypes of nicotinic receptors in mediatingcardiovascular and nociceptive responses, we treated neonatal and adult rats with capsaicin to destroy C-fibers in primaryafferent terminals. Reduction of C-fiber terminals was ascertained by the loss of isolectin B4, CGRP and vanilloid receptorsas monitored by immunofluorescence. Receptor autoradiography shows a reduction in number of epibatidine binding sitesfollowing capsaicin treatment. The reduction is particularly marked in the dorsal horn and primarily affects the class of highaffinity epibatidine binding sites thought to modulate nociceptive responses. Accompanying the loss of terminals and nicotinicbinding sites were significant reductions in the expression of α3, α4, α5, β2 and β4 nicotinic receptor subunits in the superficiallayers of the spinal cord as determined by antibody staining and confocal microscopy. The loss of nicotinic receptors thatfollows capsaicin treatment results in attenuation of the nociceptive responses to both spinal cytisine and epibatidine. Capsaicintreatment also diminishes the capacity of cytisine to desensitize nicotinic receptors mediating nociception, but it shows littleeffect on intrathecal nicotinic agonist elicited pressor and heart rate responses. Hence, our data suggest that α3, α4, α5, β2 andβ4 subunits of nicotinic receptors are localized in the spinal cord on primary afferent terminals that mediate nociceptive input.A variety of convergent data based on functional studies and subunit expression suggest that α3 and α4, in combination withβ2 and α5 subunits, form the majority of functional nicotinic receptors on C-fiber primary afferent terminals. Conversely, spinalnicotinic receptors not located on C-fibers play a primary role in the spinal pathways evoking spinally coordinated autonomiccardiovascular responses.
Abbreviations
IT: Intrathecal, SBP: systolic blood pressure, NeuN B: neuronal nuclei B, IB4: isolectin B4, CGRP: calcitonin gene related peptide.
Introduction
Intrathecal administration of nicotinic agonists elic-its distinctive cardiovascular and sensory responsesthrough stimulation of a variety of nicotinic receptorsubtypes (Buccafusco, 1996; Khan et al., 1994a, 1996,2001). Thus, nicotine, cytisine and epibatidine, in se-quentially increasing potencies, cause transient pres-sor, tachycardia and nociceptive responses that can bepreferentially blocked by selective nicotinic antagonists(Khan et al., 1997, 1998, 2001). In addition to cardio-
∗To whom correspondence should be addressed.
vascular and nociceptive responses, epibatidine andA-85380 also produce transient analgesia following in-trathecal administration (Khan et al., 2001). The noci-ceptive response elicited by spinal nicotinic agonistsmay result in reflexly driven cardiovascular responses.Our previous work also suggests that intrathecally ad-ministered nicotinic agonists directly stimulate pregan-glionic sympathetic neurons to increase blood pressureand heart rate (Khan et al., 1994b) that is mediated by
0300–4864 C© 2005 Springer Science + Business Media, Inc.

544 KHAN, WENNERHOLM, SINGLETARY ET AL.
multiple nicotinic receptor subtypes (Khan et al., 1994c,1997). This finding is consistent with a multiplicity ofspinal nicotinic receptor subtypes as shown in ligandbinding studies and by the localization of the subunitsin various spinal neurons and terminals (Khan et al.,2003). Furthermore, our previous studies indicate thatalpha-bungarotoxin does not influence the responseselicited by spinal nicotinic agonists (Khan et al., 1994a,b), indicating a limited involvement of α7 subunit inspinal nicotinic responses.
Autoradiography and immunohistochemical studiesshow that nicotinic ligand binding and the expressednicotinic receptor subunits α3, α4, α5, β2 and β4 pre-dominate in the superficial layer of the dorsal hornof the spinal cord (Khan et al., 2003). An importantquestion relates to the terminals with which these sub-units are associated. Although small, primary afferentC fibers project primarily into the superficial dorsalhorn (Lawson, 1992) and appear to express nicotinicreceptors (Roberts et al., 1995; Khan et al., 2003), otherterminals also arise from spino-fugal projection neu-rons, excitatory and inhibitory interneurons, and bul-bospinal projection terminals (Li et al., 1999). Moreover,studies suggesting localization of nicotinic ligand bind-ing on C fiber terminals have not identified the specificneuronal nicotinic receptor subunits expressed on theseterminals. To address these issues, we have employedcapsaicin to destroy a population of nociceptive pri-mary afferent terminals in order to correlate the lossof these capsaicin-sensitive primary afferents with: (i)changes in expression of nicotinic agonist binding sitesand neuronal nicotinic receptor subunits, and (ii) al-terations in spinal nicotinic agonist-elicited nociceptiveand autonomic responses. This approach has enabledus to delineate neuronal nicotinic receptor subtypes inspinal cord and to correlate their localization withinthe dorsal horn of the spinal cord with the mediationof nociceptive and cardiovascular responses.
Materials and methods
EXPERIMENTAL ANIMALS
Male Sprague-Dawley rats (300–350 g) and timed pregnantrats were purchased from Harlan Co. (Indianapolis, IN). Allanimals were housed in the animal facility and were main-tained on 12-h light/dark cycles. They received standard Pu-rina Rat Chow and water ad libitum. All studies were carriedout according to protocols reviewed and approved by theUCSD Institutional Animal Care Committee.
SELECTIVE DESTRUCTION OF SMALL PRIMARYAFFERENT TERMINALS
Systemic capsaicin treatment of neonatal animals and priorintrathecal (IT) capsaicin treatment of adult animals em-ployed distinct protocols to destroy C-fiber primary afferentterminals.
On the second day after birth, rats, usually one-half ofthe litter, were treated with 50 mg/kg body weight of cap-saicin subcutaneously. Capsaicin solutions were made in 10%ethanol, 10% Tween-80 in sterile saline and administered as aninjection of 5 µl/gm body weight. Control rats received injec-tion of vehicle only. After injection, the neonates were wipedwith moist Kim-wipes and then returned to their mother. At11 weeks of age, intrathecal catheters were chronically im-planted into the thecal space for spinal drug delivery as de-scribed previously (Khan et al., 1994a) and tested four to fivedays following IT catheter placement. Control rats used forreceptor autoradiography and immunohistochemistry werenot implanted.
To achieve C-fiber degeneration by treatment of adult an-imals, eleven-week-old rats were implanted with intrathecalcatheters. Four to five days following catheter implantation,the rats were re-anesthetized and treated intrathecally with60 µg of capsaicin in an injection volume of 10 µl (Khan et al.,1996). Control rats received vehicle intrathecally. Five to sevendays following capsaicin-treatment, cardiovascular and no-ciceptive responses to IT nicotinic agonists were examined.Separate groups of IT capsaicin treated and control rats wereused for immunohistochemical and receptor autoradiogra-phy assays.
SURGICAL PROCEDURES, ARTERIAL BLOOD PRESSUREAND HEART RATE RECORDINGS
For both the neonatal and IT capsaicin treated rats, tail ar-teries were catheterized to monitor blood pressure and heartrate as previously described (Khan et al., 1994a). Nicotinic ag-onists were administered intrathecally according to a previ-ously reported procedure (Khan et al., 1996). Cardiovascularparameters were recorded with a Gould-Polygraph and dataanalyzed with the Ponemah Physiology Platform-3 (GouldInstruments Systems, Valley View, OH).
Nociceptive or irritation responses to intrathecal nicotinicagonists were quantitated by a scoring index that was trans-formed to irritation intensity (Khan et al., 1996, 2002) by thefollowing equation:
Irritation Intensity/min, I =n∑
i=1
f xi
where, xi = the irritation score, f = frequency of irritationwithin a specific time period, i = 1 to 4.
Total irritation intensities were estimated by summing the1 min time interval intensities over 10 min following nicotinicagonist administration. Areas under the intensity-time curvewere plotted in 1 min intervals or for the entire 10 min pe-riod. They provided an estimate of the maximum irritationresponse in the period (usually 10 min) following nicotinicagonist administration.
RECEPTOR AUTORADIOGRAPHY
Procedures for tissue preparation, binding assays, emulsiondipping, processing, imaging and post-collection processingof images have been described (Khan et al., 1996, 2003). Briefly,16 µm thick lumbar and thoracic spinal cord transverse sec-tions were mounted on glass slides. The slides were incu-bated in assay buffer containing either 0.05 nM or 0.8 nMof [3H]-epibatidine for 60 min at 4◦C. Slides were washed in

Nicotinic receptors and spinal afferent neurons 545
ice-cold assay buffer followed by two dips in ice-cold deion-ized distilled water. Nonspecific binding was determined byco-incubation of tissue sections with [3H]-epibatidine and 60µM of l-nicotine. Dried slides were then dipped in Kodakphoto emulsion (NTB-2, 1:1 with water), stored in light-tightboxes for 10–12 weeks, developed and fixed. Alternatively,some slides were placed against Amersham [3H]-Hyperfilmfor 8–10 weeks, and then the films were hand processed.
Silver grain images of tissue sections obtained with a KodakSpot digital camera were then processed. Autoradiograms onfilms were scanned and processed with Adobe Photo Shop.Autoradiogram intensities between control and capsaicin-treated rats were compared with the aid of Image ProPlus(Media Cybernetics, Inc.) and NIH-Image softwares. For eachcomparison of paired samples done in the same experiment,spinal sections from capsaicin-treated and control rats werematched for treatment, post-tissue collection manipulations,assay protocol, emulsion dipping and grain development.Differences in grain counts or autoradiogram intensities fromtreated and control rats were analyzed from matched areas forstatistical significance by paired t-test. A one-tailed ‘p’ valueof <0.05 was considered as significant.
IMMUNOFLUORESCENCE AND CONFOCALMICROSCOPY
Antibodies to nAChR subunitsAntibodies to neuronal nAChR subunits (α3, α4, α5, β2 andβ4) were raised in rabbits using either synthetic peptidefragments (22 to 26 amino acids in length within the cyto-plasmic domain between transmembrane segments 3 and 4)or GST-nicotinic receptor subunit (cytoplasmic domain be-tween transmembrane segments 3 and 4) proteins (Khan etal., 2003). The polyclonal antibodies were affinity purified bycoupling the antigen to beads using an UltraLink Immobiliza-tion kit (Pierce). As determined by Western blot analysis andimmunohistochemical assays, these antibodies exhibit speci-ficity for the particular subunit and subunit sequence to whichthey were generated (Khan et al., 2003).
Immunohistochemical assaysThe procedure for immunohistochemical assays for confocalmicroscopy has been described previously (Khan et al., 2003).Briefly, spinal cord tissue fixed in 4% paraformaldehyde wassectioned (100 µm thick) with a Vibratome. Tissue sectionswere incubated overnight with anti-nAChR subunit antibod-ies (∼1 µg/ml). Other antibodies used in the assay were:anti-calcitonin-gene related peptide (CGRP) (1:200 of stock,Chemicon, CA), anti-neuron specific nuclear protein (NeuNB) (1:200 of stock, Chemicon, CA), and anti-vanilloid receptorprotein (TRPV1, Guo, 1991) (1:200 of stock, Novus Biological,CO). The primary afferent nerve terminal marker, GriffoniaSimplicifolia isolectin B4 (IB4) conjugated to fluorescein isoth-iocyanate (FITC), was used at recommended concentrations(Sigma, St. Louis, MO).
After incubation with primary antibodies, the sectionswere incubated with appropriate secondary antibodies (1:75in buffer) conjugated to CyTM5 (Cyanine), FITC or Lissamine-Rhodamine fluorescence probes (Jackson Immuno Research,PA). Immunofluorescence labeling of the spinal tissue was ob-served using a Bio-Rad MRC-1024 confocal system attached
to a Zeiss Axiovert 35 M microscope. The collected imageswere processed using Adobe Photo Shop. Immunostainingby various antibodies to the control and treated rat spinalsections were quantitated by the same protocol as previouslyindicated for comparing the autoradiograms (grain counts).For each comparison of paired samples conducted in the sameexperiment, spinal sections from capsaicin-treated and con-trol rats were matched for treatment, post-tissue collectionmanipulations and immunoassay protocols. To compare flu-orescence intensities between two images, images were col-lected with the same laser intensity and gain setting on thesame day. Moreover, the images were taken as single opticalsections with identical confocal aperture setting. Post-capturemanipulation of the images for gray levels and contrast wereconducted with identical settings. Differences in label intensi-ties (mean pixel values/area) of the anti-nAChR subunit anti-bodies between capsaicin-treated and paired control rats werecompared using the statistical criteria described for recep-tor autoradiography (see above). As samples did not exhibitsignificant changes in label intensities (for a particular anti-body) when scanned at different times or with different mi-croscopes, we believe that no significant photobleaching wasobserved in this time frame. Differences in mean pixel valuesfrom treated and control rats were analyzed from matchedareas for statistical significance by paired t-test. A one-tailed‘p’ value of <0.05 was considered as significant.
Results
SPINAL C-FIBER PRIMARY AFFERENT TERMINALMARKERS AND CAPSAICIN TREATMENT
Consistent with the location of C-fiber primary afferentterminals in dorsal spinal horn, a dense band of eitherIB4 binding sites or CGRP immunoreactivity (Figs. 1(insets A and B), 2 (insets A and B) and 3 (insets B, C, D,F and G)) and anti-vanilloid receptor protein (TRPV1)binding (Fig. 3: insets A and E) was observed in the su-perficial layer of the dorsal lumbar spinal cord (Khanet al., 2003; Guo et al., 1999). In neonatal rats treatedwith systemic capsaicin or adult rats receiving intrathe-cal capsaicin delivery, the band of IB4 sites, CGRP orTRPV1 immunoreactivity was essentially absent in thesame region as compared to vehicle-treated control rats.Anti-CGRP, anti-TRPV1 protein and IB4 binding fluo-rescence intensities in capsaicin-treated and control ratsare quantitated in Table 1. Intrathecal capsaicin treat-ment in adult rats results in greater than 60% reductionin IB4 binding sites and almost 75% reduction in CGRPimmunoreactivity compared to control rats.
SPINAL NICOTINIC AGONIST BINDING SITESAND CAPSAICIN TREATMENT
Densities of nicotinic agonist binding sites were deter-mined by receptor autoradiography. As indicated inFigure 1 (G–N), [3H]-epibatidine at 0.05 nM showedpredominant binding in the dorsal horn of the spinalcord. Following neonatal capsaicin-treatment a loss ofbinding sites primarily in the superficial dorsal horn,
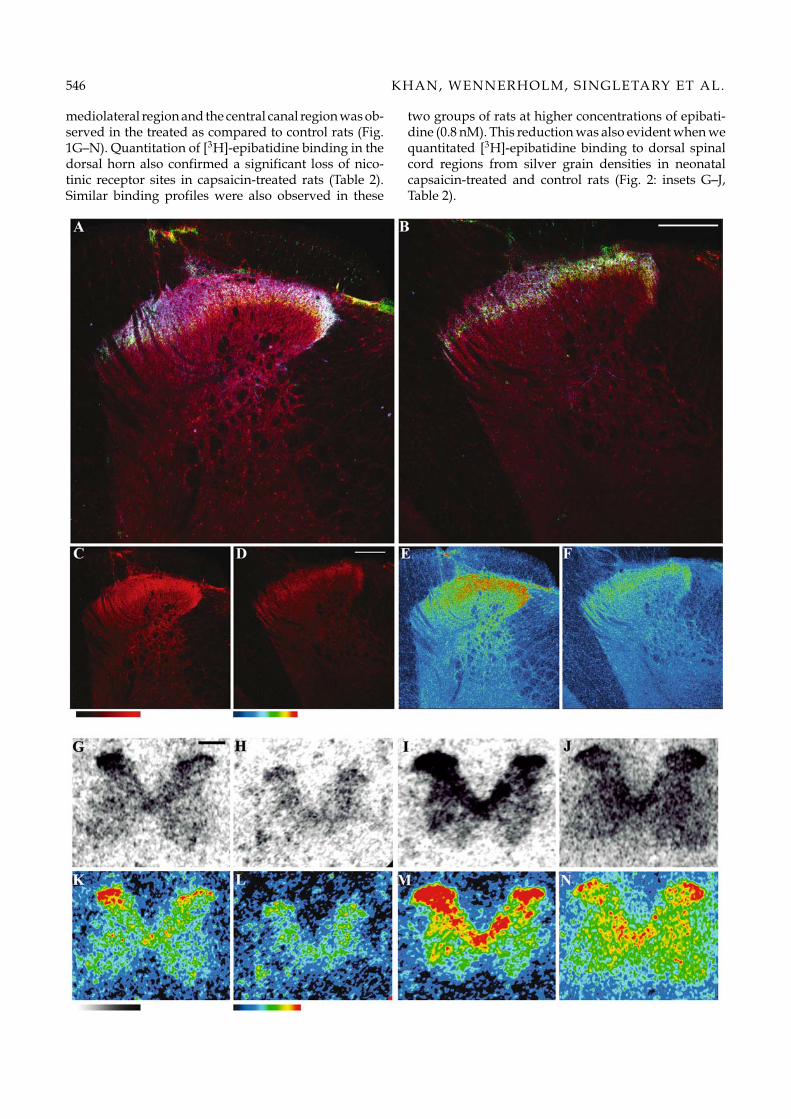
546 KHAN, WENNERHOLM, SINGLETARY ET AL.
mediolateral region and the central canal region was ob-served in the treated as compared to control rats (Fig.1G–N). Quantitation of [3H]-epibatidine binding in thedorsal horn also confirmed a significant loss of nico-tinic receptor sites in capsaicin-treated rats (Table 2).Similar binding profiles were also observed in these
two groups of rats at higher concentrations of epibati-dine (0.8 nM). This reduction was also evident when wequantitated [3H]-epibatidine binding to dorsal spinalcord regions from silver grain densities in neonatalcapsaicin-treated and control rats (Fig. 2: insets G–J,Table 2).

Nicotinic receptors and spinal afferent neurons 547
Table 1. Comparisons of specific C-fiber afferent terminal markers and nicotinic receptor subunit expression in dorsal lumbarspinal horn region between capsaicin-treated and control rats.
% ReductionControl Capsaicin versus controls ‘P’ value
Systemic treatment of neonatal ratsIB4 0.66 ± 0.03 0.23 ± 0.03 64% <0.001CGRP 0.58 ± 0.05 0.34 ± 0.06 37% <0.004
Intrathecal treatment of adult ratsIB4 0.83 ± 0.53 0.32 ± 0.05 60% <0.001CGRP 0.75 ± 0.09 0.26 ± 0.07 75% <0.001Vanilloid receptor protein 0.74 ± 0.06 0.38 ± 0.04 49% <0.001
Intrathecal treatment of adult ratsα3 0.80 ± 0.05 0.57 ± 0.04 27%† <0.001α5 0.81 ± 0.09 0.57 ± 0.10 30% <0.004β4 0.27 ± 0.02 0.19 ± 0.02 28%¶ <0.05β2 0.73 ± 0.03 0.45 ± 0.06 40% <0.002α4 0.95 ± 0.05 0.49 ± 0.04 50% <0.001
Each value, shown as a mean ± sem, represents the mean pixel intensity (0 to 255 pixels scale) per unit area in the fluorescence microscopyimages from superficial dorsal horn region of capsaicin-treated and vehicle-treated control rats. The treated and control groups were comparedby paired t-test. Each value represents at least 9 pairs of comparisons. Percent reduction indicates the reduction in fluorescence intensityin capsaicin-treated spinal region images compared to the same in control rats (approximate values). The percent reduction detected by therespective antibodies for α4 was significantly greater than the reduction for α3 († p < 0.01) and β4 (¶ p < 0.04) in capsaicin-treated comparedto control rats as determined by t-test. No significant differences in percent reduction between any other subunit pair were observed.
Receptor autoradiography also revealed a parallelreduction of [3H]-epibatidine binding in the IT cap-saicin treated rats (Table 2). Since the extents of de-pletion of C-fiber primary afferent terminals and re-ductions in [3H]-epibatidine binding sites were thesame between IT capsaicin-treated and neonatal cap-saicin treated rats, our subsequent studies focused onIT capsaicin-treatment. IT-treatment required signifi-cantly shorter treatment periods and was believed toreduce the likelihood of compensatory changes duringdevelopment.
SPINAL NICOTINIC RECEPTOR SUBUNIT DISTRIBUTIONAND CAPSAICIN TREATMENT
To determine the subtypes of nicotinic receptor sub-units expressed on primary afferent terminals, we ex-
Fig. 1. Panels A–F: Confocal microscopy images depicting immunofluorescence following incubation of lumbar spinal sectionsfrom vehicle-treated (panel A) and IT capsaicin-treated rats (panel B) with anti-nAChR α3-subunit (in red), anti-CGRP (in blue)and IB4 (in green). Panels C and D, and panels E and F are identical to the panels A and B, respectively, except they depictthe immunofluorescence to anti-α3 receptor subunits only (panels C and D) and artificial color-coded intensities of the actualfluorescence intensities of anti-α3 binding (panels E and F). White fluorescence in laminae I, II and III indicate co-localization ofthe three antigens in the control section (panel A), but colocalization is reduced substantially in the treated rats (panel B). Thecolor table gradient bar (right rectangular bar) corresponds to the red fluorescence intensity gradient bar (left rectangular bar),which represents zero to 255 pixels. White bars in panels B and D represent 200 µm. Panels G–N: Autoradiograms illustrating[3H]-epibatidine binding in neonatal vehicle-treated control (panels G, I, K and M) and capsaicin treated (panels H, J, L andN) rats. Panels G and H and panels I and J represent binding of 0.05 and 0.8 nM of [3H]-epibatidine, respectively, to lumbarspinal cord sections in the two groups of rats. Panels K and L, and panels M and N depict artificial color coded intensities ofthe actual autoradiograms in panels G and H, and in I and J, respectively. The rectangular bars in the bottom of the compositerepresent the color table gradient for the corresponding white to black gradient in the autoradiograms. The black bar in panelG represents 500 µm.
amined immunolabeling profiles for the various anti-nAChR subunit antibodies in capsaicin and controlrats by confocal fluorescence imaging microscopy. Fig-ures 1–3 illustrate the immunofluorescence by antibod-ies againstα3,α4,α5,β2 andβ4 subunits in IT capsaicin-treated and age-matched control rats. The specificitiesof these antibodies against their respective subunitshave been previously established (Khan et al., 2003).
Confocal fluorescence microscopy images of multi-labeled spinal cord sections in Figures 1 and 2 demon-strate that α3 nAChR subunits (red), CGRP (blue) andIB4 binding (green) sites are colocalized in the superfi-cial dorsal horn region. Overlap regions of fluorescenceemission of the three labels are indicated by the fluo-rescence emission of a white color in several regionsof the superficial layer of the dorsal lumbar spinal sec-tion (arrow marks, Fig. 2, panel A). The loss of IB4 and

548 KHAN, WENNERHOLM, SINGLETARY ET AL.
CGRP staining in capsaicin-treated rats correlates withsignificant reductions of α3 staining (Figs. 1 and 2, Ta-ble 1). This diminution is documented by the loss ofthe spatial overlap of fluorescence emission thereby di-minishing the fractional white fluorescence emission in
the capsaicin-treated rats as well as reducing the color-coded intensities of anti-α3 labeling (bottom panels).Although loss of staining for anti-α3 occurs throughout the dorsal horn (Fig. 1), the loss is most evident inthe superficial layer (Figs. 1 and 2).

Nicotinic receptors and spinal afferent neurons 549
Table 2. Autoradiographic intensities and grain densities in the superficial dorsal horn region in capsaicin-treated and controlrats.
High and low affinity sites High affinity sites
Capsaicin treatment Control Capsaicin Control Capsaicin
Autoradiogram intensities:Neonatal (systemic) 100 ± 4.8 74.1 ± 5.3∗ 100 ± 6.7 61.6 ± 2.9∗
Adult (intrathecal) 100 ± 6.7 62.9 ± 2.9∗ 100 ± 28.4 65.9 ± 2.9∗
Grain densities:Neonatal (systemic) 100 ± 7.1 81.3 ± 6.9∗ 100 ± 8.7 50.3 ± 6.9∗
Adult (intrathecal) ND ND ND ND
Each value, shown as a mean ± s.e.m., represents the percent intensity or grain density compared to the average intensity or grain density ofall of the control measurements in each group. *p < 0.05 as compared to control (paired t-test, one tailed), n = 3 to 6 pairs (control and treated)of animals in each treatment protocol. ND, not determined.
Similar to observations with the α3 subunit, im-munoreactivities for the other subunit targeted anti-bodies were also reduced. Quantification indicated thatthe ordering of reduction was α4 and β2 > α3, α5 andβ4 subunits in the superficial layer of the lumbar spinalcord in capsaicin-treated as compared to vehicle-treatedcontrol rats (Figs. 2 and 3, Table 1).
INTRATHECAL NICOTINIC AGONIST-ELICITEDRESPONSES AND CAPSAICIN TREATMENT
Panels A and B in Figure 4, illustrate the cardiovas-cular and nociceptive responses to sequential dosesof intrathecal cytisine and epibatidine in intrathecalcapsaicin-treated and control rats. The cardiovascularor nociceptive responses to each dose of the nicotinicagonist depict the maximum-recorded response withinthe 10 min interval following intrathecal administra-tion of the specified dose of nicotinic agonists. Whenmaximum responses are examined, no significant alter-ations in the pressor response following administrationof either nicotinic agonist are evident in the capsaicin-treated rats. However, capsaicin treated rats exhibitedaltered heart rate and nociceptive responses to certaindoses of cytisine (Fig. 4, panel A). The irritation re-sponses to both 0.5 and 5.0 µg of cytisine were sig-nificantly diminished in IT capsaicin-treated rats. Onlythe nociceptive response to various doses of spinal epi-
Fig. 2. Panels A–F: Confocal microscopy images depicting immunofluorescence following incubation of lumbar spinal sectionsfrom vehicle-treated controls (panel A) and capsaicin-treated rats (panel B) with anti-nAChR α3-subunit (in red), anti-CGRP(in blue) and IB4 (in green). Panels C and D, and panels E and F are identical to the panels A and B, respectively, except theydepict the immunofluorescence to anti-α3 receptor subunits only (panels C and D) and artificial color-coded intensities of theactual fluorescence intensities of anti-α3 binding (panels E and F). White fluorescence areas in laminae I, II and III indicateco-localization of the three antigens in the control section (arrow marks, panel A) and they are absent in the treated rats (panelB). The color table gradient bar (right rectangular bar) corresponds to the red fluorescence intensity gradient bar (left rectangularbar), which represents zero to 255 pixels. White bars in B and D represent 50 µm. Panels G–J: Dark-field photomicrographsdepicting [3H]-epibatidine binding as represented by silver grain densities in neonatal vehicle-treated control (panels G andI) and capsaicin-treated (panels H and J) rats. The upper and lower panels represent binding with low (0.05 nM, panels Gand H) and high (0.8 nM, panels I and J) concentrations of [3H]-epibatidine to dorsal horn of lumbar spinal cord regions. Therectangular box on the diagram of the cross-section of lumbar spinal cord indicates the area of the spinal cord shown in thephotomicrographs.
batidine was attenuated in the capsaicin-treated rats(Fig. 4, panel B). There was no significant difference be-tween capsaicin and control rats in the overall pressorresponse or tachycardia elicited at the doses of epibati-dine employed.
Temporal profiles of the cardiovascular and nocicep-tive responses to 0.5µg cytisine and 0.05µg epibatidine,respectively, were also compared between IT capsaicin-treated and control rats (Fig. 4, panels C and D). Boththe tachycardia and irritation responses were signifi-cantly ablated in the capsaicin-treated rats followingadministration of 0.5 µg of cytisine (Fig. 4, panel C). Ina separate group of IT capsaicin-treated rats, we alsoobserved a significant attenuation of the irritation re-sponse to 0.05 µg epibatidine. Again, there were no dif-ferences in the pressor response; however, tachycardiaelicited by the intermediate dose of epibatidine differedsignificantly between capsaicin-treated and control rats(Fig. 4, panel D).
To investigate the propensity for receptor desensiti-zation, we examined the effect of repeated exposure toagonist on the response elicited. As shown in Fig. 5 (leftpanels), the difference in nociceptive response betweencapsaicin-treated and control rats observed followinga single dose of 5 µg of cytisine disappears after thesecond dose of cytisine. Subsequent administration of0.5 µg epibatidine after the cytisine also did not showa difference in nociceptive response between the two

550 KHAN, WENNERHOLM, SINGLETARY ET AL.
groups of rats. However, using a similar experimen-tal design with epibatidine exposure, a significant dif-ference in the nociceptive responses between capsaicinand control rats was still evident after sequential epi-batidine dosing (Fig. 5, right panels).
Discussion
Both systemic capsaicin in neonatal rats or IT cap-saicin in adult rats resulted in significant reductions inIB4, anti-CGRP and anti-vanilloid receptor fluorescence

Nicotinic receptors and spinal afferent neurons 551
labeling found in the superficial dorsal horn. Parallel-ing the loss of the C-fiber primary afferent terminalsindicated by the loss of these three markers are sig-nificant reductions in [3H]-epibatidine binding sites incapsaicin-treated rats (Figs. 1 and 2). These observa-tions emphasize the prominent association of nicotinicreceptors with small primary afferents. This associa-tion permits us to determine which nicotinic receptorsubunits are in fact associated with reduction in epiba-tidine binding. Our current observations show that α3,α4, α5, β2 and β4 subunits of the nicotinic acetylcholinereceptor are expressed in the superficial dorsal horn.Following capsaicin-treatment, significant, but differ-ential reductions in expression of these five subunitsin the superficial dorsal horn region are evident. Thepresent study is the first to localize functional nicotinicreceptors as well as nicotinic receptor subunit proteinexpression to spinal C-fiber terminals in intact animals.To determine if these binding sites have functional rele-vance, we examined the nociceptive and cardiovascularresponses evoked by intrathecal cytisine and epibati-dine. Here, the loss of the epibatidine binding and nico-tinic receptor subunits is accompanied by a reductionof the evoked nociceptive responses and to a lesser ex-tent the cardiovascular responses of blood pressure andheart rate. These presynaptic effects mediated throughsmall TRPV1 bearing primary afferents are consistentwith previous in vitro studies. Puttfarcken et al. (1997)reported the release of the small afferent peptide sub-stance P by various nicotinic agonists in a clonal F11dorsal root ganglion (DRG) neuronal cell line.
NEONATAL AND INTRATHECAL CAPSAICIN ANDAFFERENT FIBER DEGENERATION
The primary afferent terminals in the dorsal lumbarspinal sections can be broadly classified into unmyeli-nated C-fiber and thinly myelinated A-fiber terminals.The C-fibers convey nociceptive and thermal stimuli toneurons in laminae I and II of the dorsal spinal horn,whereas the A-fibers convey mechanical or tactile sen-sory information typically to laminae III and IV withsome terminals also terminating in lamina I (Willis &Coggeshall, 1991). The strategy of the present stud-ies employs capsaicin to deplete selectively subpopula-
Fig. 3. Confocal microscopy images depicting immunofluorescence labeling in control (panels A–D) and IT capsaicin treated(panels E–H) rats following binding with anti-nAChR subunit antibodies (in red), anti-neuron specific nuclear protein (NeunB) (in blue) and IB4 (panels B–D and F–H, in green) or anti-vanilloid receptor protein (panels A and E, in green). In panelpairs A and E, B and F, C and G, and D and H, red color represents staining with anti-α4, anti-α5, anti-β2 and anti-β4 subunitantibodies, respectively. Panel pairs I and K, M and O, Q and R, and U and V are identical to panels A and E, B and F, C and G,and D and H, respectively except they represent fluorescence labeling by the anti-nAChR subunit only. Similarly, panel pairs Jand L, N and P, S and T, and W and X are identical to panel pairs A and E, B and F, C and G, and D and H, respectively exceptthey represent artificial color-coded intensities of the actual fluorescence intensities of anti-nAChR binding (panels A–H). Thecolor table gradient bar (right rectangular bar) corresponds to the red fluorescence intensity gradient bar (left rectangular bar),which represents zero to 255 pixels. White bars represent 50 µm.
tions of C-fibers without affecting the A-fiber terminals(Jancso & Lawson, 1990; Ribeiro-Da-Silva & Coimbra,1984).
These studies are predicated on the selective loss ofsmall afferents through the use of capsaicin treatmentin both neonatal and adult animals. With neonatal treat-ment compensatory changes can occur over time (Nagy& Hunt, 1983). These changes include (i) A-fiber ter-minals sprouting into lamina II (Mannion et al., 1998,Schicho et al., 1999). The interpretation of such workusing transport markers for large fibers is now ques-tioned (Bao et al., 2002; Hughes et al., 2003). (ii) Reten-tion of A-fiber terminals in lamina II, which are other-wise retracted to lamina III and IV during normal ag-ing of the animals (Torsney et al., 2000). (iii) Sproutingof serotonin terminals (Feldblum et al., 1998). With cap-saicin treatment in adult animals, it has been suggestedthat nerve terminals may be destroyed, but not the ax-ons or soma of dorsal root ganglion neurons (Chunget al., 1990). In spite of the differences in the models,our data indicate similar reductions in the multiple C-fiber markers occur after both systemic neonatal andIT capsaicin-treated rats and are consistent with previ-ously published studies (Jancso, 1981; Williams & Ziegl-gansberger, 1982; Jancso & Lawson, 1990). Accordingly,we feel confident that the results observed here reflecta principal loss of small afferent terminals.
LOSS OF PRIMARY AFFERENT TERMINALS AND[3H]-EPIBATIDINE BINDING IN RAT SPINAL CORD
Epibatidine binding typically displays high and lowaffinity binding components in spinal cord (Khan et al.,1997), which are believed to reflect the presence of atleast two distinct nicotinic binding sites. In the presentstudy, we show that such binding largely localizesin the superficial dorsal horn and that capsaicin re-sulted in a prominent reduction in both the high andlow affinity components. These observations supportlocalization of both components on the C fiber ter-minals. Overall, our results are consistent with ear-lier work in which a loss of nicotinic agonist bind-ing sites was observed in isolated membrane frac-tions from the spinal cord of systemic capsaicin-treatedrats (Roberts et al., 1995). Importantly, reductions in

552 KHAN, WENNERHOLM, SINGLETARY ET AL.
Fig. 4. Panels A and B: Changes in systolic blood pressure (SBP), heart rate (beats per minute) and irritation intensity responsesto intrathecal (IT) cytisine (0.05, 0.5 and 5 µg, panel A) and epibatidine (0.005, 0.05 and 0.5 µg, panel B) in IT capsaicin-treatedand vehicle-treated control rats. The responses record the maximum change in blood pressure and heart rate and the highestirritation intensity over a specific period following cytisine administration (4 min for 0.05 µg and 10 min for 0.5 and 5 µg cytisine).Each value represents the mean ± SEM. ∗p < 0.05 control versus capsaicin-treated rats at the specific dose as determined byone-way ANOVA. Panels C and D: Line graph depicting changes in SBP, heart rate and irritation responses over 10 min timeperiod following intrathecal administration of cytisine (0.5 µg, Panel C) and epibatidine (0.05 µg, Panel D) in IT capsaicin (opensymbols) and vehicle-treated control rats (closed symbols). The values represent maximum changes in SBP and heart rate andhighest irritation score recorded between each one-minute time interval. Each value represents the mean ± SEM. ∗∗∗ p < 0.001control versus IT capsaicin-treated rats as determined by two-way ANOVA over the 10 min interval.
[3H]-epibatidine binding occurred also in the deeperlayers of the dorsal horn and the central canal regionafter capsaicin. This finding is consistent with previousstudies indicating that some C-fiber primary afferentterminals, particularly those associated with visceralafferents, synapse in these spinal areas (Janig, 1996).
nAChR SUBUNIT EXPRESSION ON C-FIBER PRIMARYAFFERENT TERMINALS
Isolectin B4 and CGRP are markers for nonpeptider-gic C-fiber neurons and peptidergic C-fiber neurons,respectively (Silverman & Kruger, 1990). However, IB4may also bind to CGRP positive neurons (Averill et al.,1995), and CGRP is expressed in some A-fiber neurons(McCarthy and Lawson, 1990). We therefore employedthree markers: IB4 binding sites, CGRP and vanilloidreceptor proteins to identify the C-fiber neuronal ter-minals (Guo et al., 1999; Nasu, 1999; Ribeiro-da-Silva,1995). In control rats, we observed co-localization ofthe five nicotinic receptor subunits with CGRP, vanil-loid receptor or IB4 indicating that all five subunits ap-pear to be expressed on or near C-fiber primary af-
ferent terminals. Capsaicin-treatment resulted in sig-nificant reductions in fluorescence labeling by all fivenicotinic receptor subunit antibodies in the superficialdorsal horn area. The decreased fluorescence labelingis accompanied by substantial reductions in primaryafferent terminal markers in the same area.
Although expression of all five subunits is signif-icantly reduced in the dorsal lumbar spinal cord ofcapsaicin treated rats, fluorescence stains for the sub-units were not uniformly diminished. Accordingly, α3,α5 and β4 subunits showed limited reduction in expres-sion (≤30%, Table 2) compared to α4 and β2 subunitsthat showed a greater reduction (≥40%), but not com-plete ablation, in the capsaicin-treated than in controlrats. We previously demonstrated that except for the β4subunit, the other four subunits α3, α4, α5 and β2, havewell-defined expression on post-synaptic neurons inthe spinal cord (Khan et al., 2003). Thus, consistent withour previous observations, the present study indicatesthat a significant fraction of the immunofluorescencefor α3, α4, α5 and β2 antibodies in the superficial layeroccurs on non-C fiber terminals, and on inter-neuronslocated in spinal dorsal horn regions (Figs. 2 and 3).

Nicotinic receptors and spinal afferent neurons 553
Fig. 5. Line-graphs depicting nociceptive responses over a 10 min period following repeated administration of cytisine (5 µg, leftpanels) and epibatidine (0.5 µg, right panels) in IT capsaicin (open symbols) and vehicle-treated control (closed symbols) rats.In the left column of panels, top and middle panels present data following first and second dose of cytisine administered 25 minlater, whereas the bottom panel represents dosing by 0.5 µg of epibatidine following two doses of cytisine. In the right columnof panels, the top, middle and bottom panels present data following three consecutive doses of epibatidine every 25 mins. Eachvalue represents the mean ± s.e.m. ∗∗∗ p < 0.001 controls versus IT capsaicin-treated rats as determined by two-way ANOVAover 10 min interval.
Indeed, we have observed strong presynaptic stainingby α3, α5 and β4 subunits as compared to α4 and β2subunits in the superficial layer (Khan et al., 2003). Ac-cordingly, data from the present study are consistentwith the hypothesis that β4 subunit expression in thesuperficial layer is mainly presynaptic on C-fiber ter-minals. In as much as cholinergic neurons and termi-
nals have been identified in spinal dorsal horn regions(Khan et al., 2003; Olave et al., 2002; Naim et al., 1998;Schafer et al., 1998; Ribeiro-da-Silva & Cuello, 1990), lo-calization of nicotinic receptor subunits in the differentspinal neurons and terminals is important for under-standing the role of the nicotinic acetylcholine receptorsystem in pain processing.

554 KHAN, WENNERHOLM, SINGLETARY ET AL.
INFLUENCE OF CAPSAICIN-TREATMENT ON SPINALCYTISINE-ELICITED RESPONSES
Spinal administration of nicotinic agonists elicits au-tonomic (pressor and tachycardia) and nociceptive re-sponses (Khan et al., 1994a, 1998, 2001). These re-sponses are believed to reflect depolarization of spinalcircuitry, for example, the afferent terminal and/or apost-synaptic excitation of spinal projection neurons.The present studies, showing substantial localizationof nicotinic receptors on C fiber terminals, and the at-tenuation of the pain evoked behaviors by capsaicintreatment provide convergent support for nicotinic re-ceptors on C-fiber afferents playing a significant rolein nociception. In addition, the altered cardiovascularresponse to cytisine in capsaicin-treated compared tocontrol rats further supports our previous contentionthat some portion of nicotinic agonist-elicited tachy-cardia results from a reflex response to C-fiber afferentterminal stimulation (Khan et al., 1994, 1996, 2002).
Although the nociceptive response to both of thenicotinic agonists was significantly attenuated incapsaicin-treated, the residual nociceptive responsesuggests that nicotinic agonists may also bind to nico-tinic sites in other terminals and interneurons in thedorsal horn region of the spinal cord. This is supportedby the immunohistochemical studies where we observeexpression of nicotinic receptors in presynaptic termi-nals and interneurons in the superficial layer of the dor-sal spinal cord (Figs. 2 and 3, Khan et al., 2003). Otherstudies have also identified functional nicotinic recep-tors in non-primary afferent terminals in the superficiallayer of the dorsal horn (Kiyosawa et al., 2001; Takedaet al., 2003).
Repeated administration of cytisine resulted in dis-appearance of the differences in nociceptive responsesto subsequent nicotinic agonist dosing between cap-saicin and control rats. However, in rats receiving re-peated epibatidine administration, the difference in no-ciceptive responses between the capsaicin-treated andcontrol groups was still evident. This indicates thatnicotinic receptor subtypes expressed on primary af-ferent terminals exhibit different capacities for desensi-tization when compared to nicotinic receptors on nonC-fiber sites. Nicotinic receptors composed of α3 and β4subunits alone or in combination with α5 subunit showfaster rates of desensitization with cytisine compared toreceptors of α3 and β2 or other subunit compositions(Wang et al., 1996, 2002; Gerzanich et al., 1998). It wasalso observed that epibatidine has a higher affinity forα3 and β2 containing receptor subtypes than those com-posed of α3 and β4 (Wang et al., 1996, 2002; Gerzanichet al., 1998). These data, along with our present observa-tions, indicate that that α3 and α4 in combination withβ2 and α5 subunits form the majority of functional nico-tinic receptors on C-fiber primary afferent terminals.Thus, consistent with previous studies, these receptors
show higher affinity for epibatidine (Table 1), and ex-hibit a slower rate of desensitization by cytisine (Fig. 5).Reduction of these receptor subtypes by ablation of C-fiber terminals results in an action of nicotinic agonistonly on the residual subtypes expressed in the super-ficial layer (such as subtypes composed of α3, α5 andβ4 subunits). Consistent with this assumption, in pre-vious studies we observed that, while cytisine pretreat-ment desensitized epibatidine and A-85380 elicited no-ciceptive responses, it could distinguish between anti-nociceptive responses to these agonists. Epibatidine,but not A-85380, elicited anti-nociceptive responseswere desensitized by cytisine exposure. Epibatidineelicited antinociceptive responses involve nicotinic re-ceptors on C-fiber terminals, whereas, A-85380 requiresthe adrenergic bulbospinal terminals (Khan et al., 2001).
Overall, our data support the notion that, in addi-tion to C-fiber primary afferent terminals, nicotinic re-ceptors also reside on other sites in spinal pain cir-cuits. Stimulation of nicotinic receptors on excitatory in-terneurons may still evoke a significant nociceptive re-sponse in the absence of primary afferent terminals. In-terestingly, separate studies have also identified expres-sion of selective functional nicotinic receptor subunitsin interneurons in the superficial layer that are receptiveto the pain neurotransmitter, substance P (Khasabovet al., 2002, Khan et al., 2005, unpublished observa-tions). Furthermore, nicotinic receptors with their mul-tiple subtypes expressed in this region may be linkednot only to the release of excitatory neurotransmit-ters, but also to inhibitory neurotransmitters. Kiyosawaet al. (2001) have also demonstrated that nicotinic ago-nists stimulate pre-synaptic terminals of inhibitory in-terneurons to release glycine, an inhibitory transmitterthat may contribute to the analgesic response to certainspinal nicotinic agonists. As indicated above, cholin-ergic terminals have been identified in the superficiallayer. These terminals appear to be presynaptic to pri-mary sensory neurons (Khan et al., 2003; Olave et al.,2002; Schafer et al., 1998; Ribeiro-da-Silva & Cuello,1990). Thus, the potential role of nicotinic acetylcholinereceptor system in the spinal dorsal horn both as ‘vol-ume’ and ‘wire’ neurotransmitters is becoming increas-ingly evident.
In summary, our present observations provide evi-dence for functional nicotinic receptors on C-fiber pri-mary afferent terminals. The data also suggest that amajority of nicotinic receptor subtypes expressed onthe nociceptive sensory terminals have high affinity for[3H]-epibatidine. Although exact subunit compositionscould not be determined, the findings indicate that α3,α4, α5, β2 and β4 subunits are components of the highand low affinity nicotinic receptor subtypes expressedin the superficial layer of the spinal cord. Thus, similarto the observations of Flores et al. (1996) in trigeminalganglion, more than one subtype of nicotinic receptor

Nicotinic receptors and spinal afferent neurons 555
appear to be expressed on the primary afferent termi-nals. Future studies involving immunoprecipitation ofthe receptor oligomers will be required to determinethe subunit compositions of the nicotinic receptor sub-type(s) expressed on the primary afferent terminals.
Acknowledgments
Supported by USPHS Grant HL- 35018 and the Califor-nia Tobacco-related Disease Research Program to PT.
References
AVERILL, S., MCMAHON, S. B., CLARY, S. B.,REICHARDT, L. F. & PRIESTLEY, J. V. P. (1995)Immunocytochemical localization of trkA receptors inchemically identified subgroups of adult rat sensory neu-rons. European Journal of Neuroscience 7, 1484–1494.
BAO, L., WANG, H. F., CAI, H. J., TONG, Y. G., JIN, S.X., LU, Y. J., GRANT, G., HOKFELT, T. & ZHANG,X. (2002) Peripheral axotomy induces only very limitedsprouting of coarse myelinated afferents into inner lam-ina II of rat spinal cord. European Journal of Neuroscience16, 175–185.
BUCCAFUSCO, J. J. (1996) The role of central choliner-gic neurons in the regulation of blood pressure and inexperimental hypertension. Pharmacological Reviews 48,179–211.
CHUNG, K., KLEIN, C. M. & COGGESHALL, R. E. (1990)The receptive part of the primary afferent axon is mostvulnerable to systemic capsaicin in adult rats. Brain Re-search 511, 222–226.
FELDBLUM, S., ANOAL, M., LAPSHER, S.,DUMOULIN, A. & PRIVAT, A. (1998) Partial deaf-ferentation of the developing rat spinal cord delaysthe spontaneous repression of GAD67 mRNAs inspinal cells. Perspectives on Developmental Neurobiology 5,131–143.
FLORES, C. M., DECAMP, R. M., KILO, S., ROGERS, S.W. & HARGREAVES, M. (1996) Neuronal nicotinic re-ceptor expression in sensory neurons of the rat trigeminalganglion: Demonstration of α3β4, a novel subtype in themammalian nervous system. Journal of Neuroscience 16,7892–7901.
GERZANICH, V., WANG, F., KURYATOV, A. &LINDSTROM, J. (1998). Alpha 5 subunit alters desen-sitization, pharmacology, Ca++permeability and Ca++
modulation of human neuronal alpha 3 nicotinic recep-tors. Journal of Pharmacology and Experimental Therapeutics6, 311–320.
GUO, A., VULCHANOVA, L., LI, W. X. & ELDE, R. (1999)Immunocytochemical localization of the vanilloid recep-tor 1 (TRPV1): Relationship to neuropeptides, the P2X3purinoceptor and IB4 binding sites. European Journal ofNeuroscience 11, 946–958.
HUGHES, D. I., SCOTT, D. T., TODD, A. J. & RIDDELL,J. S. (2003) Lack of evidence for sprouting of A beta affer-ents into the superficial laminas of the spinal cord dor-sal horn after nerve section. Journal of Neuroscience 23,9491–9499.
JANCSO, G. (1981) Intracisternal capsaicin: Selective degen-eration of chemosensitive primary sensory afferents inthe adult rat. Neuroscience Letters 27, 41–45.
JANCSO, G. & LAWSON, S. N. (1990) Transganglionic de-generation of capsaicin-sensitive C-fiber primary affer-ent terminals. Neuroscience 39, 501–511.
JANIG, W. (1996) Neurobiology of visceral afferent neurons:Neuroanatomy, functions, organ regulations and sensa-tions. Biological Psychology 42(1/2), 29–51.
KHAN, I. M., BUERKLE, H., TAYLOR, P. & YAKSH, T.L. (1998) Nociceptive and antinociceptive responses tointrathecally administered nicotinic agonists. Neurophar-macology 37, 1515–1525.
KHAN, I. M., MARSALA, M., PRINTZ, M. P., TAYLOR,P. & YAKSH, T. L. (1996) Intrathecal nicotinic agonist-elicited release of excitatory amino acids as measured byin vivo spinal microdialysis in rats. Journal of Pharmacologyand Experimental Therapeutics 278, 97–106.
KHAN, I. M., OSAKA H., STANISLAUS, S., CALVO,R., DEERNICK T., YAKSH, T. L. & TAYLOR,P. (2003) Nicotinic acetylcholine receptor distribu-tion in relation to spinal neurotransmission pathways.Journal of Comparative Neurology 467, 44–59.
KHAN, I. M., SINGLETARY, E., ALEMMYU, A.,STANISLAUS, S., PRINTZ, M.P., YAKSH, T.L. &TAYLOR, P. (2002) Nicotinic receptor gene cluster onrat chromosome 8 in nociceptive and blood pressure hy-perresponsiveness. Physiological Genomics 11, 65–72.
KHAN, I. M., STANISLAUS, S., ZHANG, L., TAYLOR,P. & YAKSH, T. L. (2001) A-85380 and epibatidine eachinteract with disparate spinal nicotinic receptor subtypesto achieve analgesia and nociception. Journal of Pharma-cology and Experimental Therapeutics 297, 230–239.
KHAN, I. M., TAYLOR, P. & YAKSH, T. L. (1994a)Cardiovascular and behavioral responses to nico-tinic agonists administered intrathecally. Journal ofPharmacology and Experimental Therapeutics 270, 150–158.
KHAN, I. M., TAYLOR, P. & YAKSH, T. L. (1994b)Stimulatory pathways and sites of actions of in-trathecally administered nicotinic agonists. Journal ofPharmacology and Experimental Therapeutics 271, 1550–1557.
KHAN, I. M., YAKSH, T. L. & TAYLOR, P. (1997) Epiba-tidine binding sites and activity in the spinal cord. BrainResearch 753, 269–282.
KHAN, I. M., YAKSH, T. L. & TAYLOR, P. (1994c) Lig-and specificity of nicotinic acetylcholine receptors in ratspinal cord: Studies with nicotine and cytisine. Journal ofPharmacology and Experimental Therapeutics 270, 159–166.
KHASABOV, S. G., ROGERS, S. D., GHILARDI, J. R.,PETERS, C. M., MANTYH, P. W. & SIMONE,D. A. (2002) Spinal neurons that possess the sub-stance P receptor are required for the developmentof central sensitization. Journal of Neuroscience 2, 9086–9098.
KIYOSAWA, A., KATSURABAYASHI, S., AKAIKE, N.,PANG, Z. P. & AKAIKE, N. (2001) Nicotine facilitatesglycine release in the rat spinal dorsal horn. Journal ofPhysiology 536, 101–110.
LAWSON, S. N. (1992) Morphological and biochemical celltypes of sensory neurons. In Sensory Neurons, Diversity,

556 KHAN, WENNERHOLM, SINGLETARY ET AL.
Development and Plasticity (edited by Scott, S.A.), pp.27–59. New York: Oxford University Press.
LI, Y. Q., LI, H., KANEKO, T. & MIZUNO, N. (1999) Sub-stantia gelatinosa neurons in the medullary dorsal horn:An intracellular labeling study in the rat. Journal of Com-parative Neurology 411, 399–412.
MANNION, R. J., DOUBELL, T. P., GILL, H. & WOOLF,C. J. (1998) Deafferentation is insufficient to inducesprouting of A-fiber central terminals in the rat dor-sal horn. Journal of Comparative Neurology 393, 135–144.
MCCARTHY, P. W. & LAWSON, S. N. (1990) Cell typeand conduction velocity of rat primary afferent neuronswith calcitonin gene-related peptide-like immunoreac-tivity. Neuroscience 34, 623–632.
NAGY, J. L. & HUNT, S. P. (1983) The termination of pri-mary afferents within the rat dorsal horn: Evidence forrearrangement following capsaicin treatment. Journal ofComparative Neurology 218, 145–158.
NAIM, M., SPIKE, R. C., WATT, C., SHEHAB, S. A. S. &ANDREW J. T. (1997) Cells in laminae III and IV of therat spinal cord that possess the neurokinin-1 receptor andhave dorsally directed dendrites receive a major synap-tic input from tachykinin-containing primary afferents.Journal of Neuroscience 17, 5536–5548.
NASU, F. (1999) Analysis of calcitonin gene-related peptide(CGRP)-containing nerve fibers in the rat spinal cord us-ing light and electron microscopy. Journal of Electron Mi-croscopy 48, 267–275.
OLAVE, M. J., PURI, N., KERR, R. & MAXWELL, D.J. (2002) Myelinated and unmyelinated primary affer-ent axons form contacts with cholinergic interneurons inthe spinal dorsal horn. Experimental Brain Research 145,448–456.
PUTTFARCKEN, P. S., MANELLI, A. M., ARNERIC, S. P.& DONELLEY-ROBERTS, D. L. (1997) Evidence fornicotinic receptors potentially modulating nociceptivetransmission at the level of the primary sensory neu-rons: Studies with F11 cells. Journal of Neurochemistry 69,930–938.
RIBEIRO-DA-SILVA, A. (1995) Ultrastructural features ofthe colocalization of calcitonin gene related peptide withsubstance P or somatostatin in the dorsal horn of theSpinal Cord. Canadian Journal of Physiology and Pharma-cology 73, 940–944.
RIBEIRO-DA-SILVA, A. & COIMBRA, A. (1984) Capsaicincauses selective damage of type I synaptic glomeruli inrat substansia gelatinosa. Brain Research 290, 380–383.
RIBEIRO-DA-SILVA, A. & CUELLO, A. C. (1990) Cholineacetyltransferase profiles are presynaptic to primary sen-sory fibers in the rat superficial dorsal horn. Journal ofComparative Neurology 295, 370–384.
ROBERTS, R. G. D, STEVENSON, J. E., WESTERMAN, R.A. & PENNEFATHER, J. (1995) Nicotinic acetylcholinereceptors on capsaicin-sensitive nerves. NeuroReport 6,1578–1582.
SCHAFER, M. K.-H., EIDEN, L. E. & WEIHE, E. (1998)Cholinergic neurons and terminal fields revealed byimmunohistochemistry for the vesicular acetylcholinetransporter. I. Central nervous system. Neuroscience 84,331–359.
SCHICHO, R., SKOFITSCH, G. & DONNERER, J. (1999)Regenerative effect of human recombinant NGF oncapsaicin-lesioned sensory neurons in the adult rat. BrainResearch 815, 60–69.
SILVERMAN, J. D. & KRUGER, L. (1990) Selective neu-ronal glycoconjugate expression in sensory and auto-nomic ganglia: Relation of lectin reactivity to peptide andenzyme markers. Journal of Neurocytology 19, 789–801.
TAKEDA, D., NAKATSUK, T., PAPKE, R. & GU, J. G.(2003) Modulation of inhibitory synaptic activity by anon-α4β2, non-α7 subtype of nicotinic receptors in thesubstantia gelatinosa of adult rat spinal cord. Pain 101,13–23.
TORSNEY, C., MEREDITH-MIDDLETON, J. &FITZGERALD, M. (2000) Neonatal capsaicin treat-ment prevents the normal postnatal withdrawal of Afibers from lamina II without affecting Fos responses toinnocuous peripheral stimulation. Developmental BrainResearch 121, 55–65.
WANG, F., GERZANICH, V., WELLS, G. B., ANAND,R., PENG, X., KEYSER, K. & LINDSTROM J. (1996)Assembly of human neuronal nicotinic receptor α5 sub-units with α3, β2, and β4 subunits. Journal of BiologicalChemistry 271, 17656–17665.
WANG, N., ORR-URTREGER, A., CHAPMAN, J.,RABINOWITZ, R., NACHMAN, R. & KORCZYN,A. D. (2002) Autonomic function in mice lacking α5 neu-ronal nicotinic acetylcholine receptor subunit. Journal ofPhysiology 542, 347–354.
WILLIAMS, J. T. & ZIEGLGANSBERGER, W. (1982) Theacute effects of capsaicin on rat primary afferents andspinal neurons. Brain Research 253, 125–131.
WILLIS, W. D. & COGGESHALL, R. E. (1991) Sensorymechanisms of the spinal cord, 2nd edition. New York:Plenum.
Copyright © 2022 FDOKUMEN




















