Neurohypophyseal hormones protect against pentylenetetrazole-induced seizures in zebrafish: Role of...
7
Please cite this article in press as: Braida D, et al. Neurohypophyseal hormones protect against pentylenetetrazole-induced seizures in zebrafish: Role of oxytocin-like and V1a-like receptor. Peptides (2012), http://dx.doi.org/10.1016/j.peptides.2012.07.013 ARTICLE IN PRESS G Model PEP-68749; No. of Pages 7 Peptides xxx (2012) xxx–xxx Contents lists available at SciVerse ScienceDirect Peptides j ourna l ho me pa ge: www.elsevier.com/locate/peptides Neurohypophyseal hormones protect against pentylenetetrazole-induced seizures in zebrafish: Role of oxytocin-like and V1a-like receptor Daniela Braida a , Andrea Donzelli a , Roberta Martucci a , Luisa Ponzoni a , Alberto Pauletti a , Mariaelvina Sala a,b,∗ a Dipartimento di Biotecnologie mediche e Medicina traslazionale, Università degli Studi di Milano, Via Vanvitelli 32, Milan, Italy b CNR, Institute of Neuroscience, Via Vanvitelli 32, Milan, Italy a r t i c l e i n f o Article history: Received 26 June 2012 Received in revised form 16 July 2012 Accepted 16 July 2012 Available online xxx Keywords: Epilepsy Anticonvulsant Neurohypophyseal Hormone Teleost a b s t r a c t Oxytocin (OT) and arginine-vasopressin (AVP) are involved in the physiological response to different stressors like the occurrence of seizures which is regarded as a severe stress factor. Zebrafish (Danio rerio) is recently featured as a model of epilepsy but the role of neurohypophyseal hormones on this teleost is still unknown. We attempted to determine whether non-mammalian homologues like isotocin (IT) and vasotocin (AVT) affected pentylenetetrazole (PTZ)-induced seizures in adult zebrafish in comparison with OT/AVP. The mechanism was studied using the most selective OT and AVP receptor antagonists. Zebrafish were injected i.m. with increasing doses (0.1–40 ng/kg) of the neuropeptides 10 min before PTZ exposure. DesGly-NH2-d(CH2)5-[D-Tyr2,Thr4]OVT (desglyDTyrOVT) for OT receptor and SR49059 for V1a subtype receptor, were injected together with each agonist 20 min before PTZ exposure. All the peptides significantly decreased the number of seizures, increased the mean latency time to the first seizure and decreased lethality. This protective effect led to a dose–response curve following a U-shaped form. IT was approximately 40 times more active than OT while AVT was 20 times more potent than AVP in reducing the number of seizures. DesglyDTyrOVT was more effective in antagonizing OT/IT, while SR49059 mainly blocked AVP/AVT-induced protection against PTZ-induced seizures. The present findings provide direct evidence of an important involvement of IT/OT and AVP/AVT as anticonvulsant agents against PTZ-induced seizures with a receptor-mediated mechanism in zebrafish. These data reinforce zebrafish as an emerging experimental model to study and identify new antiepileptic drugs. © 2012 Elsevier Inc. All rights reserved. 1. Introduction Zebrafish (Danio rerio) have been recently proposed as a new genetic model to evaluate epilepsy [21,39]. Using different con- vulsive drugs, seizures were observed in embryos [43,44], larvae [4,41,45] and lately in adults fish as well [1]. The obtained findings from these studies indicate that, at least for kainate, adult zebrafish exhibit seizures that closely resemble those reported in rodents. The neurohypophyseal peptide isotocin (IT), which differs from its corresponding OT analog for the replacement of two amino acids (serine and isoleucine) and vasotocin (AVT) which differs from its corresponding AVP analog for the replacement of an amino acid Abbreviations: V1A, vasopressin receptor type 1; OT, oxytocin; IT, isotocin; AVP, vasopressin; AVT, vasotocin; PTZ, pentylenetetrazole; VLP, valproate sodium salt; HPA, hypothalamic–pituitary–adrenal axis. ∗ Corresponding author at: Dipartimento di Biotecnologie mediche e Medicina traslazionale, Università degli Studi di Milano, Via Vanvitelli 32, Milan, Italy. Tel.: +39 02 50317042; fax: +39 02 50317036. E-mail address: [email protected] (M. Sala). (phenylalanine), influence a wide range of physiological and behav- ioral responses, including aspects of sexual and social behaviors aspects, osmoregulation, stress response, metabolism, blood pres- sure, and circadian rhythms [12]. Very little is known about IT and AVT on a potential protective role in zebrafish. Instead, the non- apeptide hormone oxytocin (OT) and arginine-vasopressin (AVP) are known to be implicated in some pathological conditions and stress in mammals. Exogenous OT administration in rats alleviated tissue damage in several animal models of ischemia–reperfusion injury in liver [10], kidney [46] and heart [20] and against sepsis- induced oxidative damage [23]. In vitro, OT protected cellular viability of immature hippocampal cultures subjected to oxygen- glucose deprivation [7] and reduced the infarct size on in vitro myocardial injury [34]. Synthesis and release of both peptides are implicated in the physiological response to different stressors such as osmotic stim- ulation [25], acute or chronic restraint-stress [13,33] or forced swimming [14]. The occurrence of seizures is also regarded as a severe stres- sor. Recently, a high susceptibility to PTZ-induced tonic–clonic seizures has been found in mice mutant for OT receptor, which 0196-9781/$ – see front matter © 2012 Elsevier Inc. All rights reserved. http://dx.doi.org/10.1016/j.peptides.2012.07.013
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Neurohypophyseal hormones protect against pentylenetetrazole-induced seizures in zebrafish: Role of...

P
Ns
DMa
b
a
ARRAA
KEANHT
1
gv[fe
i(c
vH
tT
0h
ARTICLE IN PRESSG ModelEP-68749; No. of Pages 7
Peptides xxx (2012) xxx–xxx
Contents lists available at SciVerse ScienceDirect
Peptides
j ourna l ho me pa ge: www.elsev ier .com/ locate /pept ides
eurohypophyseal hormones protect against pentylenetetrazole-inducedeizures in zebrafish: Role of oxytocin-like and V1a-like receptor
aniela Braidaa, Andrea Donzelli a, Roberta Martuccia, Luisa Ponzonia, Alberto Pauletti a,ariaelvina Salaa,b,∗
Dipartimento di Biotecnologie mediche e Medicina traslazionale, Università degli Studi di Milano, Via Vanvitelli 32, Milan, ItalyCNR, Institute of Neuroscience, Via Vanvitelli 32, Milan, Italy
r t i c l e i n f o
rticle history:eceived 26 June 2012eceived in revised form 16 July 2012ccepted 16 July 2012vailable online xxx
eywords:pilepsynticonvulsanteurohypophysealormoneeleost
a b s t r a c t
Oxytocin (OT) and arginine-vasopressin (AVP) are involved in the physiological response to differentstressors like the occurrence of seizures which is regarded as a severe stress factor. Zebrafish (Danio rerio)is recently featured as a model of epilepsy but the role of neurohypophyseal hormones on this teleostis still unknown. We attempted to determine whether non-mammalian homologues like isotocin (IT)and vasotocin (AVT) affected pentylenetetrazole (PTZ)-induced seizures in adult zebrafish in comparisonwith OT/AVP. The mechanism was studied using the most selective OT and AVP receptor antagonists.Zebrafish were injected i.m. with increasing doses (0.1–40 ng/kg) of the neuropeptides 10 min beforePTZ exposure. DesGly-NH2-d(CH2)5-[D-Tyr2,Thr4]OVT (desglyDTyrOVT) for OT receptor and SR49059for V1a subtype receptor, were injected together with each agonist 20 min before PTZ exposure. All thepeptides significantly decreased the number of seizures, increased the mean latency time to the firstseizure and decreased lethality. This protective effect led to a dose–response curve following a U-shaped
form. IT was approximately 40 times more active than OT while AVT was 20 times more potent thanAVP in reducing the number of seizures. DesglyDTyrOVT was more effective in antagonizing OT/IT, whileSR49059 mainly blocked AVP/AVT-induced protection against PTZ-induced seizures. The present findingsprovide direct evidence of an important involvement of IT/OT and AVP/AVT as anticonvulsant agentsagainst PTZ-induced seizures with a receptor-mediated mechanism in zebrafish. These data reinforcezebrafish as an emerging experimental model to study and identify new antiepileptic drugs.. Introduction
Zebrafish (Danio rerio) have been recently proposed as a newenetic model to evaluate epilepsy [21,39]. Using different con-ulsive drugs, seizures were observed in embryos [43,44], larvae4,41,45] and lately in adults fish as well [1]. The obtained findingsrom these studies indicate that, at least for kainate, adult zebrafishxhibit seizures that closely resemble those reported in rodents.
The neurohypophyseal peptide isotocin (IT), which differs from
Please cite this article in press as: Braida D, et al. Neurohypophyseal hzebrafish: Role of oxytocin-like and V1a-like receptor. Peptides (2012
ts corresponding OT analog for the replacement of two amino acidsserine and isoleucine) and vasotocin (AVT) which differs from itsorresponding AVP analog for the replacement of an amino acid
Abbreviations: V1A, vasopressin receptor type 1; OT, oxytocin; IT, isotocin; AVP,asopressin; AVT, vasotocin; PTZ, pentylenetetrazole; VLP, valproate sodium salt;PA, hypothalamic–pituitary–adrenal axis.∗ Corresponding author at: Dipartimento di Biotecnologie mediche e Medicina
raslazionale, Università degli Studi di Milano, Via Vanvitelli 32, Milan, Italy.el.: +39 02 50317042; fax: +39 02 50317036.
E-mail address: [email protected] (M. Sala).
196-9781/$ – see front matter © 2012 Elsevier Inc. All rights reserved.ttp://dx.doi.org/10.1016/j.peptides.2012.07.013
© 2012 Elsevier Inc. All rights reserved.
(phenylalanine), influence a wide range of physiological and behav-ioral responses, including aspects of sexual and social behaviorsaspects, osmoregulation, stress response, metabolism, blood pres-sure, and circadian rhythms [12]. Very little is known about IT andAVT on a potential protective role in zebrafish. Instead, the non-apeptide hormone oxytocin (OT) and arginine-vasopressin (AVP)are known to be implicated in some pathological conditions andstress in mammals. Exogenous OT administration in rats alleviatedtissue damage in several animal models of ischemia–reperfusioninjury in liver [10], kidney [46] and heart [20] and against sepsis-induced oxidative damage [23]. In vitro, OT protected cellularviability of immature hippocampal cultures subjected to oxygen-glucose deprivation [7] and reduced the infarct size on in vitromyocardial injury [34].
Synthesis and release of both peptides are implicated in thephysiological response to different stressors such as osmotic stim-ulation [25], acute or chronic restraint-stress [13,33] or forced
ormones protect against pentylenetetrazole-induced seizures in), http://dx.doi.org/10.1016/j.peptides.2012.07.013
swimming [14].The occurrence of seizures is also regarded as a severe stres-
sor. Recently, a high susceptibility to PTZ-induced tonic–clonicseizures has been found in mice mutant for OT receptor, which

IN PRESSG ModelP
2 tides xxx (2012) xxx–xxx
w[ttpnic(vsipAicsaos
smtspomsTettsb
2
2
ns6eMwBt(FfiFfioet
2
[pap
Table 1Score quantification on the basis of zebrafish behavior after PTZ treatment accordingto Alfaro et al. [1].
Stage Swimming behavior
I Immobility and hyperventilation of the animalII Whirlpool-like swimming behaviorIII Rapid movements from right to leftIV Abnormal and spasmodic muscular contractionsV Rapid whole-body clonus-like convulsionsVI Sinking to the bottom of the tank and spasms for several minutes
ARTICLEEP-68749; No. of Pages 7
D. Braida et al. / Pep
as completely rescued by OT centrally administered treatment36]. Such effect was blocked by a selective V1a subtype recep-or SR49059 pre-treatment suggesting an OT protective effect byhis receptor. Surprisingly, there are divergent data about AVP/AVTro/anticonvulsant effect in rodents. AVP (1–3 �g) given subcuta-eously had potentiating effects on pilocarpine-induced seizures
n rats [9] while when given i.c.v. (0.01–1000 ng/kg) did not signifi-antly affect the seizures parameters. The V2 receptor antagonist[adamantaneacetyl1, O-Et-D-tyr2, Val4, aminobutyryl6, Arg8,9]-asopressin) prevented hyperthermic seizures but did not affecteizure latency in rat pups [18] while AVP given alone s.c. or i.c.v.n rats, did not produce any sign of seizure in centrally-inducedilocarpine seizures [17]. In contrast, Eagan et al. [11] found thatVP i.c.v. (10 ng) in rats induced absence-like seizure while AVT
ncreased the latency to convulsions [24]. Notably, plasma AVPoncentrations have been found to be markedly increased aftereizures in patients with idiopathic and pos-traumatic epilepticttacks [2]. Accordingly, Wistar audiogenic rats, a genetic modelf reflex epilepsy with high intensity sound stimulation-inducedeizures, showed a marked increase of AVP plasma levels [16].
Therefore, on the basis of the ability of OT to rescue PTZ-inducedeizures in mice [18] and since the role of neurohypophyseal hor-ones on zebrafish epilepsy models is still unknown, we attempted
o determine whether these neurohormones were able to rescueeizures in adult zebrafish. Thus, the current study investigated theotential protective effect of IT and AVT compared to OT and AVP,n PTZ-induced seizures. Additionally, to evaluate their receptor-ediated mechanism of action, we identified and used highly
elective antagonists assayed on human OT/AVP receptor subtypes.o reach this goal, adult zebrafish were pre-treated with differ-nt doses of hormones given intramuscularly and then exposedo the pro-convulsive drug PTZ, dissolved in a tank. Antiepilep-ic drug valproate sodium salt (VLP) was used as reference. Theeizure-like behavior was scored according to a scale elaboratedy [1].
. Materials and methods
.1. Subjects
Adult wild type (WT) zebrafish (D. rerio) (0.4–1 g) of heteroge-eous genetic background were obtained from a local aquariumupply store (Aquarium Center, Milan, Italy). Zebrafish were almost–12 months of age, and were about 3 and 4 cm in length. In allxperiments the sex ratio of zebrafish was approximately 50–50%.ales and females were identified according to [40]. The animalsere kept at approximately 28.5 ◦C on a 14-h light/10-h dark cycle.ehavioral testing took place during the light phase from 9.00o 14.00 h. Tank water consisted of deionized H2O and sea salts0.6 g/10 l of water; Instant Ocean, Aquarium Systems, Sarrebourg,rance). The home tanks with groups of approximately 30 adultsh were maintained with both constant filtration and aeration.ish were fed daily with brine shrimp and flake fish food (Tropicalsh food, Consorzio G5, Italy). All the drug naive fish were used oncenly. The experimental protocol was approved by the Italian Gov-rnmental Decree No. 29/2010. All efforts were made to minimizehe number of animals used and their discomfort.
.2. Treatment
Zebrafish body weight was measured as previously described
Please cite this article in press as: Braida D, et al. Neurohypophyseal hzebrafish: Role of oxytocin-like and V1a-like receptor. Peptides (2012
5,6]. Briefly, fish were removed from their tank using a net andlaced in a container with tank water, positioned on a digital bal-nce. Fish weight was determined as the weight of the containerlus the fish minus the weight of the container before the fish
VII Death
was added. The mean of three measurements was recorded. Forintramuscular administration each fish was injected in the caudalmusculature along the posterior axis. The site of injection was con-stantly kept in the area below the caudal fin on the left side ofeach fish. Each volume, depending on the fish’s weight (2 �l/g), wasgiven using a Hamilton syringe (Hamilton Bonaduz AG, Bonaduz,Switzerland). The fish were injected inside the water tank using anet. Each fish was kept immobilized in the net with two fingersof the left hand. Immediately after, the needle was positioned ata 45◦ angle in relation to the fish back with the needle pointingtowards the head. The injection was in the largest portion of thecaudal muscle, immediately posterior to the caudal fin. The needlewas inserted over the bevel into the muscle. Total time spent outof water was approximately 10 s. Each fish was removed from thenet and immediately dipped into the tank.
2.3. PTZ-induced seizures
To evoke experimental seizures, the animals were individu-ally exposed to PTZ for 20 min according to Baraban et al. in a2-l transparent observation tank (20 cm long × 10 cm wide × 15 cmtall) containing home tank water filled up to 12 cm [4]. Controlfish were exposed to equivalent volumes of water, taken from theaquarium. Then animals were moved to the observation tanks filledwith water tank only and their behaviors were videotaped by ahigh-resolution Canon MV900 camera. The degree of seizures activ-ity was monitored by trained blind observers for 6 min accordingto Alfaro et al. and the score quantified using the scale reported inTable 1 [1]. In addition, the number of seizures, the latency (s) tothe first one, and lethality were evaluated.
2.4. Drugs
Oxytocin (OT), arginine-vasopressin (AVP), isotocin ((Ser4,Ile8)-oxytocin) (IT), and vasotocin ((Arg8)-vasotocin) (AVT) (Bachem,Germany) were dissolved in saline (0.9%) and injected i.m. Therange of doses and the route of administration were the same usedin our previous work where the peptides injected intramuscularlyshowed an effective outcome on social behavior and fear responseto predator test [5]. To investigate the mechanism of action weused desglyDTyrOVT, the most selective OT receptor antagonistreported to date in rats (100-fold more selective in terms of affinityversus OT than V1a receptor) [38] and SR49059 the well known V1areceptor antagonist [42]. DesGly-NH2-d(CH2)5-[D-Tyr2,Thr4]OVT(desglyDTyrOVT) was kindly supplied by M. Manning (Toledo,USA) and ((2S)-1-[[(2R,3S)-5-chloro-3(2-chlorophenyl)-1-[(3,4-dimethoxyphenyl)sulfonyl]-2,3-dihydro-3-hydroxy-1H-indol-2-l]carbonyl]-2-pyrrolidin carboxamide) (SR 49059) was purchasedby Sigma–Aldrich, St Louis, USA [38]. All the antagonists were
ormones protect against pentylenetetrazole-induced seizures in), http://dx.doi.org/10.1016/j.peptides.2012.07.013
injected intramuscularly 10 min before each agonist. The dose ofeach antagonist was chosen on the basis of its ability to antagonizethe reduced fear response to predator induced by the neuropep-tides [5]. PTZ (Sigma–Aldrich, St Louis, USA) was dissolved in
ARTICLE IN PRESSG ModelPEP-68749; No. of Pages 7
D. Braida et al. / Peptides xxx (2012) xxx–xxx 3
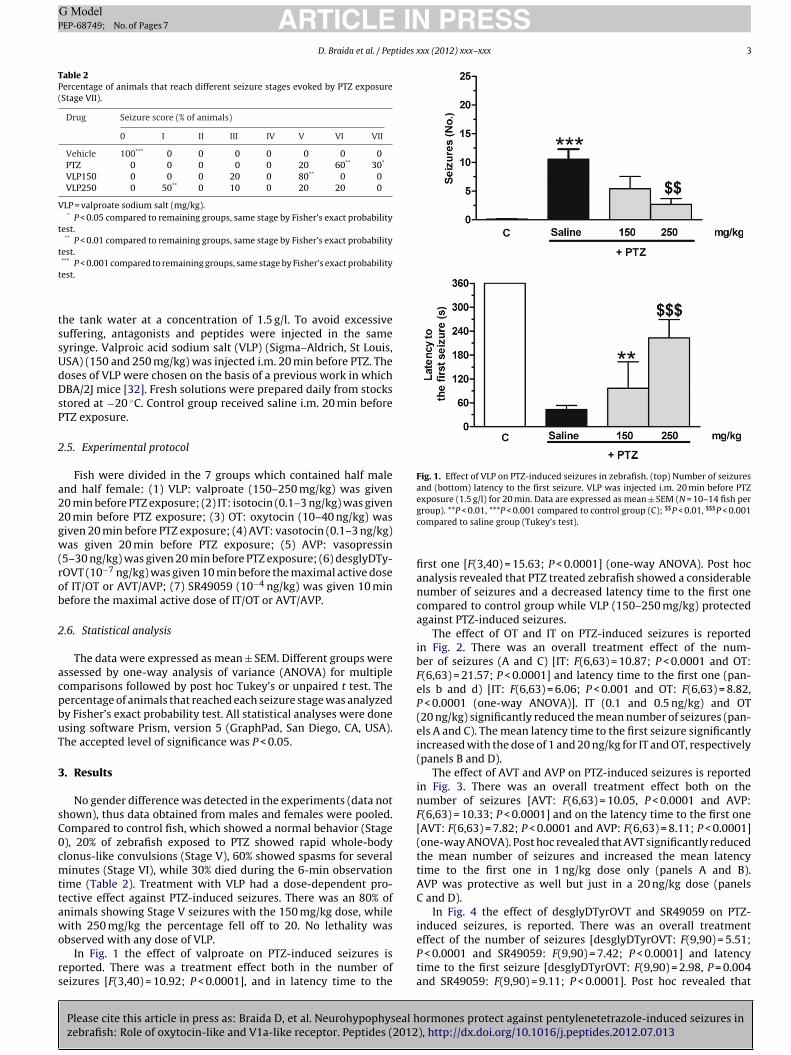
Table 2Percentage of animals that reach different seizure stages evoked by PTZ exposure(Stage VII).
Drug Seizure score (% of animals)
0 I II III IV V VI VII
Vehicle 100*** 0 0 0 0 0 0 0PTZ 0 0 0 0 0 20 60** 30*
VLP150 0 0 0 20 0 80** 0 0VLP250 0 50** 0 10 0 20 20 0
VLP = valproate sodium salt (mg/kg).* P < 0.05 compared to remaining groups, same stage by Fisher’s exact probability
test.** P < 0.01 compared to remaining groups, same stage by Fisher’s exact probability
test.***
t
tssUdDsP
2
a22gw(rob
2
acpbuT
3
sC0cmttawo
rs
Fig. 1. Effect of VLP on PTZ-induced seizures in zebrafish. (top) Number of seizuresand (bottom) latency to the first seizure. VLP was injected i.m. 20 min before PTZexposure (1.5 g/l) for 20 min. Data are expressed as mean ± SEM (N = 10–14 fish per
P < 0.001 compared to remaining groups, same stage by Fisher’s exact probabilityest.
he tank water at a concentration of 1.5 g/l. To avoid excessiveuffering, antagonists and peptides were injected in the sameyringe. Valproic acid sodium salt (VLP) (Sigma–Aldrich, St Louis,SA) (150 and 250 mg/kg) was injected i.m. 20 min before PTZ. Theoses of VLP were chosen on the basis of a previous work in whichBA/2J mice [32]. Fresh solutions were prepared daily from stocks
tored at −20 ◦C. Control group received saline i.m. 20 min beforeTZ exposure.
.5. Experimental protocol
Fish were divided in the 7 groups which contained half malend half female: (1) VLP: valproate (150–250 mg/kg) was given0 min before PTZ exposure; (2) IT: isotocin (0.1–3 ng/kg) was given0 min before PTZ exposure; (3) OT: oxytocin (10–40 ng/kg) wasiven 20 min before PTZ exposure; (4) AVT: vasotocin (0.1–3 ng/kg)as given 20 min before PTZ exposure; (5) AVP: vasopressin
5–30 ng/kg) was given 20 min before PTZ exposure; (6) desglyDTy-OVT (10−7 ng/kg) was given 10 min before the maximal active dosef IT/OT or AVT/AVP; (7) SR49059 (10−4 ng/kg) was given 10 minefore the maximal active dose of IT/OT or AVT/AVP.
.6. Statistical analysis
The data were expressed as mean ± SEM. Different groups weressessed by one-way analysis of variance (ANOVA) for multipleomparisons followed by post hoc Tukey’s or unpaired t test. Theercentage of animals that reached each seizure stage was analyzedy Fisher’s exact probability test. All statistical analyses were donesing software Prism, version 5 (GraphPad, San Diego, CA, USA).he accepted level of significance was P < 0.05.
. Results
No gender difference was detected in the experiments (data nothown), thus data obtained from males and females were pooled.ompared to control fish, which showed a normal behavior (Stage), 20% of zebrafish exposed to PTZ showed rapid whole-bodylonus-like convulsions (Stage V), 60% showed spasms for severalinutes (Stage VI), while 30% died during the 6-min observation
ime (Table 2). Treatment with VLP had a dose-dependent pro-ective effect against PTZ-induced seizures. There was an 80% ofnimals showing Stage V seizures with the 150 mg/kg dose, whileith 250 mg/kg the percentage fell off to 20. No lethality was
Please cite this article in press as: Braida D, et al. Neurohypophyseal hzebrafish: Role of oxytocin-like and V1a-like receptor. Peptides (2012
bserved with any dose of VLP.In Fig. 1 the effect of valproate on PTZ-induced seizures is
eported. There was a treatment effect both in the number ofeizures [F(3,40) = 10.92; P < 0.0001], and in latency time to the
group). **P < 0.01, ***P < 0.001 compared to control group (C); $$P < 0.01, $$$P < 0.001compared to saline group (Tukey’s test).
first one [F(3,40) = 15.63; P < 0.0001] (one-way ANOVA). Post hocanalysis revealed that PTZ treated zebrafish showed a considerablenumber of seizures and a decreased latency time to the first onecompared to control group while VLP (150–250 mg/kg) protectedagainst PTZ-induced seizures.
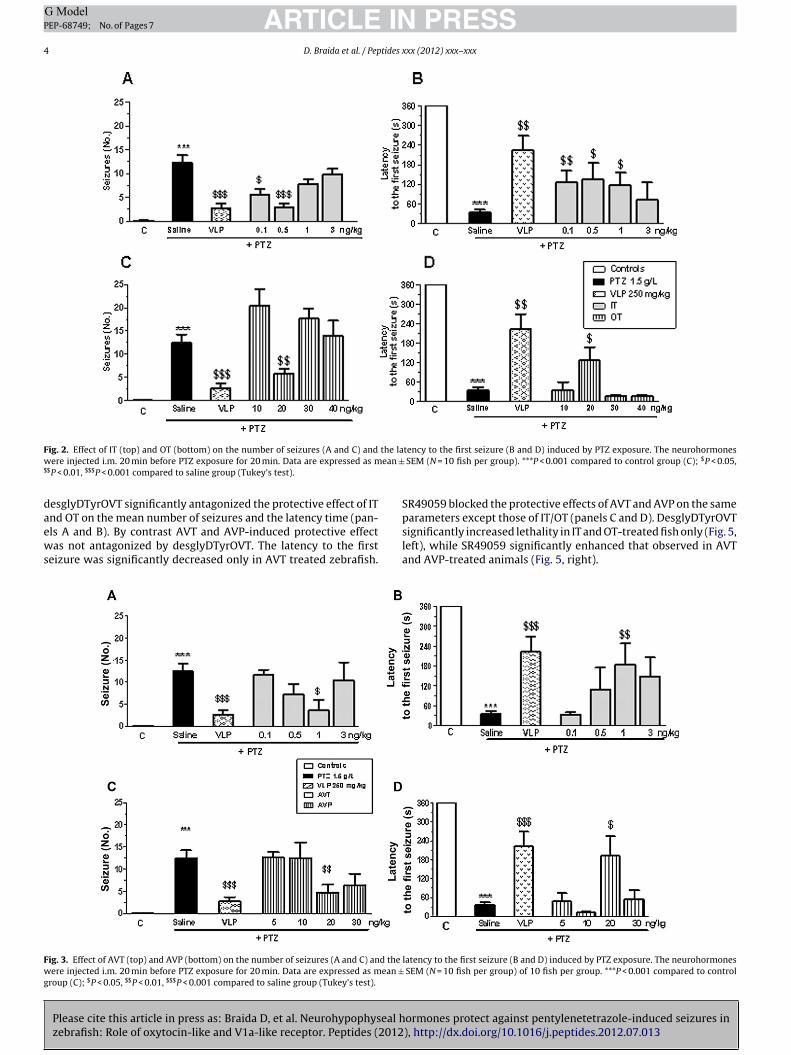
The effect of OT and IT on PTZ-induced seizures is reportedin Fig. 2. There was an overall treatment effect of the num-ber of seizures (A and C) [IT: F(6,63) = 10.87; P < 0.0001 and OT:F(6,63) = 21.57; P < 0.0001] and latency time to the first one (pan-els b and d) [IT: F(6,63) = 6.06; P < 0.001 and OT: F(6,63) = 8.82,P < 0.0001 (one-way ANOVA)]. IT (0.1 and 0.5 ng/kg) and OT(20 ng/kg) significantly reduced the mean number of seizures (pan-els A and C). The mean latency time to the first seizure significantlyincreased with the dose of 1 and 20 ng/kg for IT and OT, respectively(panels B and D).
The effect of AVT and AVP on PTZ-induced seizures is reportedin Fig. 3. There was an overall treatment effect both on thenumber of seizures [AVT: F(6,63) = 10.05, P < 0.0001 and AVP:F(6,63) = 10.33; P < 0.0001] and on the latency time to the first one[AVT: F(6,63) = 7.82; P < 0.0001 and AVP: F(6,63) = 8.11; P < 0.0001](one-way ANOVA). Post hoc revealed that AVT significantly reducedthe mean number of seizures and increased the mean latencytime to the first one in 1 ng/kg dose only (panels A and B).AVP was protective as well but just in a 20 ng/kg dose (panelsC and D).
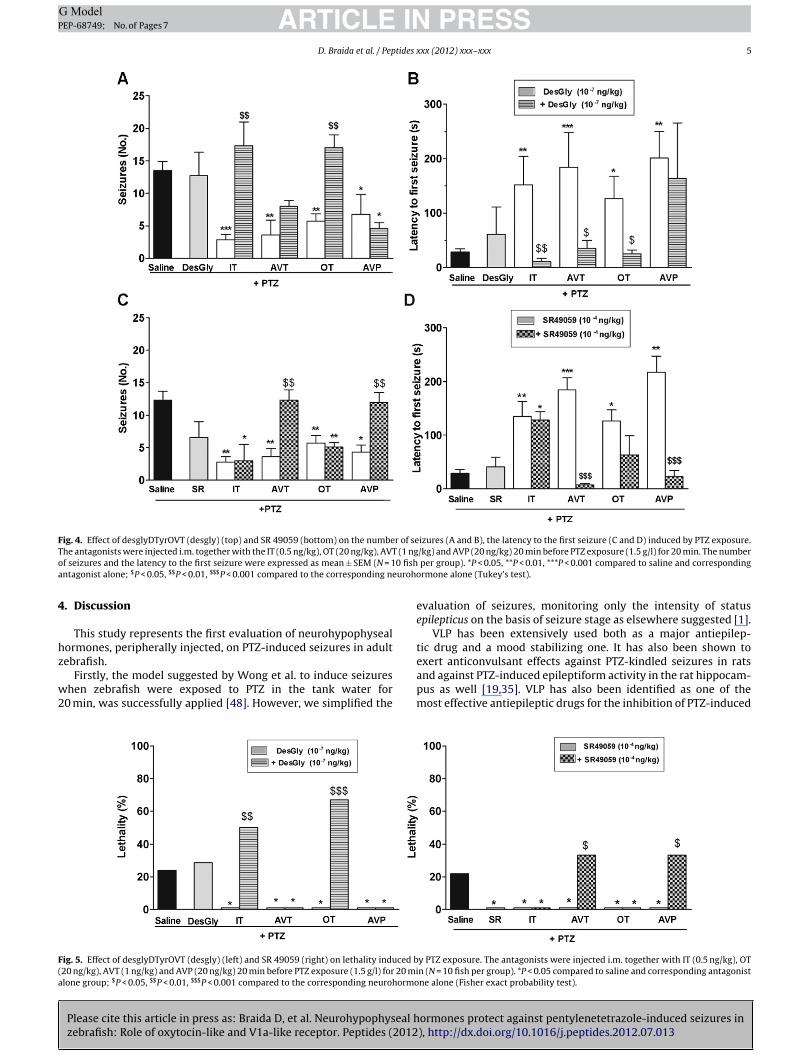
In Fig. 4 the effect of desglyDTyrOVT and SR49059 on PTZ-induced seizures, is reported. There was an overall treatmenteffect of the number of seizures [desglyDTyrOVT: F(9,90) = 5.51;
ormones protect against pentylenetetrazole-induced seizures in), http://dx.doi.org/10.1016/j.peptides.2012.07.013
P < 0.0001 and SR49059: F(9,90) = 7.42; P < 0.0001] and latencytime to the first seizure [desglyDTyrOVT: F(9,90) = 2.98, P = 0.004and SR49059: F(9,90) = 9.11; P < 0.0001]. Post hoc revealed that
ARTICLE IN PRESSG ModelPEP-68749; No. of Pages 7
4 D. Braida et al. / Peptides xxx (2012) xxx–xxx
F the law ean ±$
daews
Fwg
ig. 2. Effect of IT (top) and OT (bottom) on the number of seizures (A and C) andere injected i.m. 20 min before PTZ exposure for 20 min. Data are expressed as m
$P < 0.01, $$$P < 0.001 compared to saline group (Tukey’s test).
esglyDTyrOVT significantly antagonized the protective effect of IT
Please cite this article in press as: Braida D, et al. Neurohypophyseal hzebrafish: Role of oxytocin-like and V1a-like receptor. Peptides (2012
nd OT on the mean number of seizures and the latency time (pan-ls A and B). By contrast AVT and AVP-induced protective effectas not antagonized by desglyDTyrOVT. The latency to the first
eizure was significantly decreased only in AVT treated zebrafish.
ig. 3. Effect of AVT (top) and AVP (bottom) on the number of seizures (A and C) and the
ere injected i.m. 20 min before PTZ exposure for 20 min. Data are expressed as mean ±roup (C); $P < 0.05, $$P < 0.01, $$$P < 0.001 compared to saline group (Tukey’s test).
tency to the first seizure (B and D) induced by PTZ exposure. The neurohormones SEM (N = 10 fish per group). ***P < 0.001 compared to control group (C); $P < 0.05,
SR49059 blocked the protective effects of AVT and AVP on the same
ormones protect against pentylenetetrazole-induced seizures in), http://dx.doi.org/10.1016/j.peptides.2012.07.013
parameters except those of IT/OT (panels C and D). DesglyDTyrOVTsignificantly increased lethality in IT and OT-treated fish only (Fig. 5,left), while SR49059 significantly enhanced that observed in AVTand AVP-treated animals (Fig. 5, right).
latency to the first seizure (B and D) induced by PTZ exposure. The neurohormones SEM (N = 10 fish per group) of 10 fish per group. ***P < 0.001 compared to control
ARTICLE IN PRESSG ModelPEP-68749; No. of Pages 7
D. Braida et al. / Peptides xxx (2012) xxx–xxx 5
Fig. 4. Effect of desglyDTyrOVT (desgly) (top) and SR 49059 (bottom) on the number of seizures (A and B), the latency to the first seizure (C and D) induced by PTZ exposure.T (1 ngo 0 fisha uroho
4
hz
w2
F(a
he antagonists were injected i.m. together with the IT (0.5 ng/kg), OT (20 ng/kg), AVTf seizures and the latency to the first seizure were expressed as mean ± SEM (N = 1ntagonist alone; $P < 0.05, $$P < 0.01, $$$P < 0.001 compared to the corresponding ne
. Discussion
This study represents the first evaluation of neurohypophysealormones, peripherally injected, on PTZ-induced seizures in adult
Please cite this article in press as: Braida D, et al. Neurohypophyseal hzebrafish: Role of oxytocin-like and V1a-like receptor. Peptides (2012
ebrafish.Firstly, the model suggested by Wong et al. to induce seizures
hen zebrafish were exposed to PTZ in the tank water for0 min, was successfully applied [48]. However, we simplified the
ig. 5. Effect of desglyDTyrOVT (desgly) (left) and SR 49059 (right) on lethality induced b20 ng/kg), AVT (1 ng/kg) and AVP (20 ng/kg) 20 min before PTZ exposure (1.5 g/l) for 20 milone group; $P < 0.05, $$P < 0.01, $$$P < 0.001 compared to the corresponding neurohormo
/kg) and AVP (20 ng/kg) 20 min before PTZ exposure (1.5 g/l) for 20 min. The number per group). *P < 0.05, **P < 0.01, ***P < 0.001 compared to saline and correspondingrmone alone (Tukey’s test).
evaluation of seizures, monitoring only the intensity of statusepilepticus on the basis of seizure stage as elsewhere suggested [1].
VLP has been extensively used both as a major antiepilep-tic drug and a mood stabilizing one. It has also been shown to
ormones protect against pentylenetetrazole-induced seizures in), http://dx.doi.org/10.1016/j.peptides.2012.07.013
exert anticonvulsant effects against PTZ-kindled seizures in ratsand against PTZ-induced epileptiform activity in the rat hippocam-pus as well [19,35]. VLP has also been identified as one of themost effective antiepileptic drugs for the inhibition of PTZ-induced
y PTZ exposure. The antagonists were injected i.m. together with IT (0.5 ng/kg), OTn (N = 10 fish per group). *P < 0.05 compared to saline and corresponding antagonistne alone (Fisher exact probability test).

ING ModelP
6 tides
ezctpiafi(f
sfitot
mdt2
aalatssa
ci
mmewi
odtpSsfhbb
oaeriEfiar
iPAlt
[
[
[
[
[
[
[
[
[
[
ARTICLEEP-68749; No. of Pages 7
D. Braida et al. / Pep
pileptiform activity in the exposed brains of agar-embeddedebrafish larvae, reducing epileptiform discharge amplitude in aoncentration dependent manner [4]. Additionally, in the posi-ive control experiment carried out to validate our model, VLPre-treatment, dose-dependently reduced convulsive-like behav-
or decreasing the mean number of seizures induced by PTZnd increasing the mean latency time to the first seizure. Ourndings agree with those of Lee et al. where, however, VLP200 �M) was not injected but dissolved in aquarium wateror 1 h [26].
Secondly, we demonstrated for the first time the anticonvul-ive effect of neurohypophyseal hormones OT/AVP and their teleostsh homologues IT/AVT against PTZ-induced seizures. In detail,he four neuropeptides significantly decreased both the numberf seizures and lethality, and increased the mean latency time tohe first seizure.
Our findings on OT anticonvulsive effect appear in disagree-ent with those of Loyens et al. in mice where OT (0.5 mg/kg)
ecreased seizure thresholds after PTZ injection [29]. However,he ratio between our dose (20 ng/kg) and that of the author, was5,000 times.
IT and AVT elicited 70 and 77% of seizure inhibition, respectively,t doses lower (1–3 ng/kg), than OT and AVP (20–30 ng/kg). Both OTnd AVP induced protective effect against PTZ-induced seizures at aower efficacy (54 and 61%, respectively). Furthermore, IT resultedpproximately 40 times more active than OT while AVT was 20imes more potent than AVP in reducing the number of seizures. Aimilar range of doses was found in a previous study where a greatelectivity of teleost specific neurohormones on the prosocial andnxiolytic effect, was demonstrated [5].
AVT has been previously shown to protect against PTZ-inducedonvulsions in rats by [27]. Accordingly, AVP reduced the seizurencidence in spontaneously seizing Mongolian gerbils.
Even if is still unclear, an inverted U dose–response is quite com-on for neurohypophyseal hormones, where a low dose appearsore effective than a high. OT has been shown to exhibit a biphasic
ffect both on social behavior and anxiety in teleosts [5,42]. AVTas shown to follow a similar dose–response curve for aggression
n fish [37].To verify if the protective effect of neurohypophyseal hormones
n PTZ-induced seizures was specifically receptor-mediated, weecided to use peptidic analogs with high receptor selectivity forhe human OT/AVP receptors [5]. In particular, we employed thereviously characterized highly selective V1a receptor antagonistR49059 [38] and a peptidic analog previously reported as highlyelective in rats, desglyDTyrOVT [31]. This antagonist was beenound extremely selective on human receptors, too. It binds withigh affinity the OT receptors only. In addition, this antagonist haseen found to inhibit the pro-social and anxiolytic effect inducedy OT/IT and AVT/AVP in zebrafish.
The characterization of IT and AVT receptors was reported forther teleosts [3,8,30,47]. One IT and one AVT receptor were clonednd pharmacologically investigated [22,28,30]. Recently, one genencoding for an AVT receptor (avplr1b arginine-vasopressin-likeeceptor 1beta, corresponding to a V1a subtype), and one encod-ng for an IT receptor (Oxtl) [15,28] have been shown in zebrafish.ven if a complete pharmacological characterization of teleostsh receptors is still lacking, AVT and IT receptors acted through
PLC/IP3 intracellular signaling pathway as the mammalian OTeceptor/V1a/V1b receptor subtypes [47].
Our findings indicated that desglyDTyrOVT was more effectiven antagonizing OT/IT than AVP/AVT-induced protection against
Please cite this article in press as: Braida D, et al. Neurohypophyseal hzebrafish: Role of oxytocin-like and V1a-like receptor. Peptides (2012
TZ-induced seizures. On the other hand, SR49059 mainly blockedVP/AVT-induced protection. Only high doses of these antagonists
ost their selectivity and blocked all the peptides-induced protec-ion (data not shown). These data support a maintained selectivity
[
PRESSxxx (2012) xxx–xxx
of both desglyDTTyrOVT and SR49059 for an OT-like and a V1a-likereceptor, respectively, in zebrafish.
In conclusion, our findings provide direct evidence of animportant involvement of neurohypophyseal hormones as anti-convulsant agents against PTZ-induced seizures with a receptor-mediated mechanism in zebrafish. These data reinforce zebrafishas an emerging experimental model to study and identify newantiepileptic drugs.
Conflict of interest
Authors declare no conflict of interest.
Acknowledgments
The authors thank Bice Chini, CNR Institute, Milan for her gen-erous contribution of desglyDTyrOVT and SR49059 used in thisexperiment.
References
[1] Alfaro JM, Ripoll-Gómez J, Burgos JS. Kainate administered to adultzebrafish causes seizures similar to those in rodent models. Eur J Neurosci2011;33:1252–5.
[2] Aminoff MJ, Davis SL, Panitch HS. Serial evoked potential studies inpatients with definite multiple sclerosis. Clinical relevance. Arch Neurol1984;41:1197–202.
[3] An KW, Kim NN, Choi CY. Cloning and expression of aquaporin 1 and argininevasotocin receptor mRNA from the black porgy, Acanthopagrus schlegeli: effectof freshwater acclimation. Fish Physiol Biochem 2008;34:185–94.
[4] Baraban SC, Taylor MR, Castro PA, Baier H. Pentylenetetrazole induced changesin zebrafish behavior, neural activity and c-fos expression. Neuroscience2005;131:759–68.
[5] Braida D, Donzelli A, Martucci R, Capurro V, Busnelli M, Chini B, et al. Neu-rohypophyseal hormones manipulation modulate social and anxiety-relatedbehavior in zebrafish. Psychopharmacology 2012;220:319–30.
[6] Braida D, Limonta V, Pegorini S, Zani A, Guerini-Rocco C, Gori E, et al.Hallucinatory and rewarding effect of salvinorin A in zebrafish: kappa-opioid and CB1-cannabinoid receptor involvement. Psychopharmacology2007;190:441–8.
[7] Ceanga M, Spataru A, Zagrean AM. Oxytocin is neuroprotective against oxygen-glucose deprivation and reoxygenation in immature hippocampal cultures.Neurosci Lett 2010;477:15–8.
[8] Conklin DJ, Smith MP, Olson KR. Pharmacological characterization of argininevasotocin vascular smooth muscle receptors in the trout (Oncorhynchus mykiss)in vitro. Gen Comp Endocrinol 1999;114:36–46.
[9] Croiset G, De Wied D. Proconvulsive effect of vasopressin; mediation bya putative V2 receptor subtype in the central nervous system. Brain Res1997;759:18–23.
10] Düs ünceli F, Is eri SÖ, Ercan F, Gedik N, Yegen C, Yegen BC. Oxytocin alleviateshepatic ischemia–reperfusion injury in rats. Peptides 2008;29:1216–22.
11] Eagan PC, Veale WL, Cooper KE. Evidence for the antipyretic action of argininevasopressin the rat. Proc Can Fed Biol Soc 1980;23:40A.
12] Eaton JL, Holmqvist B, Glasgow E. Ontogeny of vasotocin-expressing cells inzebrafish: selective requirement for the transcriptional regulators orthopediaand single-minded 1 in the preoptic area. Dev Dyn 2008;237:995–1005.
13] Engelmann M, Ebner K, Landgraf R, Wotjak CT. Effects of Morris water mazetesting on the neuroendocrine stress response and intrahypothalamic releaseof vasopressin and oxytocin in the rat. Horm Behav 2006;50:496–501.
14] Engelmann M, Ludwig M. The activity of the hypothalamo-neurohypophysialsystem in response to acute stressor exposure: neuroendocrine and electro-physiological observations. Stress 2004;7:91–6.
15] Filby AL, Paull GC, Hickmore TF, Tyler CR. Unreavelling the neurophysiologicalbasis of aggression in a fish model. BMC Genomics 2010;11:498–515.
16] Fonseca AG, Santos RA, Moraes MF, Leite MF, Doretto MC. Vasopressinergichypothalamic neurons are recruited during the audiogenic seizure of WARs.Brain Res 2005;1038:32–40.
17] Gulec G, Isbil-Buyukcoskun N. Arginine vasopressin does not contribute toseizures induced by intracerebroventricularly-injected pilocarpine. Neuropep-tides 2005;39:575–9.
18] Gulec G, Noyan B. Arginine vasopressin in the pathogenesis of febrile convulsionand temporal lobe epilepsy. Neuroreport 2002;13:2045–8.
19] Gupta YK, Malhotra J. Effect of theophylline on diazepam and sodium valproate
ormones protect against pentylenetetrazole-induced seizures in), http://dx.doi.org/10.1016/j.peptides.2012.07.013
protection in pentylenetetrazole-kindled seizures in rats. Indian J Physiol Phar-macol 1997;41:280–4.
20] Hausmann H, Meyerhof W, Zwiers H, Lederis K, Richter D. Teleost isotocinreceptor: structure, functional expression, mRNA distribution and phylogeny.FEBS Lett 1995;370:227–30.

ING ModelP
tides
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[[
[
[
[
[
[
[
ARTICLEEP-68749; No. of Pages 7
D. Braida et al. / Pep
21] Hortopan GA, Dinday MT, Baraban SC. Spontaneous seizures and altered geneexpression in GABA signaling pathways in a mind bomb mutant zebrafish. JNeurosci 2010;30:13718–28.
22] Houshmand F, Faghihi M, Zahediasl S. Biphasic protective effect of oxy-tocin on cardiac ischemia/reperfusion injury in anaesthetized rats. Peptides2009;30:2301–8.
23] Is eri SO, Sener G, Saglam B, Gedik N, Ercan F, Yegen BC. Oxytocin protectsagainst sepsis-induced multiple organ damage: role of neutrophils. J Surg Res2005;126:73–81.
24] Kasting NW, Veale WL, Cooper KE. Vasotocin protects rats against convulsionsinduced by pentylenetetrazol. Experientia 1981;37:1001–2.
25] Kim JS, Kim WB, Kim YB, Lee Y, Kim YS, Shen FY, et al. Chronic hyperosmoticstress converts GABAergic inhibition into excitation in vasopressin and oxy-tocin neurons in the rat. J Neurosci 2011;31:13312–22.
26] Lee Y, Kim D, Kim YH, Lee H, Lee CJ. Improvement of pentylenetetrazol-induced learning deficits by valproic acid in the adult zebrafish. Eur J Pharmacol2010;643:225–31.
27] Lee RJ, Lomax P. The effect of spontaneous seizures on pentylenetetrazole andmaximum electroshock induced seizures in the Mongolian gerbil. Eur J Phar-macol 1984;106:91–6.
28] Lema SC. Identification of multiple vasotocin receptor cDNAs in teleost fish:sequences, phylogenetic analysis, sites of expression, and regulation in thehypothalamus and gill in response to hyperosmotic challenge. Mol CellEndocrinol 2010;321:215–30.
29] Loyens E, Vermoesen K, Schallier A, Michotte Y, Smolders I. Proconvulsiveeffects of oxytocin in the generalized pentylenetetrazol mouse model are medi-ated by vasopressin 1a receptors. Brain Res 2012;1436:43–50.
30] Mahlmann S, Meyerhof W, Hausmann H, Heierhorst J, Schönrock C, Zwiers H,et al. Structure, function, and phylogeny of [Arg8] vasotocin receptors fromteleost fish and toad. Proc Natl Acad Sci USA 1994;91:1342–5.
31] Manning M, Miteva K, Pancheva S, Stoev S, Wo NC, Chan WY. Design and syn-thesis of highly selective in vitro and in vivo uterine receptor antagonists ofoxytocin: comparisons with Atosiban. Int J Pept Protein Res 1995;46:244–52.
32] Marrosu F, Bortolato M, Frau R, Orrù M, Puligheddu M, Fà M, et al. Levetiracetamattenuates spontaneous spike-and-wave discharges in DBA/2J mice. EpilepsyRes 2007;75:224–7.
33] Ondrejcakova M, Bakos J, Garafova A, Kovacs L, Kvetnansky R, Jezova D.
Please cite this article in press as: Braida D, et al. Neurohypophyseal hzebrafish: Role of oxytocin-like and V1a-like receptor. Peptides (2012
Neuroendocrine and cardiovascular parameters during stimulation of stress-induced rise in circulating oxytocin in the rat. Stress 2010;13:324–32.
34] Ondrejcakova M, Ravingerova T, Bakos J, Pancza D, Jezova D. Oxytocin exertsprotective effects on in vitro myocardial injury induced by ischemia and reper-fusion. Can J Physiol Pharmacol 2009;87:137–42.
[
PRESSxxx (2012) xxx–xxx 7
35] Piredda S, Yonekawa W, Whittingham TS, Kupferberg HJ. Effects of antiepilep-tic drugs on pentylenetetrazole-induced epileptiform activity in the in vitrohippocampus. Epilepsia 1986;27:341–6.
36] Sala M, Braida D, Lentini D, Busnelli M, Bulgheroni E, Capurro V, et al. Phar-macologic rescue of impaired cognitive flexibility, social deficits, increasedaggression, and seizure susceptibility in oxytocin receptor null mice: a neu-robehavioral model of autism. Biol Psychiatry 2011;69:875–82.
37] Santangelo N, Bass AH. New insights into neuropeptide modulation of aggres-sion: field studies of arginine vasotocin in a territorial tropical damselfish. ProcBiol Sci 2006;273:3085–92.
38] Serradeil-Le Gal C, Raufaste D, Marty E, Garcia C, Maffrand JP, Le Fur G. Bind-ing of [3H] SR 49059, a potent nonpeptide vasopressin V1a antagonist, torat and human liver membranes. Biochem Biophys Res Commun 1994;199:353–60.
39] Stewart AM, Desmond D, Kyzar E, Gaikwad S, Roth A, Riehl R, et al. Perspectivesof zebrafish models of epilepsy: what, how and where next? Brain Res Bull2012;87:135–43.
40] Streisinger G. The zebrafish book. Eugene. Oregon: Oregon Press; 2000.41] Teng Y, Xie X, Walker S, Saxena M, Kozlowski DJ, Mumm JS, et al. Loss of
zebrafish lgi1b leads to hydrocephalus and sensitization to pentylenetetrazolinduced seizure-like behavior. PLoS One 2011;6:e24596.
42] Thompson RR, Walton JC. Peptide effects on social behavior: effects of vasotocinand isotocin on social approach behavior in male goldfish (Carassius auratus).Behav Neurosci 2004;118:620–6.
43] Tiedeken JA, Ramsdell JS. Embryonic exposure to domoic acid increases the sus-ceptibility of zebrafish larvae to the chemical convulsant pentylenetetrazole.Environ Health Perspect 2007;115:1547–52.
44] Tiedeken JA, Ramsdell JS. DDT exposure of zebrafish embryos enhances seizuresusceptibility: relationship to fetal p,p′-DDE burden and domoic acid exposureof California sea lions. Environ Health Perspect 2009;117:68–73.
45] Todd KJ, Slatter CA, Ali DW. Activation of ionotropic glutamate receptors onperipheral axons of primary motoneurons mediates transmitter release at thezebrafish NMJ. J Neurophysiol 2004;91:828–40.
46] Tugtepe H, Sener G, Biyikli NK, Yüksel M, Cetinel S, Gedik N, et al. The protectiveeffect of oxytocin on renal ischemia/reperfusion injury in rats. Regul Pept Suppl2007;140:101–8.
47] Warne JM. Cloning and characterization of an arginine vasotocin recep-
ormones protect against pentylenetetrazole-induced seizures in), http://dx.doi.org/10.1016/j.peptides.2012.07.013
tor from the euryhaline flounder Platichthys flesus. Gen Comp Endocrinol2001;22:312–9.
48] Wong K, Stewart A, Gilder T, Wu N, Frank K, Gaikwad S, et al. Modeling seizure-related behavioral and endocrine phenotypes in adult zebrafish. Brain Res2010;348:209–15.