
Activation of Protein Kinase C by Oxytocin Inhibits the Biological Activity of the Human Myometrial...
10
Activation of Protein Kinase C by Oxytocin Inhibits the Biological Activity of the Human Myometrial Corticotropin-Releasing Hormone Receptor at Term* DIMITRIS K. GRAMMATOPOULOS AND EDWARD W. HILLHOUSE The Sir Quinton Hazell Molecular Medicine Research Center, Department of Biological Sciences, University of Warwick, Coventry, United Kingdom CV4 7AL ABSTRACT The role of placental CRH in human pregnancy is currently un- known. The myometrium expresses CRH receptors that during preg- nancy become coupled to adenylate cyclase. Oxytocin (OT) is one of the main regulators of uterine activity, acting via activation of the inositol triphosphate pathway. In view of the possible cross-talk between the CRH and OT signal transduction pathways we have sought to ex- amine in more detail the second messenger mechanisms involved. CRH receptor binding affinity for CRH and activation of adenylate cyclase were reduced in the presence of OT in pregnant (at term, but not preterm) human myometrium. OT action was mediated via per- tussis toxin-sensitive G proteins, which directly inhibit adenylate cyclase and, via activation of protein kinase C, phosphorylate the CRH receptor, leading to desensitization. Activation of protein kinase C by OT could be partially inhibited in human pregnant myometrial cells by OT antagonists (F327 and CAP476; 1 mM) or phospholipase C inhibitors (U73122; 10 mM). These results suggest that in term myometrium, CRH receptor function is modulated by OT, leading to reduced biological activity, lower cAMP levels, and a subsequent shift in favor of contractility rather than relaxation. (Endocrinology 140: 585–594, 1999) I N SEARCH of potential mediators of human labor, interest has been directed toward placentally derived CRH. The biological function of this placental CRH during human pregnancy is still unknown, but we and others have devel- oped the hypothesis that CRH might influence myometrial contractility, and hence parturition. Specific CRH receptors exist within the human myometrium that increase their af- finity during pregnancy and become functionally linked to the adenylate cyclase system in the pregnant state only (1, 2). However, as term approaches, CRH appears to have a re- duced stimulatory action on myometrial adenylate cyclase (3), and one explanation is a decreased expression and down- regulation of G s a toward term and labor (4). As the uterus becomes prepared for labor, it is likely that uterine activation occurs in response to paracrine/autocrine action of a variety of uterotonins. Oxytocin (OT) is the most potent known endogenous uterotonic agent (5), and the ma- jor factor influencing its bioactivity is an increase in the number of myometrial OT receptors at term (6). Local OT production by uterine tissues is also increased (7). In mice, however, OT may not play a critical role, because the in- duction of labor and parturition can proceed normally in mice in which the OT gene has been deleted (8). In the human myometrium, the effects of OT are mediated through a sev- en-transmembrane domain receptor (9) linked to both G i and G q proteins (10), which activate phospholipase C (PLC) with subsequent production of inositol 1,4,5-triphosphate. This has two important consequences: 1) the rapid release of cal- cium ions from intracellular stores in the sarcoplasmic re- ticulum, and 2) the accumulation of diacylglycerol (DAG), which remains membrane bound. Elevated levels of intra- cellular calcium activate some types of cytosolic protein ki- nase C (PKC) and induce its translocation to the cellular membrane, where it is further activated by DAG (11). PKC is a serine/threonine kinase and acts as a key mediator in signal transduction events (12). The family of PKC has sev- eral members, which have been divided into a Ca 21 -depen- dent or conventional PKC group and a Ca 21 -independent or novel PKC group (12). One of the most important functions of PKC in signal transduction is that it mediates cross-talk between different signaling pathways. In the human myometrium one of the most important signaling pathways is the adenylate cyclase-cAMP system (13), which can be activated by the CRH receptor (2). Mo- lecular characterization has revealed that this receptor be- longs to the calcitonin/vasoactive intestinal polypeptide/ GRF/PTH subfamily of G protein-coupled receptors. There are two groups of CRH receptors (CRH-R1 and CRH-R2) arising from separate genes (14, 15), both of which have several variants arising from alternative splicing. The R1 gene encodes four known subtypes: R1a, R1b (with a 29- amino acid insertion in the first intracellular loop) (14), R1c (with a 40-amino acid exon deletion from the N-terminal domain) (16), and R1d (with a 12-amino acid exon deletion from the seventh transmembrane domain; our unpublished observations). The CRH-R2 also has three spliced variants, R2a, R2b, and R2g, each the product of differential 59-exon splicing and each with distinct tissue distributions (17–19). All of these subtypes of the CRH receptor, with the exception Received July 22, 1998. Address all correspondence and requests for reprints to: Dr. D. Gram- matopoulos, The Sir Quinton Hazell Molecular Medicine Research Cen- ter, Department of Biological Sciences, University of Warwick, Gibbet Hill Road, Coventry, United Kingdom CV4 7AL. E-mail: chdg@dna. bio.warwick.ac.uk. * This work was supported by a grant from the Wellcome Trust. Holds the WPH Charitable Trust Chair of Medicine. 0013-7227/99/$03.00/0 Vol. 140, No. 2 Endocrinology Printed in U.S.A. Copyright © 1999 by The Endocrine Society 585
Transcript of Activation of Protein Kinase C by Oxytocin Inhibits the Biological Activity of the Human Myometrial...

Activation of Protein Kinase C by Oxytocin Inhibits theBiological Activity of the Human MyometrialCorticotropin-Releasing Hormone Receptor at Term*
DIMITRIS K. GRAMMATOPOULOS AND EDWARD W. HILLHOUSE†
The Sir Quinton Hazell Molecular Medicine Research Center, Department of Biological Sciences,University of Warwick, Coventry, United Kingdom CV4 7AL
ABSTRACTThe role of placental CRH in human pregnancy is currently un-
known. The myometrium expresses CRH receptors that during preg-nancy become coupled to adenylate cyclase. Oxytocin (OT) is one of themain regulators of uterine activity, acting via activation of the inositoltriphosphate pathway. In view of the possible cross-talk between theCRH and OT signal transduction pathways we have sought to ex-amine in more detail the second messenger mechanisms involved.
CRH receptor binding affinity for CRH and activation of adenylatecyclase were reduced in the presence of OT in pregnant (at term, butnot preterm) human myometrium. OT action was mediated via per-
tussis toxin-sensitive G proteins, which directly inhibit adenylatecyclase and, via activation of protein kinase C, phosphorylate theCRH receptor, leading to desensitization. Activation of protein kinaseC by OT could be partially inhibited in human pregnant myometrialcells by OT antagonists (F327 and CAP476; 1 mM) or phospholipaseC inhibitors (U73122; 10 mM).
These results suggest that in term myometrium, CRH receptorfunction is modulated by OT, leading to reduced biological activity,lower cAMP levels, and a subsequent shift in favor of contractilityrather than relaxation. (Endocrinology 140: 585–594, 1999)
IN SEARCH of potential mediators of human labor, interesthas been directed toward placentally derived CRH. The
biological function of this placental CRH during humanpregnancy is still unknown, but we and others have devel-oped the hypothesis that CRH might influence myometrialcontractility, and hence parturition. Specific CRH receptorsexist within the human myometrium that increase their af-finity during pregnancy and become functionally linked tothe adenylate cyclase system in the pregnant state only (1, 2).However, as term approaches, CRH appears to have a re-duced stimulatory action on myometrial adenylate cyclase(3), and one explanation is a decreased expression and down-regulation of Gsa toward term and labor (4).
As the uterus becomes prepared for labor, it is likely thatuterine activation occurs in response to paracrine/autocrineaction of a variety of uterotonins. Oxytocin (OT) is the mostpotent known endogenous uterotonic agent (5), and the ma-jor factor influencing its bioactivity is an increase in thenumber of myometrial OT receptors at term (6). Local OTproduction by uterine tissues is also increased (7). In mice,however, OT may not play a critical role, because the in-duction of labor and parturition can proceed normally inmice in which the OT gene has been deleted (8). In the humanmyometrium, the effects of OT are mediated through a sev-en-transmembrane domain receptor (9) linked to both Gi andGq proteins (10), which activate phospholipase C (PLC) with
subsequent production of inositol 1,4,5-triphosphate. Thishas two important consequences: 1) the rapid release of cal-cium ions from intracellular stores in the sarcoplasmic re-ticulum, and 2) the accumulation of diacylglycerol (DAG),which remains membrane bound. Elevated levels of intra-cellular calcium activate some types of cytosolic protein ki-nase C (PKC) and induce its translocation to the cellularmembrane, where it is further activated by DAG (11). PKCis a serine/threonine kinase and acts as a key mediator insignal transduction events (12). The family of PKC has sev-eral members, which have been divided into a Ca21-depen-dent or conventional PKC group and a Ca21-independent ornovel PKC group (12). One of the most important functionsof PKC in signal transduction is that it mediates cross-talkbetween different signaling pathways.
In the human myometrium one of the most importantsignaling pathways is the adenylate cyclase-cAMP system(13), which can be activated by the CRH receptor (2). Mo-lecular characterization has revealed that this receptor be-longs to the calcitonin/vasoactive intestinal polypeptide/GRF/PTH subfamily of G protein-coupled receptors. Thereare two groups of CRH receptors (CRH-R1 and CRH-R2)arising from separate genes (14, 15), both of which haveseveral variants arising from alternative splicing. The R1gene encodes four known subtypes: R1a, R1b (with a 29-amino acid insertion in the first intracellular loop) (14), R1c(with a 40-amino acid exon deletion from the N-terminaldomain) (16), and R1d (with a 12-amino acid exon deletionfrom the seventh transmembrane domain; our unpublishedobservations). The CRH-R2 also has three spliced variants,R2a, R2b, and R2g, each the product of differential 59-exonsplicing and each with distinct tissue distributions (17–19).All of these subtypes of the CRH receptor, with the exception
Received July 22, 1998.Address all correspondence and requests for reprints to: Dr. D. Gram-
matopoulos, The Sir Quinton Hazell Molecular Medicine Research Cen-ter, Department of Biological Sciences, University of Warwick, GibbetHill Road, Coventry, United Kingdom CV4 7AL. E-mail: [email protected].
* This work was supported by a grant from the Wellcome Trust.† Holds the WPH Charitable Trust Chair of Medicine.
0013-7227/99/$03.00/0 Vol. 140, No. 2Endocrinology Printed in U.S.A.Copyright © 1999 by The Endocrine Society
585

of R2g, exist in the human pregnant myometrium and havebeen identified using PCR, sequencing of the amplified DNAfragments, and fluorescent in situ hybridization (20). Inter-estingly, the complementary DNA sequences of the CRH-R1and -R2 receptors encode four potential PKC phosphoryla-tion sites, identical in both CRH receptors, which are locatedin the first and second intracellular loops as well as in theC-terminus.
These PKC phosphorylation sites may be involved in themodulation of CRH receptor signal transduction. Their pre-cise role, however, has not yet been characterized. In view ofthis, we searched for potential mechanisms of cross-talk be-tween OT and the CRH system that might be involved in theregulation of human myometrial contractility during humanpregnancy.
Materials and MethodsExperimental subjects
Pregnant myometrial tissue was obtained from women undergoingelective caesarean section at term (.38 weeks gestation; n 5 12) orpreterm (,34 weeks gestation; n 5 9) before the onset of labor. Allcesarean sections were performed for reasons of breech presentation orfetal distress. The biopsy was standardized to the upper margin of thelower segment of the uterus in the midline. This provides the closestapproximation of the upper segment of the uterus. The tissue wasimmediately snap-frozen in liquid nitrogen and stored at 270 C untiluse.
For myocyte isolation and culture, fresh tissue was placed in 20 mlice-cold DMEM culture medium containing 200 IU penicillin/ml and200 mg streptomycin/ml. Myocytes were prepared by enzymatic dis-persion (see below).
For all biopsies ethical approval was obtained from the local ethicalcommittee, and each patient gave informed consent to the study.
Chemicals
Ovine (o) CRH, Tyr-oCRH, and all other chemicals were purchasedfrom Sigma Chemical Co. (Poole, UK). Waters Sep-Pak C18 columnswere obtained from Millipore Corp. (UK) Ltd. (Watford, UK). OT an-tagonists, CAP476 (1-deamino-2-d-Tyr-(OEt)-4-Thr-8-Orn-OT) and F327(des-Gly9-[d-Tyr(Et)2,Thr4,Orn8]dC6-OT), were gifts from Ferring Phar-maceuticals Ltd. (Malmo, Sweden).
125I-Labeled Nle21,Tyr32-oCRH (SA, 922 Ci/mmol), human/rat (h/r)CRH, and OT were obtained from Peninsula Laboratories (Merseyside,UK).
Phorbol myristate acetate (PMA), H-7 and H-8 (PKC inhibitors), andU73122 were obtained from Calbiochem (La Jolla, CA). The myristoy-lated PKC peptide (Myr-PKC) inhibitor was purchased from PromegaCorp. (Madison, WI). The cAMP assay kits were obtained from DuPont-New England Nuclear (Hertfordshire, UK). Protein A-Sepharose beads(CL-4B) were purchased from Pharmacia Biotech (Uppsala, Sweden).
A specific CRH receptor antibody was obtained from Santa CruzBiotechnology, Inc. (Santa Cruz, CA). This is a goat polyclonal antibodyraised against a peptide corresponding to amino acids 425–444 mappingat the C-terminus of the human CRH-R1 precursor. The antibody reactswith both human CRH-R1 and -R2 receptors, and this has been verifiedin preliminary studies by Western blotting and immunohistochemistry.
Myocyte isolation and culture: preparation of myometrialmembranes from biopsies
Pieces of myometrium were transferred into DMEM containing col-lagenase (300 U/ml), deoxyribonuclease (30 U/ml), penicillin (200U/ml), and streptomycin (200 mg/ml) and incubated at 37 C for 30 min.After filtration and centrifugation, cells were suspended in DMEM con-taining 10% FCS, penicillin (100 U/ml), streptomycin (100 mg/ml), andfungizone (2.5 mg/ml). The cells were kept at 37 C in a humidifiedatmosphere of 95% air and 5% CO2 until confluent (2–4 weeks). The
purity of myometrial muscle cells was assessed by immunocytochemicalstaining. Mouse antihuman smooth muscle actin-specific monoclonalantibody and peroxidase-conjugated rabbit antimouse antibody wereused. Human fibroblast cells and omission of the primary antibody wereused as negative controls; frozen myometrial tissue was used as a pos-itive control.
Myometrial membranes were prepared by homogenization and dif-ferential centrifugation as described previously (1). Protein concentra-tions were measured using the bicinchoninic acid method (21). The finalpellet was resuspended in 3 ml extraction buffer and after homogeni-zation was aliquoted in 1.5 ml Eppendorf tubes (50 mg protein/tube) andstored at 270 C until use.
Cultured human myometrial cells were used only for those experi-ments on the OT-induced activation and subsequent translocation ofPKC from the cytosol to the cell membrane. The rest of the experimentalstudies used membrane homogenates prepared from myometrial biop-sies at term or preterm.
Binding studies on myometrial membranes:Scatchard analysis
Scatchard analysis was performed as previously described (1). Allexperiments were performed at 22 C for 2 h. For the control Scatchardplot, myometrial membrane suspensions (100–150 mg protein) wereincubated with 50 ml of different concentrations of 125I-labeledNle21,Tyr32-oCRH (10,000–700,000 cpm) and unlabeled oCRH (1,000-fold molar excess) in 50 ml incubation buffer A [50 mm Tris-HCl, 2 mmEGTA, 10 mm MgCl2, 0.1% BSA (wt/vol), and 0.15 mm bacitracin, pH7.2]. In an additional series of polypropylene tubes, 10 different con-centrations of 125I-labeled Nle21,Tyr32-CRH (10,000–400,000 cpm) wereincubated in the presence of varying concentrations of OT (1–500 nm)and unlabeled CRH (1,000-fold molar excess). Binding data were ana-lyzed by computer analysis with EBDA (22) and LIGAND (23).
Isoelectric focusing
Myometrial membrane suspensions (300–350 mg protein) were in-cubated with 50 ml [125I]oCRH (200,000–300,000 cpm) in the presence orabsence of OT (100 nm) diluted in buffer A. Nonspecific binding wasdetermined in the presence of unlabeled CRH (1 mm). The reaction wascarried out at 22 C for 2 h. After the binding assay, pellets were resus-pended in 500 ml incubation buffer A containing 1% Triton X-100, in-cubated for 1–2 h at 0 C, and centrifuged at 12,000 rpm for 10 min. Thesolubilized membrane suspensions were collected, concentrated usingCentricon-30 microconcentrators, and fractionated by isoelectric focus-ing as previously described (24). Gels were stained and dried, andautoradiography was performed for 5–10 days at 270 C using Fuji PhotoFilm Co. Ltd. x-ray film (Tokyo, Japan) and intensifying screens. Therelative distribution of the human myometrial CRH receptor isoformswas measured by optical density scanning using the Image analysisprogram (Wayne Rasband, NIH, Bethesda, MD).
In preliminary experiments (24), we confirmed that the two isoformsat pI 4.6 and 4.9 represent free radioligand preparation and not receptor-ligand complex; thus, they were excluded from further analysis. Fur-thermore, no differences in the isoform profile were found when CRHwas covalently cross-linked to its binding sites using disuccinimidylsuberate (final concentration, 1.5 mm; for 10 min at room temperature);therefore, disuccinimidyl suberate was not used in subsequentexperiments.
cAMP studies
Human pregnant (at term or preterm) myometrial membrane prep-arations (50 mg protein) were preincubated with different concentrationsof h/rCRH (0.1–1000 nm) or OT (100 nm) in 50 ml extraction buffer in thepresence or absence of the Myr-PKC inhibitor (final concentration, 100mm) for 30 min at 22 C before the addition of 100 ml 50 mm Tris-HClcontaining 10 mm MgCl2, 1 mm EGTA, 0.1% BSA, 1 mm ATP, ATPregeneration system (7.4 mg/ml creatine phosphate and 1 mg/ml cre-atine phosphokinase), 100 mm isobutyrmethylxanthine (phosphodies-terase inhibitor), and 0.15 mm bacitracin, pH 7.4 (cAMP assay buffer),at 37 C. The reaction was terminated after 10 min by the addition of 1
586 INTERACTIONS BETWEEN OT AND CRH IN HUMAN PREGNANT MYOMETRIUM Endo • 1999Vol 140 • No 2

ml 0.1 m imidazole buffer, pH 7, followed by heating the tubes in boilingwater for 5 min.
For the pertussis toxin (PT) pretreatment of the myometrial mem-brane preparations, PT was preactivated with 50 mm dithiothreitol.Human pregnant myometrial membranes at term were incubated for 50min at 30 C with 500 ml 50 mm Tris-HCl containing 1 mm ATP, 1 mmthymidine, 0.1 mm GTP, 10 mm MgCl2, 1 mg/ml digitonin, 1 mm NAD,7.4 mg/ml creatine phosphate, and 1 mg/ml creatine phosphokinase,pH 7.4, in the presence or absence of PT (final concentration, 25 mg/ml)for 45 min at 30 C. The reaction was stopped by the addition of 1 mlice-cold extraction buffer, followed by centrifugation at 10,000 rpm in aBeckman Coulter, Inc., J20 centrifuge (Palo Alto, CA) for 15 min. Thepellet was then washed with extraction buffer and centrifuged at 10,000rpm for 15 min (three times). The final pellet was resuspended in ex-traction buffer and homogenized. Pretreated human myometrial mem-branes were preincubated with different concentrations of OT (1–500nm) for 30 min at 22 C before the addition of 50 ml cAMP assay bufferand initiation of the cAMP release reaction at 37 C.
The amount of cAMP in the incubate was estimated in the superna-tants by RIA using commercial cAMP RIA kits. Standard cAMP con-centrations, covering the range 100–0.5 pmol/ml, were used for deter-mination of the standard curve of the RIA. The interassay coefficient ofvariation was 5%.
PKC assay in human myometrial cell cultures andmyometrial membranes
Human pregnant myometrial cells, at a density of around 106 cells/flask, were preincubated for 15 min at 37 C with 5 ml PBS containing OT(1–100 nm) in the presence or absence of 10 mm U73122 (PLC inhibitor)or OT inhibitors (1 mm). After the incubation period, cells were washedwith 5 ml PBS (twice) and were removed from the bottom of the dishesand resuspended in 0.5 ml chilled extraction buffer [Dulbecco’s PBScontaining 10 mm MgCl2, 2 mm EGTA, 1.5 mg/liter BSA (wt/vol), 10 mmb-mercaptoethanol, 0.15 mm bacitracin, and 1 mm phenylmethylsulfo-nylfluoride, pH 7.2]. The suspensions were homogenized for a fewseconds and centrifuged for 1 h at 50,000 3 g. Supernatants were stored(at 0 C), and the membrane fraction was resuspended in 0.5 ml extractionbuffer containing 1% Triton X-100 and solubilized for 1 h at roomtemperature. After centrifugation and collection of the resulting super-natants, all samples (supernatants) were loaded onto 1-ml diethylami-noethyl (DEAE)-cellulose columns, and the PKC-containing fraction waseluted using 1 ml extraction buffer containing 200 mm NaCl. Sampleswere frozen at 270 C until assayed for PKC activity.
Human myometrial membranes (250 mg/tube) were resuspended in50 ml buffer containing 20 mm Tris-HCl and 10 mm MgCl2, pH 7.5.Membrane suspensions were incubated in the presence or absence ofPKC activators (PMA, 200 nm; OT, 1–100 nm) or inhibitors (H-7, 100 mm;H-8, 100 mm; myristoylated PKC peptide inhibitor, 100 mm) for 30 minat room temperature. Membranes were solubilized, and PKC was ex-tracted as described using DEAE-cellulose columns.
The activity of PKC recovered from either human myometrial cells ormembrane preparations was assayed in a 10 mm Tris-HCl buffer (pH 7.5)containing 10 mm MgCl2, 0.2 mm CaCl2, 0.5 mm EGTA, and 0.1 mg/mlBSA, using a commercial PKC assay system in which the incorporationof 32P into a specific PKC biotinylated peptide substrate (neurogranin-(28–43)-AAKIQASFRGHMAR KK) (25) using [g-32P]ATP (3000 Ci/mmol) as a 32P donor was measured by the binding of the substrate tostreptavidin disks and monitoring the radioactivity using a scintillationcounter.
In vitro phosphorylation of the myometrial CRH receptor
Human pregnant (at term or preterm) myometrial membrane sus-pensions (500 mg/tube) were incubated in the presence or absence of OT(100 nm) with or without myristoylated PKC peptide inhibitor (100 mm)for 10 min at 30 C. [g-32P]ATP (3000 Ci/mmol) was added as a 32P-donor (4 mCi/tube). The reaction was stopped by centrifugation (15,000rpm in a microfuge), and membranes were solubilized for 2 h on ice in500 ml solubilization buffer containing 10 mm Tris-HCl, 10 mm EDTA,500 mm NaCl, 1% Nonidet P-40, 0.5% deoxycholate, 0.1% SDS, 1 mmphenylmethylsulfonylfluoride, and 1 mm bacitracin, pH 7.4. After cen-trifugation, immunoprecipitation of the CRH-receptor complex was per-
formed by adding 25 ml CRH receptor antiserum to the supernatant andincubating overnight at 4 C. Isolation of the immune complexes wascarried out using protein A-Sepharose beads (100 ml), rotation for 20 minat 4 C, centrifugation, washing of the resulting pellet with 1 ml Tris-EDTA buffer (three times), and resuspension in SDS buffer. Proteinswere resolved on 8% SDS-PAGE, and gels were dried and subjected toautoradiography (270 C, 10–14 days) using intensifying screens.
Statistical analysis
Data are shown as the mean 6 sem of each measurement. Comparisonbetween group means was performed using the nonparametric statis-tical method of Kruskal-Wallis. P , 0.05 was considered significant.
ResultsEffect of OT on CRH binding to the CRH receptors
In preliminary experiments we found that OT was able toreduce CRH binding to human pregnant (at term) myome-trial membranes. This OT effect was time dependent; whenOT was added at equilibrium there was a lag phase of 15–20min in the OT inhibitory effect. Maximum inhibition (50%)was observed at 30 min and remained constant for at least 2 h.The lag phase might be due to our experimental conditionsor, alternatively, might be the result of factors missing in themembrane preparation that are necessary for efficient andrapid receptor desensitization and/or the presence of con-taminating adenosine triphosphatases, which contribute tothe low rate of the reaction. Although we cannot ignore thepossibility of receptor degradation, the experimental condi-tions were designed in such a way as to minimize this pos-sibility by including several protease inhibitors.
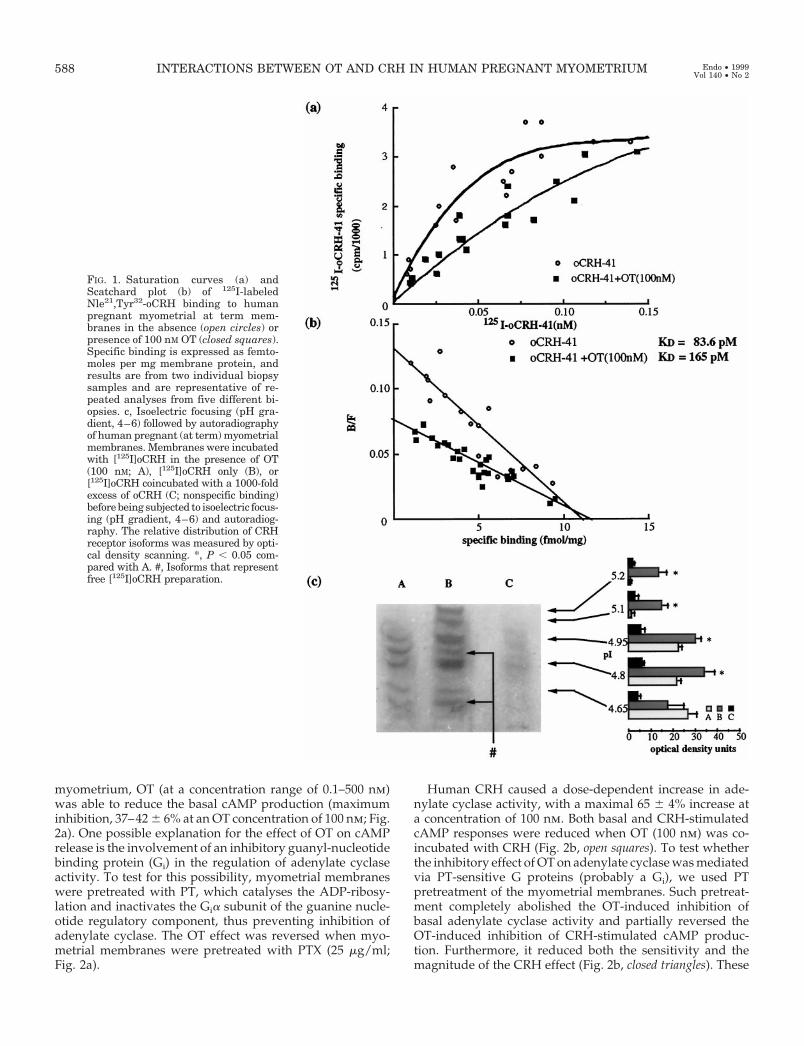
Saturation analysis of 125I-labeled Nle21,Tyr32-oCRHbinding to human pregnant myometrial membranes dem-onstrated that binding was saturable and of high affinity(Fig. 1a). When the myometrial membranes were incu-bated with radiolabeled oCRH in the presence of OT for2 h, the receptor required a greater concentration of ra-dioligand to reach saturation. Scatchard analysis of thesedata (Fig. 1b) showed that the OT effect was due to areduction in the affinity of the CRH receptor. The disso-ciation constant (Kd) was increased from 84 6 4 to 165 612 pm, whereas the maximum receptor concentration wasunchanged (11 6 2 fmol/mg membrane protein). Thiseffect was OT concentration dependent, was apparent atOT concentrations greater than 1 nm, and was maximal ata concentration around 100 nm (Table 1). This OT con-centration was used in all subsequent experiments.
In membrane preparations from human pregnant myo-metrial tissue, we identified multiple CRH-binding sites us-ing isoelectric focusing (24). The effect of OT on radiolabeledoCRH binding to each of the isoforms was analyzed. OTcaused decreased binding to four of the five isoforms; inparticular, CRH binding to the two most basic isoforms (pI5.1 and 5.2) was completely inhibited (Fig. 1c). Analysis ofmembrane preparations incubated with a 1000-fold excess ofunlabeled h/rCRH showed an equal displacement of bound[125I]oCRH from each of the five receptor isoforms.
Effect of OT on basal and CRH-stimulatedcAMP production
This OT effect was analyzed in more detail at a functionallevel. In membranes prepared from human pregnant term
INTERACTIONS BETWEEN OT AND CRH IN HUMAN PREGNANT MYOMETRIUM 587
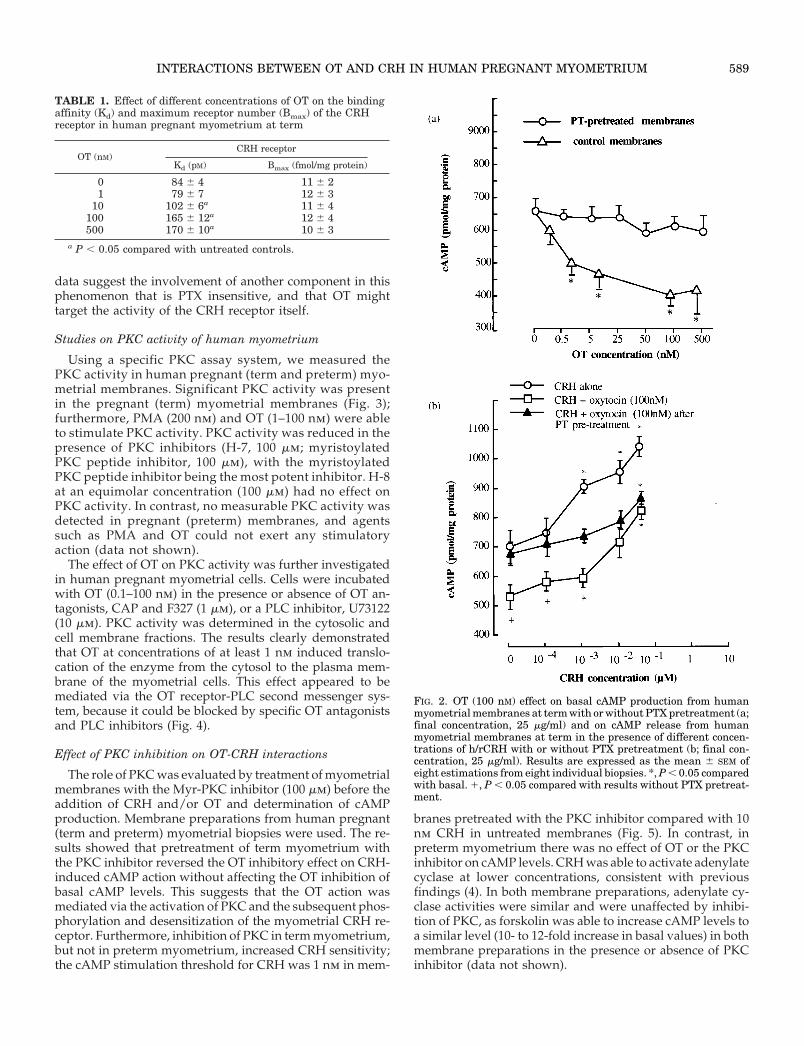
myometrium, OT (at a concentration range of 0.1–500 nm)was able to reduce the basal cAMP production (maximuminhibition, 37–42 6 6% at an OT concentration of 100 nm; Fig.2a). One possible explanation for the effect of OT on cAMPrelease is the involvement of an inhibitory guanyl-nucleotidebinding protein (Gi) in the regulation of adenylate cyclaseactivity. To test for this possibility, myometrial membraneswere pretreated with PT, which catalyses the ADP-ribosy-lation and inactivates the Gia subunit of the guanine nucle-otide regulatory component, thus preventing inhibition ofadenylate cyclase. The OT effect was reversed when myo-metrial membranes were pretreated with PTX (25 mg/ml;Fig. 2a).
Human CRH caused a dose-dependent increase in ade-nylate cyclase activity, with a maximal 65 6 4% increase ata concentration of 100 nm. Both basal and CRH-stimulatedcAMP responses were reduced when OT (100 nm) was co-incubated with CRH (Fig. 2b, open squares). To test whetherthe inhibitory effect of OT on adenylate cyclase was mediatedvia PT-sensitive G proteins (probably a Gi), we used PTpretreatment of the myometrial membranes. Such pretreat-ment completely abolished the OT-induced inhibition ofbasal adenylate cyclase activity and partially reversed theOT-induced inhibition of CRH-stimulated cAMP produc-tion. Furthermore, it reduced both the sensitivity and themagnitude of the CRH effect (Fig. 2b, closed triangles). These
FIG. 1. Saturation curves (a) andScatchard plot (b) of 125I-labeledNle21,Tyr32-oCRH binding to humanpregnant myometrial at term mem-branes in the absence (open circles) orpresence of 100 nM OT (closed squares).Specific binding is expressed as femto-moles per mg membrane protein, andresults are from two individual biopsysamples and are representative of re-peated analyses from five different bi-opsies. c, Isoelectric focusing (pH gra-dient, 4–6) followed by autoradiographyof human pregnant (at term) myometrialmembranes. Membranes were incubatedwith [125I]oCRH in the presence of OT(100 nM; A), [125I]oCRH only (B), or[125I]oCRH coincubated with a 1000-foldexcess of oCRH (C; nonspecific binding)before being subjected to isoelectric focus-ing (pH gradient, 4–6) and autoradiog-raphy. The relative distribution of CRHreceptor isoforms was measured by opti-cal density scanning. *, P , 0.05 com-pared with A. #, Isoforms that representfree [125I]oCRH preparation.
588 INTERACTIONS BETWEEN OT AND CRH IN HUMAN PREGNANT MYOMETRIUM Endo • 1999Vol 140 • No 2
data suggest the involvement of another component in thisphenomenon that is PTX insensitive, and that OT mighttarget the activity of the CRH receptor itself.
Studies on PKC activity of human myometrium
Using a specific PKC assay system, we measured thePKC activity in human pregnant (term and preterm) myo-metrial membranes. Significant PKC activity was presentin the pregnant (term) myometrial membranes (Fig. 3);furthermore, PMA (200 nm) and OT (1–100 nm) were ableto stimulate PKC activity. PKC activity was reduced in thepresence of PKC inhibitors (H-7, 100 mm; myristoylatedPKC peptide inhibitor, 100 mm), with the myristoylatedPKC peptide inhibitor being the most potent inhibitor. H-8at an equimolar concentration (100 mm) had no effect onPKC activity. In contrast, no measurable PKC activity wasdetected in pregnant (preterm) membranes, and agentssuch as PMA and OT could not exert any stimulatoryaction (data not shown).
The effect of OT on PKC activity was further investigatedin human pregnant myometrial cells. Cells were incubatedwith OT (0.1–100 nm) in the presence or absence of OT an-tagonists, CAP and F327 (1 mm), or a PLC inhibitor, U73122(10 mm). PKC activity was determined in the cytosolic andcell membrane fractions. The results clearly demonstratedthat OT at concentrations of at least 1 nm induced translo-cation of the enzyme from the cytosol to the plasma mem-brane of the myometrial cells. This effect appeared to bemediated via the OT receptor-PLC second messenger sys-tem, because it could be blocked by specific OT antagonistsand PLC inhibitors (Fig. 4).
Effect of PKC inhibition on OT-CRH interactions
The role of PKC was evaluated by treatment of myometrialmembranes with the Myr-PKC inhibitor (100 mm) before theaddition of CRH and/or OT and determination of cAMPproduction. Membrane preparations from human pregnant(term and preterm) myometrial biopsies were used. The re-sults showed that pretreatment of term myometrium withthe PKC inhibitor reversed the OT inhibitory effect on CRH-induced cAMP action without affecting the OT inhibition ofbasal cAMP levels. This suggests that the OT action wasmediated via the activation of PKC and the subsequent phos-phorylation and desensitization of the myometrial CRH re-ceptor. Furthermore, inhibition of PKC in term myometrium,but not in preterm myometrium, increased CRH sensitivity;the cAMP stimulation threshold for CRH was 1 nm in mem-
branes pretreated with the PKC inhibitor compared with 10nm CRH in untreated membranes (Fig. 5). In contrast, inpreterm myometrium there was no effect of OT or the PKCinhibitor on cAMP levels. CRH was able to activate adenylatecyclase at lower concentrations, consistent with previousfindings (4). In both membrane preparations, adenylate cy-clase activities were similar and were unaffected by inhibi-tion of PKC, as forskolin was able to increase cAMP levels toa similar level (10- to 12-fold increase in basal values) in bothmembrane preparations in the presence or absence of PKCinhibitor (data not shown).
TABLE 1. Effect of different concentrations of OT on the bindingaffinity (Kd) and maximum receptor number (Bmax) of the CRHreceptor in human pregnant myometrium at term
OT (nM)CRH receptor
Kd (pM) Bmax (fmol/mg protein)
0 84 6 4 11 6 21 79 6 7 12 6 3
10 102 6 6a 11 6 4100 165 6 12a 12 6 4500 170 6 10a 10 6 3
a P , 0.05 compared with untreated controls.
FIG. 2. OT (100 nM) effect on basal cAMP production from humanmyometrial membranes at term with or without PTX pretreatment (a;final concentration, 25 mg/ml) and on cAMP release from humanmyometrial membranes at term in the presence of different concen-trations of h/rCRH with or without PTX pretreatment (b; final con-centration, 25 mg/ml). Results are expressed as the mean 6 SEM ofeight estimations from eight individual biopsies. *, P , 0.05 comparedwith basal. 1, P , 0.05 compared with results without PTX pretreat-ment.
INTERACTIONS BETWEEN OT AND CRH IN HUMAN PREGNANT MYOMETRIUM 589
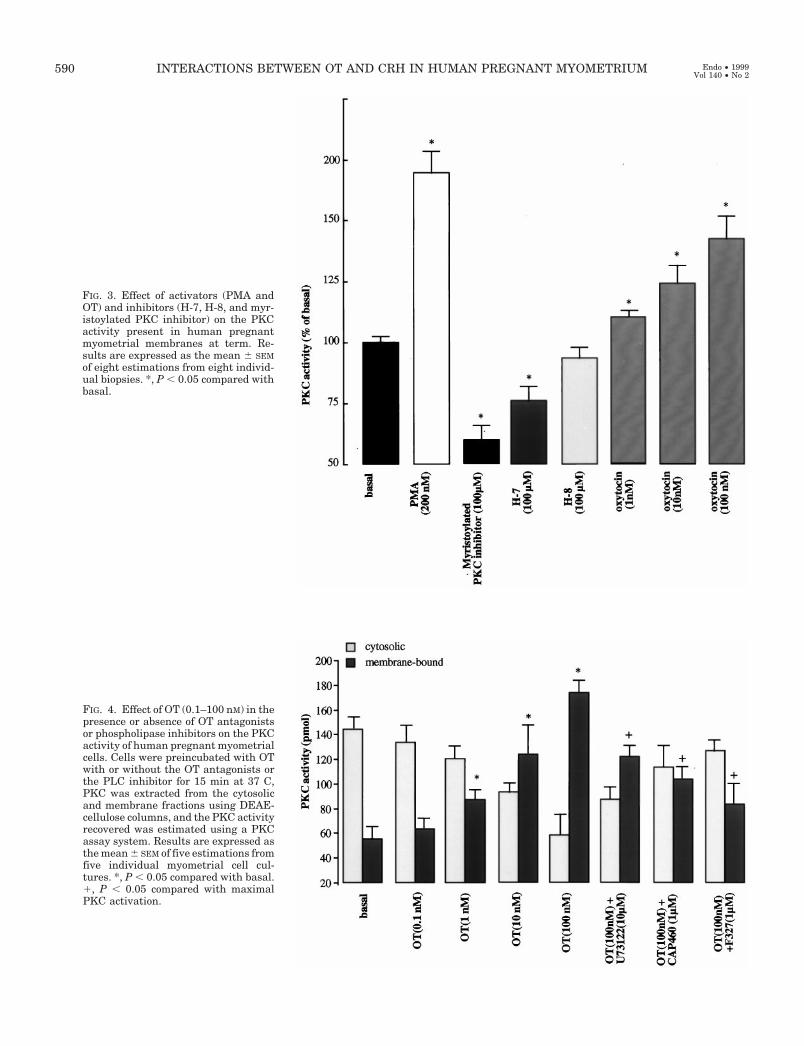
FIG. 3. Effect of activators (PMA andOT) and inhibitors (H-7, H-8, and myr-istoylated PKC inhibitor) on the PKCactivity present in human pregnantmyometrial membranes at term. Re-sults are expressed as the mean 6 SEMof eight estimations from eight individ-ual biopsies. *, P , 0.05 compared withbasal.
FIG. 4. Effect of OT (0.1–100 nM) in thepresence or absence of OT antagonistsor phospholipase inhibitors on the PKCactivity of human pregnant myometrialcells. Cells were preincubated with OTwith or without the OT antagonists orthe PLC inhibitor for 15 min at 37 C,PKC was extracted from the cytosolicand membrane fractions using DEAE-cellulose columns, and the PKC activityrecovered was estimated using a PKCassay system. Results are expressed asthe mean 6 SEM of five estimations fromfive individual myometrial cell cul-tures. *, P , 0.05 compared with basal.1, P , 0.05 compared with maximalPKC activation.
590 INTERACTIONS BETWEEN OT AND CRH IN HUMAN PREGNANT MYOMETRIUM Endo • 1999Vol 140 • No 2
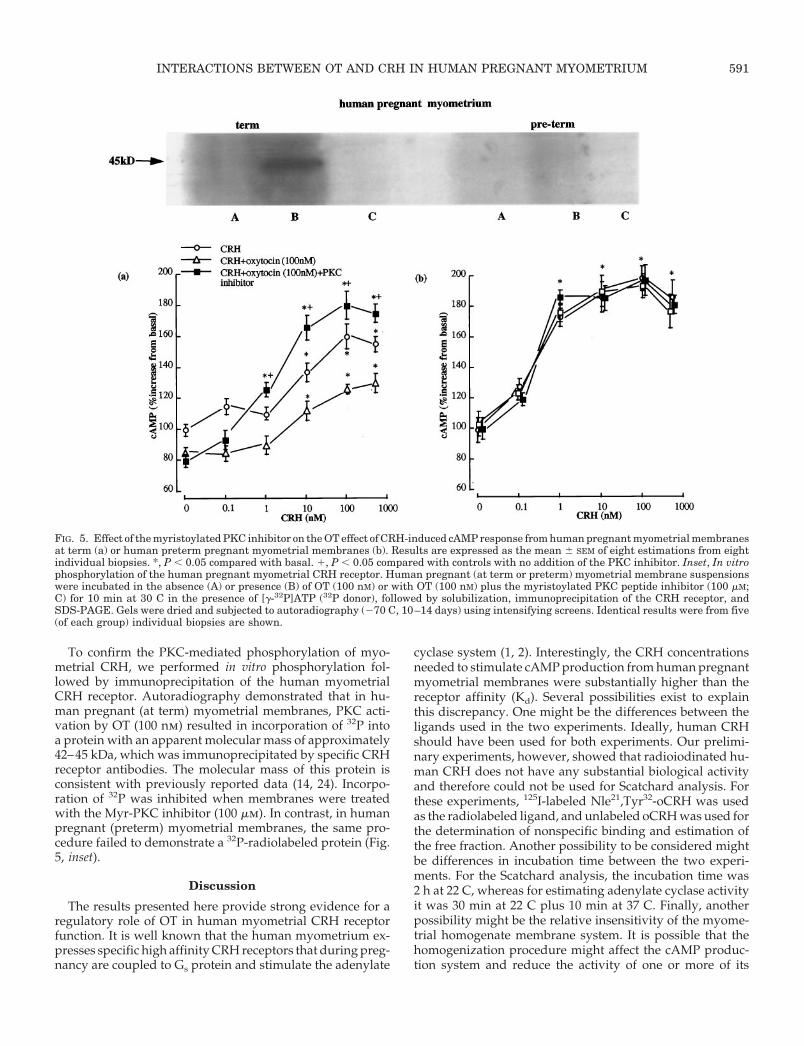
To confirm the PKC-mediated phosphorylation of myo-metrial CRH, we performed in vitro phosphorylation fol-lowed by immunoprecipitation of the human myometrialCRH receptor. Autoradiography demonstrated that in hu-man pregnant (at term) myometrial membranes, PKC acti-vation by OT (100 nm) resulted in incorporation of 32P intoa protein with an apparent molecular mass of approximately42–45 kDa, which was immunoprecipitated by specific CRHreceptor antibodies. The molecular mass of this protein isconsistent with previously reported data (14, 24). Incorpo-ration of 32P was inhibited when membranes were treatedwith the Myr-PKC inhibitor (100 mm). In contrast, in humanpregnant (preterm) myometrial membranes, the same pro-cedure failed to demonstrate a 32P-radiolabeled protein (Fig.5, inset).
Discussion
The results presented here provide strong evidence for aregulatory role of OT in human myometrial CRH receptorfunction. It is well known that the human myometrium ex-presses specific high affinity CRH receptors that during preg-nancy are coupled to Gs protein and stimulate the adenylate
cyclase system (1, 2). Interestingly, the CRH concentrationsneeded to stimulate cAMP production from human pregnantmyometrial membranes were substantially higher than thereceptor affinity (Kd). Several possibilities exist to explainthis discrepancy. One might be the differences between theligands used in the two experiments. Ideally, human CRHshould have been used for both experiments. Our prelimi-nary experiments, however, showed that radioiodinated hu-man CRH does not have any substantial biological activityand therefore could not be used for Scatchard analysis. Forthese experiments, 125I-labeled Nle21,Tyr32-oCRH was usedas the radiolabeled ligand, and unlabeled oCRH was used forthe determination of nonspecific binding and estimation ofthe free fraction. Another possibility to be considered mightbe differences in incubation time between the two experi-ments. For the Scatchard analysis, the incubation time was2 h at 22 C, whereas for estimating adenylate cyclase activityit was 30 min at 22 C plus 10 min at 37 C. Finally, anotherpossibility might be the relative insensitivity of the myome-trial homogenate membrane system. It is possible that thehomogenization procedure might affect the cAMP produc-tion system and reduce the activity of one or more of its
FIG. 5. Effect of the myristoylated PKC inhibitor on the OT effect of CRH-induced cAMP response from human pregnant myometrial membranesat term (a) or human preterm pregnant myometrial membranes (b). Results are expressed as the mean 6 SEM of eight estimations from eightindividual biopsies. *, P , 0.05 compared with basal. 1, P , 0.05 compared with controls with no addition of the PKC inhibitor. Inset, In vitrophosphorylation of the human pregnant myometrial CRH receptor. Human pregnant (at term or preterm) myometrial membrane suspensionswere incubated in the absence (A) or presence (B) of OT (100 nM) or with OT (100 nM) plus the myristoylated PKC peptide inhibitor (100 mM;C) for 10 min at 30 C in the presence of [g-32P]ATP (32P donor), followed by solubilization, immunoprecipitation of the CRH receptor, andSDS-PAGE. Gels were dried and subjected to autoradiography (270 C, 10–14 days) using intensifying screens. Identical results were from five(of each group) individual biopsies are shown.
INTERACTIONS BETWEEN OT AND CRH IN HUMAN PREGNANT MYOMETRIUM 591

components. In support of this hypothesis, we obtained datasuggesting that in cultured human myometrial cells h/rCRHis able to stimulate cAMP production at a concentration aslow as 100 pm (unpublished observations).
At term, the ability of the myometrial CRH receptors toactivate adenylate cyclase is impaired (3), and our resultsimplicate OT in this phenomenon. The decidua has beenshown to express high concentrations of messenger RNA forOT (7), and the translated peptide may reach high concen-trations locally and exert a paracrine effect. Furthermore, thenumber of OT receptors can increase as much as 200-fold (6).We now report that OT is able to reduce CRH binding to itsmyometrial receptor by reducing the affinity of the CRHreceptor without affecting the number of receptors. This ef-fect is mediated by at least four CRH receptor isoforms, asidentified by isoelectric focusing. Previously, we have shownthat the human myometrium expresses five isoforms of theCRH receptor, which can be separated by charge (24). How-ever, a paradox appears to exist, as Scatchard analysis sug-gested a single population of high affinity CRH receptors. Itis possible that the different isoforms separated with iso-electric focusing might represent receptor heterogeneity dueto different glycosylation forms of the same receptor duringposttranslational modification that display the same bindingaffinity. It is well known that there are several potentialN-glycosylation sites in both R1 and R2 CRH receptor aminoacid sequences (14, 15). In support of this hypothesis was theobservation that the isoform profile was identical in bothpregnant and nonpregnant myometrium (24), which sug-gests that the difference in CRH binding affinity and functionin these two tissues is not due to differences in the receptorisoform population.
In human pregnant myometrium at term, OT was able toreduce both basal and CRH-stimulated cAMP levels, inagreement with previous data (26). The OT effect on basaladenylate cyclase activity appears to be mediated by a PTX-sensitive G protein, most likely Gi protein, which is consis-
tent with previous observations (10). Such PTX-sensitive aswell as PTX-insensitive G proteins have been shown to beinvolved in OT stimulation of phosphoinositol hydrolysis inhuman myometrial cells (10). Inhibition of Gi activity by PTX,however, did not completely reverse the OT effect on CRH-stimulated cAMP, suggesting the presence of an additionalmechanism. Subsequently, we showed that OT as well asphorbol esters (PMA) were able to stimulate PKC in humanpregnant (at term but not preterm) myometrial membranes,an effect that could be reversed in the presence of PKCinhibitors. A possible explanation for the lack of PKC activityin preterm myometrium is the immaturity of the myome-trium, which lacks sufficient OT receptors as well as sensi-tizing stimuli capable of inducing myometrial PKC activityin the cell membrane.
Most importantly, we showed that in human pregnantmyometrial cells, OT was able to induce PKC translocationfrom the cytosol to the cell membrane. This phenomenon hasbeen suggested previously, and interactions between PKCactivation/inhibition and the OT signaling pathway havebeen described in human myometrium (27) and amnion cells(28), although no direct PKC stimulation has been shown.Several isoforms of PLC (b1, b2, b3, g1, and b2) have beendescribed in human myometrial cells (26), and this pathwaywas investigated in more detail using specific OT antagonistsand the PLC inhibitor U73122 (29); partial inhibition of theOT-induced translocation of PKC activity was achieved.Only one dose of OT antagonists was used, so as to dem-onstrate the effect of the antagonists rather than investigatedose-response characteristics. The actions of OT antagonistsused have been well described; they inhibit OT-induced cal-cium influx (30) and inositol triphosphate generation in myo-metrial cells (31).
These results in association with the presence of four po-tential PKC phosphorylation sites in each of the three intra-cellular loops as well as in the C-terminus of the comple-mentary DNA sequence of the CRH-R1 and -R2 receptors led
FIG. 6. A schematic representation ofthe proposed interactions between CRHand OT signaling pathways in the hu-man myometrium during pregnancy atterm. CRH-R, CRH receptor; AC, ade-nylate cyclase.
592 INTERACTIONS BETWEEN OT AND CRH IN HUMAN PREGNANT MYOMETRIUM Endo • 1999Vol 140 • No 2

us to investigate whether the inhibitory action of OT wasmediated by PKC-induced phosphorylation and desensiti-zation of the CRH receptor. There are several examples of7TMD receptors where PKC-induced phosphorylation anddesensitization are accompanied by a reduction in the re-ceptor affinity for the agonist, presumably by impairment ofreceptor-G protein coupling (32–34). Furthermore, interac-tions between the CRH signaling pathway and PKC havebeen described previously; in the pituitary, PKC activationpotentiates CRH-stimulated cAMP production (35). In termpregnant myometrium, the inhibitory action of OT on CRH-stimulated cAMP production was reversed in the presence ofthe Myr-PKC inhibitor, and the sensitivity of the CRH re-ceptor was increased. In preterm myometrium there was noeffect of either OT or the PKC inhibitor on CRH-stimulatedcAMP production. Although we cannot exclude other ac-tions of OT on the CRH receptor resulting in increased sen-sitivity, we obtained further evidence for a direct phosphor-ylation effect of PKC (induced by OT) on the CRH receptoronly in term myometrium with a direct phosphorylationassay.
At present, it is not clear how PKC can potentiate CRHaction in the pituitary while inhibiting it in the myometriumat term. It is possible that different PKC isoenzymes areinvolved with tissue-specific action or that the PKC actionstarget different CRH-R subtypes. Recently, in pregnant myo-metrium we have identified several CRH-R messenger RNAtranscripts (20). These correspond to four spliced variants ofthe R1 receptor (1a, 1b, 1C, and a novel form named 1d) andthe R2a receptor. At the moment it is uncertain which ofthese subtypes mediates the OT effect, and studies usingrecombinant receptor subtypes are necessary to delineatethese events.
In summary, these data suggest that in human pregnant (atterm) myometrium there is an inhibitory action of OT onCRH biological activity that is mediated via activation ofPKC and subsequent phosphorylation and desensitization ofthe CRH receptor. CRH-OT interactions may play a primary,and important, role in the fine balance of myometrial con-tractility. Furthermore, these interactions appear to be de-pendent on both OT receptor expression (36) and up-regu-lation of PKC activity at term. Currently, the role of CRHduring pregnancy is unknown, and the presence of multipleCRH receptor subtypes in the human pregnant (at term)myometrium suggests distinct functional roles for each re-ceptor during pregnancy and raises the possibility of mul-tiple roles for CRH and/or related peptides. It is attractive tospeculate that the OT-sensitive CRH receptors are respon-sible mainly for the generation of cAMP and the stimulationof myometrial relaxation; inhibition of their biological activ-ity by OT may enable CRH to play a different role in thecontrol of uterine contractility and the mechanism of labor.We speculate that the major CRH receptor subtype influ-enced by OT is R1a, as in transfection studies we have shownthat this subtype is most efficiently linked to adenylate cy-clase (unpublished observations). The interactions of theCRH and OT signal transduction pathways are shown dia-grammatically in Fig. 6. Further studies are under way toinvestigate in-depth the role of phosphorylation on the func-tion of the CRH receptor by using mutant receptor technol-
ogy where the potential PKC phosphorylation sites havebeen mutated.
References
1. Hillhouse EW, Grammatopoulos D, Milton NGN, Quartero HWP 1993 Theidentification of a human myometrial corticotropin-releasing hormone recep-tor that increases in affinity during pregnancy. J Clin Endocrinol Metab.76:736–41
2. Grammatopoulos D, Milton NGN, Hillhouse EW 1994 The human myome-trial CRH receptor: G proteins and second messengers. Mol Cel Endocrinol99:245–250
3. Grammatopoulos D, Stirrat GM, Williams SA, Hillhouse EW 1996 The bi-ological activity of the corticotropin-releasing hormone receptor-adenylatecyclase complex in human myometrium is reduced at the end of pregnancy.J Clin Endocrinol Metab. 81:745–751
4. Europe-Finner GN, Phaneuf S, Tolkovsky AM, Watson SP, Lopez-Bernal A1994 Down-regulation of Gas in human myometrium in term and pretermlabor: a mechanism for parturition. J Clin Endocrinol Metab. 79:1835–1839
5. Fuchs AR, Fuchs F 1984 Endocrinology of human parturition: a review. Br JObstet Gynaecol. 91:948–967
6. Fuchs AR, Fuchs F, Husslein P, Soloff MS, Fernstrom MJ 1982 Oxytocinreceptors and human parturition: a dual role for oxytocin in the initiation oflabor. Science 215:1396–1398
7. Chibbar R, Miller FD, Mitchell BF 1993 Synthesis of oxytocin in amnion,chorion, and decidua may influence the timing of human parturition. J ClinInvest 91:185–192
8. Nishimori K, Young LJ, Guo Q, Wang Z, Insel TR, Matzuk MM 1996 Oxy-tocin is required for nursing but is not essential for parturition or reproductivebehavior. Proc Natl Acad Sci USA 93:11699–11704
9. Kimura T, Tanizawa O, Mri K, Brownstein MJ, Okayama H 1992 Structureand expression of a human oxytocin receptor. Nature 336:526–529
10. Phaneuf S, Europe-Finner GN, Varney M, MacKenzie IZ, Watson SP, LopezBernal A 1993 Oxytocin stimulated phosphoinositide hydrolysis in humanmyometrial cells: involvement of pertussis toxin-sensitive and -insensitiveG-proteins. J Endocrinol 136:497–509
11. Berridge MJ, Irvine RF 1989 Inositol phosphates and cell signalling. Nature341:197–205
12. Nishizuka Y 1988 The molecular heterogeneity of protein kinase C and itsimplications of cellular regulation. Nature 334:661–665
13. Sutherland CW, Oye I, Butcher RW 1965 The action of epinephrine and therole of the adenylyl cyclase system in hormone action. Recent Prog Horm Res21:623–646
14. Chen R, Lewis K, Perrin MH, Vale WW 1993 Expression cloning of a humancorticotropin-releasing factor receptor. Proc Natl Acad Sci USA 90:8967–8971
15. Liaw CW, Lovenberg TW, Barry G, Oltersdorf T, Grigoriadis DE, DeSouzaEB 1996 Cloning and characterisation of a corticotropin-releasing factor-2receptor cDNA. Endocrinology 137:72–77
16. Ross PC, Kostas CM, Ramabhadran TV 1994 A variant of the human corti-cotropin-releasing factor (CRF) receptor: cloning, expression and pharmacol-ogy. Biochem Biophys Res Commun. 205:1836–1842
17. Lovenberg TW, Chalmers DT, Liu C, DeSouza EB 1995 CRF1a and CRF2b
receptor mRNAs are differentially distributed between the rat central nervoussystem and peripheral tissues. Endocrinology 136:4139–4142
18. Valdenaire O, Giller T, Breu V, Gottowik J, Kilpatrick G 1997 A new func-tional isoform of the human CRF2 receptor for corticotropin-releasing factor.Biochim Biophys Acta 1352:129–132
19. Sperle K, Chen A, Kostich W, Largent BL, CRH-2g: a novel CRH2 receptorisoform found in the human brain. Soc Neurosci Abstr 23:1765 (Abstract)
20. Grammatopoulos D, Dai Y, Chen J, Karteris E, Papadopoulou N, Easton AJ,Hillhouse EW 1998 Human CRH receptor: differences in subtype expressionbetween pregnant and non-pregnant myometrium. J Clin Endocrinol Metab.83:2539–2544
21. Smith P, Krohn R, Hermanson G, Mallia AK, Gartner FH, Provenzano MD,Fujimoto EK, Goeke NM, Olson BJ, Klenk DC 1985 Measurement of proteinusing bicinchoninic acid. Anal Biochem 150:76–85
22. McPherson G 1983 A practical computer based approach to the analysis ofradioligand binding experiments. Comput Prog Biomed 17:107–114
23. Munson P, Rodgbard D 1980 LIGAND: a versatile computerised approach forcharacterisation of ligand binding systems. Anal Biochem 107:220–239
24. Grammatopoulos D, Thompson S, Hillhouse EW 1995 The human myome-trium expresses multiple isoforms of the corticotropin releasing hormonereceptor. J Clin Endocrinol Metab 80:2388–2393
25. Chen SJ, Klann E, Gower MC, Powell CM, Sessoms JS, Sweatt JD 1993Studies with synthetic peptide substrates derived from the neuronal proteinneurogranin reveal structural determinants of potency and selectivity for pro-tein kinase C. Biochemistry 32:1032–1039
26. Quartero HWP, Srivatsa G, Gillham B 1992 Role for cyclic adenosine mono-phosphate in the synergistic interaction between oxytocin and corticotropin-releasing factor in isolated human gestational myometrium. Clin Endocrinol(Oxf) 36:141–145
INTERACTIONS BETWEEN OT AND CRH IN HUMAN PREGNANT MYOMETRIUM 593

27. Phaneuf S, Europe-Finner GN, Carasco MP, Hamilton CH, Lopez Bernal A1995 Oxytocin signalling in human myometrium. In: Ivell R, Russell J (eds)Oxytocin. Plenum Press, New York, pp 453–467
28. Moor JJ, Dubyak GR, Moore RM, Kooy DV 1988 Oxytocin activates theinositol-phospholipid-protein kinase C system and stimulates prostaglandinproduction in human amnion cells. Endocrinology 123:1771–1777
29. Yule DI, Williams JA 1992 U73122 inhibits Ca21 oscillations in response tocholecystokinin and carbachol but not to JMV-180 in rat pancreatic acinar cells.J Biol Chem 267:13830–13835
30. Thornton S, Gillespie JI, Anson LC, Greenwell JR, Melin P, Dunlop W 1993The effect of the oxytocin antagonists CAP476 and F327 on calcium mobil-isation in single cultured human myometrial cells. Br J Obstet Gynaecol.100:581–586
31. Phaneuf S, Asboth G, MacKenzie MA, Melin P, Lopez Bernal A 1994 Effectof oxytocin antagonists on the activation of human myometrium in vitro:
atosiban prevents oxytocin-induced desensitisation. Am J Obstet Gynecol.171:1627–1634
32. Findlay DM, Michelangeli VP, Robinson PJ 1989 Protein kinase C-induceddown regulation of calcitonin receptors and calcitonin-activated adenylatecyclase in T47D and BEN cells. Endocrinology 125:2656–2663
33. Bouvier M, Guilbault N, Bonin H 1991 Phorbol-ester-induced phosphoryla-tion of the b-2-adrenergic receptor decreases its coupling to Gs. FEBS Lett279:243–248
34. Harrington MA, Shaw K, Zhong P, Ciaranello RD 1994 Agonist-induceddesensitization and loss of high-affinity binding sites of stably expressedhuman 5-HT1A receptors. J Pharmacol Exp Ther 268:1098–1106
35. Cronin MJ, Zysk JR, Baertschi AJ 1986 Protein kinase C potentiates cortico-tropin releasing factor stimulated cyclic AMP in pituitary. Peptides 7:935–938
36. Soloff MS, Alexandrova M, Fernstrom J 1979 Oxytocin receptors, triggers forparturition and lactation? Science 204:1313–1315
594 INTERACTIONS BETWEEN OT AND CRH IN HUMAN PREGNANT MYOMETRIUM Endo • 1999Vol 140 • No 2




















