
Tonic and phasic descending dopaminergic controls of nociceptive transmission in the medullary...
11
Tonic and phasic descending dopaminergic controls of nociceptive transmission in the medullary dorsal horn Olivier Lapirot a,b , Céline Melin a,b,c , Alice Modolo a,b , Charline Nicolas a,b,d , Yassine Messaoudi a,b , Lénaic Monconduit a,b , Alain Artola a,b , Philippe Luccarini a,b,d , Radhouane Dallel a,b,c,⇑ a Clermont Université, Université d’Auvergne, Neurobiologie de la douleur trigéminale, BP 10448, F-63000 Clermont-Ferrand, France b Inserm, U929, F-63000 Clermont-Ferrand, France c CHU Clermont-Ferrand, Service d’Odontologie, F-63003 Clermont-Ferrand, France d IUT Génie Biologique, 63172 Aubière, France Sponsorships or competing interests that may be relevant to content are disclosed at the end of this article. article info Article history: Received 9 November 2010 Received in revised form 23 March 2011 Accepted 24 March 2011 Available online xxxx Keywords: D2-like receptor Diffuse noxious inhibiting controls Medullary dorsal horn Trigeminal Windup abstract The transfer of nociceptive information at the level of dorsal horn is subject to extensive processing by both local segmental and supraspinal mechanisms, including descending dopaminergic controls, origi- nating from the hypothalamic A11 nucleus. The inhibitory role of dopamine on evoked pain via activation of D2-like receptors at the level of the dorsal horn is well established. Here, by use of behavioral, electro- physiological, and anatomical techniques, we examined within the trigeminal sensory complex, first, whether descending dopaminergic controls also modulate pain behavior after an inflammatory insult, and second, under which physiological conditions these descending dopaminergic controls are actually recruited. We show that D2 receptors are mostly located within superficial medullary dorsal horn where trigeminal nociceptive fibers abut. Activating these D2-like receptors inhibits, whereas blocking them enhances, both formalin- and capsaicin-evoked pain behavior and C-fiber-evoked action potential firing of trigeminal wide dynamic range (WDR) neurons. Moreover, windup and diffuse noxious inhibitory con- trols (DNIC), 2 dynamic properties of C-fiber-evoked firing of WDR neurons, are inhibited by activating and blocking, respectively, these D2-like receptors. Altogether, our results are consistent with a tonic inhibition of the trigeminal nociceptive input by descending dopaminergic controls via activation of D2-like receptors at the level of superficial medullary dorsal horn. Such dopamine-dependent tonic inhi- bition of nociceptive information can be dynamically modulated by pain. This suggests that dysregulation of descending dopaminergic controls should translate in patients into diffuse, cephalic, and extracephalic pain symptoms—spontaneous pain, decreased pain thresholds, deficient DNIC, or some combination of these. Ó 2011 International Association for the Study of Pain. Published by Elsevier B.V. All rights reserved. 1. Introduction The sensory inflow generated by nociceptors activates neurons in the dorsal horn (DH), which project to the cortex via a relay in the thalamus, generating pain. The transfer of nociceptive informa- tion at the level of the DH is subject to extensive processing by both local segmental and supraspinal mechanisms [29]. In this re- gard, a network of descending pathways projecting from cerebral structures to DH act to modulate the passage of nociceptive mes- sages to the brain. Although descending noradrenergic and seroto- nergic controls have been the object of intense investigation, the actions of descending dopaminergic controls on spinal and medul- lary DHs (SDH and MDH, respectively) are less well defined (for re- view, see [43]). Nevertheless, these descending dopaminergic controls of sensory afferents might be important in maintaining normal nociceptive thresholds because abnormalities in dopami- nergic neurotransmission have been suggested to contribute to painful clinical conditions including Parkinson disease, chronic ce- phalic pain, fibromyalgia, and restless legs syndrome (for review, see [44,50,65]). Dopamine acts through 5 distinct G protein-coupled receptors (D1–D5) belonging to 2 families, D1-like (D1 and D5) and D2-like (D2, D3, and D4) receptors, which positively and negatively, respectively, regulate adenylyl-cyclase. Though all dopamine receptors are present in the spinal cord [3,8,69,70], the D2 receptor is the most prominent dopamine receptor subtype in both the SDH and MDH (eg, [3,9,70]; but see [38]), and mediates the antinocicep- tive action of dopamine [22,66]. 0304-3959/$36.00 Ó 2011 International Association for the Study of Pain. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.pain.2011.03.030 ⇑ Corresponding author. Address: Inserm U929, Neurobiologie de la douleur trigéminale, Faculté de Chirurgie Dentaire, 11 boulevard Charles de Gaulle, 63000 Clermont-Ferrand, France. Tel.: +33 4 73 17 73 13; fax: +33 4 73 17 73 06. E-mail address: [email protected] (R. Dallel). PAIN Ò xxx (2011) xxx–xxx www.elsevier.com/locate/pain Please cite this article in press as: Lapirot O et al. Tonic and phasic descending dopaminergic controls of nociceptive transmission in the medullary dorsal horn. PAIN Ò (2011), doi:10.1016/j.pain.2011.03.030
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Tonic and phasic descending dopaminergic controls of nociceptive transmission in the medullary...

PAIN�
xxx (2011) xxx–xxx
w w w . e l s e v i e r . c o m / l o c a t e / p a i n
Tonic and phasic descending dopaminergic controls of nociceptive transmissionin the medullary dorsal horn
Olivier Lapirot a,b, Céline Melin a,b,c, Alice Modolo a,b, Charline Nicolas a,b,d, Yassine Messaoudi a,b,Lénaic Monconduit a,b, Alain Artola a,b, Philippe Luccarini a,b,d, Radhouane Dallel a,b,c,⇑a Clermont Université, Université d’Auvergne, Neurobiologie de la douleur trigéminale, BP 10448, F-63000 Clermont-Ferrand, Franceb Inserm, U929, F-63000 Clermont-Ferrand, Francec CHU Clermont-Ferrand, Service d’Odontologie, F-63003 Clermont-Ferrand, Franced IUT Génie Biologique, 63172 Aubière, France
Sponsorships or competing interests that may be relevant to content are disclosed at the end of this article.
a r t i c l e i n f o a b s t r a c t
Article history:Received 9 November 2010Received in revised form 23 March 2011Accepted 24 March 2011Available online xxxx
Keywords:D2-like receptorDiffuse noxious inhibiting controlsMedullary dorsal hornTrigeminalWindup
0304-3959/$36.00 � 2011 International Associationdoi:10.1016/j.pain.2011.03.030
⇑ Corresponding author. Address: Inserm U929, Ntrigéminale, Faculté de Chirurgie Dentaire, 11 boulevClermont-Ferrand, France. Tel.: +33 4 73 17 73 13; fa
E-mail address: [email protected] (
Please cite this article in press as: Lapirot O et ahorn. PAIN
�(2011), doi:10.1016/j.pain.2011.03.
The transfer of nociceptive information at the level of dorsal horn is subject to extensive processing byboth local segmental and supraspinal mechanisms, including descending dopaminergic controls, origi-nating from the hypothalamic A11 nucleus. The inhibitory role of dopamine on evoked pain via activationof D2-like receptors at the level of the dorsal horn is well established. Here, by use of behavioral, electro-physiological, and anatomical techniques, we examined within the trigeminal sensory complex, first,whether descending dopaminergic controls also modulate pain behavior after an inflammatory insult,and second, under which physiological conditions these descending dopaminergic controls are actuallyrecruited. We show that D2 receptors are mostly located within superficial medullary dorsal horn wheretrigeminal nociceptive fibers abut. Activating these D2-like receptors inhibits, whereas blocking themenhances, both formalin- and capsaicin-evoked pain behavior and C-fiber-evoked action potential firingof trigeminal wide dynamic range (WDR) neurons. Moreover, windup and diffuse noxious inhibitory con-trols (DNIC), 2 dynamic properties of C-fiber-evoked firing of WDR neurons, are inhibited by activatingand blocking, respectively, these D2-like receptors. Altogether, our results are consistent with a tonicinhibition of the trigeminal nociceptive input by descending dopaminergic controls via activation ofD2-like receptors at the level of superficial medullary dorsal horn. Such dopamine-dependent tonic inhi-bition of nociceptive information can be dynamically modulated by pain. This suggests that dysregulationof descending dopaminergic controls should translate in patients into diffuse, cephalic, and extracephalicpain symptoms—spontaneous pain, decreased pain thresholds, deficient DNIC, or some combination ofthese.
� 2011 International Association for the Study of Pain. Published by Elsevier B.V. All rights reserved.
1. Introduction actions of descending dopaminergic controls on spinal and medul-
The sensory inflow generated by nociceptors activates neuronsin the dorsal horn (DH), which project to the cortex via a relay inthe thalamus, generating pain. The transfer of nociceptive informa-tion at the level of the DH is subject to extensive processing byboth local segmental and supraspinal mechanisms [29]. In this re-gard, a network of descending pathways projecting from cerebralstructures to DH act to modulate the passage of nociceptive mes-sages to the brain. Although descending noradrenergic and seroto-nergic controls have been the object of intense investigation, the
for the Study of Pain. Published by
eurobiologie de la douleurard Charles de Gaulle, 63000x: +33 4 73 17 73 06.R. Dallel).
l. Tonic and phasic descending030
lary DHs (SDH and MDH, respectively) are less well defined (for re-view, see [43]). Nevertheless, these descending dopaminergiccontrols of sensory afferents might be important in maintainingnormal nociceptive thresholds because abnormalities in dopami-nergic neurotransmission have been suggested to contribute topainful clinical conditions including Parkinson disease, chronic ce-phalic pain, fibromyalgia, and restless legs syndrome (for review,see [44,50,65]).
Dopamine acts through 5 distinct G protein-coupled receptors(D1–D5) belonging to 2 families, D1-like (D1 and D5) and D2-like(D2, D3, and D4) receptors, which positively and negatively,respectively, regulate adenylyl-cyclase. Though all dopaminereceptors are present in the spinal cord [3,8,69,70], the D2 receptoris the most prominent dopamine receptor subtype in both the SDHand MDH (eg, [3,9,70]; but see [38]), and mediates the antinocicep-tive action of dopamine [22,66].
Elsevier B.V. All rights reserved.
dopaminergic controls of nociceptive transmission in the medullary dorsal

2 O. Lapirot et al. / PAIN�
xxx (2011) xxx–xxx
Local administration of either dopamine or D2-like receptoragonists reduces heat sensitivity [2,24,35,39] and increasesmechanical nociceptive thresholds [1,58]. Similarly, locally applieddopamine reduces the responses of DH neurons to noxious stimulisuch as cutaneous mechanical stimulation [22,59]) and electricalstimulation of either the sciatic nerve [66] or the middle meningealartery [3]. Descending dopamine projections into the mammalianspinal cord arise from A11 cell groups in the dorsoposterior hypo-thalamus [31,56]. Accordingly, electrical activation of this regionhas been shown to inhibit responses to noxious stimuli (cutaneouspinch, dural stimulation; [8,22]) and spontaneous excitatory post-synaptic currents (EPSCs; [59]) in DH neurons.
Though the role of DH dopamine on evoked pain is well estab-lished, to our knowledge, that of pain behavior after an inflamma-tory insult has not yet been evaluated. By means of behavioral,electrophysiological, and anatomical techniques, we have ad-dressed 2 questions. First, do descending dopaminergic controlsmodulate formalin- and capsaicin-evoked pain behavior? And sec-ond, under what physiological conditions are descending dopami-nergic controls recruited? These questions were addressed in thetrigeminal system.
2. Methods
Adult male Sprague Dawley rats (200–300 g; Charles River Lab-oratories, L’Arbresle, France) were housed in a temperature-con-trolled room (22 �C; lights on at 07:00–21:00 h) with food andwater ad libitum. All procedures followed the guidelines underthe International Association for the Study of Pain and the Euro-pean Community Council (86/609/EEC) and were approved bythe local animal care and use committee (UFR d’Odontologie, Uni-versité d’Auvergne-Clermont 1). All efforts were made to minimizeanimal suffering and to reduce the number of animals used.
2.1. Behavioral analysis
2.1.1. Facial capsaicin and formalin testsRats received either formalin (50 lL, 1.5%) or capsaicin (25 lL,
0.1%) into the right vibrissa pad as previously described [48,52].After injection, animals were placed back within the test box fora 45-min observation period. The recording time was divided into15 3-min blocks. For each block, a nociceptive score was deter-mined by measuring the time (in seconds) animals spent rubbingthe injected area (vibrissa pad) with the forepaw and not the hindpaw. Behavior was analyzed by an investigator who was blinded toanimal treatment.
For intracisternal (i.c.) injection, animals were briefly (<3 min)anaesthetized with halothane (2%) and a 25-gauge needle attachedto a 10-lL Hamilton syringe inserted through the skin into cisternamagna. Five microliters of either the D2-like receptor agonist quin-pirole (1.2, 4, 12, and 20 nmol) or the D2-like receptor antagonistsulpiride (30 nmol) or vehicle solution were applied. The antago-nistic action of sulpiride on quinpirole-induced effects was as-sessed by intraperitoneal administration of sulpiride (50 mg/kg)90 min before the i.c. quinpirole (12 nmol) was administered.
2.1.2. Locomotor function, coordination, and sedation of ratsRats were tested 20 and 40 min after i.c. quinpirole (20 nmol) or
vehicle with the accelerating rotarod (LE 8500; Ugo Basile), in whichrats are required to walk against the motion of a rotating drum, withthe speed increasing from 4 to 40 rpm over 5 min (see [40]). Thetime taken to fall off the rotarod is recorded as the latency (inseconds). Twenty-four hours before drug testing, rats were trainedto stay on the accelerating rotarod for 60 s. The next day, rotarodlatencies were measured after i.c. injection of quinpirole.
Please cite this article in press as: Lapirot O et al. Tonic and phasic descendinghorn. PAIN
�(2011), doi:10.1016/j.pain.2011.03.030
2.2. Electrophysiological recordings
2.2.1. Animal preparationAs previously described [15], animals were anesthetized with
2% halothane in a NO2/O2 mixture (2:1). After intramuscular injec-tion of 100 lg atropine sulfate, the trachea was cannulated and thecarotid artery and external jugular vein catheterized. Animals werethen paralyzed by an intravenous perfusion of vecuronium bro-mide (2.4 mg/h) and artificially ventilated with a volume-con-trolled pump (54–55 strokes/min). Levels of halothane, O2, N2O,and end-tidal CO2 (3.5–4.5%) were measured by an anesthetic gasanalyser (Artema MM 200, Sundbyberg, Sweden) during the entireexperimental period. These parameters were digitally displayedand under the control of alarms. The arterial catheter was attachedto a calibrated pressure transducer (UFI, Morro Bay, CA, USA) con-nected to an amplifier (Stoelting, Wood Dale, IL, USA) for continu-ous monitoring of the mean arterial blood pressure (90–110 mmHg). The analog output from the blood pressure amplifier was con-nected to a computer data sampling system (Cambridge Electron-ics Design 1401 computer interface; Cambridge, UK). Heart ratewas continuously monitored and cutaneous vascularization peri-odically checked by observing the color of the paw extremitiesand the rapidity by which they regained normal color after pres-sure application. Colorectal temperature was kept constant at38 ± 0.5 �C by means of a feedback-controlled heating blanket. Sur-gical level of anesthesia was verified by a stable mean arterialblood pressure and a constant heart rate during noxiousstimulation.
The animals were placed in a stereotaxic frame with the headfixed in a ventroflexed position (incisor bar dropped 5 mm underthe standard position) by means of an adapted metallic bar. Forspinal trigeminal subnucleus oralis (Sp5O) recordings, a craniot-omy was performed on the right side at the level of the occipitop-arietalis suture and the dura mater removed. After surgery, thelevel of halothane was reduced to 0.6–0.7% and maintained at thislevel during the recording period.
2.2.2. MicroinjectionsDrugs were delivered in the MDH by 3-barrel glass micropi-
pettes fixed on the micromanipulator and connected to Hamiltonsyringes (1 ll) by means of polyethylene tubing [15]. Micropi-pettes and tubing were filled with either quinpirole (10 nmol in0.25 lL) or sulpiride (5 nmol in 0.25 lL) or saline pontamine skyblue (for location of the injection site) solutions. Injections of drugswere performed with a manual injector over a period of 2 min andmonitored by observing the movement of an air bubble in the tub-ing. The slow rate of injection was chosen to minimize tissuedamage.
The micropipette was positioned stereotaxically within theMDH 1 h before the injection. The micropipette tip was first low-ered 1 mm below the pial surface. The medulla was then coveredby placing 2% Ringer-agar gel. The coordinates used for microinjec-tion sites were 5.3 mm posterior to interaural plane (approxi-mately obex) and 1.0–2.8 mm lateral to midline [46].
2.2.3. Unitary extracellular recordingsWe used glass micropipettes (8–10 MX) filled with pontamine
sky blue solution to explore the Sp5O (AP interaural �1.1 to�2.6 mm, length 2.4–3.0 mm). Single unit activities were amplifiedand displayed on oscilloscopes and led into a window discrimina-tor connected to a CED 1401plus interface and a computer (Spike2.05 software) to allow sampling and analysis of the spontaneousand evoked neuronal activities. Wide dynamic range (WDR) neu-rons were recognized on the basis of their responses to mechanicaland percutaneous electrical stimulations of their receptive field[14]. Specifically, neurons that responded in a graded manner with
dopaminergic controls of nociceptive transmission in the medullary dorsal

O. Lapirot et al. / PAIN�
xxx (2011) xxx–xxx 3
increasing firing rates to the stimulus range from non-noxious tonoxious intensities were classified as WDR cells. Once a neuronhad been identified, its receptive field was mapped. Electricalsquare-wave stimuli (2-ms duration) were applied through a pairof stainless steel needle electrodes subcutaneously placed intothe center of the previously delineated receptive field and thresh-olds for eliciting A- and C-fiber-evoked responses determined. Inpoststimulus time histograms (PSTHs), C- and A-fiber-evoked re-sponses were distinguished according to their latencies: all spikesbetween 30 and 300 ms after stimulus were considered as C fiberevoked [14,33].
Recording and microinjection sites were visualized by injectionof pontamine sky blue solution at the end of the experiment.
2.2.4. C-fiber-evoked responses to peripheral electrical stimuliSequences of 30 electrical shocks were applied repeatedly
(0.66 Hz) to the excitatory receptive field at 3 times the thresholdfor C-fiber activation. Sequences were repeated at 5-min intervals.When 2 successive control sequences had been recorded with avariation of less than 10% in C-fiber-evoked responses, either quin-pirole (10 nmol in 0.25 lL), sulpiride (5 nmol in 0.25 lL) or quinpi-role with sulpiride were microinjected into the MDH. Such avolume is consistent with an easy diffusion of the drugs withinthe MDH. Indeed, autoradiographic data [6,17] have demonstratedthat 0.2 lL of morphine can diffuse in a 1.5-mm diameter sphere ofbrain tissue. Responses were recorded up to 95 min after drugapplication. Responses—either windup slope or number ofspikes—were normalized to the averaged 2 responses recorded be-fore applying the drug (baseline). Only one cell was tested in eachanimal, and only cells showing no change in spike amplitude orwaveform during the complete experimental procedure wereconsidered.
During repetitive stimulation, C-fiber-evoked responses of WDRneurons progressively increased, reached a maximum and thenplateaued. Fits of the windup plots were obtained by a least-squares fitting algorithm. The dynamic properties of the successiveresponses were best fitted by a second-order polynomial regres-sion (R2 = 0.99), indicating that in all cases there was a strong cor-relation between the amplitude of the response and the number ofrepetitive stimuli [12]. Because the initial slope of the polynomialfit curves (coefficient of the first degree of x) corresponded to theability of the neuron to amplify its response, it was used to quan-tify windup.
2.2.5. Diffuse noxious inhibitory controls (DNIC)As previously described [36], sequences of 105 electrical stimuli
(0.66 Hz) were applied to the receptive field at 3 times the thresh-old for C fibers at 15-min intervals. A 15-min interval was selectedbecause it is the shortest one to allow full recovery to baseline re-sponse between sequences. DNIC were triggered by alternativelyimmersing the right or left hind paw—only one hind paw was im-mersed during each sequence, the right one at 5, 35, 65, and 95 minand the left one at 20, 50, and 80 min after microinjection—into a48 �C water bath between the 36th and 60th stimuli (duration:37.5 s).
In each sequences, the PSTHs built from the first to 30th re-sponses were used for studying the unconditioned response, thosebuilt from the responses to 36th to 105th stimuli for studyingDNIC. To assess the inhibitory effect of noxious heat stimulation,only the 46th to 60th responses were considered, responses to ear-lier 36th to 45th stimuli during immersion being discarded owingto the latency (5–10 s) of the maximal inhibitory effects. Postef-fects were measured on the 61th to 75th and 76th to 90th re-sponses, 0–22 s and 22–44 s after the end of noxious heatstimulation, respectively. In each sequence, PSTHs, built from46th to 60th (effect), 61st to 75th (first posteffect) and 76th to
Please cite this article in press as: Lapirot O et al. Tonic and phasic descendinghorn. PAIN
�(2011), doi:10.1016/j.pain.2011.03.030
90th (second posteffect) responses, were normalized to that builtfrom the 21st to 35th responses (before noxious heat stimulation;baseline). The inhibitory effect of DNIC was computed as the per-cent decrease in the number of C-fiber-evoked action potentials.Sulpiride was microinjected within the MDH after recording 2 sta-ble (variation <10%) unconditioned C-fiber responses. There wasonly one sulpiride injection in each rat.
2.3. Immunohistochemistry
Rats were deeply anesthetized with chloral hydrate (400 mg/kg,i.p.) and perfused transcardially with 200 mL of warm (37 �C) hep-arinized, phosphate-buffered saline (PBS) followed by 500 mL of 4%paraformaldehyde, 0.03% picric acid in PBS (0.1 M, pH 7.6) for15 min. Brains were removed and cryoprotected overnight in thesame paraformaldehyde–picric acid solution containing 30% su-crose at 4 �C. Coronal frozen sections (40 lm thick) were cut andcollected in 0.05 M Tris-buffered saline (TBS). All immunolabelswere diluted in 0.25% bovine serum albumin, 0.3% Triton X-100in TBS.
2.3.1. D2. receptor immunohistochemistryAfter brain stem sections had free-floated in 1% normal horse
serum for 1 h, immunohistochemistry reaction was carried outwith a rabbit antibody against D2 receptor (1:9000, Calbiochem;overnight at room temperature) using biotinylated horse anti-rab-bit followed by avidin–biotin–peroxidase complex (Immpress,Abcys, Vector; 30 min at room temperature). D2 receptor immuno-reactivity was visualized in sections using 3,30-diaminobenzidenetetrahydrochloride (DAB) and ammonium nickel sulphate (kit Vec-tor Peroxidase substrate DAB). All sections were rinsed in TBS andtransferred to gelatinized slides before being coverslipped withDPX.
2.3.2. D2. receptor and gamma-subtype protein kinase C (PKCc)immunofluorescence
After brain stem sections had free-floated in 1% normal goat ser-um for 1 h, immunofluorescent staining reaction was carried outwith a rabbit antibody against D2 receptor (1:9000, Calbiochem;overnight at room temperature) and a mouse antibody againstPKCc (1:4000, Sigma; overnight at room temperature). Immunore-activity was revealed using Cy3 conjugated goat anti-rabbit(1:200) and Cy2 conjugated goat anti-mouse (1:200) secondaryantibodies respectively (2 h at room temperature).
Specificity controls consisted of the omission of the primaryantibodies and incubation of sections in inappropriate secondaryantibodies.
2.3.3. Data analysisComputer-assisted bright-field images of injection sites and
representative labeling were obtained with a CCD color video cam-era (Sony DXC-950P) connected to a Nikon Optiphot-2 microscope.Images were exported to Adobe PhotoShop (v 5.5) to adjust bright-ness and contrast before adjusting the image scale. PKCc and D2receptor immunofluorescence was analyzed with a motorizedZeiss Axioplan 2 Imaging microscope coupled with a HamamatsuC4742-95 digital camera, by switching between fluorescein andTexas Red filter sets. For illustrations, numerical images of immu-nofluorescence were deconvoluted by Metamorph 5.4 software.The delineation of the MDH was based on the atlas of Paxinosand Watson [46] and our own myeloarchitectural atlas.
2.4. Drugs
The following drugs were used: Quinpirole (Sigma Aldrich,France) dissolved in 0.9% saline and sulpiride (Sigma Aldrich,
dopaminergic controls of nociceptive transmission in the medullary dorsal

4 O. Lapirot et al. / PAIN�
xxx (2011) xxx–xxx
France) in 2.5% HCl, 7.5% DMSO, and 90% saline (0.9%). Fresh solu-tions were prepared every testing day. The doses for the dopami-nergic compounds were selected according to the dosage used inprevious experiments: i.c. quinpirole (1.2–20 nmol; comparedwith 30–300 nmol; [24,58]); intracerebral quinpirole (10 nmolcompared with 1–300 nmol; [1,41,53,60]); i.c. sulpiride (30 nmol;compared with 3–300 nmol; [11,26,55]); intracerebral sulpiride(5 nmol compared with 0.5–15 nmol; [45,64,68]); and systemic(i.v.) sulpiride (50 mg/kg; compared with 20–60 mg/kg; [47]). Wealso examined the effect of i.c. quinpirole on behavior of the rats.We thus noticed that large doses of i.c. quinpirole (P120 nmol,ie, much larger than the ones we used here) produced psychomo-tor impairments (as assessed with the accelerating rotarod) andyawns.
2.5. Statistical analysis
Results are presented as mean ± SEM. Data were analyzed byeither the Student t test or 1-way analysis of variance (ANOVA), or1-way repeated measures ANOVA followed by a post hoc Student-Newman–Keuls test. The level of significance was set at P < .05.
3. Results
3.1. D2-like receptor agonist/antagonist modulate formalin- andcapsaicin-evoked trigeminal pain behavior
To examine whether dopamine can modulate formalin- andcapsaicin-evoked trigeminal pain via D2-like receptors at the levelof the MDH, we tested the effect of i.c. injections of the D2-likereceptor agonist, quinpirole, and antagonist, sulpiride, on facial for-malin- and capsaicin- evoked nociceptive behaviors.
Subcutaneous injection of formalin into the vibrissa pad causesa 2-phase nociceptive rubbing response: an early, short-lastingphase 1 and a late long-lasting phase 2 (as previously described[48,52]). Control rats exhibited intense facial rubbing behavior(n = 8). Durations of phase 1 (0–3 min) and phase 2 (15–39 min)rubbing were 9 ± 5 and 225 ± 31 s, respectively (Fig. 1a and b). Pre-treatment with i.c. quinpirole dose-dependently reduced phase 2(ANOVA followed by Student-Newman–Keuls test, F6,53 = 20.148,P < .001). Thus, phase 2 was reduced by 3 ± 9 (n = 8), �38 ± 10(n = 11; P < .05), �72 ± 5 (n = 10; P < .001), and �70 ± 5% (n = 10;P < .001), after i.c. administration of 1.2, 4.0, 12.0, and 20.0 nmolof quinpirole, respectively. The ED50 value for phase 2 suppressionwas 7.1 nmol (r = 0.76, 95% confidence interval, 9.7–5.2 nmol). Weconfirmed that quinpirole acts through D2-like receptors by show-ing that systemic administration of sulpiride (50 mg/kg, i.v., 90 minbefore formalin; n = 6) significantly reduced the antinociceptive ef-fect of quinpirole (12 nmol). Indeed, in these conditions, phase 2was only decreased by �36 ± 6% (compared with �72 ± 5%;P < .001; Fig. 1a and b); that is, quinpirole-induced antinociceptionwas reduced by �51 ± 11%. Conversely, pretreatment with i.c. sul-piride (30 nmol) significantly facilitated phase 2 (64 ± 19%, n = 7;P < .002; Fig. 1a and b). Neither i.c. quinpirole nor i.c. sulpiride af-fected phase 1.
Subcutaneous injection of capsaicin into the vibrissa pad in-duced a nociceptive rubbing behavior that lasted 89 ± 14 s in i.c.vehicle pretreated rats (n = 8). Pretreatment with i.c. quinpirole(4 nmol) significantly reduced capsaicin-induced rubbing behaviorby �72 ± 7% (n = 8; P < .05) whereas i.c. sulpiride (30 nmol) signif-icantly facilitated it by 131 ± 44% (n = 6; P < .001) (Fig. 1c).
We controlled that the highest dose of i.c. quinpirole (20 nmol)had no effect on motor performance by showing that the rotarodperformance of rats was not affected (155 ± 13 and 183 ± 31 s com-pared with 161 ± 11 and 191 ± 17 s with vehicle, at 20 and 40 min
Please cite this article in press as: Lapirot O et al. Tonic and phasic descendinghorn. PAIN
�(2011), doi:10.1016/j.pain.2011.03.030
after i.c. administration, respectively). In addition, there was nei-ther macroscopic change in the aspect of the cerebrospinalfluid—being translucent, with no blood—nor brain stem lesionsafter postmortem controls.
3.2. D2 receptors are located within superficial MDH
We used an immunohistochemical approach to investigate thelocation of D2-like receptors within the trigeminal sensory com-plex (Fig. 2a and b). Whereas there was no detectable D2 recep-tor immunoreactive neuron or fiber in the Sp5O (Fig. 2a), wefound a strong D2 receptor immunostaining in the MDH(Fig. 2b). Within MDH, D2 receptor immunostaining was mostprominent in superficial laminae. In order to get more insightinto the laminar distribution of D2 receptor immunoreactivity,we performed double-staining immunohistochemistry for D2receptor and the PKCc proteins (Fig. 2b, inset). PKCc is concen-trated in interneurons localized in the inner part of lamina II(for review, see [61]). Fig. 2b (inset) shows that D2 receptorsand PKCc did not colocalize. Therefore, these results indicate thatD2 receptor immunoreactivity is mostly restricted to lamina Iand the outer part of lamina II (IIo).
We then compared the laminar distribution of D2 receptorimmunoreactivity in the MDH with that in the spinal cord(Fig. 2d). Interestingly, both areas exhibited the very same patternof D2 receptor immunoreactivity with a strong D2 receptor stain-ing in superficial (lamina I/IIo) DH (see Fig. 2d, inset).
Finally, we assessed D2 receptor immunoreactivity within thetrigeminal ganglion. D2 receptor protein was present in neuronalcell bodies distributed within the ganglion (Fig. 2c).
3.3. D2-like receptor agonist/antagonist into the MDH modulate theC-fiber-evoked response of trigeminal WDR neurons
Thus, D2-like receptors in the trigeminal sensory complex seemto be mostly located within superficial MDH. Can activation of D2-like receptors at the level of superficial MDH account for the antin-ociceptive action of quinpirole? To address this question, we devel-oped a combined microinjection/single cell recording approach inwhich microinjection and recording sites were well apart fromeach other: the D2-like receptor agonist/antagonist was microin-jected into superficial MDH while we recorded from WDR neuronsin the Sp5O. Sp5O WDR neurons are located rostrally, approxi-mately 3 mm apart from C-fiber afferent terminals that abut cau-dally in superficial MDH [13–15]. Interneurons relay thenociceptive information from superficial MDH to downstreamSp5O WDR neurons [13,63].
A total of 28 WDR neurons were recorded within the Sp5O.None of them exhibited spontaneous activity. All had an ipsilateralreceptive field that included the intraoral or perioral region. Theywere sensitive to both innocuous and noxious mechanical stimuliand responded by increasing their firing rate as stimulus intensityincreased into the noxious range. When 2-ms-long percutaneouselectrical stimuli were applied to the center of the receptive fieldof the neurons, responses attributable to peripheral activation ofA and C fibers could be observed (Fig. 3a and b and 4a). The longestlatency responses (94 ± 2 ms) were C fiber evoked. Indeed, thecomputed conduction velocity (�0.6 m s�1) was in the range ofthose previously reported for C fibers [12–14,42]. In addition, longlatency responses could only be evoked by high intensity stimula-tions (mean threshold: 8.8 ± 1.6 mA) and they exhibited windup[12–14,42]. Fig. 4 shows an example of the progressive increaseof C-fiber-evoked responses to repetitive (0.66 Hz) constant percu-taneous suprathreshold electrical stimuli applied to the receptivefield. Finally, it has already been shown that these long latency re-sponses in Sp5O neuron are selectively depressed by intravenous
dopaminergic controls of nociceptive transmission in the medullary dorsal
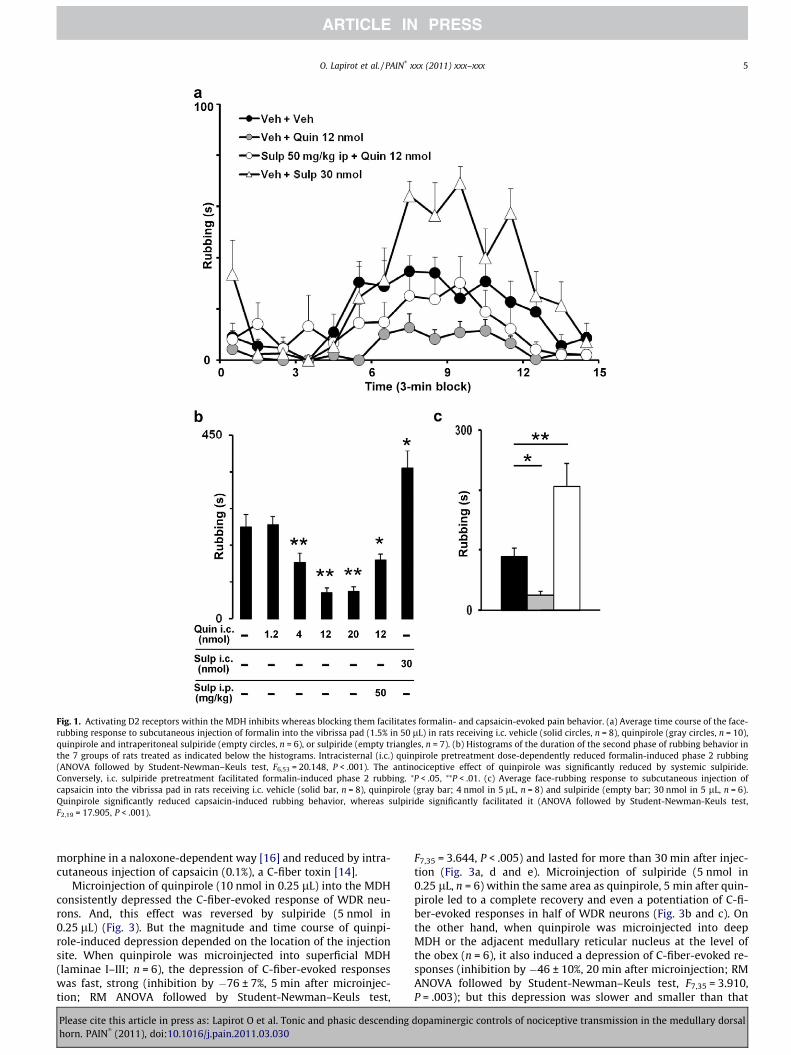
Fig. 1. Activating D2 receptors within the MDH inhibits whereas blocking them facilitates formalin- and capsaicin-evoked pain behavior. (a) Average time course of the face-rubbing response to subcutaneous injection of formalin into the vibrissa pad (1.5% in 50 lL) in rats receiving i.c. vehicle (solid circles, n = 8), quinpirole (gray circles, n = 10),quinpirole and intraperitoneal sulpiride (empty circles, n = 6), or sulpiride (empty triangles, n = 7). (b) Histograms of the duration of the second phase of rubbing behavior inthe 7 groups of rats treated as indicated below the histograms. Intracisternal (i.c.) quinpirole pretreatment dose-dependently reduced formalin-induced phase 2 rubbing(ANOVA followed by Student-Newman–Keuls test, F6,53 = 20.148, P < .001). The antinociceptive effect of quinpirole was significantly reduced by systemic sulpiride.Conversely, i.c. sulpiride pretreatment facilitated formalin-induced phase 2 rubbing. ⁄P < .05, ⁄⁄P < .01. (c) Average face-rubbing response to subcutaneous injection ofcapsaicin into the vibrissa pad in rats receiving i.c. vehicle (solid bar, n = 8), quinpirole (gray bar; 4 nmol in 5 lL, n = 8) and sulpiride (empty bar; 30 nmol in 5 lL, n = 6).Quinpirole significantly reduced capsaicin-induced rubbing behavior, whereas sulpiride significantly facilitated it (ANOVA followed by Student-Newman-Keuls test,F2,19 = 17.905, P < .001).
O. Lapirot et al. / PAIN�
xxx (2011) xxx–xxx 5
morphine in a naloxone-dependent way [16] and reduced by intra-cutaneous injection of capsaicin (0.1%), a C-fiber toxin [14].
Microinjection of quinpirole (10 nmol in 0.25 lL) into the MDHconsistently depressed the C-fiber-evoked response of WDR neu-rons. And, this effect was reversed by sulpiride (5 nmol in0.25 lL) (Fig. 3). But the magnitude and time course of quinpi-role-induced depression depended on the location of the injectionsite. When quinpirole was microinjected into superficial MDH(laminae I–III; n = 6), the depression of C-fiber-evoked responseswas fast, strong (inhibition by �76 ± 7%, 5 min after microinjec-tion; RM ANOVA followed by Student-Newman–Keuls test,
Please cite this article in press as: Lapirot O et al. Tonic and phasic descendinghorn. PAIN
�(2011), doi:10.1016/j.pain.2011.03.030
F7,35 = 3.644, P < .005) and lasted for more than 30 min after injec-tion (Fig. 3a, d and e). Microinjection of sulpiride (5 nmol in0.25 lL, n = 6) within the same area as quinpirole, 5 min after quin-pirole led to a complete recovery and even a potentiation of C-fi-ber-evoked responses in half of WDR neurons (Fig. 3b and c). Onthe other hand, when quinpirole was microinjected into deepMDH or the adjacent medullary reticular nucleus at the level ofthe obex (n = 6), it also induced a depression of C-fiber-evoked re-sponses (inhibition by �46 ± 10%, 20 min after microinjection; RMANOVA followed by Student-Newman–Keuls test, F7,35 = 3.910,P = .003); but this depression was slower and smaller than that
dopaminergic controls of nociceptive transmission in the medullary dorsal

Fig. 2. Immunohistochemical assessment of the D2 receptor distribution in the Sp5O (a), MDH (b), adult rat trigeminal ganglion (c) and lumbar spinal cord (d). (a) Lightmicrograph of D2 receptor protein immunoreactivity in the Sp5O. No D2 receptor immunoreactivity is detected. (b) Light micrograph of D2 receptor proteinimmunoreactivity in the MDH. It shows a strong D2 receptor immunostaining in superficial MDH. Inset: Double-staining immunochemistry for D2 receptor (red) and PKCc(green) proteins showing that there is no overlap between D2 receptor and PKCc immunostainings. Therefore, within MDH, D2 receptor immunoreactivity is mainlydistributed in lamina I and IIo. (c) Light micrograph of D2 receptor protein immunoreactivity in the trigeminal ganglion. Neurons in the rat trigeminal ganglion do express D2receptors. (d) Light micrograph of D2 receptor protein immunoreactivity in the lumbar spinal cord. It shows a strong D2 receptor immunostaining in superficial DH. Inset:Double staining immunochemistry for D2 receptor (red) and PKCc (green) proteins showing that there is no overlap between D2 receptor and PKCc immunostainings.Therefore, in the spinal DH, too, D2 receptor protein immunoreactivity is mainly distributed in lamina I and IIo. Scale bars: 1 mm in a, b, and d; 50 lm in c.
6 O. Lapirot et al. / PAIN�
xxx (2011) xxx–xxx
in the previous group (Fig. 3d and e). This is consistent with theinterpretation that quinpirole had first to diffuse into superficialMDH before activating D2-like receptors. Conversely, microinjec-tion of sulpiride alone (5 nmol in 0.25 lL) into superficial MDHconsistently facilitated C-fiber-evoked responses of WDR neurons(potentiation by 50 ± 12%, 20 min after microinjection, n = 8;P = .015; not illustrated).
3.4. D2-like receptor activation attenuates windup
Windup is a progressive, frequency-dependent facilitation ofthe responses of WDR neurons to C-fiber activation observed onthe application of repetitive stimuli of constant intensity (for re-view, see [12,30]; Fig. 4a). Windup was markedly attenuated(�67 ± 8%, n = 10; P = .016, paired t test) at 5 min after quinpirolemicroinjection (10 nmol in 0.25 lL) into superficial MDH (Fig. 4b).
3.5. D2-like receptor blockade inhibits DNIC
Descending inhibition of pain is involved in a naturally acti-vated control system, namely DNIC, whereby the response ofWDR neurons in the DH (for review, see [37]) and trigeminal nuclei[14] to C-fiber activation is inhibited by the application of noxiousstimuli to remote body areas. Is this modulation of C-fiber-evokedresponse also dependent on the activation of D2-like receptors?
To address this question, we investigated the effect of blockingD2-like receptors in superficial MDH (Fig. 5a) on DNIC of WDR neu-rons. During the control period, application of noxious heat (48 �C)to the right or left hind paw strongly depressed all tested units(inhibition of C-fiber-evoked responses by �88 ± 1, �52 ± 6, and�32 ± 9% during heat application and the 2 consecutive 22-s post-
Please cite this article in press as: Lapirot O et al. Tonic and phasic descendinghorn. PAIN
�(2011), doi:10.1016/j.pain.2011.03.030
conditioning periods, respectively; n = 8; Fig. 5b). After sulpiridemicroinjection into the MDH (5 nmol in 0.25 lL), DNIC triggeredby noxious heat stimulation of the right or left hind paw werestrongly reduced in all recorded neurons (n = 8; Fig. 5b). Sulpir-ide-induced reduction of DNIC was fast (already significant at5 min and maximum at 35 min after sulpiride microinjection)and lasted for at least 95 min after microinjection (Fig. 5c).Thirty-five minutes after sulpiride microinjection, heat-inducedinhibition of C-fiber-evoked responses was only �45 ± 10%,�28 ± 12%, and �22 ± 12% during heat application and the 2 post-conditioning periods, respectively; that is, DNIC was strongly re-duced by �49 ± 10% (RM ANOVA followed by Student-Newman-Keuls test, F7,42 = 7.702, P < .001), �55 ± 21% and �52 ± 26%,respectively. It is interesting to note that sulpiride also markedlyenhanced baseline C-fiber-evoked responses (Fig. 5c). But potenti-ation of baseline C-fiber-evoked responses and reduction in DNIChad different time courses. And changes in baseline C-fiber-evokedresponses and DNIC were not correlated. Finally, microinjectedvehicle into the MDH had no effect (n = 4).
4. Discussion
We show that, within the trigeminal sensory complex, D2-likereceptors are mostly located in superficial MDH where nociceptivefibers abut. Activating these D2-like receptors inhibits, whereasblocking them enhances, both formalin- and capsaicin-evoked painbehavior and C-fiber-evoked action potential firing of trigeminalWDR neurons. Windup and DNIC, 2 dynamic properties of C-fi-ber-evoked firing of WDR neurons, are inhibited by activatingand blocking, respectively, these D2-like receptors. Our resultsare consistent with a tonic inhibition of trigeminal nociceptive
dopaminergic controls of nociceptive transmission in the medullary dorsal
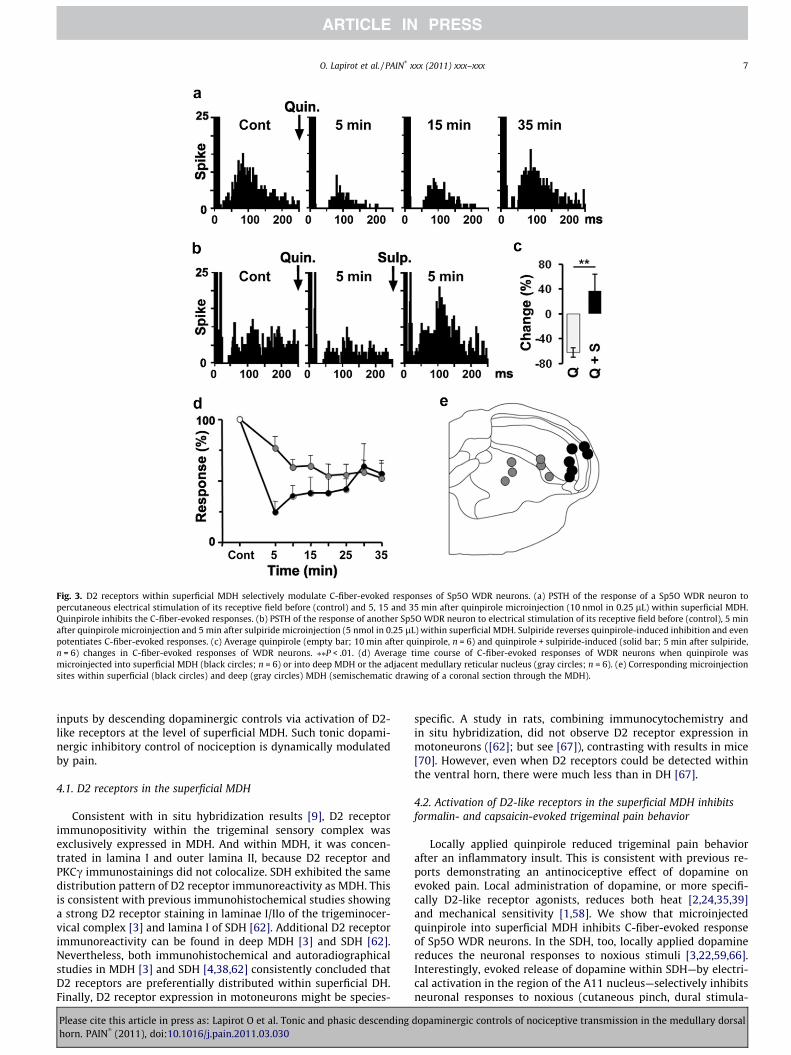
Fig. 3. D2 receptors within superficial MDH selectively modulate C-fiber-evoked responses of Sp5O WDR neurons. (a) PSTH of the response of a Sp5O WDR neuron topercutaneous electrical stimulation of its receptive field before (control) and 5, 15 and 35 min after quinpirole microinjection (10 nmol in 0.25 lL) within superficial MDH.Quinpirole inhibits the C-fiber-evoked responses. (b) PSTH of the response of another Sp5O WDR neuron to electrical stimulation of its receptive field before (control), 5 minafter quinpirole microinjection and 5 min after sulpiride microinjection (5 nmol in 0.25 lL) within superficial MDH. Sulpiride reverses quinpirole-induced inhibition and evenpotentiates C-fiber-evoked responses. (c) Average quinpirole (empty bar; 10 min after quinpirole, n = 6) and quinpirole + sulpiride-induced (solid bar; 5 min after sulpiride,n = 6) changes in C-fiber-evoked responses of WDR neurons. ⁄⁄P < .01. (d) Average time course of C-fiber-evoked responses of WDR neurons when quinpirole wasmicroinjected into superficial MDH (black circles; n = 6) or into deep MDH or the adjacent medullary reticular nucleus (gray circles; n = 6). (e) Corresponding microinjectionsites within superficial (black circles) and deep (gray circles) MDH (semischematic drawing of a coronal section through the MDH).
O. Lapirot et al. / PAIN�
xxx (2011) xxx–xxx 7
inputs by descending dopaminergic controls via activation of D2-like receptors at the level of superficial MDH. Such tonic dopami-nergic inhibitory control of nociception is dynamically modulatedby pain.
4.1. D2 receptors in the superficial MDH
Consistent with in situ hybridization results [9], D2 receptorimmunopositivity within the trigeminal sensory complex wasexclusively expressed in MDH. And within MDH, it was concen-trated in lamina I and outer lamina II, because D2 receptor andPKCc immunostainings did not colocalize. SDH exhibited the samedistribution pattern of D2 receptor immunoreactivity as MDH. Thisis consistent with previous immunohistochemical studies showinga strong D2 receptor staining in laminae I/IIo of the trigeminocer-vical complex [3] and lamina I of SDH [62]. Additional D2 receptorimmunoreactivity can be found in deep MDH [3] and SDH [62].Nevertheless, both immunohistochemical and autoradiographicalstudies in MDH [3] and SDH [4,38,62] consistently concluded thatD2 receptors are preferentially distributed within superficial DH.Finally, D2 receptor expression in motoneurons might be species-
Please cite this article in press as: Lapirot O et al. Tonic and phasic descendinghorn. PAIN
�(2011), doi:10.1016/j.pain.2011.03.030
specific. A study in rats, combining immunocytochemistry andin situ hybridization, did not observe D2 receptor expression inmotoneurons ([62]; but see [67]), contrasting with results in mice[70]. However, even when D2 receptors could be detected withinthe ventral horn, there were much less than in DH [67].
4.2. Activation of D2-like receptors in the superficial MDH inhibitsformalin- and capsaicin-evoked trigeminal pain behavior
Locally applied quinpirole reduced trigeminal pain behaviorafter an inflammatory insult. This is consistent with previous re-ports demonstrating an antinociceptive effect of dopamine onevoked pain. Local administration of dopamine, or more specifi-cally D2-like receptor agonists, reduces both heat [2,24,35,39]and mechanical sensitivity [1,58]. We show that microinjectedquinpirole into superficial MDH inhibits C-fiber-evoked responseof Sp5O WDR neurons. In the SDH, too, locally applied dopaminereduces the neuronal responses to noxious stimuli [3,22,59,66].Interestingly, evoked release of dopamine within SDH—by electri-cal activation in the region of the A11 nucleus—selectively inhibitsneuronal responses to noxious (cutaneous pinch, dural stimula-
dopaminergic controls of nociceptive transmission in the medullary dorsal

Fig. 4. Windup of Sp5O WDR neurons is D2-like receptor-dependent. (a) Typicalresponses of a Sp5O WDR neuron to the first, third, fifth, and eighth (from top tobottom) supramaximal (3� C fiber threshold intensity, 0.66 Hz) percutaneouselectrical stimulation of its receptive field. Note the progressive increase in theresponse component corresponding to C-fiber latency. (b) Total number of actionpotentials in the C-fiber-evoked component of the response to 15 successivestimulations (3� C-fiber threshold intensity, 0.66 Hz) before (control; solidsymbols) and after microinjection of quinpirole (10 nmol in 0.25 lL; emptysymbols) into the MDH. Superimposed on the plots are the polynomial fits of thedata (n = 10; before, y = �0.192x2 + 5.117x � 1.401; after, y = �0.013x2 +1.043x � 0.360).
Fig. 5. D2-like receptors within superficial MDH modulate DNIC. (a) Example of amicroinjection site within superficial MDH. Scale bar: 1 mm. (b) Peristimulushistograms showing the C-fiber-evoked responses of a Sp5O WDR neuron to 105successive percutaneous electrical stimulations (3� C-fiber threshold intensity,0.66 Hz) recorded before (left) and 35 min (right) after sulpiride microinjection(5 nmol in 0.25 lL) within superficial MDH. Between the 36th and 60th stimulation,the hind paw was immersed into 48 �C water bath. DNIC triggered by heat noxiousstimulation of the hind paw is reduced after sulpiride microinjection. (c) Averagetime course of the effect of sulpiride microinjection within superficial MDH (n = 8)on C-fiber-evoked responses of WDR neurons (empty columns) and on DNIC (solidcolumns) triggered by stimulation of the right (5, 35, 66, and 95 min) or left hindpaw (20, 50, and 80 min). Sulpiride microinjection within superficial MDHfacilitates C-fiber-evoked baseline responses and reduces DNIC. Whereas C-fiber-evoked baseline responses are back to control levels 35 min after sulpiride injection,DNIC is still reduced 95 min after it. ⁄P < .05, ⁄⁄⁄P < .001.
8 O. Lapirot et al. / PAIN�
xxx (2011) xxx–xxx
tion), but not innocuous, stimulation [8,22]. Altogether, theseresults suggest that, at the level of DH, dopamine selectively sup-presses passage of nociceptive—but not nonnociceptive—informa-tion to the brain, via D2-like receptors.
4.3. Dopamine tonically inhibits the transmission of nociceptiveinformation in the MDH
That nociceptive responses—behavioral as well as electrophysi-ological responses—are enhanced after locally applied sulpiridesuggests that there is an ongoing (tonic) stimulation of D2-likereceptors within the superficial MDH inhibiting nociceptive inputs.
Please cite this article in press as: Lapirot O et al. Tonic and phasic descendinghorn. PAIN
�(2011), doi:10.1016/j.pain.2011.03.030
In SDH, however, sulpiride fails to affect any monitored—noxiousas well as innocuous—response [22]. But, in this latter study, sul-piride was iontophoresed close to the recorded WDR neurons, thatis, in deep—and not superficial—SDH. Nevertheless, our conclusionthat there is a tonic dopaminergic inhibitory control of nociceptionis consistent with the effect of electrically lesioning the A11
dopaminergic controls of nociceptive transmission in the medullary dorsal

O. Lapirot et al. / PAIN�
xxx (2011) xxx–xxx 9
nucleus, that is facilitation of neuronal responses in the trigemi-nocervical complex to dural and noxious cutaneous stimulation[8].
4.4. Descending dopamine controls are involved in DNIC
Application of a D2-like receptor antagonist in superficial MDHstrongly attenuates ‘‘pain-inhibition of pain,’’ or DNIC. This sug-gests that D2-like receptor-mediated tonic inhibitory control ofnociception is dynamically modulated by, at least, heterotopicpain. Mechanisms underlying DNIC involve a supraspinal loop withits ascending and descending limbs traveling through the ventro-lateral and the dorsolateral funiculi, respectively (for review, see[37]). However, the supraspinal structures of DNIC are still poorlyknown. Neurokinin 1 receptor-expressing projection neurons inDH and the parabrachial area seem to be critical relays in ascend-ing pathways [36,57]. And the subnucleus reticularis dorsalis thatprojects through the dorsolateral funiculus to all rostrocaudallevels of the SDH is likely involved in descending pathways (for re-view, see [37]). Our results suggest that descending dopaminergicinputs, that also travel through the dorsolateral funiculus [32] anddistribute to all rostrocaudal levels of the SDH [31,56], also contrib-ute to DNIC.
That, in DNIC experiments, sulpiride-induced potentiation ofbaseline C-fiber-evoked responses and reduction in DNIC have dif-ferent time courses is also consistent with 2 inhibitory effects bydescending dopaminergic innervation: a tonic one, which main-tains normal nociceptive thresholds, and a phasic one, triggeredhere by heterotopic pain. Indeed, the latter, producing a strongeranalgesia than the former, likely also requires a larger ratio ofD2-like receptor occupancy. As sulpiride is progressively elimi-nated from brain tissue, D2-like receptor blockade decreases andtonic inhibition, requiring a lower ratio of D2-like receptor occu-pancy than the phasic one, recovers first.
4.5. Pre- and postsynaptic localization of D2-like receptors in DH
That activation of D2-like receptors selectively inhibits nocicep-tive transmission whilst innocuous transmission (but see [59]) isunaffected implies that these receptors are located on nociceptivefiber primary terminals and/or on neurons that are selectively acti-vated by nociceptive fibers. This raises the question as to whetherD2-like receptor-mediated inhibition of nociceptive transmissionis mediated by pre- and/or postsynaptic mechanisms. Our immu-nohistochemical study demonstrates the presence of D2 receptorswithin the trigeminal ganglion, consistent with in situ hybridiza-tion results [49]. Consistent with a presynaptic mechanism are alsoelectrophysiological results showing that dopamine produces anidentical relative depression of early, AMPA/kainate, and late,NMDA receptor-mediated, components of the responses of DHneurons to electrical stimulation of primary afferents [25] and re-duces the frequency of spontaneous and mini EPSCs in substantiagelatinosa neurons [59]. However, dopamine or D2-like receptoragonists have also been shown to have direct postsynaptic actionson substantia gelatinosa [58,59] and deep DH neurons [25] and toinhibit the responses of trigeminal neurons to microiontophoreti-cally applied glutamate [3]. Activation of both pre- and postsynap-tic D2-like receptors can account for the reduction of the windupslope after local application of a D2-like receptor agonist. Windupslope is correlated with stimulus intensity, ie, the number of re-cruited C-fiber afferents [12], and reducing the excitability ofspinal cord neurons, for instance by opening potassium channels,decreases or even abolishes windup (for review, see [30]).
That quinpirole and sulpiride are D2–D3 receptor ligands raisesthe question as to whether their actions are mediated by D2 and/orD3 receptors. Quantitative autoradiography [38] and in situ
Please cite this article in press as: Lapirot O et al. Tonic and phasic descendinghorn. PAIN
�(2011), doi:10.1016/j.pain.2011.03.030
hybridization [70] show that, in the spinal cord, D3 receptors, asD2 receptors, are predominantly distributed within superficialSDH [38] but significantly less than them. That heat thresholdsare lowered and formalin-evoked pain behavior enhanced in D3receptor knock-out mice ([20]; but see [71]) suggests that D3receptors also mediate a tonic control on nociception. However,D3 receptors might be involved in more generally limiting spinalcord excitability, impaired D3 receptor-mediated inhibition ofspinal sensory and reflex circuits likely underlying the restless legssyndrome [10].
4.6. Conclusion and clinical implications
Our results suggest that an ongoing (tonic) stimulation of D2-like receptors within DH is necessary to maintain normal nocicep-tive responses. Thus, impaired descending dopaminergic controlsshould translate into diffuse, cephalic and extracephalic, painsymptoms, ie, spontaneous pain and/or decreased heat andmechanical pain thresholds and/or deficient DNIC. Can such painsymptoms be identified in patients with impaired dopamineneurotransmission?
Patients with Parkinson disease exhibit sensory disorders, com-bining diffuse spontaneous pain (often burning; for review, see[23]) with decreased specifically heat pain thresholds [19]. Thesethresholds return to normal ranges after L-dopa administration[5]. Similarly, the burning mouth syndrome associates burningsensations in the mouth with diffuse reduced heat pain thresholds[27]. Cutaneous allodynia is also often associated with migraine,either within the referred pain area on the ipsilateral head, or with-in and outside the ipsilateral head [7]. Interestingly, cephalic cuta-neous allodynia is mostly mechanical, while the diffuseextracephalic one is mostly thermal [28]. Finally, fibromyalgia, achronic widespread pain syndrome comorbid with migraine [34],exhibits both decreased heat pain thresholds and deficient DNIC[51]. Interestingly, in most of these clinical conditions, in whichabnormalities in dopamine neurotransmission are suspected (forreview, see [50,65]), diffuse spontaneous pain is associated withrather specifically decreased heat thresholds, raising the possibilitythat dopaminergic control of nociception is modality specific. Andadministration of D2-like receptor agonists might be considered inpatients associating diffuse spontaneous pain, reduced heat painthresholds, and deficient DNIC. It is important to be aware thatdouble-blind, placebo-controlled studies on neuroleptics and pain,though very few, suggest that such D2 receptor antagonists ratherproduce analgesia at therapeutic doses [18,21,54]. Certainly, the ef-fect of neuroleptics on pain depend on many factors, including: (1)doses (low doses likely block presynaptic D2 autoreceptors, en-hance dopamine release, and are antinociceptive, while higherdoses also block postsynaptic D2 receptors and are pronocicep-tive); (2) the primary action of neuroleptics, which is usually notlimited to D2 receptors but also involves serotonin, norepineph-rine, and histamine receptors; and (3) the site of action withinthe brain.
Conflict of interest statement
None of the authors has any conflict of interest to declare.
Acknowledgments
We thank A.M. Gaydier for secretarial assistance and J.L. Molatfor technical help during electrophysiological experiments. Thiswork was supported by funding from Institut National de la Santéet de la Recherche Médicale (Inserm), Université Clermont1(France), and Région Auvergne (France).
dopaminergic controls of nociceptive transmission in the medullary dorsal

10 O. Lapirot et al. / PAIN�
xxx (2011) xxx–xxx
References
[1] Ansah OB, Leite-Almeida H, Wei H, Pertovaara A. Striatal dopamine D2receptors attenuate neuropathic hypersensitivity in the rat. Exp Neurol2007;205:536–46.
[2] Barasi S, Duggal KN. The effect of local and systemic application ofdopaminergic agents on tail flick latency in the rat. Eur J Pharmacol1985;117:87–94.
[3] Bergerot A, Storer RJ, Goadsby PJ. Dopamine inhibits trigeminovasculartransmission in the rat. Ann Neurol 2007;61:251–62.
[4] Bouthenet ML, Martres MP, Sales N, Schwartz JC. A detailed mapping ofdopamine D-2 receptors in rat central nervous system by autoradiographywith [120I]iodosulpiride. Neuroscience 1987;20:117–55.
[5] Brefel-Courbon C, Payoux P, Thalamas C, Ory F, Quelven I, Chollet F, MontastrucJL, Rascol O. Effect of levodopa on pain threshold in Parkinson’s disease: aclinical and positron emission tomography study. Mov Disord2005;20:1557–63.
[6] Burkey AR, Carstens E, Wenniger JJ, Tang J, Jasmin L. An opioidergic corticalantinociception triggering site in the agranular cortex of the rat thatcontributes to morphine antinociception. J Neurosci 1996;16:6612–23.
[7] Burstein R, Yarnitsky D, Goor-Aryeh I, Ransil BJ, Bajwa ZH. An associationbetween migraine and cutaneous allodynia. Ann Neurol 2000;47:614–24.
[8] Charbit AR, Akerman S, Holland PR, Goadsby PJ. Neurons of the dopaminergic/calcitonin gene-related peptide A11 cell group modulate neuronal firing in thetrigeminocervical complex: an electrophysiological and immunohistochemicalstudy. J Neurosci 2009;29:12532–41.
[9] Chen JF, Qin ZH, Szele F, Bai G, Weiss B. Neuronal localization and modulationof the D2 dopamine receptor mRNA in brain of normal mice and mice lesionedwith 6-hydroxydopamine. Neuropharmacology 1991;30:927–41.
[10] Clemens S, Hochman S. Conversion of the modulatory actions of dopamine onspinal reflexes from depression to facilitation in D3 receptor knock-out mice. JNeurosci 2004;24:11337–45.
[11] Cobacho N, De la Calle JL, González-Escalada JR, Paíno CL. Levodopa analgesiain experimental neuropathic pain. Brain Res Bull 2010;83:304–9.
[12] Coste J, Voisin DL, Luccarini P, Dallel R. A role for wind-up in trigeminalsensory processing: intensity coding of nociceptive stimuli in the rat.Cephalalgia 2008;28:631–9.
[13] Coste J, Voisin DL, Miraucourt LS, Dallel R, Luccarini P. Dorsal horn NK1-expressing neurons control windup of downstream trigeminal nociceptiveneurons. Pain 2008;137:340–51.
[14] Dallel R, Duale C, Luccarini P, Molat JL. Stimulus-function, wind-up andmodulation by diffuse noxious inhibitory controls of responses of convergentneurons of the spinal trigeminal nucleus oralis. Eur J Neurosci 1999;11:31–40.
[15] Dallel R, Duale C, Molat JL. Morphine administered in the substantia gelatinosaof the spinal trigeminal nucleus caudalis inhibits nociceptive activities in thespinal trigeminal nucleus oralis. J Neurosci 1998;18:3529–36.
[16] Dallel R, Luccarini P, Molat JL, Woda A. Effects of systemic morphine on theactivity of convergent neurons of spinal trigeminal nucleus oralis in the rat.Eur J Pharmacol 1996;314:19–25.
[17] Dickenson AH, Le Bars D. Supraspinal morphine and descending inhibitionsacting on the dorsal horn of the rat. J Physiol (Lond) 1987;384:81–107.
[18] Dipirro JM, Thompson AC, Suarez M, Leo RJ. Low doses of risperidone andmorphine interact to produce more analgesia and greater extrapyramidaleffects in rats. Neurosci Lett 2011;490:21–6.
[19] Djaldetti R, Shifrin A, Rogowski Z, Sprecher E, Melamed E, Yarnitsky D.Quantitative measurement of pain sensation in patients with Parkinsondisease. Neurology 2004;62:2171–5.
[20] Dowling P, Klinker F, Stadelmann C, Hasan K, Paulus W, Liebetanz D. DopamineD3 receptor specifically modulates motor and sensory symptoms in iron-deficient mice. J Neurosci 2011;31:70–7.
[21] Fishbain DA, Cutler RB, Lewis J, Cole B, Rosomoff RS, Rosomoff HL. Do thesecond-generation ‘‘atypical neuroleptics’’ have analgesic properties? Astructured evidence-based review. Pain Med 2004;5:359–65.
[22] Fleetwood-Walker SM, Hope PJ, Mitchell R. Antinociceptive actions ofdescending dopaminergic tracts on cat and rat dorsal horn somatosensoryneurons. J Physiol 1988;399:335–48.
[23] Ford B. Pain in Parkinson’s disease. Mov Disord 2010;25:S98–S103.[24] Gao X, Zhang Y, Wu G. Effects of dopaminergic agents on carrageenan
hyperalgesia after intrathecal administration to rats. Eur J Pharmacol2001;418:73–7.
[25] Garraway SM, Hochman S. Modulatory actions of serotonin, norepinephrine,dopamine, and acetylcholine in spinal cord deep dorsal horn neurons. JNeurophysiol 2001;86:2183–94.
[26] Golbidi S, Moriuchi H, Irie T, Ghafghazi T, Hajhashemi V. Involvement ofcalmodulin inhibition in analgesia induced with low doses of intrathecaltrifluoperazine. Jpn J Pharmacol 2002;88:151–7.
[27] Grémeau-Richard C, Dubray C, Aublet-Cuvelier B, Ughetto S, Woda A. Effect oflingual nerve block on burning mouth syndrome (stomatodynia): arandomized crossover trial. Pain 2010;149:27–32.
[28] Guy N, Marques AR, Orliaguet T, Lanteri-Minet M, Dallel R, Clavelou P. Arethere differences between cephalic and extracephalic cutaneous allodynia inmigraine patients? Cephalalgia 2010;30:881–6.
[29] Heinricher MM, Tavares I, Leith JL, Lumb BM. Descending control ofnociception: specificity, recruitment and plasticity. Brain Res Rev2009;60:214–25.
Please cite this article in press as: Lapirot O et al. Tonic and phasic descendinghorn. PAIN
�(2011), doi:10.1016/j.pain.2011.03.030
[30] Herrero JF, Laird JM, Lopez-Garcia JA. Windup of spinal cord neurons and painsensation: much ado about something? Prog Neurobiol 2000;61:169–203.
[31] Hökfelt T, Phillipson O, Goldstein M. Evidence for a dopaminergic pathway inthe rat descending from the A11 cell group to the spinal cord. Acta PhysiolScand 1979;107:393–5.
[32] Holstege JC, Van Dijken H, Buijs RM, Goedknegt H, Gosens T, Bongers CM.Distribution of dopamine immunoreactivity in the rat, cat and monkey spinalcord. J Comp Neurol 1996;376:631–52.
[33] Hu JW. Response properties of nociceptive and non-nociceptive neurons in therat’s trigeminal subnucleus caudalis (medullary dorsal horn) related tocutaneous and deep craniofacial afferent stimulation and modulation bydiffuse noxious inhibitory controls. Pain 1990;41:331–45.
[34] Ifergane G, Buskila D, Simiseshvely N, Zeev K, Cohen H. Prevalence offibromyalgia syndrome in migraine patients. Cephalalgia 2006;26:451–6.
[35] Jensen TS, Yaksh TL. Effects of an intrathecal dopamine agonist, apomorphine,on thermal and chemical evoked noxious responses in rats. Brain Res1984;296:285–93.
[36] Lapirot O, Chebbi R, Monconduit L, Artola A, Dallel R, Luccarini P. NK1receptor-expressing spinoparabrachial neurons trigger diffuse noxiousinhibitory controls through lateral parabrachial activation in the male rat.Pain 2009;142:245–54.
[37] Le Bars D. The whole body receptive field of dorsal horn multireceptiveneurones. Brain Res Brain Res Rev 2002;40:29–44.
[38] Levant B, McCarson KE. D3 dopamine receptors in rat spinal cord: implicationsfor sensory and motor function. Neurosci Lett 2001;303:9–12.
[39] Liu QS, Qiao JT, Dafny N. D2 dopamine receptor involvement in spinaldopamine-produced antinociception. Life Sci 1992;251:1485–92.
[40] Luccarini P, Perrier L, Dégoulange C, Gaydier AM, Dallel R. Synergisticantinociceptive effect of amitriptyline and morphine in the rat orofacialformalin test. Anesthesiology 2004;100:690–6.
[41] Magnusson JE, Fisher K. The involvement of dopamine in nociception: the roleof D1 and D2 receptors in the dorsolateral striatum. Brain Res2000;855:260–6.
[42] Mendell LM. Physiological properties of unmyelinated fiber projection to thespinal cord. Exp Neurol 1966;16:316–32.
[43] Millan MJ. Descending control of pain. Prog Neurobiol 2002;66:355–474.[44] Mascia A, Afra J, Schoenen J. Dopamine and migraine: a review of
pharmacological, biochemical, neurophysiological, and therapeutic data.Cephalalgia 1998;18:174–82.
[45] Mochizuki T, Kim J, Sasaki K. Microinjection of neuropeptide S into the ratventral tegmental area induces hyperactivity and increases extracellular levelsof dopamine metabolites in the nucleus accumbens shell. Peptides2010;31:926–31.
[46] Paxinos G, Watson C. The rat brain in stereotaxic coordinates. 3rd ed. NewYork: Academic Press; 1997.
[47] Pelissier T, Laurido C, Hernandez A, Constandil L, Eschalier A. Biphasic effect ofapomorphine on rat nociception and effect of dopamine D2 receptorantagonists. Eur J Pharmacol 2006;546:40–7.
[48] Pelissier T, Pajot J, Dallel R. The orofacial capsaicin test in rats: effects ofdifferent capsaicin concentrations and morphine. Pain 2002;96:81–7.
[49] Peterfreund RA, Kosofsky BE, Fink JS. Cellular localization of dopamine D2receptor messenger RNA in the rat trigeminal ganglion. Anesth Analg1995;81:1181–5.
[50] Potvin S, Grignon S, Marchand S. Human evidence of a supra-spinalmodulating role of dopamine on pain perception. Synapse 2009;63:390–402.
[51] Potvin S, Larouche A, Normand E, de Souza JB, Gaumond I, Grignon S,Marchand S. DRD3 Ser9Gly polymorphism is related to thermal painperception and modulation in chronic widespread pain patients and healthycontrols. J Pain 2009;10:969–75.
[52] Raboisson P, Dallel R. The orofacial formalin test. Neurosci Biobehav Rev2004;28:219–26.
[53] Saunier-Rébori BT, Pazo JH. Inhibition of jaw opening reflex and single neuronsin the trigeminal subnucleus caudalis by activation of striatal D2 dopaminereceptors. Neuropharmacology 2006;51:263–71.
[54] Seidel S, Aigner M, Ossege M, Pernicka E, Wildner B, Sycha T. Antipsychotics foracute and chronic pain in adults. J Pain Symptom Manage 2010;39:768–78.
[55] Shimizu T, Iwata S, Morioka H, Masuyama T, Fukuda T, Nomoto M.Antinociceptive mechanism of L-DOPA. Pain 2004;110:246–9.
[56] Skagerberg G, Lindvall O. Organization of diencephalic dopamine neuronesprojecting to the spinal cord in the rat. Brain Res 1985;342:340–51.
[57] Suzuki R, Morcuende S, Webber M, Hunt SP, Dickenson AH. Superficial NK1-expressing neurons control spinal excitability through activation ofdescending pathways. Nat Neurosci 2002;5:1319–26.
[58] Tamae A, Nakatsuka T, Koga K, Kato G, Furue H, Katafuchi T, Yoshimura M.Direct inhibition of substantia gelatinosa neurones in the rat spinal cord byactivation of dopamine D2-like receptors. J Physiol 2005;568:243–53.
[59] Taniguchi W, Nakatsuka T, Miyazaki N, Yamada H, Takeda D, Fujita T,Kumamoto E, Yoshida M. In vivo patch-clamp analysis of dopaminergicantinociceptive actions on substantia gelatinosa neurons in the spinal cord.Pain 2010;152:95–105.
[60] Taylor BK, Joshi C, Uppal H. Stimulation of dopamine D2 receptors in thenucleus accumbens inhibits inflammatory pain. Brain Res2003;987:135–43.
[61] Todd AJ. Anatomy of primary afferents and projection neurones in the ratspinal dorsal horn with particular emphasis on substance P and the neurokinin1 receptor. Exp Physiol 2002;87:245–9.
dopaminergic controls of nociceptive transmission in the medullary dorsal

O. Lapirot et al. / PAIN�
xxx (2011) xxx–xxx 11
[62] van Dijken H, Dijk J, Voom P, Holstege JC. Localization of dopamine D2 receptorin rat spinal cord identified with immunocytochemistry and in situhybridization. Eur J Neurosci 1996;8:621–8.
[63] Voisin DL, Doméjean-Orliaguet S, Chalus M, Dallel R, Woda A. Ascendingconnections from the caudal part to the oral part of the spinal trigeminalnucleus in the rat. Neuroscience 2002;109:183–93.
[64] Winter S, Dieckmann M, Schwabe K. Dopamine in the prefrontal cortexregulates rats behavioral flexibility to changing reward value. Behav Brain Res2009;198:206–13.
[65] Wood PB. Role of central dopamine in pain and analgesia. Expert RevNeurother 2008;8:781–97.
[66] Yang HW, Zhou LJ, Hu NW, Xin WJ, Liu XG. Activation of spinal D1/D5receptors induces late-phase LTP of C-fiber-evoked field potentials in rat spinaldorsal horn. J Neurophysiol 2005;94:961–7.
[67] Yokoyama C, Okamura H, Nakajima T, Taguchi JI, Ibata Y. Autoradiographicdistribution of [0H]YM-O915 1–2, a high-affinity and selective antagonist
Please cite this article in press as: Lapirot O et al. Tonic and phasic descendinghorn. PAIN
�(2011), doi:10.1016/j.pain.2011.03.030
ligand for the dopamine D2 receptor group, in the rat brain and spinal cord. JComp Neurol 1994;344:121–36.
[68] Zarrindast MR, Naghdi-Sedeh N, Nasehi M, Sahraei H, Bahrami F, Asadi F. Theeffects of dopaminergic drugs in the ventral hippocampus of rats in thenicotine-induced anxiogenic-like response. Neurosci Lett 2010;475:156–60.
[69] Zhao H, Zhu W, Pan T, Xie W, Zhang A, Ondo WG, Le W. Spinal cord dopaminereceptor expression and function in mice with 6-OHDA lesion of the A11nucleus and dietary iron deprivation. J Neurosci Res 2007;85:1065–76.
[70] Zhu H, Clemens S, Sawchuk M, Hochman S. Expression and distribution of alldopamine receptor subtypes (D1–D5) in the mouse lumbar spinal cord: a real-time polymerase chain reaction and non-autoradiographic in situhybridization study. Neuroscience 2007;149:885–97.
[71] Zhu J, Chen Y, Lai J, Danga Y, Yan C, Xu M, Chen T. Dopamine D3 receptorregulates basal but not amphetamine-induced changes in pain sensitivity inmice. Neurosci Lett 2010;477:134–7.
dopaminergic controls of nociceptive transmission in the medullary dorsal




















