
General Anesthetic and Specific Effects of Ethanol on Acetylcholine Receptors

Visualization and Trafficking of the Vesicular AcetylcholineTransporter in Living Cholinergic Cells
Magda S. Santos, *Jose´ Barbosa Jr., *Christopher Kushmerick, *Marcus V. Gomez,Vania F. Prado, and *Marco A. M. Prado
Departamento de Bioquı´mica-Imunologia and*Laboratorio de Neurofarmacologia, Departamento de Farmacologia, ICB,Universidade Federal de Minas Gerais, Belo Horizonte, Minas Gerais, Brazil
Abstract: The present experiments investigated the traf-ficking of the vesicular acetylcholine transporter (VAChT)tagged with the enhanced green fluorescent protein(EGFP) in living cholinergic cells (SN56). The EGFP–VAChT chimera was located in endosomal-like compart-ments in the soma of SN56 cells, and it was also targetedto varicosities of neurites. In contrast, EGFP alone in cellswas soluble in the cytoplasm. The C-terminal cytoplas-mic tail of VAChT has been implicated in targeting ofVAChT to synaptic vesicles; thus, we have examined therole of the C-terminal region in the trafficking to varicos-ities. A C-terminal fragment tagged with EGFP appearedto be selectively accumulated in varicosities when ex-pressed in SN56 cells. Interestingly, the protein was notfreely soluble in the cytosol, and it presented a punctatepattern of expression. However, EGFP–C terminus didnot present this peculiar pattern of expression in a non-neuronal cell line (HEK 293). Moreover, the C-terminalregion of VAChT did not seem to be essential for VAChTtrafficking, as a construct that lacks the C-terminal tailwas, similar to EGFP–VAChT, partially targeted to endo-cytic organelles in the soma and sorted to varicosities.These experiments visualize VAChT for the first time inliving cells and suggest that there might be multiple sig-nals that participate in trafficking of VAChT to sites ofsynaptic vesicle accumulation. Key Words: Vesicularacetylcholine transporter—Synaptic vesicle—Endocyto-sis—Cholinergic neurons—Vesicular trafficking—Synap-tic proteins.J. Neurochem. 74, 2425–2435 (2000).
Neurons are highly polarized cells, and as such theyhave to make complex decisions regarding the locationof their proteins. However, compared with the morestudied targeting of proteins to basolateral and apicalmembranes in polarized epithelial cells (for review, seeMellman, 1996), there is little information about motifsresponsible for directing synaptic proteins through theextensive pathways of endomembranes in neurons.
Studies of synaptic protein trafficking have shown thatsynaptophysin (Syp), and possibly other synaptic vesicleproteins, pass through the constitutive secretory pathwayto reach nerve terminals (Re´gnier-Vigouroux et al.,
1991). These proteins seem to travel through axons intubulovesicular organelles that may represent synapticvesicle precursors (Nakata et al., 1998). Vesicle-associ-ated membrane protein (VAMP)/synaptobrevin has beensuggested to have distinct targeting motifs: one thatdirects the protein to varicosity sites in primary culturedneurons (West et al., 1997), and another motif that par-ticipates in targeting to synaptic-like vesicles in PC12cells (Grote et al., 1995; Grote and Kelly, 1996). How-ever, information on targeting of other proteins involvedin synaptic vesicle function is lacking. The trafficking ofvesicular acetylcholine transporter (VAChT) and the ve-sicular monoamine transporters (VMATs) 1 and 2 hasbeen a subject of recent interest, because these homolo-gous proteins present distinct localization in PC12 cells(Weihe et al., 1996; Liu and Edwards, 1997; Tao-Chengand Eiden, 1998; Varoqui and Erickson, 1998). In thismodel system, VAChT appears predominantly in smallsynaptic-like vesicles, whereas VMATs are preferen-tially targeted to large dense core vesicles (Weihe et al.,1996; Liu and Edwards, 1997; Tao-Cheng and Eiden,1998; Varoqui and Erickson, 1998). Two distinct lines ofevidence indicate that the cytoplasmic C-terminal re-gions of these proteins may contribute to trafficking.First, exchange of the C-terminal tail of VAChT by thatof VMAT2 changes the localization of the latter in PC12cells, and as a consequence, VMAT2 appears in smallvesicles (Varoqui and Erickson, 1998). Second, bothproteins possess classic intracellular trafficking motifs in
Received December 15, 1999; revised manuscript received February10, 2000; accepted February 14, 2000.
Address correspondence and reprint requests to Dr. M. A. M. Pradoat Laborato´rio de Neurofarmacologia, Departamento de Farmacologia,ICB, Universidade Federal de Minas Gerais, Av. Antoˆnio Carlos, 6627,31270-901, Belo Horizonte, MG, Brazil. E-mail: [email protected]
Abbreviations used:EGFP, enhanced green fluorescent protein;EGFP–C terminus, EGFP-tagged C-terminal region of VAChT;EGFP–12TM, EGFP-tagged VAChT truncated after 12th membranedomain; HBSS, HEPES-buffered salt solution; Syp, synaptophysin;VAChT, vesicular acetylcholine transporter; VAMP, vesicle-associatedmembrane protein; VMAT, vesicular monoamine transporter.
2425
Journal of NeurochemistryLippincott Williams & Wilkins, Inc., Philadelphia© 2000 International Society for Neurochemistry

their C termini that may function in internalization (Leu-Leu for VAChT and Iso-Leu for VMAT2) (Tan et al.,1998). Adaptor protein complexes, which participate incoated vesicle formation, have been shown to interactwith this type of short cytoplasmic motifs (Tyr-based ordi-Leu) (Glickman et al., 1989; Ohno et al., 1995; Markset al., 1996); these interactions may help proteins tofind their correct cargo vesicle and consequently theirright locations in polarized cells (Bonifacino andDell’Angelica, 1999).
It is possible that the targeting signal present in thecytoplasmic tail of VAChT could have two distinct func-tions: targeting VAChT to synaptic sites and also sortingthe protein to synaptic vesicles. Alternatively, theremight be distinct trafficking signals for each step. Thepresent experiments attempt to answer whether the Cterminus may participate in targeting VAChT to varicos-ity sites in a cholinergic cell line (SN56) (Blusztajn et al.,1992; Barbosa et al., 1999) by taking advantage of theenhanced green fluorescent protein (EGFP) to tagVAChT.
MATERIALS AND METHODS
Cell cultureSN56 cells were a generous gift of Prof. Bruce Wainer
(Department of Pathology, Emory University School of Med-icine, Atlanta, GA, U.S.A.). The cells were maintained inDulbecco’s modified Eagle’s medium (Sigma), 10% fetal bo-vine serum (Gibco), 2 mM L-glutamine, and 1% penicillin/streptomycin in 50-ml culture bottles in a 5% CO2 atmosphereat 37°C. Cells were differentiated in the same medium butlacking fetal bovine serum and supplemented with 1 mM dibu-tyryl-cyclic AMP for at least 2 days. Medium was changedevery 2 days. For confocal analysis, cells were grown directlyon coverslips and were exposed to the same differentiationprotocol as described above. Cell passages up to eight wereused in the experiments.
HEK 293 cells were maintained in Dulbecco’s modifiedEagle’s medium (Sigma) supplemented with 10% fetal bovineserum (Gibco) and 1% penicillin/streptomycin in 5% CO2 at37°C.
Plasmid constructs, C-terminal fusion proteinexpression, and antibody generation
The complete coding region of the mouse VAChT gene, theC terminus (region codifying amino acids 470–530), and the12TM (region codifying amino acids 1–469) were amplified byPCR (Barbosa et al., 1999) (sequences for all the primers areavailable upon request), cloned 39 of EGFP in the pEGFP-C1vector (Living Colors, Clontech; see Fig. 1) using theEcoRIand BamHI restriction sites, and purified by CsCl gradient(Sambrook et al., 1989). All clones had their sequence con-firmed by automatic sequencing using the dideoxynucleotidechain termination reaction (Sanger et al., 1977).
The C terminus of the mouse VAChT was also cloned in thepMAL bacterial expression system (New England BioLabs) asdescribed (Barbosa et al., 1999). The purified recombinantprotein was used by a commercial supplier (Bion Ltda., BeloHorizonte, Brazil) to generate rabbit anti-VAChT antibodies.The serum was used without purification.
Cell transfectionSN56 and HEK 293 cells were plated on coverslips 24 h
before transfection with the different constructs (describedabove). Cells were transfected using either the calcium phos-phate or liposome-mediated (Tfx-50; Promega) method. For thecalcium phosphate protocol, 15mg of DNA was used for 106
cells and transfection was carried out as described by Sam-brook et al. (1989). Liposome-mediated transfections (1mg ofDNA for 104 cells) were performed according to the manufac-turer’s instructions. SN56 cells were maintained in serum-freemedium and differentiated after the transfection. Cells trans-fected with either protocol presented the same pattern of EGFPexpression; thus, we have used mainly Tfx for the experimentspresented herein.
Confocal imagingExperiments were performed at room temperature
(20–25°C). Cells on coverslips were incubated in HEPES-buffered salt solution (HBSS in mM: 124 NaCl, 4 KCl, 2CaCl2, 1.2 MgCl2, 10 glucose, 25 HEPES, pH 7.4, adjustedwith NaOH). The coverslips were washed in HBSS and thentransferred to a custom holder in which the coverslip formedthe bottom of a 400-ml bath. Imaging was performed with aBio-Rad MRC 1024 laser scanning confocal system runningthe software Lasersharp 3.0 coupled to a Zeiss microscope(Axiovert 100) with a water immersion objective (403, 1.2NA). The appropriate sets of filters and laser lines were used toavoid bleed-through between the different fluorophores used inthis work. Usually a Z series of EGFP-expressing cells wasobtained with an iris of,3 mm. However, some optical sec-tions of fixed cells were obtained with larger pinholes (iris 6).Image analysis and processing were performed with the Laser-sharp software (Bio-Rad), Adobe Photoshop, and Metamorph(Universal Imaging).
In general, fluorescence of EGFP in the soma of SN56 cellswas very strong. Therefore, in many cases, the settings toobtain images of the cell body and varicosities were different(see Fig. 8).
Labeling of organellesTo label endocytic organelles, we used the styryl dye FM4-
64. Cells were incubated with 1.5mM FM4-64 (MolecularProbes) for 16 h as described previously (Maletic-Savatic andMalinow, 1998). Then they were viewed by confocal micros-
FIG. 1. Schematic representation of the EGFP constructs. Theregions corresponding to EGFP and VAChT are represented byhatched and open bars, respectively (not in scale). The numbersat the bottom indicate the positions of residues that border eachVAChT construct used.
J. Neurochem., Vol. 74, No. 6, 2000
2426 M. S. SANTOS ET AL.

copy as described (Barbosa et al., 1999). For staining of lyso-somes in living cells, we used LysoTracker-Red (MolecularProbes). In brief, cells were incubated with LysoTracker (50nM) for 15 min, washed three times with HBSS, and visualizedimmediately by confocal microscopy. However, it was difficultto obtain Z series of cells labeled with LysoTracker because offast photobleaching. Thus for most of the experiments in whichthe dye was used, only a few optical sections (three to four)were obtained.
ImmunofluorescenceAfter visualization of cells transfected with EGFP or other
fusion proteins, cells were washed twice in 0.1M phosphatebuffer (pH 7.4) and then fixed with 4% paraformaldehyde in0.1 M phosphate buffer for 20 min. Immunolabeling with thedifferent antibodies was performed as described elsewhere(Barbosa et al., 1999). Primary antibodies used were an anti-Syp (1:150) monoclonal antibody (Sigma) and a rabbit anti-VAChT (1:2,000) antiserum. Appropriate secondary antibodiescoupled to Alexa568 (Molecular Probes) were used to visualizeimmunolabeling. In a few experiments, 4,6-diamidino-2-phe-nylindole (1:200) was added for visualization of the nucleus.Controls to test for specific labeling consisted of omission ofthe primary antibody or, in the case of anti-VAChT, incubatingthe antiserum with excess amount of recombinant fusion pro-tein or maltose binding protein (50mM). Adsorption occurredonly when the antiserum was incubated with maltose bindingprotein fused with the C-terminal tail of VAChT (not shown).
Simultaneous whole-cell voltage clamp and imagingWhole-cell patch clamp was performed on a Zeiss Axioskop
upright microscope with a 403 water immersion objective (0.5NA). A cooled MicroMax CCD Camera with a Sony 1300interlaced CCD chip (Princeton Instruments) was mounted onthe vertical port. The CCD camera was controlled by AxonImage Workbench software. Epifluorescence illumination pro-vided by an HBO mercury lamp was passed through an exci-tation filter centered at 485 nm and emitted light passed througha band-pass filter (505–530 nm).
Patch pipettes were pulled from borosilicate glass (Clark)and had resistances of 2–5 MV when filled with a pipettesolution containing the following (in mM): CsF (80), CsCl(35), NaCl (15), tetraethylammonium Cl (10), EGTA (10),HEPES (10), pH 7.4 (adjusted with NaOH), 299 mOsmol.Extracellular solution contained the following (in mM): NaCl(140), CsCl (5), CaCl2 (2), MgCl2 (1), HEPES (10), CdCl2
(0.2), pH 7.4 (adjusted with NaOH), 299 mOsmol. Thesesolutions were selected because they provided long-lived giga-ohm seals and allowed the measurement of Na1 currents as ameasure of cell viability. An Axon 200B patch clamp amplifierand pClamp software were used to control the voltage andrecord current through the patch pipette. The whole-cell con-figuration (Hamill et al., 1981) was formed in the standard wayby applying negative pressure to the patch pipette after agigaohm seal had been obtained. A holding potential of280mV was used in all experiments, and whole-cell electricalparameters (cell capacitance, access resistance, and membraneresistance) were determined by analysis of the current responseto a square wave (120 mV relative to the holding potential).Generation of the square wave and analysis of the currenttransients were performed by the Membrane Test option ofpClamp 7. The present setup allowed for the simultaneousimaging of the cells during dialysis of the cell cytosol in thepatch pipette. To analyze the fluorescence data, regions of
interest in the cells were drawn and the software reported theaverage fluorescence intensity (12-bit data) for each time point.
RESULTS
We first tested whether EGFP–VAChT chimeric pro-tein is correctly targeted in SN56 cells. NondifferentiatedSN56 cells transfected with the EGFP plasmid showed adiffuse pattern of expression for the fluorescent protein(Fig. 2A), suggesting that EGFP is soluble and not com-partmentalized in these cells. In contrast, when EGFP–VAChT was expressed by SN56 cells, the fusion proteinpresented discrete localization, observed as fluorescentpuncta in the cell cytoplasm (Fig. 2B). In;25% of theEGFP–VAChT-transfected cells, EGFP fluorescencewas also found in the nucleus, in a somewhat compart-mentalized fashion. This represented most likely an ar-tifact of overexpression, as immunolabeling of EGFP–VAChT-transfected cells with an antibody againstVAChT showed co-localization of the immunolabelingwith EGFP in the cytosol but not in the nucleus (notshown). Thus, it appears that a small part of the cellsdegrade the fusion protein and then EGFP, for an un-known reason, enters the nucleus.
Differentiated SN56 cells extend a large array of neu-rites that present several synaptic proteins in their vari-cosities (Barbosa et al., 1999). When transfected cellswere differentiated, EGFP–VAChT was also found to betargeted to varicosities, and in several cells the labelingon varicosities was punctate, suggesting its presence inmembranous compartments (Fig. 3, green). The patternof expression of EGFP–VAChT in the cell body wassimilar to that in nondifferentiated cells; fluorescencewas present in puncta throughout the soma (Fig. 3,green), and it contrasted with the soluble EGFP fluores-cence seen in differentiated cells (Fig. 3, inset). To verifythe endomembrane compartment in which VAChT was
FIG. 2. Distribution of EGFP and EGFP–VAChT fusion proteintransiently expressed in nondifferentiated SN56 cells. Forty-eight hours after transfection with the indicated constructs, livingcells were examined using confocal laser scanning microscopy.A: The EGFP fluorescence is scattered over the cell body. B:EGFP–VAChT fusion protein appears to be distributed in dis-crete subcellular compartments. N, nucleus. Bars 5 20 mm.
J. Neurochem., Vol. 74, No. 6, 2000
2427TRAFFICKING OF VAChT IN SN56 CELLS

present, we labeled living cells first with LysoTracker-Red, a weak amine that is trapped in acidic organellesand has been used to label lysosomes and late endosomes(Fig. 3, red). The overlay image (Fig. 3) indicates thatvery little overlap appears to exist between EGFP–VAChT fluorescence and lysosomes or late endosomes.In contrast, cells stained with an impermeant fluorescentprobe, FM4-64 (Fig. 4A, red), which labels endocyticorganelles, show that a large part of EGFP–VAChT (Fig.4A, green) is present in FM4-64-labeled organelles inliving cells (Fig. 4A, overlay yellow). Note that FM4-64also labels SN56 cells in a punctate pattern, and that invaricosities, part of EGFP–VAChT is also present inFM4-64-labeled organelles (Fig. 4A, arrow in the over-lay).
To test whether EGFP–VAChT co-localized withother synaptic vesicle proteins, transfected cells werefixed and processed for immunolabeling against Syp, asynaptic vesicle marker. As can be seen in Fig. 4B,EGFP–VAChT fluorescence overlapped with Syp immu-noreactivity in the soma and neuritic processes.
In an attempt to answer whether the C-terminal tail ofVAChT would participate in the trafficking of the trans-porter to varicosity sites in SN56 cells, two truncatedVAChT constructs were generated: The EGFP–C termi-nus contains the last 60 amino acids of VAChT, andEGFP–12TM is a deletion construct truncated afteramino acid 469 (the end of the 12th putative transmem-brane domain).
Surprisingly, the pattern of expression of the C-termi-nal construct in living SN56 cells was somewhat similarto that of VAChT (Fig. 5A1, A2, and B, green). Figure5A1 shows an image of the same cell as Fig. 5A2obtained with low laser power to show the discretelocalization of the C-terminal construct in the cytoplasm.A higher laser power image of the cell shows labeling ofneuritic processes (Fig. 5A2). The maximum projectionimage of another cell expressing this construct indicates
that fluorescence was punctate in the soma (Fig. 5B) andthat varicosities and growth cone also presented local-ized fluorescence (Fig. 5, arrows). Note that there wasnot much diffuse labeling of neurites (Fig. 5), but therewas clear presence of the truncated protein in varicosi-ties. However, the fluorescence was more diffuse thanthe pattern observed with EGFP–VAChT. As the C-terminal region of VAChT represents the cytoplasmictail of the VAChT protein (Gilmor et al., 1996; Weiheet al., 1996), it is possible that the fusion protein is boundto subcellular organelles at their cytoplasmic face. Thepunctate pattern of staining suggested that the EGFP–Cterminus could be located in endocytic organelles; thus,we tested whether EGFP-labeled organelles co-localizedwith FM4-64. Double-labeling experiments withFM4-64 (Fig. 5B, red and overlay) showed that, in gen-eral, co-localization of EGFP–C terminus with FM4-64-labeled organelles was not as good as that of EGFP–VAChT (just 9 of 26 cells labeled with FM4-64 showedsome degree of co-localization). However, immunofluo-rescence analysis with an antibody against Syp indicatesthat the C-terminal tail has some degree of co-localiza-tion with Syp immunoreactivity in fixed cells (Fig. 6),including their presence in varicosities (see Fig. 6B).This suggests that at least part of the fusion protein isfound in subcellular compartments that also present othersynaptic vesicle proteins.
To test whether the distribution pattern of EGFP–Cterminus is compatible with the notion that the protein isnot soluble in the cytoplasm, simultaneous imaging andwhole-cell voltage-clamp experiments were performed.First, to test whether EGFP in the absence of VAChTtargeting signals is soluble in the cytosol, we dialyzed thecell cytosol with a patch pipette in the whole-cell con-figuration while simultaneously following EGFP fluores-cence. Figure 7A and B shows an EGFP-transfected cellimmediately and several minutes after obtaining thewhole-cell configuration. EGFP rapidly diffuses to the
FIG. 3. Subcellular distribution of EGFP–VAChT fusion protein transiently expressed in differentiated SN56 cells. Forty-eight hours aftertransfection with the EGFP–VAChT, living cells were examined using confocal laser scanning microscopy. EGFP–VAChT is present ina punctate pattern and is targeted to varicosities (green). Labeling with LysoTracker-Red, a weak base that stains lysosomes and lateendosomes, also showed punctate fluorescence in the soma and varicosities (red). The overlay image shows an excluding pattern oflabeling for EGFP and LysoTracker in the optical section. Arrows indicate varicosities where the fusion protein is concentrated. Bar 5 10mm. Inset: Distribution of EGFP transiently expressed in differentiated SN56 cells. Forty-eight hours after transfection with the EGFPconstruct, living cells were examined by serial Z sectioning and the maximum Z projection is shown. Note that the fluorescence isscattered over the entire cell body and cellular processes. Bar 5 20 mm.
J. Neurochem., Vol. 74, No. 6, 2000
2428 M. S. SANTOS ET AL.

pipette, and after a few minutes the fluorescence in thecell diminishes (Fig. 7B and C). The decrease in fluo-rescence is exponential, as is quantitatively shown in Fig.7C. In contrast with EGFP-transfected cells, EGFP–VAChT did not exit to the patch pipette even afterseveral minutes in the whole-cell configuration (Fig.7D). Note that fluorescence decreases during illumina-tion at a constant rate of 1.86 0.26%/min (n5 11 forEGFP fluorescence before the whole-cell configuration),probably due to photobleaching. However, the fast ex-ponential decay in fluorescence that occurs in EGFP-transfected cells upon establishing the whole-cell config-uration [226 4.5%/min (n5 8) in the initial phase] wasdifferent from the loss of fluorescence in cells expressingEGFP–VAChT, which approached photobleaching rates[decay was 1.56 0.20%/min (n5 6)]. Indeed, the decayin EGFP–VAChT fluorescence in a patch-clamped cellwas no different from that of another cell in the samefield of view that was left intact during the experiment(Fig. 7D). Figure 7E shows Na1 currents and theirI/Vrelationship obtained for the cell from which the fluores-cence data represented in the figure (upper panel) werecollected, indicating that the whole-cell configurationwas obtained. Data from seven cells indicated that the Cterminus did not readily diffuse to the patch pipette asdid EGFP, and the rate of fluorescence decay was notdifferent from that of photobleaching [1.86 0.33%/min(n 5 7); Fig. 7E]. We interpreted this result as anindication that the chimeric protein is bound to a subcel-lular compartment. As the diameter of the patch pipette is;1 mm, the addition of 7 kDa to EGFP (VAChT–Cterminus) should not impair its dialysis out of the cell.
The above data suggest that the C-terminal VAChTtail has the potential to interact with organelles and thatit might participate in VAChT trafficking to varicosities.However, these experiments do not answer the questionof whether the C terminus contains the unique sorting/trafficking signal for VAChT to be targeted to varicosi-ties. To address whether the C terminus is essential forVAChT trafficking, we transfected SN56 cells withEGFP–12TM. Interestingly, this truncated form ofVAChT was also located in puncta in the soma of dif-ferentiated SN56 cells (Fig. 8B, green). Moreover,EGFP–12TM was also found accumulated in varicosities(Fig. 8A and B, arrows). Twenty of 28 cells (71.4%)transfected with EGFP–12TM presented partial overlapof the fusion protein with FM4-64-labeled organelles, asindicated in Fig. 8B, and this was a little less than thatobtained for EGFP–VAChT (85% of the transfected cellsexamined presented co-localization with FM4-64). Moreinterestingly, as EGFP–12TM lacks the C-terminal tail ofVAChT, which contains the region against which ourantibody was raised, we could test whether this fusionprotein was targeted in a similar way to endogenousVAChT. This experiment (Fig. 8C) indicates that there isco-localization of EGFP–12TM and endogenous VAChTin fixed SN56 cells.
To further explore the role of C terminus in VAChTtrafficking, we used the above constructs to transfect a
FIG. 4. EGFP–VAChT is present in endocytic organelles and isco-localized with Syp. A: Co-localization of EGFP–VAChT andFM4-64 in differentiated SN56 cells. Forty-eight hours aftertransfection with EGFP–VAChT constructs, living cells wereexamined by serial Z sectioning and the maximum Z projec-tion is presented. EGFP–VAChT fluorescence (green) ispresent in distinct subcellular compartments and also in var-icosities (arrow). When cells were incubated with FM4-64 for16 h, they presented staining compatible with the presence ofthe impermeant dye inside membranous structures (red). Theoverlay image shows that in many regions, EGFP–VAChT andFM4-64 fluorescence co-localize (yellow in the overlap). Arrowindicates a varicosity that was able to internalize the dye dueto constitutive endocytic activity and presented partial co-localization. The bottom right panel shows the differentialinterference contrast image for the cells. B: Co-localization ofEGFP–VAChT and Syp in SN56 cells. Optical section of a cellthat expressed the fusion protein (green) was labeled with amonoclonal antibody against Syp (red). The overlay imageshows that both proteins appear co-localized (yellow). Thenucleus was stained with 4,6-diamidino-2-phenylindole (blue).Arrows indicate co-localization in neurites. Bars 5 20 mm (A)and 10 mm (B).
J. Neurochem., Vol. 74, No. 6, 2000
2429TRAFFICKING OF VAChT IN SN56 CELLS

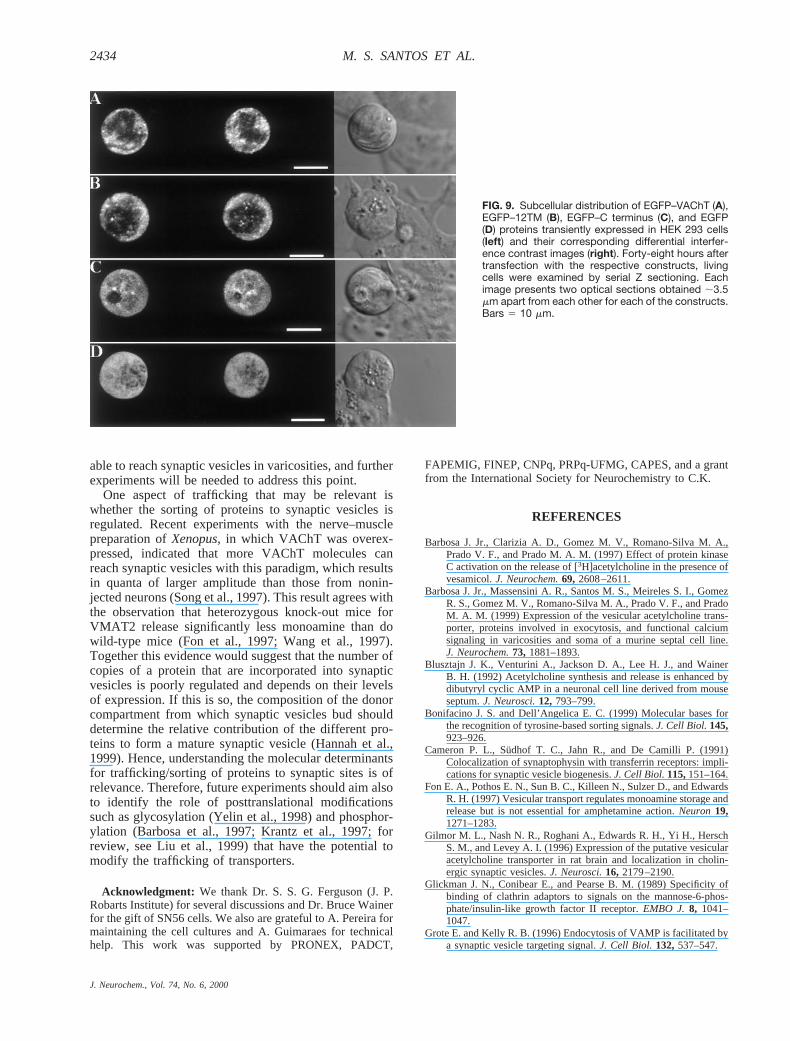
nonneuronal cell line, HEK 293. Synaptic vesicle pro-teins, including VAChT, are targeted to membranouscompartments in nonneuronal cells (Johnston et al.,1989; Cameron et al., 1991; Liu and Edwards, 1997).EGFP–VAChT in HEK 293 cells was observed in apattern compatible with labeling of membranous struc-tures close to the plasma membrane (Fig. 9A) and alsotoward the middle of the cell. A very similar result wasobtained with EGFP–12TM, which presented a predom-inant cytoplasmic localization in what appears to be alarge endomembrane compartment (Fig. 9B).
The pattern of expression of the EGFP–C terminus inSN56 cells suggests that it can accumulate in varicositysites and may be bound to an endomembrane compart-ment. To test whether the punctate pattern observed inSN56 cells corresponds to interactions that may also takeplace in nonneuronal cells, we transfected HEK 293 cellswith EGFP–C terminus. The pattern of expression of thisconstruct was different from that seen in SN56 cells (Fig.9C). The protein presented a much more diffuse expres-sion (Fig. 9C). Moreover, only very few fluorescent
puncta of small size could be detected when the Cterminus was expressed.
DISCUSSION
Localization of EGFP–VAChT in living cellsExpression of EGFP in SN56 cells shows that the
protein is soluble, whereas EGFP–VAChT presents adiscrete localization, indicating that VAChT-targetingsequences are effective in the presence of EGFP. EGFP–VAChT was targeted to endocytic-like organelles in thesoma and also to varicosities of SN56 cells. At the lightmicroscopy level, EGFP–VAChT appeared sometimesas large fluorescent puncta, but small vesicle-like struc-tures were also observed. Previous work with PC12 cellsoverexpressing VAChT has shown that the transporterco-localized with transferrin receptors and Syp, whichare found in endosomes in the cell body (Liu and Ed-wards, 1997). Our data agree with these early observa-tions, as EGFP–VAChT was not consistently found inlysosomes or late endosomes, but it did co-localize inlarge extension with organelles that stained with the
FIG. 5. The EGFP–C terminus presents a punctatepattern of expression. A: Optical sections of anSN56 cell transiently expressing the EGFP–C-termi-nal fusion protein: an image of a cell obtained at lowpower to allow better visualization of the expressionof EGFP–C terminus in the cell body (A1); same cellas in A1 but imaged with high laser power and lowermagnification for better visualization of neurites andvaricosities (A2). Arrows indicate varicosities. Alsoshown is the differential interference contrast imagefor the cell (right). B: Double-labeling experimentwith the EGFP–C terminal fusion protein and the vitaldye FM4-64. Forty-eight hours after transfectionwith the EGFP–C terminal construct and 16 h afterincubation with FM4-64, living cells were examinedby serial Z sectioning and the maximum Z projectionis shown. The cell expressing the fusion protein(green) shows a punctate fluorescence in the soma,and the varicosity presents localized EGFP fluores-cence (arrows). FM4-64 labeling of endocytic or-ganelles is red. The overlay image shows that thereis much less co-localization than observed forEGFP–VAChT (see Fig. 4A). The bottom right panelshows the correspondent differential interferencecontrast image. Bars 5 20 mm.
J. Neurochem., Vol. 74, No. 6, 2000
2430 M. S. SANTOS ET AL.

endocytic probe FM4-64. Moreover, EGFP–VAChT alsoshowed a good degree of co-localization with Syp infixed cells, similar to endogenous VAChT, which co-localizes with Syp in SN56 cells (Barbosa et al., 1999).Varicosities of SN cells possess several synaptic proteins(Barbosa et al., 1999) and also present synaptic vesicles(Hammond et al., 1990). Therefore, the simplest inter-pretation for these data is that VAChT recycles throughthe plasma membrane to endosomes, travels perhapsthrough tubulovesicular organelles to varicosities, andthen reaches synaptic vesicles. If this picture is correct,early observations of ACh release in SN56 cells that areneurochemically differentiated but do not present vari-cosities (Blusztajn et al., 1992) would be explained bythe presence of VAChT in cell body organelles that fuseto the plasma membrane and recycle in the soma. How-ever, as we used a membrane probe to label endocyticvesicles, this experiment cannot discern between thealternative possibility that VAChT is targeted from the
trans-Golgi network to an endomembrane compartmentthat receives recycling vesicles, and thus dye, but that theprotein itself does not sort to this recycling pathway andrarely reaches the plasma membrane in the soma. Despitethese two possibilities, it is clear that VAChT reachesendosomes and is also sorted to transport vesicles goingto varicosities.
Role of C-terminal VAChT tail in traffickingOverexpression of the C terminus tagged with EGFP
surprisingly suggests a pattern of expression similar tothat of EGFP–VAChT; in particular, the fragment ap-peared to label varicosities or growth cones much morethan it labeled neurites. Two lines of evidence supportthat EGFP–C terminus is not freely soluble: Most of thecells transfected presented a punctate pattern of expres-sion in the cell body, and EGFP–C terminus was notreadily dialyzed to the pipette used to obtain the whole-cell configuration in SN56 cells. In this regard, the re-sults were similar to those of VAChT and distinct fromthose obtained with EGFP, which presents a diffuseexpression pattern and is readily dialyzed to the patchpipette. It should be noticed that there was more diffusefluorescence with the C-terminal construct than withVAChT in SN56 cells, which indicates that the predictedcytosolic carboxy-tail peptide was not as effectively tar-geted to endomembranes as the intact protein. One pos-sibility is that the endogenous VAChT could bind moreeffectively to sites in endosomes than the EGFP–C ter-minus. It could also mean that the system is saturable, ashas already been shown in other trafficking processes(Marks et al., 1996). Nonetheless, it is still intriguing thatthis predicted cytosolic fragment presents such a distinctpattern of expression.
EGFP–C terminus did not seem to be largely presentin organelles that are derived by endocytic activity in thecell body of SN56 cells as VAChT is, as there was poorfluorescence overlap with the endocytic dye FM4-64.One possibility is that the C terminus is bound to thecytoplasmic face of transport vesicles or compartmentsthat participate in the trafficking of proteins to varicosi-ties, perhaps by protein–protein interaction. However,we cannot discard the possibility that overexpression ofEGFP–C terminus is able to compete in the normalpathway of VAChT trafficking, and thus it could de-crease endocytic activity in transfected cells.
Would there be a signal for VAChT trafficking tovaricosities different from the C terminus? Our resultssuggest that although the C terminus has characteristicsthat would indicate a role in VAChT transport, there maybe also other regions of the protein that could participatein such a task.
The EGFP–12TM construct generated a protein with apattern of expression similar to that of VAChT. Theprotein presented punctate labeling in the soma in endo-cytic organelles, but, more importantly, it also accumu-lated in varicosities. These results would favor the ideathat other trafficking or sorting signals may be present inthe protein and can drive it to varicosities and that the C
FIG. 6. Localization of EGFP–C terminus and Syp in transientlytransfected SN56 cells. A: Optical sections of the cell body of acell expressing the EGFP–C terminal protein (green) and labeledwith a monoclonal antibody against Syp (red). The overlay imageshows a region with similar distribution (yellow). B: Image ofvaricosities of a cell expressing EGFP–C terminus and labeledwith anti-Syp antibody. Cell expressing the EGFP–C terminus(green) shows accumulation in varicosities and the presence ofSyp (red). The overlay image shows co-localization of both pro-teins in varicosities (arrows). Bars 5 20 mm.
J. Neurochem., Vol. 74, No. 6, 2000
2431TRAFFICKING OF VAChT IN SN56 CELLS

terminus is not essential for this aspect of VAChT traf-ficking.
A potential problem for the interpretation of theseresults could arise from the transitory nature of the ex-pression of the EGFP constructs and overexpression ofthe recombinant proteins. However, is should be noticedthat expression of distinct fusion proteins (protein kinaseCbII and Rab5) tagged with EGFP generates patterns oflabeling in SN56 cells distinct from that of VAChT or itstruncated forms (our unpublished observations). Theseobservations support the notion that the localization ofEGFP–VAChT and of the truncated forms is related tothe targeting signals found in the VAChT protein.
Another interesting aspect of the cell biology ofVAChT trafficking came from experiments using HEK293 cells. The pattern of expression of EGFP–VAChTand EGFP–12TM was very similar, as seen in SN56cells. This was surprising as earlier experiments in non-
neuronal cells have suggested that the di-Leu motif in theC-terminal tail acts as an internalization signal whentransferred to a plasma membrane protein (Tan et al.,1998). Although VAChT in the experiments of Tan andcollaborators did not seem to cycle to the plasma mem-brane as effectively as VMAT2, mutation of the di-Leuresults in accumulation of VAChT in the plasma mem-brane. It is not clear why VAChT–12TM did not accu-mulate in the plasma membrane in the present experi-ments both in SN56 and in HEK 293 cells. One possi-bility is that the cells that we used (SN56 and HEK 293)have different trafficking behavior than the CHO cellsused by Tan et al. (1998). Alternatively, the C terminusmay contain additional trafficking information that couldparticipate in sorting to vesicles that fuse to the plasmamembrane. As these trafficking signals would be sup-pressed from the EGFP–12TM construct, the truncatedprotein would be impaired from leaving an intracellular
FIG. 7. EGFP is soluble in the cytosol. A: Cells expressing EGFP were patch-clamped in the whole-cell configuration. The image showsa representative cell immediately after going to whole-cell mode. Note the diffusion of EGFP to the patch pipette. B: Same cell in A after10 min of dialysis in the whole-cell mode. Bars 5 15 mm. C: Quantification of fluorescence levels from cell in A before and after obtainingthe whole-cell configuration. D: A cell expressing EGFP–VAChT was analyzed in the same way as in C. Filled circles representfluorescence from a cell that was patch-clamped, whereas open circles indicate the fluorescence from a cell that was not patch-clamped present in the same field of view. Note that there is a similar degree of decrease in fluorescence for both cells, indicatingphotobleaching during the prolonged illumination. E: Lack of dialysis of EGFP–C terminus to a patch pipette during whole-cell voltageclamp (upper panel). The experiment was performed as above, and the fluorescence was followed for a cell expressing EGFP–Cterminus. Also shown are sodium currents (middle panel) and the corresponding I/V relationship (bottom panel) recorded as describedin Materials and Methods.
J. Neurochem., Vol. 74, No. 6, 2000
2432 M. S. SANTOS ET AL.

compartment to the plasma membrane. Further experi-ments will be necessary to understand this difference.
Transfection of the C-terminal construct in HEK 293cells indicated that the protein presented a much morediffuse pattern of expression than observed in SN56cells, suggesting that if the C terminus is binding to acellular component, the latter is preferentially present inneuronal or cholinergic cells. This result agrees withrecent observations withDrosophila, which appear tohave a motor protein from the Kinesin superfamily pref-erentially expressed in cholinergic neurons. This proteinhas been implicated in the trafficking of choline acetyl-transferase (Ray et al., 1999).
The final steps of synaptic vesicle assembly arethought to occur at the nerve terminal (for review, seeHannah et al., 1999) to which synaptic vesicle proteinstravel in tubulovesicular organelles (Okada et al., 1995;Nakata et al., 1998). Moreover, VAMP seems to rely onat least two different targeting signals: one to be directedto an endosomal compartment at synaptic sites and the
other to be incorporated in synaptic-like vesicles. Thelatter trafficking signal seems to be related to the hydro-phobicity of a predicted amphipathica helix on thecytosolic region of the protein (Grote et al., 1995),whereas the former could not be identified by truncatingseveral stretches of 10 amino acids on the cytoplasmicregion of VAMP (West et al., 1997).
One of the interpretations for the present data mayfollow the proposal of different signals: one that directsthe protein to varicosities and the other to synaptic ves-icles. The C terminus, as has been shown previously,participates in targeting to synaptic vesicles (Varoquiand Erickson, 1998), and our data suggest that this frag-ment may bind to neuronal components in living cells.However, we cannot exclude that signal(s) other than theC terminus take part in the trafficking pathway ofVAChT, as evidenced by what appears to be the correctlocalization of EGFP–12TM in varicosities. These re-sults, however, give no information about whether thesefusion proteins (EGFP–12TM or EGFP–C terminus) are
FIG. 8. A: Distribution of EGFP–12TMfusion protein transiently expressed indifferentiated SN56 cells. Forty-eighthours after transfection with the EGFP–12TM construct and 16 h after incuba-tion with FM4-64, living cells were exam-ined by serial Z sectioning. The maxi-mum Z projection is shown. EGFP–12TM fusion protein is present in thesoma and is also accumulated in vari-cosities (arrows; green). The right lowerpanel shows the differential interferencecontrast image of the cells. B: Same asin A but with images obtained with lowerlaser power for better visualization of thedistribution of EGFP–12TM in the soma.Note in the overlay that there are severalregions of overlap between EGFP–12TMand FM4-64. C: Localization of EGFP–12TM (green) and immunofluorescencefor endogenous VAChT (red) are shownin optical sections. The overlay showsco-localization of the EGFP–12TM andendogenous VAChT. Bars 5 20 mm (Aand B) and 10 mm (C).
J. Neurochem., Vol. 74, No. 6, 2000
2433TRAFFICKING OF VAChT IN SN56 CELLS
able to reach synaptic vesicles in varicosities, and furtherexperiments will be needed to address this point.
One aspect of trafficking that may be relevant iswhether the sorting of proteins to synaptic vesicles isregulated. Recent experiments with the nerve–musclepreparation ofXenopus,in which VAChT was overex-pressed, indicated that more VAChT molecules canreach synaptic vesicles with this paradigm, which resultsin quanta of larger amplitude than those from nonin-jected neurons (Song et al., 1997). This result agrees withthe observation that heterozygous knock-out mice forVMAT2 release significantly less monoamine than dowild-type mice (Fon et al., 1997; Wang et al., 1997).Together this evidence would suggest that the number ofcopies of a protein that are incorporated into synapticvesicles is poorly regulated and depends on their levelsof expression. If this is so, the composition of the donorcompartment from which synaptic vesicles bud shoulddetermine the relative contribution of the different pro-teins to form a mature synaptic vesicle (Hannah et al.,1999). Hence, understanding the molecular determinantsfor trafficking/sorting of proteins to synaptic sites is ofrelevance. Therefore, future experiments should aim alsoto identify the role of posttranslational modificationssuch as glycosylation (Yelin et al., 1998) and phosphor-ylation (Barbosa et al., 1997; Krantz et al., 1997; forreview, see Liu et al., 1999) that have the potential tomodify the trafficking of transporters.
Acknowledgment: We thank Dr. S. S. G. Ferguson (J. P.Robarts Institute) for several discussions and Dr. Bruce Wainerfor the gift of SN56 cells. We also are grateful to A. Pereira formaintaining the cell cultures and A. Guimaraes for technicalhelp. This work was supported by PRONEX, PADCT,
FAPEMIG, FINEP, CNPq, PRPq-UFMG, CAPES, and a grantfrom the International Society for Neurochemistry to C.K.
REFERENCES
Barbosa J. Jr., Clarizia A. D., Gomez M. V., Romano-Silva M. A.,Prado V. F., and Prado M. A. M. (1997) Effect of protein kinaseC activation on the release of [3H]acetylcholine in the presence ofvesamicol.J. Neurochem.69, 2608–2611.
Barbosa J. Jr., Massensini A. R., Santos M. S., Meireles S. I., GomezR. S., Gomez M. V., Romano-Silva M. A., Prado V. F., and PradoM. A. M. (1999) Expression of the vesicular acetylcholine trans-porter, proteins involved in exocytosis, and functional calciumsignaling in varicosities and soma of a murine septal cell line.J. Neurochem.73, 1881–1893.
Blusztajn J. K., Venturini A., Jackson D. A., Lee H. J., and WainerB. H. (1992) Acetylcholine synthesis and release is enhanced bydibutyryl cyclic AMP in a neuronal cell line derived from mouseseptum.J. Neurosci.12, 793–799.
Bonifacino J. S. and Dell’Angelica E. C. (1999) Molecular bases forthe recognition of tyrosine-based sorting signals.J. Cell Biol.145,923–926.
Cameron P. L., Su¨dhof T. C., Jahn R., and De Camilli P. (1991)Colocalization of synaptophysin with transferrin receptors: impli-cations for synaptic vesicle biogenesis.J. Cell Biol.115,151–164.
Fon E. A., Pothos E. N., Sun B. C., Killeen N., Sulzer D., and EdwardsR. H. (1997) Vesicular transport regulates monoamine storage andrelease but is not essential for amphetamine action.Neuron19,1271–1283.
Gilmor M. L., Nash N. R., Roghani A., Edwards R. H., Yi H., HerschS. M., and Levey A. I. (1996) Expression of the putative vesicularacetylcholine transporter in rat brain and localization in cholin-ergic synaptic vesicles.J. Neurosci.16, 2179–2190.
Glickman J. N., Conibear E., and Pearse B. M. (1989) Specificity ofbinding of clathrin adaptors to signals on the mannose-6-phos-phate/insulin-like growth factor II receptor.EMBO J. 8, 1041–1047.
Grote E. and Kelly R. B. (1996) Endocytosis of VAMP is facilitated bya synaptic vesicle targeting signal.J. Cell Biol. 132,537–547.
FIG. 9. Subcellular distribution of EGFP–VAChT (A),EGFP–12TM (B), EGFP–C terminus (C), and EGFP(D) proteins transiently expressed in HEK 293 cells(left) and their corresponding differential interfer-ence contrast images (right). Forty-eight hours aftertransfection with the respective constructs, livingcells were examined by serial Z sectioning. Eachimage presents two optical sections obtained ;3.5mm apart from each other for each of the constructs.Bars 5 10 mm.
J. Neurochem., Vol. 74, No. 6, 2000
2434 M. S. SANTOS ET AL.

Grote E., Hao J. C., Bennett M. K., and Kelly R. B. (1995) A targetingsignal in VAMP regulating transport to synaptic vesicles.Cell 81,581–589.
Hamill O. P., Marty A., Neher E., Sakmann B., and Sigworth F. J.(1981) Improved patch-clamp techniques for high-resolution cur-rent recording from cells and cell-free membrane patches.PflugersArch. 391,85–100.
Hammond D. N., Lee H. J., Tonsgard J. H., and Wainer B. H. (1990)Development and characterization of clonal cell lines derivedfrom septal cholinergic neurons.Brain Res.512,190–200.
Hannah M. J., Schmidt A. A., and Huttner W. B. (1999) Synapticvesicle biogenesis.Annu. Rev. Cell Dev. Biol.15, 733–798.
Johnston P. A., Cameron P. L., Stukenbrok H., Jahn R., De Camilli P.,and Su¨dhof T. C. (1989) Synaptophysin is targeted to similarmicrovesicles in CHO and PC12 cells.EMBO J. 8, 2863–2872.
Krantz D. E., Peter D., Liu Y., and Edwards R. H. (1997) Phosphor-ylation of a vesicular monoamine transporter by casein kinase II.J. Biol. Chem.272,6752–6759.
Liu Y. and Edwards R. H. (1997) Differential localization of vesicularacetylcholine and monoamine transporters in PC12 cells but notCHO cells.J. Cell Biol. 139,907–916.
Liu Y., Krantz D. E., Waites C., and Edwards R. H. (1999) Membranetrafficking of neurotransmitter transporters in the regulation ofsynaptic transmission.Trends Cell Biol.9, 356–363.
Maletic-Savatic M. and Malinow R. (1998) Calcium-evoked dendriticexocytosis in cultured hippocampal neurons. Part I:trans-Golginetwork-derived organelles undergo regulated exocytosis.J. Neu-rosci. 18, 6803–6813.
Marks M. S., Woodruff L., Ohno H., and Bonifacino J. S. (1996)Protein targeting by tyrosine- and di-leucine-based signals: evi-dence for distinct saturable components.J. Cell Biol. 135, 341–354.
Mellman I. (1996) Endocytosis and molecular sorting.Annu. Rev. CellDev. Biol.12, 575–625.
Nakata T., Terada S., and Hirokawa N. (1998) Visualization of thedynamics of synaptic vesicle and plasma membrane proteins inliving axons.J. Cell Biol. 140,659–674.
Ohno H., Stewart J., Fournier M. C., Bosshart H., Rhee I., Miyatake S.,Saito T., Gallusser A., Kirchhausen T., and Bonifacino J. S.(1995) Interaction of tyrosine-based sorting signals with clathrin-associated proteins.Science269,1872–1875.
Okada Y., Yamazaki H., Sekine-Aizawa Y., and Hirokawa N. (1995)The neuron-specific kinesin superfamily protein KIF1A is aunique monomeric motor for anterograde axonal transport ofsynaptic vesicle precursors.Cell 81, 769–780.
Ray K., Perez S. E., Yang Z., Xu J., Ritchings B. W., Steller H., andGoldstein L. S. (1999) Kinesin-II is required for axonal transportof choline acetyltransferase in drosophila.J. Cell Biol.147,507–518.
Regnier-Vigouroux A., Tooze S. A., and Huttner W. B. (1991) Newlysynthesized synaptophysin is transported to synaptic-like mi-crovesicles via constitutive secretory vesicles and the plasmamembrane.EMBO J.10, 3589–3601.
Sambrook J., Fritsch E. F., and Maniatis T. (1989)Molecular Cloning:A Laboratory Manual,2nd ed. Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, NY.
Sanger F., Nicklen S., and Coulson A. R. (1977) DNA sequencing withchain-terminating inhibitors.Proc. Natl. Acad. Sci. USA74,5463–5467.
Song H., Ming G., Fon E., Bellocchio E., Edwards R. H., and Poo M.(1997) Expression of a putative vesicular acetylcholine transporterfacilitates quantal transmitter packaging.Neuron18, 815–826.
Tan P. K., Waites C., Liu Y., Krantz D. E., and Edwards R. H. (1998)A leucine-based motif mediates the endocytosis of vesicularmonoamine and acetylcholine transporters.J. Biol. Chem.273,17351–17360.
Tao-Cheng J. H. and Eiden L. E. (1998) The vesicular monoaminetransporter VMAT2 and vesicular acetylcholine transporterVAChT are sorted to separate vesicle populations in PC12 cells.Adv. Pharmacol.42, 250–253.
Varoqui H. and Erickson J. D. (1998) The cytoplasmic tail of thevesicular acetylcholine transporter contains a synaptic vesicletargeting signal.J. Biol. Chem.273,9094–9098.
Wang Y. M., Gainetdinov R. R., Fumagalli F., Xu F., Jones S. R., BockC. B., Miller G. W., Wightman R. M., and Caron M. G. (1997)Knockout of the vesicular monoamine transporter 2 gene results inneonatal death and supersensitivity to cocaine and amphetamine.Neuron19, 1285–1296.
Weihe E., Tao-Cheng J. H., Schafer M. K., Erickson J. D., and EidenL. E. (1996) Visualization of the vesicular acetylcholine trans-porter in cholinergic nerve terminals and its targeting to a specificpopulation of small synaptic vesicles.Proc. Natl. Acad. Sci. USA93, 3547–3552.
West A. E., Neve R. L., and Buckley K. M. (1997) Targeting of thesynaptic vesicle protein synaptobrevin in the axon of culturedhippocampal neurons: evidence for two distinct sorting steps.J. Cell Biol. 139,917–927.
Yelin R., Steiner-Mordoch S., Aroeti B., and Schuldiner S. (1998)Glycosylation of a vesicular monoamine transporter: a mutation ina conserved proline residue affects the activity, glycosylation, andlocalization of the transporter.J. Neurochem.71, 2518–2527.
J. Neurochem., Vol. 74, No. 6, 2000
2435TRAFFICKING OF VAChT IN SN56 CELLS
Copyright © 2022 FDOKUMEN




















