
Meta-analysis of Trials Comparing beta-Blockers, Calcium Antagonists, and Nitrates for Stable Angina

The e¡ectofquorum-sensing blockers on the formationofmarinemicrobial communities and larval attachmentSergey Dobretsov1,2, Hans-Uwe Dahms1, Huang YiLi1, Martin Wahl2 & Pei-Yuan Qian1
1Department of Biology, Hong Kong University of Science and Technology, Clear Water Bay, Kowloon, Hong Kong SAR, PR China; and2Benthic Ecology, Leibniz-Institut fur Meereswissenschaften, IFM-GEOMAR, Kiel University, Kiel, Germany
Correspondence: Sergey Dobretsov,
Benthic Ecology, Leibniz-Institut fur
Meereswissenschaften, IFM-GEOMAR,
Kiel University, Dusternbrooker Weg 20,
24105 Kiel, Germany. Tel.: 149 431 600
4527; fax: 149 431 600 1671;
e-mail: [email protected]
Received 15 June 2006; revised 13 October
2006; accepted 17 November 2006.
First published online 16 March 2007.
DOI:10.1111/j.1574-6941.2007.00285.x
Editor: Riks Laanbroek
Keywords
microbial communities; quorum-sensing
inhibitors; larval attachment; furanones;
antifouling.
Abstract
We studied the effect of the quorum-sensing (QS) blockers 5-hydroxy-3[(1R)-1-
hydroxypropyl]-4-methylfuran-2(5H)-one (FUR1), (5R)-3,4-dihydroxy-5-[(1S)-
1,2-dihydroxyethyl]furan-2(5H)-one (FUR2) and triclosan (TRI) on the forma-
tion of bacterial biofilms, and the effect of these biofilms on the larval attachment
of the polychaete Hydroides elegans and the bryozoan Bugula neritina. 14-day-old
subtidal biofilms were harvested from artificial substrata and were allowed to
develop in the laboratory with and without QS blockers. QS blockers inhibited the
production of violacein by the QS reporter strain Chromobacterium violaceum
CV026 and did not affect the metabolic activity of bacteria in multispecies
biofilms. At a concentration of 10�3 M all three tested compounds inhibited the
establishment of microbial communities, but at one of 10�4 M only FUR2
inhibited establishment. The tested QS blockers caused changes in bacterial density
and bacterial community structure, as revealed by terminal restriction fragment
length polymorphism and FISH. The groups most affected by QS blockers were
Alphaproteobacteria, Gammaproteobacteria and the Cytophagales. Larvae of H.
elegans and B. neritina avoided settling on biofilms that had developed in the
presence of QS blockers. Our results suggest that QS blockers directly control the
formation of multi-species biofilms, and indirectly – by means of biofilm proper-
ties � affect larval attachment on these modified biofilms.
Introduction
Natural and artificial substrata in the marine environment
are quickly colonized by marine micro- and macro-organ-
isms in a process known as ‘biofouling’. Biofouling causes
serious problems for marine industries and navies around
the world (Yebra et al., 2004). At present, the most effective
methods of biofouling control are based on the application
of substances such as tributyltin (TBT), copper or organic
compounds (e.g. Sea-Nine, Isothiazolone) (Thomas, 2001;
Yebra et al., 2004). All these chemicals are toxic and pollute
the aquatic environment (see the reviews of Evans, 1999;
Thomas, 2001). Accordingly, there is a need for the devel-
opment of ‘environmentally friendly’ nontoxic antifoulants.
Biofilms can enhance (Kirchman et al., 1982; Lau & Qian,
1997, 2001; Qian et al., 2003) or inhibit the larval settlement
of marine invertebrates and the attachment of algal spores
(Rodriguez et al., 1993; Egan et al., 2000, 2001; Dobretsov &
Qian, 2002; Huang & Hadfield, 2003). Moreover, chemical
compounds produced by bacteria can lead to the disruption
of biofilm formation and the prevention of biofouling
(Dobretsov et al., 2006). Hence, bacterial compounds may
be useful in the biotechnological development of an envir-
onmentally friendly protection against marine biofouling
(Clare et al., 1992; Holmstr�m & Kjelleberg, 1999; Arm-
strong et al., 2000).
Although bacteria are unicellular organisms, they can
control their growth and population densities (Xavier &
Bassler, 2003; March & Bentley, 2004). In order to achieve
this, bacteria have evolved a regulatory mechanism named
quorum sensing (QS) that consists of exuded info-chemicals
that activate or de-activate target bacterial genes responsible
for cell division and adhesion and, thus, control biofilm
formation and composition (Davies et al., 1998). Biofilm
formation, in turn, can control many processes at surfaces,
for example the uptake or release of compounds by host
organisms, bacterial virulence for the host, and biocorrosion
(Parsek & Greenberg, 2000; Sauer et al., 2002). The process of
QS is based on the production and release of low-molecular-
weight signal molecules (often called autoinducers). The
FEMS Microbiol Ecol 60 (2007) 177–188 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved

extracellular concentration of QS molecules reflects the
population density of the producing organism. Bacteria can
perceive and react to such signal molecules, allowing the
whole cell population to initiate a concerted action once a
critical population density has been reached (de Nys et al.,
2000).
The intercellular signalling compounds that have received
the most attention so far are the N-acetyl homoserine
lactones (AHLs) that are restricted to Gram-negative bacter-
ia (Whitehead et al., 2001). The effects of AHLs are based on
gene products of the luxR gene analogues (Gray & Garey,
1999). Each Bacterial species may produce different AHL
analogues that differ in the length of the N-acetyl chains,
ranging from 4 to 14 carbons, and in the substitution at the
3-position of the side chain (McLean et al., 1997).
A second communication molecule, a furanosyl borate
diester found in Vibrio harveyi, has been suggested to
be active in interspecific signalling (Chen et al., 2002).
A genomic database analysis has indicated that such inter-
species communication possibly occurs throughout the
Eubacteria (Sperandio et al., 2001). The QS-producing
bacteria are highly diverse and fall within a large number
of species among Alpha-, Beta- and Gammaproteobacteria
(Manefield & Turner, 2002) which are dominant in tropical
waters (Webster et al., 2004; Huang et al., in press).
In contrast to Gram-negative bacteria, Gram-positive bac-
teria exude peptides as signal molecules (Miller & Bassler,
2001). QS signals produced by bacteria may also show
transphyletic effects and induce algal spore attachment
(Joint et al., 2002).
Because QS signals are important for bacterial growth,
adhesion and biofilm formation, the inhibition of QS signals
leads to the disruption of biofilm formation (Ren et al.,
2001a) and the inhibition of bacterial pathogenicity (Ren
et al., 2001b). Many marine organisms, such as the red alga
Delisea pulchra and the bacterium Aeromonas veronii, use
QS blockers to control epibiotic biofilm formation (McLean
et al., 1997, 2004; Manefield et al., 1999; Rice et al., 1999;
Bauer & Robinson, 2002; Zhang & Dong, 2004). Delisea
pulchra produces furanones that interfere with bacterial
AHLs and inhibit the growth of Gram-negative bacteria as
well as the settlement of invertebrate larvae (Steinberg et al.,
1997; Rasmussen et al., 2000; Kjelleberg et al., 2001). At the
same time, it is possible to propose that QS blockers can
control larval settlement indirectly by regulating the micro-
bial community structure of biofilms and the density of
bacteria, which in turn affects larval behaviour.
The aims of this study were to investigate (1) the effect on
the community structure and density of bacteria in multi-
species microbial communities of QS blockers at various
concentrations, and (2) the effect on larval attachment of
microbial communities that were treated or untreated by QS
blockers.
Materials and methods
Inhibition of QS by chemical compounds
The bacterium Chromobacterium violaceum CV026 (a mini-
Tn5 mutant) was used as an indicator organism for QS by
quantifying violacein synthesis. Prior to the bioassay, the
strain was grown overnight on 0.5% yeast extract and 1%
tryptone in filtered (0.22-mm) seawater. The inhibitive
effects of QS blockers on the production of violacein by
C. violaceum cultures were tested according to Martinelli
et al. (2004). Briefly, the experiments were conducted in 96-
well microplates that contained N-hexanoylhomoserine
lactone (HHL) at 3.7� 10�8 M, 100 mL of C. violaceum, and
QS blockers. Tested concentrations of QS blockers were
10�3–10�5 M. After exposure for 16 h at 27 1C, the plates
were dried and the absorbance of each cell well was
measured with an automatic plate reader at 590 nm. Cells
without QS blockers but with HHL and without HHL were
used as the positive and negative controls. In addition, the
effect of QS blockers on bacterial growth was investigated by
a comparison of bacterial culture turbidity at 660 nm.
Microbial film development
Biofilms were developed on polystyrene Petri dishes (Falcon
no. 1006) exposed to biofouling in the subtidal zone
(maximal depth = 3 m at high tide) during February and
March 2005. Additional experiments with a QS reporter
strain were carried out in September 2006. All dishes were
attached vertically to nylon ropes, and the upper end of each
rope was tied to the pier. A weight was attached to the lower
end of the rope to keep the rope straight. After 14 days, the
dishes were retrieved and microbial films were washed from
the dishes by a sterile brush into a sterile Petri dish filled
with filtered (0.22-mm) seawater (FSW). The biofilm sus-
pension obtained was filtered through a 45-mm mesh and
subsequently through ash filters (Whatman, N1) in order to
eliminate most of the diatoms. This microbial suspension
was used in further experiments (see below).
Biofilm treatment
The QS blockers 5-hydroxy-3[(1R)-1-hydroxypropyl]-4-
methylfuran-2(5H)-one (FUR1), (5R)-3,4-dihydroxy-5-
[(1S)-1,2-dihydroxyethyl]furan-2(5H)-one (FUR2, Sigma)
and triclosan (TRI, Sigma) were dissolved in FSW at
concentrations of 10�2, 10�3 and 10�4 M. Experiments were
performed in large Petri dishes (diameter 90 mm). Before
the commencement of each experiment, sterile polystyrene
slides (washed in 70% ethanol and rinsed three times in
sterile distilled water) cut from large Petri dishes were placed
on the bottom of each Petri dish. Thereafter, 9 mL of
microbial suspension and 1 mL of one type of the QS
FEMS Microbiol Ecol 60 (2007) 177–188c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
178 S. Dobretsov et al.

blockers were added in order to obtain a final concentration
of 10�3, 10�4 and 10�5 M. Each concentration was replicated
48-fold. Petri dishes without any chemicals added were used
as the controls. Biofilms were developed under laboratory
conditions (20 1C, no light) within 48 h according to Railkin
(1998). After the experiment, solutions were discarded and
the slides were rinsed several times with FSW. For each
concentration and compound, 10 replicates were used for
larval bioassay experiments, three replicates were used for
terminal restriction fragment length polymorphism (T-
RFLP) analysis, and the remaining 35 replicates were used
for bacterial counting and FISH, as well as for quantification
of respiratory active (alive) and nonactive (dead) cells. The
numbers of metabolically active cells in biofilms treated with
QS blockers and untreated (control) cells in biofilms were
determined by staining with 6 mM 5-cyano-2, 3-ditolyl
tetrazoliumchloride (CTC, Polysciences) in FSW according
to Haglund et al. (2002).
Larval cultivation and experiments with larvae
Adult broodstocks of the polychaete Hydroides elegans
(Haswell) and the bryozoan Bugula neritina (Linnaeus) were
collected from pilings and floating rafts in Port Shelter Bay,
Hong Kong (221190N, 1141160E). Batch cultures of poly-
chaete larvae were reared for 5 days to competent stage
according to Harder & Qian (1999). Bryozoan larvae were
obtained as described by Bryan et al. (1997); newly hatched
larvae were included in the bioassays. Still-water laboratory
bioassays were performed with five replicates in sterile
polystyrene Petri dishes (no. 1006, Falcon) that contained
20 larvae and the slide with biofilm developed under various
QS treatments in 4 mL of sterile-filtered (0.22-mm) seawater
(FSW). Prior to the larval bioassays, the biofilms were
washed with several portions of sterile FSW in order to
remove remaining QS blockers. Larval attachment assays
were run at 28 1C under continuous illumination for 1 h.
After this time, attached juveniles were counted and attach-
ment was expressed as percentages.
Biofilm analysis
Terminal restriction length polymorphism
The bacterial community compositions of biofilms on
polystyrene slides were compared using T-RFLP analysis
(Liu et al., 1997). For this purpose, the entire surface area of
each slide (120 mm2) was swabbed with sterile cotton balls.
Three swabs from each treatment were individually sus-
pended in 1 mL of extraction buffer (100 mM Tris-HCl,
100 mM EDTA, 100 mM sodium phosphate, 1.5 M sodium
chloride, 1% CTAB; at pH = 8) in 2-mL microcentrifuge
tubes. For lysing, the samples were subjected to three
cycles of freezing and thawing followed by incubation
for 2 h in 20% sodium dodecylsulfate (SDS) at 65 1C.
The cotton swabs were removed, and, after centrifugation
(8832 g� 5 min�1), the total DNA in the supernatant was
extracted and purified twice in a volume of 24 : 1 chlorofor-
m:isoamyl-alcohol, followed by precipitation in isopropanol
at room temperature for 15 min. The precipitated DNA was
washed with cold 70% ethanol and resuspended in 50 mL of
autoclaved double-distilled water and then frozen at
� 80 1C until use.
PCR of 16S rRNA genes of bacterial communities was
performed in a total volume of 25mL containing 1 mL of
DNA template, 250mM of each deoxyribonucleotide tripho-
sphate (dATP, dCTP, dGTP, dTTP; Pharmacia Biotechnol-
ogy), 1 U of DNA Taq polymerase (Amersham Biosciences)
and 0.8 mM of each universal primer: 341F forward (50-
CCTACGGGAGGCAGCAG-30) and 926R reversal (50-
CCGTCAATTCCTTTRAGTTT-30). The 926R primer was
labelled at the 50 end with 6-carboxy fluorescein (FAM) dye.
The thermocycling conditions were as follows: 95 1C for
2 min (1 cycle); 15 cycles of 95 1C for 30 s, 60 1C for 3 min,
and 72 1C for 3 min (the annealing temperature started at
60 1C and was reduced to 45 1C in increments of 1 1C
cycle�1); 10 cycles of 95 1C for 30 s, 45 1C for 3 min, and
72 1C for 3 min; and 72 1C for 10 min. Amplified DNA (4 mL
of PCR mixtures) was visualized by gel electrophoresis on a
1.5% agarose gel in Tris-acetate-EDTA (TAE) buffer.
Fluorescently labelled PCR products from triplicate PCR
amplifications were combined and purified with the
Wizards PCR preps DNA purification system (Promega,
WI) according to the manufacturer’s protocol. Purified
amplicons were digested with 20 U MspI (Boehringer Man-
nheim Biochemicals, IN) at 37 1C for 6 h. Aliquots of
digested products (10 mL) were mixed with 0.5mL of internal
size standard (ET550-R, Amersham Biosciences, UK). This
mixture was denatured for 2 min at 95 1C and immediately
chilled on ice prior to capillary electrophoresis on a Mega-
BACETM genetic analyser (Amersham Biosciences) operated
in the genotyping mode. After electrophoresis, the lengths of
the fluorescently labelled terminal restriction fragments
(T-RFs) were determined by comparison with internal size
standards using the FRAGMENT PROFILER software (Amersham
Biosciences, UK). The lengths of T-RFs obtained by the
analyser were rounded up to the nearest integral values.
Peaks that were o 1.5 bp from a larger peak were classified
as its ‘shoulders’ and eliminated (Dunbar et al., 2001).
For each sample, peaks over a threshold of 50 fluorescence
units (Blackwood et al., 2003) and whose peak heights
contributed for at least 1% to the integrated height (Buchan
et al., 2003) were used for analysis. Terminal fragments
o 35 and 4 500 bp were excluded from the analysis in
order to avoid detection of primers and uncertainties of size
determination.
FEMS Microbiol Ecol 60 (2007) 177–188 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
179Antifouling activity of QS blockers

FISH
Biofilm slides were fixed in 4% paraformaldehyde sus-
pended in phosphate-buffered saline for 8 h at 4 1C and
transferred to ethanol–phosphate-buffered saline (1 : 1) at
20 1C until further processing. In order to perform FISH,
slides were dehydrated with 96% ethanol and air-dried. For
each treatment we had five replicated slides. For each slide,
only one fluorescent probe (Table 1) labelled with Cy3 was
used. Hybridization solution was mixed in a ratio of 1 : 1
with the appropriate fluorescently labelled oligonucleotide
and applied to each slide. Slides were incubated in 50-mL
Falcons tubes at 46 1C in the dark for 1.5 h. After hybridiza-
tion, slides were carefully removed and rinsed immediately
in warmed wash buffer (46 1C). Slides were rinsed in fresh
water to remove excess salt, air-dried, mounted, and stained
with the DNA-binding fluorochrome 4,6-diamidino-2-phe-
nylindole (DAPI, Vectashield, Vector Lab; concentration
0.5mg mL�1) for 15 min. The total number of bacteria and
the number of particular groups of bacteria in five randomly
selected fields of view were estimated using an epifluores-
cence microscope (Axiophot, Zeiss, Germany; magnifica-
tion � 1000 with excitation peaks at 359 and 647 nm).
Statistical analysis
The normality assumption was verified with Shapiro–Wilk’s
test (Shapiro & Wilk, 1965), and homogeneity of the data
with Barlett’s or Levene’s test (Zar, 1999). In order to
improve the normality of the data they were arcsine-
transformed. In the case of zero attached larvae, a value of
4n�1 (where n is the number of larvae in a single replicate)
was assigned to improve the arcsine transformation (Zar,
1999). The effect of concentration and chemical treatment
on larval attachment was determined by two-way ANOVA. The
differences between the treatments and the control were
determined by one-way ANOVA followed by a Dunnet posthoc
test (Quinn & Keough 2002). In all cases, the threshold for
significance was 5%. The data presented in all figures were
not transformed.
The T-RFLP patterns of different bacterial community
DNA samples were used for the construction of the Bray–
Curtis similarity matrix based on the total number of T-RFs
observed in all samples compared with the presence or
absence of these T-RFs in individual samples. The construc-
tion of a multi-dimensional scaling (MDS) plot to demar-
cate the similarity of microbial communities was performed
using the PRIMER program (Plymouth Marine Laboratory,
UK). In all cases, the threshold level for significance was 5%.
Results
Inhibition of QS by chemical compounds
All tested compounds significantly inhibited the production
of violacein by C. violaceum in the presence of HHL (Fig. 1;
ANOVA: F = 169.2, Po 0.0001). The most effective substances
were FUR1 and FUR2, which significantly (ANOVA, Dunnet
test, Po 0.05) inhibited bacterial QS at concentrations of
10�3–10�5 M. Violacein formation was reduced by up to
90% in comparison with the control, while there was no
inhibition of bacterial growth. The results of this experiment
suggest that the tested compounds do indeed interfere with
the signalling system of QS.
The effect of QS blockers on the development ofbiofilms
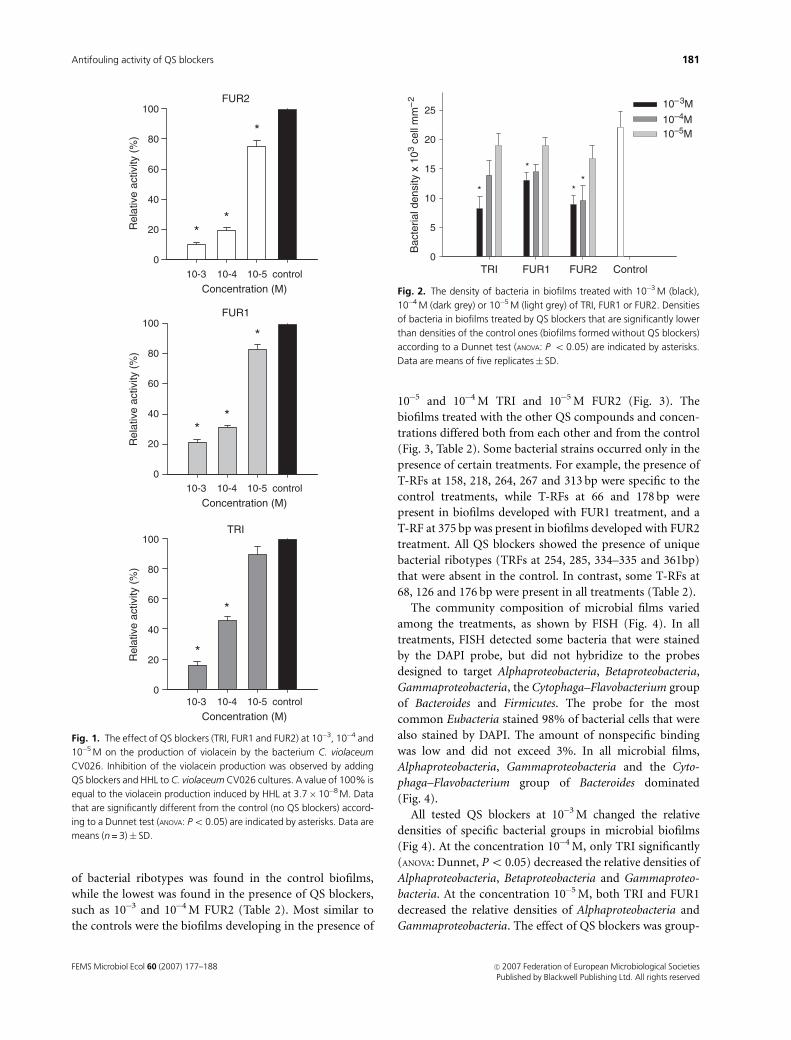
The densities of bacteria in biofilms were affected by
chemical compounds (Fig. 2). TRI and FUR1 were effective
at the highest concentration (10�3 M), while FUR2 signifi-
cantly (Po 0.05) decreased bacterial densities relative to the
control at all concentrations equal or above 10�4 M. None of
the compounds was active at 10�5 M. The percentage of
metabolically active cells (81� 3%) in biofilms treated by
chemical compounds was similar to that in the nontreated
biofilms (79� 5%), suggesting that QS blockers did not
have any toxic effects on bacteria from biofilms.
T-RFLP analysis (Table 2) revealed that the tested com-
pounds changed bacterial community profiles at some
concentrations (Fig. 3). Bacterial communities formed in
the presence of QS blockers were characterized by the
absence of certain terminal restriction fragments (T-RFs or
ribotypes) and the presence of others. The highest number
Table 1. Oligonucleotide probes used for FISH analysis
Probe Sequence Specificity
Percentage
formamide Reference
EUB338 GCTGCCTCCCGTAGGAGT Most common bacteria 20 Amann et al. (1990)
ALF968 GGTAAGGTTCTGCGCGTT Alphaproteobacteria 20 Manz et al. (1992)
BET42a GCCTTCCCACTTCGTTT Betaproteobacteria 35 Manz et al. (1992)
GAM42a GCCTTCCCACATCGTTT Gammaproteobacteria 35 Manz et al. (1992)
CF319a TGGTCCGTGTCTCAGTAC Cytophaga–Flavobacteria 35 Manz et al. (1992)
LGC354a YSGAAGATTCCCTACTGC Some Firmicutes spp. 35 Meier et al. (1999)
NonEUB338 ACTCCTACGGGAGGCAGC Negative control probe 20 Amann et al. (1990)
FEMS Microbiol Ecol 60 (2007) 177–188c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
180 S. Dobretsov et al.
of bacterial ribotypes was found in the control biofilms,
while the lowest was found in the presence of QS blockers,
such as 10�3 and 10�4 M FUR2 (Table 2). Most similar to
the controls were the biofilms developing in the presence of
10�5 and 10�4 M TRI and 10�5 M FUR2 (Fig. 3). The
biofilms treated with the other QS compounds and concen-
trations differed both from each other and from the control
(Fig. 3, Table 2). Some bacterial strains occurred only in the
presence of certain treatments. For example, the presence of
T-RFs at 158, 218, 264, 267 and 313 bp were specific to the
control treatments, while T-RFs at 66 and 178 bp were
present in biofilms developed with FUR1 treatment, and a
T-RF at 375 bp was present in biofilms developed with FUR2
treatment. All QS blockers showed the presence of unique
bacterial ribotypes (TRFs at 254, 285, 334–335 and 361bp)
that were absent in the control. In contrast, some T-RFs at
68, 126 and 176 bp were present in all treatments (Table 2).
The community composition of microbial films varied
among the treatments, as shown by FISH (Fig. 4). In all
treatments, FISH detected some bacteria that were stained
by the DAPI probe, but did not hybridize to the probes
designed to target Alphaproteobacteria, Betaproteobacteria,
Gammaproteobacteria, the Cytophaga–Flavobacterium group
of Bacteroides and Firmicutes. The probe for the most
common Eubacteria stained 98% of bacterial cells that were
also stained by DAPI. The amount of nonspecific binding
was low and did not exceed 3%. In all microbial films,
Alphaproteobacteria, Gammaproteobacteria and the Cyto-
phaga–Flavobacterium group of Bacteroides dominated
(Fig. 4).
All tested QS blockers at 10�3 M changed the relative
densities of specific bacterial groups in microbial biofilms
(Fig 4). At the concentration 10�4 M, only TRI significantly
(ANOVA: Dunnet, Po 0.05) decreased the relative densities of
Alphaproteobacteria, Betaproteobacteria and Gammaproteo-
bacteria. At the concentration 10�5 M, both TRI and FUR1
decreased the relative densities of Alphaproteobacteria and
Gammaproteobacteria. The effect of QS blockers was group-
FUR1
Rel
ativ
e ac
tivity
(%
)R
elat
ive
activ
ity (
%)
0
20
40
60
80
100
FUR2
Concentration (M)10-3 10-4 10-5 control
Concentration (M)10-3 10-4 10-5 control
Concentration (M)10-3 10-4 10-5 control
Rel
ativ
e ac
tivity
(%
)
0
20
40
60
80
100
TRI
0
20
40
60
80
100
**
*
**
*
*
*
Fig. 1. The effect of QS blockers (TRI, FUR1 and FUR2) at 10�3, 10�4 and
10�5 M on the production of violacein by the bacterium C. violaceum
CV026. Inhibition of the violacein production was observed by adding
QS blockers and HHL to C. violaceum CV026 cultures. A value of 100% is
equal to the violacein production induced by HHL at 3.7� 10�8 M. Data
that are significantly different from the control (no QS blockers) accord-
ing to a Dunnet test (ANOVA: Po 0.05) are indicated by asterisks. Data are
means (n = 3)� SD.
TRI FUR1 FUR2 Control
Bac
teria
l den
sity
x 1
03 ce
ll m
m–
2
0
5
10
15
20
2510–3M
10–4M10–5M
*
*
**
Fig. 2. The density of bacteria in biofilms treated with 10�3 M (black),
10�4 M (dark grey) or 10�5 M (light grey) of TRI, FUR1 or FUR2. Densities
of bacteria in biofilms treated by QS blockers that are significantly lower
than densities of the control ones (biofilms formed without QS blockers)
according to a Dunnet test (ANOVA: P o 0.05) are indicated by asterisks.
Data are means of five replicates� SD.
FEMS Microbiol Ecol 60 (2007) 177–188 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
181Antifouling activity of QS blockers

Table 2. T-RFLP profiles of bacterial communities formed in the pre-
sence of 10�3 M, 10�4 M and 10�5 M of TRI, FUR1 and FUR2 or without
quorum-sensing blockers (CON)
TRFs
(bp)
TRI FUR1 FUR2
CON10�3 10�4 10�5 10�3 10�4 10�5 10�3 10�4 10�5
57 Xa
59 Xb
62 Xb Xa Xb
63 Xa Xa Xb
64 Xa Xb
66 Xa Xa Xa Xa
68 Xa Xa Xa Xa Xa Xb Xa Xa Xa Xa
72 Xb
75 Xb
77 Xb Xa Xa Xb Xb Xa
79 Xa Xa Xa Xa Xa Xa
80 Xa Xa Xa Xa Xa Xa
88 Xb Xa
89 Xa Xa Xa Xa Xa Xa
90 Xa Xa Xa Xa Xa Xb Xa Xa Xa
91 Xa Xa Xa Xa Xa Xa Xb Xa Xa
92 Xa Xb
93 Xa
102 Xa Xa Xa Xb Xa Xa Xa
103 Xb
116 Xb
123 Xb Xa Xa Xa Xb
124 Xa Xa Xa Xa Xa Xa Xa Xa
125 Xa
126 Xa Xa Xa Xa Xa Xa Xa Xa Xa Xa
127 Xb Xa Xa Xb
128 Xa Xa Xb
129 Xa Xa Xb Xb Xa
130 Xb Xb Xa Xb
131 Xa Xb
142 Xb
145 Xa Xa Xa Xa
149 Xa
150 Xa
153 Xa Xa Xa Xa Xb Xa Xa Xa
154 Xa
158 Xa
163 Xa
172 Xb Xb
173 Xa
175 Xa Xa
176 Xa Xa Xa Xa Xa Xa Xb Xb Xb Xa
177 Xa Xa Xb
178 Xb Xb Xa
179 Xb
181 Xa
182 Xb
183 Xb
189 Xb
193 Xb Xb
194 Xa Xb Xa Xa
195 Xa
203 Xa
204 Xa Xa
Table 2. Continued.
TRFs
(bp)
TRI FUR1 FUR2
CON10�3 10�4 10�5 10�3 10�4 10�5 10�3 10�4 10�5
206 Xa Xa Xa Xa Xa Xa Xa Xa
207 Xa Xa Xb Xa
208 Xb Xa Xa
209 Xa
218 Xa
219 Xa Xa
220 Xb
246 Xa Xa Xa
247 Xa Xa Xa
248 Xa
253 Xb Xb Xb Xb
254 Xa Xb Xa
264 Xa
267 Xb
279 Xa Xb Xb
286 Xb
292 Xa
293 Xa Xb Xb Xb
294 Xb Xa
295 Xa Xa Xa Xb
298 Xa Xb
299 Xa Xa Xb Xb
300 Xa Xa Xb Xa Xa
302 Xa Xa
303 Xb Xa Xa Xa Xa Xb Xa Xa
310 Xa Xa Xa Xb Xa Xa
313 Xb
314 Xa Xb Xa Xa
315 Xa
329 Xa
333 Xa Xa Xa Xa
334 Xa Xa Xa Xa
335 Xa Xb Xa Xa
336 Xb Xb Xa
338 Xb Xa Xa Xb Xb
359 Xa
361 Xb Xa Xa Xa Xa
363 Xa
365 Xb Xb Xa
367 Xa Xa Xa
368 Xa Xa
370 Xa Xa Xa Xa Xa
371 Xa
372 Xa
374 Xa Xa Xa Xa Xa
375 Xa Xa Xa
397 Xa
419 Xa
420 Xa Xa Xa Xa
421 Xa Xa Xb Xa
422 Xa Xa
432 Xb
433 Xa
435 Xa Xb
436 Xb
441 Xa
FEMS Microbiol Ecol 60 (2007) 177–188c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
182 S. Dobretsov et al.

specific (Fig. 4). For example, at the concentration 10�3 M,
TRI significantly (ANOVA: Dunnet, Po 0.05) decreased the
relative density of Betaproteobacteria, while FUR1 at this
concentration decreased the relative density of Alphaproteo-
bacteria, and FUR2 at a similar concentration decreased the
density of Gammaproteobacteria as well as of the Cyto-
phaga–Flavobacterium of Bacteroides.
The effect of treated biofilms on larvalattachment
The effect of larval attachment of B. neritina varied accord-
ing to the compound tested (ANOVA: F = 5.5, Po 0.008), its
concentration (ANOVA: F = 12.9, Po 0.0001) and the combi-
nations of these factors (ANOVA: F = 2.9, Po 0.04, Fig. 5).
The percentage of larval attachment of B. neritina larvae on
the biofilms developed without QS-inhibiting compounds
(control) was 94� 5%. Larval attachment was similar to the
control on the biofilms treated with low concentrations
(10�4–10�5 M) of FUR2 and TRI and all tested concentra-
tions of FUR1 (Fig. 5). The biofilms developed in the
Table 2. Continued.
TRFs
(bp)
TRI FUR1 FUR2
CON10�3 10�4 10�5 10�3 10�4 10�5 10�3 10�4 10�5
445 Xa
451 Xa
483 Xb
Total
number TRFs
31 32 28 29 37 38 27 27 33 52
The presence and the fragment sizes (bps) of individual TRFs are denoted
as follows: Xa, present in three out of three replicates; Xb, present in two
out of three replicates; the presence in one out of three replicates is
omitted. Blank cells indicate the absence of particular TRFs in a given
biofilm.
TRI, triclosan; FUR1, 5-hydroxy-3[(1R)-1-hydroxypropyl]-4-methylfuran-
2(5H)-one; FUR2, (5R)-3,4-dihydroxy-5-[(1S)-1,2-dihydroxyethyl]furan-
2(5H)-one.
FUR1
FUR2
TRI
CON
Stress: 0.1310
1010
1010
10
10
1010
10 1010
1010
101010
10
1010
10
1010
1010
1010
ConCon
Con
Fig. 3. Multi-dimensional scaling (MDS) showing similarities in bacterial
communities formed in the presence of TRI (grey square), FUR1 (white
triangle) and FUR2 (black triangle) or without quorum-sensing blockers
(CON, white rhombus). The MDS plot is based on the Bray–Curtis
similarity matrix of the presence and absence of T-RFLP fragments in
various biofilm samples. Analysis of this matrix was based on two
dimensions. Numbers on the graph correspond to the concentrations of
the compounds used for biofilm treatment. Three replicated samples
were used for T-RFLP analysis. The stress value in the right upper corner
determines the fit of the reproduced distance matrix to the observed
distance matrix.
10–3 M
10–4 M
10–5 M
% D
AP
I cou
nt
0
20
40
60
80
100 Alpha BettaGamma CFLG
*
**
*
% D
AP
I cou
nt0
20
40
60
80
100
*
*
*
Treatment
TRI FUR1 FUR2 Control
% D
AP
I cou
nt
0
20
40
60
80
100
*
**
Fig. 4. Quantitative estimates of biofilm community composition in the
presence of TRI, FUR1 and FUR2 using group-specific FISH probes (see
Table 1) for Alphaproteobacteria (black), Betaproteobacteria (light grey),
Gammaproteobacteria (dark grey), Cytophaga–Flavobacteria (oblique
stroke) and some Firmicutes spp. (dashed). In control dishes (CON) no
QS blockers were added. All densities are expressed as a percentage
of total bacterial numbers obtained using dual hybridization by DAPI
staining. Densities of bacteria that are significantly lower than the control
ones according to a Dunnet test (ANOVA: Po 0.05) are indicated by
asterisks. Data are means (n = 5)� SD.
FEMS Microbiol Ecol 60 (2007) 177–188 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
183Antifouling activity of QS blockers

presence of high concentrations (10�3 M) of TRI and FUR2
significantly reduced (ANOVA: Dunnet test, Po 0.05) larval
attachment.
Both compounds (ANOVA: F = 15.3, Po 0.0001) and the
concentration (ANOVA: F = 32.1, Po 0.0001) significantly
affected biofilm formation, which in turn affected larval
attachment of Hydroides elegans (Fig. 6). The biofilms
formed at high concentrations (10�3 M) of all tested chemi-
cals significantly inhibited (ANOVA: F = 7.3, Dunnet test,
Po 0.05) the attachment of larvae compared with the
nontreated biofilms. Larval attachment on such biofilms
was reduced 1.3–4 times (Fig. 6). FUR2 reduced the attrac-
tiveness of biofilms more strongly than the other com-
pounds did at concentrations of 10�3–10�4 M.
Discussion
In earlier experiments, furanones produced by the red alga
Delisea pulchra were shown to inhibit larval attachment
(Steinberg et al., 1997), grazing (de Nys et al., 1996), and
microbial attachment (Manefield et al., 1999). In this study
we investigated for the first time an indirect control of larval
attachment by quorum-sensing (QS) blockers via the mod-
ification of marine bacterial communities.
Our experiments demonstrated that some chemical
compounds not only significantly decreased production of
violacein by C. violaceum induced by AHLs (quorum-
sensing signals) but also decreased bacterial densities and
modified the composition of microbial communities, as
revealed by both FISH and T-RFLP analysis. Triclosan
(TRI), 5-hydroxy-3[(1R)-1-hydroxypropyl]-4-methylfuran-
2(5H)-one (FUR1) and (5R)-3,4-dihydroxy-5-[(1S)-1,2-di-
hydroxyethyl]furan-2(5H)-one (FUR2) at 10�3 M had the
most striking effect on the density and composition of
bacterial communities. The effects of tested compounds
decreased with decreasing concentration of QS blockers.
FISH analysis showed that bacteria affected by the QS
blockers predominantly belonged to the Alphaproteobacter-
ia, Gammaproteobacteria or the Cytophaga–Flavobacterium
of the Bacteroides group. T-RFLP analysis demonstrated that
some bacterial ribotypes were absent after QS blocker
treatment, while other bacterial species were not affected by
QS blockers. This result suggests that the effect of QS
blockers was selective in our experiment. Analogously, the
AHL communication system (affected by the compounds
tested in this study) is highly species-specific (Whitehead
et al., 2001). For example, AHLs with side chains different
from the native ones interfere with signalling in the bacter-
ium Vibrio fischeri (Schaefer et al., 1996). Moreover, im-
plication of different AHLs leads to a different composition
of bacterial communities (McLean et al., 2005).
How did QS blockers affect the composition of microbial
communities in our experiments? We used two types of QS
blockers: furanones (FUR1 and FUR2), which inhibit quor-
um-sensing signals mediated by N-acylhomoserine lactones
(AHLs) (Rasmussen et al., 2000; Martinelli et al., 2004), and
triclosan (TRI), which inhibits the fatty acid biosynthesis
(Zhang & Dong, 2004). It has been shown that furanones
and triclosan interfere with AHL-mediated QS systems of
Gram-negative bacteria, in which a LuxR-type receptor
protein and a LuxI-type AHL synthase are the central
components (Fuqua & Greenberg, 2002; Zhang & Dong,
2004). Taking into account that in our experiments not all
Treatments
TRI FUR1 FUR2 Control
Atta
chm
ent %
0
20
40
60
80
100
10–3 M10–4 M10–5 M
*
*
Fig. 5. The effect of biofilms formed in the presence of TRI, FUR1 and
FUR2 at 10�3 M (black), 10�4 M (dark grey) and 10�5 M (light grey) on the
larval attachment of Bugula neritina. Attachment of larvae was ex-
pressed as a percentage. Levels of attachment of B. neritina on biofilms
treated by QS blockers that are significantly lower than levels of
attachment on the control (biofilms formed in FSW) according to a
Dunnet test (ANOVA: Po 0.05) are indicated by asterisks. Data are means
(n = 5)� SD.
TRI FUR1 FUR2 Control
Atta
chm
ent %
0
20
40
60
80
100 10–3 M10–4 M10–5 M
**
*
*
Fig. 6. The effect of biofilms formed in the presence of triclosan TRI,
FUR1 and FUR2 at 10�3 M (black), 10�4 M (dark grey) and 10�5 M (light
grey) on the larval attachment of Hydroides elegans. Attachment of
larvae was expressed as a percentage. Levels of attachment on biofilms
treated by QS blockers that are significantly lower than levels of
attachment on the control (biofilm formed with FSW) according to a
Dunnet test (ANOVA: Po 0.05) are indicated by asterisks. Data are means
(n = 5)� SD.
FEMS Microbiol Ecol 60 (2007) 177–188c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
184 S. Dobretsov et al.

tested concentrations of QS blockers were toxic to bacteria
or inhibited production of violacein by C. violaceum, we
conclude that the chemicals at concentrations of
10�3–10�5 M interact only with the quorum-sensing signal-
ling system of Gram-negative bacteria, thus inhibiting their
attachment, growth, and the formation of biofilms. Some of
bacterial strains that do not use AHL-mediated QS systems
may take advantage of the low density of competitors and
increase their growth. In our experiments, QS blockers did
not increase the density of low G1C Gram-positive bacteria
(mostly Firmicutes), as visualized by a LGC354a probe
(Meier et al., 1999). This suggests that some unique ribo-
types appearing in biofilms treated with QS blockers belong
to other bacterial groups.
A variety of QS blockers have been isolated from plants,
bacteria and fungi (reviewed by Zhang & Dong, 2004). For
example, halogenated furanones from the marine alga Deli-
sea pulchra inhibit the growth, swarming and biofilm
formation of Pseudoalteromonas aeruginosa and Escherichia
coli by interfering with bacterial AHLs (Rasmussen et al.,
2000; Hentzer et al., 2003; Ren et al., 2004). Furanones are
also produced by marine bacteria, green, red and brown
algae, sponges, fungi, and ascidians (Kjelleberg et al., 2001),
as well as by higher plants (Slaughter 1999). For example,
butenolides [2(5H)-furanones] have been isolated from
Streptomyces and from Hortonia spp. (Cho et al., 2001).
Gram-positive Bacillus spp. and Gram-negative Agrobacter-
ium tumefaciens and Arthrobacter spp. (Zhang et al., 2002,
2004) produced AHL-lactonase, which degrades bacterial
AHL signals. It is important to note that most of the QS
signals and their inhibitors were tested only in biomedical
experiments and against monospecies biofilms (for reviews
see Bauer & Robinson, 2002; Smith et al., 2003; Zhang &
Dong, 2004). Information about QS signals and their
inhibitors in the marine environment is fairly limited.
Examples include the production of AHLs by Roseobacter
spp. isolated from ‘marine snow’ (Gram et al., 2002), by
Roseobacter sp. and Vibrio sp. associated with sponges
(Taylor et al., 2004), and by marine free-living Alphaproteo-
bacteria and bacteria associated with eukaryotic algae
(Wagner-Dobler et al., 2005). It is possible to conclude that
QS signals and their inhibitors may play an important role
in aquatic microbial communities and should be a main
target of future studies.
In our experiments, the tested compounds modified
biofilms, which, in turn, affected the larval attachment of
the bryozoan B. neritina and the polychaete H. elegans. This
effect is concentration-dependent: all QS blockers at the
concentration 10�3 M affected the formation of biofilms,
which in turn inhibited larval attachment, but at the
concentration of 10�4 M only biofilms formed in the pre-
sence of FUR2 decreased the attachment of H. elegans
compared with the control. Were the concentrations of QS
blockers in our investigation realistic? Unfortunately, actual
concentrations of AHLs and QS blockers in marine biofilms
have not yet been determined. Taking into account that the
concentration of halogenated furanones in the alga Delisea
pulchra reaches 3–25 mg g�1 (Wright et al., 2000) it is
possible to propose that QS blockers in biofilms may exist
at similar concentrations. In the laboratory, halogenated
furanones from D. pulchra inhibit bacterial attachment at
10 mM (Maximilien et al., 1998), and QS signals in biofilms
of P. aeruginosa at 10 mM (Hentzer et al., 2003) and in
biofilms of C. violaceum at 100 mM (Martinelli et al., 2004).
These examples demonstrate that furanones can inhibit QS
and bacterial attachment at wide range of concentrations
that are similar to those used in this study.
Variations in the larval settlement response to the bio-
films treated with QS blockers can be explained in two ways.
First, compounds may decrease bacterial density, which in
turn causes low levels of larval attachment (Lau & Qian,
1997; Huang & Hadfield, 2003). Second, compounds may
modify the diversity of microbial communities. Bacterial
species composition and abundance in a microbial biofilm
can determine the biofilm inductiveness towards larval
settlement (Lau & Qian, 1997; Huang & Hadfield, 2003;
Qian et al., 2003; Dobretsov & Qian, 2006).
In conclusion, the experiments described here demon-
strate that QS blockers may affect larval attachment indir-
ectly by the modification of bacterial films. This finding may
inspire the search for novel antifouling technologies that
could be used in the control of both micro- and macrofoul-
ing. Overall, the indirect effects of QS blockers on larval
settlement deserve further investigation.
Acknowledgements
We thank Dr Urs Sequin (Switzerland) for providing us with
a synthetic furanone 5-hydroxy-3[(1R)-1-hydroxypropyl]-
4-methylfuran-2(5H)-one, and two anonymous reviewers
for their comments. This work was supported by RGC
grants (HKUST6402/05 M and CA04/05.SC01) to P.Y.Q.
and partially by a grant from the Alexander von Humboldt
Foundation to S.D.
References
Amann RI, Krumholz L & Stahl DA (1990) Fluorescent-
oligonucleotide probing of whole cells for determinative,
phylogenetic, and environmental studies in microbiology.
J Bacteriol 172: 762–770.
Armstrong E, Boyd KG & Burgess JG (2000) Prevention of marine
biofouling using natural compounds from marine organisms.
Biotechnol Annu Rev 6: 221–241.
Bauer W & Robinson JB (2002) Disruption of bacterial quorum
sensing by other organisms. Curr Opin Biotechnol 13: 234–237.
FEMS Microbiol Ecol 60 (2007) 177–188 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
185Antifouling activity of QS blockers

Blackwood CB, Marsh T, Kim SH & Paul EA (2003) Terminal
restriction fragment length polymorphism data analysis for
quantitative comparison of microbial communities. Appl
Environ Microbiol 69: 926–932.
Bryan PJ, Rittschof D & Qian PY (1997) Settlement inhibition of
bryozoan larvae by bacterial films and aqueous leachates. Bull
Mar Sci 61: 849–857.
Buchan A, Newell SY, Butler M, Biers EJ, Hollibaugh JT & Moran
MA (2003) Dynamics of bacterial and fungal communities on
decaying salt marsh grass. Appl Environ Microbiol 69:
6676–6687.
Chen X, Schauder S, Potier N, Van Dorsselaer, Pelczer I, Bassler
BL & Hughson FM (2002) Structural identification of a
bacterial quorum sensing signal containing boron. Nature 415:
545–549.
Cho KW, Lee HS, Rho JR, Kim TS, Mo SJ & Shin J (2001) New
lactone-containing metabolites from a marine-derived
bacterium of the genus Streptomyces. J Nat Prod 64: 664–667.
Clare AS, Rittschof D, Gerhard DJ & Maki JS (1992) Molecular
approaches to non-toxic antifouling. Invert Reprod Dev 22:
67–76.
Davies SF, Parsek JP, Pearson JP, Iglewski BH, Costerton JW &
Greenberg EP (1998) The involvement of cell-to-cell signals in
the development of a bacterium biofilm. Science 280: 295–298.
de Nys R, Steinberg PD, Rogers CN, Charlton TS & Duncan MW
(1996) Quantitative variation of secondary metabolites in the
sea hare Aplysia parvula and its host plant, Delisea pulchra.
Mar Ecol Prog Ser 130: 135–146.
de Nys R, Rice S, Manefield M, Srinivasan S, McDougald D, Loh
A, Ostling J, Lindum P, Givskov M, Steinberg P & Kjelleberg S
(2000) Cross-talk in bacterial extracellular signaling systems.
Microbial Biosystems: New Frontiers. Proceedings of the 8th
International Symposium on Microbial Ecology (Bell CR,
Brylinsky M & Johnson-Green P, eds), pp. 23–29. Atlantic
Canada Society for Microbiology, Halifax.
Dobretsov S & Qian PY (2002) Effect of bacteria associated with
the green alga Ulva reticulata on marine micro- and
macrofouling. Biofouling 18: 217–228.
Dobretsov S & Qian PY (2006) Facilitation and inhibition of
larval attachment of the bryozoan Bugula neritina in
association with mono-species and multi-species biofilms. J
Exp Mar Biol Ecol 333: 263–264.
Dobretsov S, Dahms HU & Qian PY (2006) Inhibition of
biofouling by marine microorganisms and their metabolites.
Biofouling 22: 43–54.
Dunbar J, Ticknor LO & Kuske CR (2001) Phylogenetic
specificity and reproducibility and new method for analysis of
terminal restriction fragment profiles of 16S rRNA genes from
bacterial communities. Appl Environ Microbiol 67: 190–197.
Egan S, Thomas T, Holmstr�m C & Kjelleberg S (2000)
Phylogenic relationship and antifouling activity of bacterial
epiphytes from marine algae Ulva lactuca. Environ Microbiol 2:
343–347.
Egan S, James S, Holmstr�m C & Kjelleberg S (2001) Inhibition
of algal spore germination by the marine bacterium
Pseudoalteromonas tunicata. FEMS Microbiol Ecol 35: 67–73.
Evans SM (1999) TBTor not TBT?: that is the question. Biofouling
14: 117–129.
Fuqua C & Greenberg EP (2002) Listening in on bacteria: acyl-
homoserine lactone signaling. Nat Rev Mol Cell Biol 3:
685–695.
Gram L, Grossart HP, Schlingloff A & Kio+rboe T (2002) Possible
quorum sensing in marine snow bacteria: production of
acylated homoserine lactones by Roseobacter strains isolated
from marine snow. Appl Environ Microbiol 68: 4111–4116.
Gray KM & Garey JR (1999) The evolution of bacterial Luxl and
LuxR quorum sensing regulators. Microbiology 147:
2379–2387.
Haglund AL, Trnblom E, Bostrm B & Tranvik L (2002) Large
differences in the fraction of active bacteria in plankton,
sediments, and biofilm. Microbial Ecol 43: 232–241.
Harder T & Qian PY (1999) Induction of larval attachment and
metamorphosis in the serpulid polychaete Hydroides elegans by
dissolved free amino acids: isolation and identification. Mar
Ecol Prog Ser 179: 259–271.
Hentzer M, Wu H, Andersen JB et al. (2003) Attenuation of
Pseudomonas aeruginosa virulence by quorum sensing
inhibitors. EMBO J 22: 3803–3815.
Holmstr�m C & Kjelleberg S (1999) Marine Pseudoalteromonas
species are associated with higher organisms and produce
active extracellular compounds. FEMS Microbiol Ecol 30:
285–293.
Huang S & Hadfield M (2003) Composition and density of
bacterial biofilms determine larval settlement of the polychaete
Hydroides elegans. Mar Ecol Prog Ser 260: 161–172.
Huang OS, Thiyagarajan V, Zhang R, Wu RSS & Qian PY
Attachment of Balanus amphitrite larvae to biofilms originated
from contrasting environments in Hong Kong: results of
laboratory and field experiments. Mar Ecol Prog Ser, in press.
Joint I, Tait K, Callow ME, Callow JA, Milton D, Williams P &
Camara M (2002) Cell-to-Cell communication across the
Prokaryote–Eukaryote boundary. Science 298: 1207.
Kirchman D, Graham D, Reish D & Mitchell R (1982) Lectins
may mediate in the settlement and metamorphosis of Janua
(Dexiospira) brasiliensis Grube (Polychaetea: Spirorbidae).
Mar Biol Lett 3: 201–222.
Kjelleberg S, Steinberg P, Givskov M, Gram L, Manefield M & De
Nys R (2001) Do marine natural products interfere with
prokaryotic AHL regulatory systems? Aquat Microb Ecol 13:
85–93.
Lau SCK & Qian PY (1997) Phlorotannins and related
compounds as larval settlement inhibitors of a tube-building
polychaete Hydroides elegans (Haswell). Mar Ecol Prog Ser 159:
219–227.
Lau SCK & Qian PY (2001) Larval settlement in the serpulid
polychaete Hydroides elegans in response to bacterial films: an
investigation of the nature of putative larval settlement cue.
Mar Biol 138: 321–328.
FEMS Microbiol Ecol 60 (2007) 177–188c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
186 S. Dobretsov et al.

Liu WT, Marsh TL, Cheng H & Forney LJ (1997)
Characterization of microbial diversity by determining
terminal restriction fragment length polymorphisms of genes
encoding 16S rRNA. Appl Environ Microbiol 63: 4516–4522.
Manefield M & Turner SL (2002) Quorum sensing in context: out
of molecular biology and into microbial ecology. Microbiology
148: 3762–3764.
Manefield M, de Nys R, Kumar N, Read R, Givskov M, Steinberg
P & Kjelleberg S (1999) Evidence that halogenated furanones
from Delisea pulchra inhibit acylated homoserine lactone
(AHL)-mediated gene expression by displacing the AHL signal
from its receptor protein. Microbiology 145: 283–291.
Manz W, Amann R, Ludwig W, Wagner M & Schleifer KH (1992)
Phylogenetic oligodeoxynucleotide probes for the major
subclasses of proteobacteria: problems and solutions. Syst Appl
Microbiol 15: 593–600.
March JC & Bentley WE (2004) Quorum sensing and bacterial
cross-talk in biotechnology. Curr Opin Biotechnol 15: 495–502.
Martinelli D, Grossmann G, Sequin U, Brandl H & Bachofen R
(2004) Effects of natural and chemically synthesized furanones
on quorum sensing in Chromobacterium violaceum. BMC
Microbiol 4: 1–10.
Maximilien R, De Nys R, Holmstr�m C, Gram L, Givskov M,
Crass K, Kjelleberg S & Steinberg P (1998) Chemical
mediation of bacterial surface colonisation by the secondary
metabolites from the red algae Delisea pulchra. Aquat Microb
Ecol 15: 233–246.
McLean RJ, Pierson LS & Fuqua C (2004) A simple screening
protocol for the identification of quorum sensing antagonists.
J Microbiol Methods 58: 351–360.
McLean RJC, Barnes MB, Windham MK, Merchant M, Forstner
MRJ & Fuqua C (2005) Cell–cell influences on bacterial
community development in aquatic biofilms. Appl Environ
Microbiol 71: 8987–8990.
McLean KH, McClean KH, Winson MK et al. (1997) Quorum
sensing and Chromobacterium violaceum: exploitation of
violacein production and inhibition for the detection of N-
acylhomoserine lactones. Microbiology 143: 3703–3711.
Meier H, Amann R, Ludwig W & Schleifer KH (1999) Specific
oligonucleotide probes for in situ detection of a major group
of Gram-positive bacteria with low DNA G-C content. Syst
Appl Microbiol 22: 186–196.
Miller MB & Bassler BL (2001) Quorum sensing in bacteria. Annu
Rev Microbiol 55: 165–199.
Parsek MR & Greenberg EP (2000) Acyl-homoserine lactone
quorum sensing in gram-negative bacteria: a signaling
mechanism involved in associations with higher organisms.
Proc Natl Acad Sci USA 97: 8789–8793.
Qian PY, Thiyagarajan V, Lau SCK & Cheung SCK (2003)
Relationship between bacterial community profile in biofilm
and attachment of the acorn barnacle Balanus amphitrite.
Aquat Microb Ecol 33: 225–237.
Quinn GP & Keough MJ (2002) Experimental Design and Data
Analysis for Biologists. Cambridge University Press,
Cambridge.
Railkin AI (1998) The pattern of recovery of disturbed microbial
communities inhabiting hard substrates. Hydrobiologia 385:
47–57.
Rasmussen TB, Manefield M, Andersen JB, Eberl L, Anthoni U,
Christopersen C, Steinberg P, Ratnayake R, Karunaratne V,
Ratnayake BBM, Kumar V, MacLeod JK & Simmonds P (2000)
Two new lactones with mosquito larvicidal activity from three
Hortonia species. J Nat Prod 64: 376–378.
Ren D, Sims JJ & Wood TK (2001a) Inhibition of biofilms
formation and swarming of Escherichia coli by (5Z)-4-Bromo-
5-(Bromomethylene)-3-Buthyl-2(5H)-Furanone. Environ
Microbiol 3: 731–736.
Ren D, Sims JJ & Wood TK (2001b) Synthetic furanones inhibit
quorum-sensing and enhance bacterial clearance in
Pseudomonas aeruginosa lung infection in mice. J Antimicrob
Chemother 53: 1054–1061.
Ren D, Bedzyk LA, Thomas SM, Ye RW & Wood TK (2004)
Differential gene expression shows natural brominated
furanones interfere with the autoinducer-2 bacterial signalling
system of Escherichia coli. Biotechnol Bioeng 1: 23–31.
Rice SA, Givskov M, Steinberg P & Kjelleberg S (1999) Bacterial
signals and antagonists: the interaction between bacteria and
higher organisms. J Mol Microbiol Biotechnol 1: 23–31.
Rodriguez SR, Ojeda FP & Inestrosa NC (1993) Settlement of
benthic marine invertebrates. Mar Ecol Prog Ser 97: 193–207.
Sauer K, Camper AK, Ehrlich GD, Costerton JW & Davies DG
(2002) Pseudomonas aeruginosa displays multiple phenotypes
during development as a biofilm. J Bacteriol 184: 1140–1154.
Schaefer AL, Hanzelka BL, Eberhard A & Greenberg EP (1996)
Quorum sensing in Vibrio fischeri: probing autoinducer –
LuxR interactions with autoinducer analogs. J Bacteriol 178:
2897–2901.
Shapiro SS & Wilk MB (1965) An analysis of variance test for
normality (complete samples). Biometrika 52: 591–611.
Slaughter CJ (1999) The naturally occurring furanones:
formation and function from pheromone to food. Biol Rev
Cambridge Philos Soc 74: 259–276.
Smith KM, Bu Y & Suga H (2003) Library screening for synthetic
agonists and antagonists of a Pseudomonas aeruginosa
autoinducer. Chem Biol 10: 563–571.
Sperandio V, Torres AG, Giron JA & Kaper JB (2001) Quorum
sensing is a global regulatory mechanism in
enterohemorrhagic Escherichia coli. J Bacteriol 183: 5187–5197.
Steinberg PD, Schneider R & Kjelleberg S (1997) Chemical
defenses of seaweeds against microbial colonization.
Biodegradation 8: 211–220.
Taylor MW, Schupp PJ, Baillie HJ, Charlton TS, de Nys R,
Kjelleberg S & Steinberg PD (2004) Evidence for acyl
homoserine lactone signal production in bacteria associated
with marine sponges. Appl Environ Microbiol 70: 4387–4389.
Thomas KV (2001) The environmental fate and behaviour of
antifouling paint booster biocides: a review. Biofouling 17:
73–86.
Wagner-Dobler I, Thiel V, Eberl L, Allgaier M, Bodor A, Meyer S,
Ebner S, Hennig A, Pukall R & Schulz S (2005) Discovery of
FEMS Microbiol Ecol 60 (2007) 177–188 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
187Antifouling activity of QS blockers

complex mixtures of novel long-chain quorum sensing signals
in free-living and host-associated marine alphaproteobacteria.
Chem Biochem 6: 2195–2206.
Webster NS, Smith LD, Heyward AJ, Watts JEM, Webb RI,
Blackall LL & Negri AP (2004) Metamorphosis of a
scleratinian coral in response to microbial biofilms. Appl
Environ Microbiol 70: 1213–1221.
Whitehead NA, Barnard AM, Slater H, Simpson NJ & Salmond
GP (2001) Quorum sensing in Gram-negative bacteria. FEMS
Microbiol Rev 25: 365–404.
Wright JT, de Nys R & Steinberg P (2000) Geographic variation in
halogenated furanones from the red alga Delisea pulchra and
associated herbivores and epiphytes. Mar Ecol Prog Ser 207:
227–241.
Xavier KB & Bassler BL (2003) LuxS quorum sensing: more than
just a numbers game. Curr Opin Microbiol 6: 191–197.
Yebra DM, Kiil S & Dam-Johansen K (2004) Antifouling
technology – past, present and future steps towards efficient
and environmentally friendly antifouling coatings. Prog Org
Coat 50: 75–104.
Zar JH (1999) Biostatistical Analysis, 4th edn. Prentice Hall
International, Inc., Upper Saddle River, NJ.
Zhang LH & Dong YH (2004) Quorum sensing and signal
interference: diverse implications. Mol Microbiol 53:
1563–1571.
Zhang HB, Wang C & Zhang LH (2002) Genetic control of
quorum sensing signal turnover in Agrobactrium tumefaciens.
Proc Natl Acad Sci USA 99: 4638–4643.
Zhang HB, Wang C & Zhang LH (2004) The quormone
degradation system of Agrobacterium tumefaciens is regulated
by starvation signal and stress alarmone. Mol Microbiol 52:
1389–1401.
FEMS Microbiol Ecol 60 (2007) 177–188c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
188 S. Dobretsov et al.
Copyright © 2022 FDOKUMEN







![A Study On Agus Musthafa's Interpretation Over Qs. Al-Anfal [8]](https://static.fdokumen.com/doc/165x107/6315486ac72bc2f2dd04a6c7/a-study-on-agus-musthafas-interpretation-over-qs-al-anfal-8.jpg)







![STAM New Sample Qs - Home [howardmahler.com]](https://static.fdokumen.com/doc/165x107/632a4ed37e519a0bdc068b7b/stam-new-sample-qs-home-howardmahlercom.jpg)




