
Toluene inhalation modulates dentate gyrus granule cell output in vivo
10
Toluene inhalation modulates dentate gyrus granule cell output in vivo Jimmie M. Gmaz, Brittany A. Matthews, Bruce E. McKay ⁎ Department of Psychology, Wilfrid Laurier University, Waterloo, Ontario, Canada N2L 3C5 abstract article info Article history: Received 11 February 2012 Received in revised form 30 April 2012 Accepted 30 April 2012 Available online 8 May 2012 Keywords: Dentate gyrus fEPSP Perforant path Population spike Rat Toluene Toluene, a psychoactive volatile solvent found in adhesives and other products, is inhaled for its euphoric and intoxicating effects. Toluene inhalation additionally results in cognitive disturbances including impairments in select types of spatial and non-spatial memory, which converging evidence suggests may involve neurons of the dentate gyrus. In the present study we examined the effects of acute binge-like (~ 5000 ppm) toluene inhalation on dentate gyrus granule cell output and perforant path synaptic transmission, using extracellular field potential recordings in anesthetized adult rats in vivo. We found that toluene rapidly and reversibly increased or decreased the amplitudes of evoked population spikes from granule cells over a wide range of stimulation intensities. These changes in granule cell output could not be accounted for by changes in perforant path action potential discharge or presynaptic neurotransmitter release. A marked decrease in the power of the theta rhythm measured within the dentate gyrus was additionally noted. Overall our results suggest that inhalation of abuse-relevant concentrations of toluene changes the readout of perforant path inputs by dentate gyrus granule cells, putatively through a postsynaptic mechanism, and may contribute to explanations for specific learning and memory deficits associated with toluene inhalation. © 2012 Elsevier Inc. All rights reserved. 1. Introduction Toluene is the principal rewarding volatile chemical found in many household products such as adhesives and paint thinner. Toluene vapors, when inhaled from these products, produce a euphoric, in- toxicated state accompanied by numerous cognitive impairments (Bowen et al., 2006). The recreational use of toluene is widespread, with alarmingly high rates of abuse in impoverished socioeconomic groups (McDonald, 1999; Paglia-Boak et al., 2011). Despite aware- ness of the behavioral and cognitive deficits associated with toluene use, and evidence that the recreational use of toluene-containing products is increasing, the neurobiological mechanisms underlying the effects of toluene are incompletely understood. The use and abuse of toluene-containing products has particularly pronounced effects on learning and memory (Filley et al., 2004), with converging evidence that these impairments may be explained in part by changes in hippocampal function. For instance, acute toluene inhalation results in impairments in hippocampal-dependent refer- ence memory tasks, including those with spatial and non-spatial task demands (Andersen et al., 2007; Baydas et al., 2005; Rosenberg et al., 2002; von Euler et al., 2000; Win-Shwe and Fujimaki, 2012; Win-shwe et al., 2009). Additionally, within the hippocampus, toluene exposure increases extracellular glutamate and taurine levels, de- creases extracellular acetylcholine levels, and decreases amounts of NR1 and NR2B transcripts (Honma and Suda, 2004; Perrine et al., 2011; Win-Shwe and Fujimaki, 2012; Win-Shwe et al., 2007). Granule cell atrophy and even degeneration of hippocampal neurons has been reported for longer-term binge-like exposures to toluene (Gotohda et al., 2002; Kanter, 2008). Together this evidence suggests that toluene should evoke measurable changes in the physiology of hippocampal neurons, and indeed, in vitro experiments have demonstrated that toluene inhibits synaptically-evoked population spikes in select hip- pocampal regions, likely due to enhanced GABA-mediated inhibition (Ikeuchi et al., 1993; MacIver, 2009). However, these experiments were completed in acute brain slices, and thus the effects of toluene on the physiology of hippocampal neurons in intact neural networks remain to be shown. The purpose of the present series of experiments was to evaluate the physiological effects of an acute abuse-relevant (binge-like) con- centration of toluene vapor (~ 5000 ppm) in vivo, on population spike discharge from dentate gyrus (DG) granule cells and the characteris- tics of synaptic transmission from perforant path (PP) inputs. The DG was chosen as the region of interest as it is the entry point to the hippocampus, plays an important role in filtering incoming synaptic input that will be processed and distributed to other regions of the hippocampus, and contributes to the performance of many of the learning and memory tasks which are impaired by acute toluene in- halation (Andersen et al., 2007; Bowen et al., 2006). Our experiments, which utilized extracellular field potential recordings in anesthetized rats, revealed that during toluene vapor inhalation there are robust Neurotoxicology and Teratology 34 (2012) 403–412 ⁎ Corresponding author at: Department of Psychology, Wilfrid Laurier University, 75 University Ave W., Waterloo, ON, Canada N2L 3C5. Tel.: +1 519 884 0710x3033; fax: +1 519 746 7605. E-mail address: [email protected] (B.E. McKay). 0892-0362/$ – see front matter © 2012 Elsevier Inc. All rights reserved. doi:10.1016/j.ntt.2012.04.011 Contents lists available at SciVerse ScienceDirect Neurotoxicology and Teratology journal homepage: www.elsevier.com/locate/neutera
Transcript of Toluene inhalation modulates dentate gyrus granule cell output in vivo

Neurotoxicology and Teratology 34 (2012) 403–412
Contents lists available at SciVerse ScienceDirect
Neurotoxicology and Teratology
j ourna l homepage: www.e lsev ie r .com/ locate /neutera
Toluene inhalation modulates dentate gyrus granule cell output in vivo
Jimmie M. Gmaz, Brittany A. Matthews, Bruce E. McKay ⁎Department of Psychology, Wilfrid Laurier University, Waterloo, Ontario, Canada N2L 3C5
⁎ Corresponding author at: Department of Psycholo75 University Ave W., Waterloo, ON, Canada N2L 3C5.fax: +1 519 746 7605.
E-mail address: [email protected] (B.E. McKay).
0892-0362/$ – see front matter © 2012 Elsevier Inc. Alldoi:10.1016/j.ntt.2012.04.011
a b s t r a c t
a r t i c l e i n f oArticle history:Received 11 February 2012Received in revised form 30 April 2012Accepted 30 April 2012Available online 8 May 2012
Keywords:Dentate gyrusfEPSPPerforant pathPopulation spikeRatToluene
Toluene, a psychoactive volatile solvent found in adhesives and other products, is inhaled for its euphoric andintoxicating effects. Toluene inhalation additionally results in cognitive disturbances including impairmentsin select types of spatial and non-spatial memory, which converging evidence suggests may involve neuronsof the dentate gyrus. In the present study we examined the effects of acute binge-like (~5000 ppm) tolueneinhalation on dentate gyrus granule cell output and perforant path synaptic transmission, using extracellularfield potential recordings in anesthetized adult rats in vivo. We found that toluene rapidly and reversiblyincreased or decreased the amplitudes of evoked population spikes from granule cells over a wide range ofstimulation intensities. These changes in granule cell output could not be accounted for by changes inperforant path action potential discharge or presynaptic neurotransmitter release. A marked decrease inthe power of the theta rhythmmeasured within the dentate gyrus was additionally noted. Overall our resultssuggest that inhalation of abuse-relevant concentrations of toluene changes the readout of perforant pathinputs by dentate gyrus granule cells, putatively through a postsynaptic mechanism, and may contribute toexplanations for specific learning and memory deficits associated with toluene inhalation.
© 2012 Elsevier Inc. All rights reserved.
1. Introduction
Toluene is the principal rewarding volatile chemical found inmanyhousehold products such as adhesives and paint thinner. Toluenevapors, when inhaled from these products, produce a euphoric, in-toxicated state accompanied by numerous cognitive impairments(Bowen et al., 2006). The recreational use of toluene is widespread,with alarmingly high rates of abuse in impoverished socioeconomicgroups (McDonald, 1999; Paglia-Boak et al., 2011). Despite aware-ness of the behavioral and cognitive deficits associated with tolueneuse, and evidence that the recreational use of toluene-containingproducts is increasing, the neurobiological mechanisms underlyingthe effects of toluene are incompletely understood.
The use and abuse of toluene-containing products has particularlypronounced effects on learning and memory (Filley et al., 2004), withconverging evidence that these impairments may be explained inpart by changes in hippocampal function. For instance, acute tolueneinhalation results in impairments in hippocampal-dependent refer-ence memory tasks, including those with spatial and non-spatialtask demands (Andersen et al., 2007; Baydas et al., 2005; Rosenberget al., 2002; von Euler et al., 2000; Win-Shwe and Fujimaki, 2012;Win-shwe et al., 2009). Additionally, within the hippocampus, toluene
gy, Wilfrid Laurier University,Tel.: +1 519 884 0710x3033;
rights reserved.
exposure increases extracellular glutamate and taurine levels, de-creases extracellular acetylcholine levels, and decreases amountsof NR1 and NR2B transcripts (Honma and Suda, 2004; Perrine et al.,2011;Win-Shwe and Fujimaki, 2012;Win-Shwe et al., 2007). Granulecell atrophy and even degeneration of hippocampal neurons has beenreported for longer-term binge-like exposures to toluene (Gotohda etal., 2002; Kanter, 2008). Together this evidence suggests that tolueneshould evoke measurable changes in the physiology of hippocampalneurons, and indeed, in vitro experiments have demonstrated thattoluene inhibits synaptically-evoked population spikes in select hip-pocampal regions, likely due to enhanced GABA-mediated inhibition(Ikeuchi et al., 1993; MacIver, 2009). However, these experimentswere completed in acute brain slices, and thus the effects of tolueneon the physiology of hippocampal neurons in intact neural networksremain to be shown.
The purpose of the present series of experiments was to evaluatethe physiological effects of an acute abuse-relevant (binge-like) con-centration of toluene vapor (~5000 ppm) in vivo, on population spikedischarge from dentate gyrus (DG) granule cells and the characteris-tics of synaptic transmission from perforant path (PP) inputs. The DGwas chosen as the region of interest as it is the entry point to thehippocampus, plays an important role in filtering incoming synapticinput that will be processed and distributed to other regions of thehippocampus, and contributes to the performance of many of thelearning and memory tasks which are impaired by acute toluene in-halation (Andersen et al., 2007; Bowen et al., 2006). Our experiments,which utilized extracellular field potential recordings in anesthetizedrats, revealed that during toluene vapor inhalation there are robust

Fig. 1. Schematic of the sequence of electrophysiological protocols used in the experi-ments. I/O = test of input/output relations; Sp = recording of spontaneous(unstimulated) activity; and PPS = paired-pulse stimulation protocols. During control,continuous toluene, and phasic toluene, black bars indicate the presence of toluenevapor. * Phasic groups received an additional 5 min of toluene exposure.
404 J.M. Gmaz et al. / Neurotoxicology and Teratology 34 (2012) 403–412
reversible changes in the excitability of DG granule cells, mediatedostensibly by postsynaptic mechanisms. Our results may accountfor some of the observed learning and memory deficits produced bythis widely abused organic solvent.
2. Materials and methods
2.1. Animals
Male and female adult (2–3.5 months old) Long–Evans rats (CharlesRiver Laboratories, Saint-Constant, QC) were used in the experiments.Rats were single or pair housed and maintained under standard condi-tions (12:12 photoperiod with lights on at 0700 h local time; constanttemperature of 21 °C; food and water available ad libitum). All experi-ments were approved by the Wilfrid Laurier University Animal CareCommittee in accordance with the guidelines of the Canadian Councilon Animal Care.
2.2. Chemicals
Chemicals andothermaterialswere obtained fromVWR(Mississauga,ON) unless stated otherwise. All compressed gases were obtained fromPraxair Canada Inc. (Kitchener, ON).
2.3. Anesthesia
Rats were anesthetized in a custom-constructed induction cham-ber with 5% isoflurane (Baxter Healthcare Corporation, Deerfield, IL)delivered by a T3 isoflurane vaporizer (Smiths Medical Canada Ltd,Markham, ON); oxygen was provided at 550 ml/min. Rats remained inthe induction chamber until full inhibition of the hind-pawwithdrawalreflex was achieved. Rats were then transferred and mounted into astereotaxic frame (ASI Instruments, Warren, MI) and continuously pro-vided isoflurane through a nosecone. After ~5 min the isoflurane wasreduced to 2.25 to 2.5% and maintained at this level for the remainderof the experiment. Body temperature was monitored rectally andmaintained at ~37 °C with the use of a heating pad.
2.4. Surgery for electrophysiology
Burr holes were drilled into the dorsal skull at the following loca-tions: male rats (Paxinos and Watson, 2005): 7.9 to 8.1 mm posteriorand 4.3 mm lateral to bregma (above right medial PP) and 3.5 to3.55 mm posterior and 2.0 to 2.05 mm lateral to bregma (above rightDG, granule cell body layer); female rats: 7.3 mm posterior and 3.9 to3.95 mm lateral (above right medial PP) and 3.1 to 3.3 mm posteriorand 1.9 to 2.15 mm lateral to bregma (above right DG, granule cellbody layer). A reference electrode was inserted through a third burrhole and onto the mid-parietal region of the contralateral hemisphere.
2.5. Electrophysiology
Stimulating and recording electrodes were custom-constructedfrom 75 μm diameter Teflon-coated tungsten wire inserted into 26gauge stainless steel cannulae (A-M Systems, Carlsborg, WA). Thestimulating electrode consisted of a single-barrel cannula, whereaseither 1-barrel or 4-barrel cannulae were used for recordings. Elec-trodes were lowered into the brain until fEPSP characteristics wereoptimized (final depth for electrodes in male rats were approximately2.4 to 3.8 mm (recording) and 1.65 to 3.5 mm (stimulating) as mea-sured from the dura; final electrode depths for female rats were 2.4to 4.0 mm (recording) and 1.45 to 2.75 mm(stimulating) asmeasuredfrom the dura. fEPSPs were evoked by 100 μs square pulses at 0.05 Hz(1 per 20 s) via an A365 isolated current stimulator (World PrecisionInstruments, Sarasota, FL). fEPSPs were measured via an EX-1 singlechannel differential amplifier (Dagan Corporation, Minneapolis, MN)
or a Model 1700 four channel differential amplifier (A-M Systems)with low and high pass filters set to achieve a bandwidth of 0.1 to20 kHz; signals were digitized and sent to the host computer via aDigidata 1440A (Danaher Corporation, Sunnyvale, CA), and recordedwith pCLAMP 10 software (Danaher Corporation).
The timeline for our electrophysiological protocols is illustrated inFig. 1. fEPSPs were monitored and allowed to stabilize for approxi-mately 1 h, after which the input–output relation was assessed.The input–output relation typically consisted of 15 equal-sized stepsfrom 0 μA through an intensity that saturated the amplitude of thepopulation spike; responses were obtained at 0.05 Hz. A 2 min periodof stimulation-free recording was subsequently obtained for thetaband (8–13 Hz) frequency analysis (not done for all rats). Next,paired-pulse stimulation protocols were conducted using a stimulusintensity subthreshold for eliciting population spikes; paired-pulsestimuli were delivered once every 25 s (0.04 Hz) using inter-pulse in-tervals of 20, 50, 100, 200, 500, 1000 and 5000 ms; the same inter-pulse intervals were immediately repeated using a stimulus intensitysufficient to elicit population spikes. The stimulation intensity thatevoked a population spike of ~50–75% of the maximal amplitudeduring assessment of the input–output relation was then used toobtain a 15 min baseline (no toluene) recording (stimulation at0.05 Hz). Following baseline recording, rats were exposed to toluenevapor through the nosecone continuously for 30 min, or phasically for30 min (5 min toluene vapor then 5 min no toluene vapor, repeatedthree times). Controls did not receive toluene. After this 30 minperiod the toluene vapor was applied for the remainder of the re-cording session (control rats received no toluene during this period;rats in the phasic groups received an extra 5 min of toluene beforeproceeding); at the onset of this period a second input–output relationwas measured, followed by 2 min of stimulation-free recording forfrequency analysis, and then the paired-pulse stimulation protocolsdescribed above. At the completion of the experiment each rat wasremoved from the stereotaxic frame and rapidly decapitated whilestill anesthetized.
2.6. Electrolytic brain lesions and cresyl violet staining
Histological confirmation of recording and stimulating electrodeplacement sites were completed for a subset of male and female rats(n=6 total, including rats in both phasic-increasing and phasic-decreasing groups). Lesionswere produced by passing 6 mAof currentfor 10 s through each electrode. Brains were rapidly extracted, fixedand cryoprotected, sectioned on a cryostat (Leica Microsystems,Concord, ON), and stained with cresyl violet according to a standardprotocol (Gerfen, 1997). Lesion sites were visualized with an OlympusBX43 research light microscope (4× objective) equipped with anOlympus XM10 Monochrome fluorescence CCD camera controlledby cellSens imaging software (Olympus, Markham, ON). In every ratexamined (n=6 of 6) the histology confirmed that the recording elec-trodes had successfully been placed into the granule cell layer of thesuprapyramidal blade of the dorsal DG (Fig. 2A) and the stimulating

Fig. 2. Histological verification of electrode placements. Black dots indicate the loca-tions of the recording (A) and stimulating (B) electrodes in 6 rats.Rat brain line art adapted from Paxinos and Watson (2005).
405J.M. Gmaz et al. / Neurotoxicology and Teratology 34 (2012) 403–412
electrode had been placed into the medial PP (Fig. 2B). Because of theconsistency, only a subset of rats was assessed histologically.
2.7. Electrophysiological data analysis
All electrophysiological analyses were completed in Clampfit 10(Danaher Corporation). For multi-channel recordings, only a singlechannel from each rat was used (for all rats, all responses were simi-lar on all channels in the dentate gyrus). Fiber volley amplitude wasmeasured as the voltage difference between the initial positive peakvoltage after stimulation and the following antipeak voltage; thestimulus artifact occluded the fiber volley in a few recordings, andthus sometimes could not be measured. Fiber volley latency-to-peak-amplitude wasmeasured as the latency from the onset of stimu-lation to the peak positive voltage of the fiber volley. fEPSP rise slopeswere measured via a linear fit to the rising edge of the fEPSP. fEPSPlatency-to-peak-amplitude was taken as the time from the onset ofstimulation to the peak positive voltage of the fEPSP. Populationspike amplitude was calculated from the peak negative voltage ofthe population spike relative to the midpoint voltage interpolated be-tween the peak voltage of the rising phase of the fEPSP and the peakvoltage of the onset of the decaying phase of the fEPSP. Populationspike latency-to-peak-amplitude was measured as the latency fromthe onset of stimulation to the peak of the population spike. Paired-pulse ratios (PPRs) were calculated as the second response dividedby the first response (P2/P1); these calculations were completed forfEPSP rise slopes and fEPSP amplitudes. Fourier analyses to assess thepower of theta frequency during spontaneous recordings (see Fig. 1)were also completed (n=16 rats) in Clampfit 10.
Only recordings for which fEPSP rise slopes and population spikeamplitudes were stable over the 15 min baseline period were usedfor further analyses. All data was divided into 5 min bins and stabilitywas defined in the following way: mean fEPSP rise slope and meanpopulation spike amplitude during each 5 min bin during baselinewere required to be within 15% of the overall mean for all baselinetimepoints, and for recordings that satisfied the first criteria, therecould be no consistent change (increase or decrease) in fEPSP riseslope or population spike amplitude from one bin to the next.
2.8. Toluene vapor delivery
A T-junctionwas inserted into the tubing (0.25 inch inner diameter)that connected the isoflurane and oxygen supply to the nosecone onthe stereotaxic frame, thereby creating two paths for gas flow to thenosecone. Each path was constructed identically and included a sealedreservoir (90 ml capacity) as well as a pair of manually-operatedvalves to direct gas flow exclusively through one path or the other.The reservoir inlet and outlet lines were installed just above the levelof the liquid in the reservoirs. To one reservoir we added 20 mlof water as a control, and to the other reservoir we added 20 ml oftoluene. One path thus provided the rat with isoflurane, and oxygen,and the other path provided the rat with isoflurane, oxygen andtoluene vapor. As a control we filled both reservoirs with water, and
noted no obvious differences in fEPSP characteristics when switchingbetween the two reservoirs (data not shown).
2.9. Gas chromatography
2.9.1. InstrumentationA Varian Saturn CP-3800 gas chromatograph equipped with a
flame ionization detector (FID; Varian, Palo Alto, CA) and a DB5MScapillary column (Agilent Technologies, Santa Clara, CA) was used tomeasure toluene levels in vapor samples from the nosecone of thestereotaxic frame. The gas chromatograph was operated by SaturnGC/MSWorkstation System Control software (v. 5.51). All instrumentsettings were identical to those we have previously reported, whichoptimally discriminate toluene and ortho-xylene peaks at ~5.0 and~8.6 min, respectively (Perit et al., 2012).
2.9.2. Determination of toluene concentration in the noseconeTo measure the concentration of toluene in the nosecone of the
stereotaxic apparatus we first sealed the nosecone with Parafilm.Gas flow (2.5% isoflurane, 550 ml/min oxygen) was directed throughthe path containing the toluene-filled reservoir and into the sealednosecone. Vapor samples were obtained from the nosecone by piercingthe Parafilm wrapping with the needle of a gastight syringe (Hamilton#81317, VWR). For further details see Perit et al. (2012). Air sampleswere also collected immediately adjacent to the sealed nosecone anddid not have detectable levels of toluene, confirming that the noseconewas tightly sealed by the Parafilm (n=6). Gas chromatograms werecompared against standard samples of known concentrations of tolueneand ortho-xylene that we reported previously (Perit et al., 2012).
2.10. Statistical analyses
All statistical analyses were completed with IBM SPSS Statisticsv.19 (IBM, Armonk, NY). Independent samples t-tests were used forthe PPR analysis and to assess age-related effects on populationspike amplitude, paired-samples t-tests were used for the frequencyanalyses, and analyses of variance were used for all other analyses.No systematic differences were noted between male and female ratsfor any of the measures reported with respect to outcomes of tolueneexposure, and thus data for male and female rats has been pooled.Data are presented as mean and standard error of the mean.
3. Results
3.1. Toluene concentration in the nosecone
Toluene vapor (mixed with isoflurane and oxygen) or the controlcondition (isoflurane and oxygen only) was delivered to each ratthrough the nosecone of the stereotaxic apparatus. Vapor sampleswere extracted from the nosecone and submitted for gas chromatog-raphy analysis. By comparison to a previously generated standardcurve of samples consisting of various concentrations of toluene vapor(0 to 10,000 ppm) in ortho-xylene (10,000 ppm) (Perit et al., 2012)the concentration of toluene within the nosecone was ~5000 ppm(M=4700 ppm, SEM=560 ppm; n=10). This vapor concentration iswithin the typical range for human abusers directly inhaling toluenevapor from a number of toluene-containing products (Bukowski,2001). No toluene vapor could be detected in the nosecone when airflow was directed through the path containing the water-filled reser-voir (data not shown).
3.2. Toluene changes the characteristics of PP-DG fEPSPs
A series of experiments (see Fig. 1 for sequence) were completedto assess the effects of toluene vapor inhalation on fEPSPs at PP-DGsynapses. Following at least 1 h of stabilization recording, and then
406 J.M. Gmaz et al. / Neurotoxicology and Teratology 34 (2012) 403–412
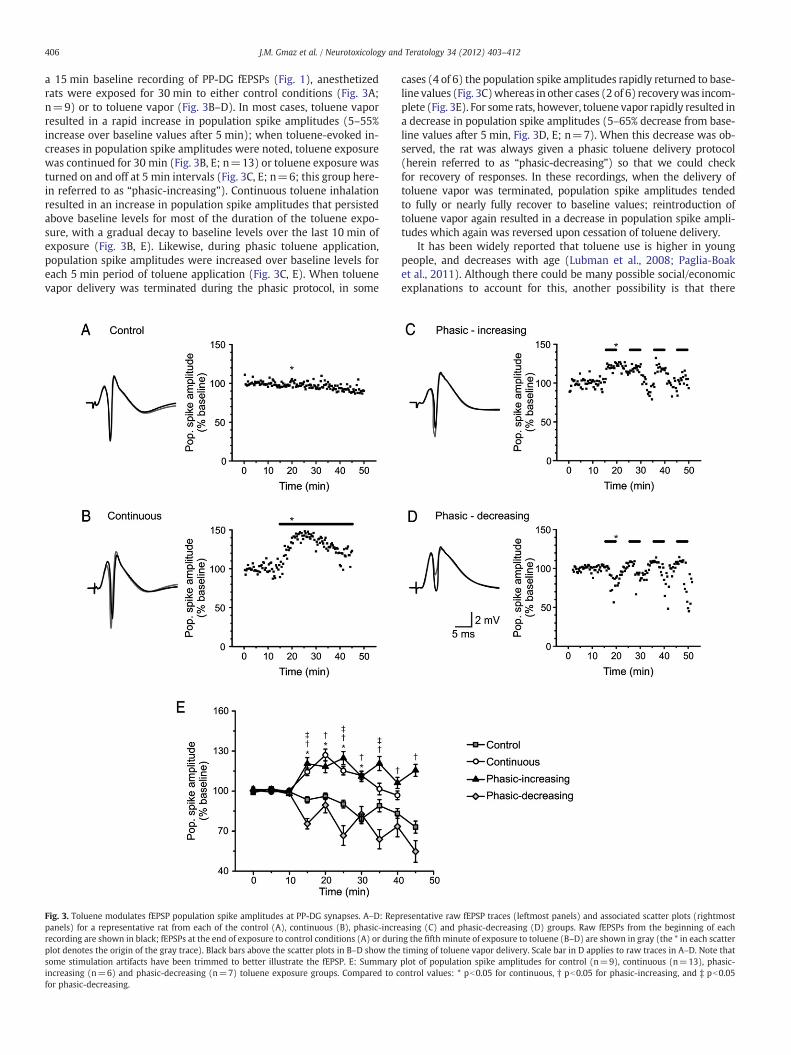
a 15 min baseline recording of PP-DG fEPSPs (Fig. 1), anesthetizedrats were exposed for 30 min to either control conditions (Fig. 3A;n=9) or to toluene vapor (Fig. 3B–D). In most cases, toluene vaporresulted in a rapid increase in population spike amplitudes (5–55%increase over baseline values after 5 min); when toluene-evoked in-creases in population spike amplitudes were noted, toluene exposurewas continued for 30 min (Fig. 3B, E; n=13) or toluene exposure wasturned on and off at 5 min intervals (Fig. 3C, E; n=6; this group here-in referred to as “phasic-increasing”). Continuous toluene inhalationresulted in an increase in population spike amplitudes that persistedabove baseline levels for most of the duration of the toluene expo-sure, with a gradual decay to baseline levels over the last 10 min ofexposure (Fig. 3B, E). Likewise, during phasic toluene application,population spike amplitudes were increased over baseline levels foreach 5 min period of toluene application (Fig. 3C, E). When toluenevapor delivery was terminated during the phasic protocol, in some
Fig. 3. Toluene modulates fEPSP population spike amplitudes at PP-DG synapses. A–D: Reppanels) for a representative rat from each of the control (A), continuous (B), phasic-increrecording are shown in black; fEPSPs at the end of exposure to control conditions (A) or duriplot denotes the origin of the gray trace). Black bars above the scatter plots in B–D show thesome stimulation artifacts have been trimmed to better illustrate the fEPSP. E: Summaryincreasing (n=6) and phasic-decreasing (n=7) toluene exposure groups. Compared to cfor phasic-decreasing.
cases (4 of 6) the population spike amplitudes rapidly returned to base-line values (Fig. 3C)whereas in other cases (2 of 6) recoverywas incom-plete (Fig. 3E). For some rats, however, toluene vapor rapidly resulted ina decrease in population spike amplitudes (5–65% decrease from base-line values after 5 min, Fig. 3D, E; n=7). When this decrease was ob-served, the rat was always given a phasic toluene delivery protocol(herein referred to as “phasic-decreasing”) so that we could checkfor recovery of responses. In these recordings, when the delivery oftoluene vapor was terminated, population spike amplitudes tendedto fully or nearly fully recover to baseline values; reintroduction oftoluene vapor again resulted in a decrease in population spike ampli-tudes which again was reversed upon cessation of toluene delivery.
It has been widely reported that toluene use is higher in youngpeople, and decreases with age (Lubman et al., 2008; Paglia-Boaket al., 2011). Although there could be many possible social/economicexplanations to account for this, another possibility is that there
resentative raw fEPSP traces (leftmost panels) and associated scatter plots (rightmostasing (C) and phasic-decreasing (D) groups. Raw fEPSPs from the beginning of eachng the fifth minute of exposure to toluene (B–D) are shown in gray (the * in each scattertiming of toluene vapor delivery. Scale bar in D applies to raw traces in A–D. Note thatplot of population spike amplitudes for control (n=9), continuous (n=13), phasic-ontrol values: * pb0.05 for continuous, † pb0.05 for phasic-increasing, and ‡ pb0.05

407J.M. Gmaz et al. / Neurotoxicology and Teratology 34 (2012) 403–412
are neurobiological differences in the effects of toluene on youngervs. older brains. We found that the amount of change in populationspike amplitude following toluene administration was negatively cor-related with the age of our rats [r(24)=−0.61, pb0.01]. Specifically,rats whose population spikes increased in the presence of toluenetended to be younger (mean=71 days old; SD=8; n=19) than ratswhose population spikes decreased when toluene vapors were inhaled(mean=86 days old; SD=12; n=7) [t(24)=3.4, pb0.01]. Althoughthe age difference is statistically significant, the variance suggests thatthere is a slow transition between toluene as an enhancer vs. inhibitorof population spike amplitudes.
Fig. 4. Effects of toluene vapor on fiber volley and synaptic transmission characteristics. Summ(A), fiber volley amplitude (B), fiber volley latency-to-peak-amplitude (C), fEPSP rise slopamplitude to fiber volley amplitude (F), and ratio of population spike latency to fiber voincreasing, and ‡ pb0.05 for phasic-decreasing. Legend applies to A–G.
Whereas toluene either increased or decreased population spikeamplitudes, in all cases toluene decreased the latencies to the peaksof the population spikes (Fig. 4A). This combination of results sug-gests that toluene may have dissociable effects on the mechanismsthat may lead to the numbers of units recruited into the populationof granule cells being recorded (population spike amplitudes), andthe excitability of the granule cells in the population (latency forthe population spike to be generated). To rule out the potential con-tribution of toluene-evoked changes in the characteristics of PP fibers,we measured the fiber volley amplitude, an index of the number of PPfibers recruited by electrical stimulation, and fiber volley latency, an
ary plots showing the effects of toluene on population spike latency-to-peak-amplitudee (D), ratio of fEPSP rise slope to fiber volley amplitude (E), ratio of population spikelley latency (G). Compared to control: * pb0.05 for continuous, † pb0.05 for phasic-

408 J.M. Gmaz et al. / Neurotoxicology and Teratology 34 (2012) 403–412
index of the speed of conduction down PP fibers (Lomo, 1971). Wefound that continuous toluene application after manyminutes resultedin small but statistically significant increases in the amplitude of thefiber volley (Fig. 4B). However, phasic toluene application had no sucheffect (Fig. 4B), suggesting that only prolonged toluene applicationchanges the recruitment of PP fibers. Toluene did, however, result ina slight, but statistically significant, decrease in fiber volley latencies,revealing that toluene causes a slight (~5%) increase in the speed ofaction potential conduction in PP axons (Fig. 4C). Additionally weconsidered the fEPSP rise slope, generally taken to be a measure of theefficacy of excitatory synaptic transmission (Johnston and Wu, 2001).We found that toluene vapor inhalation did not consistently changefEPSP rise slope in any of the toluene exposure groups (Fig. 4D), andthat the few changes observed in fEPSP rise slope were nullified whenfEPSP rise slopes were normalized against fiber volley amplitudes(Fig. 4E). In contrast, changes in fiber volley amplitudes could notaccount for toluene-evoked changes in population spike amplitudes(Fig. 4F), providing early evidence for a primarily postsynaptic locus oftoluene's effects. Finally, the decreases in fiber volley latencies couldnot completely account for the decreases in population spike latencies(Fig. 4G). Together these results suggest that toluene-evoked changesin the production of population spikes in DG granule cells are not
Fig. 5. Effects of toluene on input–output relations. A: Change in maximal population spike alation intensity to evoke a population spike. C–F: Representative input–output curves for sin(F) treatment groups. * pb0.05 and ** pb0.01. Legend in C applies to C–F.
explained by changes in the characteristics of PP fibers, or synapticefficacy associated with very low frequency synaptic activation.
3.3. Effects of toluene on input–output relations
It was next determined whether the effects of toluene on popula-tion spike amplitudes were restricted to a specific stimulus intensity(e.g., the intensity that evoked a half-maximal population spike), oroccurred over a broad range of stimulation intensities. Moreover,to assess potential changes in the stimulation threshold to evokepopulation spikes, input–output relations were recorded for each ratby applying incrementally greater stimulus intensities to PP fibersand measuring the resulting amplitudes of population spikes in thegranule cells of the DG (Fig. 5). The input–output relations wereassessed before and during toluene exposure (or during the equiva-lent time periods for toluene-free control recordings; see Fig. 1).Given the variation between rats with respect to the specific stimu-lation intensities (and increments) required to produce a typicalsigmoid-shaped input–output relation, it was not possible to averagetogether recordings from different rats. Fig. 5 thus illustrates mea-sures readily extracted from all input–output relations (maximumpopulation spike amplitude (Fig. 5A) and stimulation threshold to
mplitudes evoked at the highest stimulation intensities. B: Change in minimum stimu-gle rats in the control (C), phasic-increasing (D), phasic-decreasing (E) and continuous

409J.M. Gmaz et al. / Neurotoxicology and Teratology 34 (2012) 403–412
elicit population spikes (Fig. 5B), and representative input–outputrelations for a single rat in each treatment group (Fig. 5C–F).
Phasic-increasing and phasic-decreasing recordings both showedchanges in population spike amplitudes (increases and decreases, re-spectively) across a broad range of stimulus intensities (Fig. 5A, D, E).However, compared to the phasic-increasing recordings, phasic-decreasing recordings underwent a marked increase in the thresholdintensity of stimulation to elicit population spikes (Fig. 5B, D, E).In phasic-decreasing recordings, increasing the stimulation intensitywell beyond the maximal intensity used during the pre-tolueneperiod was insufficient in any recording to evoke a populationspike equivalent in amplitude to the maximum recorded in the pre-toluene period (data not shown). In contrast to phasic toluene appli-cation, there was little difference in the input–output relations afterrats had been exposed to toluene continuously (Fig. 5A, B, F). Thislatter result again suggests that there may be cumulative effectsto prolonged toluene inhalation distinct from that observed duringphasic toluene inhalation.
3.4. Effects of toluene on paired-pulse ratios
To identify any potential contribution of presynaptic mechanismsto the effects of toluene at PP-DG synapses, a series of paired-pulse in-terval stimulation protocols were used. Paired-pulses were appliedwith inter-pulse intervals ranging from 20 through 5000 ms. Forthese experiments the stimulator was set at an intensity sufficientto produce a robust fEPSP, but below the threshold to generate apopulation spike. Paired-pulse ratios were first calculated by dividingthe characteristic of interest (e.g., fEPSP amplitude) for the secondpulse (P2) by the same characteristic measured from the first pulse(P1) (e.g., P2/P1). Synapses showed weak paired-pulse facilitationfor 20 and 50 ms inter-pulse intervals; at intervals greater than50 ms fEPSP slopes and amplitudes evoked by the second pulse wereequivalent to the first pulse. To compare paired-pulse ratios beforeand during toluene inhalation, we computed the ratio (P2/P1)Toluene/(P2/P1)Baseline (Fig. 6). This calculation did not differ among any ofthe toluene exposure groups (phasic-increasing, phasic-decreasing,
Fig. 6. Toluene does not alter paired-pulse ratios. A: Representative recordings ofPP-evoked fEPSPs elicited by paired-pulse stimulation (shown here for 20 ms ISI)before and during toluene administration. B: During-toluene paired-pulse ratios relativeto before-toluene paired-pulse ratios (PPRToluene/PPRBaseline) for control (white bars;n=9) and toluene-treated (gray bars; n=26) rats for fEPSP amplitudes.
and continuous exposure), and therefore these groups were collapsedinto a single ‘toluene group’ (n=26; Fig. 6) and compared againstcontrol (n=9; no toluene) recordings. For the fEPSP characteristicsmeasured, including fEPSP amplitude (Fig. 6A, B) and fEPSP rise slope(data not shown), there were no consistent statistically significantdifferences produced by toluene inhalation.
We also examined these same paired pulse inter-pulse intervalswith stimuli of sufficient intensity to evoke a population spike. Ata 20 ms inter-pulse interval, there was a robust paired-pulse depres-sion, in that population spikes were only evoked by the first stimuluspulse. At inter-pulse intervals of 100 ms and longer, population spikeswere of equivalent size for the first and second pulses. This pattern offindings was apparent during baseline recordings and during tolueneexposure (data not shown) suggesting that toluene did not changethe characteristics of repetitive synchronous action potential outputfrom these synapses and cells. Together, these results suggest thatany effect of toluene on synaptic transmission at PP-DG synapses, orgranule cell output, is not likely to be mediated through a presynapticmechanism.
3.5. Effects of toluene on hippocampal oscillations
Theta frequency oscillations contribute importantly to hippocampal-mediated learning tasks, many of which have been shown to be im-paired by toluene inhalation (Bowen et al., 2006; Buzsaki, 2002). Thusas a final measure of the effects of toluene on DG physiology, thepower of spontaneous theta frequency oscillation was measured in asubset of control rats and toluene-treated rats that showed an increasein population spike amplitude. Theta power during baseline recordingswas 0.003 and 0.005 mV2/Hz for control- and toluene-treated rats, re-spectively [t(14)=2.0, ns]. We found that toluene vapor inhalation(n=8) significantly reduced power in the 8–13 Hz frequency band(Fig. 7B, C). No differences in the power of the theta frequency wereseen in control-exposed rats (n=8) over the equivalent period of time(Fig. 7A, C).
4. Discussion
4.1. Effects of toluene on granule cells
The present study was conducted to assess the effects of an abuse-relevant concentration of toluene vapor (~5000 ppm) on granule celloutput and synaptic transmission at PP-DG synapses in the anesthe-tized rat hippocampus in vivo. In the majority of our experiments,a toluene-induced increase in population spike amplitude that wasat least partially, and often fully, reversible upon brief washout oftoluene was observed. This potentiation in population spike ampli-tude occurred alongside a reduction of population spike latencyand could not be accounted for by any increase in synaptic drive.Moreover, the increase in population spike amplitude was nearlyinstantaneous, whereas the small enhancement of fEPSP rise slopewas gradual and only apparent in the continuous toluene exposuregroup. Further, a wide range of stimulus intensities was used in thepresent study (from suprathreshold to produce a fEPSP, to sufficientto evoke a robust population spike), and toluene vapor inhalationchanged evoked output from granule cells across this entire range.This suggests that many strengths of afferent input, and thereforeamounts or complexity of information from the entorhinal cortex,may be affected by toluene inhalation. Together, these results suggestthat toluene increased the amount of synchronous granule cell dis-charge in response to afferent stimulation from the PP, possibly throughenhanced granule cell excitability, or by an alteration in the relativeweighting of excitatory and inhibitory inputs (Andersen et al., 1971;Bliss and Lomo, 1973).
Interestingly, a subset of rats showed a decrease in populationspike amplitude. Within this subset of experiments there was also a

Fig. 7. Toluene decreased the power of the theta rhythm in the DG. A–B: Representative power spectrums from a rat in the control (A) and toluene-treated (B) groups. Baselinerecording is represented by black circles and the period after toluene exposure, or an equivalent period of time for controls, is represented by white circles. C: Power of thetheta rhythm (8–13 Hz), relative to baseline, in control (n=8) and toluene-treated (n=8) rats. ** pb0.01.
410 J.M. Gmaz et al. / Neurotoxicology and Teratology 34 (2012) 403–412
decrease in the population spike/fEPSP rise slope ratio, and the stim-ulation intensity required to evoke a population spike was higher,compared to the phasic-increasing group. Our analyses revealedthat these rats were on average about two weeks older than thoserats whose population spikes increased during the inhalation of tolu-ene vapors, although it is important to note that there was substantialoverlap in the distributions of the ages of each of these groups. Inhumans, inhalation of toluene-containing products tends to be mostprominent in youth (Lubman et al., 2008; Paglia-Boak et al., 2011),and our results suggest that there may be important neurobiologicaldifferences as age progresses that lead to differential effects of tolu-ene. Rigorous developmental studies of the neurobiological effectsof toluene are clearly warranted.
The phasic-decreasing recordings may be explained by differencesin feed-forward inhibition (Buzsaki and Eidelberg, 1982; Wilson et al.,1981). Glutamatergic afferents from the PP to the DG project to DGgranule cells and basket cells, which then form inhibitory synapses onthe granule cells (Hjorth-Simonsen and Jeune, 1972). The basket cellinterneurons have a lower threshold for action potential discharge andthus fire before the appearance of a population spike in extracellularrecordings, suggesting that they could exert their inhibitory influenceon granule cells during the evoked response (Buzsaki and Eidelberg,1982). Since toluene is known to enhance GABA-mediated currentsin various preparations (Bale et al., 2005b; Beckley and Woodward,2011; Beckstead et al., 2000; MacIver, 2009), the biphasic effects oftoluene on population spike amplitude could be due to variationsbetween experiments in the relative numbers of granule and basketcells recruited during PP stimulation. Because stimulating electrodeswere in the medial, and not lateral, PP in the small sample of ratsassessed histologically, methodological differences related to electrodeplacement may not explain the noted biphasic effects (McNaughtonand Barnes, 1977), although it is possible that unresolvable differencesin the distribution of recording electrodes along the mediolateralaxis in the granule cell layermight account for some of the observed dif-ferences. Furthermore, since the gas chromatographywork showed thatthe concentration of toluene in the nosecone was always ~5000 ppm,variability in the inhaled dose of toluene (Ikeuchi et al., 1993) is notlikely to account for the biphasic effects of toluene.
4.2. Effects of toluene on perforant path axons and synapses
Whereas the principal effects reported in the present study revealeda probable postsynaptic effect of toluene on granule cells, we uncovered
some evidence suggesting toluene-evoked changes in the characteris-tics of PP fibers as well. First, toluene vapor inhalation increased fibervolley amplitude in a gradual and persistentmanner, implying a greaterrecruitment of PP fibers after prolonged exposures (Lomo, 1971). Thisdata is in agreement with a previous finding in guinea pig hippocampalslices, where it was shown that bath application of 20 μg/ml toluene onhippocampal slices produces an enhancement of the PP-evoked fibervolley (Ikeuchi et al., 1993). Second, toluene decreased the latency ofthe fiber volley to arrive at the recording electrode, suggesting an in-crease in the velocity of conduction of action potentials down PPaxons. Additionally, changes observed in fEPSP rise slopes, which is acommon indication of change in synaptic efficacy (Johnston and Wu,2001) could be accounted for by the above-noted changes in PP activity.However, we did measure a gradual decrease in the fEPSP rise slope/fiber volley ratio after prolonged toluene exposure, suggesting adelayed synaptic effect independent of the effects of toluene onPP axons. Beckley and Woodward (2011) have recently shown thattoluene dose-dependently decreases AMPA-mediated currents via theendocannabinoid system, a residual effect that only becomes apparentafter long periods. This may explain why a gradual decrease in fEPSPslopes was observed in rats who received continuous, but not phasic,exposure to toluene. Overall, however, contributions of presynapticeffects to our results may be minimal, given that no changes in thepaired-pulse ratio for fEPSP amplitudes were observed.
4.3. Effects of toluene on the theta rhythm
Exposure to toluene decreased the power of 8 to13 Hz oscillations,suggesting that toluene decreased the power of the theta rhythm(Buzsaki, 2002). Cholinergic and GABAergic inputs from the medialseptal nucleus act as rhythm generators for hippocampal theta, anddisruption of the septohippocampal pathway decreases or abolishesthe theta rhythm (Freund and Antal, 1988). Notably, both cholinergicand GABAergic signaling are disrupted by toluene exposure (Baleet al., 2002, 2005a, 2005b; Beckley and Woodward, 2011; Becksteadet al., 2000; MacIver, 2009). Additionally, Perit et al. (2012) observeda robust increase in c-Fos immunoreactivity in the septohippocampalnucleus during toluene exposure, suggesting one possible anatomicallocation for toluene's disruptive effects on hippocampal theta. Inter-estingly, stimulation of the medial septum alongside the PP results inan enhancement of the PP-evoked population spike without affectingthe initial rise slope of the fEPSP (Fantie and Goddard, 1982). This fa-cilitation is the result of GABAergic projections from the septum that

411J.M. Gmaz et al. / Neurotoxicology and Teratology 34 (2012) 403–412
terminate on DG basket cells, resulting in an increase in the excitabilityof granule cells due to a decrease in inhibition (Bilkey and Goddard,1985; Freund and Antal, 1988). Given that toluene disrupted thetheta rhythm, it is thus possible that the effects observed on granulecell excitability could in part be due to alterations in inhibitory inputfrom the medial septal area.
4.4. Comparison of toluene effects to other psychoactive drugs
Toluene is often likened to central nervous system depressantsthat enhance GABAergic activity, such as alcohol and benzodiazepines(Bowen et al., 2006). However, in contrast to our present results, in theanesthetized rat, intraperitoneal injections of ethanol or diazepam de-creased population spike amplitudes in the DG and increased paired-pulse inhibition (Albertson and Joy, 1989; Steffensen and Henriksen,1992). Toluene is also known to enhance serotonergic currents(Lopreato et al., 2003). In the anesthetized rat in vivo preparation, intra-peritoneal injections of the serotonin precursor 5-hydroxytryptophanincreased PP-evoked population spikes in the DG, similar to the major-ity of results in the current study (Klancnik et al., 1989). Intraperitoneallow dose nicotine injections facilitate synaptic transmission at manytypes of synapses, and result in marked increases in the amplitudeof population spikes in the DG (Matsuyama et al., 2000; McKay et al.,2007). Although superficially similar to the present observations,these results are incompatible with the known nicotinic acetylcholinereceptor inhibiting properties of toluene (Bale et al., 2002, 2005a),suggesting that our observed results are not likely working through anAChR-mediated mechanism. Finally, amphetamine has been shownto increase fEPSP rise slope in the DG without affecting populationspike amplitude in vivo (Morimoto et al., 1987). Thus it would appearthat toluene may work through unique mechanisms in the DG in com-parison to other psychoactive drugs.
4.5. Neurobiology of toluene effects
One important question not addressed by the present study is theidentification of the specific molecular target (or targets) affected bytoluene that underlie the reported effects. When rats are exposed to~5000 ppm toluene through the nosecone on our stereotaxic frame,a brain concentration of toluene of ~830 μM (SD=205 μM; n=19)is achieved (Gmaz, Matthews and McKay, unpublished observations).The brain concentration of toluene rises very rapidly, and there are nodifferences in brain concentrations between rats exposed phasicallyor continuously to toluene. Studies done primarily in heterologousexpression systems have identified a number of ion channels andneurotransmitter receptors that may be activated or inhibited bytoluene at approximately this brain concentration, including NMDA,AMPA, GABAA, glycine, 5HT3, and nicotinic acetylcholine receptors,Na+, Ca2+, and K+ channels, and internal calcium stores (Bale et al.,2002; Beckley and Woodward, 2011; Beckstead et al., 2000; Cruzet al., 1998, 2003; Del Re et al., 2006; Lopreato et al., 2003; MacIver,2009; Shafer et al., 2005; Tillar et al., 2002). For these neurotransmitterreceptors and ion channels, those typically associated with inhibitionare activated by toluene, and those typically associated with excitationare inhibited by toluene. This provides a challenge to incorporateone of these mechanisms into an explanation of our observed effectswhich by-and-large show that toluene increases output from DGgranule cells; a previous report has also demonstrated that toluenehad a net excitatory effect on neuronal output, in that case from dopa-mine neurons in the ventral tegmental area (Riegel et al., 2007). It isalso important to note that the time course of the effects of toluenein the current experiments differed for population spikes (rapid) andPP fiber volleys (slow), and thus there may be several mechanismswhich account for specific aspects of the observed effects. Determiningwhich mechanismmay account for the present results will be of signif-icant future interest.
4.6. Caveats of isoflurane anesthesia
The present study was completed in the rat DG in vivo, providingthe benefits of a synaptically intact system. However, rats were anes-thetized with isoflurane, and it is important to acknowledge thatisoflurane itself produces some changes in the physiology of neuronsand synapses. For instance, isoflurane depresses glutamate releaseand inhibits NMDA-mediated currents, and decreases the amplitudeand conduction velocity of presynaptic compound potentials (Berg-Johnsen and Langmoen, 1986; MacIver et al., 1996; Nishikawa andMacIver, 2000). Isoflurane has further been shown to decrease popula-tion spike amplitude and increase latencywhile minimally affecting thefEPSP at PP-DG synapses in vitro (MacIver and Roth, 1988). Isofluraneadditionally increases paired-pulse depression and paired-pulse fa-cilitation of population spikes and fEPSPs, respectively, in the CA1(Nishikawa and MacIver, 2000; Pearce et al., 1989). Therefore thepotential exists that our report of toluene-evoked increases in fibervolley amplitudes and conduction velocities may be an underestimateof the true increases in these parameters, given the influence of theanesthetic. Importantly, theta rhythm, which is normally presentduring voluntary motor activity and rapid eye movement sleep, ispresent with the same power, but is slightly slowed, under isofluraneanesthesia (Perouansky et al., 2010). The neurophysiological effects oftoluene inhalation in awake and behaving ratswill be required to assessthe potential significance of these anesthetic effects.
4.7. Functional and behavioral implications
The hippocampus forms distinct representations of unique envi-ronments, and the DG, specifically, is responsible for distinguishingbetween similar environments via pattern separation (Leutgeb et al.,2007). In any given environment, a small fraction (2–5%) of the DGgranule cell population is activated, whereas up to half of the CApyramidal cells are recruited (Schmidt et al., 2012); connectionsformed between the dorsal DG and the CA3 are sparse, but powerfullyexcitatory. The current study showed an alteration in granule cellexcitability in response to toluene exposure (typically an increase inaction potential discharge), which has implications for pattern sepa-ration. By increasing the number of granule cells that fire in responseto PP stimulation, the CA3 may receive a larger input from the DG,resulting in impairments in distinguishing among similar environmentsor differing aspects of the same environment. In support of this, in acontextual fear conditioning task which requires distinguishing a fear-associated environment from a safe environment, pretreatment withtoluene impaired freezing when tested 24 h later, suggesting impairedability to distinguish these environments (Seo et al., 2010). Addition-ally, toluene reduces the preference for a novel object compared to afamiliar object in a hippocampal-dependent object recognition task(Broadbent et al., 2004; Seo et al., 2010). Taken together, our electro-physiological results in the DG, with implications for pattern separationand the processing of environmental information by the hippocampus,may provide a novel explanation for the behavioral impairments associ-ated with toluene intoxication.
Conflict of interest statement
There are no conflicts of interest.
Acknowledgments
This research was supported by a Discovery Grant from the NaturalSciences and Engineering Research Council of Canada (NSERC) (B.E.M.).J.M.G. was supported by an Ontario Graduate Scholarship (OGS) anda Faculty of Science Student Association (FOSSA) grant from WilfridLaurier University. B.A.M. was supported by an Undergraduate Student

412 J.M. Gmaz et al. / Neurotoxicology and Teratology 34 (2012) 403–412
Research Award from NSERC. The authors thank Ms. Gena Braun fortechnical assistance with gas chromatography.
References
Albertson TE, Joy RM. Modification of evoked hippocampal dentate inhibition bydiazepam and three antagonists in urethane-anesthetized rats. Exp Neurol 1989;106:142–9.
Andersen P, Bliss TV, Skrede KK. Unit analysis of hippocampal population spikes. ExpBrain Res 1971;13:208–21.
Andersen P, Morris RG, Amaral D, Bliss TV, O'Keefe J. The hippocampus book; OxfordUniversity Press, Inc, New York: Oxford; 2007.
Bale AS, Smothers CT, Woodward JJ. Inhibition of neuronal nicotinic acetylcholinereceptors by the abused solvent, toluene. Br J Pharmacol 2002;137:375–83.
Bale AS, Meacham CA, Benignus VA, Bushnell PJ, Shafer TJ. Volatile organic compoundsinhibit human and rat neuronal nicotinic acetylcholine receptors expressed inXenopus oocytes. Toxicol Appl Pharmacol 2005a;205:77–88.
Bale AS, Tu Y, Carpenter-Hyland EP, Chandler LJ, Woodward JJ. Alterations inglutamatergic and gabaergic ion channel activity in hippocampal neurons followingexposure to the abused inhalant toluene. Neuroscience 2005b;130:197–206.
Baydas G, Ozveren F, Tuzcu M, Yasar A. Effects of thinner exposure on the expressionpattern of neural cell adhesion molecules, level of lipid peroxidation in the brainand cognitive function in rats. Eur J Pharmacol 2005;512:181–7.
Beckley JT, Woodward JJ. The abused inhalant toluene differentially modulates excitatoryand inhibitory synaptic transmission in deep-layer neurons of the medial prefrontalcortex. Neuropsychopharmacology 2011;36:1531–42.
Beckstead MJ, Weiner JL, Eger II EI, Gong DH, Mihic SJ. Glycine and gamma-aminobutyricacid(A) receptor function is enhanced by inhaled drugs of abuse. Mol Pharmacol2000;57:1199–205.
Berg-Johnsen J, Langmoen IA. The effect of isoflurane on unmyelinated and myelinatedfibres in the rat brain. Acta Physiol Scand 1986;127:87–93.
Bilkey DK, Goddard GV. Medial septal facilitation of hippocampal granule cell activity ismediated by inhibition of inhibitory interneurons. Brain Res 1985;361:99-106.
Bliss TV, Lomo T. Long-lasting potentiation of synaptic transmission in the dentate areaof the anaesthetized rabbit following stimulation of the perforant path. J Physiol1973;232:331–56.
Bowen SE, Batis JC, Paez-Martinez N, Cruz SL. The last decade of solvent research inanimal models of abuse: mechanistic and behavioral studies. Neurotoxicol Teratol2006;28:636–47.
Broadbent NJ, Squire LR, Clark RE. Spatial memory, recognition memory, and thehippocampus. Proc Natl Acad Sci U S A 2004;101:14515–20.
Bukowski JA. Review of the epidemiological evidence relating toluene to reproductiveoutcomes. Regul Toxicol Pharmacol 2001;33:147–56.
Buzsaki G. Theta oscillations in the hippocampus. Neuron 2002;33:325–40.Buzsaki G, Eidelberg E. Direct afferent excitation and long-term potentiation of hippo-
campal interneurons. J Neurophysiol 1982;48:597–607.Cruz SL, Mirshahi T, Thomas B, Balster RL, Woodward JJ. Effects of the abused solvent
toluene on recombinant N-methyl-D-aspartate and non-N-methyl-D-aspartatereceptors expressed in Xenopus oocytes. J Pharmacol Exp Ther 1998;286:334–40.
Cruz SL, Orta-Salazar G, Gauthereau MY, Millan-Perez Pena L, Salinas-Stefanon EM.Inhibition of cardiac sodium currents by toluene exposure. Br J Pharmacol 2003;140:653–60.
Del Re AM, Dopico AM, Woodward JJ. Effects of the abused inhalant toluene onethanol-sensitive potassium channels expressed in oocytes. Brain Res 2006;1087:75–82.
Fantie BD, Goddard GV. Septal modulation of the population spike in the fascia dentataproduced by perforant path stimulation in the rat. Brain Res 1982;252:227–37.
Filley CM, Halliday W, Kleinschmidt-DeMasters BK. The effects of toluene on thecentral nervous system. J Neuropathol Exp Neurol 2004;63:1-12.
Freund TF, Antal M. GABA-containing neurons in the septum control inhibitory inter-neurons in the hippocampus. Nature 1988;336:170–3.
Gerfen CR. Basic neuroanatomical methods. Current protocols in neuroscience. JohnWiley & Sons, Inc; 1997. pp. 1..-..11.
Gotohda T, Tokunaga I, Kubo S, Kitamura O, Ishigami A. Toluene inhalation induces glialcell line-derived neurotrophic factor, transforming growth factor and tumor necrosisfactor in rat cerebellum. Leg Med (Tokyo) 2002;4:21–8.
Hjorth-Simonsen A, Jeune B. Origin and termination of the hippocampal perforant pathin the rat studied by silver impregnation. J Comp Neurol 1972;144:215–32.
Honma T, Suda M. Brain microdialysis study of the effects of hazardous chemicalson the central nervous system 2. Toluene exposure and cerebral acetylcholine.Ind Health 2004;42:336–47.
Ikeuchi Y, Hirai H, Okada Y, Mio T, Matsuda T. Excitatory and inhibitory effects oftoluene on neural activity in guinea pig hippocampal slices. Neurosci Lett 1993;158:63–6.
Johnston D, Wu SM. Foundations of cellular neurophysiology. Cambridge: The MITPress; 2001. p. 676.
Kanter M. Nigella sativa and derived thymoquinone prevents hippocampal neu-rodegeneration after chronic toluene exposure in rats. Neurochem Res 2008;33:579–88.
Klancnik JM, Baimbridge KG, Phillips AG. Increased population spike amplitude inthe dentate gyrus following systemic administration of 5-hydroxytryptophan or8-hydroxy-2-(di-n-propylamino)tetralin. Brain Res 1989;505:145–8.
Leutgeb JK, Leutgeb S, Moser MB, Moser EI. Pattern separation in the dentate gyrus andCA3 of the hippocampus. Science 2007;315:961–6.
Lomo T. Patterns of activation in a monosynaptic cortical pathway: the perforant pathinput to the dentate area of the hippocampal formation. Exp Brain Res 1971;12:18–45.
Lopreato GF, Phelan R, Borghese CM, Beckstead MJ, Mihic SJ. Inhaled drugs of abuseenhance serotonin-3 receptor function. Drug Alcohol Depend 2003;70:11–5.
Lubman DI, Yucel M, Lawrence AJ. Inhalant abuse among adolescents: neurobiologicalconsiderations. Br J Pharmacol 2008;154:316–26.
MacIver MB. Abused inhalants enhance GABA-mediated synaptic inhibition. Neuro-psychopharmacology 2009;34:2296–304.
MacIver MB, Roth SH. Inhalation anaesthetics exhibit pathway-specific and differentialactions on hippocampal synaptic responses in vitro. Br J Anaesth 1988;60:680–91.
MacIver MB, Mikulec AA, Amagasu SM, Monroe FA. Volatile anesthetics depress gluta-mate transmission via presynaptic actions. Anesthesiology 1996;85:823–34.
Matsuyama S, Matsumoto A, Enomoto T, Nishizaki T. Activation of nicotinic acetylcho-line receptors induces long-term potentiation in vivo in the intact mouse dentategyrus. Eur J Neurosci 2000;12:3741–7.
McDonald G. First Nations and Inuit Regional Health Survey. Ottawa: First Nations andInuit Regional Health Survey National Steering Committee; 1999.
McKay BE, Placzek AN, Dani JA. Regulation of synaptic transmission and plasticity byneuronal nicotinic acetylcholine receptors. Biochem Pharmacol 2007;74:1120–33.
McNaughton BL, Barnes CA. Physiological identification and analysis of dentate granulecell responses to stimulation of the medial and lateral perforant pathways in therat. J Comp Neurol 1977;175:439–54.
Morimoto K, Otani S, Goddard GV. Effects of acute and long-term treatment withamphetamine on evoked responses and long-term potentiation in the dentategyrus of anesthetized rats. Brain Res 1987;407:137–43.
Nishikawa K,MacIver MB. Excitatory synaptic transmissionmediated by NMDA receptorsis more sensitive to isoflurane than are non-NMDA receptor-mediated responses.Anesthesiology 2000;92:228–36.
Paglia-Boak A, Adlaf EM, Mann RE. Drug use among Ontario students 1977–2011.Toronto: Centre for Addiction and Mental Health; 2011.
Paxinos G, Watson C. The rat brain in stereotaxic coordinates. San Diego: ElsevierAcademic Press; 2005.
Pearce RA, Stringer JL, Lothman EW. Effect of volatile anesthetics on synaptic transmis-sion in the rat hippocampus. Anesthesiology 1989;71:591–8.
Perit KE, Gmaz JM, Browne JDC, Matthews BA, Dunn MBF, Yang L, et al. Distributionof c-Fos immunoreactivity in the rat brain following abuse-like toluene vapor inha-lation. Neurotoxicol Teratol 2012;34:37–46.
Perouansky M, Rau V, Ford T, Oh SI, Perkins M, Eger II EI, et al. Slowing of the hippo-campal theta rhythm correlates with anesthetic-induced amnesia. Anesthesiology2010;113:1299–309.
Perrine SA, O'Leary-Moore SK, Galloway MP, Hannigan JH, Bowen SE. Binge tolueneexposure alters glutamate, glutamine and GABA in the adolescent rat brain asmeasured by proton magnetic resonance spectroscopy. Drug Alcohol Depend2011;115:101–6.
Riegel AC, Zapata A, Shippenberg TS, French ED. The abused inhalant toluene increasesdopamine release in the nucleus accumbens by directly stimulating ventral tegmentalarea neurons. Neuropsychopharmacology 2007;32:1558–69.
Rosenberg NL, Grigsby J, Dreisbach J, Busenbark D, Grigsby P. Neuropsychologic impair-ment and MRI abnormalities associated with chronic solvent abuse. J Toxicol ClinToxicol 2002;40:21–34.
Schmidt B, Marrone DF, Markus EJ. Disambiguating the similar: the dentate gyrus andpattern separation. Behav Brain Res 2012;226:56–65.
Seo HS, Yang M, Song MS, Kim JS, Kim SH, Kim JC, et al. Toluene inhibits hippocampalneurogenesis in adult mice. Pharmacol Biochem Behav 2010;94:588–94.
Shafer TJ, Bushnell PJ, Benignus VA, Woodward JJ. Perturbation of voltage-sensitive Ca2+ channel function by volatile organic solvents. J Pharmacol Exp Ther 2005;315:1109–18.
Steffensen SC, Henriksen SJ. Comparison of the effects of ethanol and chlordiazepoxide onelectrophysiological activity in the fascia dentata and hippocampus regio superior.Hippocampus 1992;2:201–11.
Tillar R, Shafer TJ, Woodward JJ. Toluene inhibits voltage-sensitive calcium channelsexpressed in pheochromocytoma cells. Neurochem Int 2002;41:391–7.
von Euler M, Pham TM, Hillefors M, Bjelke B, Henriksson B, von Euler G. Inhalation oflow concentrations of toluene induces persistent effects on a learning retentiontask, beam-walk performance, and cerebrocortical size in the rat. Exp Neurol2000;163:1–8.
Wilson RC, Levy WB, Steward O. Changes in translation of synaptic excitation to den-tate granule cell discharge accompanying long-term potentiation. II. An evaluationof mechanisms utilizing dentate gyrus dually innervated by surviving ipsilateraland sprouted crossed temporodentate inputs. J Neurophysiol 1981;46:339–55.
Win-Shwe TT, Fujimaki H. Acute administration of toluene affects memory retentionin novel object recognition test and memory function-related gene expression inmice. J Appl Toxicol 2012;32:300–4.
Win-Shwe TT, Mitsushima D, Nakajima D, Ahmed S, Yamamoto S, Tsukahara S, et al.Toluene induces rapid and reversible rise of hippocampal glutamate and taurineneurotransmitter levels in mice. Toxicol Lett 2007;168:75–82.
Win-shwe TT, Hojo R, Mitsushima D, Nakajima D, Yamamoto S, Fujimaki H. Establishmentof a mouse model to assess brain neurotransmitter level and learning performancesimultaneously following toxic chemical exposure: using in vivo microdialysis andschedule-controlled operant behavior. J UOEH 2009;31:1-11.




















