
Acute Restraint Stress in Zebrafish: Behavioral Parameters and Purinergic Signaling
Upload
independentCategory
view
3download
0
,1 ,1
a
*Departamento de Bioquımica, Facultad de Veterinaria, Universidad Complutense de Madrid, Madrid, Spain
�Departamento de Toxicologıa y Farmacologıa, Facultad de Veterinaria, Universidad Complutense de Madrid, Madrid, Spain
Glial cells and, particularly, astrocytes exhibit a form ofexcitability based on variations of the intracellular calciumconcentration (Perea and Araque 2005). Among the stimuliable to raise the intracellular calcium concentration, ATP andother nucleotides deserve special attention. These com-pounds are involved in intercellular communication in thebrain through the activation of specific membrane receptorscalled P2 purinoceptors (Fields and Burnstock 2006). Twodistinct subclasses of P2 receptors, namely the metabotropic,P2Y, and the ionotropic, P2X, families, are currently known.The former family, which includes eight subtypes withdifferent sensitivities for adenine and/or uridine nucleotidesare mainly coupled to phospholipase C (PLC) activation(Abbracchio et al. 2006), whereas the latter are ligand-operated channels that consist of homo- and hetero-oligo-mers formed by the assembly of different subunits (P2X1
through P2X7) (North 2002). Activation of P2X receptorscauses the opening of a channel permeable to small cations(Ca2+, Na+, and K+). Thus, intracellular calcium mobilization
constitutes one of the first steps in the signaling pathway ofboth metabotropic and ionotropic purinergic receptors. Thisfact together with the lack of specific pharmacological toolsfor the different receptor subtypes have made an arduous taskthe identification of the nucleotide receptors mediating thissort of effect in native tissues.
Received May 8, 2009; accepted May 11, 2009.Address correspondence and reprint requests to Esmerilda Garcıa
Delicado, Departamento de Bioquımica, Facultad de Veterinaria, Uni-versidad Complutense, 28040 Madrid, Spain.E-mail: [email protected] authors equally contributed to this work.Abbreviations used: A438079, 3-(5-(2,3-dichlorophenyl)-1H-tetrazol-
1-yl)methyl pyridine; BBG, brilliant blue G; BzATP, 3¢-O-(4-benzoyl)benzoyl ATP; MRS 2179, 2¢-deoxy-N6-methyl adenosine 3¢,5¢-diphos-phate diammonium salt; MRS 2211, pyridoxalphosphate-5¢-phosphate-6-azo-(-2-chloro-5-nitrophenyl)-2,4-disulfonate; PLC, phospholipase C;PPADS, pyridoxalphosphate-6-azophenyl-2¢,4¢-disulfonic acid.
Abstract
Previous work has established the presence of functional
P2X7 subunits in rat cerebellar astrocytes, which after stimu-
lation with 3¢-O-(4-benzoyl)benzoyl ATP (BzATP) evoked
morphological changes that were not reproduced by any other
nucleotide. To further characterize the receptor(s) and sig-
naling mechanisms involved in the action of BzATP, we have
employed fura-2 microfluorometry and the patch-clamp tech-
nique. BzATP elicited intracellular calcium responses that
typically exhibited two components: the first one was transient
and metabotropic in nature – sensitive to phospholipase C
inhibition and pertussis toxin treatment –, whereas the second
one was sustained and depended on the presence of extra-
cellular calcium. The ionotropic nature of this latter compo-
nent was corroborated by measurements of Mn2+ entry and
macroscopic non-selective cation currents evoked by either
BzATP (100 lM) or ATP (1 mM). The two components of the
calcium response to BzATP differed in their pharmacological
sensitivity. The metabotropic component was partially sensi-
tive to pyridoxalphosphate-5¢-phosphate-6-azo-(-2-chloro-
5-nitrophenyl)-2,4-disulfonate, a selective antagonist of P2Y13
receptors, while the ionotropic component was modulated by
external magnesium and markedly reduced by brilliant blue G
and 3-(5-(2,3-dichlorophenyl)-1H-tetrazol-1-yl)methyl pyridine
(A438079), thus implying the involvement of P2X7 purinergic
receptors. It is concluded that P2Y13 and P2X7 purinergic
receptors are functionally expressed in rat cerebellar astro-
cytes and mediate the increase in intracellular calcium elicited
by BzATP in these cells.
Keywords: astrocytes, BzATP, calcium signaling, glial cells,
ionotropic receptors, nucleotide receptors.
J. Neurochem. (2009) 110, 879–889.
JOURNAL OF NEUROCHEMISTRY | 2009 | 110 | 879–889 doi: 10.1111/j.1471-4159.2009.06179.x
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 879–889 879

Both P2X and P2Y receptors are expressed on all types ofbrain cells, including neurons, astrocytes, microglial cellsand oligodendrocytes. One of the P2X receptor subtypes, theP2X7 one, has attracted much interest for its uniqueproperties (Anderson and Nedergaard 2006). The P2X7
receptor was originally identified as the cytolytic P2Zreceptor of the monocyte-macrophage lineage and wasinitially considered exclusive of this immune lineage(Surprenant et al. 1996). However, over the last years, thepresence of functional P2X7 receptors has also beendescribed in a variety of cellular systems including neuralcells, in which they mediate, among other effects, celldifferentiation and proliferation as well as the modulation ofsynaptic transmission (Solini et al. 1999; Deuchars et al.2001; Fumagalli et al. 2003; Hervas et al. 2003; Miras-Portugal et al. 2003; Wang et al. 2004; Adinolfi et al. 2005;Dıaz-Hernandez et al. 2008).
In a preliminary study, we demonstrated the existence offunctional P2X7 subunits in rat cerebellar astrocytes, whichafter stimulation with the selective agonist 3¢-O-(4-ben-zoyl)benzoyl ATP (BzATP) experienced morphologicalchanges instead of undergoing cell lysis (Delicado et al.2005). In the current study we aim to go further into theunderstanding of the function of P2X7 receptors in thesecells, mainly by conducting a characterization of BzATP-induced changes in the intracellular calcium concentration(‘calcium responses’). We report that BzATP-induced cal-cium responses exhibit two components: the first one istransient and metabotropic in nature whereas the second oneis ionotropic, sustained or slowly rising during the stimula-tion period. Furthermore, non-selective ionic currents, pos-sibly underlying the ionotropic component of the calciumresponse, can also be recorded in cerebellar astrocytes uponBzATP application. The two components are mediated bydistinct purinergic receptors, identified as the P2Y13 andP2X7 subtypes. These results confirm the expression offunctional P2X7 receptors in cerebellar astrocytes from therat and extend the spectrum of actions of BzATP onpurinergic receptors.
Materials and methods
Astrocyte-purified culturesCerebella were obtained from 7- to 8-day-old Wistar rats and
astrocyte-purified cultures were established as previously described
(Jimenez et al. 1999). Briefly, cerebellar cells were dissociated by
trypsin digestion and astrocytes purified by orbital shaking. Once
plated on culture flasks, cerebellar astrocytes were grown until
confluence in Dulbecco’s modified Eagle’s medium supplemented
with 10% (v/v) fetal calf serum, 30 mM glucose, 100 U/mL
penicillin, 100 lg/mL streptomycin, 100 lg/mL kanamycin and
2.5 lg/mL amphotericin at 37�C in a humidified atmosphere of 5%
CO2/95% air. Finally, type 1 astrocytes were harvested by treating
the culture dish with a commercial trypsin/EDTA solution. Collected
cells were plated onto 15 mm-diameter coverslips in 35 mm Petri
dishes at a density of 3 · 105 cells in 3 mL of culture medium for
calcium imaging and at a density of 104 cells in 3 mL for
electrophysiological recordings. Cells were used within two days
after plating.
Fura-2 microfluorometry
Calcium imagingCalcium imaging experiments were carried out basically as
previously described (Carrasquero et al. 2005). Astrocytes attachedto coverslips were incubated in Locke’s solution (composition in
mM: 140 NaCl, 4.5 KCl, 2.5 CaCl2, 1.2 KH2PO4, 1.2 MgSO4, 5.5
glucose, 10 HEPES, pH 7.4) supplemented with 1 mg/mL bovine
serum albumin and loaded with 5–7 lM fura-2/AM for 45 min at
37�C. Once washed in fresh Locke¢s solution, a coverslip was
placed in a small superfusion chamber and the cells were imaged
through a Nikon TE-200 microscope (Nikon, Madrid, Spain) with a
Plan Fluor 20X/0.5 objective. Cells were illuminated at 340 and
380 nm and the emitted light isolated by a dichroic mirror (430 nm)
and a 510 nm band-pass filter (Omega Optical, Brattleboro, VT,
USA). Images were obtained with an ORCA-ER C 47 42–80 camera
from Hamamatsu (Hamamatsu City, Japan) controlled by MetaFluor
6.2r & PC software (Universal Imaging Corp., Cambridge, UK).
Exposure time was 100 ms and the change in the wavelength of
incoming light from 340 to 380 nm was carried out in less than
5 ms. Fluorescence images were acquired at a sampling frequency
of 2 Hz and processed by averaging signals from small elliptical
regions within individual cells. Background signals were subtracted
from each wavelength and the F340/F380 ratio was calculated. The
maximal effect of a given agonist concentration was estimated by
subtracting the basal F340/F380 value from the largest F340/F380observed following drug application (DF340/F380). EC50 values of
BzATP were derived from the best fit of a four parameter logistic
equation (to determine the basal and maximal responses, and EC50
and slope values) to concentration-response data by using Origin 7.0
software.
Manganese entryThe divalent cation Mn2+ was used as a Ca2+ surrogate to trace Ca2+
fluxes. Since the affinity of fura-2 for Mn2+ is very high, essentially
all Mn2+ ions entering a fura-2-loaded cell are trapped as fura-2-
Mn2+ complexes. These complexes are virtually non-fluorescent at
all wavelengths. Thus, the rate of loss of total fura-2 fluorescence
provides an estimate of the rate of Mn2+ influx into the cell,
irrespectively of the magnitude of calcium changes that may occur
simultaneously (Alonso et al. 1989). To conduct Mn2+ uptake
experiments, astrocytes plated on coverslips were loaded with fura-
2/AM as described above and, subsequently, placed in a superfusion
chamber containing Locke¢s solution, this saline being replaced by a
Ca2+ -free Locke¢s buffer containing 0.5 mM Mn2+ at the beginning
of fluorescence recording. Fluorescence images were obtained with
an Olympus IX inverted microscope equipped with a thermostatic
plate. Excitation and emission filters had similar spectra to those
described for calcium imaging experiments. Digital images were
taken with a Hamamatsu EM-CCD C9100 camera and analyzed
with the CellR software (Olympus Digital Imaging solutions,
Hamburg, Germany). Total fluorescence of fura-2 was calculated as
Ftot = (F340 + F380 · k), where k stands for the ratio of the values of
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 879–889� 2009 The Authors
880 | L. M. G. Carrasquero et al.

Fmax-Fmin at 340 nm and Fmax-Fmin at 380 nm (Alonso et al.1989). Ftot values were normalized to those calculated at the time of
introducing Mn2+ into the superfusate. The rate of Mn2+ entry was
estimated by fitting two linear functions over a 60 s period
immediately before and after agonist application. The change in
the rate of Mn2+ entry induced by agonists was calculated as the
difference between the slopes of the two lines and expressed as D%Ftot/min.
In microfluorometry experiments, drugs were added to Locke¢ssolution at 37�C and applied to the cells by bath superfusion.
Electrophysiological recordingsElectrophysiological recordings were performed with an EPC9
patch-clamp amplifier using the PULSE software (HEKA Elec-
tronic, Lambrecht, Germany). Pipettes were pulled from Kimax
borosilicate glass (Witz Scientific, Holland, OH, USA) and
subsequently wax-coated and fire-polished to obtain a final
resistance of 2–3 MW when filled with standard solutions. Mem-
brane currents were measured in either the whole-cell configuration
of the patch-clamp technique or its perforated-patch variant, filtered
at 3 kHz and sampled at 10 KHz. Once established electrical access
to the cytoplasm, cells were held at a voltage (Vh) of )70 mV and
those with holding currents larger than 20 pA (conventional whole-
cell) or series resistance above 30 MW (perforated-patch) were
rejected. Series resistance was compensated by 80% and monitored
throughout the experiment together with the cell membrane
capacitance.
The input resistance and the resting membrane potential of rat
cerebellar astrocytes were estimated by measuring in voltage-clamp
the current evoked by 20 mV hyperpolarizing/depolarizing pulses
from a Vh of )70 mV, and finding the potential at which current
level was zero during a 500 ms voltage ramp from )90 mV to
+50 mV, respectively. Reversal potential values from individual
cells were corrected for the junction potential between the pipette’s
solution and the extracellular solution, which was calculated to be
+4.42 mV by using the Patcher’s Tools module included in Igor-
Pro software (WaveMetrics, Inc. Lake Oswego, OR, USA).
Ligand-gated currents were activated by P2X receptor agonists
applied onto the cell under investigation by means of a gravity-
driven perfusion system with five independent lines controlled by
electronic valves (The Lee Company, Westbrook, CO, USA). This
system allowed the exchange of the medium surrounding a cell in
less than 100 ms. Current-voltage curves for BzATP-evoked
responses were obtained by sequentially stepping the holding
voltage for 2 s to the indicated values during the stationary phase of
the current response. Leak currents recorded at the same voltages
prior to BzATP application were subtracted from the corresponding
currents in the presence of the agonist.
All recordings were obtained at room temperature (21–24�C).
SolutionsThe extracellular (bath) solution contained (in mM) 140 NaCl, 2.8
KCl, 2 CaCl2, 10 HEPES and 10 glucose (pH 7.2, adjusted with
NaOH; 298 mOsm). Recording pipettes were filled with a solution
containing (in mM) 140 KCl, 1 MgCl2, and 10 HEPES (pH 7.2,
adjusted with KOH; 289 mOsm). The perforated-patch configura-
tion was obtained using amphotericin B (0.25 mg/mL). Amphoter-
icin B was dissolved in dimethylsulfoxide and stored at )20�C in
aliquots at 50 mg/mL. Fresh solution of amphotericin B in the
internal solution was prepared every 4 h. To more easily seal the
cells, the patch pipette was immersed a few seconds in an internal
solution without amphotericin B and then backfilled with the
internal solution containing amphotericin B. Cells attached to glass
coverslips were transferred to a recording chamber placed in the
stage of an inverted Zeiss Axiovert 100 microscope and continu-
ously superfused with bath solution (perfusion rate of 1 mL/min).
Stock solutions of drugs were diluted daily in extracellular saline
and incorporated into the perfusion system a few minutes before
starting the experiments.
Statistical analysisResults are expressed as the mean ± SEM calculated from at least
three experiments performed in cells from different cultures.
Comparisons were usually made by one-way ANOVA, and Dunnett
post-test analysis was only applied when a significant (p < 0.05)
effect was in evidence from ANOVA.
MaterialsAmphotericin B, bovine serum albumin, trypsin/EDTA solution,
nucleotides, brilliant blue G (BBG), 2¢-deoxy-N6-methyl adenosine
3¢,5¢-diphosphate diammonium salt (MRS 2179) and pyridoxalphos-
phate-6-azophenyl-2¢,4¢-disulfonic acid (PPADS) were purchased
from Sigma Chemicals (Alcobendas, Spain); A438079, 3-(5-(2,3-
dichlorophenyl)-1H-tetrazol-1-yl)methyl pyridine was from Abbott
Laboratories, Dulbecco’s modified Eagle’s medium and fetal calf
serum from Gibco (Barcelona, Spain). Pyridoxalphosphate-5¢-phos-phate-6-azo-(-2-chloro-5-nitrophenyl)-2,4-disulfonate (MRS 2211)
was from Tocris (Biogen Cientıfica, Madrid, Spain). U 73122, its
inactive analogue U 73343, and pertussis toxin were from Calbio-
chem (Madrid, Spain). Fura-2/AM was from Molecular Probes
(Invitrogen, Barcelona, Spain). All other reagents were from Merck
(Madrid, Spain).
All experiments were carried out at the Universidad Complutense
de Madrid following the guidelines of the International Council for
Laboratory Animal Science (ICLAS).
Results
The two main characteristics of P2X7 receptors are the lowaffinity – in the millimolar range – for the physiologicalligand, ATP, and the lack of desensitization in the continuouspresence of the agonist (North 2002). In previous studies, wedemonstrated that cerebellar astrocytes exhibited metabo-tropic purinergic calcium responses mediated by P2Y2/P2Y4
nucleotide receptors, at which ATP and UTP were equi-potent. These results, together with the insensitivity ofastrocytes to the P2X1 and P2X3 agonist, alpha-beta-methylene-ATP, led us to suggest that cerebellar astrocyteswould lack ionotropic P2X receptors capable of inducinga rise of intracellular calcium (Jimenez et al. 2000). Never-theless, those studies were carried out using ATP concentra-tions of up to 100 lM, which are enough to activate all P2Xreceptors except those of the P2X7 subtype. Taken this factinto consideration and in view of our recent results involving
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 879–889
Calcium responses to BzATP in individual astrocytes | 881

P2X7 receptors in morphological changes experienced bycerebellar astrocytes following treatment with BzATP(Delicado et al. 2005), we have analyzed calcium responsesin single cerebellar astrocytes evoked byBzATP and comparedthem with those triggered by mM concentrations of ATP.
Calcium responses evoked by nucleotides in singlecerebellar astrocytesThe initial characterization of calcium responses to nucleo-tides was performed by applying in random order ATP orBzATP at 8–10 min intervals. Practically, all tested cellsresponded to 1 mM ATP and to 100 lM BzATP stimula-tions. As shown in Fig. 1(a), the shape of ATP calcium
responses was nevertheless markedly different to thatobserved after BzATP applications. When challenged withATP, cells responded with elevations of intracellular calciumthat peaked (DF340/F380 of 0.28 ± 0.02, from basal levelsbetween 0.6 and 0.8; n = 372 cells) and returned to the basallevel within the first 2 min of stimulation. Calcium responsesto ATP exhibited a kinetics similar to that of responsesinduced by 100 lM UTP (data not shown), which indicatedthat ATP was mainly interacting with metabotropic P2Yreceptors (Jimenez et al. 2000). In contrast, BzATP calciumresponses were biphasic in the majority of the cells (83%),displaying an initial transient component (DF340/F380 of0.29 ± 0.02; n = 758 cells) followed by a sustained phase,which often rose in the continuous presence of the agonist.Calcium responses to BzATP were concentration-dependent.The EC50 of BzATP producing the initial transient andsustained components were 15.3 ± 3.5 lM and 80.9 ±6.8 lM, respectively (Fig. S1).
To investigate the metabotropic and/or ionotropic nature ofnucleotide responses, similar experiments were carried out inthe absence of extracellular calcium (by adding a 6 mMTris/6 mM EGTA mix to achieve a free calcium concentration inthe Locke¢s solution of about 200 nM). Extracellular calciumremoval had no significant effect (13% inhibition, p > 0.05)on calcium responses to ATP, thus suggesting it is metab-otropic. Interestingly, this maneuver abolished the secondcomponent of the calcium response to BzATP but leftunaffected the initial one, hence evidencing the existence ofboth a metabotropic and an ionotropic mechanism in theresponse to this nucleotide (Fig. 1a).
Additional evidence suggesting the involvement ofionotropic purinergic receptors, particularly of those belong-ing to the P2X7 subtype, in the calcium response to BzATPderived from experiments in which the extracellularconcentration of Mg2+ was modified. It is known thatextracellular Mg2+ inhibits P2X7-mediated responses bothby combining with ATP-, the active form of ATP on P2Xreceptors, and by an allosteric action on the receptor itself(Virginio et al. 1997; Acuna-Castillo et al. 2007). Eitherremoving or increasing the extracellular concentration ofMg2+ in the Locke’s solution produced a change in theshape of calcium responses to BzATP. So, in experimentsperformed in the same cells, deleting Mg2+ from the bathsolution induced an enlargement of the second componentof the response, whereas increasing the Mg2+ content from1.2 to 4.3 mM was associated to a pronounced reduction ofsuch a component (Fig. 1b).
Next, we examined the effect of BBG, a selectiveantagonist of the rat P2X7 receptor (Bianchi et al. 1999;Jiang et al. 2000), on calcium responses to ATP andBzATP. As it was the case when external calcium wasremoved, pre-incubation of astrocytes for 3 min with 1 lMBBG depressed the sustained component of BzATP calciumresponses while the transient component elicited by either
F34
0/F
380
0.7
0.8
0.9
1.0
1.1
1.2
1.3
100 μM BzATP
100 μM BzATP
0 Mg2+
100 μM BzATP
4.3 mM Mg2+
50 s
1.2 mM Mg2+
100 s
1 mM ATP
100 μM BzATP
1 mM ATP
EGTA
F34
0/F
380
F34
0/F
380
0.7
0.8
0.9
1.0
1.1
1.2
1.3
0.7
0.8
0.9
1.0
1.1
1.2
1.3
100 μM BzATP
EGTA
(b)
(a)
Fig. 1 Intracellular calcium responses to BzATP and ATP in single
cerebellar astrocytes. (a) Dependence on extracellular calcium of
calcium responses induced by BzATP (100 lM; upper panel) and ATP
(1 mM; lower panel). Cells were superfused with Ca2+-free Locke’s
solution (by adding Tris/EGTA buffer to get a free calcium concen-
tration of calcium concentration of about 200 nM) for 5 min before and
during nucleotide applications. (b) Modulation by extracellular Mg2+ of
calcium responses induced by BzATP. Calcium responses elicited by
successive stimulations with BzATP (100 lM) to a cell initially bathed
in standard saline (1.2 mM Mg2+) and subsequently exposed to an
external solution containing no Mg2+ or high Mg2+ (4.3 mM). Fura-2
fluorescence ratio (F340/F380) increases were taken as indicative of
rises in intracellular calcium concentration (see Methods). Drugs were
applied during the time indicated by horizontal bars. Recordings rep-
resent typical responses observed in > 200 cells from at least three
different cultures for each condition.
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 879–889� 2009 The Authors
882 | L. M. G. Carrasquero et al.

ATP or BzATP was for its major part preserved (Fig. 2).So, DF340/F380 corresponding to the initial transient evokedby ATP and BzATP were reduced by 28% and 30%,respectively. A qualitative similar effect was also observedby using the novel P2X7 receptor antagonist A438079. At aconcentration 1 lM, A438079 curtailed the persistentcomponent of the calcium response to ATP (1 mM) orBzATP (100 lM) while barely affecting the initial transientcomponent (17% and 14% inhibition, respectively) of theresponse (Fig. S2).
Mn2+ influx triggered by nucleotidesTo further prove the involvement of P2X7 receptors in thecalcium responses induced by nucleotides in cerebellarastrocytes, we conducted Mn2+ entry experiments. To thisend, cells were stimulated with 100 lM BzATP or 1 mMATP in Ca2+ -free Locke¢s solution containing 0.5 mM Mn2+
and total fluorescence decay determined as described inMethods. Since Mn2+ is a strong quencher of fura-2fluorescence, the fluorescence decay in fura-2 loaded cellsin the presence of extracellular Mn2+ is commonly taken asindicative of Mn2+ influx through Ca2+ -permeable channels.As shown in Fig. 3(a), unstimulated cells displayed avariable but continuous decrease in fluorescence (18.57 ±1.4% Ftot/min; n = 375 cells), reflecting basal Mn2+ entry.The addition of BzATP markedly accelerated the fluores-cence decay (50.34 ± 1.5% Ftot/min; n = 199 cells). Inter-estingly, in experiments carried out with 1 mM ATP, we
also observed an increase in the rate of Mn2+ entry(37.11 ± 0.83% Ftot/min; n = 176 cells), though this effectwas less pronounced than that induced by BzATP (Fig. 3aand b). More importantly, Mn2+ entry stimulated by eitherBzATP or ATP was markedly reduced by BBG, thusimplicating both nucleotides in the activation of P2X7
receptors in our cell system. It is also important to noticethat Mn2+ entry evoked by BzATP or ATP was not fullyblocked by BBG, implying the existence of other mecha-nisms, most likely capacitative calcium entry activated bydepletion of intracellular calcium stores, contributing to theionotropic component of the calcium response elicited by
100 μM BzATP
0.7
0.8
0.9
1.0
1.1
1.2
100 μM BzATP
1 μM BBG
0.7
0.8
0.9
1.0
1.1
1.2 1 mM ATP 1 mM ATP
1 μM BBG
F34
0/F
380
F34
0/F
380
100 s
Fig. 2 Effect of BBG on calcium responses to BzATP and ATP in
individual cerebellar astrocytes. Cells were exposed to BBG (1 lM)
3 min before and during the stimulation with BzATP (100 lM) or ATP
(1 mM). Drugs were applied during the time indicated by horizontal
bars. Recordings represent typical responses observed in 80 and 65
cells stimulated with BzATP or ATP, respectively.
0
10
20
30
BzATP BBG +BzATP
ATP BBG +ATP
* *
Δ %
Fto
t/min
100 μM BzATP
Fto
t (%
)F
tot (
%)
+ 1 μM BBG
0
20
40
60
80
100
30 s0
20
40
60
80
100
+ 1 μM BBG
1 mM ATP
(a)
(b)
Fig. 3 BzATP and ATP stimulate BBG-sensitive Mn2+ influx in cere-
bellar astrocytes. (a) Effect of different purinergic drugs on the decay
of total fura-2 fluorescence as an assay of Mn2+ influx. Cells were
stimulated with BzATP (100 lM) or ATP (1 mM) in a Ca2+-free Locke’s
solution containing 0.5 mM Mn2+, and the decay of total fura-2 fluo-
rescence (Ftot) calculated as described in Methods. Where indicated,
cells were treated with BBG (1 lM) for 3 min before and during
application of nucleotides. Traces show data from single cells stimu-
lated with BzATP alone or BzATP in the presence of BBG (upper
panel), and with ATP alone or ATP in the presence of BBG (lower
panel). (b) Net effect of BzATP (100 lM) or ATP (1 mM) in the rate of
Mn2+ influx in the absence and the presence of BBG (1 lM). Values
are means ± SEM of pooled data from 189 cells treated with BzATP
alone, 80 cells treated with BBG plus BzATP, 176 cells treated with
ATP alone, and 87 cells treated with BBG plus ATP. *p < 0.05.
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 879–889
Calcium responses to BzATP in individual astrocytes | 883
purinergic agonists in cerebellar astrocytes. In this latterregard, it is worth to mention that BBG (1 lM) did not affectcapacitative calcium entry as assayed by measuring theintracellular calcium rise that appears upon reintroducingexternal calcium in the bath solution of cells previouslytreated with BzATP (100 lM for 1 min) in Ca2+-free Locke¢ssolution (Fig. S3).
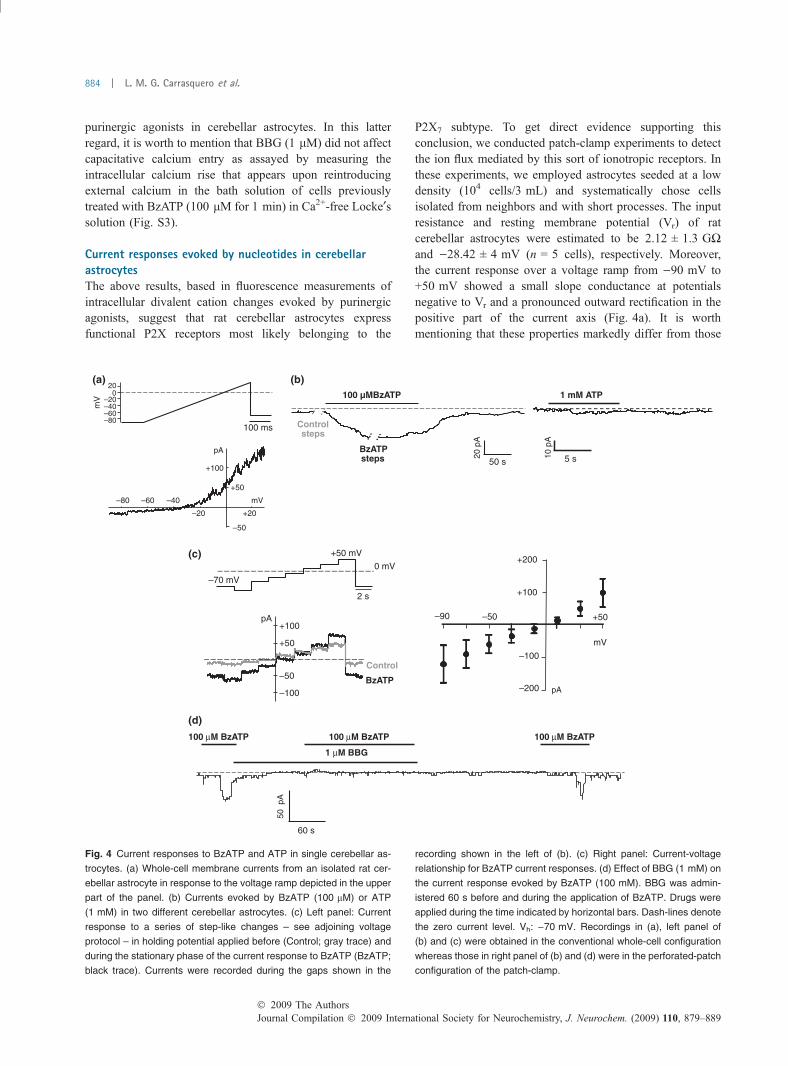
Current responses evoked by nucleotides in cerebellarastrocytesThe above results, based in fluorescence measurements ofintracellular divalent cation changes evoked by purinergicagonists, suggest that rat cerebellar astrocytes expressfunctional P2X receptors most likely belonging to the
P2X7 subtype. To get direct evidence supporting thisconclusion, we conducted patch-clamp experiments to detectthe ion flux mediated by this sort of ionotropic receptors. Inthese experiments, we employed astrocytes seeded at a lowdensity (104 cells/3 mL) and systematically chose cellsisolated from neighbors and with short processes. The inputresistance and resting membrane potential (Vr) of ratcerebellar astrocytes were estimated to be 2.12 ± 1.3 GWand )28.42 ± 4 mV (n = 5 cells), respectively. Moreover,the current response over a voltage ramp from )90 mV to+50 mV showed a small slope conductance at potentialsnegative to Vr and a pronounced outward rectification in thepositive part of the current axis (Fig. 4a). It is worthmentioning that these properties markedly differ from those
60 s
50 p
A
100 μM BzATP 100 μM BzATP 100 μM BzATP
1 μM BBG
–80–60–40–20
020
mV
100 ms
mV
+100
+50
pA
–50
–80 –60 –40
–20 +20
5 s 10 p
A
1 mM ATP100 µMBzATP
50 s20
pA
Controlsteps
BzATPsteps
+50
pA
–90 –50
mV
–200
–100
+100
+200
–100
–50
+50
+100pA
Control
BzATP
–70 mV
+50 mV
2 s
0 mV
(a) (b)
(c)
(d)
Fig. 4 Current responses to BzATP and ATP in single cerebellar as-
trocytes. (a) Whole-cell membrane currents from an isolated rat cer-
ebellar astrocyte in response to the voltage ramp depicted in the upper
part of the panel. (b) Currents evoked by BzATP (100 lM) or ATP
(1 mM) in two different cerebellar astrocytes. (c) Left panel: Current
response to a series of step-like changes – see adjoining voltage
protocol – in holding potential applied before (Control; gray trace) and
during the stationary phase of the current response to BzATP (BzATP;
black trace). Currents were recorded during the gaps shown in the
recording shown in the left of (b). (c) Right panel: Current-voltage
relationship for BzATP current responses. (d) Effect of BBG (1 mM) on
the current response evoked by BzATP (100 mM). BBG was admin-
istered 60 s before and during the application of BzATP. Drugs were
applied during the time indicated by horizontal bars. Dash-lines denote
the zero current level. Vh: )70 mV. Recordings in (a), left panel of
(b) and (c) were obtained in the conventional whole-cell configuration
whereas those in right panel of (b) and (d) were in the perforated-patch
configuration of the patch-clamp.
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 879–889� 2009 The Authors
884 | L. M. G. Carrasquero et al.

of the so-called ‘passive’ astrocytes but are very similar tothose reported for a subpopulation of astrocytes from mousehippocampus characterized by expressing a-amino-3-hydro-xy-5-methylisoxazole-4-propionate-type of glutamate recep-tors (Matthias et al. 2003).
Ligand-gated ionic currents were elicited by challengingthe cells with BzATP (100 lM) or ATP (1 mM) for varyingperiods of time, ranging from 10 to 300 s. BzATP applica-tions were associated with the appearance of inwardly-directed currents in 63% of the cells tested (33 out of 52). Nodifferences in the frequency of appearance of currentresponses were observed in relation with the recordingconfiguration used (conventional whole-cell or perforated-
patch), thus indicating that cell dialysis with the internalsolution did not influence the probability of detectingcurrents activated by BzATP. BzATP-induced currents acti-vated with variable latency ranging from 1 to 20 s, did notdesensitize in the presence of the agonist and could persistfor several seconds after stopping the application of thestimulus (Fig. 4b). Peak currents evoked by BzATP were of77.67 ± 12.44 pA (n = 33 cells), which considering anaverage membrane capacitance of 25.39 ± 1.77 pF (n = 52cells), implies a mean current density of about 3 pA/pF. Thepurinergic nature of BzATP-induced currents was evaluatedby assaying their sensitivity to BBG (1 lM). In cells pre-treated for 60 s with BBG, this drug fully and reversibly
100 μM BzATP 100 μM BzATP 100 μM BzATP
100 s
3 μM U 73122 3 μM U 73343
F34
0/F
380
0.7
0.9
1.1
1.3
0.5
100 s
100 μM BzATP
10 μM MRS 2179
100 μM BzATP
10 μM MRS 2211
100 μM BzATP
F34
0/ F
380
0.7
0.8
0.9
1.0
1.1
1.2
0
20
40
60
80
100
120
Contro
lU 7
3122
U 733
43PT
XPP
ADSM
RS 21
79M
RS 22
11
*** ***
**
% o
f tra
nsie
nt c
alci
umre
spon
se to
BzA
TP
0.7
0.8
0.9
1.0
1.1
1.2
F34
0/F
380
100 μM BzATP 100 μM BzATP
30 μM PPADS
0.8
0.9
1.0
1.1
1.2
F34
0/F
380
100 s
1 mM ATP 1 mM ATP
30 μM PPADS
0.7
(a)
(b)
(c)
(d)
Fig. 5 Characterization of the metabotropic component of the calcium
response to BzATP and ATP in cerebellar astrocytes. (a) Effect of U
73122 and U 73343 on calcium responses to BzATP. (b) Effect of
MRS 2179 and MRS 2211 on calcium responses to BzATP. (c) Effect
of PPADS on calcium responses to BzATP or ATP. Cells were treated
with each of the drugs (3 lM U 731223, 3 lM U 73343, 30 lM
PPADS, 10 lM MRS 2179 or 10 lM MRS 2211) for 3 min and sub-
sequently rechallenged with BzATP (100 lM) or ATP (1 mM) in the
presence of the same compound. Recordings represent typical res-
ponses. (d) Summarized effects of U 73122 (3 lM), U 73343 (3 lM),
pertussis toxin (100 ng/mL, overnight), PPADS (30 lM), MRS 2179
(10 lM) and MRS 2211 (10 lM) on the initial component of the
calcium response to BzATP (100 lM). Values are normalized with
respect to data obtained with BzATP alone. Values are means ± SEM
of pooled data from cells treated with BzATP alone (n = 758; Control),
U 73122 plus BzATP (n = 107; U 73122), U 73343 plus BzATP
(n = 118; U 73343), pertussis toxin plus BzATP (n = 94; pertussis
toxin), PPADS plus BzATP (n = 125; PPADS), MRS 2179 plus BzATP
(n = 86; MRS 2179) and MRS 2211 plus BzATP (n = 153; MRS
2211).**p < 0.01, ***p < 0.001, with respect to Control.
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 879–889
Calcium responses to BzATP in individual astrocytes | 885

suppressed the current response to BzATP (n = 6 cells)(Fig. 4d). Left panel in Fig. 4(c) shows the results of arepresentative experiment employed to estimate the reversalpotential of BzATP-activated currents. The current responseto a series of step-like changes (2 s long, in 20 mVincrements) in Vh applied before and during the stationaryphase of the response to BzATP are depicted. By digitalsubtraction of the current response in the absence of BzATPfrom that obtained in its presence, the net response to BzATPwas subsequently obtained. Then, the mean current duringeach voltage step was plotted against the correspondingvoltage and the reversal potential was estimated by curve-fitting. Current-voltage relationships were approximatelylinear between )90 and +50 mV, and reversed at+5.1 ± 2.5 mV (n = 3 cells; individual values corrected forthe junction potential) (Fig. 4c, right panel). Currentresponses to ATP (1 mM) applications were observed in asmall fraction of the cells tested (� 35%; n = 17 cells) and,on average, their amplitudes were half ()35.44 ± 5.08; n = 6cells) of that recorded after BzATP application (Fig. 4b).These results demonstrate that, in addition to mobilizecalcium from intracellular stores, purinergic agonists activateionotropic P2X7 receptors mediating non-selective cationcurrents in cerebellar astrocytes.
Characterization of the metabotropic component of thecalcium response to BzATP in cerebellar astrocytesConsidering that the initial transient component of calciumresponses to BzATP likely involved a metabotropic mech-anism, we decided to test the participation of PLC by usingthe PLC inhibitor, U 73122. After 3 min pre-incubation with3 lM U 73122, the calcium responses elicited by BzATPonly displayed the second component. Similar pre-incuba-tions carried out with 3 lM U 73343, an inactive analogue ofU 73122, did not modify the shape of calcium responses(Fig. 5a and d). Experiments involving the use of a puremetabotropic purinergic agonist, UTP, were also conducted.As expected, U 73122 completely prevented the calciumresponses to UTP (100 lM), whereas U 73343 had no effect(data not shown). Furthermore, pre-treatment of astrocyteswith pertussis toxin (100 ng/mL, overnight) slightly reducedthe size of the initial component of BzATP calciumresponses, hence suggesting the involvement of a Gq/11
protein (Fig. 5d).In an attempt to identify the receptor(s) mediating the
metabotropic effect of BzATP, we tested several purinergicantagonists, other than BBG. First, we evaluated the effect ofPPADS. PPADS has been reported to depress responsesmediated by metabotropic (P2Y1, P2Y4, P2Y13) as well asionotropic (P2X1-3, P2X5, P2X7) receptors (Communi et al.1996, 1999; Chessell et al. 1998a; Garcıa-Lecea et al. 1999;North 2002; von Kugelgen 2006). Figure 5(b) shows thatboth, the metabotropic and ionotropic, components of theresponse to BzATP were suppressed by PPADS (10 lM),
whereas the response to ATP remained unaffected. Theseresults are consistent with our previous findings indicatingthat ATP activates the PPADS-insensitive P2Y2 nucleotidereceptors in cerebellar astrocytes (Jimenez et al. 2000), andrule out the involvement of such a receptor in the metab-otropic response triggered by BzATP. Since P2Y1 and/orP2Y13 receptors are expressed in our preparation (Jimenezet al. 2000; Carrasquero et al. 2005), we decided to explorewhether these receptors could be the targets of BzATP. Tothis aim, we assayed the effect of MRS 2179, a selectiveP2Y1 antagonist, and MRS 2211, a selective P2Y13 antag-onist, on BzATP calcium responses. As it is shown inFig. 5(c) and (d), pre-treatment of the cells for 3 min withMRS 2179 (10 lM) did not significantly modify BzATPcalcium responses. On the contrary, MRS 2211 (10 lM)depressed both the early and sustained components of thecalcium response to BzATP. Strikingly, MRS 2211 alsoreduced the rate of Mn2+ entry induced by BzATP (Fig. 6).While the effect of MRS 2211 on the calcium response isconsistent with P2Y13 receptors being targeted by BzATP, thelarge effect of this antagonist on Mn2+ entry also pointstoward a potential blocking effect of MRS 2211 on P2X7
receptors. On the other hand, although MRS 2211 does nor
0
20
40
60
80
100
Fto
t (%
)Δ
% F
tot/m
in
30 s
100 μM BzATP
+ 10 μM MRS 2211
0
10
20
30
BzATP MRS 2211 +BzATP
*
(a)
(b)
Fig. 6 Effect of MRS 2211 on Mn2+ influx stimulated by BzATP in
cerebellar astrocytes. (a) Decay of total fura-2 fluorescence (Ftot) of
astrocytes stimulated with BzATP (100 lM) in the absence and the
presence of MRS 2211 (10 lM). Cells were bathed in a Ca2+-free
Locke’s solution containing 0.5 mM Mn2+ and treated with MRS 2211
for 3 min before and during BzATP application. Traces show averaged
data from 199 and 135 cells stimulated with BzATP in the absence or
presence of MRS 2211, respectively. (b) Effect of BzATP (100 lM) in
the rate of Mn2+ influx in the absence or presence of MRS 2211
(1 lM). *p < 0.05.
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 879–889� 2009 The Authors
886 | L. M. G. Carrasquero et al.

inhibit directly capacitative calcium entry (see Fig. S3), it isexpected to inhibit this sort of calcium entry indirectly as aconsequence of a diminished intracellular calcium mobiliza-tion brought about by a weaker metabotropic stimulation.
Discussion
In this study, we report that BzATP-induced calciumresponses exhibit a biphasic kinetics, which is not repro-duced by any other nucleotide tested so far in cerebellarastrocytes (ATP, UTP, 2-methylthioadenosine diphosphate,2-methylthioadenosine triphosphate, alpha-beta-methylene-ATP) (Jimenez et al. 2000). While the early component ofthe calcium response showed the usual features of ametabotropic mechanism – lack of dependence on extra-cellular calcium, sensitivity to PLC inhibition and pertussistoxin treatment – the delayed but sustained componentexhibited all the expected characteristics of an ionotropicresponse: dependence on external calcium and temporalcorrelation with transmembrane ion fluxes evidenced byboth microfluorometric and electrophysiological techniques.Moreover, its modulation by external Mg2+ and sensitivityto BBG, A438079 and PPADS, specifically implicate P2X7
receptors in the calcium response to BzATP.Rat cerebellar astrocytes co-express P2Y1, P2Y2, P2Y6
and P2Y13 receptors, but neither P2Y11 nor P2X1–P2X6
receptors are present in these cells (Jimenez et al. 2000;Carrasquero et al. 2005). In addition, we have recentlydescribed the expression in this experimental system of aP2X7-like receptor (Delicado et al. 2005). Hence, the resultspresented in this report extend and confirm our own findingsregarding the existence of functional P2X7 receptors incerebellar astrocytes. Our study concentrated in the initialevents (increase in plasma membrane permeability, calciumentry and elevation of intracellular calcium levels) of thesignaling pathway of ionotropic purinergic receptors. Byusing fura-2 microfluorometry to monitor changes in intra-cellular calcium or Mn2+ entry, and patch-clamp to recordwhole-cell ionic currents, we have demonstrated that BzATPand also high concentrations (1 mM) of ATP are capable ofeliciting an ionotropic response in cerebellar astrocytes fromthe rat. Moreover, the pharmacological and biophysicalevidence obtained, particularly the sensitivity to BBG andexternal Mg2+, and the lack of cationic selectivity of thechannels (reversal potential close to 0 mV), support thenotion that BzATP-induced current is mediated by genuineP2X receptors, most likely belonging to the P2X7 subtype.
Our results differ, in part, to those commonly reported inthe literature, in which BzATP calcium responses areabrogated by quelation of extracellular calcium (Solini et al.1999; Parvathenani et al. 2003; Hervas et al. 2005).Although BzATP is widely used for the characterization ofP2X7 receptors, it also activates other nucleotide receptors. Inthis regard, Bianchi and colleagues reported that BzATP is a
more potent agonist at the P2X1 and P2X3 receptors (nMrange) than at the P2X7 ones (lM range) (Bianchi et al.1999). Likewise, BzATP can also act as an agonist orantagonist of P2Y11 and P2Y1 receptors, respectively(Communi et al. 1999; Vigne et al. 1999). In cerebellarastrocytes, BzATP not only stimulated Ca2+ entry from theextracellular space but also mobilized Ca2+ from intracellularstores. This type of effect was observed in the vast majorityof the cells and could be unequivocally separated from thecomponent of the calcium response mediated by P2X7
receptors. Besides removing external calcium, the use ofU-compounds was instrumental in dissecting the two com-ponents of the response both from the kinetic and mecha-nistic point of view. So, the transient component waspreserved by removing external calcium but was selectivelyaffected by treatment with U 73122. Likewise, this com-pound was similarly effective when tested against UTP, apure metabotropic purinergic agonist. Importantly, U 73343neither modified the transient nor the sustained component ofthe response to BzATP, hence ruling out any PLC-indepen-dent action of the U-compounds, as that reported on P2X7
receptors from a mouse microglial cell line (Takenouchiet al. 2005).
We have also addressed the question of the receptor(s)presumably involved in this metabotropic component of thecalcium response to BzATP. This component was barelyaffected by treatment with BBG and A438079, a result thatcan be explained by a partial overlap between the metabo-tropic and ionotropic components of the response. At vari-ance, the non-selective P2 antagonist, PPADS, suppressedthe transient component of the response to BzATP but not ofthat induced by ATP, hence indicating that BzATP and ATPactivate different metabotropic receptors. This result, whichis in good agreement with the expression of diversemetabotropic purinergic receptors in rat cerebellar astrocytes,supports the contention that ATP mainly activates PPADS-insensitive P2Y2 receptors in this cell system. A furtherattempt to pharmacologically identify the P2Y receptormediating the effect of BzATP relied on the use of novelcompounds, namely MRS 2179 and MRS 2211, withpurported selectivity to block P2Y1 and P2Y13 receptors,respectively. The lack of effect of MRS 2179 on BzATPcalcium responses allows the P2Y1 receptor to be discarded.In contrast, MRS 2211 markedly reduced the transientcomponent of the calcium response to BzATP, therebyinvolving P2Y13 receptors in its generation. In this regard, itis worth mentioning that the presence of P2Y13 receptors incerebellar astrocytes and the blocking effectiveness of MRS2211 have been corroborated by the ability of this compoundto suppress the calcium response to 2-methylthioadenosinediphosphate, a selective agonist of P2Y1 and P2Y13 receptors(Carrasquero et al. unpublished results). It should be noticed,however, that the effect of MRS 2211 was a partial one,which leads to consider that other metabotropic receptors
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 879–889
Calcium responses to BzATP in individual astrocytes | 887

could participate in the initial calcium response to BzATPtoo. MRS 2211 also reduced the sustained component of thecalcium response and the entry of Mn2+ induced by BzATP.Taking into account that MRS 2211 is a PPADS derivative(Kim et al. 2005), the latter effect could, at least in part, bebecause of the antagonism of P2X7 receptors. While thispossibility should be investigated in a cell model expressingP2X7 receptors in isolation – i.e. HEK293 cells or 1321N1astrocytoma cells (Chessell et al. 1998b; Gendron et al.2003) – we also endorse the interpretation that by limiting theproduction of inositol (1,4,5) trisphosphate mediated byP2Y13 receptors, MRS 2211 would also reduce Mn2+ entrythrough store-operated channels. It should be reminded thatactivation of this sort of channels can be an all-or-noneprocess, requiring large concentrations of inositol (1,4,5)trisphosphate (Parekh et al. 1997). Thus, a reduced activa-tion of a store-operated calcium channels and of the ensuedcapacitative calcium entry could account for a significant partof the effect of MRS 2211 on the sustained component ofBzATP calcium response.
In this respect, our data provide evidence favoring theexistence of a capacitative calcium entry following intracel-lular calcium store depletion in cerebellar astrocytes. Thisstatement is based on measurements of intracellular calciumincreases elicited by reintroducing calcium in the bathsolution of cells previously exposed to BzATP in the absenceof external calcium. Two types of pharmacological evidencefurther suggest that capacitative calcium entry contribute tocalcium entry following metabotropic purinergic receptorstimulation: i) despite that BBG suppresses ionic currentsinduced by BzATP, it only partially blocks Mn2+ entryevoked by this nucleotide, and ii) neither BBG nor MRS2211 directly block capacitative calcium entry triggered bycalcium reintroduction. Defining the relative contribution ofcapacitative calcium entry secondary to intracellular storedepletion and of P2X7 receptor-mediated calcium entry to theelevation of intracellular calcium elicited by a mixed,metabotropic and ionotropic purinergic agonist, like BzATP,would nevertheless require of additional experiments.
The actions of BzATP on the calcium signal of cerebellarastrocytes can be easily fitted into a model of nearlysimultaneous activation of both metabotropic and ionotropicpurinergic receptors. On a short-term basis, intracellularcalcium release and calcium entry would possibly cooperatein shaping the calcium signal and sustaining its oscillatorybehavior over longer periods of time (Haydon 2001;Fumagalli et al. 2003); moreover, we also envisage asynergistic role of the two classes of purinergic receptors inthe control of astrocyte¢s secretory response and, thus, ofintercellular communication within brain (Duan et al. 2003;Suadicani et al. 2006; Bowser and Khakh 2007). Last, P2X7
receptors may act on a longer-term basis by inducingmorphological changes that possibly underlie reactive gliosisafter cerebral trauma or ischemia (Franke et al. 2004).
Through the expression of a diversity of purinergic receptorscoupled to distinct signaling pathways and displayingdifferent affinities for endogenous agonists, astrocytes, maydynamically adapt their activity to subtle variations in thenucleotidic input.
Acknowledgements
This work was supported by grants from the Ministerio
de Educacion y Ciencia (BFU2008-02699; BFU2005-0634),
Comunidad de Madrid (S-SAL-0253-2006), Fundacion Marcelino
Botın and the Spanish Ion Channel Initiative (CSD2008-00005).
L.M.G. was a research fellow of Ministerio de Educacion y Ciencia,
D.B. is a research fellow of Basque Government and Y.G.M. has a
post-doctoral contract (SAL-0253-2006) fromComunidad deMadrid.
Supporting Information
Additional Supporting Information may be found in the onlineversion of this article:
Figure S1. Concentration-response curves for the calciumresponse to BzATP.
Figure S2. Effect of A438079 on calcium responses toBzATP and ATP in individual cerebellar astrocytes.
Figure S3. Effect of BBG and MRS 2211 on capacitativecalcium entry evoked by BzATP in rat cerebellar astrocytes.
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting materials supplied bythe authors. Any queries (other than missing material) should bedirected to the corresponding author for the article.
References
Abbracchio M. P., Burnstock G., Boeynaems J. M. et al. (2006) Inter-national Union of Pharmacology LVIII: update on the P2Y Gprotein-coupled nucleotide receptors: from molecular mechanismsand pathophysiology to therapy. Pharmacol. Rev. 58, 281–341.
Acuna-Castillo C., Coddou C., Bull P., Brito J. and Huidobro-Toro J. P.(2007) Differential role of extracellular histidines in copper, zinc,magnesium and proton modulation of the P2X7 purinergic receptor.J. Neurochem. 101, 17–26.
Adinolfi E., Callegari M. G., Ferrari D., Bolognesi C., Minelli M.,Wieckowski M. R., Pinton P., Rizzuto R. and Di Virgilio F. (2005)Basal activation of the P2X7 ATP receptor elevates mitochondrialcalcium and potential, increases cellular ATP levels, and promotesserum-independent growth. Mol. Biol. Cell 16, 3260–3272.
Alonso M. T., Sanchez A. and Garcia-Sancho J. (1989) Monitoring ofthe activation of receptor-operated calcium channels in humanplatelets. Biochem. Biophys. Res. Commun. 162, 24–29.
Anderson C. M. and Nedergaard M. (2006) Emerging challenges ofassigning P2X7 receptor function and immunoreactivity in neurons.TINS 29, 257–262.
Bianchi B. R., Lynch K. J., Touma E. et al. (1999) Pharmacologicalcharacterization of recombinant human and rat P2X receptor sub-types. Eur. J. Pharmacol. 376, 127–138.
Bowser D. N. and Khakh B. S. (2007) Vesicular ATP is the predominantcause of intercellular calcium waves in astrocytes. J. Gen. Physiol.129, 485–491.
Carrasquero L. M., Delicado E. G., Jimenez A. I., Perez-Sen R. andMiras-Portugal M. T. (2005) Cerebellar astrocytes co-express
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 879–889� 2009 The Authors
888 | L. M. G. Carrasquero et al.

several ADP receptors. Presence of functional P2Y13-like recep-tors. Purinergic Signal. 1, 153–159.
Chessell I. P., Michel A. D. and Humphrey P. P. (1998a) Effects ofantagonists at the human recombinant P2X7 receptor. Br. J.Pharmacol. 124, 1314–1320.
Chessell I. P., Simon J., Hibell A. D., Michel A. D., Barnard E. A. andHumphrey P. P. (1998b) Cloning and functional characterisation ofthe mouse P2X7 receptor. FEBS Lett. 439, 26–30.
Communi D., Motte S., Boeynaems J. M. and Pirotton S. (1996) Phar-macological characterization of the human P2Y4 receptor. Eur. J.Pharmacol. 317, 383–389.
Communi D., Robaye B. and Boeynaems J. M. (1999) Pharmacologicalcharacterization of the human P2Y11 receptor. Br. J. Pharmacol.128, 1199–1206.
Delicado E. G., Carrasquero L. M., Perez-Sen R. and Miras-PortugalM. A. (2005) Identification of functional P2X7 receptor in ratcerebellas astrocytes. International Proceedings of VII EuropeanMeeting on Glial Cell Function in Health and Disease, pp. 71–78.Medimond, Bologna.
Deuchars S. A., Atkinson L., BrookeR. E.,MusaH.,Milligan C. J., BattenT. F., Buckley N. J., Parson S. H. and Deuchars J. (2001) NeuronalP2X7 receptors are targeted to presynaptic terminals in the centraland peripheral nervous systems. J. Neurosci. 21, 7143–7152.
Dıaz-Hernandez M., Del Puerto A., Dıaz-Hernandez J. I., Dıez-Zaera M.,Lucas J. J.,Garrido J. J. andMiras-PortugalM.T. (2008) InhibitionoftheATP-gated P2X7 receptor promotes axonal growth and branchingin cultured hippocampal neurons. J. Cell Sci. 121, 3717–3728.
Duan S., Anderson C. M., Keung E. C., Chen Y., Chen Y. and SwansonR. A. (2003) P2X7 receptor-mediated release of excitatory aminoacids from astrocytes. J. Neurosci. 23, 1320–1328.
Fields R. D. and Burnstock G. (2006) Purinergic signalling in neuron-glia interactions. Nature Rev. 7, 423–436.
Franke H., Gunther A., Grosche J., Schmidt R., Rossner S., ReinhardtR., Faber-Zuschratter H., Schneider D. and Illes P. (2004) P2X7
receptor expression after ischemia in the cerebral cortex of rats.J. Neuropathol. Exp. Neurol. 63, 686–699.
Fumagalli M., Brambilla R., D’Ambrosi N., Volonte C., Matteoli M.,Verderio C. and Abbracchio M. P. (2003) Nucleotide-mediatedcalcium signaling in rat cortical astrocytes: role of P2X and P2Yreceptors. Glia 43, 218–230.
Garcıa-Lecea M., Delicado E. G., Miras-Portugal M. T. and Castro E.(1999) P2X2 characteristics of the ATP receptor coupled to [Ca2+]iincreases in cultured Purkinje neurons from neonatal rat cere-bellum. Neuropharmacology 38, 699–706.
Gendron F. P., Neary J. T., Theiss P. M., Sun G. Y., Gonzalez F. A. andWeisman G. A. (2003) Mechanisms of P2X7 receptor-mediatedERK1/2 phosphorylation in human astrocytoma cells. Am. J.Physiol. Cell Physiol. 284, C571–C581.
Haydon P. G. (2001) GLIA: listening and talking to the synapse.Nat. Rev. 2, 185–193.
Hervas C., Perez-Sen R. and Miras-Portugal M. T. (2003) Coexpressionof functional P2X and P2Y nucleotide receptors in single cerebellargranule cells. J. Neurosci. Res. 73, 384–399.
Hervas C., Perez-Sen R. and Miras-Portugal M. T. (2005) Presence ofdiverse functional P2X receptors in rat cerebellar synaptic termi-nals. Biochem. Pharmacol. 70, 770–785.
Jiang L. H., Mackenzie A. B., North R. A. and Surprenant A. (2000)Brilliant blue G selectively blocks ATP-gated rat P2X7 receptors.Mol. Pharmacol. 58, 82–88.
Jimenez A. I., Castro E., Mirabet M., Franco R., Delicado E. G. andMiras-Portugal M. T. (1999) Potentiation of ATP calcium res-ponses by A2B receptor stimulation and other signals coupled to Gsproteins in type-1 cerebellar astrocytes. Glia 26, 119–128.
Jimenez A. I., Castro E., Communi D., Boeynaems J. M., Delicado E. G.and Miras-Portugal M. T. (2000) Coexpression of several types ofmetabotropic nucleotide receptors in single cerebellar astrocytes.J. Neurochem. 75, 2071–2079.
Kim Y. C., Lee J. S., Sak K., Marteau F., Mamedova L., BoeynaemsJ. M. and Jacobson K. A. (2005) Synthesis of pyridoxal phosphatederivatives with antagonist activity at the P2Y13 receptor. Biochem.Pharmacol. 70, 266–274.
von Kugelgen I. (2006) Pharmacological profiles of cloned mammalianP2Y-receptor subtypes. Pharmacol. Ther. 110, 415–432.
Matthias K., Kirchhoff F., Seifert G., Huttmann K., Matyash M.,Kettenmann H. and Steinhauser C. (2003) Segregated expressionof AMPA-type glutamate receptors and glutamate transportersdefines distinct astrocyte populations in the mouse hippocampus.J. Neurosci. 23, 1750–1758.
Miras-Portugal M. T., Dıaz-Hernandez M., Giraldez L., Hervas C.,Gomez-Villafuertes R., Sen R. P., Gualix J. and Pintor J. (2003)P2X7 receptors in rat brain: presence in synaptic terminals andgranule cells. Neurochem. Res. 28, 1597–1605.
North R. A. (2002) Molecular physiology of P2X receptors. Physiol.Rev. 82, 1013–1067.
Parekh A. B., Fleig A. and Penner R. (1997) The store-operated calciumcurrent I(CRAC): nonlinear activation by InsP3 and dissociationfrom calcium release. Cell 89, 973–980.
Parvathenani L. K., Tertyshnikova S., Greco C. R., Roberts S. B.,Robertson B. and Posmantur R. (2003) P2X7 mediates superoxideproduction in primary microglia and is up-regulated in a transgenicmouse model of Alzheimer’s disease. J. Biol. Chem. 278, 13309–13317.
Perea G. and Araque A. (2005) Properties of synaptically evokedastrocyte calcium signal reveal synaptic information processing byastrocytes. J. Neurosci. 25, 2192–2203.
Solini A., Chiozzi P., Morelli A., Fellin R. and Di Virgilio F. (1999)Human primary fibroblasts in vitro express a purinergic P2X7
receptor coupled to ion fluxes, microvesicle formation and IL-6release. J. Cell Sci. 112, 297–305.
Suadicani S. O., Brosnan C. F. and Scemes E. (2006) P2X7 receptorsmediate ATP release and amplification of astrocytic intercellularCa2+ signaling. J. Neurosci. 26, 1378–1385.
Surprenant A., Rassendren F., Kawashima E., North R. A. and Buell G.(1996) The cytolytic P2Z receptor for extracellular ATP identifiedas a P2X receptor (P2X7). Science 272, 735–738.
Takenouchi T., Ogihara K., Sato M. and Kitani H. (2005) Inhibitoryeffects of U73122 and U73343 on Ca2+ influx and pore formationinduced by the activation of P2X7 nucleotide receptors in mousemicroglial cell line. Biochim. Biophys. Acta 1726, 177–186.
Vigne P., Hechler B., Gachet C., Breittmayer J. P. and Frelin C. (1999)Benzoyl ATP is an antagonist of rat and human P2Y1 receptors andof platelet aggregation. Biochem. Biophys. Res. Commun. 256,94–97.
Virginio C., Church D., North R. A. and Surprenant A. (1997) Effects ofdivalent cations, protons and calmidazolium at the rat P2X7
receptor. Neuropharmacology 36, 1285–1294.Wang X., Arcuino G., Takano T. et al. (2004) P2X7 receptor inhibition
improves recovery after spinal cord injury. Nat. Med. 10, 821–827.
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 879–889
Calcium responses to BzATP in individual astrocytes | 889
Copyright © 2022 FDOKUMEN



![Nucleotide analogues containing 2-oxa-bicyclo[2.2.1]heptane and l-α-threofuranosyl ring systems: interactions with P2Y receptors](https://static.fdokumen.com/doc/165x107/6336ee671f95e36b5d086b6e/nucleotide-analogues-containing-2-oxa-bicyclo221heptane-and-l-threofuranosyl-1682844496.jpg)
















