
The Gi-coupled P2Y12 Receptor Regulates Diacylglycerol-mediated Signaling in Human Platelets

P1: GIU/HAK P2: HAK
Cellular and Molecular Neurobiology [cemn] PP353-365663 January 8, 2002 17:27 Style file version Oct 23, 2000
Cellular and Molecular Neurobiology, Vol. 21, No. 5, October 2001 ( C© 2001)
Regulation of Corticotropin-Releasing Hormone(CRH) Transcription and CRH mRNAStability by Glucocorticoids
Xin-Ming Ma,1,2 Cheri Camacho,1 and Greti Aguilera1,3
Received October 17, 2001; accepted October 22, 2001
SUMMARY
1. The increases in corticotropin-releasing hormone (CRH) mRNA following long-term adrenalectomy are associated with low levels of CRH gene transcription, suggestingthat glucocorticoids regulate CRH mRNA at the posttranscriptional level. In this studywe determined the time course of transcriptional activation after early adrenalectomy byintronic in situ hybridization, and evaluated the effects of glucocorticoids on CRH mRNAstability.
2. Plasma corticosterone was undetectable 3 h after adrenalectomy, but CRH hnRNAincreased only by 12 h, and remained elevated for the next 72 h. CRH mRNA increased18 h after adrenalectomy and reached a plateau lasting from 2 to 6 days, despite very lowCRH hnRNA levels.
3. Assessment of CRH mRNA stability, by incubation of slide-mounted hypothalamicsections in an intracellular-like medium at 37◦C, prior to measuring CRH mRNA levelsby in situ hybridization, revealed a half-life (t1/2) of 11.5 min in sham-operated rats, anda slower decrease adrenalectomized rats (t1/2-26.3 min). Corticosterone administration for3 days markedly decreased CRH mRNA t1/2 in both sham-operated and adrenalectomizedrats (6.5 and 5.0 min, respectively).
4. The data show that adrenalectomy causes transient increases in CRH mRNA tran-scription, followed by decreases in the rate of CRH mRNA degradation. This suggeststhat glucocorticoids regulate CRH mRNA at two sites, by inhibiting transcription and bydecreasing mRNA stability.
KEY WORDS: corticotropin-releasing hormone; hypothalamic paraventricular nucleus;transcription; mRNA stability; glucocorticoids; adrenalectomy.
INTRODUCTION
Corticotropin-releasing hormone (CRH) produced by parvocellular neurons of thehypothalamic paraventricular nucleus (PVN) is a major regulator of ACTH secretion
1 Section on Endocrine Physiology, Developmental Endocrinology Branch, National Institute of ChildHealth and Human Development, National Institutes of Health, Bethesda, Maryland.
2 Present address: Department of Neuroscience, University of Connecticut Health Center, Farmington,Connecticut 06030.
3 To whom correspondence should be addressed at Section on Endocrine Physiology, DevelopmentalEndocrinology Branch, National Institute of Child Health and Human Development, National Institutesof Health, Bldg 10 Rm 10N 262, 10 Center Drive, MSC 1862, Bethesda, Maryland 20892-1862.
465
0272-4340/01/1000-0465$19.50/0 C© 2001 Plenum Publishing Corporation

P1: GIU/HAK P2: HAK
Cellular and Molecular Neurobiology [cemn] PP353-365663 January 8, 2002 17:27 Style file version Oct 23, 2000
466 Ma, Camacho, and Aguilera
during stress (Aguilera, 1998; Vale et al., 1983). A number of immunohistochemi-cal and in situ hybridization studies have shown that basal levels of CRH expres-sion are under negative feedback inhibition by glucocorticoids. For example, CRHmRNA levels in the PVN decrease after glucocorticoid administration or implan-tation of dexamethasone in the PVN (Kovacs et al., 1986; Lightman and Young,1989; Sawchenko, 1997), but increase markedly following adrenalectomy, the lattereffect being prevented by glucocorticoid replacement (Beyer et al., 1988; Jingamiet al. 1985; Swanson and Simmonds, 1989; Young et al., 1986). The increases in CRHmRNA in the PVN after adrenalectomy are relatively slow, requiring 48 h to be-come detectable and reaching stable levels of about threefold the controls after 72 h(Brown and Sawchenko, 1997; Lightman and Young, 1989; Swanson and Simmonds,1989). Recent studies using in situ hybridization with exonic and intronic probes havedemonstrated that the increases in CRH mRNA following long-term adrenalectomyare associated with low levels of CRH transcription, not different from control val-ues, as shown by the very low levels of primary transcript (Imaki et al., 1995; Maet al., 1997). This suggests that glucocorticoid withdrawal must initially activate tran-scription but later CRH mRNA levels are mainly maintained by increasing mRNAstability. The use of intronic in situ hybridization (Herman et al., 1992) showed rapidincreases in CRH transcription during the first 30 min following blockade of adrenalcorticosterone production by metyrapone injection, but no information is availableon the time course of the changes in CRH gene transcription or CRH mRNA stabilityduring adrenalectomy.
Measurement of CRH mRNA stability using conventional methods would re-quire isolation of hypothalamic polyribosomes or blocking transcription in vivo(Kren et al., 1996; Nickenig and Murphy, 1996). Because of the difficulties in apply-ing these procedures to small brain nuclei which are little accessible to transcriptioninhibiting drugs, we used in situ hybridization techniques to evaluate the effect of glu-cocorticoids on CRH mRNA stability by adapting published procedures to measuremRNA half-life in polyribosomes in vitro (Nickenig and Murphy, 1996). In addition,we determined the time of transcriptional activation after adrenalectomy by mea-suring levels of CRH hn RNA during the first 3 days following adrenalectomy. Theresults suggest that glucocorticoids decrease CRH mRNA levels at transcriptionaland posttranscriptional sites.
MATERIALS AND METHODS
Animals and In Vivo Procedures
Adult male Sprague-Dawley rats weighing 250–300 g were housed three percage with a 14–10-h light–dark cycle with food and water available ad libitum, forat least 5 days prior to study. All procedures were performed according to the NIHguidelines, and experimental protocols were approved by the NICHD Animal Careand Use Committee.
Rats were adrenalectomized bilaterally via a dorsal approach or sham-operatedunder ketamine/xylazine anesthesia. For determination of the time course of changes

P1: GIU/HAK P2: HAK
Cellular and Molecular Neurobiology [cemn] PP353-365663 January 8, 2002 17:27 Style file version Oct 23, 2000
CRH mRNA Regulation by Glucocorticoids 467
in CRH transcription, groups of six adrenalectomized or sham-operated rats werekilled by decapitation at the times indicated in the figures, between 3 and 72 h aftersurgery.
To evaluate the effect of glucocorticoids on CRH mRNA stability, groups ofsham-operated and adrenalectomized rats were subjected to twice daily injections ofcorticosterone, 300 ¹g/100 g at 8:00 A.M. and 600 ¹g/100 g body weight at 6:00 P.M.,subcutaneously in peanut oil, or vehicle alone for 3 days starting the 3rd day aftersurgery, and killed in the morning on the 7th day. Previous studies have shown that thisdose of corticosterone injected in the evening increases circulating corticosterone tolevels in the stress range, followed by a progressive decline for the first 6 h andremaining near basal levels for up to 10 h (Ochedalski et al., 1998).
Rats were killed by decapitation between 0900 and 1200 h. Trunk blood wascollected into ice-cold tubes containing EDTA for corticosterone determination byradioimmunoassay using 50 ¹L of plasma and the rat corticosterone kit (DPC, LosAngeles, CA). The sensitivity of the assay was 7.5 ng/mL. Brains were removed,frozen on dry ice, and stored at−80◦C. Twelve-micrometer sections were cut thoughthe medial parvocellular subdivision of the PVN in a cryostat at−20◦C, thaw mountedon poly-L-lysine (Sigma, St. Louis, MO) coated slides, and stored at−80◦C until usedfor in situ hybridization.
CRH mRNA Stability
Six sets of consecutive coronal hypothalamic sections through the medial parvo-cellular PVN were obtained from each rat in a cryostat at−20◦C and stored at−70◦Cand used to evaluate mRNA stability by modification of the procedure described tomeasure mRNA half-life in polyribosomes (Nickenig and Murphy, 1996). At the timeof the experiment, sections were thawed in a dessicator under vacuum and incubatedat 37◦C in an intracellular-like medium containing 10 mM Tris-HCl (pH 7.5), 100 mMpotassium acetate, 10 mM magnesium acetate, 2 mM DTT, RNasin 3 units/mL, andyeast tRNA 4 mg/mL, for 0, 15, 30, 60, and 120 min. Slides for the different timepoints were added sequentially, and at 120-min incubation of all slides was terminatedsimultaneously by fixation in 4% formaldehyde for 5 min to stop mRNA degradation.After fixation slides were washed three times in PBS and immediately processed forevaluation of CRH mRNA levels by in situ hybridization (Luo et al., 1994).
In Situ Hybridization
The rat CRH intron (CRHin) probe (kindly supplied by Dr Robert Thompson,University of Michigan, Ann Arbor, MI), was a 530 bp pvuII fragment of the CRHgene subcloned into pGEM-3 (Promega, Madison, WI) and linearized with XbaI.The rat CRH (CRHex 2) cDNA (Dr Robert Thompson) was a 770 bp BamHIfragment subcloned in pGEM-3Z (Promega), linearized by HindIII. High specificactivity antisense cRNA probes for CRHin and CRHex were produced using35S-ATP and 35S-UTP, as previously described (Luo et al., 1994). In situ hybridiza-tion was performed as previously described (Ma and Aguilera, 1999a). Briefly, priorto hybridization, stored sections were air-dried at room temperature, fixed with 4%

P1: GIU/HAK P2: HAK
Cellular and Molecular Neurobiology [cemn] PP353-365663 January 8, 2002 17:27 Style file version Oct 23, 2000
468 Ma, Camacho, and Aguilera
formaldehyde for 5 min at room temperature, washed three times with PBS, and thenacetylated in 0.25% acetic anhydride in 0.1 M triethanolamine/0.9% NaCl (pH 8.0)for 10 min at room temperature. Slides used for mRNA stability evaluation followedthe same prehybridization and hybridization procedures after fixation. Sections weredehydrated in serial ethanol solutions, defated in chloroform, and dried before hy-bridization with the radiolabeled probes. Sections were hybridized overnight at 55◦Cwith 2× 106 cpm labeled CRHin or CRHex probe per slide containing four sectionsrespectively. Nonspecifically hybridized probe was removed by washing with 50%formamide/250 mM NaCl at 60oC for 10–15 min, ribonuclease A treatment for 30min at 37◦C, followed by three washes with 0.1 × SSC.
For quantification of CRH hnRNA and CRH mRNA in the PVN, sections wereexposed to Kodak BIOMAX film (Kodak, Rochester, NY) together with 14C-labeledstandards (American Radiochemical, St. Louis, MO), for 10 h (CRHex) or for 15days (CRHin). For cellular localization of CRH mRNA or CRH hnRNA, slides weresubsequently dipped in nuclear emulsion diluted 1:1 in distilled water (NTB2, Kodak,Rochester, NY), exposed for appropriate times (CRHin, 40 days; CRHex, 4 days),and counterstained with cresyl violet acetate (Sigma, St. Louis, MO). The opticaldensity of film autoradiographic images of parvocellular CRH mRNA and CRHhnRNA was measured in a computerized image analysis system (Imaging Research,St. Catherines, Ontario, Canada), using the public domain NIH Image program (de-veloped at the National Institutes of Health, and available at http://rsb.info.nih.gov/nih-image). Optical densities obtained in two consecutive sections per rat were aver-aged and used to calculate group means. The results are presented as mean and SEof the percentage of change from basal levels in sham-operated rats.
Statistical analysis was performed by one-way analysis of variance (ANOVA)followed by Fisher’s least significant difference procedure (PLSD) test to assessstatistical significance between control and experimental groups at each time point.p < 0.05 was considered to be statistically significant.
RESULTS
Plasma Corticosterone Levels
In sham-operated rats, plasma corticosterone was markedly increased 3 h aftersurgery and declined to remain at levels near basal after 12 h, while in adrenalec-tomized rats plasma corticosterone had decreased to almost undetectable levels by3 h (Fig. 1(A)). Plasma corticosterone levels were below the detection limit of theassay in 6-day adrenalectomized rats receiving oil injection, while levels were 31.3±11.7 ng/mL in rats receiving corticosterone injections, 18 h after the last injection.
CRH mRNA and CRH hnRNA Levels
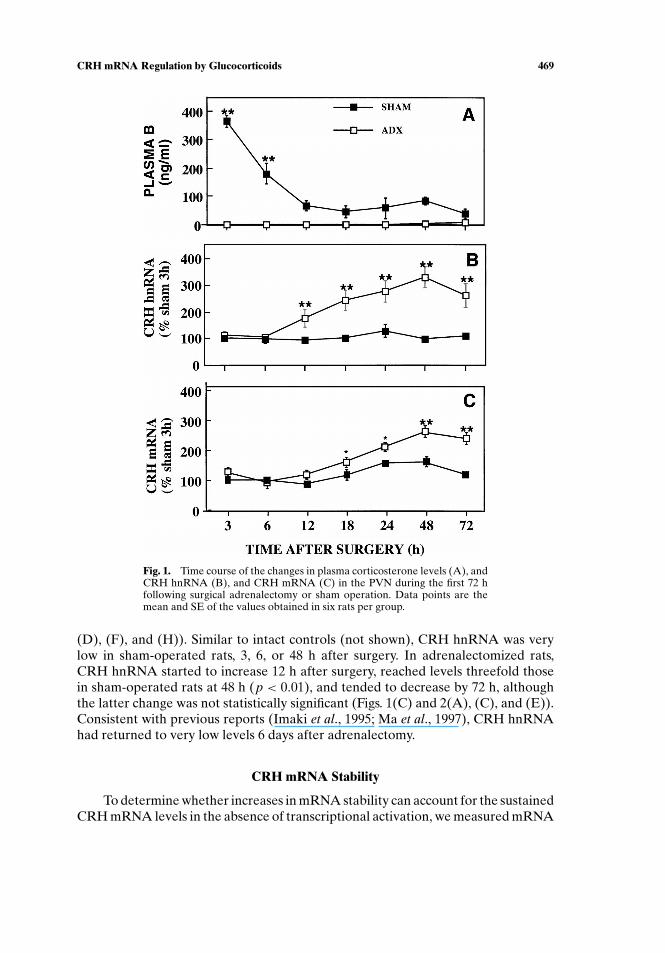
CRH mRNA levels increased significantly over the values in sham-operatedcontrols between 18 (1.7-fold, p < 0.05) and 48 h after adrenalectomy, to remain atthese levels (2.5-fold, p < 0.01) at longer times after surgery (Figs. 1(B) and 2(B),
P1: GIU/HAK P2: HAK
Cellular and Molecular Neurobiology [cemn] PP353-365663 January 8, 2002 17:27 Style file version Oct 23, 2000
CRH mRNA Regulation by Glucocorticoids 469
Fig. 1. Time course of the changes in plasma corticosterone levels (A), andCRH hnRNA (B), and CRH mRNA (C) in the PVN during the first 72 hfollowing surgical adrenalectomy or sham operation. Data points are themean and SE of the values obtained in six rats per group.
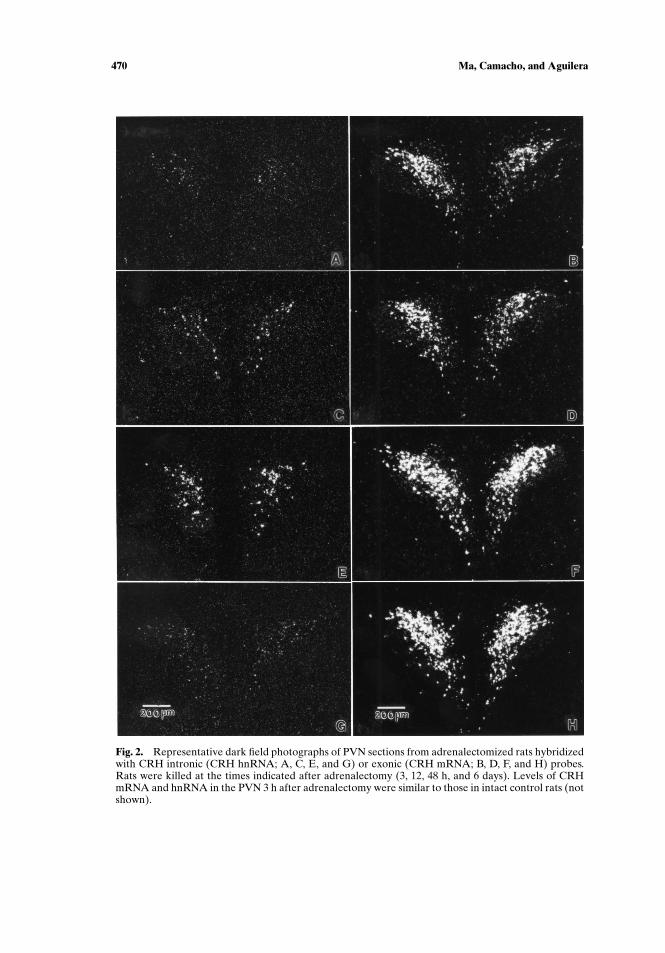
(D), (F), and (H)). Similar to intact controls (not shown), CRH hnRNA was verylow in sham-operated rats, 3, 6, or 48 h after surgery. In adrenalectomized rats,CRH hnRNA started to increase 12 h after surgery, reached levels threefold thosein sham-operated rats at 48 h (p < 0.01), and tended to decrease by 72 h, althoughthe latter change was not statistically significant (Figs. 1(C) and 2(A), (C), and (E)).Consistent with previous reports (Imaki et al., 1995; Ma et al., 1997), CRH hnRNAhad returned to very low levels 6 days after adrenalectomy.
CRH mRNA Stability
To determine whether increases in mRNA stability can account for the sustainedCRH mRNA levels in the absence of transcriptional activation, we measured mRNA
P1: GIU/HAK P2: HAK
Cellular and Molecular Neurobiology [cemn] PP353-365663 January 8, 2002 17:27 Style file version Oct 23, 2000
470 Ma, Camacho, and Aguilera
Fig. 2. Representative dark field photographs of PVN sections from adrenalectomized rats hybridizedwith CRH intronic (CRH hnRNA; A, C, E, and G) or exonic (CRH mRNA; B, D, F, and H) probes.Rats were killed at the times indicated after adrenalectomy (3, 12, 48 h, and 6 days). Levels of CRHmRNA and hnRNA in the PVN 3 h after adrenalectomy were similar to those in intact control rats (notshown).
P1: GIU/HAK P2: HAK
Cellular and Molecular Neurobiology [cemn] PP353-365663 January 8, 2002 17:27 Style file version Oct 23, 2000
CRH mRNA Regulation by Glucocorticoids 471
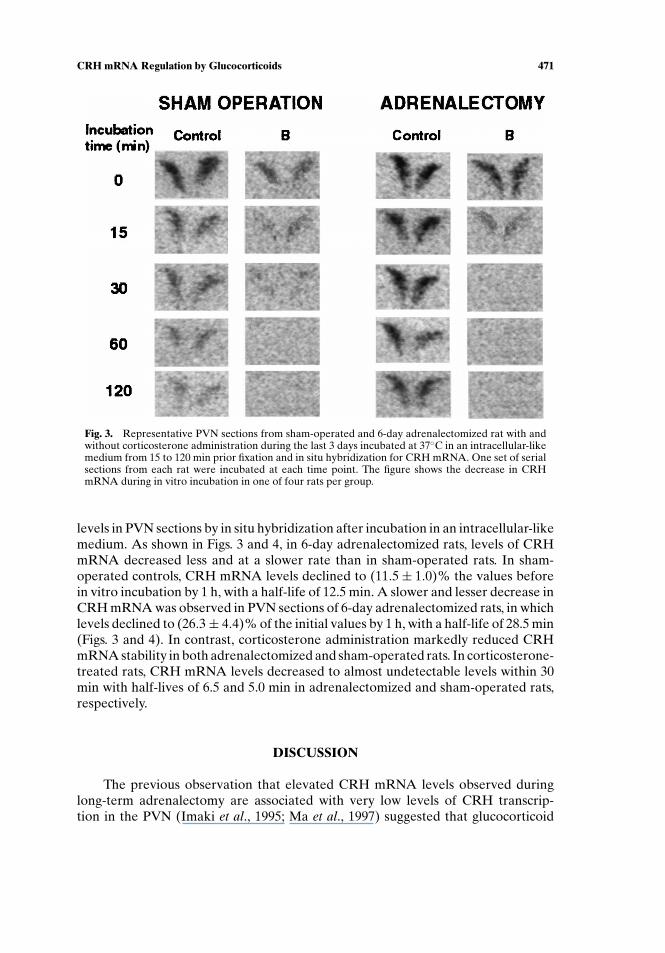
Fig. 3. Representative PVN sections from sham-operated and 6-day adrenalectomized rat with andwithout corticosterone administration during the last 3 days incubated at 37◦C in an intracellular-likemedium from 15 to 120 min prior fixation and in situ hybridization for CRH mRNA. One set of serialsections from each rat were incubated at each time point. The figure shows the decrease in CRHmRNA during in vitro incubation in one of four rats per group.
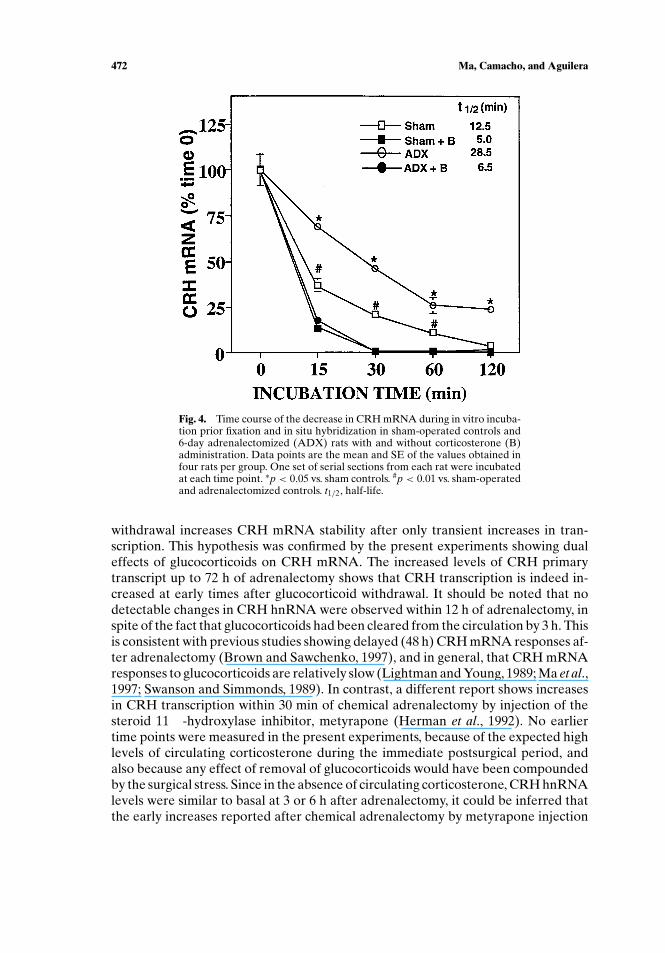
levels in PVN sections by in situ hybridization after incubation in an intracellular-likemedium. As shown in Figs. 3 and 4, in 6-day adrenalectomized rats, levels of CRHmRNA decreased less and at a slower rate than in sham-operated rats. In sham-operated controls, CRH mRNA levels declined to (11.5± 1.0)% the values beforein vitro incubation by 1 h, with a half-life of 12.5 min. A slower and lesser decrease inCRH mRNA was observed in PVN sections of 6-day adrenalectomized rats, in whichlevels declined to (26.3± 4.4)% of the initial values by 1 h, with a half-life of 28.5 min(Figs. 3 and 4). In contrast, corticosterone administration markedly reduced CRHmRNA stability in both adrenalectomized and sham-operated rats. In corticosterone-treated rats, CRH mRNA levels decreased to almost undetectable levels within 30min with half-lives of 6.5 and 5.0 min in adrenalectomized and sham-operated rats,respectively.
DISCUSSION
The previous observation that elevated CRH mRNA levels observed duringlong-term adrenalectomy are associated with very low levels of CRH transcrip-tion in the PVN (Imaki et al., 1995; Ma et al., 1997) suggested that glucocorticoid
P1: GIU/HAK P2: HAK
Cellular and Molecular Neurobiology [cemn] PP353-365663 January 8, 2002 17:27 Style file version Oct 23, 2000
472 Ma, Camacho, and Aguilera
Fig. 4. Time course of the decrease in CRH mRNA during in vitro incuba-tion prior fixation and in situ hybridization in sham-operated controls and6-day adrenalectomized (ADX) rats with and without corticosterone (B)administration. Data points are the mean and SE of the values obtained infour rats per group. One set of serial sections from each rat were incubatedat each time point. ∗p < 0.05 vs. sham controls. #p < 0.01 vs. sham-operatedand adrenalectomized controls. t1/2, half-life.
withdrawal increases CRH mRNA stability after only transient increases in tran-scription. This hypothesis was confirmed by the present experiments showing dualeffects of glucocorticoids on CRH mRNA. The increased levels of CRH primarytranscript up to 72 h of adrenalectomy shows that CRH transcription is indeed in-creased at early times after glucocorticoid withdrawal. It should be noted that nodetectable changes in CRH hnRNA were observed within 12 h of adrenalectomy, inspite of the fact that glucocorticoids had been cleared from the circulation by 3 h. Thisis consistent with previous studies showing delayed (48 h) CRH mRNA responses af-ter adrenalectomy (Brown and Sawchenko, 1997), and in general, that CRH mRNAresponses to glucocorticoids are relatively slow (Lightman and Young, 1989; Ma et al.,1997; Swanson and Simmonds, 1989). In contrast, a different report shows increasesin CRH transcription within 30 min of chemical adrenalectomy by injection of thesteroid 11 ¯-hydroxylase inhibitor, metyrapone (Herman et al., 1992). No earliertime points were measured in the present experiments, because of the expected highlevels of circulating corticosterone during the immediate postsurgical period, andalso because any effect of removal of glucocorticoids would have been compoundedby the surgical stress. Since in the absence of circulating corticosterone, CRH hnRNAlevels were similar to basal at 3 or 6 h after adrenalectomy, it could be inferred thatthe early increases reported after chemical adrenalectomy by metyrapone injection

P1: GIU/HAK P2: HAK
Cellular and Molecular Neurobiology [cemn] PP353-365663 January 8, 2002 17:27 Style file version Oct 23, 2000
CRH mRNA Regulation by Glucocorticoids 473
reflect biphasic changes in CRH transcription (Herman et al., 1992). However, it isalso possible that the stress caused by the metyrapone injection is sufficient to in-crease CRH transcription under conditions in which circulating glucocorticoids startto decrease because of the blockade of the steroidogenic pathway. In this regard, re-cent studies in our laboratory show rapid and marked increases in CRH hnRNA inresponse of a minor stress in 48-h adrenalectomized rats while no effect is observedin intact rats (Ma and Aguilera, 1999b). The mechanisms by which transcriptiondecreases after long-term adrenalectomy in spite of the continuous absence of glu-cocorticoid feedback are not clear but suggest the presence of intracellular feedbackmechanisms by which transcription is inhibited to prevent uncontrolled increases inmRNA levels.
The use of in situ hybridization techniques after in vitro incubation of hy-pothalamic sections in these experiments made it possible to identify changes inthe rate of CRH mRNA degradation in the PVN during adrenalectomy and glu-cocorticoid administration. Although the rate of mRNA degradation measured inthese experiments is unlikely to represent the half-life of CRH mRNA in vivo, itis clear from these experiments that glucocorticoids affect CRH mRNA stability.Thus, while removal of glucocorticoids by adrenalectomy increases CRH mRNAhalf-life, administration of corticosterone markedly reduces mRNA stability, sug-gesting that glucocorticoids facilitate CRH mRNA degradation. This marked ef-fect of glucocorticoids on CRH mRNA stability was observed with relatively lowdoses of corticosterone known to transiently elevate circulating levels (Ochedalskiet al., 1998). This suggests that mRNA stability is a major site of regulation of CRHmRNA levels by glucocorticoids, and it may explain the slow effect of glucocorti-coids on CRH mRNA levels in the PVN. A number of studies in other systems haveshown that glucocorticoids can affect mRNA stability. Similar to the present effectson CRH mRNA, glucocorticoids have been shown to decrease the stability of CRHreceptor-1 mRNA in AtT-20 cells (Iredale and Duman, 1997), IL-4 mRNA in lympho-cytes (Mozo et al., 1998), procollagen mRNA in osteosarcoma cells (Mahonen et al.,1998) and monocyte chemotactic protein mRNA in the lung (Cooper et al., 1998). Inother systems such as mRNA for glutamine synthetase and fattyacid synthetase inthe lung (Xu and Rooney, 1997), glucocorticoids increase mRNA stability for theseproteins.
Several factors, such as the length of the poly(A) tail and changes in the levels ofproteins interacting with the RNA or RNA-degrading enzymes, can influence the sen-sitivity of mRNAs to nucleases and affect mRNA stability (Sacks, 1993). Concerninghypothalamic neuropeptides, increases in vasopressin mRNA and oxytocin transla-tion during osmotic stimulation or circadian rhythm (Carranza et al., 1988; Carterand Murphy, 1989; Robinson et al., 1988) are associated with increase in length inthe poly(A) tail.
In summary, this study shows that long-term adrenalectomy transiently increasesCRH transcription while decreasing the rate of mRNA degradation. This suggeststhat glucocorticoids have dual regulatory actions on CRH mRNA levels by inhibit-ing transcription and decreasing mRNA stability, and that the latter mechanism ismainly responsible for the maintenance of high CRH mRNA levels during long-termadrenalectomy.

P1: GIU/HAK P2: HAK
Cellular and Molecular Neurobiology [cemn] PP353-365663 January 8, 2002 17:27 Style file version Oct 23, 2000
474 Ma, Camacho, and Aguilera
REFERENCES
Aguilera, G. (1998). Corticotropin releasing hormone, receptor regulation and the stress response. TrendsEndocrinol. Metab. 9:329–336.
Beyer, H. S., Matta, S. G., and Sharp, B. M. (1988). Regulation of the messenger ribonucleic acid forcorticotropin releasing factor in the paraventricular nucleus and other brain regions. Endocrinology123:2117–2123.
Brown, E. R., and Sawchenko, P. E. (1997). Hypophysiotropic CRF neurons display a sustained immediate-early gene response to chronic stress but not to adrenalectomy. J. Neuroendocrinol. 9:307–316.
Carranza, E. J., Pasieka, K. B., and Majzoub, J. A. (1988). The vasopressin mRNA poly(A) tract is unusuallylong and increases during stimulation of vaopressin gene expression in vivo. Mol. Cell. Biol. 8:2267–2274.
Carter, D. A., and Murphy, D. (1989). Independent regulation of neuropeptide mRNA level and poly(A)tail length. J. Biol. Chem. 264:6601–6603.
Cooper, J. A., Fuller, J. M., McMinn, K. M., and Culbreth, R. R. (1998). Modulation of monocyte chemo-tactic protein-1 production by hyperoxia: Importance of mRNA stability in control of cytokine pro-duction. Am. Respir. Cell. Mol. Biol. 18:521–525.
Herman, J. P., Schafer, M. K., Thompson, R. C., and Watson, S. J. (1992). Rapid regulation of corticotropin-releasing hormone gene transcription in vivo. Mol. Endocrinol. 6:1061–1069.
Imaki, T., Wang, X.-Q., Shibasaki, T., Yamada, K., Harada, S., Chikada, N., Naruse, M., and Demura,H. (1995). Stress induced activation of neuronal activity and corticotropin-releasing factor geneexpression in the paraventricular nucleus is modulated by glucocorticoid in rats. J. Clin. Invest. 96:231–328.
Iredale, P. A., and Duman, R. S. (1997). Glucocorticoid regulation of corticotropin releasing factor-1receptor expression in pituitary-derived AtT-20 cells. Mol. Pharmacol. 51:794–799.
Jingami, H., Matsukura, S., Numa, S., and Imura H. (1985). Effects of adrenalectomy and dexametha-sone administration on the levels of prepro-corticotropin releasing factor messenger ribonucleic acid(mRNA) in the hypothalamus and adrenocorticotropin/B-lipotropin processor mRNA in the pitu-itary in rats. Endocrinology 117:1314–1320.
Kovacs, K., Kiss, J. Z., and Makara, G. B. (1986). Glucocorticoid implants around the hypothalamicparaventricular nucleus prevent the increase of corticotropin releasing factor and arginine vasopressinimmunostaining induced by adrenalectomy. Neuroendocrinology 44:229–234.
Kren, B. T., Trembley, J. H., and Steer, C. J. (1996). Alterations in mRNA stability during rat liver regen-eration. Am. J. Physiol. 270:G763–G777.
Lightman, S. L., and Young, W. S. (1989). Influence of steroids on the hypothalamic corticotropin releasingfactor and proenkephalin mRNA responses to stress. Proc. Natl. Acad. Sci. USA 86:4306–4310.
Luo, X., Kiss, A., Makara, G. B., Lolait, S. J., and Aguilera, G. (1994). Stress-specific regulation of corti-cotropin releasing hormone receptor expression in the paraventricular and supraoptic nuclei of thehypothalamus in the rat. J. Neuroendocrinol. 6:689–696.
Ma, X.-M., and Aguilera G. (1999a). Transcriptional responses of the vasopressin and corticotropin re-leasing hormone genes to acute and repeated intraperitoneal hypertonic saline injection. Mol. BrainRes. 68:129–140.
Ma, X.-M., and Aguilera, G. (1999b). Differential regulation of corticotropin releasing hormone andvasopressin transcription by glucocorticoids. Endocrinology 140:5642–5650.
Ma, X.-M., Levy, A., and Lightman, S. L. (1997). Rapid changes of heteronuclear RNA for argininevasopressin but not for corticotropin releasing hormone in response to acute corticosterone admin-istration. J. Neuroendocrinol. 9:723–728.
Mahonen, A., Jukkola, A., Risteli, L., Risteli, J., and Maenpaa, P. H. (1998). Type I procollagen synthesis isregulated by steroids and related hormones in human osteosarcoma cells. J. Cell. Biochem. 68:151–163.
Mozo, L., Gayo, A., Suarez, A., Rivas, D., Zamorano, J., and Gutierrez, C. (1998). Glucocorticoids inhibitIl-4 and mitogen-induced IL-4R alpha chain by different post-transcriptional mechanisms. J. AllergyClin. Immunol. 102:968–976.
Nickenig, G., and Murphy, T. J. (1996). Enhanced angiotensin receptor type-1 mRNA degradation andinduction of polyribosomal mRNA binding proteins by angiotensin II in vascular smooth musclecells. Mol. Pharmacol. 50:743–751.
Ochedalski, T., Rabadan-Diehl, C., and Aguilera, G. (1998). Interaction between glucocorticoids andcorticotropin releasing hormone in the regulation of pituitary CRH receptor in vivo in the rat.J. Neuroendocrinol. 10:363–369.
Robinson, B. G., Frim, D. M., Schwartz, W. J., and Majzoub, J. A. (1988). Vasopressin mRNA insuprachiasmatic nuclei: Daily regulation of polyadenylate tail length. Science 241:342–344.

P1: GIU/HAK P2: HAK
Cellular and Molecular Neurobiology [cemn] PP353-365663 January 8, 2002 17:27 Style file version Oct 23, 2000
CRH mRNA Regulation by Glucocorticoids 475
Sacks, A. B. (1993). Messenger RNA degradation in eukaryotes. Cell 74:413–421.Sawchenko, P. E. (1997). Evidence for a local site of action for glucocorticoids in inhibiting CRF and
vasopressin expression in the paraventricular nucleus. Brain Res. 403:213–224.Swanson, L. W., and Simmonds, D. M. (1989). Differential steroid hormone and neural influence on
peptide mRNA levels in CRH cells of the paraventricular nucleus: A hybridization histochemistrystudy in the rat. J. Comp. Neurol. 285:413–435.
Vale, W. W., Rivier, C., Brown, M. R., Spiess, J., Koob. G., Swanson, L., Bilezikjian, L., Bloom, F., andRivier, J. (1983). Chemical and biological characterization of corticotropin releasing factor. Rec.Prog. Horm. Res. 39:245–270.
Xu, Z. X., and Rooney, S. A. (1997). Glucocorticoids increase fatty acid synthetase mRNA stability infetal rat lung. Am. J. Physiol. 272:L860–L864.
Young, W. S., Mezey, E., and Siegel, R. E. (1986). Quantitative in situ hybridization histochemistry revealsincreased levels of corticotropin releasing factor mRNA after adrenalectomy in rats. Neurosci. Lett.70:198–203.
Copyright © 2022 FDOKUMEN




















