
Gonadal steroids influence the involvement of arginine vasopressin in social recognition in mice
13
Psychoneuroendocrinology, Vol. 18, No. 4, pp. 323-335, 1993 0306-4530/93 $6.00 + .00 Printed in the U.S.A. © 1993 Pergamon Press Ltd. GONADAL STEROIDS INFLUENCE THE INVOLVEMENT OF ARGININE VASOPRESSIN IN SOCIAL RECOGNITION IN MICE R.M. BLUTHI~, G. GHEUSI and R. DANTZER INRA-INSERM U176, Rue Camille Saint-Sa~ns, 33077 Bordeaux Cedex, France (Received 17 August 1992; in final form 10 January 1993) SUMMARY Gonadal steroids have been shown to modulate the involvement of vasopressinergic neurotransmis- sion in social recognition in rats. To assess whether the same phenomenon occurs in another species showing sexual dimorphism of vasopressinergic neurons, social recognition was studied in DBA2 male mice. Social recognition was inferred from the reduction in investigation time of a juvenile conspecific when this social stimulus was presented for the second time at different intervening intervals after the initial exposure. Such a reduction occurred when the interval was 20 min or 1 hr but not 2 hr. This effect was stimulus-specific, because it did not occur when a different juvenile was presented on the second exposure. AVP (0.4 ~g/mouse, SC) injected immediately after the first exposure to the juvenile prolonged social recognition, whereas SC injection of an antagonist of the vasopressor receptors of AVP, dPTyr(Me)AVP (2/.~g/mouse) impaired it. Compared with intact males, castrated mice spent less time investigating juveniles hut were still able to recognize them after a 2-hr interval. However, this recognition was no longer sensitive to dPTyr(Me)AVP. These results confirm that androgen-dependent vasopressinergic transmission modulates social recognition in mice. INTRODUCTION IN RATS, ARGININEvasopressin (AVP) containing neurons projecting from the bed nucleus of the stria terminalis (BNST) to the lateral septum and the lateral habenular nucleus are sexually dimorphic. Male rats have more immunoreactive neurons in the BNST and a higher density of immunoreactive projections of these neurons to the septum (De Vries et al., 1981). This sex difference is dependent on gonadal steroids--it is reduced by castration and restored to normal following testosterone replacement therapy (De Vries et al., 1984). Testosterone is thought to act via aromatase-coupled androgen receptors, because treatment of castrated male rats with oestradiol-17fl results in a normal AVP fibre distribution, but with less dense staining than that observed in intact animals or in animals treated with testosterone (De Vries et al., 1986). Gonadal steroids regulate AVP Address correspondence and reprint requests to: R.M. Bluth6, INSERM U176, Rue Camille Saint- Sa~ns, 33077 Bordeaux Cedex, France. 323
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Gonadal steroids influence the involvement of arginine vasopressin in social recognition in mice

Psychoneuroendocrinology, Vol. 18, No. 4, pp. 323-335, 1993 0306-4530/93 $6.00 + .00 Printed in the U.S.A. © 1993 Pergamon Press Ltd.
G O N A D A L S T E R O I D S I N F L U E N C E T H E I N V O L V E M E N T O F A R G I N I N E V A S O P R E S S I N I N S O C I A L R E C O G N I T I O N
I N M I C E
R.M. BLUTHI~, G. GHEUSI and R. DANTZER INRA-INSERM U176, Rue Camille Saint-Sa~ns,
33077 Bordeaux Cedex, France
(Received 17 August 1992; in final form 10 January 1993)
SUMMARY
Gonadal steroids have been shown to modulate the involvement of vasopressinergic neurotransmis- sion in social recognition in rats. To assess whether the same phenomenon occurs in another species showing sexual dimorphism of vasopressinergic neurons, social recognition was studied in DBA2 male mice. Social recognition was inferred from the reduction in investigation time of a juvenile conspecific when this social stimulus was presented for the second time at different intervening intervals after the initial exposure. Such a reduction occurred when the interval was 20 min or 1 hr but not 2 hr. This effect was stimulus-specific, because it did not occur when a different juvenile was presented on the second exposure. AVP (0.4 ~g/mouse, SC) injected immediately after the first exposure to the juvenile prolonged social recognition, whereas SC injection of an antagonist of the vasopressor receptors of AVP, dPTyr(Me)AVP (2/.~g/mouse) impaired it. Compared with intact males, castrated mice spent less time investigating juveniles hut were still able to recognize them after a 2-hr interval. However, this recognition was no longer sensitive to dPTyr(Me)AVP. These results confirm that androgen-dependent vasopressinergic transmission modulates social recognition in mice.
INTRODUCTION
IN RATS, ARGININE vasopressin (AVP) containing neurons projecting from the bed nucleus of the stria terminalis (BNST) to the lateral septum and the lateral habenular nucleus are sexually dimorphic. Male rats have more immunoreactive neurons in the BNST and a higher density of immunoreactive projections of these neurons to the septum (De Vries et al., 1981). This sex difference is dependent on gonadal steroids--it is reduced by castration and restored to normal following testosterone replacement therapy (De Vries et al., 1984). Testosterone is thought to act via aromatase-coupled androgen receptors, because treatment of castrated male rats with oestradiol-17fl results in a normal AVP fibre distribution, but with less dense staining than that observed in intact animals or in animals treated with testosterone (De Vries et al., 1986). Gonadal steroids regulate AVP
Addre s s c o r r e s p o n d e n c e and repr in t r eques t s to: R.M. Bluth6, I N S E R M U176, Rue Camil le Saint- Sa~ns, 33077 B o r d e a u x Cedex , F rance .
323

324 R.M. BLUTHf~ et al.
synthesis at the gene level, as revealed by the higher levels of AVP mRNA in intact males as compared with females (Miller et al., 1989b) and the increased expression of AVP mRNA in castrated animals implanted with a testosterone capsule, as compared to castrates implanted with an empty capsule (Miller et al., 1989a).
The same sex difference has been found in mice (Mayes et al., 1988). Dense neurophy- sin II (the carrier protein associated with AVP) immunoreactivity appears earlier in the lateral septum and the lateral habenula during postnatal development in male than in female mice and the subsequent density is higher in males. Gonadectomy results in a gradual loss of neurophysin II immunoreactivity in these structures, which is restored to normal following testosterone implants. Decreased neurophysin II immunoreactivity is also observed in mutant hypogonadal mice (Mayes et al., 1988).
In rats, androgen-dependent vasopressinergic neurons have been implicated in social recognition, a form of short-term memory. When presented with a juvenile conspecific, rats engage in bouts of investigation of this social stimulus, the duration of which depends on the familiarity with the conspecific. Since novel animals are explored longer than familiar ones, the difference between durations of social investigation of the same stimulus animal presented at various time intervals can be used as a convenient index of the memory for this particular individual. In intact male rats, this form of memory lasts for less than l hr (Thor & Holloway, 1982; Dantzer et al., 1987; Sekiguchi et al., 1991). It is prolonged by administration of AVP and blocked by administration of a specifc antagonist of the vasopressor receptors (V 1) ofAVP, dPTyr(Me)AVP. However, adminis- tration of the V1 receptor antagonist has no effect on social recognition in castrated males (Bluth6 et al., 1990) and in females (Bluth6 & Dantzer, 1990). Implantation of a testosterone-filled capsule in castrated male rats restores sensitivity of social recognition to the action of the V1 receptor antagonist (Bluth6 et al., 1990).
The present experiment was carried out to test whether androgen-dependent vasopres- sinergic neurons are similarly involved in the modulation of social recognition in mice. In the absence of any published study on social recognition in mice, we first assessed the ability of mice to recognize previously presented juveniles and the sensitivity of this recognition to different intervening treatments, including repeated presentations of the same juvenile, presentation of a different juvenile and injection of AVP (Dantzer et al. , 1987). We then tested the sensitivity of mice to the V1 receptor antagonist before and after castration. We confirm that mice are able to form a memory for previously presented juveniles, which influence their subsequent social interactions. We also show that this form of memory is modulated by androgen-dependent vasopressinergic neurons since the VI receptor antagonist which blocks social recognition in intact male mice loses its effect after castration.
ANIMALS AND METHODS
Animal s Male mice of the DBA/2 inbred strain were used. They were purchased from Charles
River (France) at 6 weeks of age. Mice were housed in groups of three in 26.6 × 21.4 × 14.3 cm polypropylene cages under a 12:12 hr light:dark cycle (lights on at 1600h). Food and water were available ad lib. All behavioural tests were conducted when the subjects were 8-11 weeks of age (25-30 g). Castration was performed under chloral hydrate anesthesia in 8-week-old mice (6%, 0.06 ml/10 g mouse) and testing occurred from 2 weeks after surgery. Juvenile male mice (21-35 days of age) of the same strain served as stimulus animals. They were housed in groups of 10 in the same room.

SOCIAL RECOGNITION AND A V P IN MICE 325
General Procedure The procedure described previously for rats (Dantzer et al., 1987) was slightly modified
to adapt it to mice. Social isolation was kept to a minimum to avoid the development of aggression (Brain, 1975). Beginning on the day before the test session and continuing for the next 6 days, mice were individually housed. After this period, they were again housed in groups of three before further testing if necessary.
All behavioral tests took place in the home cage of the test animals during the dark phase of the cycle using a red light, and were recorded with a video camera. Each test consisted of a 4-rain exposure to a juvenile, followed by a second exposure to the same or a different juvenile, at different intervening intervals. Between two successive presentations, juveniles were kept individually in a small cage. During each 4-min test, total duration of active social investigation was measured by typing preset keys on the keyboard of an Apple II computer. Social investigation consisted mainly of anogenital sniffing, face and neck grooming and close-following of the juvenile (Bluth6 et al., 1991). During the recognition test which took place following a defined intervening interval, social investigation was continuously observed until the criterion of neglect was met or up to a maximum of 4 min. The criterion of neglect was defined as 20 continuous seconds with no further investigation of the juvenile by the adult.
Peptides Arginine vasopressin (AVP), a gift from Clin-Midy, Montpellier, and the V 1 receptor
antagonist 1-deaminopenicillamine 2-O-methyltyrosine arginine-vasopressin (dPTyr (Me)AVP, AAVP) (Manning & Sawyer, 1984), a gift from Dr. Jean Rivier (Salk Institute, San Diego, California, USA), were dissolved (1 /zg//~l) in 0.01 M acetic acid and then stored at -80°C in 20-tzl aliquots. On the day of use, each aliquot was diluted in 0.9% saline. Injections were given subcutaneously (SC) in a volume of 100 tzl per mouse. Control animals received the same volume of saline. The dose of AVP (0.4/zg/mouse) was selected on the basis of already published experiments on the memory-enhancing effects of this peptide in mice (Siegfried et al., 1984). The V1 receptor antagonist was injected at five times the dose of AVP, as this ratio fully blocks the effects of AVP on extinction of active avoidance in rats (Le Moal et al., 1981). Treatments were administered SC immediately after the first observation session and were spaced 3-day apart.
Time and Stimulus Dependency of Social Recognition in Intact Male Mice To assess the duration during which mice are able to recognize a juvenile, male mice
(n = 11) were exposed twice to the same juvenile with a 20, 60, 120, or 180 min intervening interval. A different interval was tested every day, in a random order. To assess specificity of social recognition, the same mice were submitted to a presentation of a juvenile followed after 20 rain by presentation of the same or a different juvenile. This last phase of the experiment took place immediately after the preceding one and lasted 2 days.
Facilitation and Inhibition o f Social Recognition To assess the possibility of improving social recognition, male mice (n = 6) were
submitted to two successive presentations of the same juvenile at a 30-min interval. This was followed 90 rain later by a recognition test in which the same juvenile was again presented. To assess the possibility of impairing social recognition, male mice (n = 8) were presented with a different juvenile 5 min after having been exposed to a first juvenile, and their ability to recognize the first juvenile was tested 20 min later.

326 R.M. BLUTH~ et al.
Effects o f A VP on Social Recognition To assess the ability of AVP to improve social recognition (Dantzer et al., 1987), male
mice (n = 8) were submitted to two presentations of the same or different juveniles with a 120-min intervening interval. AVP or saline was injected immediately after the first exposure.
Effects o f AA VP on Social Recognition To assess the ability of the V 1 receptor antagonist to impair social recognition (Dantzer
et al., 1987; Bluth6 et al., 1990), male mice (n = 14) were submitted to two presentations of the same juvenile with a 20-min intervening interval. AAVP or saline was injected immediately after the first exposure.
Effects o f Castration on Social Recognition To assess the effects of castration on the temporal characteristics of social recognition,
and its sensitivity to the VI receptor antagonist, 14 male mice were castrated and allowed 2 weeks to recover. To assess their ability to recognize previously presented juveniles, castrated mice were first submitted to two presentations of the same juvenile with a 20 min intervening interval. This was followed by two presentations of the same juvenile with a 20, 60, 120, or 180 min intervening interval. A different interval was tested every day in a random order.
To assess their sensitivity to the V1 receptor antagonist, the same mice were submitted to two presentations of the same juvenile with a 20-min intervening interval. AAVP or saline was injected immediately after the first exposure. This experiment began 3 weeks after surgery. Intact male mice (n = 14) were used as a positive control group.
Statistics Investigation times were analysed with standard 2- or 3-way ANOVA with repeated
measurements on the time and the treatment factors. Comparisons of individual group means were carried out using the least significant difference test.
RESULTS
Time and Stimulus Dependency o f Social Recognition in Intact Male Mice Means and standard errors of social investigation times are illustrated in Fig. 1. A two-
way ANOVA with repeated measurements (2 exposures × 4 intervals) on the duration of investigation of the juvenile by intact male mice revealed a significant effect of both factors [F(1, 10) = 27.17, p < .001 and F(3, 30) = 5.46, p < .01, respectively] and their interaction [F(3, 30) = 24.68, p < .001]. Mean investigation times were significantly lower when the second exposure to the juvenile took place 20 min (p < .001) or 60 rain (p < .001), but not 120 or 180 min after the first exposure. These results indicate that intact male mice explored less a previously presented juvenile conspecific which had been presented 20 or 60 min earlier but not 2 or 3 hr earlier.
To determine whether this decrease in investigation time is specific to a familiar juvenile, mice were presented with a familiar or an unfamiliar juvenile 20 rain after the initial exposure. Means and standard errors of social investigation times are presented in the right part of Fig. 1. A two-way ANOVA with repeated measurements (2 exposures × 2 juveniles) on the duration of investigation of the juvenile by intact mice revealed a significant effect of both factors IF(l , 10) = 28.95, p < .001 and F(1, 10) = 13.28, p <

SOCIAL RECOGNITION AND AVP IN MICE 327
¢..)
09
t - O
. , I - , , ,*
03
09
t -
' , , , I , - -
0
E I - -
150
100
50_
O_
150_
,0o t 5O
o 20 60 120 180 20 20
IEI (min) IEI (min)
FIG. 1: Left panel: Effect of different inter-exposure intervals (IEI) on social recognition in intact male mice. Each bar represents the mean ( - SEM) investigation time by adult mice (n = 11) on the first exposure (white columns) and the second exposure (black columns) to an identical juvenile (***p < .001 compared with first exposure to the juvenile). Right panel: Specificity of social recognition in intact male mice (n = 11) exposed to the same juvenile (black column) or to a different juvenile (hatched column) on the recognition test (***p < .001 compared with first exposure to the juvenile).
.01, respectively] and their interaction [F(1, 10) = 34.31, p < .001]. Mean investigation times on the second exposure decreased only when a familiar juvenile was presented (p < .001). These results indicate that the decrease in investigation time observed when the same juvenile was presented 20 min after the initial exposure is specific to this particular juvenile.
Facilitation and Inhibition of Social Recognition Means and standard errors of social investigation times are illustrated in Fig. 2. Mice
submitted to two successive presentations of the same juvenile at a 30-min interval, followed by a third presentation of the same juvenile 120 min after the initial presentation, displayed a marked decrease in investigation time of the previously presented juvenile [F(2, 15) = 22.62, p < .001]. However , when mice were presented with a different juvenile 5 min after the initial presentation, investigation time of the originally presented juvenile remained the same as on the first presentation IF(2, 14) = 2.35, p > .05]. These results indicate that repeated exposure to the same juvenile was sufficient to prolong the duration of recognition of this juvenile, whereas exposure to another juvenile interfered with subsequent recognition of the first juvenile.
Effects of A VP on Social Recognition Means and standard errors of social investigation times during the first and second
exposures to a familiar or to a different juvenile in mice treated with saline (SAL) or AVP immediately after the first presentation are illustrated in Fig. 3. In mice reexposed 120 rain later to the same juvenile, a two-way ANOVA (2 exposures x 2 treatments) on investigation times revealed a significant effect of both factors and their interaction [F(1, 7) = 51.15, p < .001; F(1, 7) = 10.06, p < .05; and F(1,7) = 13.95, p < .01, respectively].

328 R.M. BLUTH~ et al.
o 160.
12C .o
= 8C > f-
° ~ ~- 4C. O
._E 0 ~ _ -
"r
160 -
120 I ~
* * 80. * "g-
H 40.
0 . - - 0 30 120
IEI (min) 0 IEI
½ h " / / / / / / / /
/ i
~4
",4, % 15 30 (min)
FIG. 2: Left panel: Facilitation of social recognition. The same juvenile was presented twice at 30- and 120-min intervals to intact male mice (n = 6) following the first exposure (**p < .01; ***p < .001 compared to time 0). Right panel: Impairment of social recognition. The same juvenile was presented twice to intact male mice (n = 8) at 30-min interval, interrupted by a 5-min exposure to a different juvenile. White columns represent investigation time of the original juvenile, black columns investigation time of the same juvenile on the 2nd exposure test, and hatched columns investigation time of the different juvenile.
Mean investigation times were significantly lower during the second exposure to the same juvenile in AVP- but not in saline-treated mice (p < .001). In mice reexposed to a different juvenile, a two-way ANOVA (2 exposures × 2 treatments) on investigation times revealed no effect of exposure or treatment and their interaction in AVP-treated mice (p > . 10). These results indicate that AVP enhanced the recognition of a familiar juvenile but did not interfer with the recognition of a novel juvenile.
O ,~ SAL AVP SAL AVP 150
-- iii
--- iii 111 i11 i11 i11
100 " ' " ._ //. ;-;
~ I l l / , i l l ~ i / / # i l l
i l l l l i > ~/½ / z /
t - y /
- i "
i i i / . . . i "6 50 . . . . . .
111 .,../ I~ i.i il/
""" i l l
• -- ~
ii/
I.- 0 120 120 120 20
IEI (rain) FIG. 3: Effects of AVP on social recognition. Means (± SEM) social investigation times by adult male mice on first and second exposure to the same juvenile (left panel) or to a different juvenile (right panel). In each case, AVP (0.4 ~g/mouse) or saline was injected immediately after the first exposure (***p < .001 compared with time 0).
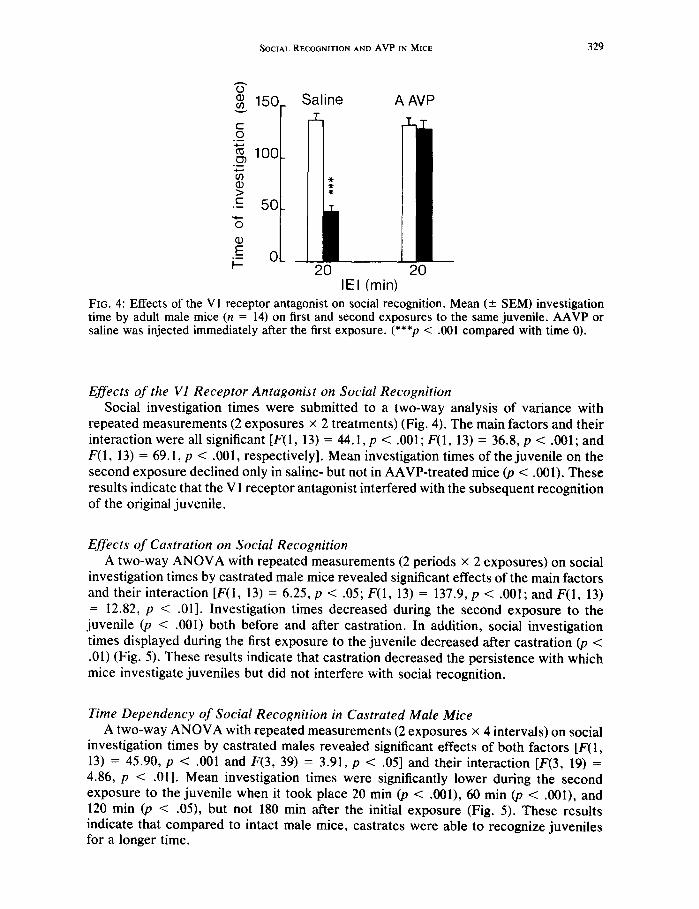
SOCIAL RECOGNITION AND A V P IN MICE 329
O (1.)
v
150 Saline A AVP
c O . m
100
• ~ 50 O
E 0 20 20
IEI (min) FIG. 4: Effects of the VI receptor antagonist on social recognition. Mean (___ SEM) investigation time by adult male mice (n = 14) on first and second exposures to the same juvenile. AAVP or saline was injected immediately after the first exposure. (***p < .001 compared with time 0).
Effects of the V1 Receptor Antagonist on Social Recognition Social investigation times were submitted to a two-way analysis of variance with
repeated measurements (2 exposures × 2 treatments) (Fig. 4). The main factors and their interaction were all significant [F(1, 13) = 44.1, p < .001; F(1, 13) = 36.8, p < .001; and F(1, 13) = 69. I, p < .001, respectively]. Mean investigation times of the juvenile on the second exposure declined only in saline- but not in AAVP-treated mice (p < .001). These results indicate that the V 1 receptor antagonist interfered with the subsequent recognition of the original juvenile.
Effects o f Castration on Social Recognition A two-way ANOVA with repeated measurements (2 periods × 2 exposures) on social
investigation times by castrated male mice revealed significant effects of the main factors and their interaction [F(1, 13) = 6.25, p < .05; F(1, 13) = 137.9, p < .001; and F(1, 13) = 12.82, p < .01]. Investigation times decreased during the second exposure to the
juvenile (p < .001) both before and after castration. In addition, social investigation times displayed during the first exposure to the juvenile decreased after castration (p < .01) (Fig. 5). These results indicate that castration decreased the persistence with which mice investigate juveniles but did not interfere with social recognition.
Time Dependency of Social Recognition in Castrated Male Mice A two-way ANOVA with repeated measurements (2 exposures × 4 intervals) on social
investigation times by castrated males revealed significant effects of both factors [F(1, 13) = 45.90, p < .001 and F(3, 39) = 3.91, p < .05] and their interaction [F(3, 19) = 4.86, p < .01]. Mean investigation times were significantly lower during the second exposure to the juvenile when it took place 20 rain (/9 < .001), 60 min (p < .001), and 120 min (p < .05), but not 180 min after the initial exposure (Fig. 5). These results indicate that compared to intact male mice, castrates were able to recognize juveniles for a longer time.

330 R.M. BLUTHI~ et al.
t IE~_2?_~in /
I1! i
Bef Aft 20 60 120 180 castration IEI (min)
FIG. 5: Left panel: Effects of castration on social recognition in male mice. Mean (-+ SEM) investigation time on the first and second exposures to the same juvenile, before and after castration (***p < .001 compared with first exposure to the juvenile). Right panel: Time-effect dependency of social recognition in castrated male mice. Same conventions as in Fig. 1 (*p < .05; ***p < .001 compared with first exposure to the juvenile).
Sensitivity of Castrated Mice to the VI Receptor Antagonist In intact male mice (Fig. 6), a two-way A N O V A (2 exposures x 2 t reatments) on
investigation times revealed a significant effect of the main factors and their interaction [F(1, 13) = 23.40, p < .001; F(1, 13) = 7.24, p < .05; and F(1, 13) = 13.30, p < .01, respectively]. Investigation times decreased in saline- but not in AAVP-t rea ted male mice exposed for the second time to the same juvenile (p < .001).
In castra ted male mice, a two-way A N O V A (2 exposures x 2 t reatments) on investiga- tion times revealed a significant effect of the exposure factor, but no effect of the t rea tment factor and no interaction IF( l , 13) = 26.2, p < .001; F(1, 13) = 1.11, NS; F(1, 13) = 0.52, NS, respectively].
- - - 1 5 0 . o
t--
.o 100.
> 5 0 ¢-
o
E 0 b-
FIG. 6: Effects of conventions as in
Intact Castrated
I] 20 20 20 20 IEI (min)
SAL AAVP SAL AAVP Treatment the Vl receptor antagonist on social recognition in castrated male mice. Same Fig. 4 (**p < .01 ; ***p < .001 compared with first exposure to the juvenile).

SOCIAL RECOGNITION AND AVP IN MICE 331
These results indicate that administration of the V 1 receptor antagonist impaired social recognition in normal male mice, but was ineffective in castrated mice.
DISCUSSION
The present findings show that male mice of the DBA strain spent longer investigating new juvenile conspecifics than familiar ones. These differences can be used to infer the duration during which they are able to recognize juveniles. Social recognition is based on some form of memory of the stimulus characteristics of the juvenile since it was enhanced by two successive presentations of the same juvenile and blocked by presenta- tion of a novel juvenile in the interval between two presentations of the same juvenile. Social recognition was enhanced by administration of vasopressin and blocked by admin- istration of an antagonist of the vasopressor receptors of vasopressin. This last effect is modulated by androgens since it was abrogated by castration.
The ability of mice to discriminate the social characteristics of conspecifics has been mainly studied in the context of sexual behaviour and aggression and chemosensory cues have been identified as the primary stimuli that permit such discriminations. For example, Dixon and Mackintosh (1975) have demonstrated that urine from female mice has aggres- sion-reducing properties for male mice and that urine from estrous mice elicits greater sexual responding in males than does urine from diestrous females. However, only a few experiments indicate that besides their signalling properties, chemoceptive cues can be used for social recognition. The first evidence in favor of this possibility came from the work of Bruce and Parrott (1960) on the blockade of pregnancy by exposure of pregnant females to unfamiliar males. It is now apparent that this phenomenon represents a protective mechanism by which pregnant females are made insensitive to the estrus- promoting properties of the odor of the stud male (Keverne & de la Riva, 1982). It is based on the short-term memory of the odor of the stud male, that the female forms at the time of copulation. Besides the Bruce effect, other evidence in favor of the ability of mice to discriminate between conspecifics of the same strain is based on learning techniques in which animals have to make use of the odor of conspecifics as a cue to choose the correct goal arm in an olfactory discrimination apparatus (e.g., Bowers & Alexander, 1967).
Our findings indicate that mice have the capacity to make use of conspecifics' character- istics in a more natural context and that they are actually able to form a memory of conspecifics in a very short amount of time. The ability of an animal to recognize familiar from novel conspecifics is certainly of great adaptive value because it assists the individual in regulating social interactions. Although we have not tried to determine the sensory nature of the stimuli used for social recognition, it is likely to be based on olfaction, because adult male mice spend most of the interaction time sniffing the anogenital area of the juvenile. In rats, the decreased persistence with which adult males investigate a juvenile conspecific was significantly reduced following preexposure not only to the juvenile but also to chemosensory stimuli (soiled bedding or urine) from that juvenile (Sawyer et al . , 1984). In addition, social recognition was blocked by olfactory bulbectomy (Dantzer et al . , 1990).
Social recognition appears to hold longer in mice than in rats since it is still present after 1 hr in mice, whereas it lasts for less than 1 hr in rats (Dantzer et al . , 1987; Sekiguchi et al . , 1991). However, in the absence of any systematic investigation of the temporal characteristics of social recognition in different strains of mice and rats, it is premature

332 R.M. BLUTHt~ et al.
to conclude that this is a between-species difference. From this perspective, it would be interesting to assess social recognition in strains of mice and rats with different learning abilities.
The persistence with which male mice investigate a juvenile conspecific is modulated not only by familiarity with this particular juvenile but also by levels of sex steroids. Castra- tion of adult males resulted in a marked decrease in social investigation time. The same phenomenon has also been observed in rats (Sawyer e t al . , 1984; Bluth6 e t al . , 1990). Al- though the decreased investigation time of castrates is consistent with several earlier studies that reported that the castration of male rats decreased their preference for estrous odors (Brown, 1977; Carr et al . , 1965; Le Magnen, 1952), it is important to note that this effect was specific to sexually experienced males (Brown, 1977; Carr e t al . , 1965; Lydell & Doty, 1972; Stern, 1970). Since the mice used in the present experiment were sexually naive, the influence of androgens on odor preferences is unlikely to account for the longer social investigation times displayed by intact males. In rats, a systematic study of investigation times of novel juveniles by intact and castrated males from 40 to 80 days of age revealed the existence of significant differences between the two groups from day 50 (Thor et al . , 1982). The prolongation of investigation times by androgens has been interpreted as another example of the persistence effect of these hormones. According to this interpretation, ani- mals with high testosterone levels are more persistent in their search pattern and less dis- tracted by irrelevant stimuli than animals with low testosterone levels (Andrew, 1972, 1976; Archer, 1977). The generality of this phenomenon across different species and testing pro- cedures makes unlikely an explanation in terms of differential sensitivity to odors. The fact that in mice as well as in rats social recognition is better in castrates than in intact male mice despite shorter investigation times in the former animals appears to be paradoxical, especially in view of earlier studies suggesting that castration can impair learning. For exam- ple, although castration did not impair odor detection performance of male rats to ethyl acetate, as measured by high precision olfactometry and ago-no go signal detection task, the ability to improve detection performance over time was reduced (Doty & Ferguson-Segall, 1989). However, in a direct test of possible influences of sex hormones on memory, van Hest e t al. (1988) were unable to find any difference in the response accuracy of intact versus gonadectomized male rats in an operant delayed spatial response alternation procedure as a function of the duration of delay interval, despite a more rapid acquisition of the task by intact males.
An important clue for understanding how androgens affect social recognition is the identification of the specific neuronal systems which are regulated by sex steroids. One of the key candidates for this function is represented by the sexually dimorphic vasopres- sinergic neurons which, in rats and mice, project from the bed nucleus of the stria terminalis to the septum. In both species, the density of AVP-immunoreactive fibers in the lateral septum is higher in males than in females. It is reduced by castration and restored to normal following testosterone replacement therapy (De Vries et al . , 1984; Mayes et al . , 1988). Mice injected with AVP immediately after the first exposure to the juvenile displayed a decreased investigation of this juvenile when it was presented 2 hr later. This was not due to any residual toxic effect of the peptide treatment, because there was no decrease in the investigation of a novel juvenile by mice injected 2 hr earlier with the same dose of AVP. These findings suggest that vasopressin enhances social recognition in mice in the same manner as in rats (Dantzer et al . , 1987). However, a better demonstration of the physiological involvement of AVP in social recognition was the observation that administration of the V1 receptor antagonist immediately after the

SOCIAL RECOGNITION AND AVP 1N MICE 333
first exposure to the juvenile made mice behave as if they no longer recognized this particular juvenile. Although the antagonist peptide was injected SC, its effects are likely to be mediated centrally, as this particular peptide crosses the blood-brain barrier and is active in the same manner when injected directly into the lateral ventricle of the brain (Le Moal et al., 1987; Bluth6 & Dantzer, 1992) or into the lateral septum (Dantzer et al. , 1988).
Administration of the V1 receptor antagonist to castrated male mice 3 weeks after gonadectomy was no longer effective in impairing social recognition. This might corre- spond to the loss of functional vasopressin following castration in the AVP-containing neurons, which project from the BNST to the lateral septum. However, the time course of disappearance of AVP immunoreactivity following castration appears to be extremely slow because 8-12 weeks are needed for the complete disappearance of AVP-immunore- active fibers in the septum of rats (De Vries et al., 1984). The same appears to be true in mice since there were no changes in AVP-related neurophysin II in the lateral habenula after 2 weeks, a noticeable decrease in the intensity of immunoreactivity at 4 weeks, and a complete disappearance of immunoreactivity at 15 weeks (Mayes et al. , 1988). It could, therefore, be argued that a time interval of 2-3 weeks after castration is too short to demonstrate effects ofgonadectomy on androgen-dependent vasopressinergic neurons. However, using in situ hybridization techniques, Miller et al. (1992) observed that AVP gene expression declined much more rapidly than AVP immunoreactivity, since both the total number of labelled cells and the average number of grains per cell were significantly reduced in the BNST by I week postcastration. In addition, in previous experiments with rats we have been unable to observe any difference in the sensitivity of castrated males to the V1 receptor antagonist, regardless of whether the animals were tested 3 weeks or more than 5 weeks following castration (Bluth6 et al. , 1990). Therefore, our findings suggest that the lost of functional activity of androgen dependent AVP neurons occurs earlier than the disappearance of AVP immunoreactivity.
In conclusion, the demonstration of the behavioral activity of the V1 receptor antago- nist on social recognition in male mice is more than just a replication of our previous work in rats (Dantzer et al. , 1987). There are not many observations showing that blockade of vasopressinergic transmission by antagonists acting at the level of vasopres- sor receptors has functional consequences. In addition, because of the commonalities that exist between mice and rats in the extrahypothalamic distribution of vasopressin (De Vries et al. , 1984; Mayes et al. , 1988), the finding that the involvement of vasopressin in social recognition is modulated by sex steroids in mice as well as in rats allows more confidence to be put on the generality of this phenomenon and its physiological importance.
Acknowledgments: We gratefully acknowledge Dr. Jean Rivier (The Clayton Foundation Laboratories For Peptide Biology, The Salk Institute, San Diego), for providing the antagonist of vasopressor-like receptors of vasopressin used in these studies. We also thank Sandra Suarez for her assistance with behavioural observations and Dr. Quentin Pittman for reviewing the manuscript.
REFERENCES
Andrew RJ (1972) Recognition processes and behavior with special reference to effects of testoster- one on persistence. In: Lehrman DS, Hinde RA, Shaw E (Eds) Advances in the Study o f Behavior, Vol 4. Academic Press, New York.
Andrew RJ (1976) Attentional processes and animal behaviour. In: Hinde RA, Bateson PPG (Eds) Growing Points in Ethology. Cambridge University Press, Cambridge, U.K.

334 R.M. BLUTHf ~- et al.
Archer J (1977) Testosterone and persistence in mice. Anim Behav 25:479-488. Bluth6 RM, Dantzer R (1990) Social memory does not involve vasopressinergic neurotransmission
in female rats. Brain Res 535:301-304. Bluth6 RM, Dantzer R (1992) Chronic intracerebral infusions of vasopressin and vasopressin
antagonist modulate social recognition in rat. Brain Res 572:261-264. Bluth6 RM, Parnet P, Dantzer R, Kelley KW (1991) Interleukin-1 receptor antagonist blocks effects
of IL- 1 a and IL- lfl on social behaviour and body weight in mice. Neurosci Res Comm 9:151-158. Bluth6 RM, Schoenen J, Dantzer R (1990) Androgen-dependent vasopressinergic neurons are
involved in social recognition in rats. Brain Res 519:150-157. Bowers JM, Alexander BK (1967) Mice: Individual recognition by olfactory cues. Science
158:1208-1210. Brain P (1975) What does individual housing mean to a mouse? Life Sci 16:187-200. Brown RE (1977) Odor preference and urine-marking scales in male and female rats: Effects of
gonadectomy and sexual experience on responses to conspecific odors. J Comp Physiol Psychol 91:1190-1206.
Bruce HM, Parrott DMV (1960) Role of olfactory sense in pregnancy block by strange males. Science 131:1526.
Carr WJ, Loeb LS, Dissinger ML (1965) Responses of rats to sex odors. J Comp Physiol Psychol 59:370-377.
Dantzer R, Bluth6 RM, Koob GF, Le Moal M (1987) Modulation of social memory in male rats by neurohypophyseal peptides. Psychopharmacol 91:363-368.
Dantzer R, Koob GF, Bluth6 RM, Le Moal M (1988) Septal vasopressin modulates social memory in male rats. Brain Res 457:143-147.
Dantzer R, Tazi A, Bluth6 RM (1990) Cerebral lateralization of olfactory-mediated affective pro- cesses in rats. Behav Brain Res 40:53-60.
Dixon AK, Mackintosh JH (1975) The relationship between the physiological condition of female mice and the effects of their urine on the social behavior of adult males. Anim Behav 19:153-155.
De Vries GJ, Buijs RM, Sluiter AA (1984) Gonadal hormone actions on the morphology of the vasopressinergic innervation of the adult rat brain. Brain Res 298:141-145.
De Vries GJ, Buijs RM, Swaab DF (1981) Ontogeny of the vasopressinergic neurons of the suprachiasmatic nucleus and their extrahypothalamic projections in the rat brain. Presence of a sex difference in the lateral septum. Brain Res 218:67-78.
De Vries GJ, Duetz W, Buijs RM, Van Heerikhuize J, Vreeburg JTM (1986) Effects of androgens and estrogens on the vasopressin and oxytocin innervation of the adult rat brain. Brain Res 399:296-302.
Doty RL, Ferguson-Segall M (1989) Influence of adult castration on the olfactory sensitivity of the male rat: A signal detection analysis. Behav Neurosci 103:691-694.
Keverne EB, De la Riva C (1982) Pheromones in mice: Reciprocal interaction between the nose and brain. Nature 296:148-150.
Le Magnen J (1952) Les ph6nom6nes olfacto-sexuels chez le rat blanc. Arch Sci Physiol 6:295-331. Le Moal M, Dantzer R, Michaud B, Koob GF (1987) Centrally injected arginine vasopressin (AVP)
facilitates social memory in rats. Neurosci Lett 77:353-359. Le Moal M, Koob GF, Koda LY, Bloom FE Manning M, Sawyer WH, Rivier J (1981) Vasopressin
antagonist peptide: Blockade of pressor receptor prevents behavioral action of vasopressin. Nature 291:491-493.
Lydell K, Doty RL (1972) Male rat odor preferences for female urine as a function of sexual experience, urine age and urine source. Hormone Behav 3:205-212.
Manning M, Sawyer WH (1984) Design and uses of selective agonistic and antagonistic analogs of the neuropeptides oxytocin and vasopressin. Trends Neurosci 7:6-9.
Mayes CR, Watts AG, McQueen JK, Fink G, Charlton HM (1988) Gonadal steroids influence neurophysin II distribution in the forebrain of normal and mutant mice. Neurosci 25:1013-1022.
Miller MA, De Vries G J, AI-Shamma HA, Dorsa DM (1992) Decline of vasopressin immunoreactiv- ity and mRNA levels in the bed nucleus of the stria terminalis following castration. J Neurosci 12:2881-2887.
Miller MA, Urban JH, Dorsa DM (1989a) Steroid dependency of vasopressin neurons in the bed nucleus of the stria terminalis by in situ hybridization. Endocrinology 125:2335-2340.
Miller MA, Vician L, Clifton DK, Dorsa DM (1989b) Sex differences in vasopressin neurons in the bed nucleus of the stria terminalis by in situ hybridization. Peptides 10:615-619.

SOCIAL RECOGNITION AND AVP IN MICE 335
Sawyer TF, Hengehold AK, Perez WA (1984) Chemosensory and hormonal mediation of social memory in male rats. Behav Neurosci 98:908-913.
Sekiguchi R, Wolterink G, Van Ree JM (1991) Short duration of retroactive facilitation of social recognition in rats. Physiol Behav 50:1253-1256.
Siegfried B, Frischknecht H-R, Waser PG (1984) Vasopresin impairs or enhances retention of learned submissive behavior in mice depending on the time of application. Behav Brain Res 11:259-269.
Stern JJ (1970) Responses of male rats to sex odors. Physiol Behav 5:519-524. Thor DH, Holloway WR (1982) Social memory of the male laboratory rat. J Comp Physiol Psychol
96:1000-1006. Thor DH, Wainwright KL, Holloway WR (1982) Persistence of attention to a novel conspecific:
Some development variables in laboratory rats. Dev Psychobiol 15:1-8. van Hest A, van Kempen M, van Haaren F, van de Poll NE (1988) Memory in male and female
Wistar rats: Effects ofgonadectomy and stimulus presentations during the delay interval. Behav Brain Res 29:103-110.




















