
Production of Monoclonal Antibodies Directed against the Microsporidium Enterocytozoon bieneusi
Upload
independentCategory
view
0download
0
Cell Motility and the Cytoskeleton 20:109-120 (1991)
Inhibition of lntracellular Granule Movement by Microinjection of Monoclonal Antibodies
Against Caldesmon
Theresa E. Hegmann, Douglas L. Schulte, Jenny Li-Chun Lin, and Jim Jung-Ching Lin
Department of Biology, University of Iowa, Iowa City
Monoclonal antibodies, C2, C9, C18, and C21, against chicken gizzard caldes- mon (called high molecular weight isoform) were shown to crossreact with a low molecular weight isoform of caldesmon in chicken embryo fibroblasts (CEF). These antibodies were used in a microinjection study to investigate the in vivo function of caldesmon in nonmuscle cell motility. Injected cells did not appear to change their morphology significantly; the cells displayed a flat appearance and were able to ruffle and locomote normally. However, in the C21 injected cells, saltatory movements of granules and organelles appeared to be greatly inhibited. This inhibition of granule movement was reversible, so that by 3 hr after injection, granules in injected cells had already recovered to normal speed. The inhibition of granule movement by C21 antibody was also very specific; the average speeds of granule movement in cells injected with C2, C9, or C18 antibody, or with C21 antibody preabsorbed with caldesmon, were not significantly different from that in uninjected cells. In a previous epitope study, we demonstrated that, of the antibodies used in this study, only C21 antibody was able to compete with the binding of caldesmon to Caf +/calmodulin and to F-actin, although both C21 and C2 antibodies recognized the same carboxyl-terminal 10K fragment of gizzard caldesmon [Lin et al., 1991: Cell Motil. Cytoskeleton 20:95-1081. The caldes- mon distribution in C21 injected cells changed from stress-fiber localization to a more diffuse appearance, when the injection was performed at 10-30 mg/ml of C21 antibody. We have previously shown that a monoclonal anti-tropomyosin antibody exhibited motility-dependent recognition of an epitope, and that micro- injection of this antibody specifically inhibited intracellular granule movements of CEF cells [Hegmann et al., 1989: J . Cell Biol. 109:1141-11521. Therefore, it is likely that tropomyosin and caldesmon may both function in intracellular granule movement by regulating the contractile system in response to [Ca++] change inside nonmuscle cells.
Key words: nonmuscle tropomyosin, Ca+ +/calmodulin-binding protein, actin-binding protein, epitope, actomyosin
INTRODUCTION
Caldesmon is a protein that binds both C a + + / calmodulin and actin filaments [Sobue et al., 19811. In addition to myosin light chain phosphorylation, caldes- mon has been implicated in another mechanism respon- sible for smooth muscle contraction. Caldesmon has been shown to be associated with thin filaments of smooth muscle cells [Furst et al., 1986; Sobue et al., 198 11 and will inhibit both actomyosin interaction
0 1991 Wiley-Liss, Inc.
[Chalovich, 1988; Sobue et al., 1981, 19821 and acto- myosin ATPase activity [Chalovich et al., 1987; Ngai and Walsh, 1984; Sobue et al., 19821 in vitro in the absence of C a t + and calmodulin. Addition of C a + + /
Received January 3, 1991; accepted June 7, 1991.
Address reprint request to Dr. Jim Jung-Ching Lin, Department of Biology, University of Iowa, Iowa City, Iowa 52242.

110 Hegmann et al.
calmodulin can release the inhibition imposed by caldes- Anti-Caldesmon Antibodies
The preparation and characterization of mono- mon [Sobue et al., 1981, 1982; Ngai and Walsh, 19841. The apparent clonal anti-caldesmon antibodies, C2, (79, C 18, and (72 1 require the presence Of tropomyosin et 1981, were reported previously [Lin et al., 19881. Their
properties Of cladesmon Seem to
1982; Smith et al., 1987; Fujii et al., 1988; Horiuchi and Chacko, 19881. epitopes were mapped by Western blot analysis on
NTCB (2-nitro-5-thiocyanobenzoic acid) and CNBr (cy- Nonmuscle and tissues, as many types Of anogen bromide) fragments of chicken gizzard ca]des-
[Bretscher and Lynch, 1 9 8 6 i 9 human plate- [so-
T-lYmPhocYtes rMizushima mon [Lin et a]. , 19911. All these antibodies were from ascites fluid by protein A-sepharose column chro- matography as described previously [Lin et a]. , 19851.
lets [Dingus et 3 986i 9 bovine et 7 i985i 7 and
et al., 19871, have been shown to contain a low molec-
function of nonmuscle caldesmon remains to be deter- mined. Recently, in vitro reconstitution experiments have demonstrated that nonmuscle caldesmon can en- hance the binding of nonmuscle tropomyosin to F-actin filaments [Yamashiro-Matsumura and Matsumura, 19881, suggesting an interaction between these two pro- teins. It has also been shown that both the high and
purified antibodies were conjugated to lissamine
[Hegmann et
Microinjection, Time-Lapse Photography, and ~ ~ ~ l ~ ~ i ~ of G~~~~~~ Movements
CEF cells grown on glass coverslips were micro- injected with rhodamine-conjugated antibodies as de-
'lar weight Of caldesmon. The PhYsiol0gica1 rhodamine sulfonyl chloride as described previously 19891 for use in microinjection.
molecular weight isoforms of lymphocyte caldesmon may be critically important in regulating actomyosin in-
scribed Previously [Hegmann et al. 3 19891. The cover- 'lips were then quickly transferred to a perfusion
teraction, which is responsible for the capping processes of surface receptors [Mizushima et al., 1987; Walker et
chamber [Berg and B1och, 19841 and viewed with an epifluorescence microscope. Injected cells were located
al., 19891. In the present study, we have used the tech- by and the phase-contrast image was nique of microinjection to introduce monoclonal anti- caldesmon antibody into CEF cells and to thus assess the
of our antibodies C21, which was previously shown to times speed for Of granule movements.
eotaped with a Nuvicon camera (Dage/MIT, Inc. 3 Mich- igan City, IN) connected to a Panasonic time-lapse video
in viva function of ca]desmon. Microinjection of one
be able to inhibit the binding of caldesmon to C a t + /
revealed a drastic reduction of instantaneous speed and
similar effect on granule movement has been previously
cassette recorder. The videotape was played back at six
Intracellular granule movements were measured
et al., 19891. Instanteous speeds were calculated by mea-
tion. Measurements were done for eight randomly cho-
calmodulin and to F-actin filaments [Lin et al., 19911, according to the method PrevioUs1Y IHegmann
saltation distance of intracellular granule movement. A suring the duration and length Of sing1e
observed in experiments with anti-tropomyosin antibody injection [Hegmann et a]., 19891. Therefore, these re-
sen moving granules in each Of at least ten treatment. The average distance moved in a sing1e
Per
tation (saltation distance) was calculated by measuring 20 saltations in each of four cells per treatment.
For locomotion analysis, NRK cells were grown on glass coverslips to confluence, and then a strip of cells was removed from the middle of the coverslips by scratching the cell monolayer with a sterile needle. As the cells on the edges of the wound line started to migrate into the empty space, several migrating cells in the same field were microinjected with C21 antibody and the mi- gration was videotaped for more than 3 hr.
sults suggest that nonmuscle caldesmon and tropomyosin may interact with each other and participate in the reg- ulation of an actomyosin system involved in intracellular granule movement.
MATERIALS AND METHODS Cell Cultures
Primary cultures of fibroblast cells were prepared from the skin of 10-d-old chicken embryos by dissection and trypsinization as described previously [Lin et al., 19841. CEF cells between the second and sixth passaging For indirect double-label immunofluorescence,
~mmunofluorescence Microscopy
were used for microinjection and staining. Normal rat kidney cells (NRK, American Type Culture Collection CRL-1570) were used for the locomotion study. All cells were cultured in Dubecco's modified Eagle (DME) me- dium with 10% fetal bovine serum (FBS) and were kept in a humidified incubator at 37°C and 5% CO,.
cells grown on coverslips were microinjected with puri- fied monoclonal antibodies. After 30 min, cells were fixed and permeabilized as described previously [Lin and Feramisco, 19811. The coverslips were then incubated for 30 min each in FITC-conjugated goat anti-mouse IgG, rabbit antisera to caldesmon, vimentin, or tubulin,
Role of Nonmuscle Caldesmon 111
and finally TRTTC-conjugated goat anti-rabbit IgG, with 30 min washes in two changs of phosphate-buffered sa- line (PBS) between each of these steps. After a last wash in PBS, coverslips were mounted on glass slides, and viewed and recorded with a Zeiss epifluorescence phot- omicroscope 111 as described previously [Hegmann et al., 19891. To visualize actin filaments, injected cells were fixed and permeabilized as before, incubated for 30 ’ .5 * min in FITC-conjugated goat anti-mouse IgG, washed in 9 4 K * PBS, incubated in rhodamine-conjugated phalloidin (Molecular Probes Inc., Junction City, OR) for 20 min, 6 8 K * dipped into PBS and then distilled water, and mounted.
2 OOK*
A B C D E F
4 x
4Cald esrno n 4Lamin A
4 3 K * Western Blotting CEF cells grown on a 100 mm dish were washed
three times with PBS and harvested in 100 p1 of SDS PAGE gel sample buffer (2% SDS, 15% glycerol, 100 mM DTT, 80 mM Tris, pH 6.8, 0.001% bromophenol blue). After homogenizing by passage through a number 27 needle at least six times, the cell lysates were boiled at 100°C for 3-5 min and used for immunoblotting. Western blotting was performed as described previously
3 0 K *
[Lin et al., 19851. 21 K* 1 4 . 3 K *
RESULTS Monoclonal Antibody Specificity
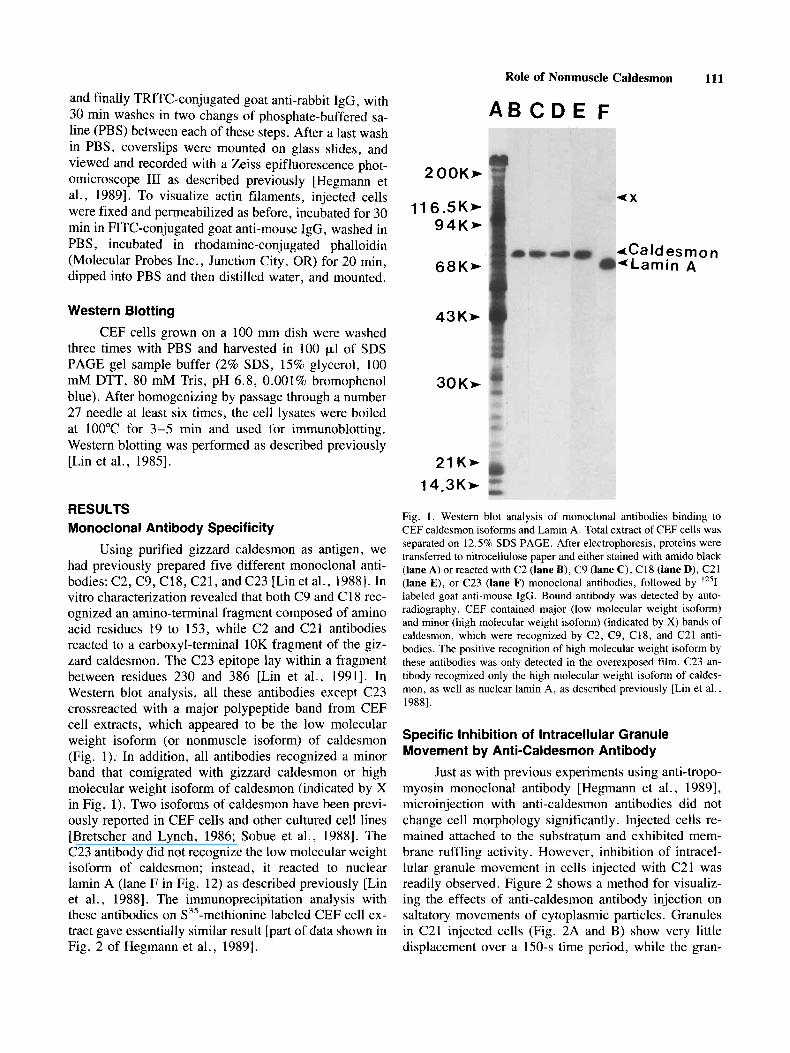
Using purified gizzard caldesmon as antigen, we had previously prepared five different monoclonal anti- bodies: C2, C9, C18, C21, and C23 [Lin et al., 19881. In vitro characterization revealed that both C9 and C 18 rec- ognized an amino-terminal fragment composed of amino acid residues 19 to 153, while C2 and C21 antibodies reacted to a carboxyl-terminal 10K fragment of the giz- zard caldesmon. The C23 epitope lay within a fragment between residues 230 and 386 [Lin et al., 19911. In Western blot analysis, all these antibodies except C23 crossreacted with a major polypeptide band from CEF cell extracts, which appeared to be the low molecular weight isoform (or nonmuscle isoform) of caldesmon (Fig. 1). In addition, all antibodies recognized a minor band that comigrated with gizzard caldesmon or high molecular weight isoform of caldesmon (indicated by X in Fig. 1). Two isoforms of caldesmon have been previ- ously reported in CEF cells and other cultured cell lines [Bretscher and Lynch, 1986; Sobue et al., 19881. The C23 antibody did not recognize the low molecular weight isofom of caldesmon; instead, it reacted to nuclear lamin A (lane F in Fig. 12) as described previously [Lin et al., 19881. The immunoprecipitation analysis with these antibodies on S3’-methionine labeled CEF cell ex- tract gave essentially similar result [part of data shown in Fig. 2 of Hegmann et al., 19891.
Fig. 1. Western blot analysis of monoclonal antibodies binding to CEF caldesmon isoforms and Lamin A. Total extract of CEF cells was separated on 12.5% SDS PAGE. After electrophoresis, proteins were transferred to nitrocellulose paper and either stained with amido black (lane A) or reacted with C2 (lane B), C9 (lane C), C18 (lane D), C21 (lane E), or C23 (lane F) monoclonal antibodies, followed by 1251- labeled goat anti-mouse IgG. Bound antibody was detected by auto- radiography. CEF contained major (low molecular weight isoform) and minor (high molecular weight isoform) (indicated by X) bands of caldesmon, which were recognized by C2, C9, C18, and C21 anti- bodies. The positive recognition of high molecular weight isoform by these antibodies was only detected in the overexposed film. C23 an- tibody recognized only the high molecular weight isoform of caldes- mon, as well as nuclear lamin A, as described previously [Lin et al., 19881.
Specific Inhibition of lntracellular Granule Movement by Anti-Caldesmon Antibody
Just as with previous experiments using anti-tropo- myosin monoclonal antibody [Hegmann et al., 19891, microinjection with anti-caldesmon antibodies did not change cell morphology significantly. Injected cells re- mained attached to the substratum and exhibited mem- brane ruffling activity. However, inhibition of intracel- Mar granule movement in cells injected with C21 was readily observed. Figure 2 shows a method for visualiz- ing the effects of anti-caldesmon antibody injection on saltatory movements of cytoplasmic particles. Granules in C21 injected cells (Fig. 2A and B) show very little displacement over a 150-s time period, while the gran-
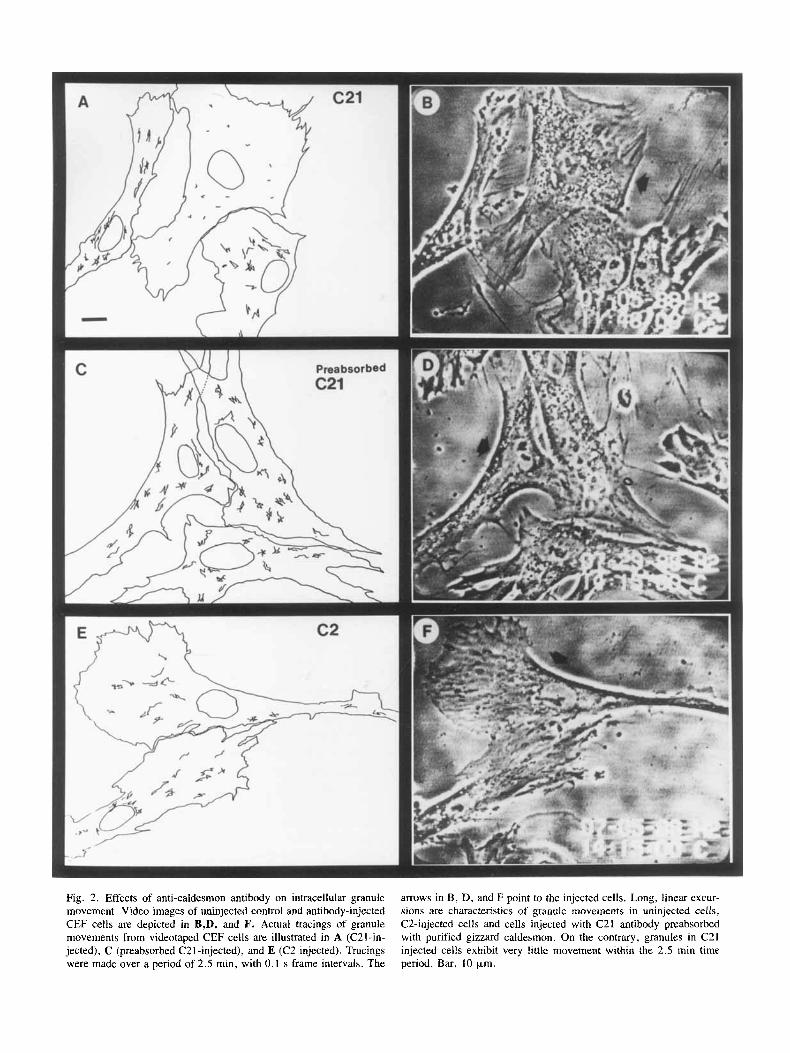
Fig. 2. Effects of anti-caldesmon antibody on intracellular granule movement. Video images of uninjected control and antibody-injected CEF cells are depicted in B,D, and F. Actual tracings of granule movements from videotaped CEF cells are illustrated in A (C21-in- jected), C (preabsorbed C21-injected), and E (C2-injected). Tracings were made over a period of 2.5 min, with 0.1 s frame intervals. The
arrows in B, D, and F point to the injected cells. Long, linear excur- sions are characteristics of granule movements in uninjected cells, C2-injected cells and cells injected with C21 antibody preabsorbed with purified gizzard caldesmon. On the contrary, granules in C21 injected cells exhibit very little movement within the 2.5 min time period. Bar, 10 p,m.
Role of Nonmuscle Caldesmon 113
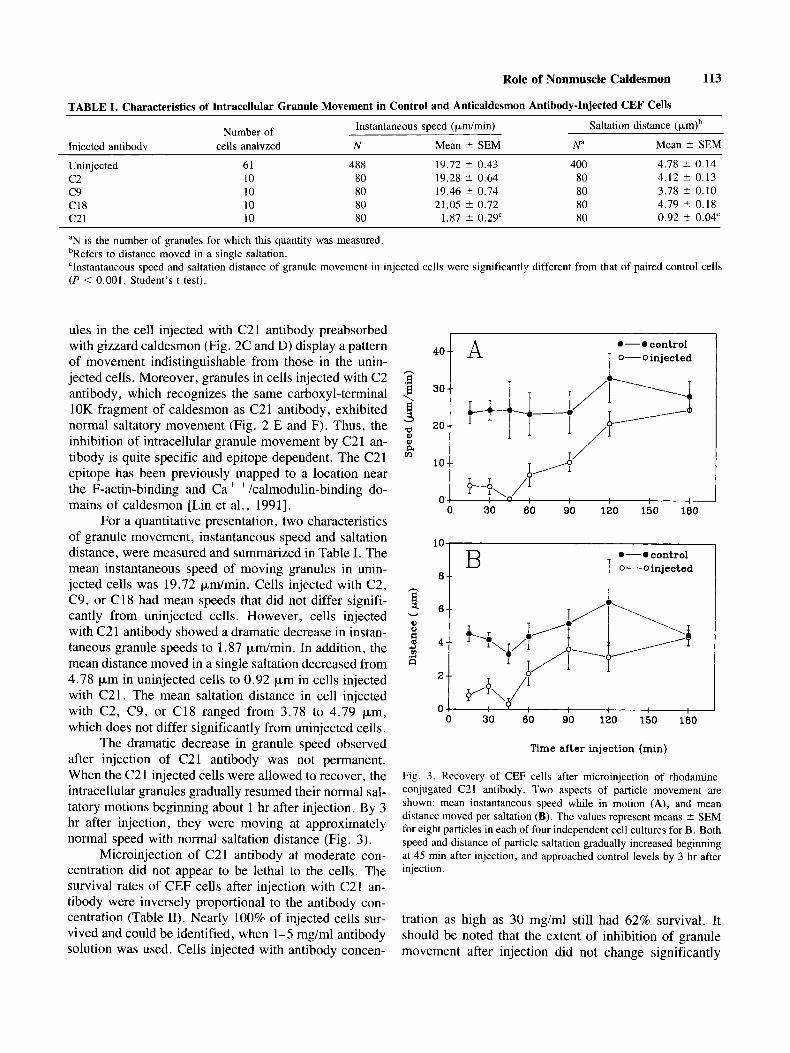
TABLE I. Characteristics of Intracellular Granule Movement in Control and Anticaldesmon Antibody-Injected CEF Cells Instantaneous speed (pdmin) Saltation distance (p,m)b
Number of cells analyzed N Mean 2 SEM N" Mean t SEM Iniected antibodv
400 4.78 k 0.14 Uninjected 61 488 19.72 * 0.43 80 4.12 t 0.13 c 2 10 80 19.28 -+ 0.64
c 9 10 80 19.46 t 0.74 80 3.78 2 0.10 C18 10 80 21.05 t 0.72 80 4.79 t 0.18 c 2 1 10 80 1.87 t 0.29' 80 0.92 ? 0.04'
"N is the number of granules for which this quantity was measured. bRefers to distance moved in a single saltation. 'Instantaneous speed and saltation distance of granule movement in injected cells were significantly different from that of paired control cells (P < 0.001, Student's t test).
~~ ~
ules in the cell injected with C21 antibody preabsorbed with gizzard caldesmon (Fig. 2C and D) display a pattern of movement indistinguishable from those in the unin- jected cells. Moreover, granules in cells injected with C2 antibody, which recognizes the same carboxyl-terminal 10K fragment of caldesmon as C21 antibody, exhibited normal saltatory movement (Fig. 2 E and F). Thus, the inhibition of intracellular granule movement by C21 an- tibody is quite specific and epitope dependent. The C21 epitope has been previously mapped to a location near the F-actin-binding and Ca+ +/calmodulin-binding do- mains of caldesmon [Lin et al., 19911.
For a quantitative presentation, two characteristics of granule movement, instantaneous speed and saltation distance, were measured and summarized in Table I . The mean instantaneous speed of moving granules in unin- jected cells was 19.72 p d m i n . Cells injected with C2, C9, or C 18 had mean speeds that did not differ signifi- cantly from uninjected cells. However, cells injected with C21 antibody showed a dramatic decrease in instan- taneous granule speeds to 1.87 p d m i n . In addition, the mean distance moved in a single saltation decreased from 4.78 pm in uninjected cells to 0.92 pm in cells injected with C2 1. The mean saltation distance in cell injected with C2, C9, or C18 ranged from 3.78 to 4.79 pm, which does not differ significantly from uninjected cells.
The dramatic decrease in granule speed observed after injection of C21 antibody was not permanent. When the C21 injected cells were allowed to recover, the intracellular granules gradually resumed their normal sal- tatory motions beginning about 1 hr after injection. By 3 hr after injection, they were moving at approximately normal speed with normal saltation distance (Fig. 3).
Microinjection of C21 antibody at moderate con- centration did not appear to be lethal to the cells. The survival rates of CEF cells after injection with C21 an- tibody were inversely proportional to the antibody con- centration (Table 11). Nearly 100% of injected cells sur- vived and could be identified, when 1-5 mg/ml antibody solution was used. Cells injected with antibody concen-
.-@control T 0-oinjected
10 o-ocontrol
o-oinjected B 8 --
0 30 60 90 120 150 180
Time after injection (min)
Fig. 3. Recovery of CEF cells after microinjection of rhodamine- conjugated C21 antibody. Two aspects of particle movement are shown: mean instantaneous speed while in motion (A), and mean distance moved per saltation (B). The values represent means ? SEM for eight particles in each of four independent cell cultures for B. Both speed and distance of particle saltation gradually increased beginning at 45 min after injection, and approached control levels by 3 hr after injection.
tration as high as 30 mg/ml still had 62% survival. It should be noted that the extent of inhibition of granule movement after injection did not change significantly

114 Hegmann et al.
TABLE 11. Survival of CEF Cells After C21 Anticaldesmon Ca + +/calmodulin binding and with F-actin binding may Antibody Injection account for the inhibition of granule movement. The in- Injected-antibody Number of Survival jected cells shown in Figure 4 still contained few stress- concentration (mg/ml) cells injected (96) fibers as revealed by polyclonal antibody staining (Fig.
1 5
10 20 30
50 50
100 50
100
94 96
70 62
78
with the concentration of C21 antibody between 1 and 30 mg/ml .
Caldesmon Localization in Injected Cells As described in the accompanying paper, the C21
epitope is proximal to the functional domains for Ca+ +/ calmodulin-binding and F-actin-binding sites on gizzard caldesmon. Therefore, C21 antibody at high concentra- tion in the in vitro reconstitution system is able to strip caldesmon molecules from F-actin filaments and to com- pete with the binding of caldesmon to Ca+ + /calmodulin [Lin et al., 19911. The mechanism by which C21 anti- body interferes with intracellular granule movements may be through this same pathway.
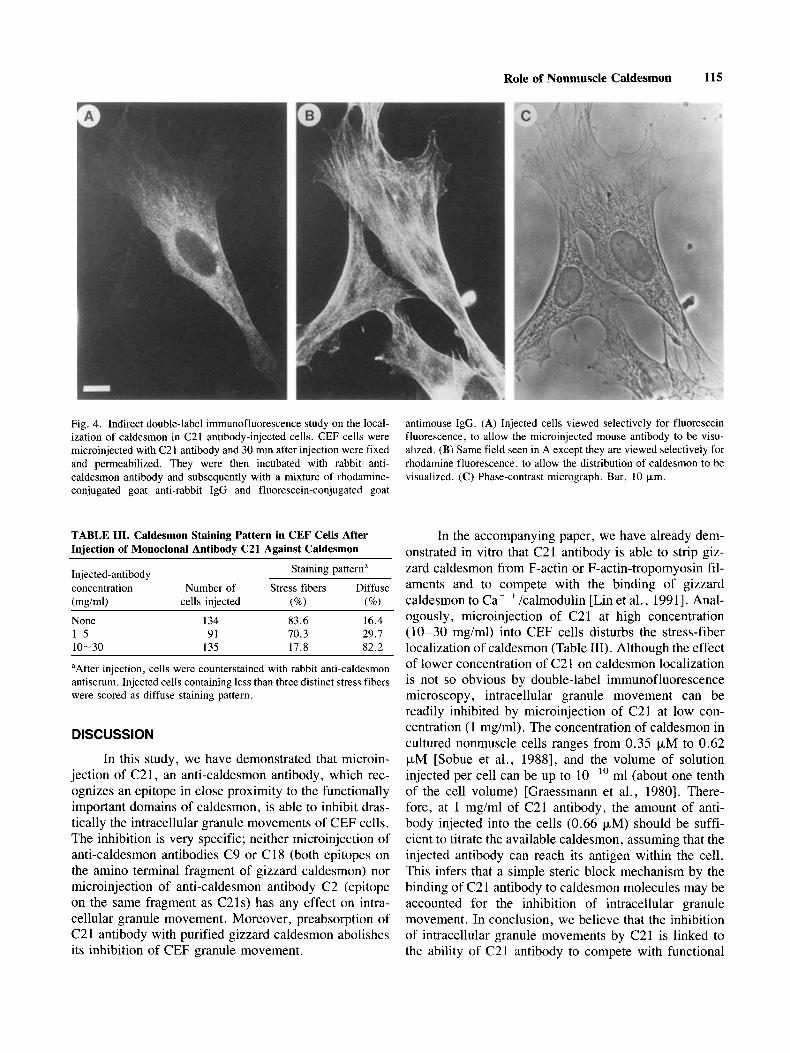
To investigate this idea, we have performed dou- ble-label immunofluorescence experiments to follow the fate of caldesmon molecules in C21 injected cells. At 30 min after microinjection with known amounts of C21 antibody, cells were fixed, permeabilized, and counter- stained with rabbit polyclonal anti-caldesmon antibody. The C21 injected cells were located by fluorescence mi- croscopy and were classified into two populations based on the staining pattern of the rabbit anti-caldesmon an- tibody. Injected cells containing less than three distinct stress fibers stained by rabbit anti-caldesmon antibody were scored as having a diffuse staining pattern; other- wise, they were judged to have a stress-fiber-like stain- ing pattern. Figure 4 shows an example of a stress-fiber- like staining pattern in a C21 injected cell. Using these criteria to score uninjected CEF cells, we found that 83.6% of the cells showed a stress-fiber staining pattern and 16.4% showed diffuse staining (Table 111). The cells injected with 1-5 mg/ml of C21 antibody had about 30% in the population of cells with diffuse staining pattern, and an even higher percentage (82.2%) of diffuse stain- ing was seen in cells injected with 10-30 mg/ml of C21 antibody. This result suggests that, just as in the in vitro system, C21 antibody at high concentration is able to disturb the intracellular localization of caldesmon. Since the inhibition of intracellular granule movements was seen in the cells injected with from 1 to 30 mg/ml of C21 antibody, the ability of C21 antibody to interfere with
4B). In contrast, the injected C21 antibody showed-a diffuse localization (Fig. 4A). This difference in staining pattern was seen more prominently in the cells injected with low amounts of antibody and there was no signifi- cant improvement observed in the injected cells pre- treated with a 0.1% Triton X-100 prior to counterstain- ing. This is consistent with our in vitro observation that C21 antibody is able to strip off caldesmon molecules from actidactin-tropomyosin filaments without depoly- merization of filaments [Lin et al., 19911.
Locomotory Activity of C21 Injected Cell To examine whether anti-caldesmon antibody in-
jection has any effect on cell locomotion, NRK cells recovering at the edge of a wounded confluent mono- layer culture were microinjected with monoclonal anti- body against caldesmon. The behavior of injected cells was videotaped for more than 3 hr. The advantages of this system are that i) all cells migrate in the same di- rection, and ii) in the same microscopic field, 5 to 6 injected cells can be followed and analyzed at the same time. When migration of cells was traced at 1 hr intervals over a 3 hr period starting immediately after injection, there was no apparent difference in migration distance and direction between C2 1 -injected and uninjected cells or cells injected with C18 antibody (Fig. 5). Alterna- tively, cells were digitized at 2 min intervals from 30 min after injection for at least 15 time segments for each condition. Cell centroid, centroid movement, and direc- tion were analyzed with a computer-assisted system (Dy- namic Morphology System) [Soll, 19881. Again, there was no significant difference in these parameters in C21 injected cells and control C18 injected cells.
Microtubules, Microfilaments, and Intermediate Filaments in C21 Injected Cells
To examine whether the injection of C21 antibody has any effect on the cell’s filamentous networks, in- jected CEF cells were double-labeled with rabbit anti- bodies against vimentin or tubulin, or else stained with rhodamino-conjugated phalloidin to label microfilaments (Fig. 6). Microinjection of C21 antibody ranging from 1 mg/ml to 20 mg/ml did not appear to have any disruptive effects on the microfilament bundles as revealed by phal- loidin staining pattern (Fig. 6B). Similarly, there was no detectable disturbance of the microtubule and intermedi- ate filament networks in the C21 injected cell (Fig. 6E and H).
Role of Nonmuscle Caldesmon 115
Fig. 4. Indirect double-label immunofluorescence study on the local- ization of caldesmon in C21 antibody-injected cells. CEF cells were microinjected with C21 antibody and 30 min after injection were fixed and permeabilized. They were then incubated with rabbit anti- caldesmon antibody and subsequently with a mixture of rhodamine- conjugated goat anti-rabbit IgG and fluorescein-conjugated goat
antimouse IgG. (A) Injected cells viewed selectively for fluorescein fluorescence, to allow the microinjected mouse antibody to be visu- alized. (B) Same field seen in A except they are viewed selectively for rhodamine fluorescence, to allow the distribution of caldesmon to be visualized. (C) Phase-contrast micrograph. Bar, 10 pm.
TABLE 111. Caldesmon Staining Pattern in CEF Cells After Iniection of Monoclonal Antibody C21 Against Caldesmon
Staining pattern" Injected-antibody concentration Number of Stress fibers Diffuse (mg/ml) cells injected (%) (%I None 1-5 10-30
134 83.6 16.4 91 70.3 29.1
135 17.8 82.2
"After injection, cells were counterstained with rabbit anti-caldesmon antiserum. Injected cells containing less than three distinct stress fibers were scored as diffuse staining pattern.
DISCUSSION
In this study, we have demonstrated that microin- jection of C21, an anti-caldesmon antibody, which rec- ognizes an epitope in close proximity to the functionally important domains of caldesmon, is able to inhibit dras- tically the intracellular granule movements of CEF cells. The inhibition is very specific; neither microinjection of anti-caldesmon antibodies C9 or C18 (both epitopes on the amino terminal fragment of gizzard caldesmon) nor microinjection of anti-caldesmon antibody C2 (epitope on the same fragment as C21s) has any effect on intra- cellular granule movement. Moreover, preabsorption of C2 1 antibody with purified gizzard caldesmon abolishes its inhibition of CEF granule movement.
In the accompanying paper, we have already dem- onstrated in vitro that C21 antibody is able to strip giz- zard caldesmon from F-actin or F-actin-tropomyosin fil- aments and to compete with the binding of gizzard caldesmon to Cat +/calmodulin [Lin et al., 19911. Anal- ogously, microinjection of C21 at high concentration (10-30 mgiml) into CEF cells disturbs the stress-fiber localization of caldesmon (Table 111). Although the effect of lower concentration of C21 on caldesmon localization is not so obvious by double-label immunofluorescence microscopy, intracellular granule movement can be readily inhibited by microinjection of C21 at low con- centration (1 mg/ml). The concentration of caldesmon in cultured nonmuscle cells ranges from 0.35 pM to 0.62 pM [Sobue et al., 19881, and the volume of solution injected per cell can be up to lo-'' ml (about one tenth of the cell volume) [Graessmann et al., 19801. There- fore, at 1 mg/ml of C21 antibody, the amount of anti- body injected into the cells (0.66 pM) should be suffi- cient to titrate the available caldesmon, assuming that the injected antibody can reach its antigen within the cell. This infers that a simple steric block mechanism by the binding of C2 1 antibody to caldesmon molecules may be accounted for the inhibition of intracellular granule movement. In conclusion, we believe that the inhibition of intracellular granule movements by C21 is linked to the ability of C21 antibody to compete with functional
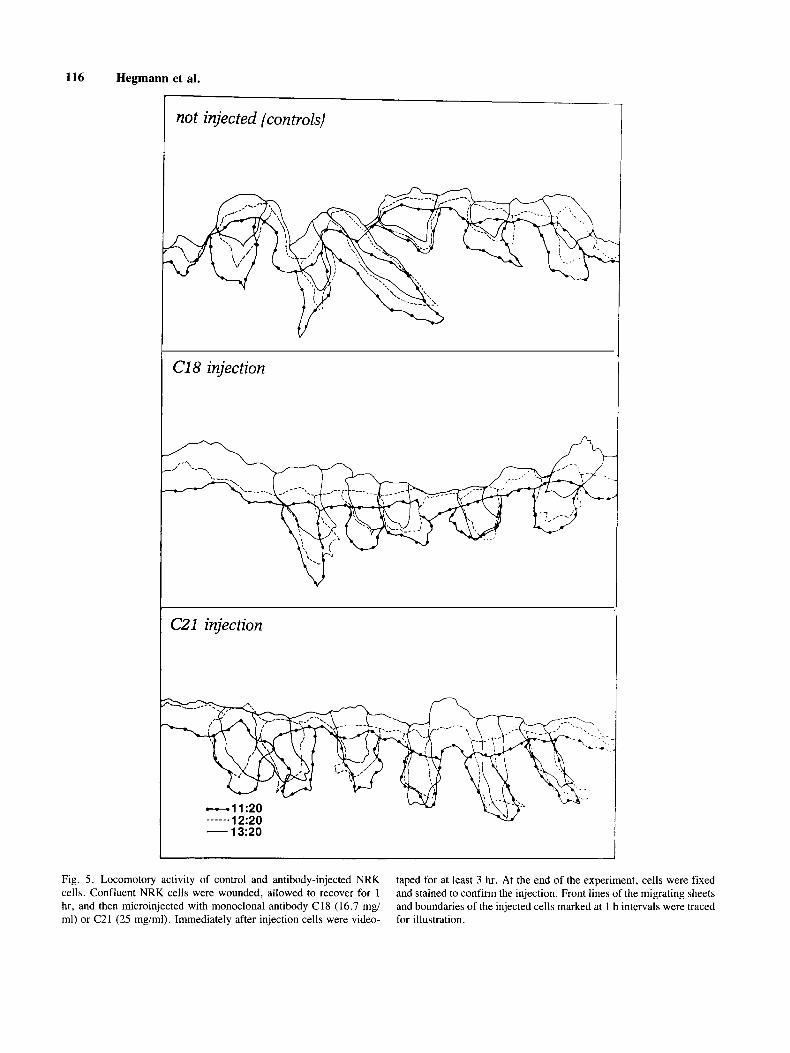
116 Hegmann et al.
not injected (controls)
C18 injection
CZl injection
- 13~20 I
Fig. 5 . Locomotory activity of control and antibody-injected NRK cells. Confluent NRK cells were wounded, allowed to recover for 1 hr, and then microinjected with monoclonal antibody C18 (16.7 mg/ ml) or C21 (25 mg/ml). Immediately after injection cells were video-
taped for at least 3 hr. At the end of the experiment, cells were fixed and stained to confirm the injection. Front lines of the migrating sheets and boundaries of the injected cells marked at 1 h intervals were traced for illustration.

Fig. 6. Indirect double-label immunofluorescence of C2 1 antibody injected cells. CEF cells were microinjected with C21 antibody and, 30 min after injection, were fixed and permeabilized. They were then incubated first with rhodamine-conjugated phalloidin (A-C), rabbit anti-tubulin antibody (D-F), or rabbit anti-vimentin antibody (G-I) and subsequently with a mixture of rhodamine-conjugated goat anti- rabbit IgG and fluorescein-conjugated goat anti-mouse IgG. (C, F, and I) phase-contrast micrographs. (A, D, and G) Injected cells
viewed selectively for fluorescein fluorescence, to allow the microin- jected mouse antibody to be visualized. (B, E, and H) Same fields seen in A, D, and G, respectively, except they are viewed selectively for rhodamine fluorescence, to allow the distribution of actin (B), tubulin (E), and vimentin (H) to be visualized. Note that apparently normal distributions of microfilament bundles, microtubules, and in- termediate filaments are observed in the C21 injected cells. Bar, 10 CLm.

118 Hegmann et al.
(actin-binding and Ca+ c calmoddin-binding) domains of caldesmon. Although our data do not allow us to distinguish clearly which domain is involved in intracel- lular granule movement, it is likely that both domains are required for the normal functions of caldesmon within CEF cells. In this regard, many investigators have pre- viously demonstrated by in vitro reconstitution studies that a concerted and opposite effect on the actin-myosin interaction can only be achieved through the binding of caldesmon to both actin filaments and Ca + +/calmodu- lin .
Anti-caldesmon antibody injection experiments have led us to suspect that nonmuscle caldesmon may play an essential role in regulating intracellular granule movement of nonmuscle cells. In support of this notion, Burgoyne et al. [1986] have previously discovered that a low molecular weight isoform of caldesmon appears to be associated with secretory granules of adrenal chro- maffin cells. Furthermore, both isolated chromaffin granules and lysosomal membranes have been demon- strated to interact with actin filaments in vitro [Araki and Ogawa, 1987b; Burridge and Phillips, 1975; Fowler and Pollard, 1982; Mehrabian et al., 19841. In addition, cy- tochalasins, which destabilize actin filaments, have been shown to inhibit lysosomal movements in rat alveolar macrophages [Araki and Ogawa, 1987al. CAMP-medi- ated inhibition of cellular translocation and intracellular particle movement is always accompanied by the rapid reorganization of actin filaments in Dictyostelium [Wes- sels et al., 19891. It has also been reported that micro- injection of DNase I, which binds to and depolymerizes actin, inhibits fast axonal transport [Goldberg et al., 1980; Isenberg et al., 19801 and that gelsolin in the pres- ence of C a + + inhibits the movement of membranous organelles in isolated axoplasm [Brady et al., 19841. All these results suggest that microfilaments are involved in intracellular granule movement. Additionally, we have recently shown that recognition of an epitope by CG1, a monoclonal anti-tropomyosin antibody, is dependent upon the motility state of the cell [Hegmann et al., 19881 and that microinjection of this CG1 antibody also inhibits intracellular granule movements CEF cells [Heg- mann et al., 19891. Therefore, nonmuscle tropomyosin and caldesmon may have a role in intracellular granule movement by regulating the contractile system of the cell in response to change in intracellular Ca+ + concentra- tion.
The pattern of the microtubule network is unaf- fected by the injection of anti-tropomyosin antibody CG1 [Hegmann et al., 19891 or anti-caldesmon antibody C21 (Fig. 6), while in both cases intracellular granule move- ment is greatly impeded. This in no way implies that cytoplasmic microtubules are not involved in intracellu- lar granule movement. On the contrary, there is a great
deal of evidence for the involvement of microtubules in organelle and granule transport [Dabora and Sheetz, 1988; Hayden, 1988; Sheetz et al., 1989; and see Heg- mann et al., 1989 for more references]. In particular, the recently identified microtubule-based motor molecules , kinesin and MAPIC/cytoplasmic dynein, which are able to promote the movement of membrane organelles on microtubules [Paschal et al., 1987; Vale et al., 19851, have clearly demonstrated the requirement for microtu- bule networks in organelle motility. However, experi- ments with purified protein reveal that kinesin alone is not sufficient for organelle motility; additional soluble factors are needed [Schroer et al., 1988; Sheetz et al., 19891. Moreover, there is evidence to suggest that the same factors are required for both kinesin- and cytoplas- mic dynein-driven organelle motility [Sheetz et al., 19891. What is the nature of these soluble factors? The results from our antibody injection studies provide strong evidence that intracellular granule movement requires actin filaments and their associated proteins, tropomyo- sin and caldesmon. It is possible that actin filaments and their associated proteins may represent the so-called sol- uble factors in microtubule-based organelle motility. The microfilament system may define low viscosity channels through the cytoplasm to facilitate organelle movement along the microtubules. In addition, it might provide a regulatory mechanism for directionality switching in re- sponse to Ca+ + concentration change.
Microinjection of either anti-tropomyosin or anti- caldesmon monoclonal antibodies into well-spread CEF cells does not seem to change cell shape or to inhibit locomotory activity. This is not surprising at all, since these antibodies do not induce breakdown of F-actin fil- aments in vivo or in vitro. In contrast, Honer et al. 119881 have shown that microinjection of monoclonal anti- myosin antibody into CEF cells induces breakdown of stress fibers, resulting in a drastic change of morphol- ogy. Moreover, after injection of anti-myosin antibodies, the cells show increased locomotory activity [Honer et al., 19881. Therefore, the actomyosin system appears to be involved in the maintenance of cell shape and the cell locomotion.
ACKNOWLEDGMENTS
We would like to thank R. E. Novy for critical reading and suggestions of the manuscript. This work was supported in part by grants HD18577, GM40580 from the National Institutes of Health, and by grants from the Muscular Dystrophy Association and the Pew Memorial Trust. Dr. J.J.-C. Lin is a recipient of a Pew Scholarship in Biomedical Sciences from the Pew Me- morial Trust.

Role of Nonmuscle Caldesmon 119
REFERENCES Araki, N., and Ogawa, K. (1987a): Regulation of intracellular lyso-
soma1 movements by the cytoskeletal system in rat alveolar macrophages. Acta Histochem. Cytochem. 20:659-678.
Araki, N. , and Ogawa, K. (1987b): In situ and in vitro morphological evidence for the interaction of actin filaments with lysosomes in rat alveolar macrophages. Acta Histochem. Cytochem. 20:
Berg, H.C., and Bloch, S.M. (1984): A miniature flow cell designed for rapid exchange of media under high-power microscope ob- jectives. I . Gen. Microbiol. 130:2915-2920.
Brady, S.T., Lasek, R.J., Allen, R.D., Yin, H.L., and Strossel, T.P. (1984): Gelsolin inhibition of fast axonal transport indicates a requirement for actin microfilaments. Nature 310:56-58.
Bretscher, A,, and Lynch, W. (1986): Identification and localization of immunoreactive forms of caldesmon in smooth and nonmus- cle cells: A comparison with the distributions of tropomyosin and a-actinin. J . Cell Biol. 100:1656-1663.
Burgoyne, R.D., Cheek, T.R., and Norman, K.-M. (1986): Identifi- cation of a secretory granule-binding protein as caldesmon. Nature 3 19:68-70.
Burridge, K., and Phillips, J.H. (1975): Association of actin and myosin with secretory granule membranes. Nature 254:526- 529.
Chalovich, J.M. (1988): Caldesmon and thin-filament regulation of muscle contraction. Cell Biophys. 12:73-85.
Chalovich, J.M., Cornelius, P., and Benson, C.E. (1987): Caldesmon inhibits skeletal actomyosin subfragment- 1 ATPase activity and the binding of myosin subfragment-1 to actin. J. Biol. Chem. 262571 1-5716.
Dabora, S.L., and Sheetz, M.P. (1988): Cultured cell extracts support organelle movement on microtubules in vitro. Cell Motil. Cy- toskeleton 10:482-495.
Dingus, J., Hwo, S . , and Bryan, J. (1986): Identification by mono- clonal antibodies and characterization of human platelet caldes- mon. J. Cell. Biol. 102:1748-1757.
Fowler, V.M., and Pollard, H.B. (1982): Chromaffin granule mem- brane: F-actin interactions are calcium sensitive. Nature 295:
Fujii, T., Ozawa, J., Ogoma, Y . , and Kondo, Y. (1988): Interaction between chicken gizzard caldesmon and tropomyosin. J. Bio- chem. 104:734-737.
Furst, D.O., Cross, R.A., deMey, J., and Small, J.V. (1986): Caldes- mon is an elongated flexible molecule localized in the actomy- osin domains of smooth muscle. EMBO J. 5:251-257.
Goldberg, D.J., Harris, D.A., Lubit, B.W., and Schwartz, J.H. (1980): Analysis of the mechanism of fast axonal transport by intracellular injection of potentially inhibitory macromolecules: Evidence for a possible role of actin filaments. Proc. Natl. Acad. Sci. U.S.A. 77:7448-7452.
Graessmann, A., Graessmann, M., and Mueller, C. (1980): Microin- jection of early SV40 DNA fragments and T-antigen. Meth. Enzymol. 652316-826.
Hayden, J.H. (1988): Microtubule-associated organelle and vesicle transport in fibroblasts. Cell Motil. Cytoskeleton 10:255- 262.
Hegmann, T.E., Lin, J.L.-C., and Lin, J.J.-C. (1988): Motility-de- pendence of the heterogeneous staining of culture cells by a monoclonal anti-tropomyosin antibody. J. Cell Biol. 106:385- 393.
Hegmann, T.E., Lin, J.L.-C., and Lin, J.J.-C. (1989): Probing the role of nommuscle tropomyosin isoforms in intracellular gran- ule movement by microinjection of monoclonal antibodies. J. Cell Biol. 109:1141-1152.
679-691.
336-339.
Honer, B., Citi, S . , Kendrick-Jones, J . , and Jockusch, B.M. (1988): Modulation of cellular morphology and locomotory activity by antibodies against myosin. J. Cell Biol. 107:2181-2189.
Horiuchi, K.Y., and Chacko, S. (1988): Interaction between caldes- mon and tropomyosin in the presence and absence of smooth muscle actin. Biochemistry 27:8388-8393.
Isenberg, G., Schubert, P., and Kreutzberg, G.W. (1980): Experi- mental approach to test the role of actin in axonal transport. Brain Res. 194:588-593.
Lin, J.J.-C., and Feramisco, J.R. (1981): Disruption of the in vivo distribution of the intermediate filaments in fibroblasts through the microinjection of a specific monoclonal antibody. Cell 24: 185-193.
Lin, J .J .X. , Matsumura, F., and Yamashiro-Matsumura, S. (1984): Tropomyosin-enriched and a-actinin-enriched microfilaments isolated from chicken embryo fibroblasts by monoclonal anti- bodies. J . Cell Biol. 98:116-127.
Lin, J J - C . , Chou, C.-S., and Lin, J .L.X. (1985): Monoclonal an- tibodies against chicken tropomyosin isoforms: production, characterization, and application. Hybridoma 4:223-242.
Lin, J . J .-C., Lin, J .L.-C., Davis-Nanthakumar, E. J . , and Lourim, D. (1988): Monoclonal antibodies against caldesmon, a Cat + / calmodulin- and actin-binding protein of smooth muscle and nonmuscle cells. Hybridoma 7:272-288.
Lin, J .J.-C., Davis-Nanthakumar, E.J., Jin, J.-P., Lourim, D., Novy , R.E., and Lin, J.L.-C. (1991): Epitope mapping of mono- clonal antibodies against caldesmon and their effects on the binding of caldesmon to Ca+ +/calmodulin and to actin or ac- tin-tropomyosin filaments. Cell Motil. Cytoskeleton 20:
Marston, S.B., and Smith, C.W.J. (1985): The thin filaments of smooth muscles. J . Muscl. Res. Cell. Motil. 6:669-708.
Mehrabian, M., Bame, K.J., and Rome, L.H. (1984): Interaction of rat liver lysosomal membranes with actin. J. Cell Biol. 99:
Mizushima, Y., Kanda, K., Hamaoka, T., Fujiwara, H., and Sobue, K. (1987): Resdistribution of caldesmon and tropomyosin as- sociated with concanavalin A receptor capping on splenic T- lymphocytes. Biomed. Res. 8:73-78.
Ngai, P.K., and Walsh, M.P. (1984): Inhibition of smooth muscle actin-activated myosin MgATPase activity by caldesmon. J . Biol. Chem. 259:13656-13659.
Paschal, B.M., Shpetner, H.S., and Vallee, R.B. (1987): MAPIC is a microtubule-activated ATPase which translocates microtubules in vitro and has dynein-like properties. J . Cell. Biol. 105: 1273-1282.
Schroer, T.A., Schnapp, B.J., Reese, T.S., and Sheetz, M.P. (1 988): The role of kinesin and other soluble factors in organelle move- ment along microtubules. J. Cell. Biol. 107:1785-1792.
Sheetz, M.P., Steur, E.R., and Schroer, T.A. (1989): The mechanism and regulation of fast axonal transport. Trend Neurosci. 12: 474 - 47 8.
Smith, C.W.J., Pritchard, K., and Marston, S.B. (1987): The mech- anism of Cat + regulation of vascular smooth muscle thin fil- aments by caldesmon and calmodulin. J. Biol. Chem. 262: 116-122.
Sobue, K., Muramoto, Y., Fujita, M., and Kakiuchi, S. (1981): Pu- rification of a calmodulin-binding protein from chicken gizzard that interacts with F-actin. Proc. Natl. Acad. Sci. U.S.A. 78: 5652-5655.
Sobue, K., Morimoto, K., Inui, M., Kanada, K., and Kakiuchi, S. (1982): Control of actin-myosin interaction of gizzard smooth muscle by calmodulin- and caldesmon-linked flip-flop mecha- nism. Biomed. Res., 3:188-196.
Sobue, K., Tanaka, T. , Kanda, K., Ashino, N., and Kakiuchi, S.
95-108.
680-685.

120 Hegmann et al.
(1985): Purification and characterization of caldesmon,,: A calmodulin-binding protein that interacts with actin filaments from bovine adrenal medulla. Proc. Natl. Acad. Sci. U.S.A. 825025-5029.
Sobue, K., Kanda, K., Tanaka, T., and Uebi, N. (1988): Caldesmon: A common actin-linked regulatory protein in the smooth mus- cle and nonmuscle contractile system. J. Cell. Biochem. 37: 3 17-325.
Soll, D.R. (1988): DMS (Dynamic Morphology System), a computer assisted system for quantitating motility, the dynamics of cy- toplasmic flow and pseudopod formation: Its application to Dictyostelium chemotaxis. Cell Motil. Cytoskeleton 10: 9 1- 106.
Vale, R.D., Reese, T.S., and Sheetz, M.P. (1985): Identification of a
novel force-generating protein, kinesin, involved in microtu- bule-based motility. Cell 42:39-50.
Walker. G., Kerrick, W.G.U., and Bourguignon, L.Y.W. (1989): The role of caldesmon in the regulation of receptor capping in mouse T-lymphoma cells. J. Biol. Chem. 264496-500.
Wessels, D., Schroeder, N.A., Voss, E., Hall, A.L., Condeelis, J . , and Soll, D.R. (1989): CAMP-mediated inhibition of intracel- Mar particle movement and actin reorganization in Dictoyo- stelium. J . Cell. Biol. 109:2841-2851.
Yamashiro-Matsumura, S. , and Matsumura, F. (1988): Characteriza- tion of 83-kilodalton nonmuscle caldesmon from cultured rat cell: Stimulation of actin binding of nonmuscle tropomyosin and periodic localization along microfilaments like tropomyo- sin. J. Cell. Biol. 106:1973-1983.
Copyright © 2022 FDOKUMEN




















