
Naturally occurring variations in defensive burying behavior are associated with differences in...
12
Naturally occurring variations in defensive burying behavior are associated with differences in vasopressin, oxytocin, and androgen receptors in the male rat Ian Linfoot a , Megan Gray a , Brenda Bingham a , Martin Williamson a , John P.J. Pinel b , Victor Viau a, ⁎ a Department of Cellular and Physiological Sciences, University of British Columbia, Vancouver, BC, Canada V6T 1Z3 b Department of Psychology, University of British Columbia, Vancouver, BC, Canada V6T 1Z3 abstract article info Article history: Received 2 February 2009 Received in revised form 10 June 2009 Accepted 10 June 2009 Available online 18 June 2009 Keywords: Bed nuclei of the stria terminalis Defensive burying Medial amygdale Oxytocin Paraventricular nucleus of the hypothalamus Vasopressin Largely ignored in tests of defensive burying is the capacity for individual animals to display marked variations in active coping behaviors. To expose the neurobiological correlates of this behavioral differ- entiation rats were exposed to a mousetrap that was remotely triggered upon approach to remove the quality of pain. Relative to animals showing no significant levels of defensive burying activity, rats showing sustained elevations in defensive burying displayed higher levels of arginine vasopressin (AVP) mRNA and increased numbers of androgen receptor positive cells in the medial amygdala and posterior bed nuclei of the stria terminalis, brain regions that integrate emotional appraisal and sensory information. In contrast, animals showing little to no defensive burying responses displayed relatively higher levels of AVP and oxytocin (OT) mRNA within the supraoptic nucleus and subregions of the paraventricular nucleus of the hypothalamus responsible for neuroendocrine and autonomic function. Finally, animals showing sustained levels of burying also displayed increased levels of testosterone and pituitary-adrenal hormones under stress conditions. These findings implicate roles for central AVP and OT in mediating differential avoidance behaviors and demonstrate the utility of using a pain-free test of defensive burying as a framework for exploring naturally occurring differences in coping style and neuroendocrine capacity. © 2009 Elsevier Inc. All rights reserved. 1. Introduction Modeling the basis for individual differences in stress-induced psychopathology or recovery is difficult on both practical and theo- retical grounds because normal subjects are used. Thus, there exists a real need to tackle this problem in animal models of experimentally induced or innate differences in psychological state. Several emerging models indicate that differences in coping style provide a strong basis for determining potential vulnerability to stress-related disorders. Thus, rodents exhibiting spontaneous individual differences in novelty seeking behavior (Kabbaj et al., 2000; Walker et al., 2008), behavioral inhibition in response to predator odor (Roseboom et al., 2007), or genetically selected for variations in anxiety traits (Landgraf and Wigger, 2002) and aggressive behavior (Koolhaas et al., 1999), show remarkable differences in anxiety- and depressive-like beha- viors and predisposition to drug self administration (Taghzouti et al., 1999; Yilmazer-Hanke et al., 2004; Kabbaj et al., 2007). Since its introduction, the shock prod defensive burying test has proven incredibly reliable and informative with respect to determin- ing the neurobiological properties of avoidance, fear, and anxiety- related behavior. First noted by Hudson (1950), and extensively developed by Pinel and colleagues (Pinel and Treit, 1978; Treit et al., 1981), rodents show a remarkable propensity for using available bedding material to bury a variety of noxious stimuli. What has emerged from some of these studies, however largely ignored, is that animals have the capacity to enter the experimental environment with different tendencies to bury as a conditioned response. In contrast to rats, for example, hamsters and gerbils do not display significant levels of conditioned defensive burying following shock prod exposure (Treit et al., 1980b). Rats selected for differences in aggressive behav- ior also display extreme variations in shock-induced defensive burying activity (De Boer and Koolhaas, 2003). While these studies underscore how social, environmental, and evolutionary pressures can direct the capacity of individual behavioral responses to challenging situations, the basis for this phenomenon is not understood. The regulation and influence of individual differences in coping styles on stress adaptation may be mediated by multiple neuropeptide systems and in brain regions implicated in fear and anxiety in both experimental animals and in humans, and may thus be important for coping style. However, only a few of these systems have been system- atically linked to the formation of individual behavioral character- istics, including in animals selectively bred for avoidance and anxiety behaviors or selected for differences in exploratory and aggressive Progress in Neuro-Psychopharmacology & Biological Psychiatry 33 (2009) 1129–1140 Abbreviations: ACTH, adrenocorticotropic hormone; AR, androgen receptor; AVP, arginine vasopressin; BST, bed nuclei of the stria terminalis; CRH, corticotropin releasing hormone; OT, oxytocin; MeA, medial nucleus of the amygdala; PVN, paraventricular nucleus of the hypothalamus. ⁎ Corresponding author. Department of Cellular and Physiological Sciences, Life Sciences Centre, The University of British Columbia, 2350 Health Sciences Mall, Vancouver, BC, Canada V6T 1Z3. Tel.: +1 604 822 3899; fax: +1 604 822 2316. E-mail address: [email protected] (V. Viau). 0278-5846/$ – see front matter © 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.pnpbp.2009.06.008 Contents lists available at ScienceDirect Progress in Neuro-Psychopharmacology & Biological Psychiatry journal homepage: www.elsevier.com/locate/pnp
Transcript of Naturally occurring variations in defensive burying behavior are associated with differences in...

Progress in Neuro-Psychopharmacology & Biological Psychiatry 33 (2009) 1129–1140
Contents lists available at ScienceDirect
Progress in Neuro-Psychopharmacology & BiologicalPsychiatry
j ourna l homepage: www.e lsev ie r.com/ locate /pnp
Naturally occurring variations in defensive burying behavior are associated withdifferences in vasopressin, oxytocin, and androgen receptors in the male rat
Ian Linfoot a, Megan Gray a, Brenda Bingham a, Martin Williamson a, John P.J. Pinel b, Victor Viau a,⁎a Department of Cellular and Physiological Sciences, University of British Columbia, Vancouver, BC, Canada V6T 1Z3b Department of Psychology, University of British Columbia, Vancouver, BC, Canada V6T 1Z3
Abbreviations: ACTH, adrenocorticotropic hormonearginine vasopressin; BST, bed nuclei of the stria treleasing hormone; OT, oxytocin; MeA, medial nucparaventricular nucleus of the hypothalamus.⁎ Corresponding author. Department of Cellular an
Sciences Centre, The University of British ColumbiaVancouver, BC, Canada V6T 1Z3. Tel.: +1 604 822 3899;
E-mail address: [email protected] (V. Viau).
0278-5846/$ – see front matter © 2009 Elsevier Inc. Aldoi:10.1016/j.pnpbp.2009.06.008
a b s t r a c t
a r t i c l e i n f oArticle history:Received 2 February 2009Received in revised form 10 June 2009Accepted 10 June 2009Available online 18 June 2009
Keywords:Bed nuclei of the stria terminalisDefensive buryingMedial amygdaleOxytocinParaventricular nucleus of the hypothalamusVasopressin
Largely ignored in tests of defensive burying is the capacity for individual animals to display markedvariations in active coping behaviors. To expose the neurobiological correlates of this behavioral differ-entiation rats were exposed to a mousetrap that was remotely triggered upon approach to remove the qualityof pain. Relative to animals showing no significant levels of defensive burying activity, rats showing sustainedelevations in defensive burying displayed higher levels of arginine vasopressin (AVP) mRNA and increasednumbers of androgen receptor positive cells in the medial amygdala and posterior bed nuclei of the striaterminalis, brain regions that integrate emotional appraisal and sensory information. In contrast, animalsshowing little to no defensive burying responses displayed relatively higher levels of AVP and oxytocin (OT)mRNA within the supraoptic nucleus and subregions of the paraventricular nucleus of the hypothalamusresponsible for neuroendocrine and autonomic function. Finally, animals showing sustained levels of buryingalso displayed increased levels of testosterone and pituitary-adrenal hormones under stress conditions.These findings implicate roles for central AVP and OT in mediating differential avoidance behaviors anddemonstrate the utility of using a pain-free test of defensive burying as a framework for exploring naturallyoccurring differences in coping style and neuroendocrine capacity.
© 2009 Elsevier Inc. All rights reserved.
1. Introduction
Modeling the basis for individual differences in stress-inducedpsychopathology or recovery is difficult on both practical and theo-retical grounds because normal subjects are used. Thus, there exists areal need to tackle this problem in animal models of experimentallyinduced or innate differences in psychological state. Several emergingmodels indicate that differences in coping style provide a strong basisfor determining potential vulnerability to stress-related disorders.Thus, rodents exhibiting spontaneous individual differences innovelty seeking behavior (Kabbaj et al., 2000; Walker et al., 2008),behavioral inhibition in response to predator odor (Roseboom et al.,2007), or genetically selected for variations in anxiety traits (Landgrafand Wigger, 2002) and aggressive behavior (Koolhaas et al., 1999),show remarkable differences in anxiety- and depressive-like beha-viors and predisposition to drug self administration (Taghzouti et al.,1999; Yilmazer-Hanke et al., 2004; Kabbaj et al., 2007).
; AR, androgen receptor; AVP,erminalis; CRH, corticotropinleus of the amygdala; PVN,
d Physiological Sciences, Life, 2350 Health Sciences Mall,fax: +1 604 822 2316.
l rights reserved.
Since its introduction, the shock prod defensive burying test hasproven incredibly reliable and informative with respect to determin-ing the neurobiological properties of avoidance, fear, and anxiety-related behavior. First noted by Hudson (1950), and extensivelydeveloped by Pinel and colleagues (Pinel and Treit, 1978; Treit et al.,1981), rodents show a remarkable propensity for using availablebedding material to bury a variety of noxious stimuli. What hasemerged from some of these studies, however largely ignored, is thatanimals have the capacity to enter the experimental environmentwith different tendencies to buryas a conditioned response. In contrastto rats, for example, hamsters and gerbils do not display significantlevels of conditioned defensive burying following shock prod exposure(Treit et al., 1980b). Rats selected for differences in aggressive behav-ior also display extreme variations in shock-induced defensive buryingactivity (De Boer and Koolhaas, 2003).While these studies underscorehow social, environmental, and evolutionary pressures can direct thecapacity of individual behavioral responses to challenging situations,the basis for this phenomenon is not understood.
The regulation and influence of individual differences in copingstyles on stress adaptationmay bemediated bymultiple neuropeptidesystems and in brain regions implicated in fear and anxiety in bothexperimental animals and in humans, and may thus be important forcoping style. However, only a few of these systems have been system-atically linked to the formation of individual behavioral character-istics, including in animals selectively bred for avoidance and anxietybehaviors or selected for differences in exploratory and aggressive

1130 I. Linfoot et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 33 (2009) 1129–1140
behaviors (Reviewed in Pawlak et al., 2008, and see Roozendaal et al.,1992; Wiersma et al., 1997; Koolhaas et al., 1998; Menard and Treit,1999; Landgraf and Wigger, 2002; Steimer and Driscoll, 2003; Kabbajet al., 2007; Veenema and Neumann, 2007). Foremost among theseneuropeptide systems include corticotropin releasing hormone (CRH)and arginine vasopressin (AVP) emanating from the amygdala andbed nuclei of the stria terminalis (De Vries and Miller, 1998; Hermanet al., 2003), and forebrain oxytocin (OT) projections originating inthe paraventricular nucleus of the hypothalamus (Sofroniew, 1983;Marroni et al., 2007). In humans and rodents, CRH and AVP maycontribute to extreme trait anxiety and the comorbid expression ofdepression-like symptoms (Keck, 2006; Landgraf, 2006), whereasOT appears to possess anxiolytic properties (de Wied et al., 1993;Neumann et al., 2000; Marroni et al., 2007).
Extrahypothalamic and hypothalamic CRH, AVP, and OT pathwaysrespond to and/or are in a position to regulate the hypothalamic-pituitary-adrenal and -gonadal neuroendocrine axes. It is perhaps notentirely surprising that individual variations in coping styles are alsomet by variations in glucocorticoid and sex steroid hormone levels,both under basal and stress conditions (reviewed in Koolhaas et al.,2007; Pawlak et al., 2008). Thus, variations in corticosterone andtestosterone release in the rodent may be important for the formationof both individual behavioral characteristics and predisposition tostress-related disorders.
In the following studies we developed a pain-free variant of thedefensive burying test aimed at inviting differences in active avoid-ance behaviors. Because anxiety-like, exploration, and defensive bury-ing behaviors sort with differences in CRH, AVP, and OT expressionand activity, we compared the expression levels of these neuropep-tides, as well as adrenal and gonadal status, in animals identified asshowing high and low defensive burying characteristics.
2. Materials and methods
2.1. Animals
Sixty adult male Sprague Dawley rats (UBC Animal Care Centre,Vancouver, Canada) were used, 55–57 days of age on the first day ofbehavioral testing. Animals were pair-housed under controlled tem-perature and lighting conditions (12:12 h light: dark cycle, lights on at0600 h), with food and water available ad libitum. All testing wasperformed during the light phase of the cycle, beginning at 0900 h.The University of British Columbia Animal Care Committee approvedall experimental protocols.
Table 1Behavioral parameters in animals identified as showing High and Low defensiveburying behaviors.
Parameter High- (H) versusLow-bury (L)
Psychological Indexa
Trap reactivity H=L Sensitivity/fear/reactivityStretch attend postures HNL Risk assessmentApproach latency HbL Active avoidanceBury latency HbL Anxiety/reactivityBury duration HNL Anxiety/active avoidanceBury (pile) height HNL Anxiety/active avoidanceFreezing HNL Fear/anxiety/passive avoidanceGrooming HbL ConflictRearing HbL Explorative escapeAmbulation H=L General exploration
a Based on Koolhaas et al. (1999) and De Boer and Koolhaas (2003).
2.2. General treatment schedule
Two days after arrival, rats were weighed and handled on each of7 consecutive days prior to behavioral testing, involving daily removalfrom the home cage room and transport to an adjacent behavioralsuite. The rats were then habituated to the defensive burying chamberfor 30 min on each of 4 consecutive days. During this phase, rats wereexposed to a dummy (unloaded) single spring mousetrap (Victor EasySetMouse TrapModelM035,Woodstream Corporation, Lititz, PA) thatwas bolted to the floor of the chamber to prevent movement whentriggered, at a height of 5 cm. Rats were then exposed to a loadedmousetrap for 15 min on each of 3 consecutive days. Under these testconditions, the trap was remotely triggered by an observer as the ratapproached no closer than approximately 2 cm from the device. Onday 4, the animals were then again exposed to the chamber-trapassembly, introducing an unloaded mousetrap for 15 min as a testof conditioned aversion. 24 h after the initiation of the conditionedaversion test, rats were anesthetized for perfusion and tissuecollection.
2.3. Behavioral testing
The burying test apparatus consisted of an opaque Plexiglaschamber measuring 30×44×44 cm (W×L×H) that was uniformlyfilled with ß wood chip bedding material to a depth of 5 cm. Thechamber was cleaned and fresh bedding was applied between testing.As described above, the apparatus contained a mousetrap that wasunloaded during habituation and a single test of conditioned aversionor remotely triggered during conditioned defensive burying. Habitua-tion and test trial behaviors were recorded using a JVC video camera,initiated as soon as the rat was placed in the chamber, away from themousetrap, and analyzed using the event recorder Hindsight program(v1.5). Acclimatization and behavioral habituation and testing wererestricted to the light phase of the cycle (0900 to1200 h), and theorder of presentation was counterbalanced across animals on eachconsecutive day.
2.4. Behavioral analysis
Several behavioral dimensions were monitored to explore indivi-dual differences in spontaneous and conditioned defensive buryingresponses (see Table 1). These measures included temporal andquantitative assessments (latency, duration, pile height) of defensiveburying, defined as pushing or spraying bedding material directlytowards the trap, trap exploration (approach latency, stretch attendposture), field exploration (rearing, center chamber crossings), andpassive avoidance (freezing, grooming). To accurately gauge theintensity of each rat's immediate reaction to the trap once remotelytriggered, several additional behaviors were analyzed to provideindividual trap-reaction scores. Based on a five-point scale (Gray et al.,1981), trap reactivity scores were: no discernible reaction (score=0);startle, but no immediate withdrawal (score=1); startle and with-drawal to the far end of the chamber (score=2); jumping and/orsquealing followed by rapid withdrawal (score=3); and finally, areflexive jump to the far end of the chamber (score=4).
2.5. Peptide in situ hybridization
To correlate levels of CRH, AVP, and OT mRNA and androgenreceptor immunoreactivity as a function of burying, 24 h after the lastexposure to the defensive burying chamber animals were deeplyanesthetized with a lethal dose of chlorohydrate (35% w/v, 350 mg/kg,intraperitoneal) for perfusion. Deep anesthesia occurredwithin approx-imately 2 min of chloral hydrate administration, as verified by corneal,pedal, and tail-pinch reflexes. Saline (125ml) and 4% paraformaldehydefixative (500 ml) were sequentially perfused via the ascending aorta ata flow rate of 20–25 ml/min. Brains were postfixed for 4 h in fixativeand then cryoprotected overnight in 15% sucrose, 0.1 M potassium

1131I. Linfoot et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 33 (2009) 1129–1140
phosphate-buffered saline (KPBS). Five 1/5 series of frozen coronalsections (30 µm) were collected and stored in antifreeze (30% ethyleneglycol and 20% glycerol in 0.05M sterile KPBS) at−20 C until processingas previously described (Viau and Sawchenko, 2002).
The in situ hybridization approach employed to measure therelative expression levels of neuropeptide transcripts used [33P] UTP-labeled (Amersham, Arlington Heights, IL) antisense cRNA probesencoding for CRH, AVP, and OT. The CRH probe was transcribed from afull-length (1.2-kb) cDNA encoding CRH mRNA (Dr K. Mayo, North-western University, Evanston, IL, USA); the AVP probe from a 230-bpcDNA fragment encoding the vasopressin-specific 3′ end of AVP (Dr. D.Richter, University of Hamburg, Germany); and the OT probe from a190-bp cDNA fragment containing exon C and part of the 2nd intron ofthe rat OT gene (Dr. D. Richter). Techniques for riboprobe synthesisand hybridization are described in extensive detail elsewhere(Simmons et al., 1989; Viau and Sawchenko, 2002). Adjoining seriesof sections were stained with thionin for anatomical referencepurposes. Labeled sense probes did not yield any positive hybridiza-tion signal (data not shown). Based on the strength of the autora-diographic signal (probe and region dependent), exposure time toemulsionwas optimized to ensure that the transcripts of interest werewithin the linear range of the assay and could be quantified bymakingrelative comparisons in optical density (OD) measures in complete,regularly spaced (150 µm) intervals through the cell groups of interest.Relative intensities in hybridization are expressed as mean (±SEM)OD values, determined bilaterally by averaging background-correctedODvalues for the transcriptwithin individual animals (Viauet al., 2001).
2.6. Androgen receptor immunohistochemistry
Based on the group differences in AVP mRNA expression in the BSTand medial amygdala (see Results), and the acknowledged sensitivityof this transcript to androgens (De Vries and Miller, 1998), we thenreasoned that the differences in AVP could be accounted for by dif-ferences in androgen receptors. Androgen receptor (AR) staining wasconducted on the same brains used to assay neuropeptide mRNAlevels, localized in an adjacent series of sections through the posteriordivision of the BST and the medial nucleus of the amygdala using apolyclonal antibody raised in rabbit against amino acids 1–21 ofthe rat androgen receptor (Santa Cruz Biotechnology Inc., SantaCruz, CA) and a conventional nickel-intensified, avidin–biotin-immunoperoxidase procedure (Vectastain Elite ABC kit; VectorLaboratories, Burlington, CA). Tests for immunolabeling specificityand androgen receptor localization are described in greater detailelsewhere (Williamson and Viau, 2007). The number of AR positivecells were derived from cell counts taken in complete, regularly spaced(150 µm) intervals through the posterior BST and medial amygdala,and corrected for double-counting error using Abercrombie's formula(Guillery, 2002). Quantitative data are extrapolated estimates derivedfrom the total corrected number of profiles encountered in each seriesof sections multiplied by the sectioning interval of 5.
Light- and dark-level images were captured using a Retiga 1300 CCDdigital camera (QImaging, Burnaby, BC), cataloged using a MacintoshOS X-driven Open Lab Image Improvision software v 3.0.9 (QuorumTechnologies, Guelph, ON), and exported toAdobePhotoshop (v. 7.0) forfinal assembly at a resolution of 300 dpi. Captured darkfield images ofCRH,AVP, andOTmRNAhybridizationwere exported to Image J (v.1.3.5)for densitometric analysis. AR cell count estimates were determinedusing the Improvision software. Parceling of the rat brain followedthe mapping of AR immunoreactivity and CRH, AVP, and OT mRNAhybridization, as defined by the morphological features provided bythionin staining of adjacent series of tissue, based on the terminology ofSwanson (2004), and of Swanson andKuypers (1980) and Swanson andSimmons (1989) to describe the PVN, and of Dong et al., (2001) andDong and Swanson (2004) to describe the medial amygdala andposterior BST.
2.7. Neuroendocrine function
In a separate experiment, animals were screened and selected fortheir propensity to show stable individual differences in defensiveburying, as described below (Statistical analysis). Three days afterbehavioral testing, animals were exposed to a single period of re-straint, which involved placing rats into flat bottom Plexiglasrestrainers (8.5×21.5 cm; Kent Scientific, Litchfield, CT) for 30 minand then returning them to their home cage for an additional 30 minperiod. Tail vein blood samples were obtained immediately followingremoval from the home cage (0 min), at the end of restraint (30 min),and 30 min following the termination of restraint (60 min). The bloodsamples were collected into ice-chilled Aprotinin-EDTA-treated tubes(3.75 mg EDTA/100 uL of blood), centrifuged at 3000 ×g for 20 min,and stored at −20 °C until assayed. All testing was performed duringthe light phase of the cycle, beginning at 0800 h.
Plasma ACTH (25 µl), corticosterone (5 µl), and testosterone(25 µl) concentrations were measured using commercial RIA kits (MPBiomedicals, Costa Mesa, CA) and 125I-labeled ligands as previouslydescribed (Viau et al., 2005). For standards and samples, 3 and 2replicates, respectively, were used in all assays. For corticosterone, theplasma samples were diluted 1:100 and 1:200 for pre-stress and post-stress time intervals, respectively, to render hormone detectionwithinthe linear part of the standard curve. The intra- and interassay coef-ficients of variation for these assays ranged from 4–6 to 10–12% res-pectively. The ACTH antibody cross-reacts 100% with ACTH1–39 andACTH1–24, but not with ß-endorphin, ß-MSH, or ß-lipotropin (allb0.8%). The standard curve ED50 for ACTHwas 89.1 pg/ml, with upperand lower detection limits of 700 pg/ml and 5.7 pg/ml, respectively.The corticosterone antibody cross-reacts 100% with corticosterone,and slightly with desoxycorcticosterone (0.34%), and testosterone andcortisol (0.10%). The standard curve ED50 for the corticosterone RIAwas 15 µg/dl, with upper and lower detection limits of 87 µg/dl and0.2 µg/dl, respectively.
The testosterone antibody cross-reacts 100% with testosterone,and slightly with 5α-dihydrotestosterone (3.40%), 5α-androstane-3β,17-β-diol (2.2%) and 11-oxotestosterone (2%). The standard curveED50 for the testosterone RIA was 1.2 ng/ml, with upper and lowerdetection limits of 7.5 ng/ml and 0.1 ng/ml, respectively.
2.8. Statistical analysis
Because our model was amenable to repeated testing, this allowedus to assess and make certain the stability of individual responsetendencies. We restricted our analyses only to those rats displayingstable defensive burying responses, defined as those showing lessthan a 1 order of magnitude shift in the duration of burying (min) over3 consecutive test trials. Thus, behavioral data and tissue sets derivedfrom individual rats displaying any combination of burying times thatdeviated more than 1 order of magnitude between trials (e.g., 5 minon one trial, but 0.5 min on another) were not analyzed. We per-formed a median split of the total population to classify animals ashigh and low buriers (designated High- and Low-bury, respectively),and by maximizing the number of animals studied, to increase thepower of tests involving repeated measures (within-subject effects).In the first study, 20 rats were identified as showing stable patterns inbury duration profiles, and a median split of this data yielded twogroups of rats (10 animals each). Fourteen rats were identified in thesecond (neuroendocrine) study, yielding two groups of rats showingstable, low and high tendencies in defensive burying (7 animals each).The inter-observer reliability of the burying duration measure wasextremely high (r=0.90).
As shown in Fig. 1, the median split produced groups of animalsshowing very high to very low levels of burying, with considerablymore spread in the High-bury group. As expected, Bartlett's testfor homogeneity of variance indicated unequal variances between the
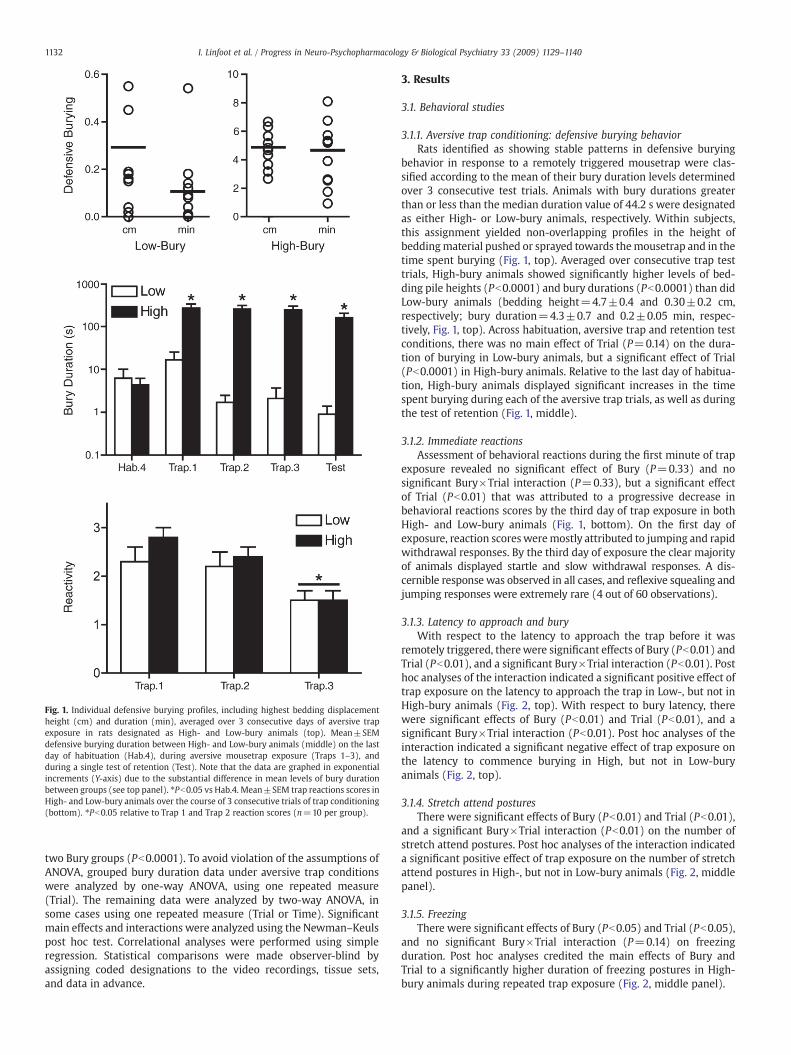
Fig. 1. Individual defensive burying profiles, including highest bedding displacementheight (cm) and duration (min), averaged over 3 consecutive days of aversive trapexposure in rats designated as High- and Low-bury animals (top). Mean±SEMdefensive burying duration between High- and Low-bury animals (middle) on the lastday of habituation (Hab.4), during aversive mousetrap exposure (Traps 1–3), andduring a single test of retention (Test). Note that the data are graphed in exponentialincrements (Y-axis) due to the substantial difference in mean levels of bury durationbetween groups (see top panel). ⁎Pb0.05 vs Hab.4. Mean±SEM trap reactions scores inHigh- and Low-bury animals over the course of 3 consecutive trials of trap conditioning(bottom). ⁎Pb0.05 relative to Trap 1 and Trap 2 reaction scores (n=10 per group).
1132 I. Linfoot et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 33 (2009) 1129–1140
two Bury groups (Pb0.0001). To avoid violation of the assumptions ofANOVA, grouped bury duration data under aversive trap conditionswere analyzed by one-way ANOVA, using one repeated measure(Trial). The remaining data were analyzed by two-way ANOVA, insome cases using one repeated measure (Trial or Time). Significantmain effects and interactions were analyzed using the Newman–Keulspost hoc test. Correlational analyses were performed using simpleregression. Statistical comparisons were made observer-blind byassigning coded designations to the video recordings, tissue sets,and data in advance.
3. Results
3.1. Behavioral studies
3.1.1. Aversive trap conditioning: defensive burying behaviorRats identified as showing stable patterns in defensive burying
behavior in response to a remotely triggered mousetrap were clas-sified according to the mean of their bury duration levels determinedover 3 consecutive test trials. Animals with bury durations greaterthan or less than the median duration value of 44.2 s were designatedas either High- or Low-bury animals, respectively. Within subjects,this assignment yielded non-overlapping profiles in the height ofbeddingmaterial pushed or sprayed towards themousetrap and in thetime spent burying (Fig. 1, top). Averaged over consecutive trap testtrials, High-bury animals showed significantly higher levels of bed-ding pile heights (Pb0.0001) and bury durations (Pb0.0001) than didLow-bury animals (bedding height=4.7±0.4 and 0.30±0.2 cm,respectively; bury duration=4.3±0.7 and 0.2±0.05 min, respec-tively, Fig. 1, top). Across habituation, aversive trap and retention testconditions, there was no main effect of Trial (P=0.14) on the dura-tion of burying in Low-bury animals, but a significant effect of Trial(Pb0.0001) in High-bury animals. Relative to the last day of habitua-tion, High-bury animals displayed significant increases in the timespent burying during each of the aversive trap trials, as well as duringthe test of retention (Fig. 1, middle).
3.1.2. Immediate reactionsAssessment of behavioral reactions during the first minute of trap
exposure revealed no significant effect of Bury (P=0.33) and nosignificant Bury×Trial interaction (P=0.33), but a significant effectof Trial (Pb0.01) that was attributed to a progressive decrease inbehavioral reactions scores by the third day of trap exposure in bothHigh- and Low-bury animals (Fig. 1, bottom). On the first day ofexposure, reaction scoresweremostly attributed to jumping and rapidwithdrawal responses. By the third day of exposure the clear majorityof animals displayed startle and slow withdrawal responses. A dis-cernible response was observed in all cases, and reflexive squealing andjumping responses were extremely rare (4 out of 60 observations).
3.1.3. Latency to approach and buryWith respect to the latency to approach the trap before it was
remotely triggered, therewere significant effects of Bury (Pb0.01) andTrial (Pb0.01), and a significant Bury×Trial interaction (Pb0.01). Posthoc analyses of the interaction indicated a significant positive effect oftrap exposure on the latency to approach the trap in Low-, but not inHigh-bury animals (Fig. 2, top). With respect to bury latency, therewere significant effects of Bury (Pb0.01) and Trial (Pb0.01), and asignificant Bury×Trial interaction (Pb0.01). Post hoc analyses of theinteraction indicated a significant negative effect of trap exposure onthe latency to commence burying in High, but not in Low-buryanimals (Fig. 2, top).
3.1.4. Stretch attend posturesThere were significant effects of Bury (Pb0.01) and Trial (Pb0.01),
and a significant Bury×Trial interaction (Pb0.01) on the number ofstretch attend postures. Post hoc analyses of the interaction indicateda significant positive effect of trap exposure on the number of stretchattend postures in High-, but not in Low-bury animals (Fig. 2, middlepanel).
3.1.5. FreezingThere were significant effects of Bury (Pb0.05) and Trial (Pb0.05),
and no significant Bury×Trial interaction (P=0.14) on freezingduration. Post hoc analyses credited the main effects of Bury andTrial to a significantly higher duration of freezing postures in High-bury animals during repeated trap exposure (Fig. 2, middle panel).

Fig. 2. Mean±SEM trap approach and bury latencies (top), stretch attend and freezing(middle), and grooming and rearing postures (bottom) averaged over 4 consecutivedays of habituation (Hab), 3 consecutive days of conditioning (Trap), and during asingle test of retention (Test) in High- and Low-bury animals. ⁎Pb0.05 vs. Low-bury;tPb0.05 vs. Hab (n=10 per group).
Fig. 3. Mean±SEM defensive burying duration (top), stretch attend (middle), andfreezing postures (bottom) in High- and Low-bury animals during the first (Hab.1) andlast (Hab.4) days of neutral mousetrap exposure. ⁎Pb0.05 vs. Low-bury; tPb0.05 vs.Hab.1 (n=10 per group). nd=not detected.
1133I. Linfoot et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 33 (2009) 1129–1140
3.1.6. Grooming and rearingThere was a significant effect of Bury (Pb0.01), no significant effect
of Trial (P=0.65), but a significant Bury×Trial interaction (Pb0.01)on grooming behavior. Post hoc analyses of the interaction indicated asignificant positive effect of trap exposure on the duration ofgrooming in Low-, but not in High-bury animals (Fig. 2, bottom).With respect to rearing behavior, there was a significant effect of Bury(P=0.02), no significant effect of Trial (P=0.06), and no significantBury×Trial interaction (P=0.60). Post hoc analysis credited the maineffect of Bury to a significantly higher duration of rearing in Low-buryanimals (Fig. 2, bottom).
3.1.7. Field and trap explorationMultiple parameters were used to study exploratory behavior,
including the number of chamber (center-line) crossings, and therelative time spent occupying the proximal and distal halves of thechamber. Assessment of the number of center chamber crossings
revealed no significant effects of Bury (P=0.40) and Trial (P=0.15),and no significant Bury×Trial interaction (P=0.56). Determining theratio of the time spent occupying the proximal and distal halves of thechamber revealed no significant effect of Bury (P=0.83), a significanteffect of Trial (Pb0.05), and no significant Bury×Trial interaction(P=0.12). The effect of trial was credited to a significant increase(Pb0.05) in the relative time spent within the proximal half of thechamber during repeated trap exposure.
In summary (see Table 1), in order of sequence High- and Low-bury animals first showed comparable behavioral reactions (i.e.reflexive startle, withdrawal, and running responses) during the firstminute after the trap was remotely triggered. Marked groupdepartures in stretched approaches and defensive burying wereinitiated thereafter. Beyond this interval, and throughout the buryingtest, High-bury animals displayed higher levels of freezing and lowerlevels of grooming postures than did Low-bury animals. Takentogether, these findings indicate that the High-bury group of animals“avoid” the trap by actively burying it, whereas the Low-bury groupappear to avoid the trap altogether, consistent with their greatertendency to display rearing postures, taken as an index of explorativeescape.
1134 I. Linfoot et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 33 (2009) 1129–1140
3.1.8. Non-aversive trap habituationTo explore differences in unconditioned behavior, behavioral
responses over the course of repeated exposure to a neutral mousetrapduring habituationwere retrospectively analyzed within individual ratsdesignated as High- and Low-bury animals during the conditioningphase of the paradigm. No significant effects of Bury and Trial, and nosignificant Bury×Trial interactions were detected with respect to fieldand trap exploration parameters (P≥0.15 in all cases), although animalstended to display an increase in trap contact frequency (P=0.06) overthe course of habituation. Significant effects were revealed, however,with respect to bury duration and the frequency/duration of stretchattend, grooming, rearing, and freezing postures (Fig. 3).
3.1.9. Bury durationThere was a significant effect of Bury (Pb0.05), no significant effect
of Trial (P=0.11), and no significant Bury×Trial interaction (P=0.14).The main effect of Bury was credited to significantly higher buryduration levels in High-bury animals during initial neutral trapexposure (Fig. 3, top panel). By the fourth day of trap habituation
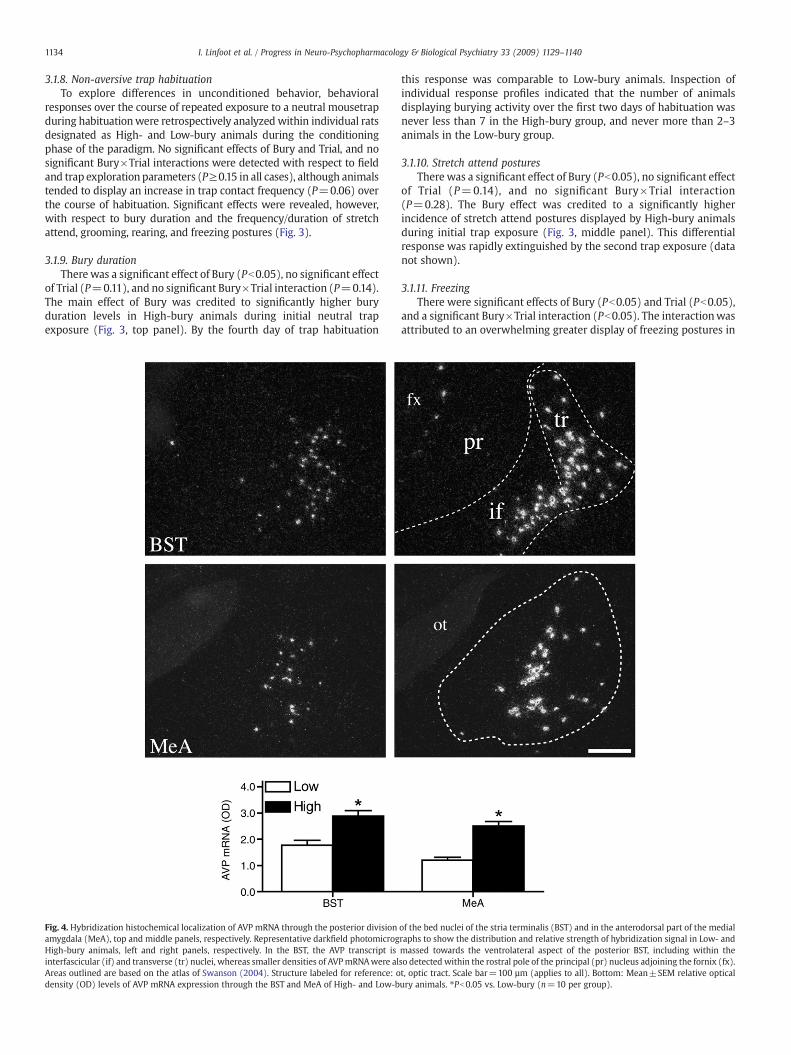
Fig. 4. Hybridization histochemical localization of AVP mRNA through the posterior divisionamygdala (MeA), top and middle panels, respectively. Representative darkfield photomicrogHigh-bury animals, left and right panels, respectively. In the BST, the AVP transcript isinterfascicular (if) and transverse (tr) nuclei, whereas smaller densities of AVPmRNAwere alAreas outlined are based on the atlas of Swanson (2004). Structure labeled for reference: odensity (OD) levels of AVP mRNA expression through the BST and MeA of High- and Low-b
this response was comparable to Low-bury animals. Inspection ofindividual response profiles indicated that the number of animalsdisplaying burying activity over the first two days of habituation wasnever less than 7 in the High-bury group, and never more than 2–3animals in the Low-bury group.
3.1.10. Stretch attend posturesThere was a significant effect of Bury (Pb0.05), no significant effect
of Trial (P=0.14), and no significant Bury×Trial interaction(P=0.28). The Bury effect was credited to a significantly higherincidence of stretch attend postures displayed by High-bury animalsduring initial trap exposure (Fig. 3, middle panel). This differentialresponse was rapidly extinguished by the second trap exposure (datanot shown).
3.1.11. FreezingThere were significant effects of Bury (Pb0.05) and Trial (Pb0.05),
and a significant Bury×Trial interaction (Pb0.05). The interactionwasattributed to an overwhelming greater display of freezing postures in
of the bed nuclei of the stria terminalis (BST) and in the anterodorsal part of the medialraphs to show the distribution and relative strength of hybridization signal in Low- andmassed towards the ventrolateral aspect of the posterior BST, including within theso detected within the rostral pole of the principal (pr) nucleus adjoining the fornix (fx).t, optic tract. Scale bar=100 µm (applies to all). Bottom: Mean±SEM relative opticalury animals. ⁎Pb0.05 vs. Low-bury (n=10 per group).

1135I. Linfoot et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 33 (2009) 1129–1140
High-bury animals on the first day that was nearly abolished by thefourth day of neutral trap exposure (Fig. 3, bottom panel).
3.1.12. Grooming and rearingThere was no significant effect of Bury (P=0.39), a significant
effect of Trial (Pb0.01), and no significant Bury×Trial interaction(P=0.39) on grooming. Post hoc analysis attributed the Trial effect toan overall positive effect (Pb0.05) of trap habituation on groomingfrequency. With respect to rearing, there were significant effects ofBury (Pb0.05) and Trial (Pb0.05), and no significant Bury×Trialinteraction (P=0.68). Post hoc analyses attributed the Bury and Trialeffects to a significantly higher display of rearing in Low-bury animals(Pb0.05) and an overall negative effect of trap habituation on rearingfrequency (Pb0.05).
To explore the extent to which differences in conditioned defen-sive burying could be accounted for by differences in unconditionedbehavior, the duration of burying during aversive trap conditions wasanalyzed as a function of individual behaviors recorded during thehabituation phase of the paradigm. No reliable relationship was foundwith respect to bury duration (P=0.29), but reliable positive rela-tionships were found with respect to stretch attend (r=+0.49;Pb0.05) and freezing (r=+0.57; Pb0.01) postures. Despite the sig-nificance of these correlations, qualitative examination of the scat-tered plots suggested that disproportionate outliers of High-burycasesmight have influenced the slopes of the relationships; at least forstretch attend postures. Nonetheless, the bulk of the Bury-groupdifferences during habituation suggest that defensive burying levelsunder aversive testing could be predicted by variations in behaviorunder non-aversive conditions.
3.2. CRH, AVP, and OT in situ hybridization
Densitometric analysis of AVP mRNA through the extent of theposterior division of the bed nuclei of the stria terminalis (BST) andthe medial nucleus of the amygdala revealed significantly higherlevels of the transcript in both of these regions in High-bury animalscompared to those identified as Low-bury animals (Fig. 4). The groupdifference in AVP mRNA in the posterior BST was most apparentwithin the interfascicular and transverse nuclei and through therostrocaudal extent of the medial amygdala, including its anterodorsalpart.
Within the central nucleus of the amygdala and through theanterior division of the BST, including within the anteromedial,fusiform, and oval nuclei, there was no significant group differences inCRH mRNA levels (data not shown). Within the PVN, there was nosignificant group difference in AVP (Fig. 5) or OT (Fig. 6) mRNA levelsin the medial parvocellular, dorsal (mpd) part of the nucleus.Likewise, there was no significant difference in the relative levels ofCRH mRNA within the mpd region (data not shown). Compared toLow-bury rats, High-bury animals showed significantly lower levels ofAVP and OTmRNAwithin both the posterior magnocellular part of thePVN and the supraoptic nucleus. The dorsal parvocellular part of thePVN was the only subregion of the parvicellular division to show agroup difference in OT mRNA levels, significantly higher in Low-buryrats (Fig. 6).
3.3. Androgen receptor immunocytochemistry
In line with our prediction that the group differences in AVP couldbe accounted for by differences in androgen receptors, total cell countestimates derived from androgen receptor staining through the extentof themedial nucleus of the amygdala and the posterior division of theBST revealed significantly higher numbers of androgenpositive cells inHigh- compared to Low-bury animals (Fig. 7) in both of these brainregions. The group difference in androgen receptor cell numbers wasevident throughout the posterior BST division, including within the
principle, interfascicular, and transverse nuclei, whereas the groupdifference in androgen receptor staining in the medial amygdala wasmost apparent within the posterodorsal part of the nucleus.
3.4. Regression analyses
To further explore relationships between neuropeptide mRNA andandrogen receptor levels as a function of burying, simple regressionanalyses of these parameters were performed using the time spentburying as the independent variable. Significant positive correlationswere detected between defensive burying and AVP mRNA in the BST(r=+0.50, Pb0.01) and MeA (r=+0.63, Pb0.05), whereas signifi-cant negative relationswere found for OTmRNA in the SON (r=−0.58,Pb0.01), aswell aswithin the dp and PM regions of the PVN (r=−0.64,Pb0.01; r=−0.50, Pb0.05, respectively). With respect to AR cellnumbers, significant positive correlations were detected in both theBST (r=+0.45, Pb0.05) and MeA (r=+0.68, Pb0.01). Despite thestrength of these correlations, closer inspection of scatterplots fordisproportionate outliers and/or inordinate clusters of cases revealedonly a discernable relationship for AR cell numbers in the MeA. Indeed,when Low-buryanimalswere removed fromall of the regressionplots, asingle reliable relationship remained with respect to bury duration andAR in this region (r=+0.69, Pb0.01). This suggests that the differencein androgen receptor levels in the medial amygdala could be accountedfor by individual variations in defensive burying, at least amongst High-bury animals. Identical resultswere found in all cases using bedding pileheight as the independent variable, in line with the high correlationbetween bury duration and pile height (r=0.96) across animals.
3.5. Neuroendocrine function
Based on previous findings that differences in HPA activity andgonadal function occur in animals showing different coping behaviors,we then measured in a separate group of animals how plasmapituitary-adrenal hormones and testosterone vary in rats identified asshowing stable individual differences in defensive burying levels(Fig. 8). Analyses of plasma ACTH and corticosterone responses to asingle 30 min episode of restraint revealed significant main effects ofBury (Pb0.01) and Restraint (Pb0.01), and significant bury×restraintinteractions (Pb0.01). These interactions were attributed to signifi-cantly higher ACTH and corticosterone levels in High- compared toLow-bury animals, both during and following restraint. For testoster-one, there was a significant effect of bury (Pb0.05), but no significanteffect of restraint (P=0.11), and no significant bury×restraintinteraction (P=0.11).
4. Discussion
The defensive burying response evoked by the mousetrap used inthe current studymodels the natural tendencies of rodents to respondto aversive stimuli by pushing, spraying or otherwisemoving availablematerials towards sources of threats (Pinel and Mana, 1989). Studiesemploying shock-induced defensive burying and less intense oraversive behavioral paradigms have made several important inroadson the neurobiological correlates of anxiety. Our findings underscorethe utility of using a variant of the defensive burying paradigm foridentifying rats that reliably chose to bury and those that did not.
We are not the first to notice this phenomenon, as similarvariability in the tendency to bury has been described in numerousstudies using the shock prod test (De Boer and Koolhaas, 2003),although often ignored in tests using noxious materials and harmlessnovel objects. In the traditional version of the shock prod test, ratstend to avoid further contact with an electrified probe once they'vebeen shocked, making repeated testing problematic. Similar to othermodels employing harmless objects, including marble burying(Njung'e and Handley, 1991), a key feature of our remotely controlled

Fig. 5.Hybridization histochemical localization of AVPmRNA in the caudal half of the paraventricular nucleus (PVN) of the hypothalmus. Representative darkfield photomicrographsto show the distribution and relative strength of hybridization signal in Low- and High-bury animals, left and right panels, respectively. At the PVN level shown, the AVP transcript isconcentrated within the posterior magnocellular (PM) part, with substantially lower densities in the dorsal body of the medial parvocellular (mpd) part and in the medialparvocellular ventral (mpv) region. Within the rostral half of the PVN, AVP mRNA was also detected within the medial magnocellular (MM) part (not shown). Scale bar=100 µm(applies to all). Bottom: Mean±SEM relative optical density (OD) levels of AVP mRNA expression within the supraoptic nucleus (SON) and different compartments of the PVN inHigh- and Low-bury animals. ⁎Pb0.05 vs. Low-bury (n=10 per group).
Fig. 6. Hybridization histochemical localization of OT mRNA in the caudal half of the paraventricular nucleus (PVN) of the hypothalmus. Representative darkfield photomicrographsto show the distribution and relative strength of hybridization signal in Low- and High-bury animals, left and right panels, respectively. Compared to AVP, the OT transcript shows arelatively wider distribution, including within the dorsal parvocellular (dp) part, the periventricular part lining the third ventricle (3 V), and a noticeable distribution within thelateral and anteroventral aspects of the posterior magnocellular (PM) division adjoining the medial parvocellular ventral (mpv) region. OT mRNA was also detected to varyingdegrees within the rostral half of the PVN, including within anterior and medial magnocellular (AM, MM) parts, and within the lateral parvocellular (lp) part occupying the caudalextent of the PVN (not shown). Scale bar=100 µm (applies to all). Bottom: Mean±SEM relative optical density (OD) levels of OT mRNA expression within the supraoptic nucleus(SON) and different compartments of the PVN in High- and Low-bury animals. ⁎Pb0.05 vs. Low-bury (n=10 per group).
1136 I. Linfoot et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 33 (2009) 1129–1140

Fig. 7. Immunohistochemical localization of androgen receptor (AR) immunoreactivity through the posterior division of the bed nuclei of the stria terminalis (BST) and in theposterodorsal part of the medial amygdala (MeA), top andmiddle panels, respectively. Representative brightfield photomicrographs to show the distribution and relative numbers ofAR positive cells in Low- and High-bury animals, left and right panels, respectively. In the BST, AR staining was most concentrated within the principal (pr) nucleus, with asubstantially lower density in the interfascicular (if) and transverse (tr) nuclei. In the MeA, AR staining was most concentrated within the posterodorsal part, with a lower densitywithin the anterodorsal part (not shown). Structures labeled for reference: cpd, cerebral peduncle; fx, fornix; ot, optic tract; sm, stria medullaris. Bottom: Mean±SEM relativenumber of androgen receptor (AR) positive cells through the extent of the posterior division of the bed nuclei of the stria terminalis (BST) and the medial nucleus of the amygdala(MeA) in High- and Low-bury animals. ⁎Pb0.05 vs. Low-bury (n=10 per group).
1137I. Linfoot et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 33 (2009) 1129–1140
and pain-free mousetrap model is that it is amenable to repeatedtesting, both under neutral and aversive conditions, allowing thestability of individual behavioral response tendencies to be assessedand quantified.
In contrast to other models employing harmless objects, however,the mousetrap represents a complex object and provides, in the veryleast, an auditory stimulus (loud audible snap). Rats bury complexnovel objects placed for the first time in a familiar environment, buteven more so if paired with an aversive stimulus (Terlecki et al., 1979).This could provide a basis for the range and amplitudes of defensiveburying achieved in the current study, significantly higher than whatis commonly observed for rodents in response to other harmlessobjects. As we had hoped for, this enhanced our capacity for detectingindividual differences in defensive behaviors. Our data show that AVP,OT, and androgen receptors sort with variations in coping style andunderscore different neuroendocrine capacities in rats identified asHigh- and Low-bury animals.
Rats seem to have entered the experimental paradigmwith alreadyestablished tendencies to display defensive burying or not at all. This
was observed when the mousetrap was introduced as an aversivestimulus source (remotely triggered) or under more neutral condi-tions during the habituation phase of the paradigm. It is important tonote that the pushing and/or spraying of bedding material was alwaysdirected towards the trap and that this occurred even in rats showingmajor differences in burying activity, indicating that the mousetrapwas readily identified as the specific source of aversive stimulation.Further, the magnitude of the behavioral responses observed, werewell within the range observed in previous studies employing shockprod, but significantly higher thanwhat is normally observed for otherharmless objects. Thus, the present findings reflect individual tenden-cies in defensive behavior that were not shaped by any underlyinglimitations of using a pain-free test of defensive burying.
High- and Low-bury animals displayed similar trap-reaction scores(and adaptation) during repeated testing, providing some indicationthat the information gained between the groups was equivalent, atleast in terms of the threatening nature, location, and regularityby which the mousetrap dispensed an aversive stimulus. High-buryanimals displayed substantially higher numbers of stretched
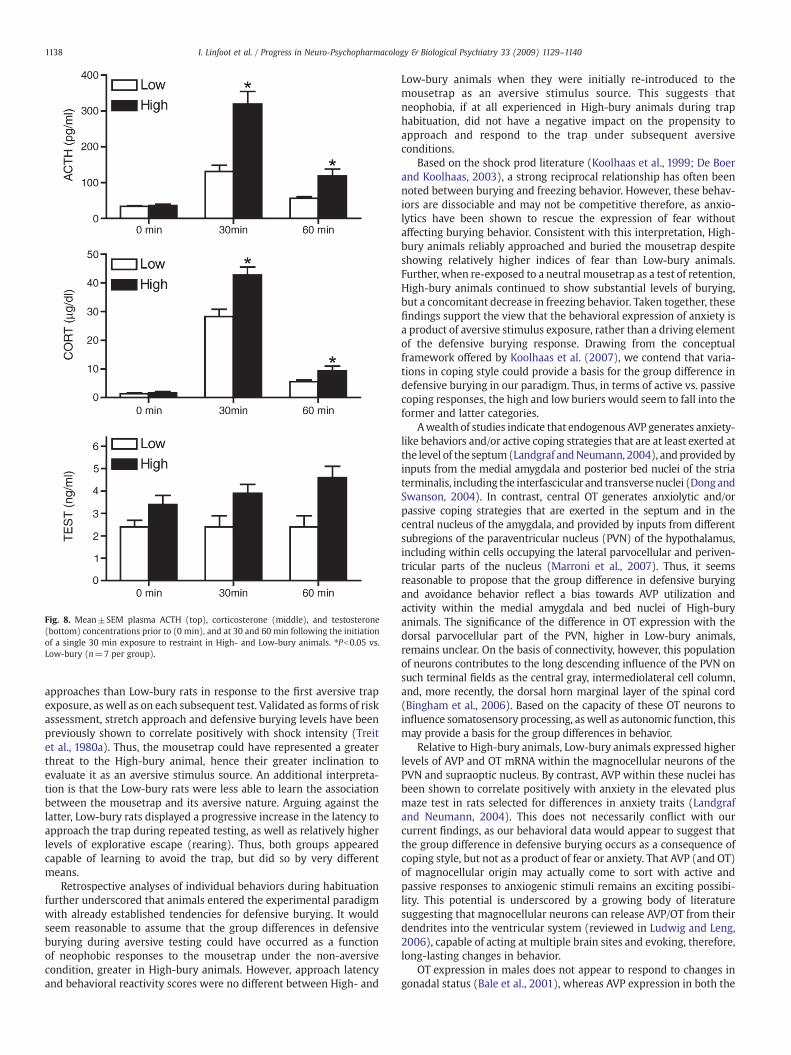
Fig. 8. Mean±SEM plasma ACTH (top), corticosterone (middle), and testosterone(bottom) concentrations prior to (0 min), and at 30 and 60 min following the initiationof a single 30 min exposure to restraint in High- and Low-bury animals. ⁎Pb0.05 vs.Low-bury (n=7 per group).
1138 I. Linfoot et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 33 (2009) 1129–1140
approaches than Low-bury rats in response to the first aversive trapexposure, as well as on each subsequent test. Validated as forms of riskassessment, stretch approach and defensive burying levels have beenpreviously shown to correlate positively with shock intensity (Treitet al., 1980a). Thus, the mousetrap could have represented a greaterthreat to the High-bury animal, hence their greater inclination toevaluate it as an aversive stimulus source. An additional interpreta-tion is that the Low-bury rats were less able to learn the associationbetween the mousetrap and its aversive nature. Arguing against thelatter, Low-bury rats displayed a progressive increase in the latency toapproach the trap during repeated testing, as well as relatively higherlevels of explorative escape (rearing). Thus, both groups appearedcapable of learning to avoid the trap, but did so by very differentmeans.
Retrospective analyses of individual behaviors during habituationfurther underscored that animals entered the experimental paradigmwith already established tendencies for defensive burying. It wouldseem reasonable to assume that the group differences in defensiveburying during aversive testing could have occurred as a functionof neophobic responses to the mousetrap under the non-aversivecondition, greater in High-bury animals. However, approach latencyand behavioral reactivity scores were no different between High- and
Low-bury animals when they were initially re-introduced to themousetrap as an aversive stimulus source. This suggests thatneophobia, if at all experienced in High-bury animals during traphabituation, did not have a negative impact on the propensity toapproach and respond to the trap under subsequent aversiveconditions.
Based on the shock prod literature (Koolhaas et al., 1999; De Boerand Koolhaas, 2003), a strong reciprocal relationship has often beennoted between burying and freezing behavior. However, these behav-iors are dissociable and may not be competitive therefore, as anxio-lytics have been shown to rescue the expression of fear withoutaffecting burying behavior. Consistent with this interpretation, High-bury animals reliably approached and buried the mousetrap despiteshowing relatively higher indices of fear than Low-bury animals.Further, when re-exposed to a neutral mousetrap as a test of retention,High-bury animals continued to show substantial levels of burying,but a concomitant decrease in freezing behavior. Taken together, thesefindings support the view that the behavioral expression of anxiety isa product of aversive stimulus exposure, rather than a driving elementof the defensive burying response. Drawing from the conceptualframework offered by Koolhaas et al. (2007), we contend that varia-tions in coping style could provide a basis for the group difference indefensive burying in our paradigm. Thus, in terms of active vs. passivecoping responses, the high and low buriers would seem to fall into theformer and latter categories.
Awealth of studies indicate that endogenous AVP generates anxiety-like behaviors and/or active coping strategies that are at least exerted atthe level of the septum(Landgraf andNeumann, 2004), and providedbyinputs from the medial amygdala and posterior bed nuclei of the striaterminalis, including the interfascicular and transversenuclei (DongandSwanson, 2004). In contrast, central OT generates anxiolytic and/orpassive coping strategies that are exerted in the septum and in thecentral nucleus of the amygdala, and provided by inputs from differentsubregions of the paraventricular nucleus (PVN) of the hypothalamus,including within cells occupying the lateral parvocellular and periven-tricular parts of the nucleus (Marroni et al., 2007). Thus, it seemsreasonable to propose that the group difference in defensive buryingand avoidance behavior reflect a bias towards AVP utilization andactivity within the medial amygdala and bed nuclei of High-buryanimals. The significance of the difference in OT expression with thedorsal parvocellular part of the PVN, higher in Low-bury animals,remains unclear. On the basis of connectivity, however, this populationof neurons contributes to the long descending influence of the PVN onsuch terminal fields as the central gray, intermediolateral cell column,and, more recently, the dorsal horn marginal layer of the spinal cord(Bingham et al., 2006). Based on the capacity of these OT neurons toinfluence somatosensory processing, aswell as autonomic function, thismay provide a basis for the group differences in behavior.
Relative to High-bury animals, Low-bury animals expressed higherlevels of AVP and OT mRNA within the magnocellular neurons of thePVN and supraoptic nucleus. By contrast, AVP within these nuclei hasbeen shown to correlate positively with anxiety in the elevated plusmaze test in rats selected for differences in anxiety traits (Landgrafand Neumann, 2004). This does not necessarily conflict with ourcurrent findings, as our behavioral data would appear to suggest thatthe group difference in defensive burying occurs as a consequence ofcoping style, but not as a product of fear or anxiety. That AVP (and OT)of magnocellular origin may actually come to sort with active andpassive responses to anxiogenic stimuli remains an exciting possibi-lity. This potential is underscored by a growing body of literaturesuggesting that magnocellular neurons can release AVP/OT from theirdendrites into the ventricular system (reviewed in Ludwig and Leng,2006), capable of acting at multiple brain sites and evoking, therefore,long-lasting changes in behavior.
OT expression in males does not appear to respond to changes ingonadal status (Bale et al., 2001), whereas AVP expression in both the

1139I. Linfoot et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 33 (2009) 1129–1140
medial amygdala and posterior BST is extremely dependent ontestosterone (De Vries and Miller, 1998). In line with this AVPrequirement for testosterone, High-bury animals displayed highernumbers of AR positive cells than did Low-bury animals in these samebrain regions. Within the posterior division of the BST, the groupdifference in AR cell numbers was evident within the principalnucleus, but also within the interfascicular, and transverse nuclei,where AVP cells are concentrated. The extent to which the groupdifference in AR actually occurs within AVP cells remains to be seen,and worthy of pursuit, as previous reports have indicated thatvirtually all AVP cells in this region of the BST, as well as within themedial amygdala, stain for AR (Zhou et al, 1994).
High-bury animals appeared to show higher levels of testosteronein circulation under restraint stress conditions. If this reflects a generaltendency for High-bury animals to secrete testosterone under chal-lenging situations, this could provide a means of sustaining higherlevels of AVP within the medial amygdala and posterior BST. Con-sistent with the relations observed between AR, testosterone, anddefensive burying in the present study, rodents showing higher levelsof aggressive behavior display increased levels of testosterone, as wellas shock-induced defensive burying (De Boer and Koolhaas, 2003).Taken together, these findings suggest a potential role for testosteroneon defensive burying (see Frye and Seliga, 2001) that may rest withinthese central AVP pathways.
Importantly, the size and distribution of the group difference inAR cell numbers vastly exceeded the number of cells capable ofexpressing AVP within the posterior BST and medial amygdala. Thus,several additional phenotypes may come to sort with individual dif-ferences in coping style within the BST and amygdala. On the bases ofprevious behavioral and anatomical studies, several candidateneuropeptides stand out in this regard (reviewed in Barrera et al.,2005; Harro, 2006; Mathew et al., 2008, and see Kask et al., 2002;Khoshbouei et al., 2002; Polston and Simerly, 2003; Ebner et al.,2004), including cholecystokinin galanin, neuropeptide Y, and sub-stance P, which have been shown to be relevant to anxiety and maythus be important for coping behavior. However, their exact distribu-tion within cells identified as expressing AR and containment withinafferentmediators of defensive behavior has yet to be fully considered.
In addition to targeting to the septal region, the principal nucleusof the posterior BST contains AR positive cells that project to the mpdpart of the PVN to regulate the expression of CRH and AVP (Choi et al.,2007;Williamson and Viau, 2007). Thus, this region of the BSTmay bean important neural substrate for the relations observed between AR,defensive burying, and HPA output in the present study. Relativeassessments of CRH, AVP, and OT mRNA expression within the mpdpart of the PVN failed to yield significant group differences for thesetranscripts. Nonetheless, as we observed in a separate group ofanimals, High-bury animals displayed higher pituitary-adrenalresponses to acute restraint exposure. Dynamic studies on the relativerelease patterns and contributions of these principal ACTH secreta-gogues in animals identified as showing differences in defensiveburying warrant further consideration. Gleaned from connectional,phenotypic, and morphological studies, the topographic segregationof different functional OT cell types in the PVN is not absolute (seeBingham et al., 2006). Thus, future studies employing a retrogradetracer approach could provide the resolution required for localizing,perhaps, even greater group differences in OT expression, not onlywith respect to those neurons governing adrenal output, but also tothose giving rise to forebrain projections.
We have only just begun to understand the bases for individualdifferences in defensive burying and coping style. As neuropeptideand androgen receptor assessments were made within 24 h of behav-ioral testing, causality remains an important issue. Based on thestrength of our current findings, worthy of pursuit is how these factorsactually respond to behavioral testing, sort as a function of behavior inanimals under non-aversive conditions, andwhether pharmacological
antagonism of these candidate neuropeptide and sex steroid hormonereceptors alters behavior in animals identified as showing differencesin defensive burying behaviors. Nevertheless, our results are in manyways reminiscent of the neurobiological correlates of rats showingindividual variations in novelty seeking behavior and coping stylethat are linked to differences in HPA output and central AVP and OTfunction. As these studies provide important models for understand-ing individual differences in stress-induced psychopathology andrecovery, our paradigm provides a new entry point for unmaskinginnate differences in psychological state that we can reliably detect,as well as predict.
Acknowledgements
This study was supported by the Canadian Institutes of HealthResearch and the Michael Smith Foundation for Health Research (VV),and the Canadian Graduate Student Scholarships Doctoral Awardsfrom the Canadian Institutes of Health Research (BB and MG). We arealso grateful for the expert statistical assistance of Dr. Stan Florescoand Dr. Liissa Galea (Department of Psychology, UBC).
References
Bale TL, Davis AM, Auger AP, Dorsa DM, McCarthy MM. CNS region-specific oxytocinreceptor expression: importance in regulation of anxiety and sex behavior.J Neurosci 2001;21:2546–52.
Barrera G, Echevarria DJ, Poulin JF, Laforest S, Drolet G, Morilak DA. One for all or one forone: does co-transmission unify the concept of a brain galanin “system” or clarifyany consistent role in anxiety? Neuropeptides 2005;39:289–92.
Bingham B, Williamson M, Viau V. Androgen and estrogen receptor-beta distributionwithin spinal-projecting and neurosecretory neurons in the paraventricularnucleus of the male rat. J Comp Neurol 2006;499:911–23.
Choi DC, Furay AR, Evanson NK, Ostrander MM, Ulrich-Lai YM, Herman JP. Bed nucleusof the stria terminalis subregions differentially regulate hypothalamic-pituitary-adrenal axis activity: implications for the integration of limbic inputs. J Neurosci2007;27:2025–34.
De Boer SF, Koolhaas JM. Defensive burying in rodents: ethology, neurobiology andpsychopharmacology. Eur J Pharmacol 2003;463:145–61.
De Vries GJ, Miller MA. Anatomy and function of extrahypothalamic vasopressinsystems in the brain. Prog Brain Res 1998;119:3-20.
de Wied D, Diamant M, Fodor M. Central nervous system effects of the neurohypo-physeal hormones and related peptides. Front Neuroendocrinol 1993;14:251–302.
Dong HW, Swanson LW. Projections from bed nuclei of the stria terminalis, posteriordivision: implications for cerebral hemisphere regulation of defensive andreproductive behaviors. J Comp Neurol 2004;471:396–433.
Dong HW, Petrovich GD, Swanson LW. Topography of projections from amygdala to bednuclei of the stria terminalis. Brain Res Brain Res Rev 2001;38:192–246.
Ebner K, Rupniak NM, Saria A, Singewald N. Substance P in the medial amygdala:emotional stress-sensitive release and modulation of anxiety-related behavior inrats. Proc Natl Acad Sci USA 2004;101:4280–5.
Frye CA, Seliga AM. Testosterone increases analgesia, anxiolysis, and cognitive perfor-mance of male rats. Cogn Affect Behav Neurosci 2001;1:371–81.
Gray DS, Terlecki LJ, Treit D, Pinel JP. Effect of septal lesions on conditioned defensiveburying. Physiol Behav 1981;27:1051–6.
Guillery RW. On counting and counting errors. J Comp Neurol 2002;447:1–7.Harro J. CCK and NPY as anti-anxiety treatment targets: promises, pitfalls, and strategies.
Amino Acids 2006;31:215–30.Herman JP, Figueiredo H, Mueller NK, Ulrich-Lai Y, OstranderMM, Choi DC, et al. Central
mechanisms of stress integration: hierarchical circuitry controlling hypothalamo-pituitary-adrenocortical responsiveness. Front Neuroendocrinol 2003;24:151–80.
Hudson BB. One trial learning in the domestic rat. Gen Psychol Monographs 1950;41:99-145.
Kabbaj M, Devine DP, Savage VR, Akil H. Neurobiological correlates of individualdifferences in novelty-seeking behavior in the rat: differential expression of stress-related molecules. J Neurosci 2000;20:6983–8.
Kabbaj M, Morley-Fletcher S, Le Moal M, Maccari S. Individual differences in the effectsof chronic prazosin hydrochloride treatment on hippocampal mineralocorticoidand glucocorticoid receptors. Eur J Neurosci 2007;25:3312–8.
Kask A, Harro J, von Hörsten S, Redrobe JP, Dumont Y, Quirion R. The neurocircuitry andreceptor subtypes mediating anxiolytic-like effects of neuropeptide Y. NeurosciBiobehav Rev 2002;26:259–83.
Keck ME. Corticotropin-releasing factor, vasopressin and receptor systems in depres-sion and anxiety. Amino Acids 2006;31:241–50.
Khoshbouei H, Cecchi M, Morilak DA. Modulatory effects of galanin in the lateral bednucleus of the stria terminalis on behavioral and neuroendocrine responses toacute stress. Neuropsychopharmacology 2002;27:25–34.
Koolhaas JM, Everts H, de Ruiter AJ, de Boer SF, Bohus B. Coping with stress in rats andmice: differential peptidergic modulation of the amygdala-lateral septum complex.Prog Brain Res 1998;119:437–48.

1140 I. Linfoot et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 33 (2009) 1129–1140
Koolhaas JM, Korte SM, De Boer SF, Van Der Vegt BJ, Van Reenen CG, Hopster H, et al.Coping styles in animals: current status in behavior and stress-physiology. NeurosciBiobehav Rev 1999;23:925–35.
Koolhaas JM, de Boer SF, Buwalda B, van Reenen K. Individual variation in coping withstress: a multidimensional approach of ultimate and proximate mechanisms. BrainBehav Evol 2007;70:218–26.
Landgraf R. The involvement of the vasopressin system in stress-related disorders. CNSNeurol Disord Drug Targets 2006;5:167–79.
Landgraf R, Wigger A. High vs low anxiety-related behavior rats: an animal model ofextremes in trait anxiety. Behav Genet 2002;32:301–14.
Landgraf R, Neumann ID. Vasopressin and oxytocin release within the brain: a dynamicconcept of multiple and variable modes of neuropeptide communication. FrontNeuroendocrinol 2004;25:150–76.
Ludwig M, Leng G. Dendritic peptide release and peptide-dependent behaviours. NatRev Neurosci 2006;7:126–36.
Marroni SS, Nakano FN, Gati CD, Oliveira JA, Antunes-Rodrigues J, Garcia-Cairasco N.Neuroanatomical and cellular substrates of hypergrooming induced by microinjec-tion of oxytocin in central nucleus of amygdala, an experimental model of compul-sive behavior. Mol Psychiatry 2007;12:1103–17.
MathewSJ, Price RB, CharneyDS. Recent advances in the neurobiologyof anxiety disorders:implications for novel therapeutics. J Med Genet C Semin Med Genet 2008;148C:89–98.
Menard J, Treit D. Effects of centrally administered anxiolytic compounds in animalmodels of anxiety. Neurosci Biobehav Rev 1999;23:591–613.
Neumann ID, Kromer SA, Toschi N, Ebner K. Brain oxytocin inhibits the (re)activity ofthe hypothalamo-pituitary-adrenal axis in male rats: involvement of hypothalamicand limbic brain regions. Regul Pept 2000;96:31–8.
Njung'e K, Handley SL. Evaluation of marble-burying behavior as a model of anxiety.Pharmacol Biochem Behav 1991;38:63–7.
Pawlak CR, Ho YJ, Schwarting RK. Animal models of human psychopathology based onindividual differences in novelty-seeking and anxiety. Neurosci Biobehav Rev 20082008;32:1544–68.
Pinel JP, Treit D. Burying as a defensive response in rats. J Comp Physiol Psychology1978;92:708–12.
Pinel JPJ, Mana MJ. Adaptive interactions of rats with dangerous in animate objects:support for a cognitive theory of defensive behavior. In: Blanchard RJ, Brain PF,Blanchard DC, Parmigiani S, editors. Ethoexperimental approaches to the study ofbehavior. Dordecht: Kluwer Academic; 1989. p. 137–50.
Polston EK, Simerly RB. Sex-specific patterns of galanin, cholecystokinin, and substanceP expression in neurons of the principal bed nucleus of the stria terminalis aredifferentially reflected within three efferent preoptic pathways in the juvenile rat. JComp Neurol 2003;465:551–9.
Roozendaal B, Wiersma A, Driscoll P, Koolhaas JM, Bohus B. Vasopressinergicmodulation of stress responses in the central amygdala of the Roman high-avoidance and low avoidance rat. Brain Res 1992;596:35–40.
Roseboom PH, Nanda SA, Bakshi VP, Trentani A, Newman SM, Kalin NH. Predator threatinduces behavioral inhibition, pituitary-adrenal activation and changes in amygdalaCRF-binding protein gene expression. Psychoneuroendocrinology 2007;32:44–55.
Simmons DM, Arriza JL, Swanson LW. A complete protocol for in situ hybridization ofmessenger RNAs in brain and other tissues with radiolabeled single-stranded RNAprobes. J Histotechnol 1989;12:169–81.
SofroniewMV. Morphology of vasopressin and oxytocin neurones and their central andvascular projections. Prog Brain Res 1983;60:101–14.
Steimer T, Driscoll P. Divergent stress responses and coping styles in psychogeneticallyselected Roman high-(RHA) and low-(RLA) avoidance rats: behavioural, neuroen-docrine and developmental aspects. Stress 2003;6(2):87-100 [Jun].
Swanson LW. Brain maps: structure of the rat brain. third Edition. San Diego: AcademicPress; 2004.
Swanson LW, Kuypers HG. The paraventricular nucleus of the hypothalamus:cytoarchitectonic subdivisions and organization of projections to the pituitary,dorsal vagal complex, and spinal cord as demonstrated by retrograde fluorescencedouble-labeling methods. J Comp Neurol 1980;194:555–70.
Swanson LW, Simmons DM. Differential steroid hormone and neural influences onpeptide mRNA levels in CRH cells of the paraventricular nucleus: a hybridizationhistochemical study in the rat. J Comp Neurol 1989;285:413–35.
Taghzouti K, Lamarque S, Kharouby M, Simon H. Interindividual differences in activeand passive behaviors in the forced-swimming test: implications for animal modelsof psychopathology. Biol Psychiatry 1999;45:750–8.
Terlecki LJ, Pinel JPJ, Treit D. Conditioned and unconditioned defensive burying in therat. Learn Motivat 1979;10:337–50.
Treit D, Pinel JPJ, Terlecki LJ. Shock intensity and conditioned defensive burying in rats.Bull Psychosom Soc 1980a;16:5–7.
Treit D, Terlecki LJ, Pinel JPJ. Conditioned defensive burying in rodents: organismicvariables. Bull Psychosom Soc 1980b;16:451–4.
Treit D, Pinel JPJ, Fibiger HC. Conditioned defensive burying: a new paradigm for thestudy of anxiolytic agents. Pharmacol Biochem Behav 1981;15:619–26.
VeenemaAH,Neumann ID.Neurobiologicalmechanismsof aggressionand stress coping: acomparative study inmouse and rat selection lines. Brain Behav Evol 2007;70:274–85.
Viau V, Sawchenko PE. Hypophysiotropic neurons of the paraventricular nucleus respondin spatially, temporally, and phenotypically differentiated manners to acute vs. re-peated restraint stress: rapid publication. J Comp Neurol 2002;445:293–307.
Viau V, Soriano L, Dallman MF. Androgens alter corticotropin releasing hormone andarginine vasopressin mRNAwithin forebrain sites known to regulate activity in thehypothalamic-pituitary-adrenal axis. J Neuroendocrinol 2001;13:442–52.
Viau V, Bingham B, Davis J, Lee P, Wong M. Gender and puberty interact on the stress-induced activation of parvocellular neurosecretory neurons and corticotropin-releasing hormonemessenger ribonucleic acid expression in the rat. Endocrinology2005;146:137–46.
Walker FR, HinwoodM, Masters L, Deilenberg RA, Day TA. Individual differences predictsusceptibility to conditioned fear arising from psychosocial trauma. J Psychiatr Res2008;42:371–83.
Wiersma A, Knollema S, Konsman JP, Bohus B, Koolhaas JM. Corticotropin-releasinghormone modulation of a conditioned stress response in the central amygdala ofRoman high (RHA/Verh)-avoidance and low (RLA/Verh)-avoidance rats. BehavGenet 1997;27:547–55.
Williamson M, Viau V. Androgen receptor expressing neurons that project to theparaventricular nucleus of the hypothalamus in the male rat. J Comp Neurol2007;503:717–40.
Yilmazer-Hanke DM, Wigger A, Linke R, Landgraf R, Schwegler H. Two Wistar rat linesselectively bred for anxiety-related behavior show opposite reactions in elevatedplus maze and fear-sensitized acoustic startle tests. Behav Genet 2004;34:309–18.
Zhou L, Blaustein JD, De Vries GJ. Distribution of androgen receptor immunoreactivity invasopressin- and oxytocin-immunoreactive neurons in the male rat brain. Endocri-nology 1994;134:2622–7.




















