
Maternal Olfaction Differentially Modulates Oxytocin and Prolactin Release during Suckling in Goats
Upload
independentCategory
view
1download
0
Rd
GD
a
ARRAA
KHPRONS
1
irmn
fci([tpippm[
dP
0d
Peptides 31 (2010) 1847–1852
Contents lists available at ScienceDirect
Peptides
journa l homepage: www.e lsev ier .com/ locate /pept ides
ole of central NO-cGMP pathway in vasopressin and oxytocin gene expressionuring sepsis
abriela Ravanelli Oliveira-Pelegrin, Fábio Alves Aguila, Paulo José Basso, Maria José Alves Rocha ∗
epartamento de Morfologia, Estomatologia e Fisiologia, Faculdade de Odontologia de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto, São Paulo, Brazil
r t i c l e i n f o
rticle history:eceived 29 March 2010eceived in revised form 26 June 2010ccepted 28 June 2010vailable online 16 July 2010
eywords:
a b s t r a c t
Sepsis induces massive production of inflammatory mediators, such as nitric oxide (NO), and causesneuroendocrine and cardiovascular alterations. This study investigates the involvement of the central NO-cGMP pathway in arginine vasopressin (AVP) and oxytocin (OXY) gene expression during sepsis inducedby cecal ligation and puncture (CLP). Male Wistar rats received an i.c.v. injection of ODQ (0.25 �g/�L),a selective inhibitor of the heme site of soluble guanylate cyclase, or of 1% dymethilsulfoxide (DMSO),as vehicle. Thirty minutes after the injections, sepsis was induced by cecal ligation and puncture or the
ypothalamusolymicrobial sepsiseal-time PCRDQitric oxideoluble guanylate cyclase
animals were sham operated. The ODQ pre-treatment did not alter the progressive NO increase observedafter CLP. In the supraoptic nucleus (SON), this pretreatment increased the relative gene expressionratio of AVP and OXY in the initial phase of sepsis, but in the late phase, the gene expression of bothhormones was reduced. In the paraventricular nucleus (PVN), soluble guanylate cyclase inhibition causedan even larger decrease in the relative gene expression ratio of AVP and OXY during sepsis. These resultsare indicative of a role of the NO-cGMP pathway in hormonal synthesis in the SON and PVN of the
ymic
hypothalamus during pol. Introduction
Sepsis is the systemic response that results from a complexnteraction between host and infectious agents. The excessiveesponse to infection leads to a high production of inflammatoryediators such as nitric oxide (NO) that is reported to affect auto-
omic and neuroendocrine functions during sepsis [6,19,23,26,28].NO is a gaseous mediator synthesized by NO synthases (NOS)
rom the semi-essential amino acid l-arginine [11,24,27]. In theentral nervous system, NO has been reported to participaten the modulation of arginine vasopressin (AVP) and oxytocinOXY) secretion, showing either stimulatory or inhibitory effects5,6,14,39,40]. AVP and OXY are nonapeptides hormones syn-hesized in magnocellular neurons of the supraoptic (SON) andaraventricular (PVN) nuclei of the hypothalamus [7,10]. Clin-
cal and experimental sepsis studies have reported high AVP
lasma levels in the initial phase of sepsis, whereas in the latehase, despite continued hypotension, the levels of these hor-ones are inappropriately low, contributing to vasodilatory shock6,21,26,28,34]. As regards OXY secretion, similar patterns release
∗ Corresponding author at: Departamento de MEF – Faculdade de Odontologiae Ribeirão Preto, USP, Avenida do Café s/n, Monte Alegre, CEP 14040-904 Ribeirãoreto, SP, Brazil. Tel.: +55 16 3602 3974; fax: +55 16 3633 0999.
E-mail address: [email protected] (M.J.A. Rocha).
196-9781/$ – see front matter © 2010 Elsevier Inc. All rights reserved.oi:10.1016/j.peptides.2010.06.031
robial sepsis.© 2010 Elsevier Inc. All rights reserved.
was observed during lipopolysaccharides-induced endotoxemia[2,5,8,14]. The reasons underlying the relative deficient secretion ofneurohypophyseal hormones during sepsis remain unclear [15,32].One hypothesis suggests that hormonal synthesis is impaired dueto NO overproduction by inducible NOS (iNOS), which, is activatedunder sepsis conditions [5,6,13,15,32]. A recent study using in situhybridization and immunocytochemistry showed that there are noapparent differences in AVP mRNA content between SON and PVNin clinical and experimental septic shock, but revealed differencesin hormone content [36]. The authors suggested that translation ofAVP mRNA and/or transport of the peptide may be differentiallyregulated in SON and PVN magnocellular neurons [36].
The modulatory action of NO can be mediated by its majorphysiological receptor, the soluble guanylate cyclase (sGC).When generated, NO rapidly diffuses to adjacent cells where itactivates sGC and increases the formation of cyclic guanosine-3′,5′-monophosphate (cGMP) [24,37]. The interaction of NO with theheme moiety of sGC results in a conformational change whichactivates the catalysis of guanosine-5′-triphosphate to cGMP, anintracellular second messenger, which in turn mediates NO-relatedphysiological events [11,20]. There is, however, also evidence that
the magnocellular neuroendocrine response following an osmoticstimulus can occurs independent of the cGMP-pathway [18,39].Since there is no information, so far, whether this may also apply tosepsis condition, our aim was to evaluate the role of the central NO-cGMP pathway in the regulation of AVP and OXY transcript levels in
1 / Pept
ts
2
2
tiltaCHts
2
1cifc
2
kadnato
2
gawrbstbwattaama
2
aoupa
848 G.R. Oliveira-Pelegrin et al.
he SON and PVN nuclei during cecal ligation and puncture-inducedepsis.
. Materials and methods
.1. Animals
Male Wistar rats (200–250 g) provided by the Animal Facility ofhe Campus of Ribeirão Preto, University of São Paulo, were housedn controlled temperature (25 ± 1 ◦C) and photoperiodic (12:12 hight:day cycle) conditions, with food (Nuvilab CR-1, Nuvital) andap water available ad libitum. All experimental protocols werepproved and performed according to the guidelines of the Ethicsommittee of the University of São Paulo – Campus Ribeirão Preto.umane endpoints in shock research [25] were used as criterion
o euthanize CLP-animals in high suffering, immediately before oroon after the studied time-points defined in this study.
.2. Drugs
The selective inhibitor of heme site of soluble guanylate cyclaseH-[1,2,4] oxadiazole [4,3,-a] quinoxalin-1-one (ODQ) was pur-hased from Tocris Bioscience (St. Louis, MO, USA) and dissolvedn vehicle consisting of 1% dymethilsulfoxide (DMSO) in pyrogen-ree sterile saline at concentrations of 0.25 �g/�L. The dose washosen on basis of previous studies [30].
.3. Lateral ventricle cannulation procedure
Animals were anesthetized by intraperitoneal injection ofetamine-xylazine (100 and 14 mg/kg, respectively) and fixed inKopf stereotaxic frame. A stainless guide cannula (0.7 mm outeriameter) was introduced into the right lateral ventricle (coordi-ates: A: −0.8 mm, L: 1.4 mm, D: 3.2–3.7 mm). The cannula wasttached to the bone with stainless screws and acrylic cement. Aight-fitting stylet was kept inside the guide cannula to preventcclusion.
.4. Cecal ligation and puncture surgery
Animals were randomly assigned to one of two groups, CLProup and sham-operated group. All experiments were performedt the same time of day (9:00–11:00 AM). Induction of severe sepsisas performed by a CLP model as previously described [26]. Briefly,
ats were anesthetized with a short-acting anesthetic agent (tri-romoethanol; 2.5%, 250 mg/kg i.p.; Acros Organics). Under sterileurgical conditions the animals were subjected to a midline laparo-omy, and the cecum was carefully isolated to avoid damage to thelood vessels. The cecum was then ligated below the ileocecal valveithout causing bowel obstruction and punctured ten times with16-gauge needle allowing the fecal contents to spill into the peri-
oneum. Sham-operated animals were submitted to laparotomy,he cecum was manipulated but neither ligated nor punctured. Thebdominal cavity was closed in two layers, and all animals receivedsubcutaneous injection of saline (20 mL/kg body weight). The ani-als were allowed to recover in their cages with free access to food
nd water.
.5. Experimental protocol
Intracerebroventricular (i.c.v.) injections were performed using
10 �L Hamilton syringe and a dental injection needle (200 �muter diameter; Mizzy, Brazil) connected to a PE-10 tube. The vol-me of each injection was 1.0 �L for all protocols. Injections wereerformed over a period of 1 min, and an additional minute wasllowed to elapse before the injection needle was removed from the
ides 31 (2010) 1847–1852
guide cannula to avoid reflux. The animals were kept in their boxesduring the entire procedure. They received an i.c.v. injection ofODQ (0.25 �g/�L) or 1% DMSO 30 min before surgeries. The sham-operated or CLP-induced sepsis animals were decapitated at 4, 6,or 24 h for blood and brain collection. Hematocrit was measured bycentrifugation, plasma osmolality was assessed by freezing-pointdepression (Precision System, Inc.), and plasma nitrate was quan-tified by chemoluminescence (Sievers 280 Noa, Sievers, Boulder,Co, Eua). The SON and PVN regions were carefully microdissectedfrom the hypothalamus for total RNA extraction and processed forquantitative polymerase chain reaction (qPCR) to evaluate AVP andOXY relative gene expression ratios in these nuclei.
2.6. Tissue collection, RNA extraction and reverse transcription
Rats were decapitated and the brain was rapidly removed fromthe cranium, frozen in dry ice and then stored at −80 ◦C untilrequired. For cryostat sectioning, the brains were placed in a brainmatrix (Insight Equipment LtdA, Ribeirao Preto, Brazil) and 1 mmof the hypothalamus were removed en bloc. The SON and PVN ofeach brain were identified with the help of the stereotaxic atlas [38]bilaterally punched out by using a needle of 1200 �m and 1400 �mdiameter, respectively.
The tissues punched were carefully homogenized and the totalRNA extracted on 1 mL of TRIzol Reagent (Invitrogen, Carlsbad, CA,USA). All samples were treated with RNase-free DNAse I (Invit-rogen, Carlsbad, CA, USA) to remove any contaminating genomicDNA. The purity and quantification of total RNA isolated wereassessed by spectrophotometry using a NanoDrop® ND-1000 sys-tem (Nanodrop Technologies). First-strand cDNA synthesis wascarried out using the following protocol. Two �g of total RNA, 1 �Lof oligo(dT)12–18 primer (0.5 �g/�L, Invitrogen, Carlsbad, CA, USA)and 1 �L of dNTP mix (10 mM) (Invitrogen, Carlsbad, CA, USA) wereincubated at 69 ◦C for 5 min and chilled on ice. Subsequently, 4 �L of5× First Strand Buffer, 2 �L of DTT (0.1 M) and 1 �L of RNAseOUTTM
Ribonuclease Inhibitor (Invitrogen, Carlsbad, CA, USA) were addedand the samples incubated for 2 min at 42 ◦C. Next, SuperScriptTM
II Reverse Transcriptase (200 U, Invitrogen, Carlsbad, CA, USA) wasadded and the reaction incubated at 42 ◦C for 50 min followed by15 min at 70 ◦C. cDNA was stored undiluted at −20 ◦C until analysis.All cDNA samples were diluted 1:5 with DEPC-treated water beforebeing used as templates in quantitative PCR assays.
2.7. Primer design
Gene-specific primers (AVP: (+) atc cga cat gga gct gag ac, (−)aaa aac cct ctc gac act cg; OXY: (+) tgc ccc agt ctc gct tgc, (−) cgaagc agc cca gct cgt c; GAPDH: (+) tca cca cca tgg aga agg c, (−)gct aag cag ttg gtg gtg ca; ACTb: (+) tgt cac caa ctg gga cga ta,(−) ggg gtg ttg aag gtc tca aa; 18S: (+) acg gaa ggg cac cac cag ga,(−) cac cac cac cca cgg aat cg) were designed based on GenBanksequences (see Supplementary Material). BLAST results show thatthe primers specifically amplified AVP and OXY gene fragments.The reference gene (GAPDH, ACTb and 18S) primer combinationshad already been validated in previous studies [1].
2.8. Quantitative real-time PCR (qPCR)
Quantitative real-time PCR studies were performed using SYBR®
Green methodology (Applied Biosystems) in a Mastercycler® eprealplex (Eppendorf, Hamburg, Germany). qPCR reactions, per-
formed in 96-well 0.2 mL thin-wall PCR microplates (Axygen)sealed with film, consisted of 8 �L of SYBR Green PCR Master Mix,2.5 �L of each forward and reverse primer (10 �M), and 2 �L of1:5-diluted template cDNA in a total volume of 20 �L. Cycling wasperformed using the optimized conditions in a four-step exper-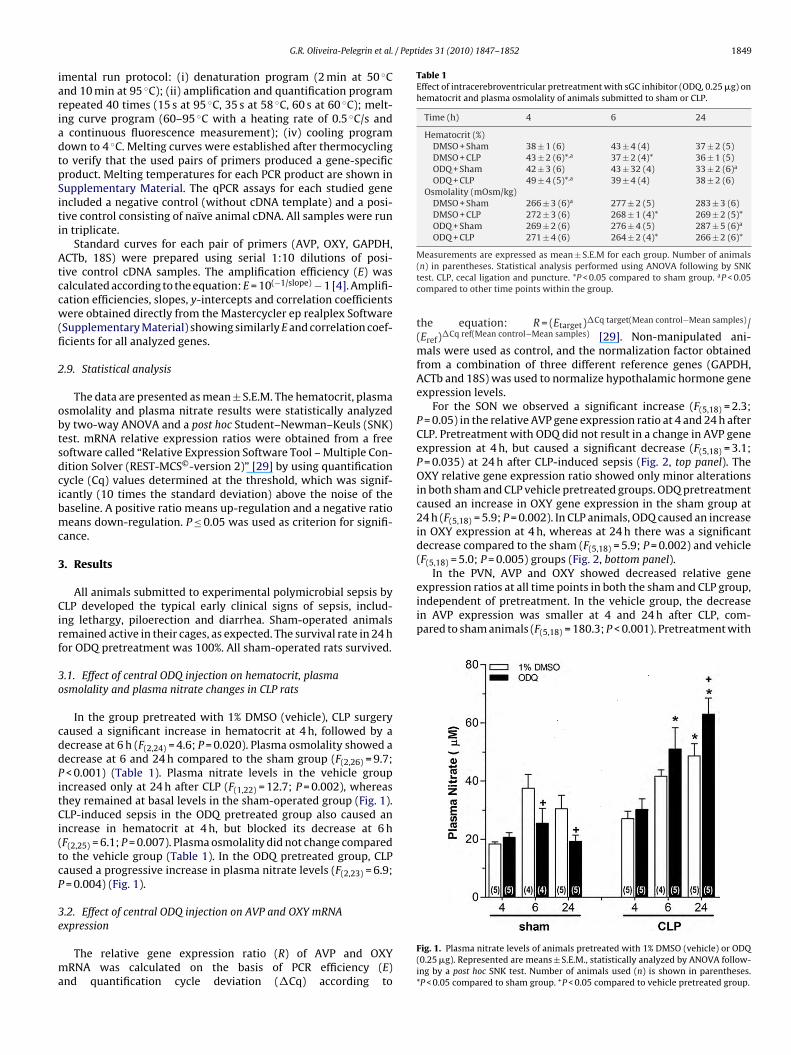
/ Peptides 31 (2010) 1847–1852 1849
iariadtpSiti
Atccw(fi
2
obtsdcibmc
3
Cirf
3o
cddPitCi(tcP
3e
ma
Table 1Effect of intracerebroventricular pretreatment with sGC inhibitor (ODQ, 0.25 �g) onhematocrit and plasma osmolality of animals submitted to sham or CLP.
Time (h) 4 6 24
Hematocrit (%)DMSO + Sham 38 ± 1 (6) 43 ± 4 (4) 37 ± 2 (5)DMSO + CLP 43 ± 2 (6)*,a 37 ± 2 (4)* 36 ± 1 (5)ODQ + Sham 42 ± 3 (6) 43 ± 32 (4) 33 ± 2 (6)a
ODQ + CLP 49 ± 4 (5)*,a 39 ± 4 (4) 38 ± 2 (6)Osmolality (mOsm/kg)
DMSO + Sham 266 ± 3 (6)a 277 ± 2 (5) 283 ± 3 (6)DMSO + CLP 272 ± 3 (6) 268 ± 1 (4)* 269 ± 2 (5)*ODQ + Sham 269 ± 2 (6) 276 ± 4 (5) 287 ± 5 (6)a
ODQ + CLP 271 ± 4 (6) 264 ± 2 (4)* 266 ± 2 (6)*
expression ratios at all time points in both the sham and CLP group,independent of pretreatment. In the vehicle group, the decreasein AVP expression was smaller at 4 and 24 h after CLP, com-pared to sham animals (F(5,18) = 180.3; P < 0.001). Pretreatment with
G.R. Oliveira-Pelegrin et al.
mental run protocol: (i) denaturation program (2 min at 50 ◦Cnd 10 min at 95 ◦C); (ii) amplification and quantification programepeated 40 times (15 s at 95 ◦C, 35 s at 58 ◦C, 60 s at 60 ◦C); melt-ng curve program (60–95 ◦C with a heating rate of 0.5 ◦C/s and
continuous fluorescence measurement); (iv) cooling programown to 4 ◦C. Melting curves were established after thermocyclingo verify that the used pairs of primers produced a gene-specificroduct. Melting temperatures for each PCR product are shown inupplementary Material. The qPCR assays for each studied genencluded a negative control (without cDNA template) and a posi-ive control consisting of naïve animal cDNA. All samples were runn triplicate.
Standard curves for each pair of primers (AVP, OXY, GAPDH,CTb, 18S) were prepared using serial 1:10 dilutions of posi-
ive control cDNA samples. The amplification efficiency (E) wasalculated according to the equation: E = 10(−1/slope) − 1 [4]. Amplifi-ation efficiencies, slopes, y-intercepts and correlation coefficientsere obtained directly from the Mastercycler ep realplex Software
Supplementary Material) showing similarly E and correlation coef-cients for all analyzed genes.
.9. Statistical analysis
The data are presented as mean ± S.E.M. The hematocrit, plasmasmolality and plasma nitrate results were statistically analyzedy two-way ANOVA and a post hoc Student–Newman–Keuls (SNK)est. mRNA relative expression ratios were obtained from a freeoftware called “Relative Expression Software Tool – Multiple Con-ition Solver (REST-MCS©-version 2)” [29] by using quantificationycle (Cq) values determined at the threshold, which was signif-cantly (10 times the standard deviation) above the noise of theaseline. A positive ratio means up-regulation and a negative ratioeans down-regulation. P ≤ 0.05 was used as criterion for signifi-
ance.
. Results
All animals submitted to experimental polymicrobial sepsis byLP developed the typical early clinical signs of sepsis, includ-
ng lethargy, piloerection and diarrhea. Sham-operated animalsemained active in their cages, as expected. The survival rate in 24 hor ODQ pretreatment was 100%. All sham-operated rats survived.
.1. Effect of central ODQ injection on hematocrit, plasmasmolality and plasma nitrate changes in CLP rats
In the group pretreated with 1% DMSO (vehicle), CLP surgeryaused a significant increase in hematocrit at 4 h, followed by aecrease at 6 h (F(2,24) = 4.6; P = 0.020). Plasma osmolality showed aecrease at 6 and 24 h compared to the sham group (F(2,26) = 9.7;< 0.001) (Table 1). Plasma nitrate levels in the vehicle group
ncreased only at 24 h after CLP (F(1,22) = 12.7; P = 0.002), whereashey remained at basal levels in the sham-operated group (Fig. 1).LP-induced sepsis in the ODQ pretreated group also caused an
ncrease in hematocrit at 4 h, but blocked its decrease at 6 hF(2,25) = 6.1; P = 0.007). Plasma osmolality did not change comparedo the vehicle group (Table 1). In the ODQ pretreated group, CLPaused a progressive increase in plasma nitrate levels (F(2,23) = 6.9;= 0.004) (Fig. 1).
.2. Effect of central ODQ injection on AVP and OXY mRNA
xpressionThe relative gene expression ratio (R) of AVP and OXYRNA was calculated on the basis of PCR efficiency (E)
nd quantification cycle deviation (�Cq) according to
Measurements are expressed as mean ± S.E.M for each group. Number of animals(n) in parentheses. Statistical analysis performed using ANOVA following by SNKtest. CLP, cecal ligation and puncture. *P < 0.05 compared to sham group. aP < 0.05compared to other time points within the group.
the equation: R = (Etarget)�Cq target(Mean control−Mean samples)/(Eref)�Cq ref(Mean control−Mean samples) [29]. Non-manipulated ani-mals were used as control, and the normalization factor obtainedfrom a combination of three different reference genes (GAPDH,ACTb and 18S) was used to normalize hypothalamic hormone geneexpression levels.
For the SON we observed a significant increase (F(5,18) = 2.3;P = 0.05) in the relative AVP gene expression ratio at 4 and 24 h afterCLP. Pretreatment with ODQ did not result in a change in AVP geneexpression at 4 h, but caused a significant decrease (F(5,18) = 3.1;P = 0.035) at 24 h after CLP-induced sepsis (Fig. 2, top panel). TheOXY relative gene expression ratio showed only minor alterationsin both sham and CLP vehicle pretreated groups. ODQ pretreatmentcaused an increase in OXY gene expression in the sham group at24 h (F(5,18) = 5.9; P = 0.002). In CLP animals, ODQ caused an increasein OXY expression at 4 h, whereas at 24 h there was a significantdecrease compared to the sham (F(5,18) = 5.9; P = 0.002) and vehicle(F(5,18) = 5.0; P = 0.005) groups (Fig. 2, bottom panel).
In the PVN, AVP and OXY showed decreased relative gene
Fig. 1. Plasma nitrate levels of animals pretreated with 1% DMSO (vehicle) or ODQ(0.25 �g). Represented are means ± S.E.M., statistically analyzed by ANOVA follow-ing by a post hoc SNK test. Number of animals used (n) is shown in parentheses.*P < 0.05 compared to sham group. +P < 0.05 compared to vehicle pretreated group.

1850 G.R. Oliveira-Pelegrin et al. / Peptides 31 (2010) 1847–1852
Fig. 2. Effect of intracerebroventricular pretreatment with the soluble guanylatecyclase inhibitor ODQ (0.25 �g) on vasopressin (AVP, top panel) and oxytocin (OXY,bfbc
O(gPi(ssCOPp
4
pSueN
i
Fig. 3. Effect of intracerebroventricular pretreatment with the soluble guanylatecyclase inhibitor ODQ (0.25 �g) on vasopressin (AVP, top panel) and oxytocin (OXY,
on AVP and OXY transcript levels under conditions of polymicro-
ottom panel) relative gene expression ratios in the supraoptic nucleus (SON) (n = 4or each group). The data show means ± S.E.M. Ratios were obtained and analyzedy using Relative Expression Software Tool – Multiple Condition Solver. *P < 0.05ompared to sham group. +P < 0.05 compared to vehicle pretreated group.
DQ, when compared with vehicle, promoted a smaller decreaseF(5,18) = 95.5; P < 0.001) on AVP relative expression in the shamroup, but caused a significant decrease after CLP (F(5,15) = 3.5;= 0.021) (Fig. 3, top panel). The gene expression ratio of OXY
n the CLP group pretreated with vehicle showed a reductionF(5,18) = 69.2; P < 0.001) only after 4 h. ODQ pretreatment in theham group caused a smaller decrease in the OXY gene expres-ion than vehicle (F(5,18) = 47.7; P < 0.001) at all time points. InLP animals, however ODQ promoted a further reduction in theXY expression when compared with the vehicle (F(5,18) = 132.5;< 0.001) and sham (F(5,18) = 176.8; P < 0.001) groups (Fig. 3, bottomanel).
. Discussion
In our experiments, we observed that the central NO-cGMPathway affects AVP and OXY relative gene expression ratio in theON and PVN of the hypothalamus during polymicrobial sepsis. Wesed ODQ, a potent and highly selective inhibitor of sGC, which is
stablished as a powerful tool to investigate the function of theO-cGMP pathway [12].ODQ competes with NO for the heme site of sGC and causes anrreversible inhibition of the NO-cGMP pathway [12,33]. Moreover,
bottom panel) relative gene expression ratios in the paraventricular nucleus (PVN)(n = 4 for each group). The data show means ± S.E.M. Ratios were obtained andanalyzed by using Relative Expression Software Tool – Multiple Condition Solver.*P < 0.05 compared to sham group. +P < 0.05 compared to vehicle pretreated group.
ODQ also substantially reduces the nitration of proteins causedby peroxynitrite and/or reactive oxygen species [41]. This sGCinhibitor does not act on particulate guanylate cyclase or on adeny-late cyclase, nor does it affect the activity of NO synthases. It alsodoes not inactivate NO nor does it block actions of NO that areunrelated to sGC [12].
We could show the CLP surgery induced NO formation as evi-denced by the progressive increase in nitrate plasma levels. Asexpected an i.c.v. injection of ODQ did not affect the nitrate levels.Increased NO formation may alter vascular permeability and conse-quently hematocrit levels and plasma osmolality, which were alsonot affected by pretreatment with ODQ.
In the initial phase of sepsis besides high AVP secretion thereis also neuronal activation of the SON and PVN, while in the latephase, when the plasma level of this hormone is inappropriatelybasal, a reduced activation of the magnocellular neurons was alsoreported [6,21,26,28,34]. The neuronal activation of the hypotha-lamus nuclei may be correlated with hormonal gene expression. Inour experiments we investigated the role of the NO-cGMP pathway
bial sepsis. In the initial phase of sepsis, the central inhibition ofsGC by ODQ increased the relative expression ratio of AVP and OXYin relation to sham in the SON. But in relation to the vehicle thechange was seen only for OXY. In the late phase (24 h), there was,

/ Pept
hcOewcrt
sisisams[
cSectto
lbasfecfr
roicaes
5
is
A
eiF#
A
t
[
[
[
[
[
[
[
[[
[
[
[
[
[
[
[
[
[
[
[
[
G.R. Oliveira-Pelegrin et al.
owever, a reduction in transcript levels of both hormones, whenompared to the sham-operated and vehicle groups. In the PVN,DQ injection caused an even larger decrease in the relative genexpression ratio of AVP and OXY during sepsis when comparedith the sham and vehicle groups. Therefore these hypothalamic
hanges suggest that there is a neuronal impairment that may beegulated independently of the NO-cGMP pathway, especially inhe SON.
Apoptosis has been reported to occur in the hypothalamus ofeptic patients [35] and appointed as responsible for neuronalmpairment seen by the decrease of c-fos expression in braintructures in septic rats [3,6]. During sepsis, the large increase inNOS-derived NO irreversibly inhibits cytochrome c oxidase anduppresses mitochondrial respiration, leading to ATP depletionnd glutamate release, which, in turn, may compromise cellularetabolism and to cause neuronal death [22]. Furthermore, the
uppression of mitochondrial respiration is not affected by ODQ42], indicating that it is a cGMP-independent process.
The differential hormone gene expression in the SON and PVNan be explained by the low activity of sGC in the PVN observed byoutham and Garthwaite [37]. These authors showed that the PVNxhibited intense staining for NADPH diaphorase, but was low forGMP after sodium nitroprusside stimulation [37]. However, sincehe PVN is closer to the third ventricle than the SON, it is possiblehat the observed difference may partly due to the higher sensitivityf PVN to the i.c.v. injection of the ODQ.
DMSO, the vehicle required to dissolve ODQ, had itself a stimu-atory effect on AVP and OXY secretion [39]. Moreover, it has otheriological effects, being analgesic, anti-inflammatory, antioxidantnd acting as a vasoconstrictor by antagonizing vasoactive sub-tances [17,31]. These intrinsic effects of DMSO may be responsibleor some of the changes observed in hypothalamic hormone genexpression, mainly in the SON. However, the inhibition of the NO-GMP pathway by ODQ caused changes that were clearly differentrom the intrinsic effects of DMSO, attributing significance to ouresults.
Although polymicrobial sepsis induced by CLP may not fully rep-esent a typical clinical situation, it closely resembles the diversityf infectious agents and the progression of peritoneal contam-nation [9,16]. In this study, using this experimental model inombination with a sGC inhibitor we observed for the first timerole of the NO-cGMP pathway on AVP and OXY transcript lev-
ls in the SON and PVN hypothalamic nuclei during polymicrobialepsis.
. Conclusion
Our results are indicative of a role of the NO-cGMP pathwayn hormonal synthesis in the SON and PVN during polymicrobialepsis.
cknowledgments
We thank Nadir Martins Fernandes and Antonio Zanardo for thexcellent technical assistance. Sérgio Akira Uyemura provided thenfrastructure for quantitative PCR assays. Financial support fromundacão de Amparo à Pesquisa do Estado de São Paulo (FAPESP,05/50162-0, #07/08532-0) is gratefully acknowledged.
ppendix A. Supplementary data
Supplementary data associated with this article can be found, inhe online version, at doi:10.1016/j.peptides.2010.06.031.
[
[
ides 31 (2010) 1847–1852 1851
References
[1] Bonefeld BE, Elfving B, Wegener G. Reference genes for normalization: a studyof rat brain tissue. Synapse 2008;62:302–9.
[2] Borges BC, da Rocha MJ. Participation of the subfornical nucleus inhypothalamic-neurohypophyseal axis activation during the early phase ofendotoxic shock. Neurosci Lett 2006;404:227–31.
[3] Bruhn FH, Correa PB, Oliveira-Pelegrin GR, Rocha MJ. Blocking systemicnitric oxide production alters neuronal activation in brain structures involvedin cardiovascular regulation during polymicrobial sepsis. Neurosci Lett2009;453:141–6.
[4] Bustin SA, Benes V, Garson JA, Hellemans J, Huggett J, Kubista M, et al. The MIQEguidelines: minimum information for publication of quantitative real-time PCRexperiments. Clin Chem 2009;55(4):611–22.
[5] Carnio EC, Moreto V, Giusti-Paiva A, Antunes-Rodrigues J. Neuro-immune-endocrine mechanisms during septic shock: role for nitric oxide invasopressin and oxytocin release. Endocr Metab Immune Disord Drug Targets2006;6:137–42.
[6] Correa PB, Pancoto JA, de Oliveira-Pelegrin GR, Carnio EC, Rocha MJ. Participa-tion of iNOS-derived NO in hypothalamic activation and vasopressin releaseduring polymicrobial sepsis. J Neuroimmunol 2007;183:17–25.
[7] Cunningham Jr ET, Sawchenko PE. Reflex control of magnocellular vasopressinand oxytocin secretion. Trends Neurosci 1991;14:406–11.
[8] de Carvalho Borges B, Carnio EC, Elias LL, Antunes-Rodrigues J, Branco LG,da Rocha MJ. Lesion of the anteroventral third ventricle (AV3V) reduceshypothalamic activation and hypophyseal hormone secretion induced bylipopolysaccharide in rats. Brain Res 2006;1115:83–91.
[9] Deitch EA. Rodent models of intra-abdominal infection. Shock 2005;24(Suppl.1):19–23.
10] Du Vigneaud V. Hormones of the posterior pituitary gland: oxytocin and vaso-pressin. Harvey Lect 1954;50:1–26.
11] Feihl F, Waeber B, Liaudet L. Is nitric oxide overproduction the target of choicefor the management of septic shock? Pharmacol Ther 2001;91:179–213.
12] Garthwaite J, Southam E, Boulton CL, Nielsen EB, Schmidt K, Mayer B. Potentand selective inhibition of nitric oxide-sensitive guanylyl cyclase by 1H-[1,2,4]oxadiazolo[4,3-a]quinoxalin-1-one. Mol Pharmacol 1995;48:184–8.
13] Giusti-Paiva A, De Castro M, Antunes-Rodrigues J, Carnio EC. Inducible nitricoxide synthase pathway in the central nervous system and vasopressin releaseduring experimental septic shock. Crit Care Med 2002;30:1306–10.
14] Giusti-Paiva A, Elias LL, Antunes-Rodrigues J. Inhibitory effect of gaseous neu-romodulators in vasopressin and oxytocin release induced by endotoxin in rats.Neurosci Lett 2005;381:320–4.
15] Holmes CL, Patel BM, Russell JA, Walley KR. Physiology of vasopressin relevantto management of septic shock. Chest 2001;120:989–1002.
16] Hubbard WJ, Choudhry M, Schwacha MG, Kerby JD, Rue 3rd LW, Bland KI, et al.Cecal ligation and puncture. Shock 2005;24(Suppl. 1):52–7.
17] Jacob SW, Herschler R. Pharmacology of DMSO. Cryobiology 1986;23:14–27.18] Kadekaro M. Nitric oxide modulation of the hypothalamo-neurohypophyseal
system. Braz J Med Biol Res 2004;37:441–50.19] Kovacs KJ. Neurohypophyseal hormones in the integration of physiological
responses to immune challenges. Prog Brain Res 2002;139:127–46.20] Krumenacker JS, Hanafy KA, Murad F. Regulation of nitric oxide and soluble
guanylyl cyclase. Brain Res Bull 2004;62:505–15.21] Landry DW, Levin HR, Gallant EM, Ashton Jr RC, Seo S, D’Alessandro D, et al.
Vasopressin deficiency contributes to the vasodilation of septic shock. Circula-tion 1997;95:1122–5.
22] Mander P, Brown GC. Nitric oxide, hypoxia and brain inflammation. BiochemSoc Trans 2004;32:1068–9.
23] McCann SM, Kimura M, Karanth S, Yu WH, Mastronardi CA, Rettori V. The mech-anism of action of cytokines to control the release of hypothalamic and pituitaryhormones in infection. Ann N Y Acad Sci 2000;917:4–18.
24] Moncada S, Palmer RM, Higgs EA. Nitric oxide: physiology, pathophysiology,and pharmacology. Pharmacol Rev 1991;43:109–42.
25] Nemzek JA, Xiao HY, Minard AE, Bolgos GL, Remick DG. Humane endpoints inshock research. Shock 2004;21:17–25.
26] Oliveira-Pelegrin GR, Ravanelli MI, Branco LG, Rocha MJ. Thermoregulation andvasopressin secretion during polymicrobial sepsis. Neuroimmunomodulation2009;16:45–53.
27] Palmer RM, Moncada S. A novel citrulline-forming enzyme implicated in theformation of nitric oxide by vascular endothelial cells. Biochem Biophys ResCommun 1989;158:348–52.
28] Pancoto JA, Correa PB, Oliveira-Pelegrin GR, Rocha MJ. Autonomic dysfunctionin experimental sepsis induced by cecal ligation and puncture. Auton Neurosci2008;138:57–63.
29] Pfaffl MW, Horgan GW, Dempfle L. Relative expression software tool (REST) forgroup-wise comparison and statistical analysis of relative expression results inreal-time PCR. Nucleic Acids Res 2002;30:e36.
30] Ravanelli MI, Branco LG. Role of locus coeruleus heme oxygenase-carbonmonoxide-cGMP pathway during hypothermic response to restraint. Brain ResBull 2008;75:526–32.
31] Remick DG, Villarete L. Regulation of cytokine gene expression by reactiveoxygen and reactive nitrogen intermediates. J Leukoc Biol 1996;59:471–5.
32] Rocha MJA, Oliveira GR, Farias-Corrêa PB. Neurohypophyseal hormone secre-tion during septic shock. In: Chen FJ, editor. New trends in brain research. NewYork: Nova Science Publishers; 2006. p. 75–94.

1 / Pept
[
[
[
[
[
[
[
[
[
in rodent models of Gram-positive and Gram-negative shock. Crit Care Med2001;29:1599–608.
[42] Zingarelli B, Hasko G, Salzman AL, Szabo C. Effects of a novel guanylyl
852 G.R. Oliveira-Pelegrin et al.
33] Schrammel A, Behrends S, Schmidt K, Koesling D, Mayer B. Characterization of1H-[1,2,4]oxadiazolo[4,3-a]quinoxalin-1-one as a heme-site inhibitor of nitricoxide-sensitive guanylyl cyclase. Mol Pharmacol 1996;50:1–5.
34] Sharshar T, Blanchard A, Paillard M, Raphael JC, Gajdos P, Annane D. Circulatingvasopressin levels in septic shock. Crit Care Med 2003;31:1752–8.
35] Sharshar T, Gray F, Lorin de la Grandmaison G, Hopkinson NS, Ross E, DorandeuA, et al. Apoptosis of neurons in cardiovascular autonomic centres triggeredby inducible nitric oxide synthase after death from septic shock. Lancet2003;362:1799–805.
36] Sonneville R, Guidoux C, Barrett L, Viltart O, Mattot V, Polito A, et al. Vasopressinsynthesis by the magnocellular neurons is different in the supraoptic nucleus
and in the paraventricular nucleus in human and experimental septic shock.Brain Pathol 2010;20:613–22.37] Southam E, Garthwaite J. The nitric oxide-cyclic GMP signalling pathway in ratbrain. Neuropharmacology 1993;32:1267–77.
38] Swanson LW. Brain maps: structure of the rat brain. Amsterdam: Elsevier Sci-ence; 1998.
ides 31 (2010) 1847–1852
39] Terrell ML, Salas N, Bui V, Summy-Long JY, Kadekaro M. NO inhibition of themagnocellular neuroendocrine system in rats is independent of cGMP signalingpathway. Exp Neurol 2003;184:846–56.
40] Yasin S, Costa A, Trainer P, Windle R, Forsling ML, Grossman A. Nitricoxide modulates the release of vasopressin from rat hypothalamic explants.Endocrinology 1993;133:1466–9.
41] Zacharowski K, Berkels R, Olbrich A, Chatterjee PK, Cuzzocrea S, Foster SJ, et al.The selective guanylate cyclase inhibitor ODQ reduces multiple organ injury
cyclase inhibitor on the vascular actions of nitric oxide and peroxynitrite inimmunostimulated smooth muscle cells and in endotoxic shock. Crit Care Med1999;27:1701–7.
Copyright © 2022 FDOKUMEN




















