
Characterization and expression analysis of AGAMOUS-like, SEEDSTICK-like, and SEPALLATA-like...
9
Characterization and expression analysis of AcSERK2, a somatic embryogenesis and stress resistance related gene in pineapple Jun Ma a, b , Yehua He a, ⁎, Zhongyi Hu a , Wentian Xu a , Jingxian Xia a , Cuihong Guo a , Shunquan Lin a , Li Cao a , Chengjie Chen a , Chenghou Wu a , Junli Zhang b a Horticultural Biotechnology College of South China Agricultural University, Guangzhou, Guangdong 510642, China b College of Landscape Architecture of Sichuan Agricultural University, Chengdu, Sichuan 611100, China abstract article info Article history: Accepted 4 March 2012 Available online 21 March 2012 Keywords: Ananas comosus SERK Somatic embryogenesis Expression pattern The isolation and characterization of AcSERK2, another somatic embryogenesis receptor-like kinase (SERK) gene homolog, confirmed that there was a SERK gene family in pineapple. AcSERK2 shared high similarity to AcSERK1 and other SERKs, containing all the characteristic domains of the SERK family. The expressed recombinant pro- tein of pET-AcSERK2 was soluble. AcSERK2 expression was effectively induced by 2,4-dichlorophenoxyacetic acid (2,4-D) and maintained in high level at the early stage of somatic embryogenesis (SE). The results of in situ hybridization showed that AcSERK2 expressed at high level in the competent cells under the induction of 2, 4-D and at low level during the development of meristematic center to globular embryo. This indicated that AcSERK2 could be used as a potential marker gene to monitor the acquisition of embryogenic competence. How- ever, AcSERK2 expression was also detected in all the tested organs and the expression level decreased along with the development of calyx, ovule and ovary. These results suggested that AcSERK2 maybe play a broader role in organ development. AcSERK2 transcription could be induced by exogenous hormones and abiotic stress. It was deduced that AcSERK2 may participate in response to exogenous hormones and abiotic stress regulation in pine- apple. The expression pattern of AcSERK2 during SE differed from that of AcSERK1, and this indicated that the SERK genes may be non-redundant in somatic embryogenesis. © 2012 Elsevier B.V. All rights reserved. 1. Introduction Somatic embryogenesis was first demonstrated by Reinert (1958) and Steward et al. (1958). SE can be used both for rapid propagation of valuable clones and also for gene transfer and regeneration of genet- ically modified plants (Li et al., 2011). Research into SE intensified as plant regeneration in vitro came to be widely used in transformation (Nolan et al., 2003). Chimera formation is a common problem associat- ed with the propagation of transgenic plants derived using explant- based vegetative regeneration protocols. However, the origin of pineap- ple somatic embryos has been demonstrated to be unicellular (He et al., 2010), and thus the use of SE based transformation system may over- come the problem of chimera formation (Sanjeev et al., 2008). The cyto- metric analysis revealed that somatic embryo-derived plantlets have no chromosomal absence, changes or alterations in ploidy (Fiuk et al., 2010). The pre-requisite for a somatic cell to change its usual fate onto an embryogenic pathway, thereby transforming itself into a complete embryo, is poorly understood (Mordhorst et al., 1997). The molecular basis of genetic and biochemical mechanisms leading to SE are not well understood (Nolan et al., 2003; Pérez-Núňez et al., 2009). There are many reports of genes involved in the regulation of the later stages of SE (Schmidt et al., 1997), such as LEC1(Lotan et al., 1998), LEC2 (Stone et al., 2001), BBM (Boutilier et al., 2002), WU (Zuo et al., 2002) and AGL15 (Harding et al., 2003). They can enhance the frequency of SE and prolong the ability of SE. But much less is known of the induction process (Nolan et al., 2003). Many studies attempted to identify the re- lationship of triggering SE with specific gene expression patterns led to the isolation of SE receptor-like kinase (SERK) gene (Baudino et al., 2001; Sanjeev et al., 2008). SERK belongs to Leu-rich repeat, receptor- like kinases (LRR-RLKs), a subgroup of protein kinases, characterized by an extracellular domain with at least five Leu-rich repeats (LRRs), a transmembrane domain and an intracellular kinase domain. They are thought to play key roles in the transduction of extracellular signals in processes as diverse as plant development, disease resistance or self- incompatibility (Baudino et al., 2001; Krupa et al., 2006). One such pro- cess is SE where the embryogenic response of cultured cells depends not only on the cell type used but also on the composition of the culture medium. SERK genes were considered to be multicellular-specific Gene 500 (2012) 115–123 Abbreviations: SERK, somatic embryogenesis receptor-like kinase; 2,4-D, 2,4- dichlorophenoxyacetic acid; SE, somatic embryogenesis; LEC, Leafy cotyledon; BBM, Baby Boom; AGL15, AGAMOUS-like 15; WU, WUSCHEL; LRR-RLKs, Leu-rich repeat, receptor-like kinases; LRRs, Leu-rich repeats; SA, salicylic acid; JA, jasmonic acid; FAA, formalin acetic acid; CTAB, cetyl trimethylammonium bromide; RACE, rapid amplification of cDNA ends; GSP, gene specific primers; SP, signal peptide; SDS-PAGE, sodium dodecyl sulfate- polyacrylamide gel electrophoresis; ORF, open reading frame; SPP, serine-proline-proline; ZIP, zipper; TM, trans-membrane; A-loop, activation loop. ⁎ Corresponding author. Tel.: + 86 20 85288262. E-mail addresses: [email protected] (J. Ma), [email protected] (Y. He). 0378-1119/$ – see front matter © 2012 Elsevier B.V. All rights reserved. doi:10.1016/j.gene.2012.03.013 Contents lists available at SciVerse ScienceDirect Gene journal homepage: www.elsevier.com/locate/gene
Transcript of Characterization and expression analysis of AGAMOUS-like, SEEDSTICK-like, and SEPALLATA-like...

Gene 500 (2012) 115–123
Contents lists available at SciVerse ScienceDirect
Gene
j ourna l homepage: www.e lsev ie r .com/ locate /gene
Characterization and expression analysis of AcSERK2, a somatic embryogenesis andstress resistance related gene in pineapple
Jun Ma a,b, Yehua He a,⁎, Zhongyi Hu a, Wentian Xu a, Jingxian Xia a, Cuihong Guo a, Shunquan Lin a, Li Cao a,Chengjie Chen a, Chenghou Wu a, Junli Zhang b
a Horticultural Biotechnology College of South China Agricultural University, Guangzhou, Guangdong 510642, Chinab College of Landscape Architecture of Sichuan Agricultural University, Chengdu, Sichuan 611100, China
Abbreviations: SERK, somatic embryogenesis recedichlorophenoxyacetic acid; SE, somatic embryogenesis;Boom; AGL15, AGAMOUS-like 15; WU, WUSCHEL; LRR-RLKkinases; LRRs, Leu-rich repeats; SA, salicylic acid; JA, jasmacid; CTAB, cetyl trimethylammonium bromide; RACE, rapGSP, gene specific primers; SP, signal peptide; SDS-Ppolyacrylamide gel electrophoresis; ORF, open reading fraZIP, zipper; TM, trans-membrane; A-loop, activation loop.⁎ Corresponding author. Tel.: +86 20 85288262.
E-mail addresses: [email protected] (J. Ma), h
0378-1119/$ – see front matter © 2012 Elsevier B.V. Alldoi:10.1016/j.gene.2012.03.013
a b s t r a c t
a r t i c l e i n f oArticle history:Accepted 4 March 2012Available online 21 March 2012
Keywords:Ananas comosusSERKSomatic embryogenesisExpression pattern
The isolation and characterization of AcSERK2, another somatic embryogenesis receptor-like kinase (SERK) genehomolog, confirmed that there was a SERK gene family in pineapple. AcSERK2 shared high similarity to AcSERK1and other SERKs, containing all the characteristic domains of the SERK family. The expressed recombinant pro-tein of pET-AcSERK2 was soluble. AcSERK2 expression was effectively induced by 2,4-dichlorophenoxyaceticacid (2,4-D) and maintained in high level at the early stage of somatic embryogenesis (SE). The results of insitu hybridization showed that AcSERK2 expressed at high level in the competent cells under the induction of2, 4-D and at low level during the development of meristematic center to globular embryo. This indicated thatAcSERK2 could be used as a potential marker gene tomonitor the acquisition of embryogenic competence. How-ever, AcSERK2 expressionwas also detected in all the tested organs and the expression level decreased alongwiththe development of calyx, ovule and ovary. These results suggested that AcSERK2 maybe play a broader role inorgan development. AcSERK2 transcription could be induced by exogenous hormones and abiotic stress. It wasdeduced that AcSERK2may participate in response to exogenous hormones and abiotic stress regulation in pine-apple. The expression pattern of AcSERK2 during SE differed from that of AcSERK1, and this indicated that theSERK genes may be non-redundant in somatic embryogenesis.
© 2012 Elsevier B.V. All rights reserved.
1. Introduction
Somatic embryogenesis was first demonstrated by Reinert (1958)and Steward et al. (1958). SE can be used both for rapid propagationof valuable clones and also for gene transfer and regeneration of genet-ically modified plants (Li et al., 2011). Research into SE intensified asplant regeneration in vitro came to be widely used in transformation(Nolan et al., 2003). Chimera formation is a common problem associat-ed with the propagation of transgenic plants derived using explant-based vegetative regeneration protocols. However, the origin of pineap-ple somatic embryos has been demonstrated to be unicellular (He et al.,2010), and thus the use of SE based transformation system may over-come the problemof chimera formation (Sanjeev et al., 2008). The cyto-metric analysis revealed that somatic embryo-derived plantlets have no
ptor-like kinase; 2,4-D, 2,4-LEC, Leafy cotyledon; BBM, Babys, Leu-rich repeat, receptor-likeonic acid; FAA, formalin aceticid amplification of cDNA ends;AGE, sodium dodecyl sulfate-me; SPP, serine-proline-proline;
[email protected] (Y. He).
rights reserved.
chromosomal absence, changes or alterations in ploidy (Fiuk et al.,2010). The pre-requisite for a somatic cell to change its usual fate ontoan embryogenic pathway, thereby transforming itself into a completeembryo, is poorly understood (Mordhorst et al., 1997). The molecularbasis of genetic and biochemical mechanisms leading to SE are notwell understood (Nolan et al., 2003; Pérez-Núňez et al., 2009). Thereare many reports of genes involved in the regulation of the later stagesof SE (Schmidt et al., 1997), such as LEC1 (Lotan et al., 1998), LEC2(Stone et al., 2001), BBM (Boutilier et al., 2002), WU (Zuo et al., 2002)and AGL15 (Harding et al., 2003). They can enhance the frequency ofSE and prolong the ability of SE. Butmuch less is known of the inductionprocess (Nolan et al., 2003). Many studies attempted to identify the re-lationship of triggering SE with specific gene expression patterns led tothe isolation of SE receptor-like kinase (SERK) gene (Baudino et al.,2001; Sanjeev et al., 2008). SERK belongs to Leu-rich repeat, receptor-like kinases (LRR-RLKs), a subgroup of protein kinases, characterizedby an extracellular domain with at least five Leu-rich repeats (LRRs), atransmembrane domain and an intracellular kinase domain. They arethought to play key roles in the transduction of extracellular signals inprocesses as diverse as plant development, disease resistance or self-incompatibility (Baudino et al., 2001; Krupa et al., 2006). One such pro-cess is SE where the embryogenic response of cultured cells dependsnot only on the cell type used but also on the composition of the culturemedium. SERK genes were considered to be multicellular-specific

116 J. Ma et al. / Gene 500 (2012) 115–123
genes. However, the finding of a close relative gene from unicellularalga implied that an SERK related gene that had been functioning inancestral conjugate algae had been recruited as a gene with a novelfunction related to SE during evolution from unicellular algae to multi-cellular plants (Sasaki et al., 2007). The first SERK genewas identified incarrot (DcSERK), as a result of the screening of differentially and specif-ically expressed genes in carrot embryogenic cell cultures (Schmidtet al., 1997). SERK expression starts in “competent” single cells andlasts up to the globular stage of embryogenesis. The expression ofDcSERK suggests that the SERK protein may transfer a signal in theculture medium and induce embryonic development while that proce-dure does not occur in cell without SERK expression (Baudino et al.,2001). Expressions of Arabidopsis AtSERK1 (Hecht et al., 2001), maizeZmSERK (Baudino et al., 2001) and Ananas comosus AcSERK1were sim-ilar to DcSERK in embryogenic structures, except ZmSERK and AcSERK1showed diverse expression pattern than that of DcSERK and AtSERKonly expressed in embryogenic structures. Plants transformed withAtSERK1 showed a marked increase in embryogenic capacity overwild-type cultures (Hecht et al., 2001). The absence of detectableLsSERK gene transcripts presented a reduction in their ability to formin vitro somatic embryonic strctures (Santos et al., 2009).The subcellu-lar location of AtSERK1was verified as amembrane protein (Shah et al.,2001a). Kinase activities were confirmed using kinase-associated pro-tein phosphatase (Shah et al., 2002). The ubiquitous presence of asmall SERK gene family has been identified in many plants, such asMedicago truncatula (Nolan et al., 2003), Helianthus annuus (Thomas etal., 2004), Oryza sativa (Hu et al., 2005), Citrus unshui (Shimada et al.,2005), soybean (Huang et al., 2010) and banana (Yang et al., 2011). Itwas reported that the SERK gene family probably played roles in em-bryogenesis as well as other developmental processes (Alam et al.,2010; Sanjeev et al., 2008; Song et al., 2008). Some of the SERKs havebeen demonstrated as a marker of embryogenic cells (Baudino et al.,2001; Hecht et al., 2001; Schmidt et al., 1997). Usually, an auxin is re-quired to induce SE, and subsequent auxin withdrawal or lowering ofthe auxin concentration is required for embryo maturation (Dudits etal., 1991). For example, 2,4-D is necessary and essential for Ananascomosus SE (He et al., 2010; Ma et al., 2012). In addition to their basicrole in somatic embryogenesis, OsSERK1, OsBISERK1 and MaSERK1arereported to partially mediate defense signal transduction leading to dis-ease resistance (Hu et al., 2005; Huang et al., 2010; Song et al., 2008).The transgenic lines absence of detectable LsSERK gene transcriptsshowed enhanced susceptibility to the pathogenic fungus (Santos etal., 2009).
Pineapple is a vegetative propagated crop depending on the suckersin practice for its self-sterile. But therewere only five suckers on a pine-apple plant per year on average, so the propagation ratio was slow. Wehave established an effective SE system of pineapple under 2,4-D induc-tion, the somatic embryo induction frequency reached 95%, and thenumber of somatic embryos per callus (3 mm×3 mm×3 mm) after40 days of culture was about 48 (He et al., 2007). These results con-firmed that SEwas a high efficientway for pineapple propagation. Infor-mation on the function of SERK in the enhancement of SE and itsdesignation as a molecular marker for SE in other plant species led usto hypothesize a significant role for AcSERK2 as factors contributingtowards SE in pineapple.
In our previous study, a SERK gene (AcSERK1) was isolated frompineapple, and the expression of AcSERK1 was effectively induced by2,4-D during SE. This gene can be used as a marker of embryogeniccompetent cells (Ma et al., 2012). In present study, another SERKgene, AcSERK2, was identified in pineapple. Similar to AcSERK1, it con-tains all the characteristics of SERK gene family. The expression ofAcSERK2 in response to 2, 4-D in culture callus was also investigatedto provide some insight into the regulation of SE. AcSERK2 was syner-gistically increased when 2, 4-D was supplied in the culture medium.The expression of AcSERK2was not specifically associated with SE andit may play a broader role in morphogenesis. It also can be detected in
somatic organs and can be induced in the callus after treatment withexogenous hormones or abiotic stress.
2. Materials and methods
2.1. Plant material
Non-embryogenic callus derived from leaf-base of Ananas comosus.cv. Shenwan on 2, 4-D free medium was randomly divided into twogroups. One was transferred to 2, 4-D-containing medium for SE in-duction, and the other was maintained on 2, 4-D-free medium actingas negative control for real-time PCR. Batches of cultured callus wereperiodically used to extract total RNA for the relevant stage, at 0, 5,10, 15, 20, 25, 30, 35, 40, 45 and 50 days after incubation on 2, 4-Dcontaining medium. Negative control samples were taken from calluscultured without 2, 4-D at 0, 15, 25, 35 and 45 days. Different organs(root, stem, leaf, calyx, bract, petal, ovary, ovule and anther) wereused to extract total RNA to detect the AcSERK2 expression level byreal-time PCR. The ovule, ovary and calyx were periodically used toextract total RNA for the relevant stage, a week before flower bloom-ing, at flower blooming, a week after flower blooming, two weeksafter flower blooming and four weeks after flower blooming, tostudy the AcSERK2 expression during the development of these or-gans. Callus underwent different treatments with hormones andabiotic stress for 24 hours were used to isolate RNA for AcSERK2expression study: 0.1 mM salicylic acid (SA), 0.1 mM jasmonic acid(JA), NaCl (3%w/v) and ethylene (1000 mg L−1) were used by sprayingonto the callus. Callus were incubated in high temperature (40 °C) orlow temperature (4 °C), or in room temperature (25 °C) after injurytreatment. All the tissue samples obtained were immersed into liquidnitrogen immediately for quick-freeze and then stored at −80 °C forRNA extraction. Embryogenic callus under 2,4-D induction were fixedin formalin acetic acid (FAA) for in situ hybridization.
2.2. RNA/DNA extraction and cDNA synthesis
Total RNA extractions from the tissuematerial mentioned in “PlantMaterial” were carried out using TRIzol reagents (Takara) followingthe recommendations of themanufacturer. Each RNA samplewas sub-jected to DNase digestion (Takara) to remove any remaining DNA.Poly (A)+ RNA was isolated from total RNA using mRNA PurificationKit (Takara). For first strand cDNA synthesis, 2 μg of Poly (A)+ RNAwas reverse transcribed in a final volume of 20 μl using oligodT (18)and M-MLV reverse transcriptase (Invitrogen). Genomic DNA wasextracted from callus cultured in vitro by cetyI trimethylammoniumbromide (CTAB) following the method of (Murray and Thompson,1980) and treated with RNase A to eliminate the presence of RNA.
2.3. Isolation of AcSERK2
Degenerate primers (P1, P2, P3 and P4, listed in SupplementaryTable 1), designed to conserve SERK regions, and were employed toamplify pineapple SERK fragments. The 5′ and 3′ sequences of SERKwas amplified by rapid amplification of cDNA ends (RACE) using the3′-Full RACE Core Set Ver.2.0 kit and 5′-Full RACE Core Set Ver.2.0kit (Takara) according to the manufacturer's instructions. The nestedgene specific primers (GSP) for 3′ RACE were P5-P6 and the primersfor 5′ RACE were P7-P8. Sequences were edited, aligned and analyzedusing DNAMAN and CLUSTAL software tools. Specific primers P9 andP10 were designed according to the results of the RACE to amplify thefull-length cDNA sequence and genomic sequence.
2.4. Bioinformatics analysis
Open reading frame (ORF) was predicted using the “ORF Finder”programme (http://www.ncbi.nlm.nih.gov/gorf/gorf.html). The signal

117J. Ma et al. / Gene 500 (2012) 115–123
peptide was predicted by using the “SignalP 3.0 Server” tool (http://www.cbs.dtu.dk/servicesSignalP/). The transmembrane region (TM) waspredicted by TMpred (http://www.ch.embnet.org/software/TMPRED_form.html). Protein predictions were performed using the prediction ofprotein sorting signals and localisation sites in amino acid sequences(PSORT) available at http://psort.nibb.ac.jp/. Use InterProScan (Zdobnovand Apweiler, 2001), PlantsP (http://plantsp.genomics.purdue.edu/html/feature_scan.html) and ScanProsite (http://us.expasy.org/prosite/) to an-alyze the function structure of AcSERK2 protein (Zdobnov and Apweiler,2001). Multiple sequence aligment was performed with DNAMAN soft-ware. Phylogenetic trees were constructed using the MEGA 4 program.
2.5. Construction of the expression vector pET-AcSERK2
After digested with two restriction enzymes (EcoRI and HindIII), thefull-length AcSERK2 gene without the signal peptide (SP) was sub-cloned into the pET-30a vector. The vectorwas digested by the same en-zymes. The recombinant plasmid (pET-AcSERK2) was then introducedinto E. coli BL21(DE3) pLysS competent cells and then sequenced fromboth sides.
2.6. Expression of the recombinant protein from E. coli
A single colony of an E. coli strain BL21 (DE) harboring the pET-30aplasmid was cultured at 37 °C overnight in Luriabetani liquid mediumcontaining 50 μg/ml kanamycin. Then the cultured cells were trans-ferred to fresh medium and shaken at 150 rpm for another 2–3 huntil the optical density (OD600) reached 0.5. Then the Isopropyl-β-D-thiogalactoside was added to the medium to induce the expression ofthe protein. Bacteria were harvested by centrifugation at 4000 rpm for5 min and resuspended in 2 ml splitting buffer, and then ultrasonicatedat 4 °C. The ultrasonication product was centrifuged at 12,000 rpmfor 10 min at 4 °C. 50 μl of precipitation and supernatant respectivelywere assessed using sodium dodecyl sulfate–polyacrylamide gel elec-trophoresis (SDS–PAGE). The expressed pET-AcSERK2 fusion proteinas separated by 12% SDS-PAGE. The protein bands were visualizedafter being stained with 0.1 M KCl solution pre-cooled.
2.7. Quantitative real time PCR analysis
For gene expression analysis, first-stranded cDNAs from the callusdescribed in “Plant material” were used as templates. The relative ex-pression of AcSERK2 was measured using THUNDERBIRD SYBR qPCRMix (Toyobo) according to the manufacturer's instructions performedon the iQ5 real-time PCR System (Biorad). Real time PCR was per-formed with gene-specific primers of AcSERK2. A gene encoding apineapple β-actin was used as the endogenous control. Amplificationwas carried out with the following cycling parameters: heating for2 min at 95 °C, 40 cycles of denaturation at 95 °C for 10s, annealingfor 20 s at 52 °C, and extension at 72 °C for 35 s. Triplicate quantitativePCR experiments were performed for each sample, and the expressionvalues obtained were normalized against β-actin. Analysis of the rela-tive gene expression data was conducted using the 2−ΔΔCt method(Livak and Schmittgen, 2001). To prevent amplification of any con-taminating genomic DNA, RNA preparations were treated with DNaseIand the probe primers were designed over an exon/intron boundaryin the cDNA sequence. Specificity of the amplifications was verifiedby electrophoresis and at the end of the PCR run by melting curveanalysis. Primer sequences used for AcSERK2 and endogenous controlare listed in Table 1. Real-time PCR results were analyzed using theiQ5 System Software.
2.8. RNA in situ hybridization and detection
RNA in situ hybridizationwas performed according to the proceduresfor in situ hybridization to chromosomes, cells, and tissue sections
(Roche). Different tissues were fixed overnight in FAA (100% formalin:100% acetic acid: 50% ethanol, in proportions of 5:5:90) at 4 °C for 12 h,and dehydrated through ethanol. Fixed tissues were embedded in paraf-fin and sectioned at 7–8 μm with a rotary microtome. The sense andantisense probes were transcribed with cDNA fragments cloned inpSPT 19 vector (Roche) and were labeled with the digoxigenin-UTP bySP6 or T7 RNApolymerase in vitro transcription kit according to the tech-nical manual (Roche). Detection was carried out with anti-digoxigenin-AP and NBT/BCIP ready-to-use tablets (Roche), and the hybridizationsignal was observed by microscope (Olympus).
3. Results
3.1. Cloning of AcSERK2
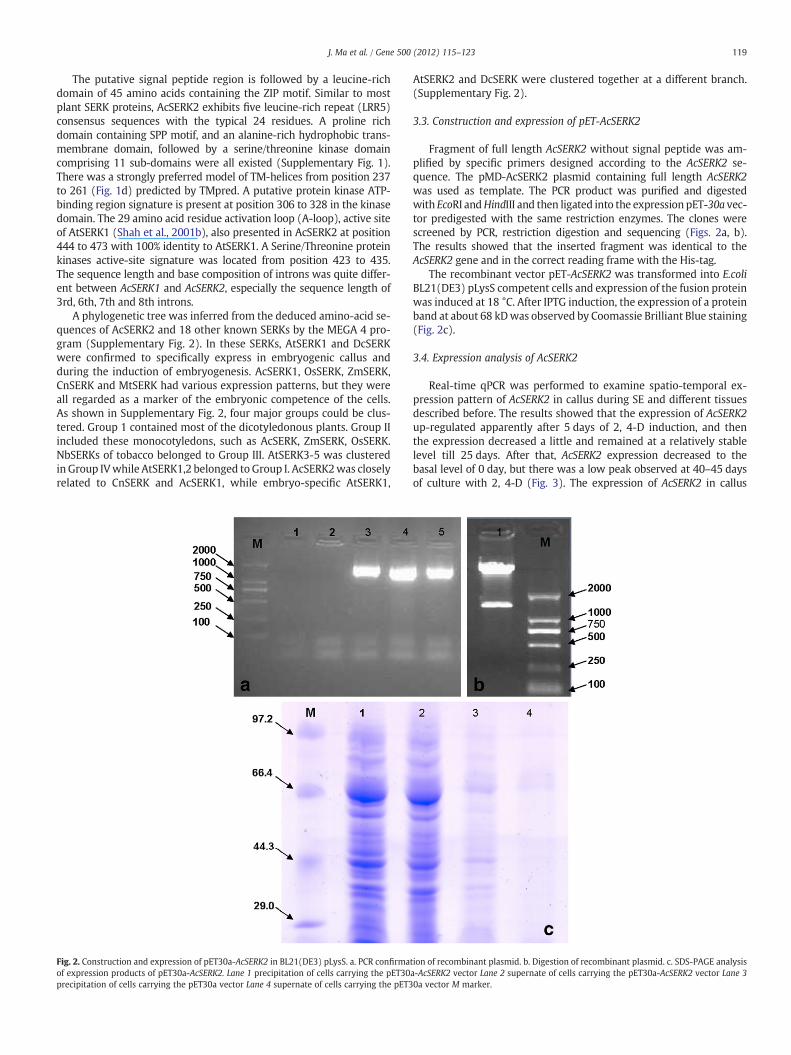
Degenerate primers designed from SERK conserved domains wereused to amplify potential SERK gene fragments from pineapple. Specificprimers designed from the putative AcSERK fragment was used toamplify both 5′ and 3′ ends of AcSERK by rapid-amplification of cDNAends (RACE). The sequence was then verified by obtaining a full-length cDNA in a single PCR reaction. Amplified PCR products werepurified, cloned, and then sequenced. The full-length mRNA sequenceof AcSERK2 (Genbank accession number HM236376) was comprisedof 2045 bp, containing an 1875-bp open reading frame. Subsequently,the full-length sequence of genomic DNA containing 5716 bp was am-plified using the same primer pair as cDNA, and its accession numberwas JN672682.
3.2. Sequence analysis
The in silico translation of AcSERK2 yields an ORF contains 624amino acid. The complete coding sequence encodes a putative68.9kD protein with a theoretical pI of 5.38. The protein contains 56strongly basic (+) amino acids (K+R), 71 strongly acidic (−) aminoacids (D+E), 229 hydrophobic amino acids (A, I, L, F, W and V), and152 polar amino acids (N, C, Q, S, T and Y). The molecular formula isC3074H4845N843O910S24.
A BLAST search of AcSERK2 resulted in several SERK similarity hitsfrom other plant species, including Cocos nucifera (AY791293.2),O. sativa (AB188247.1), Zea mays (NM001111663.1), Hordeum vulgaresubsp. Vulgare (EF216861.1) and Triticum aestivum (EU679373.1).AcSERK2 showed an identity of more than 75% with other SERKs,such as CnSERK (Cocos nucifera), HvSERK (Hordeum vulgare), OsSERK1(O. sativa Japonica Group) and ZmSERK1 (Zea mays) (SupplementaryFig. 1). The identity of nucleotide sequence of AcSERK2 with AcSERK1was 81%, while that of the putative amino acid sequence was 92%.High similarity of AcSERK2 to other SERKs was observed in the LeucineZipper region and Kinase domains, while weak similarity was observedin the serine-proline-proline (SPP) region (Supplementary Fig. 1).
A comparative alignment of genomic DNA sequence of AcSERK2with the corresponding predicted coding sequences revealed thatAcSERK2 consisted of 11 exons interspersed by 10 introns (Fig. 1a).This exon/intron structure is quite similar to AcSERK1 (Ma et al., 2012)and other SERKs (Sharma et al., 2008; Stone et al., 2001). Each LRR do-main is encoded by a separate exon, except for LRR2 and LRR3, whichshared a single exon. The 11 kinase sub-domains are contained inexons 9–11 with the Xth sub-domain sharing the last two exons.
AcSERK2 contained all the conserved domains present in the SERKprotein from other species (Fig. 1b). The SignalP3.0 program was usedto detect signal peptide sequence and the predicted first 23 aminoacids (MVVMAPWAVWFMLVIHPFLRVWA) of AcSERK2 at the N-terminus were found to correspond to a signal peptide, with a mostlikely signal peptide cleavage site between the amino acid residues 23and 24 (Fig. 1c). Compared to AcSERK1, AcSERK2 lacked six aminoacids in signal peptide region. This may influence the subcellular local-ization of AcSERK2.
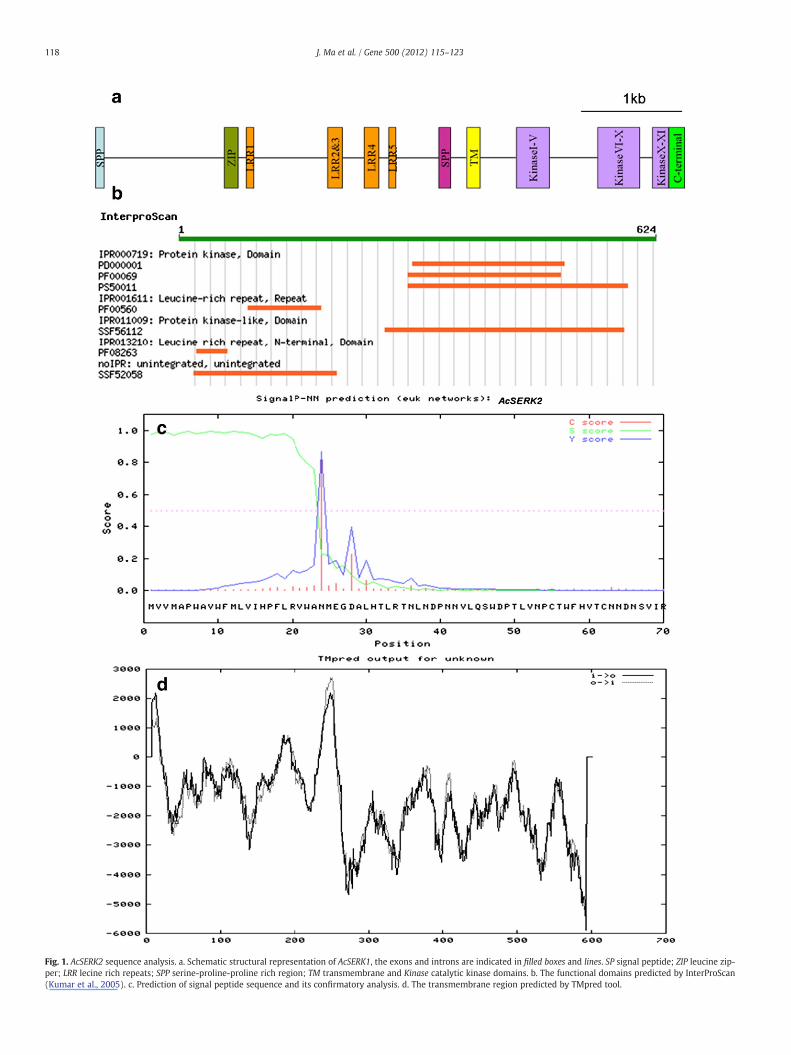
Fig. 1. AcSERK2 sequence analysis. a. Schematic structural representation of AcSERK1, the exons and introns are indicated in filled boxes and lines. SP signal peptide; ZIP leucine zip-per; LRR lecine rich repeats; SPP serine-proline-proline rich region; TM transmembrane and Kinase catalytic kinase domains. b. The functional domains predicted by InterProScan(Kumar et al., 2005). c. Prediction of signal peptide sequence and its confirmatory analysis. d. The transmembrane region predicted by TMpred tool.
118 J. Ma et al. / Gene 500 (2012) 115–123
119J. Ma et al. / Gene 500 (2012) 115–123
The putative signal peptide region is followed by a leucine-richdomain of 45 amino acids containing the ZIP motif. Similar to mostplant SERK proteins, AcSERK2 exhibits five leucine-rich repeat (LRR5)consensus sequences with the typical 24 residues. A proline richdomain containing SPP motif, and an alanine-rich hydrophobic trans-membrane domain, followed by a serine/threonine kinase domaincomprising 11 sub-domains were all existed (Supplementary Fig. 1).There was a strongly preferred model of TM-helices from position 237to 261 (Fig. 1d) predicted by TMpred. A putative protein kinase ATP-binding region signature is present at position 306 to 328 in the kinasedomain. The 29 amino acid residue activation loop (A-loop), active siteof AtSERK1 (Shah et al., 2001b), also presented in AcSERK2 at position444 to 473 with 100% identity to AtSERK1. A Serine/Threonine proteinkinases active-site signature was located from position 423 to 435.The sequence length and base composition of introns was quite differ-ent between AcSERK1 and AcSERK2, especially the sequence length of3rd, 6th, 7th and 8th introns.
A phylogenetic tree was inferred from the deduced amino-acid se-quences of AcSERK2 and 18 other known SERKs by the MEGA 4 pro-gram (Supplementary Fig. 2). In these SERKs, AtSERK1 and DcSERKwere confirmed to specifically express in embryogenic callus andduring the induction of embryogenesis. AcSERK1, OsSERK, ZmSERK,CnSERK and MtSERK had various expression patterns, but they wereall regarded as a marker of the embryonic competence of the cells.As shown in Supplementary Fig. 2, four major groups could be clus-tered. Group 1 contained most of the dicotyledonous plants. Group IIincluded these monocotyledons, such as AcSERK, ZmSERK, OsSERK.NbSERKs of tobacco belonged to Group III. AtSERK3-5 was clusteredin Group IVwhile AtSERK1,2 belonged to Group I. AcSERK2was closelyrelated to CnSERK and AcSERK1, while embryo-specific AtSERK1,
Fig. 2. Construction and expression of pET30a-AcSERK2 in BL21(DE3) pLysS. a. PCR confirmaof expression products of pET30a-AcSERK2. Lane 1 precipitation of cells carrying the pET30precipitation of cells carrying the pET30a vector Lane 4 supernate of cells carrying the pET3
AtSERK2 and DcSERK were clustered together at a different branch.(Supplementary Fig. 2).
3.3. Construction and expression of pET-AcSERK2
Fragment of full length AcSERK2 without signal peptide was am-plified by specific primers designed according to the AcSERK2 se-quence. The pMD-AcSERK2 plasmid containing full length AcSERK2was used as template. The PCR product was purified and digestedwith EcoRI andHindIII and then ligated into the expression pET-30a vec-tor predigested with the same restriction enzymes. The clones werescreened by PCR, restriction digestion and sequencing (Figs. 2a, b).The results showed that the inserted fragment was identical to theAcSERK2 gene and in the correct reading frame with the His-tag.
The recombinant vector pET-AcSERK2 was transformed into E.coliBL21(DE3) pLysS competent cells and expression of the fusion proteinwas induced at 18 °C. After IPTG induction, the expression of a proteinband at about 68 kDwas observed by Coomassie Brilliant Blue staining(Fig. 2c).
3.4. Expression analysis of AcSERK2
Real-time qPCR was performed to examine spatio-temporal ex-pression pattern of AcSERK2 in callus during SE and different tissuesdescribed before. The results showed that the expression of AcSERK2up-regulated apparently after 5 days of 2, 4-D induction, and thenthe expression decreased a little and remained at a relatively stablelevel till 25 days. After that, AcSERK2 expression decreased to thebasal level of 0 day, but there was a low peak observed at 40–45 daysof culture with 2, 4-D (Fig. 3). The expression of AcSERK2 in callus
tion of recombinant plasmid. b. Digestion of recombinant plasmid. c. SDS-PAGE analysisa-AcSERK2 vector Lane 2 supernate of cells carrying the pET30a-AcSERK2 vector Lane 30a vector M marker.

120 J. Ma et al. / Gene 500 (2012) 115–123
cultured on 2, 4-D free culturemedium,whichwere used to be negativecontrol, was relatively stable, the expression levels were close to that of0 day, and these calluses were regarded as non-embryonic callus (Heet al., 2007).
The expression of AcSERK2 in stem, leaf, calyx, petal and bract werein the similar level, andwere higher than that in ovary and anther. Thehighest expression level was observed in the roots while the lowestlevel was in the anther. Along with the development of ovule, ovaryand calyx, the expression of AcSERK2 in all these tissues decreased, es-pecially in the calyx. After the bloom of the flower, the expression ofAcSERK2 in the calyx decreased to 1/4 of the level before bloom. How-ever, the expression of AcSERK2 along with the development of ovulewas relatively stable, and maintained at relative high level (Fig. 3).
The AcSERK2 transcript expression was increased following treat-ment with JA, NaCl, SA and after the treatment of low temperatureand injure. However, the expression of AcSERK2 was decreased afterthe treatment of ethylene and high temperature (Fig. 4).
Results of in situ hybridization showed that intense AcSERK2 ex-pression signal was observed in the single competent cells (Fig. 5a)and competent cell clusters after 5 days of culture on 2, 4-D containingmedium (Fig. 5b). After 10 days of culture, these competent cell clus-ters grew bigger through cell division to form meristematic center(Fig. 5c). At the subsequent 30 days of culturewith 2, 4-D, themeriste-matic centers grew outwards to form pro-embryos (Figs. 5d, e) andthen broke through the periphery of the callus to form globular em-bryos (Figs. 5f, g). However, the AcSERK2 expressing signal tested inthis process was weak (Figs. 5d–g). The cells with AcSERK2 expressionwere small, isodiametric, non-vacuolated and cytoplasm-rich, indicat-ing a high metabolic activity. When use the sense probe for hybridiza-tion no signal was detected in any region of the calluses (Fig. 5h).
4. Disscussion
SE is an effective protocol for plant propagation and genetic trans-formation (Firoozabady et al., 2006; Kumar et al., 2005; Wu et al.,2008). Study on the molecular mechanism of SE in cultures will aidin the understanding of embryogenesis and the establishment ofmore effective regeneration and transformation system of pineapple.
The isolation of AcSERK1 and AcSERK2 confirmed that there was agene family of SERK in pineapple. Sequence analysis suggested that,similar to AcSERK1, AcSERK2 encodes a SERK protein belonging to theLRR-RLK family (Walker, 1994), with all the characteristic domainsof the SERK family. The high sequence similarity (84.4%) of AcSERK2with AtSERK1may suggest that the corresponding regions in AcSERK2may have similar role to AtSERK1. An interesting question has beenput forward whether these sequence similarities can account for
Fig. 3. Relative expression of AcSERK2measured using real-time PCR. Embryogenic callus showcallus shows AcSERK2 expression in callus cultured on medium without 2,4-D used as negatof ovule, ovary and calyx shows AcSERK2 expression during the development of ovule, ovary
their functional redundancy (Cai et al., 2011). The TM region separatesthe extracellular domains from the intracellular region. The two thre-onine residues in the A-loop have been regarded as essential for bothautophosphorylation and phosphorylation of artificial substrates(Shah et al., 2001b). The C-terminal leucine-rich domain, consideredas a specific region of SERK, is suggested to play a role in protein-protein interactions (Schmidt et al., 1997). RLKs exist as monomersuntil binding of an extracellular signal molecule induces receptor di-merization. This brings the intracellular kinase domains of individualmonomers into close proximity, allowing transphosphorylation, whichactivates the kinase domains and causes the regulation of cellular re-sponse (Becraft, 2002).
AcSERK2 clustered together with CnSERK (Pérez-Núňez et al.,2009), AcSERK1 (Ma et al., 2012), OsSERK (Hu et al., 2005) andZmSERK1 (Baudino et al., 2001), and it clustered closer to AtSERK1,2 than AtSERK3-5. These results suggested that AcSERK2 probablyplay a role in SE as which has been proposed for the orthologs inthese species. Alignment of AcSERK1 and AcSERK2 showed that theidentity of the two sequences was 92% and most of the differentialamino acids were located in the signal peptide, the SPP domain andthe C terminal region. The AcSERK2 lacked 6 amino acid residues atthe signal peptide region compared to AcSERK1. This may affect theexpression patterns and subcellular localization of AcSERK2. It wouldbe of interest to knowwhether they have similar or different functionsto perform. It is notable that AcSERK2 clustered closer to CnSERK thanto AcSERK1. This may indicate that AcSERK2 may have some differentexpression patterns or functions from AcSERK1. The highly conservedexons and exon/intron structure points to the functional significanceof this gene family (Sharma et al., 2008). However, the sequencelength and base composition of the introns of AcSERK2 were quitedifferent from AcSERK1 and other SERKs (Ma et al., 2012; Sharma etal., 2008; Stone et al., 2001). It is reported that the introns may playa role in the mediation of gene expression (Gregoise and Romeo,1999; Tosi, 1998).
We constructed an expression vector pET-AcSERK2 to detect thecharacteristic of AcSERK2 protein. A band of about 68 kDwas observedby SDS-PAGE. The observed molecular size was coincident with thesize predicted from sequence data. The corresponding band was notfound in the total cellular protein of vector pET-30a. Therefore the pro-tein was considered to be the product of recombinant AcSERK2. Thetarget band was observed both in precipitation and supernatant ofcell lysate after sonication. It was confirmed that the expressed recom-binant protein was soluble, and the expressed recombinant proteinwas distributed in soluble and insoluble fractions.
AcSERK2 expression was effectively induced after 5 days of culturewith 2, 4-D, which was earlier than AcSERK1, and this was the highest
s AcSERK2 expression in callus cultured on 2,4-D containing medium. Non-embryogenicive control. Different organs shows AcSERK2 expression in different organs. Developmentand calyx.
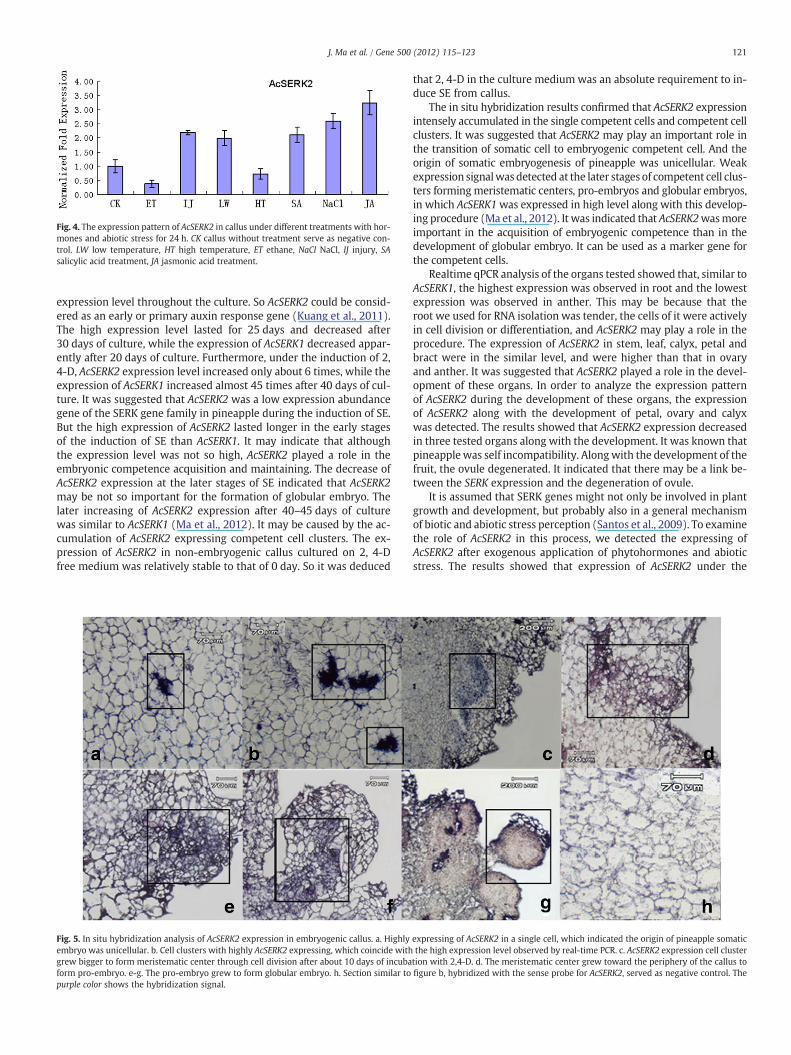
Fig. 4. The expression pattern of AcSERK2 in callus under different treatments with hor-mones and abiotic stress for 24 h. CK callus without treatment serve as negative con-trol. LW low temperature, HT high temperature, ET ethane, NaCl NaCl, IJ injury, SAsalicylic acid treatment, JA jasmonic acid treatment.
121J. Ma et al. / Gene 500 (2012) 115–123
expression level throughout the culture. So AcSERK2 could be consid-ered as an early or primary auxin response gene (Kuang et al., 2011).The high expression level lasted for 25 days and decreased after30 days of culture, while the expression of AcSERK1 decreased appar-ently after 20 days of culture. Furthermore, under the induction of 2,4-D, AcSERK2 expression level increased only about 6 times, while theexpression of AcSERK1 increased almost 45 times after 40 days of cul-ture. It was suggested that AcSERK2 was a low expression abundancegene of the SERK gene family in pineapple during the induction of SE.But the high expression of AcSERK2 lasted longer in the early stagesof the induction of SE than AcSERK1. It may indicate that althoughthe expression level was not so high, AcSERK2 played a role in theembryonic competence acquisition and maintaining. The decrease ofAcSERK2 expression at the later stages of SE indicated that AcSERK2may be not so important for the formation of globular embryo. Thelater increasing of AcSERK2 expression after 40–45 days of culturewas similar to AcSERK1 (Ma et al., 2012). It may be caused by the ac-cumulation of AcSERK2 expressing competent cell clusters. The ex-pression of AcSERK2 in non-embryogenic callus cultured on 2, 4-Dfree medium was relatively stable to that of 0 day. So it was deduced
Fig. 5. In situ hybridization analysis of AcSERK2 expression in embryogenic callus. a. Highlyembryo was unicellular. b. Cell clusters with highly AcSERK2 expressing, which coincide withgrew bigger to form meristematic center through cell division after about 10 days of incubaform pro-embryo. e-g. The pro-embryo grew to form globular embryo. h. Section similar topurple color shows the hybridization signal.
that 2, 4-D in the culture medium was an absolute requirement to in-duce SE from callus.
The in situ hybridization results confirmed that AcSERK2 expressionintensely accumulated in the single competent cells and competent cellclusters. It was suggested that AcSERK2 may play an important role inthe transition of somatic cell to embryogenic competent cell. And theorigin of somatic embryogenesis of pineapple was unicellular. Weakexpression signalwasdetected at the later stages of competent cell clus-ters forming meristematic centers, pro-embryos and globular embryos,in which AcSERK1was expressed in high level along with this develop-ing procedure (Ma et al., 2012). It was indicated that AcSERK2wasmoreimportant in the acquisition of embryogenic competence than in thedevelopment of globular embryo. It can be used as a marker gene forthe competent cells.
Realtime qPCR analysis of the organs tested showed that, similar toAcSERK1, the highest expression was observed in root and the lowestexpression was observed in anther. This may be because that theroot we used for RNA isolation was tender, the cells of it were activelyin cell division or differentiation, and AcSERK2 may play a role in theprocedure. The expression of AcSERK2 in stem, leaf, calyx, petal andbract were in the similar level, and were higher than that in ovaryand anther. It was suggested that AcSERK2 played a role in the devel-opment of these organs. In order to analyze the expression patternof AcSERK2 during the development of these organs, the expressionof AcSERK2 along with the development of petal, ovary and calyxwas detected. The results showed that AcSERK2 expression decreasedin three tested organs along with the development. It was known thatpineapplewas self incompatibility. Alongwith the development of thefruit, the ovule degenerated. It indicated that there may be a link be-tween the SERK expression and the degeneration of ovule.
It is assumed that SERK genes might not only be involved in plantgrowth and development, but probably also in a general mechanismof biotic and abiotic stress perception (Santos et al., 2009). To examinethe role of AcSERK2 in this process, we detected the expressing ofAcSERK2 after exogenous application of phytohormones and abioticstress. The results showed that expression of AcSERK2 under the
expressing of AcSERK2 in a single cell, which indicated the origin of pineapple somaticthe high expression level observed by real-time PCR. c. AcSERK2 expression cell clustertion with 2,4-D. d. The meristematic center grew toward the periphery of the callus tofigure b, hybridized with the sense probe for AcSERK2, served as negative control. The

122 J. Ma et al. / Gene 500 (2012) 115–123
treatment of JA or SA was up-regulated. This is similar to patterns ofOsSERK1 (Hu et al., 2005) and MaSERK1 (Huang et al., 2010). SA andJA were assumed to act as a central regulator of defense against path-ogens and insect attack (Huang et al., 2010; Thomma et al., 1998). Soit was suggested that AcSERK2 may partially mediate the defense sig-nal transduction and play a role in disease resistance response. Thiswas coinciding with the opinion of Baudino et al. (2001) and Krupaet al. (2006).We also tested expression pattern of AcSERK2 in responseto abiotic stress. The results showed that AcSERK2 expression wasup-regulated under cold, salt and injure stress. This indicated thatAcSERK2 may participate in abiotic stress regulation in pineapple.However, the expression of AcSERK2 under the stress of high temper-ature only decreased a little. This may be because pineapple is a tropicplant, it is insensitive to high temperature or it has high tolerance tohigh temperature. The decreasing of AcSERK2 under stress of ethyleneindicated that enthane may suppress the transcription of AcSERK2.
In conclusion, the expression of AcSERK2 accumulated during the in-duction of SE. It can be used as a mark gene for competence acquisition.AcSERK2 was expressed in somatic organs. It may have complicatedfunction in different aspects of the plant growth and development.AcSERK2 transcripts can be induced by exogenous hormones and abioticstress. Both AcSERK2 and AcSERK1 expressed during embryogenesis andsomatic organ development, but there were differences in sequencestructure and expression pattern. It was indicated that SERK membersmay play different roles in the development of plant and the SERKs inpineapple probably were non-redundant. It was meaningful to studyif there were other SERKmembers in pineapple and the relationship be-tween the SERKs.
Acknowledgments
This research was supported by the Natural Science Foundation ofChina (30971984), Project 948 of Ministry of Agriculture (2010-G2-11), Commonweal Industry Scientific Research Poject of Ministry ofAgriculture (nyhyzx07-30), and Open Found Project of Key Laboratoryof Utilization of Tropical Crop Germplasm Resources, Ministry of Agri-culture (KFKT-2010-07).
Appendix A. Supplementary data
Supplementary data to this article can be found online at doi:10.1016/j.gene.2012.03.013.
References
Alam, M.M., et al., 2010. A putatuve leucine-rich repeat receptor-like kinase of juteinvolved in stress response. Plant Mol. Biol. Rep. 28, 394–402.
Baudino, S., et al., 2001. Molecular characterization of two novel maize LRR receptor-like kinases, which belong to the SERK gene family. Planta 213, 1–10.
Becraft, P.W., 2002. Receptor kinase signaling in plant development. Annu. Rev. CellDev. Biol. 18, 163–192.
Boutilier, K., et al., 2002. Ectopic expression of BABYBOOM triggers a conversion fromvegetative to embryonic growth. Plant Cell 14, 1737–1749.
Cai, H.S., Tian, S., Liu, C.L., Dong, H.S., 2011. Identification of a MYB3R gene involved indrought, salt and cold stress in wheat (Triticum aestivum L.). Gene 485, 146–152.
Dudits, D., Bogre, L., Gyorgyey, J., 1991. Molecular and cellular approaches to the anal-ysis of plant embryo development from somatic cells in vitro. J. Cell Sci. 99,475–484.
Firoozabady, E., Heckert, M., Gutterson, N., 2006. Transformation and regeneration ofpineapple. Plant Cell Tissue Organ Cult. 84, 1–16.
Fiuk, A., Bednarek, P.T., Rybczynski, J.J., 2010. Flowcytometry, HPLC-RP, andmetAFLP anal-yses to assess genetic variability in somatic embryo-derived plantlets of Gentianapannonica Scop. Plant Mol. Biol. Rep. 28, 413–420.
Gregoise, J.M., Romeo, P.H., 1999. T-cell expression of the human GATA-3 gene is regu-lated by a non-lineage-specific silencer. J. Biol. Chem. 274, 6567–6578.
Harding, E.W., Tang, W.N., Nichols, K.W., Fernandez, D.E., Perry, S.E., 2003. Expressionand maintenance of embryogenic potential is enhanced through constitutive ex-pression of AGAMOUS-Like15. Plant Physiol. 133, 653–663.
He, Y.H., et al., 2007. Somatic embryogenesis from leaf base callus of Ananas comosus.J. Fruit Sci. 24, 59–63 (in Chinese).
He, Y.H., Fang, S.Q., Ma, J., Hu, Z.Y., Lu, M., Peng, B., 2010. Histocytology Observation onthe Somatic Embryogenesis in Ananas comosus Callus. Acta Hortic. Sin. 37, 689–696(in Chinese).
Hecht, V., et al., 2001. The Arabidopsis SOMATIC EMBRYOGENESIS RECEPTOR KINASE 1gene is expressed in developing ovules and embryos and enhances embryogeniccompetence in culture. Plant Physiol. 127, 803–816.
Hu, H., Xiong, L., Ynag, Y., 2005. Rice SERK1 gene positively regulates somatic embryo-genesis of cultured cell and host defense response against fungal infection. Planta222, 107–117.
Huang, X., et al., 2010.MaSERK1 Gene Expression Associated with Somatic EmbryogenicCompetence and Disease Resistance Response in Banana (Musa spp.). Plant Mol.Biol. Rep. 28, 309–316.
Krupa, A., Anamika, Srinivasan, N., 2006. Genome-wide comparative analyses of do-main organization of repertoires of protein kinases of Arabidopsis thaliana andOryza sativa. Gene 380, 1–13.
Kuang, J.F., et al., 2011. Two GH3 genes from longan are differentially regulated duringfruit growth and development. Gene 485, 1–6.
Kumar, K.K., Maruthasalam, S., Loganathan, M., Sudhakar, D., Balasubramanian, P.,2005. An Improved Agrobacterium-Mediated Transformation Protocol for Recalci-trant Elite Indica Rice Cultivars. Plant Mol. Biol. Rep. 23, 67–73.
Li, H.L., Wang, Y., Guo, D., Tian, W.M., Peng, S.Q., 2011. Three MADS-box genes of Heveabrasiliensis expressed during somatic embryogenesis and in the laticifer cells. Mol.Biol. Rep. 38, 4045–4052.
Livak, K.J., Schmittgen, T.D., 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2−△△Ct method. Methods 25, 402–408.
Lotan, T., et al., 1998. Arabidopsis LEAFYCOTYLEDON1 is sufficient to induce embryo de-velopment in vegetative cells. Cell 93, 1195–1205.
Ma, J., He, Y.H., Wu, C.H., Liu, H.P., Hu, Z.Y., Shun, G.M., 2012. Cloning and molecularcharacterization of a SERK gene transcriptionally induced during somatic embryo-genesis in Ananas comosus. cv. Shenwan. Plant Mol. Biol. Rep. 30, 195–203.
Mordhorst, A.P., Toonen, M.A.J., de Vries, S.C., 1997. Plant embryogenesis. Crit. Rev.Plant Sci. 16, 535–576.
Murray, M.G., Thompson, W.F., 1980. Rapid isolation of high molecular weight plantDNA. Nucleic Acids Res. 8, 4321–4325.
Nolan, K.E., Irwanto, R.R., Rose, R.J., 2003. Auxin up-regulates MtSERK1 expression inboth Medicago truncatula root-forming and embryogenic cultures. Plant Physiol.133, 218–230.
Pérez-Núňez, M.T., Souza, R., Sáenz, L., Chan, J.L., Zúňiga-Aguilar, J.J., Oropeza, C., 2009.Detection of a SERK-like gene in coconut and analysis of its expression duringthe formation of embryogenic callus and somatic embryos. Plant Cell Rep. 28,11–19.
Reinert, J., 1958. Morphogenese und ihre kontrolle a gewebekulturen aus carotten.Naturwissenschaften 45, 344–345.
Sanjeev, K.S., Steve, M., Ingo, H., Glenn, J.B., 2008. Cloning and molecular characterisa-tion of a potato SERK gene transcriptionally induced during initiation of somaticembryogenesis. Planta 228, 319–330.
Santos, M.O., Romano, E., Vieira, L.S., Baldoni, A.B., Aragao, F.J.L., 2009. Suppression ofSERK gene expression affects fungus tolerance and somatic embryogenesis intransgenic lettuce. Plant Biol. 11, 83–89.
Sasaki, G., et al., 2007. Multiple receptor-like kinase cDNAs from liverwort Marchantiapolymorpha and two charophycean green algae, Closterium ehrenbergii and Nitellaaxillaris: Extensive gene duplications and gene shufflings in the early evolutionof streptophytes. Gene 401, 135–144.
Schmidt, E.D.L., Guzzo, F., Toonen, M.A.J., de Vries, S.C., 1997. A leucine rich repeatcontaining receptor-like kinase marks somatic plant cells competent to form em-bryos. Development 124, 2049–2062.
Shah, H., Gadella, T.W.J., van Erp, H., Hecht, V., de Vries, S.C., 2001a. Subcellular locali-zation and oligomerization of the Arabidopsis thaliana somatic embryogenesis re-ceptor kinase 1 protein. J. Mol. Biol. 309, 641–655.
Shah, K., Vervoort, J., de Vries, S.C., 2001b. Role of threonines in the Arabidopsis thalianasomatic embryogenesis receptor kinase 1 activation loop in phosphorylation.J. Biol. Chem. 276, 41263–41269.
Shah, K., Russinova, E., Gadella, T.W.J., Willemse, J., de Vries, S.C., 2002. The Arabidopsiskinase-associated protein phosphatase controls internalization of the somatic em-bryogenesis receptor kinase 1. Genes Dev. 16, 1707–1720.
Sharma, S.K., Millam, S., Hein, I., Bryan, G.J., 2008. Cloning and molecular characteriza-tion of a potato SERK gene transcriptionally induced during initiation of somaticembryogenesis. Planta 228, 319–330.
Shimada, T., Hirabayashi, T., Endo, T., Fujii, H., Kita, M., Omura, M., 2005. Isolation andcharacterization of the somatic embryogenesis receptor-like kinase gene homo-logue (CitSERK1) from Citrus unshui. Marc. Sci. Hortic. 103, 233–238.
Song, D.H., Li, G.J., Song, F.M., Zheng, Z., 2008. Molecular characterization andexpression analysis of OsBISERK1, a gene encoding a leucine-rich repeat receptor-like kinase, during disease resistance responses in rice. Mol. Biol. Rep. 35,275–283.
Steward, F.C., Mapes, M.O., Mears, K., 1958. Growth and organized development of cul-tured cells:II. Organization in cultures grown from freely suspended cells. Am. J.Bot. 45, 705–708.
Stone, S.L., et al., 2001. LEAFYCOTYLEDON2 encodes a B3 domain transcription factorthat induces embryo development. Proc. Natl. Acad. Sci. U. S. A. 98, 11806–11811.
Thomas, C., Meyer, D., Himber, C., Steinmetz, A., 2004. Spatial expression of a sunflowerSERK gene during induction of somatic embryogenesis and shoot organogenesis.Plant Physiol. Biochem. 42, 35–42.
Thomma, B.P., et al., 1998. Separate jasmonate-dependent and salicylate-dependentdefense-response pathways in Arabidopsis are essential for resistance to distinctmicrobial pathogens. Proc. Natl. Acad. Sci. U. S. A. 95, 15107–15111.
123J. Ma et al. / Gene 500 (2012) 115–123
Tosi, M., 1998. Molecular genetics of C1 inhibitor. Immunobiology 199, 358–365.Walker, J.C., 1994. Structure and function of the receptor-like protein kinases of higher
plants. Plant Mol. Biol. 26, 1599–1609.Wu, S.J., et al., 2008. Enhanced Agrobacterium-mediated Transformation of Embryogenic
Calli of Upland Cotton via Efficient Selection and Timely Subculture of Somatic Em-bryos. Plant Mol. Biol. Rep. 26, 174–185.
Yang, C., Zhao, T.J., Yu, D.Y., Gai, J.Y., 2011. Isolation and Functional Characterization of aSERK Gene from Soybean (Glycinemax (L.) Merr.). PlantMol. Biol. Rep. 29, 334–344.
Zdobnov, E.M., Apweiler, R., 2001. InterProScan-an integration platform for thesignature-recognition methods in InterPro. Bioinformatics 17, 847–848.
Zuo, J.R., Niu, Q.W., Frugis, G., Chua, N.H., 2002. The WUSCHEL gene promotesvegetative-to-embryonic transition in Arabidopsis. Plant J. 30, 349–359.









![Phenotypic diversity and relationships of fruit quality traits in peach and nectarine [Prunus persica (L.) Batsch] breeding progenies](https://static.fdokumen.com/doc/165x107/6324232e4d8439cb620d3dc3/phenotypic-diversity-and-relationships-of-fruit-quality-traits-in-peach-and-nectarine.jpg)










