
Medullary structure of the Umbilicariaceae
11
Lichenologist 27(3): 189-199 (1995) MEDULLARY STRUCTURE OF THE UMBILICARIACEAE Fernando VALLADARES* and Leopold0 G. SANCHO# Abstract: The medullary structure of 24 Umbilicariaceae species was studied by light microscopy. The objective was to investigate the variability of the medulla of these lichens and to assess the possible implications for thallus ecophysiology and the taxonomic relevance of the anatomy of this layer. Several methods of embed- ding, sectioning and staining together with the comparison of radial with transverse sections showed important differences in the degree of cohesion of hyphae and their spatial orientation within the medulla. Other important characteristics were the existence of more than one structurally homogeneous layer within the medulla, the frequency of hyphal branching, the relative thickness of the cell wall and lumen and the abundance of gelatinous matter in the intercellular spaces. The medullae studied were grouped into seven main types. The constancy of the types of medulla for certain groups of species makes it a useful secondary character to be considered in the taxonomy of the Umbilicariaceae. The important role of the structure of the medulla in the water relations of the Umbilicariaceae is discussed and a general relationship between medulla type and hydration strategy is suggested. Introduction In the family Umbilicariaceae, as in most foliose lichens, the medulla is the thickest layer of the thallus. The two aspects of major interest concerning the structural features of the medulla of these lichens are their possible implica- tions for thallus ecophysiology and their taxonomic relevance. The crucial influence of the medulla in thallus water relations was mentioned in early studies of the Umbilicariaceae (Backmann 1923), and has been demonstrated more recently (Sancho & Kappen 1989). Some detailed morphological and anatomical studies concerning a smaller number of species (e.g., Scott & Larson 1984, five species of Umbilicaria; Codogno et al. 1989, three species) mention little structural variability for this thallus layer. However, a study that included a larger number of species has indicated the existence of several medullary types according to the hyphal arrangement within this layer (Sancho 1986). Clear implications of some of these different kinds of medulla for some physical and ecophysiological features of the Umbilicariaceae have been shown (Sancho & Kappen 1989; Valladares 1993; Valladares et al. 1993, 1994). The medulla has been used as a taxonomic character in several genera of macrolichens. For instance, a taxonomic rearrangement in which the medul- lary features played a relatively important role was the clustering within the genus Brodoa of most Hypogymnia species possessing some solid medullae *Centro de Ciencias Medioambientales, CSIC, Serrano 115 dup. 28006 Madrid, Spain. Present address: Department of Botany, University of California, Davis, California 95616, U.S.A. SDepartamento de Biologia Vegetal II, Facultad de Farmacia, Universidad Complutense de Madrid, 28040 Madrid, Spain. 0024-2829/95/030189+ 11 $8.0010 0 1995 The British Lichen Society
Transcript of Medullary structure of the Umbilicariaceae

Lichenologist 27(3): 189-199 (1995)
MEDULLARY STRUCTURE OF THE UMBILICARIACEAE
Fernando VALLADARES* and Leopold0 G. SANCHO#
Abstract: The medullary structure of 24 Umbilicariaceae species was studied by light microscopy. The objective was to investigate the variability of the medulla of these lichens and to assess the possible implications for thallus ecophysiology and the taxonomic relevance of the anatomy of this layer. Several methods of embed- ding, sectioning and staining together with the comparison of radial with transverse sections showed important differences in the degree of cohesion of hyphae and their spatial orientation within the medulla. Other important characteristics were the existence of more than one structurally homogeneous layer within the medulla, the frequency of hyphal branching, the relative thickness of the cell wall and lumen and the abundance of gelatinous matter in the intercellular spaces. The medullae studied were grouped into seven main types. The constancy of the types of medulla for certain groups of species makes it a useful secondary character to be considered in the taxonomy of the Umbilicariaceae. The important role of the structure of the medulla in the water relations of the Umbilicariaceae is discussed and a general relationship between medulla type and hydration strategy is suggested.
Introduction In the family Umbilicariaceae, as in most foliose lichens, the medulla is the thickest layer of the thallus. The two aspects of major interest concerning the structural features of the medulla of these lichens are their possible implica- tions for thallus ecophysiology and their taxonomic relevance. The crucial influence of the medulla in thallus water relations was mentioned in early studies of the Umbilicariaceae (Backmann 1923), and has been demonstrated more recently (Sancho & Kappen 1989). Some detailed morphological and anatomical studies concerning a smaller number of species (e.g., Scott & Larson 1984, five species of Umbilicaria; Codogno et al. 1989, three species) mention little structural variability for this thallus layer. However, a study that included a larger number of species has indicated the existence of several medullary types according to the hyphal arrangement within this layer (Sancho 1986). Clear implications of some of these different kinds of medulla for some physical and ecophysiological features of the Umbilicariaceae have been shown (Sancho & Kappen 1989; Valladares 1993; Valladares et al. 1993, 1994).
The medulla has been used as a taxonomic character in several genera of macrolichens. For instance, a taxonomic rearrangement in which the medul- lary features played a relatively important role was the clustering within the genus Brodoa of most Hypogymnia species possessing some solid medullae
*Centro de Ciencias Medioambientales, CSIC, Serrano 115 dup. 28006 Madrid, Spain. Present address: Department of Botany, University of California, Davis, California 95616, U.S.A. SDepartamento de Biologia Vegetal II, Facultad de Farmacia, Universidad Complutense de Madrid, 28040 Madrid, Spain.
0024-2829/95/030189+ 11 $8.0010 0 1995 The British Lichen Society

190 THE LICHENOLOGIST Vol. 27
(Goward 1986). However, the anatomy of the medulla has never been given extensive importance in the taxonomy of the Umbilicariaceae. Frey (1929) was the first to describe certain anatomical types within the family, but his interest focused on the upper and lower cortices, with few references to the medulla. Llano (1950) and Poelt (1977) also considered the structure of the medulla a secondary taxonomic character for these lichens. Nevertheless, the medullary structure has been useful as a complementary feature in distinguishing the genera Lasallia and Umbilicaria (Sancho & Balaguer 1989; Sancho & Crespo 1989), and more recently it has been considered in the description of the new species U. dendrophora (Hestmark 1993).
The goal of the present study was to describe more exactly the anatomy of the medulla of a large group of Umbilicariaceae, in order to assess the taxonomic importance of the medulla and its possible role in thallus eco- physiological behaviour. The main hypothesis on which the study was based is the relative constancy of the medullary structure for each species, but that important differences also exist between species or groups of species within this lichen family.
Materials and Methods Thalli of 24 Umbilicariaceae species were collected in dry conditions between 1980 and 1992. A complete list of localities is given in Sancho (1986) and Valladares (1993) and voucher specimens are in MAF herbarium. The taxa studied and the representative collection areas were as follows. Lasallia hispnnica (Frey) Sancho & Crespo: Spain; Avila, Leon, Madrid and Segovia. L. pustulata (L.) Merat: Spain, Madrid. Umbilicaria antarctica Frey & Lamb: Antarctica, South Shetland Islands. U. cinereontfescens (Schaer.) Frey: Spain, Avila. U. crusrttlosa (Ach.) Frey: Spain, Avila and Madrid. U. cylindrica (L.) Del. ex Duby: Spain, Avila and Madrid. U. decussata (Vill.) Zahlbr.: Spain, Avila. U. deusta (L.) Baumg.: Spain, Avila and Madrid. U. jreyi Codogno, Poeh & Puntillo: Spain, Avila, Leon, Madrid and Zaragoza. U. grisea Hoffm.: Portugal, Serra du Monchique; Spain, Avila, Caceres, Madrid and Toledo. U. havaasii Llano: Spain, Avila. U. hirsuta (SW. ex Westr.) Hoffm.: Spain, Avila and Madrid. U. hyperborea (Ach.) Hoffm.: Norway, Nordland and Hordaland; Spain, Asturias. U. leiocarpa DC. in Lam. & DC: Spain, Avila. U. nylandencwa (Zahlbr.) H. Magn.: Spain, Avila and Madrid. U. polyphylla (L.) Baumg.: Spain, Avila and Madrid. U. polyrrhiza (L.) Fr.: Spain, Avila, Leon and Madrid. U. proboscidea (L.) Schrad.: Norway, Nordland and Hordaland; Spain, Asturias, Avila, Caceres and Madrid. U. ruebeliana (DuRietz & Frey) Frey: Spain, Zaragoza. U. spodochroa (Ehrh. ex Hoffm.) DC.: Portugal, Serra da Estrela; Spain, Avila, Guadalajara and Segovia. U. spodochroa var. carpetann nom prov. (undescribed variety given this provisional name by Sancho 1986): Spain, Madrid. U. subglabra (Nyl.) Harm.: Spain, Avila. U. tolrefacta (Lightf.) Schrad.: Spain, Asturias, Avila, Leon and Madrid. U. vellea (L.) Ach.: Spain, Avila and Madrid. U. virginis Schaer.: Spain, Granada.
Thalli of average size for each species were taken in order to avoid the effects of thallus size and age (Larson 1984; Codogno et al. 1989). Thallus fragments were always taken from the intermediate zone between the margin and the umbilicus to avoid intrathalline variability (Valladares et al. 1994). The study of the medulla was carried out by light microscopy. Up to three different methods of sample preparation and sectioning were used. Thalli of all species were studied within a few days of collection from 5-urn-thick sections obtained with a Slee MTC cryostat and stained with lactophenol cotton blue. Samples of the most representative species of each medullary type were embedded in paraffin (as in Asta et al. 1986) and stained with Congo Red and lactophenol cotton blue. Serial sections of about 10 urn thick each were made, to give three-dimensional information on the arrangements of hyphae and bunches of hyphae within the medulla. Some samples were embedded in Spurr’s resin (as in Ascaso et al. 1986), to obtain 0.8 pm-thick sections. These sections were stained with methylene blue and used for detailed observation of the relative thickness of fungal cell walls and degree of conglutination of hyphae. Sections were made perpendicular to thallus surfaces following either of two orientations (Fig. 1).

1995 Umbilicariaceae medullary structure-V&z&-es & Sancho 191
FIG. 1. Sketch of an Umbilicariaceae thallus showing the situation of the umbilicus (u) and the two orientations of light microscopy sections considered in the present study: parallel or
perpendicular to the radii (radial, R, and transverse, T, respectively).
Radial sections were those parallel to the radii, whereas transverse sections were those perpen- dicular to the radii and consequently parallel to the thallus margins. All sections were examined with a Zeiss Axiophot light microscope.
In the use of anatomical concepts and terms we follow Frey (1936), Hale (1976), Poelt (1977) and Jahns (1973, 1988), but we include some particular terms used for the Umbilicariaceae, as employed in Sancho (1986), Sancho & Crespo (1989), Sancho & Kappen (1989), and Valladares er al. (1993).
Results
The two main characteristics that better differentiated the medullary types of the species of Umbilicariaceae were the degree of cohesion of hyphae and their spatial orientation. Other important characters for the description of the medulla of these lichens were the number of structurally different layers that can be distinguished within the medulla, the frequency of branching of the hyphae, the relative thickness of cell wall and lumen and the abundance of gelatinous matter in the intercellular spaces.
Most of the species of Umbilicariaceae posessed a medulla formed by a single layer of sparsely branched hyphae. These hyphae were generally meso- or pachydermatous (sensu Frey 1936) and loosely interwoven, as in many heteromerous lichens exhibiting the so-called medullary plectenchyma (sensu Hale 1976). However, even if some species of Umbilicariaceae can be included in this general medullary pattern (Fig. 2), many variations and exceptions of this model have been found in the species studied. Seven principal types of medullary anatomy were found in the Umbilicariaceae studied. We refer to each type by the name of one representative species.
1. Deustu type of medulla. Among those with a dense medulla (scleroplect- enchyma), this type can be recognized by the radial arrangement of periclinal hyphae made up of very long cells. In this radial scleroplectenchyma or prosoplectenchyma (equivalent to chondroid prosoplectenchyma sensu Ozenda & Clauzade 1970), the hyphae are agglutinated, with their walls frequently fused. The radial orientation of the hyphae is especially evident when radial and transverse sections of the same sample are compared (Figs 3A and B), The few intercellular spaces are usually filled with gelatinous matter. Besides U. deustu, this type of medulla has been found in U. polyphyllu, U. proboscideu and U. spodochrou (Fig. 4A).
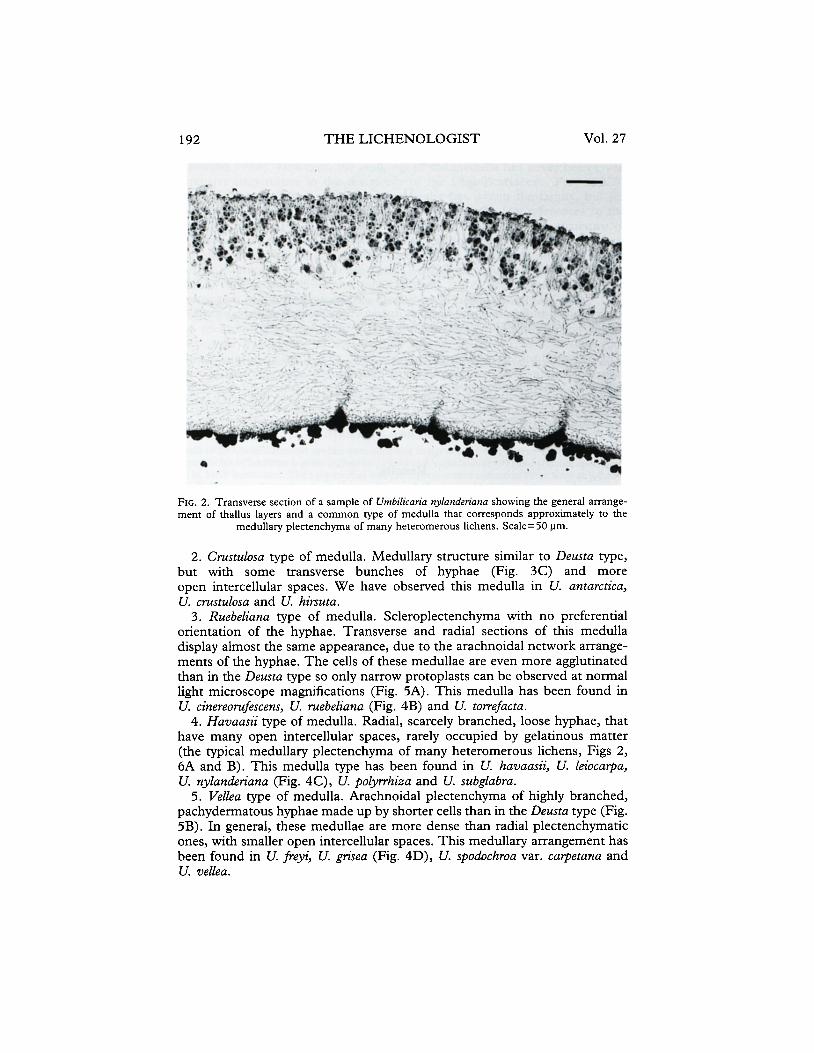
192 THE LICHENOLOGIST Vol. 27
FIG. 2. Transverse section of a sample of Utnbilicaria nylmdenma showing the general arrange- ment of thallus layers and a common type of medulla that corresponds approximately to the
medullary plecrenchyma of many heteromerous lichens. Scale= 50 pm.
2. Crustulosa type of medulla. Medullary structure similar to Deusta type, but with some transverse bunches of hyphae (Fig. 3C) and more open intercellular spaces. We have observed this medulla in U. antarctica, U. crustulosa and U. hirsuta.
3. Ruebeliuna type of medulla. Scleroplectenchyma with no preferential orientation of the hyphae. Transverse and radial sections of this medulla display almost the same appearance, due to the arachnoidal network arrange- ments of the hyphae. The cells of these medullae are even more agglutinated than in the Deusta type so only narrow protoplasts can be observed at normal light microscope magnifications (Fig. 5A). This medulla has been found in U. cinereomfescens, U. ruebeliana (Fig. 4B) and U. torrefacta.
4. Huvaasii type of medulla. Radial, scarcely branched, loose hyphae, that have many open intercellular spaces, rarely occupied by gelatinous matter (the typical medullary plectenchyma of many heteromerous lichens, Figs 2, 6A and B). This medulla type has been found in U. havaasii, U. leiocarpa, U. nylanderiana (Fig. 4C), U. polyrrhiza and U. subglabra.
5. Vellea type of medulla. Arachnoidal plectenchyma of highly branched, pachydermatous hyphae made up by shorter cells than in the Deustu type (Fig. 5B). In general, these medullae are more dense than radial plectenchymatic ones, with smaller open intercellular spaces. This medullary arrangement has been found in U. jkyi, U. grisea (Fig. 4D), U. spodochroa var. carpetana and U. vellea.

1995 Umbilicariaceae medullary structure-Vu&fares & Sancho 193
FIG. 3. Camera lucida drawings of two types of Umbilicariaceae medulla. A, Radial section of Dema type of medulla (radial scleroplectenchyma). B, Transverse section of Deusra type of medulla. C, Radial section of Cmsttrlosa type of medulla (a variation of the previous one with transverse bunches of hyphae). Solid black represents algal and fimgal cell protoplasts and fine
dots indicate gelatinous matter in intercellular spaces. Scale=20 urn.
6. Pustulutu type of medulla. A two-layered type of medulla. The upper part of the medulla is formed by a loosely arachnoidal plectenchyma, whereas the lower layer is constituted by a dense prosoplectenchyma (Fig. 6C). The lower part of the medulla is not present in the pustules, as has been already described (Sancho & Balaguer 1989). The two Spanish species of Lusullia, L. pustzrlatu and L. hispanica (Figs 3 and 4E), together with L. btiguntium Zschacke and
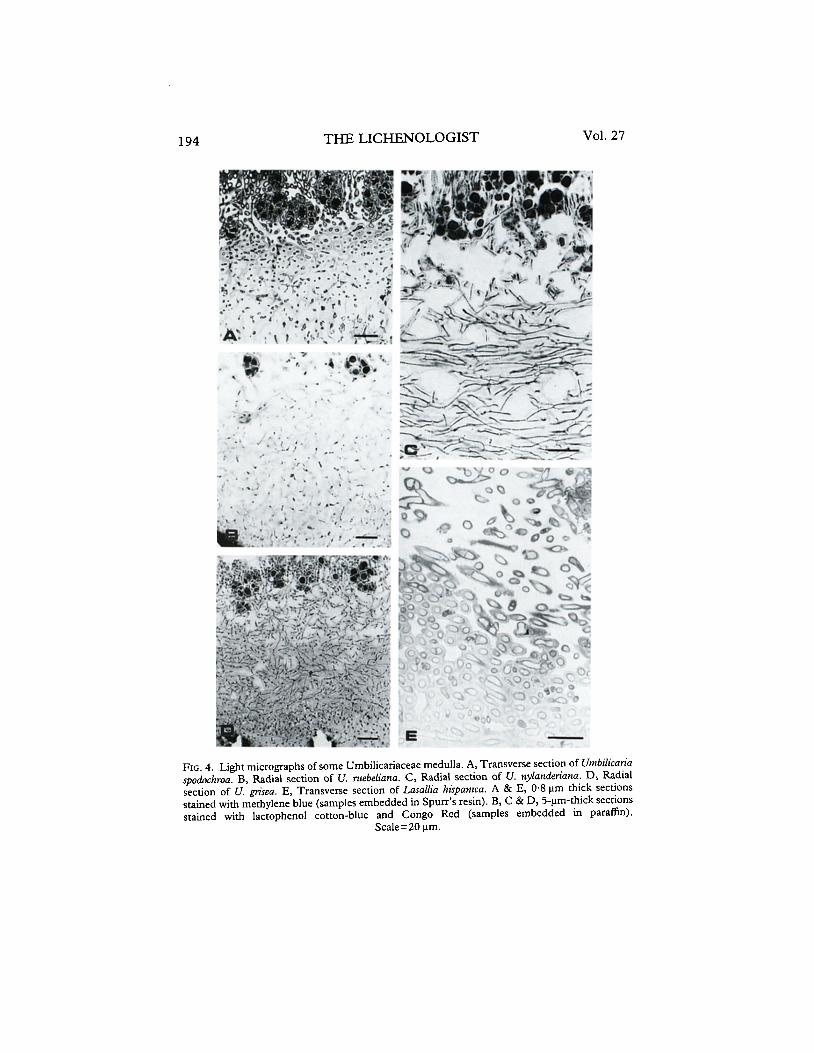
THE LICHENOLOGIST Vol. 27
FIG. 4. Light micrographs of some Umbilicariaceae medulla. A, Transverse section of Untbilicarik spodochroa. B, Radial section of U. ruebeliana. C, Radial section of U. nylamieriana. D, Radial section of U. grisea. E, Transverse section of Lasallia hispanica. A & E, 0.8 urn thick sections stained with methylene blue (samples embedded in Spurr’s resin). B, C & D, S-urn-thick sections stained with lactophenol cotton-blue and Congo Red (samples embedded in paraffin).
Scale=20 urn.

1995 Umbilicariaceae medullary structure-Vulladures & Sancho 195
FIG. 5. Camera lucida drawings of two types of Umbilicariaceae medulla. A, Radial section of Ruebelimza (arachnoidal scleroplectenchyma) type of medulla. B, Radial section of Vellea (arachnoidal plectenchyma) type of medulla. Solid black represents algal and fungal cell
proroplasts and fine dots indicate gelatinous matter in intercellular spaces. Scale=20 pm.
L. roissicu Drombovskaja (according to Sancho & Ballaguer 1989 and Sancho & Crespo 1989), show this type of medulla.
7. Decussutu type of medulla. This three-layered type of medulla consists of a central dense prosoplectenchymatic zone between two loosely arachnoidal layers that connect it with the algal layer and the lower cortex (Fig. 6D). This type of medulla has been found in U. cylindricu, U. decussutu, U. hyperboreu and U. virginis.
Discussion
Qualitative characteristics of the medulla similar to those mentioned here have been used to segregate species or groups of species within other genera of macrolichens such as Rumulinu (Krog & James 1977; Arroyo 1991) or Usneu (Clerc 1984, 1987; Ktirnefelt 1986). The medulla has been neglected in Umbilicariaceae taxonomy because it was considered similar for all the lichens of this family. On the contrary, we found a relatively constant and distinctive medullary pattern for each of several groups of species. However, the medullary typology we present here is not in disagreement with the obser- vations of other authors. For example, the relative structural uniformity found by Codogno et al. (1989) in the medulla of three species of U. hirsutu agg. could be explained by the fact that two of them (U. griseu and U. fiey;) share the VeZZeu type of medulla, and the third (U. hirsutu) exhibits a prosoplecten- chymatic medulla with some transverse bunches of hyphae (Crustulosu type)

196 THE LICHENOLOGIST Vol. 27
FIG. 6. Camera lucida drawings of three types of Umbilicariaceae medulla. A, Radial section of Havaasii (radial plectenchyma) type of medulla. B, Transverse section of Havaasii type of medulla. C, Radial section of the two-layered Pusculara type of medulla (arachnoidal plecten- chyma in the upper part and prosoplectenchyma in the lower part). D, Radial section of the three-layered Decussara type of medulla (central prosoplectenchymatic layer surrounded by two loosely arachnoidal layers). Solid black represents algal and timgal cell protoplasts and fine dots
indicate gelatinous matter in intercellular spaces. Scale=20 urn.

1995 Umbilicariaceae medullary structure-~&&ares & Sancho 197
that can present a similar image to that of the arachnoidal medullae of the other two species if the section is not completely radial or transverse. Scott & Larson (1984), despite having observed a constant medullary structure in the five species of Umbilicuria studied, found the medulla of U. muhlenbergii to be very distinctive due to its apparently great density, suggesting that structural differences in the medulla of Umbilicariaceae could be noted if this layer is studied thoroughly enough. The anatomy of the medulla in Umbilicariaceae can be used as a taxonomic feature to be evaluated with other morphological characters, since it can be useful for the segregation of some groups of taxa, as happens with the distinctive medulla of many species of the genus Lusullia (Sancho & Balaguer 1989; Sancho & Crespo 1989) or of the new species U. dendrophora (Hestmark 1993).
Our results provide additional support to the idea of Frey (1936), Schade (1955) and Poelt & Vezda (1981) that the genus Umbilicaria should not be divided into the genera Asyrophoru Nyl., Gyrophoru Ach., Omphalodiscus Schol. and Actinogyru Schol., as suggested by Scholander (1934), Llano (1950) and Thomson (1984), since none of the medullary types described here is restricted to any of these four taxa. The phylogenetic implications of the different types of medulla observed in the genus Umbilicariu are uncertain. It seems plausible that there was an early differentiation into two main groups of species, one with radial and another with arachnoidal medullae, from which the actual types evolved. However, there is scant agreement with the other characters considered in previous phylogenetic proposals for this lichen genus.
The medulla does not confer upon the thalli of Umbilicariaceae the same rigidity and hardness as the dense central cord of some fruticose lichens (e.g., Neuropogon sp.) but the thickness and density of this layer are responsible for most mechanical properties of the thallus of these lichens, as well as for certain physical features of the thallus such as its porosity (Valladares et al. 1993). Nevertheless, in our opinion, the most important aspect of medullary struc- ture for the biology of the lichens studied is its influence on thallus water relations. Honegger (1991) considered that there is no capillary water on the hydrophobic wall surfaces of the thalline interior, even in a fully hydrated thallus. This could be the case for certain loosely arachnoidal medullae studied here, where the accumulation of lichen phenolics, which could contribute to this possible hydrophobicity, was marked (e.g., the upper part of the arach- noidal layer of Lusulliu sp.). However, the hydrophilic nature of the gelatinous matter that is present in the medullae of many Umbilicariaceae species cannot be disregarded, although it remains unsubstantiated. The clear relationships between medullary anatomy and water-holding capacity (Sancho & Kappen 1989) and the significant correlations found between thallus porosity, type of medullae and water storage capacity (Valladares et al. 1993) are clear indications of the importance of this layer for the water relations of Umbili- cariaceae lichens. In addition, general hydration strategies can be associated with groups of species sharing a similar type of medulla. Umbilicuriu species with a prosoplectenchymatic (Deustu and Crustulosu types) or arachnoidal (VeZZue type) medulla usually grow in run-offs, obtaining liquid water directly from the substratum. Species with a radial plectenchymatic medulla (Huvausii type) are characteristic of vertical rocky surfaces or exposed localities where

198 THE LICHENOLOGIST Vol. 27
the major source of water is vapour contained in the air. Multilayered medullae correspond with widespread and abundant species (e.g., U. decus- sum, LusaZZiu pusntlutu), or lichens with remarkable ecological plasticity (e.g., U. cylindricu), suggesting that the combination of several types of medulla provides the thallus great adaptability. In agreement with these roles of the medulla in the hydration strategy of Umbilicariaceae are the observations of Larson (1984) and Scott & Larson (1984), which showed a relationship between the dense medulla of U. muhlenbergii and its preference for wet localities with liquid water available. However, U. grikeu, U. polyphyllu and U. polyrrhizu together with the three species with a scleroplectenchymatic, arachnoidal type of medulla cannot be easily included in this medulla-based hydration strategy presented. This sort of exception within the general trend in the family could be interpreted if other structural factors of clear relevance for thallus water relations (morphology, structure of both cortices) are included, as has been tentatively done elsewhere (Valladares 1994).
We are grateful to Mme. Letrouit-Galinou and her technician Monique Avnaim for valuable help with light microscopy techniques. Thanks are due to Carmen Ascaso and Esteban Manrique for helpful discussions and to two anonymous referees for their comments. Financial support was provided by The Spanish DGICYT (PB 90/0615).
REFERENCES Arroyo, R. (1991) El g&zero Ramalina Ach. en la Peninsula Ibkrica: quimica, quimioraxonomia,
mo+logia, anaromiu y disrribucidn. Tesis Doctoral, Universidad Complutense de Madrid. Ascaso, C., Brown, D. H. & Rapsch, S. (1986) The ultrastructure of the phycobiont of desiccated
and hydrated lichens. Lichenologisr 18: 37-46. Asta, J., Avnaim, M. & Letrouit, M. A. (1986) Colonisation dun sol verge par Baeomyces rufus
(Huds.) Rebent. (lichen) et Pogonarum urnigerum (Hedw.) I’. Beauv. (mousse): aspects morphologique et structural. Travaux Sciennjiques du Part National de la Vanoise 15: 83-10 1.
Bachmann, E. (1923) Untersuchungen iiber den Wasserhaushalt einiger Felsenflechten. Jahrbuch jiir Wissenschaftliche Boranik 62: 20-64.
Clerc, I’. (1984) Contribution a la revision de la systematique des usnies (Ascomycotina, Usnea) d’Europa. I. Usneaflorida (L.) Wigg. emend. Clerc. Cyptogamie, Byologie et Licht+nologie 34: 333-360.
Clerc, P. (1987) Systematics of the Usneafragilescens aggregate and its distribution in Scandinavia. Nordic Journal of Borany 7: 479-495.
Codogno, M., Poelt, J. & Puntillo, D. (1989) Umbilicaria jmyii spec. nova und der Formenkreis von Umbilicaria hirsura in Europe (Lichenes, Umbilicariaceae). Planr Sysrematics and Evolution 165: 55-69.
Frey, E. (1929) Beitriige zur Biologie, Morphologie und Systematik der Umbilicariaceen. Hedwigia 69: 219-252.
Frey, E. (1936) Vorarbeiten zu einer Monographie der Umbilicariaceae. Berichte Schweizer Boranische Gesellschaft 45: 198-230.
Goward, T. (1986) Brodoa, a new lichen genus in the Parmeliaceae. Bryologist 89: 219-223. Hale, M. E. (1976) Lichen structure viewed with the scanning electron microscope. In
Lichenology: Progress and Problems (D. H. Brown, D. L. Hawksworth & R. H. Bailey, eds): 1-16. London: Academic Press.
Hestmark, G. (1993) Umbilicaria dendrophora. Mycotaxon 46: 2 1 l-2 15. Honnegger, R. (1991) Functional aspects of the lichen symbiosis. Annual Review of Plant
Physiology and Planr Molecular Biology 42: 553-578. Jahns, H. M. (1973) Anatomy, morphology and development. In The Lichens (V. Ahmadjian St
M. E. Hale, eds) 3-57. New York: Academic Press. Jahns, H. M. (1988) The lichen thallus. In CRC Handbook of Lichenology 1 (M. Galun, ed.):
95-146. Boca Raton, FL: CRC Press, Inc.

1995 Umbilicariaceae medullary structure-V&x&n-es Q Szncho 199
KBmefelt, I. (1986) The genera Bryocaulon, Coelocaulon and Corniculuria and formerly associated taxa. Opera Boranica 86: l-90.
Ktog, H. & James, P. W. (1977) The genus Ramalina in Fennoscandia and the British Isles. Norwegian Journal of Botany 24: 15-43.
Larson, D. W. (1984) Thallus size as a complicating factor in the physiological ecology of lichens. New Phytologist 91: 87-97.
Llano, G. A. (1950) A Monograph of the Lichen Family Umbilicariaceae in the Western Hemisphere. Washington: Navexos. Office of Naval Research.
Ozenda, P. & Clauzade, G. (1970) Les Lichens. Paris: Masson et Cie. Poelt, J. (1977) Die Gattung Umbilicaria. Flechten des Himalaya 14. Khumbu Himal 6: 397435. Poelt, J. & Vezda, A. (198 1) Bestimmungschliissel europiiischer Flechten. Ergiinzungsheft II. Lehre:
J. Cramer. Sancho, L. G. (1986) Flora y vegetacidn liqueniuca saxicola de 10s pisos ore- y crioromediterrcineo de1
Sistema Central espariol. Tesis Doctoral, Universidad Complutense de Madrid. Sancho, L. G. & Balaguer, L. (1989) Estudio comparado de la anatomia de1 talo en el genera
Lassalia Merat (Umbilicariaceae; Lichenes). Anales Jardin BotPnico de Madrid 46: 273-28 1. Sancho, L. G. & Crespo, A. (1989) Lasallia hispanica and related species. Lichenologist 21: 45-58. Sancho, L. G. & Kappen, L. (1989) Photosynthesis and water relations and the role of anatomy
in Umbilicariaceae (Lichenes) from Central Spain. Oecologiu 81: 473-480. Schade, A. (1955) Zur sachischen Flechtenfiora insbesondere aus der Familie der Umbilicari-
aceen. Nova Acta Leopoldina 119: 195-255. Scholander, I’. F. (1934) On the apothecia in the lichen family Umbilicariaceae. Nytt Magasin for
Natuwidenskapene IS: l-3 1. Scott, M. G. & Larson, D. W. (1984) Comparative morphology and fine structure of a group of
Umbilican~ lichens. Canadian Journal of Botany 62: 1947-1964. Thomson, J. W. (1984) American Arctic Lichens. 1. The Macrolichens. New York: Columbia
University Press. Valladares, F. (1993) Estructura y propiedadesfiscas de1 talo liquenico en la familia Umbilicariaceae.
Tesis Doctoral, Universidad Complutense de Madrid. Valladares, F. (1994) Form-functional trends in Spanish Umbilicariaceae with special reference
to water relations. Cyptogamie, Byologie et Lichenologie (in press). Valladares, F., Wierzchos, J. & Ascaso, C. (1993) Porosimetric study of the lichen family
Umbilicariaceae. Anatomical interpretation and implications for the water storage capacity of the thallus. American youma of Botany 80: 263-272.
Valladares, F., Ascaso, C. & Sancho, L. G. (1994) Intrathalline variability of some structural and physical parameters in the lichen genus Lusallia. Canadian 3oumal of Botany (in press).
Accepted for publication 14 June 1994




















