
Novel Pheochromocytoma Susceptibility Loci Identified by Integrative Genomics
Upload
independentCategory
view
1download
0
Neurochemical Research, VoL 16, No. 4, 1991, pp. 489-500
Nicotinic Receptor-Elicited Sodium Flux in Rat Pheochromocytoma PC12 Cells: Effects of Agonists, Antagonists, and Noncompetitive Blockers
John W. Daly 1, Yukio Nishizawa 1, Michael W. Edwards 1, James A. Waters 1, and Robert S. Aronstam 2
(Accepted February 5, 1991)
Nicotinic agonists stimulate 22Na flux in rat pheochromoeytoma PC12 cells. The stimulatory effect of carbamylcholine is maximal at 1 mM, while the stimulatory effect of nicotine and anatoxin maximize at the same level at 100 ~M and 10 p~M, respectively. The tertiary amines arecolone and isoarecolone have no effect on flux at 100 IxM, while the methiodides at 100 ixM stimulate flux to an extent similar to 1 mM carbamylcholine. Dihydro and alcohol analogues of isoarecolone methiodide have markedly smaller effects on flux. A preincubation for 2 to 20 rain with carba- mylcholine (2 mM), nicotine (300 rzM), anatoxin (30 IxM) or isoarecolone methiodide (100 IxM) causes marked desensitization to a subsequent carbamylcholine-elicited stimulation of flux. d- Tubocurarine, mecamylamine, hexamethonium, and chlorisondamine inhibit carbamylcholine-elic- ited flux with ICso values of 1.0, 0.8, 43, and 0.020 IxM, respectively. Atropine has no effect at 1 ~M, but reduces the response to carbamylcholine by 50% at 8.6 IxM, presumably as a noncom- petitive blocker. Other noncompetitive blockers of nicotinic acetylcholine-receptors, such as his- trionicotoxins, gephyrotoxin, pumiliotoxin C, phencyclidine, bupivacaine and piperocaine, inhibit carbamylcholine-elicited stimulation of ZZNa flux with ICso values from 0.3 to 1.8 IxM. In contrast to d-tubocurarine, which inhibits carbamylcholine-elicited desensitization, and mecamylamine, which has no apparent effect on desensitization, chlorisondamine and certain noncompetitive block- ers appear to enhance desensitization. The effects of agonists, antagonists and noncompetitive blockers at the neuronal nicotinic acetylcholine receptor-channel of PC12 cells are compared to their effects on binding of [125I]e~-bungarotoxin to agonist-recognit ion sites and of [3H]perhydrohistrionicotoxin to noncompetitive blocker sites of the nicotinic acetylcholine receptor- channel of electric ray (Torpedo) electroplax membranes. There are marked differences in relative potencies for the two types of nicotinic acetylcholine receptor-channel.
KEY WORDS: Acetylcholine receptors; nicotine; desensitization.
I N T R O D U C T I O N
Nicotinic acetylcholine receptor-channels occur in neurons, in ganglia and in muscles. The subtype that occurs in muscle has been studied extensively. It is com-
i Laboratory of Bioorganic Chemistry, National Institutes of Health, Bldg. 8, Room 1A17, Bethesda, Maryland 20892 Department of Pharmacology and Toxicology, Medical College of Georgia, Augusta, Georgia 30912-3368
489
prised of two c~-subunits and single 13-, -/-, and ~-sub- units that are arranged to form a transmembrane pore, whose conducting properties are governed by the binding of nicotinic agonists to a site on each of the two o~- subunits (26). Binding of agonists and activation of the channel is blocked by d-tubocurarine and c~-bungaro- toxin. In addition, so-called noncompetitive blockers in- teract with the nicotinic receptor to block conductance through the activated channel, often by enhancing rates
0364-3190/91/0400-0489506.50/0 �9 1991 Plenum Publishing Corporation

490 Daly, Nishizawa, Edwards, Waters, and Aronstam
of desensitization (12). The electroplax of electric rays (Torpedo) contains a high density of nicotinic receptors analogous to the muscle nicotinic receptor channel of other vertebrates. The effect of agonists, antagonists and noncompetitive blockers on binding of an antagonist, [lzsI]c~-bungarotoxin and binding of noncompetitive blockers, such as [3H]perhydrohistrionicotoxin, to ni- cotinic receptor-channel complex in membranes from ray electroplax tiave been extensively studied (6-9, 20, 23) and often compared with electrophysiological data on muscle nicotinic receptor-channels (1, 5, 24, 47, 54, 55, 58, 59). In addition, other classes of nicotinic acetyl- choline receptor-channels are present in the central ner- vous system and in ganglia. Such nicotinic receptor channels differ in subunits from the muscle receptor and are insensitive to c~-bungarotoxin (61). Activation of ganglionic nicotinic receptor-channels by agonists is blocked by d-tubocurarine and by ganglionic blocking agents, such as hexamethonium, mecamylamine and chlorisondamine (32). Noncompetitive blockers interact with the neuronal and ganglionic subtypes of nicotinic receptor-channels in the central nervous system (22, 40), ganglia (37), and adrenal chromaffin cells (29, 33, 56, 57), but the interactions have not been studied in detail. The nicotinic receptor-channels are not identical in the central nervous system and ganglia, particularly with re- spect to agonist potency (28, 32).
The rat pheochromocytoma PC12 cell represents a model system for the study of effects of agents on a ganglionic nicotinic acetylcholine receptor-channel (30, 35, 38, 49, 50). Nicotine elicits a marked flux of ions through such channels and thereby triggers a release of norepinephrine (25, 34, 39, 42). d-Tubocurarine, but not c~-bungarotoxin block the response (27, 38). Ganglionic blockers are more potent in PC12 cells than in a med- ulloblastoma cell line that expresses a central nervous system-type of neuronal nicotinic receptor-channel (44). The receptor in PC12 cells can be blocked by noncom- petitive blockers (36, 50) and undergoes desensitization (36, 43, 50). The present study concerns the effect of a variety of agonists, antagonists and noncompetitive blockers on flux of 2ZNa through ganglionic-type nico- tinic receptor-channels of PC12 cells and on desensiti- zation of such receptors, and a comparison of the effects to binding affinities at the muscle-type nicotinic recep- tor-channels in Torpedo electroplax membranes.
EXPERIMENTAL PROCEDURE
Cell Cultures. Pheochromocytoma PC12 cells, provided by Dr. G. Guroff (NIH) were grown in Dulbecco's modified Eagles medium
with 5% fetal calf serum, 5% horse serum and penicillin (100 U/mI) and streptomycin (i00 Ixg/ml). Cells were grown at 37~ in an at- mosphere enriched in CO2. Cells were prelabelled with [3H]leucine for two days prior to final plating in order to be able to normalize 22Na ion flux to protein content for each well.
Assay of22Na § Flux Elicited by Activation of Nicotinic Receptor- Gated Ion Channels in PC12 Cells. [aH]Leucine-labeled PC12 (1.2- 1.3 million cells/well, 7000-12000 cpm all/well) were plated in six- well plates precoated with poly-D-lysine and cultured at least 24 hr. Media was removed by aspiration and 0.5 ml of preincubation buffer (concentrations in mM: NaC1 150; KC1 5.4; CaC12 2; HEPES/Tris, pH 7.4, 50; glucose 5) containing various agents was added at 22~ After 10 min, the preincubation buffer was replaced with influx buffer (concentration in mM: NaC1 50; KC1 5.4; CaC12 2; HEPES/Tris, pH 7.4, 50; glucose 5; sucrose 179; ouabain 5) containing 0.7 ixCi 2zNaCl, and in most cases 2 mM carbamylcholine and various agents. After 2 rain at 22~ the influx buffer was removed by aspiration and the cells washed three times with wash buffer (same composition as for prein- cubation buffer). Cells were sohbilized with 0.5 ml 1% SDS-0.5 N NaOH for 30-60 rain, pipetted in counting vials with 8 ml Hydrofluor and 0.25 ml 1 N HC1, and radioactivity (3H, Z~Na) determined in a scintillation counter. For desensitization experiments, cells were prein- cubated in desensitization buffer (composition in mM; NaC1, 10; KCI, 5.4; CaCl2, 2; HEPES/Tris, pH 7.4, 50; glucose 5; sucrose, 240) with or without various concentrations of carbamylcholine (1 IxM to 10 mM) and agents for 20 rain at 22~ The cells were washed one time with wash buffer, followed by incubation with influx buffer with 22NaC1 and 1 mM carbamylcholine for 2 min, and ZZNa+ influx assay as described above.
Dose response curves were analyzed for Ecso and ICso values by computer using a non-linear regression formula.
Tissue Preparation. Electric organs from Torpedo californica were purchased from Pacific Biomarine (Venice, CA) and stored at -70~ for up to 4 months before use. Electric organ was minced and then homogenized in a blender in 5 volumes of 50 mM Tris-C1, pH 7.4, containing 1 mM phenylmethylsuffonyl fluoride to prevent proteolysis. The homogenate was filtered through four layers of cheese cloth to remove undismpted material and then centrifuged at 20,000 g for 20 min. The pellets were resuspended in 50 mM Tris-C1 buffer and used without further treatment. Protein content was estimated by the method of Lowry et al. (31).
Binding of Radioligands to the Nicotinic Receptor Channel Com- plex in Torpedo Electroplax Membranes. [lzsI]-Alpha-bungarotoxin (pzSI]BGT, 10-20 la.Ci/lxg, Dupont-NEN) was used as a probe for receptor (i.e., acetylcholine) binding sites, torpedo membranes (5-15 Ixg protein) was incubated in the absence or presence of a competing agent for 15 min in 1 ml of 50 mM Tris-HC1, pH 7.4, at room tem- perature. [IZ~I]BGT (5 nM) was then added and the incubation contin- ued for 20 min. The reaction was quenched by the addition of 0.5 volumes of a solution containing 5 mg/ml of methylated bovine serum albumin. The suspensions were filtered on glass fiber filters (Whatman GF/B) that had been soaked in the albumin solution. The filters were washed once with 5 ml buffer, and their radioactivity content deter- mined by liquid scintillation counting. Nonspecific binding was de- termined in the presence of 30 ~M cx-tubocurarine.
Ion channel interactions were measured using [~H]perhydrohis- trionicotoxirk ([3H]H~2-HTX; 54.5 Ci/mmol), prepared by reduction of 6.6 mg of (_+)-oetahydmhistrionicotoxin with 25 Ci of tritium gas by New England Nuclear. Torpedo membranes (20-50 ~g protein) were incubated with 2 nM [3H]H12-HTX in the absence or presence of a competing agent in 50 mM Tris-HC1, pH 7.4, in a final volume of 1 ml at room temperature for 60 rain. The suspensions were filtered on
Nicot in ic Agen t s in P C 1 2 Cells 491
glass fiber filters (Whatman GF/B), and the filters were washed with 5 ml buffer and their radioactivity content determined by liquid scin- tillation counting. The filters were pretreated with a 1% organosilane solution (Sigmacote) to eliminate [3H]H12-HTX binding to the filters. Non-specific binding was determined in the presence of 100 ~M un- labeled histrionicotoxin. Receptor and ion channel binding values were determined in triplicate in each experiment, and each experiment was performed at least three times using tissue prepared from different fish.
Materials. Isoarecolone methiodide, 3,4-dihydroisoarecolone methiodide, isoarecolol methiodide, 3,4-dihydroisoarecolol methiod- ide, arecolone methiodide and isoarecolone HC1 were prepared as de- scribed (41, 48, 60). An alternate, more satisfactory route to isoarecolone methiodide was as follows: The ethylene ketal of 4-acetylpyridine was prepared based on the literature procedure (13). A solution of 65.5 g (0.5 mole) of 4-acetylpyridine and 46.6 g (0.75 mole) of ethylene glycol in 175 ml toluene containing 11.4 g (0.06 mole) p-toluenesul- fonic acid was refluxed using a Dean-Starke water separator for 28 hrs. Distillation in vacuo afforded pure ketal (b.p. 79-80 ~ 2.5 mm Hg), analyzing correctly for C, H, N. Reduction with NaBH4 in the usual manner (58) afforded nearly pure ethylene ketal of 1-methyl-4- acetyl-3,4-dehydropiperidine. Silica gel chromatography with 1.5 to 8% methanol in CHzClz:diethyl ether (3:1) yielded pure ethylene ketal as a pale yellow oil. A solution of 611 mg (3.3 mmol) of this ketal in 25 ml 50% aqueous ethanol containing 0.35 ml concentrated H2SO 4
was concentrated after 45 min at room temperature to less than 10 ml in vacu6 and the remaining solution neutralized with K2CO 3. A chlo- rof6rm extract (3 • 10 ml) was washed with water, dried over MgSO4 and evaporated in vacuo to yield 430 mg crude product. Silica gel chromatography with 2.5 to 4% methanol in CHzCl2:diethyl ether (3:1) gave pure isoarecolone. Quartemization with methyl iodide afforded isoareeolone methiodide (m.p. 153-155~ analyzing correctly for C, H, N and identical with material synthesized by a previous route (60).
Arecolone HC1 was synthesized as follows: l-Methyl-3-acetyl- pyridinium (1.55 g) in 35 ml of methanol was reduced with 1.0 g NaBH4, followed by oxidation with CrO3 in the manner previously developed for synthesis of isoarecolone (58). Ethereal HC1 was added to the crude arecolone, followed by silica gel chromatography with 6 to 8% methanol in CH2C12. After recrystallization from ethanol-diethyl ether the hygroscopic arecolone HCI salt had a m.p. 186-196~
dl-Anatoxin-a hydrochloride was prepared essentially as described by Danheiser et al (19): 9-t-Butoxycarbonyl(_+)anatoxin-a (74 mg) was generously provided by H. Danheiser (MIT) and yielded 43 mg of (_+_)anatoxin-a, which was dissolved in 5 ml diethyl ether and ethan- olic HC1 was added dropwise until precipitation was complete. After decanting the ether, followed by an ether wash, there was obtained 43 mg (-)anatoxin-a HCI, homogeneous by thin-layer chromatography. The hygroscopic salt was stored cold under Nz.
(-)Histrionicotoxin, (-)isodihydrohistrionicotoxin, and ( - ) in- dolizidine 239AB were isolated from dendrobatid frog skins as de- scribed (16, 17, 53). (-)Perhydrohistrionicotoxin was prepared by catalytic reduction of ( -)isodihydrohistrionicotoxin (17). The synthe- sis of (-4")piperidine 241D and biological activity of it and various analogs will be reported elsewhere. ( --- )Piperidine 241D has been iso- lated from a poison frog Dendrobates speciosus (18). (_)Octahy- drohistrionicotoxin and (+)gephyrotoxin were provided by Y. Kishi (Harvard University); (_-+-)indolizidine 223AB by O.E. Edwards (Na- tional Research Council, Ottawa, Canada), (+)pumiliotoxin C by A.G. Schultz (Rensselaer Polytechnic Institute, Troy, N.Y.) and cis/trans- 2-butyl-5-pentylpyrrolidine (pyrrolidine 197B) by T.H. Jones (NIH). (For a review on structures and biological activity of dendrobatid al- kaloids see ref. 15).
Phencyclidine HCI was provided by P. Skolnick (N/H); bupiva-
caine by Sterling-Winthrop Res. Inst. (Rensselaer, NY); piperocaine by Eli Lilly Co. (Indianapolis, IN), chlorisondamine HCI by Ciba- Geigy Corp. (Ardsley, NY). Carbamylcholine chloride, (-)nicotine, mecamylamine HC1, and hexamethonium chloride were from Sigma Chemical Co. (St. Louis, MO) and d-tubocurarine chloride from Boeh- ringer Mannheim (West Germany). Starting materials for syntheses were obtained from Aldrich Chemical Co. (Milwaukee, WI).
R E S U L T S
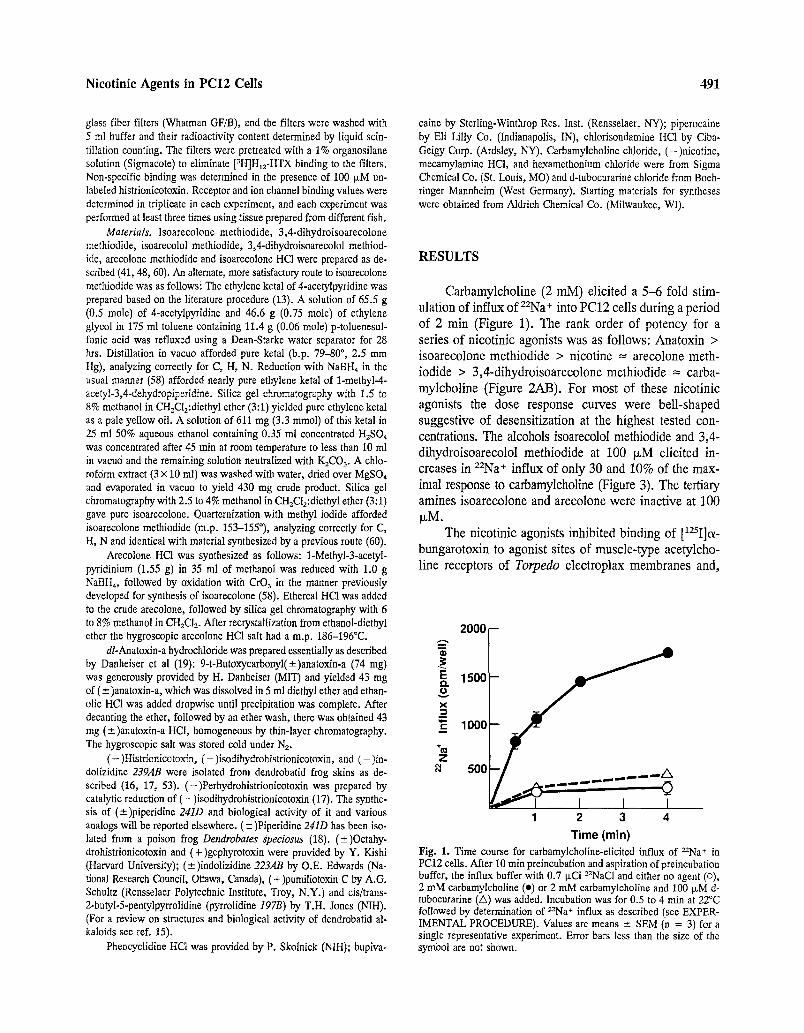
Carbamylcholine (2 raM) elicited a 5-6 fold stim- ulation of influx of ZZNa+ into PC12 cells during a period of 2 rain (Figure 1). The rank order of potency for a series of nicotinic agonists was as follows: Anatoxin > isoarecolone methiodide > nicotine = arecolone meth-
iodide > 3 ,4-dihydroisoarecolone methiodide = carba-
mylchol ine (Figure 2AB). For most of these nicot inic agonists the dose response curves were bel l -shaped suggestive of desensit izat ion at the highest tested con-
centrations. The alcohols isoarecolol methiodide and 3,4- dihydroisoarecolol methiodide at 100 txM elicited in- creases in 22Na+ inf lux of on ly 30 and 10% of the max-
imal response to carbamylcholine (Figure 3). The tertiary amines isoarecolone and arecolone were inactive at 100
IxM. The nicot inic agonists inhibited b ind ing of [125I]a-
bungarotoxin to agonist sites of muscle- type acetylcho-
l ine receptors of Torpedo electroplax membranes and,
2000 I
x~ 1500 I
iooo-
500
I I I I 1 2 3 4
Time (min) Fig. 1. Time course for carbamylcholine-elicited influx of 22Na+ in PC12 cells. After 10 rain preincubation and aspiration of preincubation buffer, the influx buffer with 0.7 txCi 22NaC1 and either no agent (o), 2 mM carbamylcholine (e) or 2 mM carbamylcholine and 100 ~M d- tnbocurarine (A) was added. Incubation was for 0.5 to 4 min at 22~ followed by determination of 22Na+ influx as described (see EXPER- IMENTAL PROCEDURE). Values are means _+ SEM (n = 3) for a single representative experiment. Error bars less than the size of the symbol are not shown.

492 Daly, Nishizawa, Edwards, Waters, and Aronstam
.c
o o
100 c o
s0 E
E 4 0
*~ z
A
-6 -5 -4 -3 -2
log [Agonist l , M
J~ o 8 o
100 o
80
E 6Q
E "6 40
20 ==
Z
I I -6 -5 -4
log [Agonist l , M
I I - 3 - 2
Fig. 2. Effect of nicotinic agonists on influx of 22Na§ in PC12 cells. Agonists at various concentrations were incubated for 2 min at 22~ with 2zNaC1 and influx of ~Na+ determined as described (see EX- PERIMENTAL PROCEDURES). A. (• (~); (-)nicotine (o); and carbamylcholine (CaCh o). B. Isoarecolone methiodide (&); arecolone methiodide (t~); 3,4-dihydroisoarecolone methiodide (=). The methiodides were not tested at > 100 ~M. The responses to 100 ~zM (• (-)nicotine and isoarecolone methiodide were blocked by 100 ~M d-tubocurarine (data not shown). Values are means • SEM (n = 3). Error bars less than the size of the symbol are not shown.
l ike c a r b a m y l c h o l i n e , e n h a n c e d b i n d i n g of [3H]perhydrohistrionicotoxin to noncompetitive blocker sites (Figure 4, Table I). The rank order of potency as inhibitors at the agonist site was as follows: 3,4-Dihy- droisoarecolone methiodide > arecolone methiodide > isoarecolone methiodide > isoarecolol methiodide = an- atoxin = nicotine > carbamylcholine > 3,4-dihydro- isoarecolol methiodide. The tertiary amines isoarecolone and arecolone had very low activity (Table I).
The response to 2 mM carbamylcholine was blocked by the nicotinic antagonist d-tubocurarine with a ICso of 1.0 txM (Figure 5). The ganglionic blocker chlorison- damine was very potent with an ICso of 20 nM. The ganglionic blockers mecamylamine and hexamethonium chloride were less potent with ICso values of 0.25 txM
120
8 100
8o
6o
20
H,c C)~ NC~ OH,
= = o.o. c.o.
c~ c~ c~
IsoAr Ar IsoAr Ar H2-1soAr I$oAml H=-IsoArol Mel Mel MOI Mel Mel
Fig. 3. Effect of isoarecolone, arecolone and methiodides on influx of ~Na + in PC12 cells. Isoarecolone (IsoAr), arecolone (At), their methiodides (MeI), 3,4-dihydro(H2-)-isoarecolone methiodides and isoarecolol (IsoArol) methiodides (100 ~M) were incubated for 2 min at 22~ with 2zNaC1 and influx of 22Na determined as described (see EXPERIMENTAL PROCEDURES). Values relative to 1 mM carba- mylcholine (eaCh) are means --- SEM (n = 3).
Arecolone Mel 1.0 ~ �9 Isoarecolone Mel
~ ~ Nicotine
0 . 8 .
~- o.B
0 .4
0 .2
0 .0 -9 -8 -7 -6 -5 -4 -3
log Concentration, k,1
Fig. 4. Inhibition of [lz~I]BuTx binding to the nicotinic acetylcholine receptor-channel complex from Torpedo electric organ by nicotinic agonists. The binding of 5 nM [lzSI]BuTx was measured in the pres- ence of arecolone methiodide (o), isoarecolone methiodide (e), nico- tine (A), anatoxin-a (&) or carbamytcholine (cz). Binding is expressed as fraction of total specific binding measured in the absence of com- peting ligands. Each point and bar represents the mean and standard deviation from 3 experiments.
and 43 IxM, respectively. The Hill coefficients for these blockers were near unity (0.8 to 1.1).
The interactions of the antagonists with the agonist recognition site of nicotinic receptor (i.e., [lzSI]BuTx binding site) and associated sites that recognize noncom- petitive blockers (i.e., [3H]Ht2-HTX binding site) of
Nicotinic Agents in PC12 Cells 493
Table I. Effect of Nicotinic Agonists on Binding of [xz~I] ot-Bungarotoxin (p~I]) and [3H]Perhydrohistrionicotoxin ([3H]H12-
HTX) Binding to the Nicotinic Receptor-Channel Complex in Torpedo Electroplax Membranes
[ iii i
ICso, ~M ECso, I~M v e r s u s v e r s u s
[125I]BuTx [3H]H~z-HTX Agonist Binding Binding
Carbamylcholine 2.8 • 0.5 0.18 --- 0.04 (-)Nicotine 1.0 • 0.4 0.21 • 0.05 (• 1.1 • 0.3 0.22 • 0.08 Isoarecolone methiodide 0.48 • 0.06 0.021 • 0.009 Arecolone methiodide 0.32 • 0.07 0.07 • 0.02 3,4-Dihydroisoarecolone 0.21 • 0.04 0.05 • 0.01
methiodide Isoarecolol methiodide 1.2 --- 0.5 0.11 --- 0.04 3,4-Dihydroisoarecolol 15 • 6 2.1 • 0.4
methiodide
Binding assays were as described in EXPERIMENTAL PROCE- DURES. The ICso values are means - SEM (n = 3) and were de- termined in competition experiments with [lzsI]BuTx. The Ecs~ values are means • SEM (n = 3) and were derived from agonist activation of [3H]H12-HTX binding; i.e., the concentration which stimulated 1 nM [3H]Hx2-HTX binding to one-half of the maximal extent.
~" 100 F
F sol-
| ~ 40[-- z = /
~ 20
-9 -8 -7 -6 -5 -4 -3
-log [Agent], M
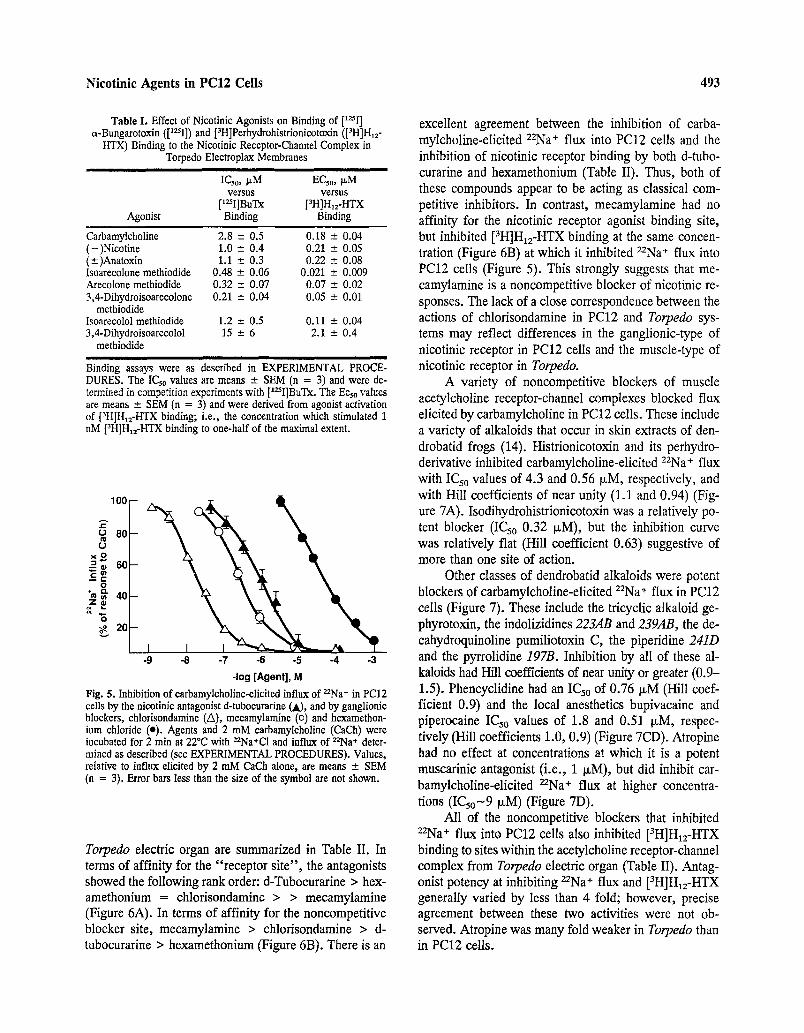
Fig. 5. Inhibition of carbamylcholine-elicited influx of ~Na § in PC12 cells by the nicotinic antagonist d-tubocurarine (A), and by ganglionic blockers, chlorisondamine (A), meeamylamine (o) and hexamethon- ium chloride (e). Agents and 2 mM carbamylcholine (CaCh) were incubated for 2 min at 22~ with 22Na+CI and influx of 22Na* deter- mined as described (see EXPERIMENTAL PROCEDURES). Values, relative to influx elicited by 2 mM CaCh alone, are means - SEM (n = 3). Error bars less than the size of the symbol are not shown.
Torpedo electric organ are summarized in Table II. In terms of affinity for the "receptor site", the antagonists showed the following rank order: d-Tubocurarine > hex- amethonium = chlorisondamine > > mecamylamine (Figure 6A). In terms of affinity for the noncompetitive blocker site, mecamylamine > chlorisondamine > d- tubocurarine > hexamethonium (Figure 6B). There is an
excellent agreement between the inhibition of carba- mylcholine-elicited ZZNa* flux into PC12 cells and the inhibition of nicotinic receptor binding by both d-tubo- curarine and hexamethonium (Table II). Thus, both of these compounds appear to be acting as classical com- petitive inhibitors. In contrast, mecamylamine had no affinity for the nicotinic receptor agonist binding site, but inhibited [3H]H12-HTX binding at the same concen- tration (Figure 6B) at which it inhibited 2ZNa+ flux into PC12 cells (Figure 5). This strongly suggests that me- camylamine is a noncompetitive blocker of nicotinic re- sponses. The lack of a close correspondence between the actions of chlorisondamine in PC12 and Torpedo sys- tems may reflect differences in the ganglionic-type of nicotinic receptor in PC12 cells and the muscle-type of nicotinic receptor in Torpedo.
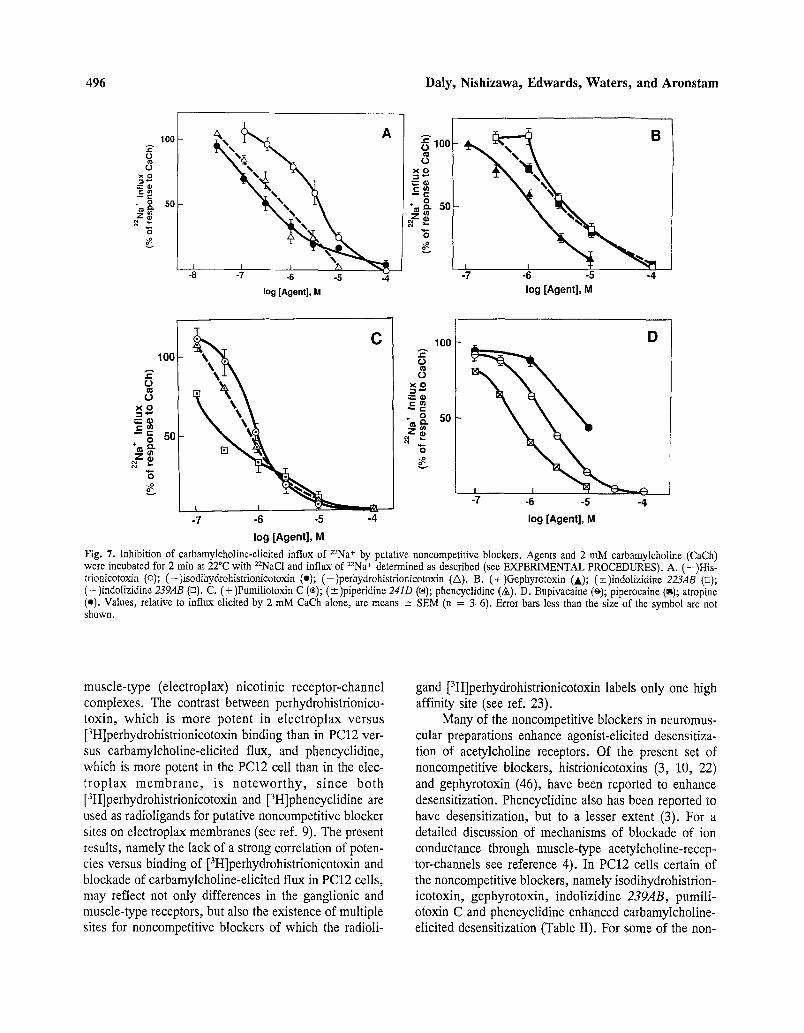
A variety of noncompetitive blockers of muscle acetylcholine receptor-channel complexes blocked flux elicited by carbamylcholine in PC12 cells. These include a variety of alkaloids that occur in skin extracts of den- drobatid frogs (14). Histrionicotoxin and its perhydro- derivative inhibited carbamylcholine-elicited Z2Na+ flux with ICso values of 4.3 and 0.56 IxM, respectively, and with Hill coefficients of near unity (1.1 and 0.94) (Fig- ure 7A). Isodihydrohistrionicotoxin was a relatively po- tent blocker (IC5o 0.32 ixM), but the inhibition curve was relatively flat (Hill coefficient 0.63) suggestive of more than one site of action.
Other classes of dendrobatid alkaloids were potent blockers of carbamylcholine-elicited 22Na+ flux in PC12 cells (Figure 7). These include the tricyclic alkaloid ge- phyrotoxin, the indolizidines 223AB and 239AB, the de- cahydroquinoline pumiliotoxin C, the piperidine 241D and the pyrrolidine 197B. Inhibition by all of these al- kaloids had Hill coefficients of near unity or greater (0.9- 1.5). Phencyclidine had an IC5o of 0.76 IxM (Hill coef- ficient 0.9) and the local anesthetics bupivacaine and piperocaine IC5o values of 1.8 and 0.51 txM, respec- tively (Hill coefficients 1.0, 0.9) (Figure 7CD). Atropine had no effect at concentrations at which it is a potent muscarinic antagonist (i.e., 1 IxM), but did inhibit car- bamylcholine-elicited 2ZNa* flux at higher concentra- tions (IC5o-9 ~M) (Figure 7D).
All of the noncompetitive blockers that inhibited 22Na§ flux into PC12 cells also inhibited [3H]Hlz-HTX binding to sites within the acetylcholine receptor-channel complex from Torpedo electric organ (Table II). Antag- onist potency at inhibiting 22Na+ flux and [3H]Hlz-HTX generally varied by less than 4 fold; however, precise agreement between these two activities were not ob- served. Atropine was many fold weaker in Torpedo than in PC12 cells.
494 Daly, Nishizawa, Edwards, Waters, and Aronstam
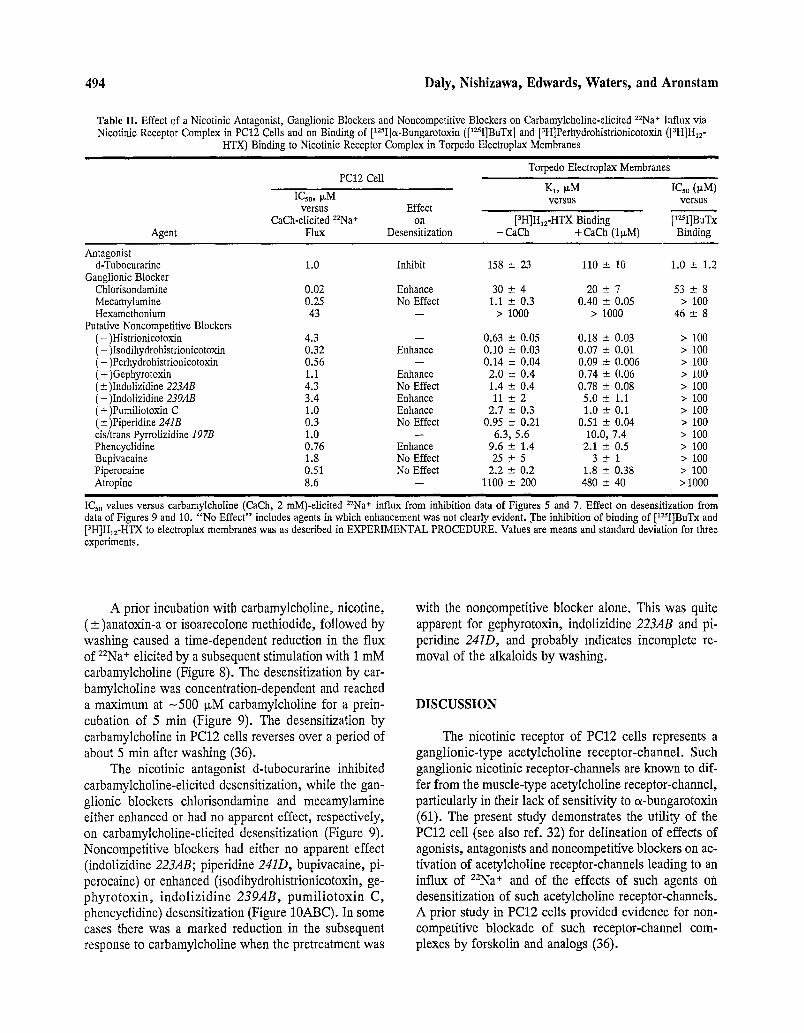
Table II. Effect of a Nicotinic Antagonist, Ganglionic Blockers and Noncompetitive Blockers on Carbamylcholine-elicited 2ZNa+ Influx via Nicotinic Receptor Complex in PC12 Cells and on Binding of [lz~I]a-Bungarotoxin ([lzSI]BuTx] and [3H]Perhydrohistrionicotoxin ([3H]H12-
HTX) Binding to Nicotinic Receptor Complex in Torpedo Electroplax Membranes
PC12 Cell Torpedo Electroplax Membranes
K1, p~M IC30 (p,M) ICso, I~M versus versus
versus Effect CaCh-elicited 22Na+ on [3H]HI,-HTX Binding [lzSI]BuTx
Agent Flux Desensitization - CaCh + CaCh (I~M) Binding
Antagonist d-Tubocurarine 1.0 Inhibit 158 -- 23 110 -- 10 1.0 -+ 1.2
Ganglionic Blocker Chlorisondamine 0.02 Enhance 30 +-- 4 20 -+ 7 53 + 8 Mecamylamine 0.25 No Effect 1.1 -- 0.3 0.40 -2_ 0.05 > 100 Hexamethonium 43 - - > 1000 > 1000 46 -+ 8
Putative Noncompetitive Blockers (-)Histrionicotoxin 4.3 - - 0.63 -2_ 0.05 0.18 -+ 0.03 > 100 (-)Isodihydrohistrionicotoxin 0.32 Enhance 0.10 - 0.03 0.07 -- 0.01 > 100 (-)Perhydrohistrionicotoxin 0.56 - - 0.14 --- 0.04 0.09 +- 0.006 > 100 (+)Gephyrotoxin 1.1 Enhance 2.0 - 0.4 0.74 - 0.06 > 100 (__)Indolizidine 223AB 4.3 No Effect 1.4 -4- 0.4 0.78 - 0.08 > 100 (-)Indolizidine 239AB 3.4 Enhance 11 _-2 2 5.0 +- 1.1 > 100 (+)Pumiliotoxin C 1.0 Enhance 2.7 --+ 0.3 1.0 -- 0.1 > 100 (--_)Piperidine 24IB 0.3 No Effect 0.95 -+ 0.21 0.51 --- 0.04 > 100 cis/trans Pyrrolizidine 197B 1.0 - - 6.3, 5.6 10.0, 7.4 > 100 Phencyclidine 0.76 Enhance 9.6 -+ 1.4 2.1 -+ 0.5 > 100 Bupivacaine 1.8 No Effect 25 -+ 5 3 - 1 > 100 Piperocaine 0.51 No Effect 2.2 -- 0.2 1.8 -+ 0.38 > 100 Atropine 8.6 - - 1100 +- 200 480 -- 40 >1000
IC5o values versus carbamylcholine (CaCh, 2 mM)-elicited 22Na* influx from inhibition data of Figures 5 and 7. Effect on desensitization from data of Figures 9 and 10. "No Effect" includes agents in which enhancement was not clearly evident. ,The inhibition of binding of [12SI]BuTx and [3H]Hlz-HTX to electroplax membranes was as described in EXPERIMENTAL PROCEDURE. Values are means and standard deviation for three experiments.
A prior incubation with carbamylcholine, nicotine, (_)anatoxin-a or isoarecolone methiodide, followed by washing caused a time-dependent reduction in the flux of 22Na+ elicited by a subsequent stimulation with 1 mM carbamylcholine (Figure 8). The desensitization by car- bamylcholine was concentration-dependent and reached a maximum at -500 t~M carbamylcholine for a prein- cubation of 5 min (Figure 9). The desensitization by carbamylcholine in PC12 cells reverses over a period of about 5 min after washing (36).
The nicotinic antagonist d-tubocurarine inhibited carbamylcholine-elicited desensitization, while the gan- glionic blockers chlorisondamine and mecamylamine either enhanced or had no apparent effect, respectively, on carbamylcholine-elicited desensitization (Figure 9). Noncompetitive blockers had either no apparent effect (indolizidine 223AB; piperidine 241D, bupivacaine, pi- perocaine) or enhanced (isodihydrohistrionicotoxin, ge- phyrotoxin, indolizidine 239AB, pumiliotoxin C, phencyclidine) desensitization (Figure 10ABC). In some cases there was a marked reduction in the subsequent response to carbamylcholine when the pretreatment was
with the noncompetitive blocker alone. This was quite apparent for gephyrotoxin, indolizidine 223AB and pi- peridine 241D, and probably indicates incomplete re- moval of the alkaloids by washing.
DISCUSSION
The nicotinic receptor of PC12 cells represents a ganglionic-type acetylcholine receptor-channel. Such ganglionic nicotinic receptor-channels are known to dif- fer from the muscle-type acetylcholine receptor-channel, particularly in their lack of sensitivity to o~-bungarotoxin (61). The present study demonstrates the utility of the PC12 cell (see also ref. 32) for delineation of effects of agonists, antagonists and noncompetitive blockers on ac- tivation of acetylcholine receptor-channels leading to an influx of 22Na+ and of the effects of such agents on desensitization of such acetylcholine receptor-channels. A prior study in PC12 cells provided evidence for non- competitive blockade of such receptor-channel com- plexes by forskolin and analogs (36).

Nicotinic Agents in PC12 Cells 495
1.0
�9 ~ 0.8 "o .=_ 113
~. 0.6 g~ tO 0.4 :,"z
0.2
0.0 -9 -2
�9 Hexamelhoniurn ~ ~. C h l o r i s o n d a m i n e ~ I I N
-'- , ~,my,a~,.o. , " " e ~ "t,
-8 -7 -6 -5 -4 -3
Log Concentration, M
i J i J
B. 1.o
0.8 m
x P" 0.6 z,
0 . 4 I Mec 9~. " ~ Chlorisondamine ~ k
0 . 2 - - - - ~ Tubocurarine k K ta ~k Hexamethonium ~ .
0.0 ' ~ ' ' ' ~
-10 -9 -8 -7 -6 -5 -4 -3
Log Concentration, M Fig. 6. Effect of nicotinic antagonists on ligand binding to the nicotinic acetylcholine receptor-channel complex from Torpedo electric organ. A. The binding of 5 nM [125I]BuTx to the nicotinic receptor (i.e., to the acetylcholine binding site) and B. the binding of 2 nM [3H]H12- HTX to a site for noncompetitive blockers were measured in the pres- ence of the indicated concentrations of d-tubocurarine (o), hexame- thonium (e), chlorisondamine (A), or mecamylamine (A). Binding is expressed as fraction of total specific binding measured in the absence
3 of competing ligands. [ H]H12-HTX binding was measured in the pres- ence of 1 ~,M carbamylcholine. Each point and bar represents the mean and standard deviation from 3 experiments.
-2
The activity of a series of quaternary and tertiary amines as activators of sodium flux in PC12 cells is reminiscent of the effects of these agents in neuromus- cular preparations (48, 60) and in antagonism of binding of [12SI]ct-bungarotoxin to agonist recognition sites on the acetylcholine receptor of Torpedo electroplax (Table I). Thus, the tertiary amines (_)anatoxin-a and ( - )ni - cotine are very potent (see ref. 10,51) while the tertiary amines isoarecolone and arecolone are nearly inactive. Of the quaternary amines isoarecolone methiodide was very potent as was the case in the neuromuscular prep- aration (48, 60) and in the electroplax membranes (Table I). However, the correlation between potencies in PC12
cells and electroplax membranes for this series of eight agonists was relatively weak (see Table I), suggesting significant differences in agonist recognition sites in the ganglionic (PC12 cells) and muscle-type (electroplax membranes) acetylcholine receptor-channels.
The response to carbamylcholine was blocked by the nicotinic antagonist d-tubocurarine and by the gan- glionic blockers chlorisondamine > mecamylamine > hexamethonium in PC12 cells (Figure 5). d-Tubocurar- ine shows comparable potency in PC12 cells and in elec- troplax membranes, while the ganglionic blockers are more potent versus the neuronal (ganglionic) acetylcho- line receptor in PC12 cells than for the muscle acetyl- choline receptor of electroplax (Table II). The lack of correspondence between potencies of ganglionic block- ers at the ganglionic (PC12 cells) and muscle-type (elec- troplax) nicotinic acetylcholine receptor-channels, suggests differences in the two classes of nicotinic receptors with regard to these ganglionic blockers. The low affinity of mecamylamine versus [125I]BuTx binding and relatively high potency versus binding of the noncompetitive blocker [3H]H12-HTX suggest that this ganglionic blocker acts not at the receptor, but as a noncompetitive blocker (see below).
The noncompetitive blockers utilized in the present study have all been established as such in studies with neuromuscular preparations and with electroplax mem- branes. These include alkaloids from dendrobatid frogs, such as the histrionicotoxins (9.47), gephyrotoxin (46) and pumiliotoxin C (58), and, in addition, phencyclidine (5), bupivacaine (24), piperocaine (52) and atropine (2). Mecamylamine also causes a noncompetitive block in neuromuscular preparations and antagonizes binding of [3H]perhydrohistrionicotoxin to sites in electroplax membranes (55, and Table II). Mecamylamine also blocks effects of nicotine at central acetylcholine receptors (21). It seems likely that mecamylamine acts as a noncom- petitive blocker at all of the nicotinic receptor-channel complexes. Other alkaloids from dendrobatid frogs, namely indolizidines 223AB and 239AB, the piperidine 241D and the pyrrolidine 197B, act as noncompetitive blockers in binding studies with electroplax membranes (6,7, this paper), but have not as yet been studied elec- trophysiologically in neuromuscular preparations. Cer- tain piperidines, similar in structure to 241D do act as noncompetitive blockers in neuromuscular preparations (62). There is no clear correlation between potencies of the thirteen noncompetitive blockers for the neuronal (PC12 cells) and potencies in binding studies for muscle (electroplax membranes) acetylcholine receptor-chan- nels (see Table II), suggesting differences in the non- competitive blocker sites on ganglionic (PC12 cell) and
496 Daly, Nishizawa, Edwards, Waters, and Aronstam
= 1oo ~ x ~ " A
~ .~o
. o = s 0 o .
I I
-a -7 -6 -s -~ - log [Agent], M
100 ~"
,-,= , ,1 ,
xo "E~ = " | 7o so
a | ~ ,,7-=,.--- ~ 7 6 5 4
-7 -6 -5 -4 log [Agent], M log [Agent], M
Fig. 7. Inhibition of carbamylcholine-elicited influx of 22Na+ by putative noncompetitive blockers. Agents and 2 mM carbamylcholine (CaCh) were incubated for 2 min at 22~ with 22NaCI and influx of 22Na+ determined as described (see EXPERIMENTAL PROCEDURES). A. (-)His- trionicotoxin (o); (-)isodihydrohistrionicotoxin (o); (-)perhydrohistrionicotoxin (A). B. (+)aephyrotoxin (A); (+_)indotizidine 223AB (•); (-)indolizidine 239AB (rn). C. (+)Pumiliotoxin C (| (_+)piperidine 241D (r~); phencyclidine (&). D. Bupivacaine (~,); piperocaine (s); atropine (e). Values, relative to influx elicited by 2 mM CaCh alone, are means • SEM (n = 3-6). Error bars less than the size of the symbol are not shown.
muscle-type (electroplax) nicotinic receptor-channel complexes. The contrast between perhydrohistrionico- toxin, which is more potent in electroplax versus [3H]perhydrohistrionicotoxin binding than in PC12 ver- sus carbamylcholine-elicited flux, and phencyclidine, which is more potent in the PC12 cell than in the elec- troplax membrane, is noteworthy, since both [3H]perhydrohistrionicotoxin and [3H]phencyclidine are used as radioligands for putative noncompetitive blocker sites on electroplax membranes (see ref. 9). The present results, namely the lack of a strong correlation of poten- cies versus binding of [3H]perhydrohistrionicotoxin and blockade of carbamylcholine-elicited flux in PC12 cells, may reflect not only differences in the ganglionic and muscle-type receptors, but also the existence of multiple sites for noncompetitive blockers of which the radioli-
gand [3H]perhydrohistrionicotoxin labels only one high affinity site (see ref. 23).
Many of the noncompetitive blockers in neuromus- cular preparations enhance agonist-elicited desensitiza- tion of acetylcholine receptors. Of the present set of noncompetitive blockers, histrionicotoxins (3, 10, 22) and gephyrotoxin (46), have been reported to enhance desensitization. Phencyclidine also has been reported to have desensitization, but to a lesser extent (3). For a detailed discussion of mechanisms of blockade of ion conductance through muscle-type acetylcholine-recep- tor-channels see reference 4). In PC12 cells certain of the noncompetitive blockers, namely isodihydrohistrion- icotoxin, gephyrotoxin, indolizidine 239AB, pumili- otoxin C and phencyclidine enhanced carbamylcholine- elicited desensitization (Table II). For some of the non-
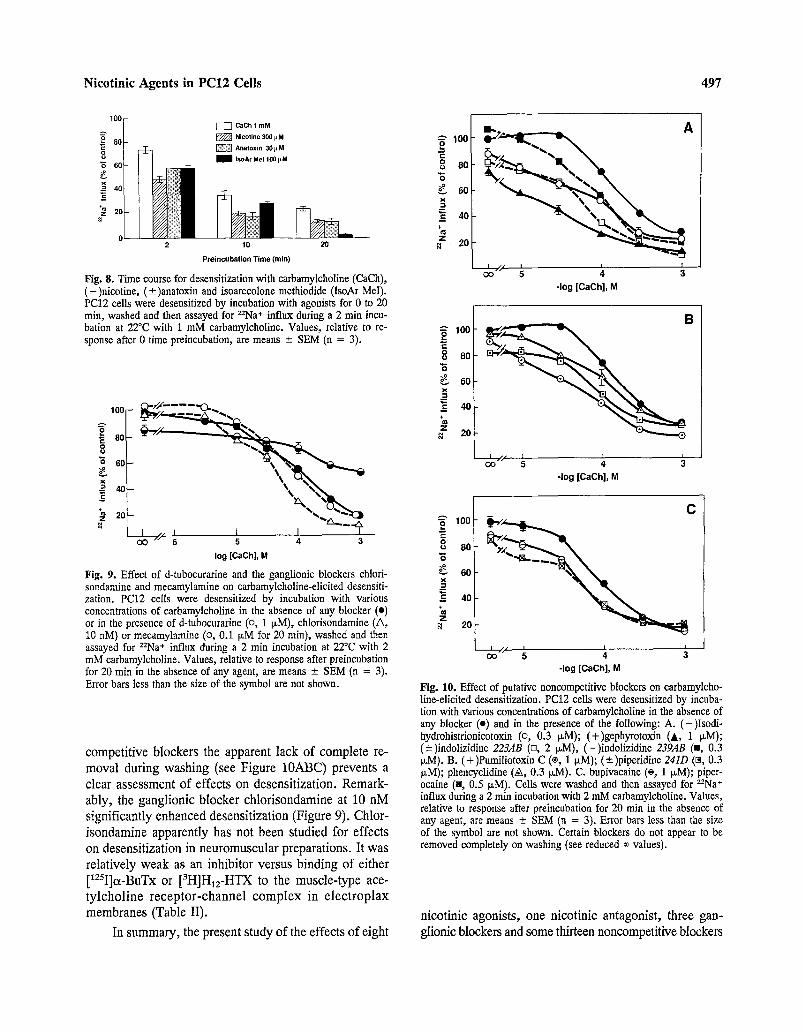
Nico t in i c A g e n t s in P C 1 2 Cel ls 497
100 ~ CaChl rnM [ A ~ Nicotine300pM ~ 100 I
~O ~ Anatoxin 30~M 60 I ~ IsoAr Me1100 ~M
{ 40 -- ~ 60
20 "E 40 %
o 2 # 20 Preincubation Time (min)
Fig. 8. Time course for desensitization with carbamylcholine (CaCh), (-)nicotine, (--+)anatoxin and isoarecolone metbiodide (lsoAr MeI). PC12 cells were desensitized by incubation with agonists for 0 to 20 rain, washed and then assayed for =Na § influx during a 2 rain incu- bation at 22~ with 1 mM carbamylcholine. Values, relative to re- sponse after 0 time preincubation, are means -4- SEM (n = 3). o 100
o
'0o r
60 /
OO 6 5 4 3 log [CaCh], M "~
Fig. 9. Effect of d-tubocurarine and the ganglionic blockers chlori- *~ sondamine and mecamylamine on carbamylcholine-elicited desensiti- zation. PC12 cells were desensitized by incubation with various concentrations of carbamylcholine in the absence of any blocker (o) or in the presence of d-tubocurarine (% 1 ~M), chlorisondamine (A, 10 nM) or mecamytamine (o, 0.1 ~M for 20 rain), washed and then assayed for 22Na* influx during a 2 min incubation at 22~ with 2 L , / ~ ~ mM carbamylcholine. Values, relative to response after preincubation for 20 min in the absence of any agent, are means - SEM (n -- 3). Error bars less than the size of the symbol are not shown.
compet i t ive b lockers the apparent lack of comple te re- moval dur ing wash ing (see F igure 10ABC) prevents a
c lear assessment of effects on desensi t iza t ion. Remark- ably, the gangl ion ic b locker ch lo r i sondamine at 10 nM s igni f icant ly enhanced desens i t iza t ion (Figure 9). Chlor - i sondamine apparent ly has not been s tudied for effects on desensi t iza t ion in neuromuscu la r prepara t ions . It was re la t ive ly w e a k as an inhibi tor versus b ind ing of ei ther [125I]~x-BuTx or [3H]H12-HTX to the musc le - type ace- t y l c h o l i n e r e c e p t o r - c h a n n e l c o m p l e x in e l e c t r o p l a x membranes (Table II).
In summary , the present s tudy of the effects of eight
Oo
-log leaCh], M
Fig. 10. Effect of putative noncompetitive blockers on carbamylcho- line-elicited desensitization. PCI2 cells were desensitized by incuba- tion with various concentrations of carbamylcholine in the absence of any blocker (e) and in the presence of the following: A. (-)Isodi- hydrohistrionicotoxin (% 0.3 ~M); (+)gephyrotoxin (A, 1 ixM); (___)indolizidine 223AB (n, 2 I~M), (-)indolizidine 239AB (. , 0.3 ixM). B. (+)Pumiliotoxin C (| 1 t~M); (-)pipefidine 241D ([], 0.3 ~M); phencyclidine (&, 0.3 ixM). C. bupivacaine (~, 1 I~M); piper- ocaine (If, 0.5 I~M). Ceils were washed and then assayed for 2~Na* influx during a 2 min incubation with 2 mM carbamylcholine. Values, relative to response after preincubation for 20 min in the absence of any agent, are means -+ SEM (n = 3). Error bars less than the size of the symbol are not shown. Certain blockers do not appear to be removed completely on washing (see reduced 00 values).
n icot inic agonis ts , one nicot inic antagonis t , three gan- gl ionic blockers and some thirteen noncompeti t ive blockers

498 Daly, Nishizawa, Edwards, Waters, and Aronstam
on 22Na+ flux through ganglionic-type nicotinic receptor channel complexes in PC12 cells reveals basic similar- ities to the effects of the same agents on muscle-type nicotinic receptor-channel in neuromuscular preparations and in binding studies to muscle-type nicotinic receptor- channel in Torpedo electroplax membranes. Some of the differences may reflect differences in assay paradigms rather than differences in receptors. Assay of nicotinic agonist-evoked 22Na+ flux in PC12 cell represents a valuable model system for further study of function and properties of ganglionic-type nicotinic receptors. Parallel studies on nicotonic agonist-evoked 22Na+-flux in my- ocyte-derived nicotinic receptors (see ref. 11,45) should provide for a better delineation of differences between the ganglionic and muscle-type nicotinic receptors and the development of selective agonists, antagonists and noncompetitive blockers. Parallel comparisons for cen- tral neuronal and ganglionic subclasses of the nicotinic receptor-channels in cultured cells should provide for a better delineation of differences in agonist, antagonist and noncompetitive blocker sites at such nicotinic re- ceptors. One such recent study (32) indicates that ago- nists, such as carbamylcholine, isoareeolone, and nicotine, are more potent in the TEGYI cell containing a central- type neuronal receptor than in the PC12 cell containing a ganglionic-type receptor. The antagonist d-tubocurar- ine was equipotent in both cell types. Ganglionic block- ers, such as mecamylamine and hexamethonium, as expected, were more potent in the PC12 cell. Noncom- petitive bIockers were not investigated per se. Atropine, which is known to have activity as a noncompetitive blocker at muscle-type receptors (2) was slightly more potent in the PC12 cells than in the TEGYI cells. The results suggest differences in sites that interact with ag- onists, antagonists and noncompetitive blocker for gan- g l ion ic - type and cen t ra l - type neuronal n ico t in ic acetylcholine receptor-channels, but further more de- tailed comparisons are needed.
R E F E R E N C E S
1. Adam, L. P., and Henderson, E. G. 1990. Calcium channel ef- feetors are potent non-competitive blockers of acetylcholine re- ceptors. Pfltigers Arch. 416:586-593.
2. Adler, M., Albuquerque, E. X., and Lebeda, F. J. 1978. Kinetic analysis of end plate currents altered by atropine and scopolamine. Mol. Pharmacol. 14:514-529.
3. Aguayo, L. G., Witkop, B., and Albuquerque, E. X. 1986. Volt- age- and time-dependent effects of phencyclidines on the endplate current arise from open and dosed channel blockade. Proc. Natl. Acad. Sci. USA 83:3523-3527.
4. Albuquerque, E. X., Daly, J. W., and Warnick, J. E. 1988. Macromolecular sites for specific neurotoxins and drugs on chro- mosensitive synapses and electrical excitation in biological mere-
brane. In "Ion Channels", Narahashi, T. (Ed.). Plenum Publishing Corp., Vol. 1, pp. 95-162.
5. Albuquerque, E. X., Tsai, M.-C., Aronstam, R. S., Eldefrawi, A. T., and Eldefrawi, M. E. 1980. Sites of action of phencycli- dine. II. Interaction with the ionic channel of the nicotinic recep- tor. Mol. Pharmacol. 18:167-178.
6. Aronstam, R. S., Daly, J. W., Spande, T. F., Narayanan, T. K., and Albuquerque, E. X. 1986. Interaction of gephyrotoxin and indolizidine alkaloids with the nicotinic acetylcholine receptor-ion channel complex of Torpedo electroplax. Neurochem. Res. 11:1227-1240.
7. Aronstam, R. S., Edwards, M. W., Daly, J. W., and Albuquer- que, E. X. 1988. Interactions of piperidine derivatives with the nicotinic cholinergic receptor complex from Torpedo electric or- gan. Neurochem. Res. 13:171-176.
8. Aronstam, R. S., Eldefrawi, A. T., Pessah, I.N., Daly, J. W., Albuquerque, E. X., and Eldefrawi, M. E. 1981. Regulation of [3H]perhydrohistrionicotoxin binding to Torpedo ocellata electro- plax by effectors of the acetylcholine receptor. J. Biol. Chem. 256:2843-2850.
9. Aronstam, R. S., King, C. T., Albuquerque, E. X., Daly, J. W., and Feigl, D. M. 1985. Binding of [3H]perhydrohistrionicotoxin and [3H]phencyclidine to the nicotinic receptor ion channel com- plex of Torpedo electroplax: Inhibition of histfionicotoxin and derivatives. Biochem. Pharmacol. 34:3037-3047.
10. Aronstam, R. S., and Witkop, B. 1981. Anatoxin-a interactions with cholinergic synaptic molecules. Proc. Natl. Acad. Sci. USA 78:4639--4643.
11. Burgermeister, W., Catterall, W. A., and Witkop, B. 1977. His- trionicotoxin enhances agonist-induced desensitization of acetyl- choline receptor, Proc. Natl. Acad. Sci. USA 74:5754--5758.
12. Changeux, J.-P., Devillers-Thiery, A., and Chemouilli, P. 1984. Acetylcholine receptor: An allosteric protein. Science 225:1335- 1345.
13. Coffen, D. L., Hengartner, W., Katonah, D. A., Mulligan, M. E., Burdich, D. C., Olson G. L., and Todaro, L. T. 1984. Syn- thesis of an antipsychotic pyrrolo[2,3-g]isoquinoline from Areca alkaloids. J. Org. Chem. 49:5109-5113.
14. Daly, J. W., Myers, C. W., and Whittaker, N. 1987. Further classification of skin alkaloids from neotropical poison frogs (Dendrobatidae), with a general survey of toxic/noxious sub- stances in the amphibia. Toxicon 25:1023-1095.
15. Daly, J. W., and Spande, T. F. 1986. Amphibian alkaloids: Chemistry pharmacology and biology. In "Alkaloids: Chemical and Biological Perspectives", S. W. Pelletier (Ed.), Vol. 4, pp. 1-274.
16. Daly, J. W., Whittaker, N., Spande, T. F., Higher, R. J., Feigl, D., Nishimori, N., Tokuyama, T., and Myers, C. W. 1986. Al- kaloids from dendrobatid frogs: structures of two w-hydroxy con- geners of 3-butyl-5-propylindolizidine and occurrence of 2,5- disubstituted pyrrolidines and a 2,6-disubstituted piperidine. J. Natural Prod. 49:265-280.
17. Daly, J. W., Witkop, B., Tokuyama, T., Nishikawa, T., and Karle, L L. 1977. Gephyrotoxins, histrionicotoxins, and pumili- otoxins from the neotropical frog Dendrobates histrionicus. Helv. Chim. Aeta 60:1128--1140.
18. Edwards, M. W., and Daly, J. W. 1988. Alkaloids from a Pan- amanian poison-frog, Dendrobates speciosus (Dendrobatidae): Identification of pumiliotoxin-A and allopumiliotoxin class alka- loids, 3,5-disubstituted indolizidines, 5-substituted-8-methylin- dolizidines, and a 2-methyl-6-nonyl-4-hydroxypiperidine. J. Nat. Products 51:1188--1197.
19. Danheiser, R. L., Morin, J. M., and Salaski, E. J. 1985. An efficient total synthesis of (• J. Am. Chem. Soe. 107:8066-8073.
20. Eldefrawi, M, E., Eldefrawi, A. T., Aronstam, R. S., Maleque, M. A., Warnick, J. E., and Albuquerque, E. X. 1980. [3H]Phencyclidine-a probe of the ionic channel of the nicotinic receptor. Proc. Natl. Acad. Sci. USA 77:7458-7462.

Nicot in ic A g e n t s in P C 1 2 Cel ls 499
21. Freund, R. K., Jungschaffer, D. A., and Collins, A. C. 1990. Nicotine effects in mouse hippoeampus are blocked by mecamyl- amine, but not other nicotinic antagonists. Brain Res. 511:187- 191.
22. Glavinovic, M., Henry, J. L., Kato, G., Krnjevfc, K., and Puff, E. 1974. Histrionicotoxin: Effects on some central and peripheral excitable cells. Can. J. Physiol. Pharmacol. 52:1220-1225.
23. Heidmann, T., Oswald, R. E., and Changeux, J.-P. 1983. Mul- tiple sites of action for noncompetitive blockers on acetylcholine receptor rich membrane fragments from Torpedo marmorata. Bio- chem.istry 22:3112-3127.
24. Ikeda, S. R., Aronstam, R. S., Daly, J. W., Aracava, Y., and Albuquerque, E. X. 1984. Interactions of bupivacaine with ionic channels of the nicotinic receptor. Electrophysiological and bio- chemical studies. Mol. Pharmacol. 26:293-303.
25. Inoue, K., and Kenimer, J. G. 1988. Muscarinic stimulation of calcium influx and norepinephrine release in PC12 cells. J. Biol. Chem. 263:8157-8161.
26. Karlin, A. 1983. The anatomy of a receptor. Neurosci. Commen- taries 1:111-123.
27. Kemp, G. and Edge, M. 1989. Cholinergic function and ~t-bun- garotoxin binding in PC12 cells. Mol. Pharmacol. 32:356-363.
28. Kemp, G., and Morley, B. J. 1986. Ganglionic nAChRs and high- affinity nicotinic binding sites are not equivalent. FEBS Lett. 205:265-268.
29. Kilpatrick, D. L., Slepetis, R., and Kirshner, N. I981. Inhibition of catecholamine secretion from adrenal medulla cells by neuro- toxins and cholinergic antagonists. J. Neurochem. 37:125-131.
30. LePrince, P. 1983. Chemical modification of the nicotinic cho- linergic receptor of PC-12 nerve cells. Biochemistry 22:5551- 5556.
31. Lowry, O. H., Rosenbrough, N. J., Farr, A. L., and Randall, R. J. 1951. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193:265-275.
32. Lukas, R. J. 1989. Pharmacological distinctions between func- tional nicotinic acetylcholine receptors on the PC12 rat pheo- chromocytoma and the TE671 human neuroblastoma. J. Pharmacol. Exp. Therap. 251:175-182.
33. McKay, D. B., and Trent-Sanchez, P. 1990. Effect of noncom- petitive nicotinic receptor blockers on catecholamine release from cultured adrenal chromaffin cells. Pharmacology 40:224-230.
34. Meldolesi, J., Gatti, G., Ambrosini, A., Pozzan, T., and West- head, E. W. 1988. Second-messenger control of catecholamine release from PC12 cells. Role of muscarinic receptors and nerve- growth-factor-induced cell differentiation. Biochem. J. 255:761- 768.
35. Mitsuka, M., and Hatanaka, H. 1983. Selective loss of acetyl- choline sensitivity in a nerve cell line cultured in hormone-sup- plemented serum-free medium. J. Neurochem. 3:1785-1790.
36. Nishizawa, Y., Seamon, K. B., Daly, J. W., and Aronstam, R. S. 1990. Effects of forskolin and analogues on nicotinic receptor- mediated sodium flux, voltage-dependent calcium efflux and volt- age-dependent rubidium flux in pheochromocytoma PC12 ceils. Cell. Mol. Neurobiol. 10:351-368.
37. Ogura, T., and Warashina, A. 1987. Effects of histrionicotoxin derivatives on ion channels and acetylcholine receptor-channel complexes in bull frog sympathetic ganglia. Comp. Biochem. Biophysiol. 886:249-254.
38. Patrick, J., and Stallcup, B. 1977. a-Bungarotoxin binding and cholinergic receptor function on a rat sympathetic nerve line. J. Biol. Chem. 252:8629-8633.
39. Rabe, C. S., Schneider, J., and McGee Jr., R. 1983. Regulation of depolarization-dependent release of neurotransmitters by aden- osines: Cyclic AMP-dependent enhancement of release from PC12 cells. J. Neurochem. 41:1623-1634.
40. Rapier, C., Wonnacutt, S., Lunt, G. G., and Albuquerque, E. X. 1987. The neurotoxin histrionicotoxin interacts with the putative ion channel of the nicotinic acetylcholine receptors in the central nervous system. FEBS Lett. 212:292-296.
4i. Reavill, C., Spivak, C. E., Stolerman, I. P., and Waters, J. A. 1987. Isoarecolone can inhibit nicotine binding and produce ni- cotine-like discriminative stimulus effects in rats. Neuropharma- cology 26:789-792.
42. Robinson, D., and McGee, R. 1985. Agonist-induced regulation of the neuronal nicotinic acetylcholine receptor of PC-12 cells. MN. Pharmacol. 26:409-417.
43. Simasko, S. M., Soares, J. R., and Weiland, G. A. 1986. Two components of carbamylcholine-induced loss of nicotinic acetyl- choline receptor function in the neuronal line PC12. Mol. Phar- macol. 30:6-12.
44. Simasko, S. M., Soares, J. R., and Weiland, G. A. 1985. Struc- ture-activity relationship for substance P inhibition of carbamyl- choline-stimulated ~Na § flux in neuronal (PC12) and non-neuronal (BC3H1) cell lines. J. Pharmacol. Exp. Therap. 235:601-605.
45. Sine, S. M., and Taylor, P. 1982. Local anesthetics and histrio- nicotoxin are allosteric inhibitors of the acetylcholine receptor. J. Biol. Chem. 257:8106-8114.
46. Souccar, C., Varanda, W. A., Aronstam, R. S., Daly, J. W., and Albuquerque, E. X. 1984. Interactions of gephyrotoxin with the acetylcholine receptor-ionic channel complex. II. Enhance- ment of desensitization. Mol. Pharmacol. 25:395-400.
47. Spivak, C. E., Maleque, M. A., Oliveira, A. C., Masukawa, L., Tokuyama, T., Daly, J. W., and Albuquerque, E. X. 1981. Ac- tions of the histrionicotoxins at the ion channel of the nicotinic acetylcholine receptor and at the voltage-sensitive ion channels of muscle membranes. Mol. Pharmacol. 21:351-361.
48. Spivak, C. E., Waters, J. A., and Aronstam, R. S. 1989. Binding of semirigid, nicotinic agonists to nicotinic and muscarinic recep- tors. Mol. Pharmacol. 36:177-184.
49. StaUcup, W. A. 1979. Sodium and calcium fluxes in a clonal nerve cell line. J. Physiol. 286:525-540.
50. Stallcup, W. B., and Patrick, J. 1980. Substance P enhances cholinergic receptor desensitization in a clonal nerve cell line. Proc. Natl. Acad. Sci. USA 77:634--638.
51. Swanson, K. L., Allen, C. N., Aronstam, R. S., Rapoport, H., and Albuquerque, E. X. 1986. Molecular mechanisms of the po- tent and stereospecific nicotinic receptor agonist (+)-anatoxin-a, Mol. Pharmacol. 29:250-257.
52. Tiedt, T. N., Albuquerque, E. X., Bakry, N. M., Eldefrawi, M. E., and Eldefrawi, A. T. 1979. Voltage- and time-dependent ac- tions of pipemcaine on the ion channel of the acetylcholine re- ceptor, Mol. Pharmacol. 16:909-921.
53. Tokuyama, T., Nishimori, N., Shimada, A., Edwards, M. V., and Daly, J. W. 1987. New classes of amidine, indolizidine, and quinolizidine alkaloids from a poison-frog, Dendrobates pumilio (Dendrobatidae). Tetrahedmn 43:643-652.
54. Tsai, M.-C., Mansour, N. A., Eldefrawi, A. T., Eldefrawi, M. E., and Albuquerque, E. X. 1978. Mechanism of action of aman- tadine on neuromuscular transmission. Mol. Pharmacol. 14:787- 803.
55. Varanda, W. A., Aracava, Y., Sherby, S. M., Van Meter, W. G., Eldefrawi, M. E., and Albuquerque, E. X. 1985. The ace- tylcholine receptor of the neuromuscular junction recognizes me- camylamine as a noncompetitive antagonist. Mol. Pharmacol. 28:128-137.
56. Wada, A., Arita, M., Yangihara, N., and Izumi, F. 1988. Binding of [3H]phencyclidine to adrenal medullary cells: Inhibition of 22Na influx, 4~Ca influx, S'~Rb efflux and catecholamine secretion caused by carbachol and veratridine. Neuroscience 25:687-696.
57. Wada, A., Uezono, Y., Arita, M., Tsuji, K., Yanagihara, N., Kobayashi, H., and Izumi, F. 1989. High-affinity and selectivity of neosurfigatoxin for the inhibition of Z2Na influx via nicotinic receptor-ion channel in cultured bovine adrenal medullary cells: Comparative study with histrionicotoxin. Neuroscience 33:333- 339.
58. Warnick, J. E., Jessup, P. J., Overman, L. E., Eldefrawi, M. E., Nimit, Y., Daly, J. W., and Albuquerque, E. X. 1982. Pum-

500 Daly, Nishizawa, Edwards, Waters, and Aronstam
iliotoxin-C and synthetic analogs. A new class of nicotinic antag- onists. Mol. Pharmacol. 22:565-573.
59. Warnick, J. E., Maleque, M. A., Bakry, N., Eldefrawi, A. T., and Albuquerque, E. X. 1982. Structure-activity relationships of amantadine. I. Interaction of the N-alkyl analogues with the ionic channels of the nicotinic acetylcholine receptor and electrically excitable membrane. Mol. Pharmacol. 22:82-93.
60. Waters, J. A., Spivak, C. E., Hermsmeier, M., Yadov, J. S., Liang, R. F., and Gund, T. M. 1988. Synthesis, pharmacology,
and molecular modeling studies of semirigid, nicotinic agonists. J. Med. Chem. 31:545-554.
61. Whiting, P., and Lindstrom, J. 1987. Purification and character- ization of a nicotinic acetylcholine receptor from rat brain. Proc. Natl. Acad. Sci. USA 84:595-599.
62. Yeh, J. Z., Narahashi, J., and Almon, R. R. 1975. Characteri- zation of neuromuscular blocking action of piperidine derivatives. J. Pharmacol. Exp. Therap. 194:373-383.
Copyright © 2022 FDOKUMEN




















