
Suppression of a Vegetative MADS Box Gene of Potato Activates Axillary Meristem Development1
Upload
independentCategory
view
0download
0
A brassinolide-suppressed rice MADS-box transcriptionfactor, OsMDP1, has a negative regulatory role in BR signaling
Ke Duan1,2,3,†, Li Li2,3,†, Peng Hu2,3, Shu-Ping Xu2,3, Zhi-Hong Xu2 and Hong-Wei Xue2,3,*
1Key Laboratory of Photosynthesis and EnvironmentalMolecular Physiology, Institute of Botany, Chinese Academy of Sciences,
20 Nanxincun Road, 100093 Beijing, China,2National Key Laboratory of Plant Molecular Genetics, Institute of Plant Physiology and Ecology, Shanghai Institutes for
Biological Science (SiBS), Chinese Academy of Sciences, 300 Fenglin Road, 200032 Shanghai, China, and3Partner Group of Max-Planck-Institute of Molecular Plant Physiology (MPI-MP) on Plant Molecular Physiology and Signal
Transduction, 300 Fenglin Road, 200032 Shanghai, China
Received 8 November 2005; revised 22 March 2006; accepted 7 April 2006.*For correspondence (fax þ86 21 5492 4060; e-mail [email protected]).†These authors contributed equally to this study.
Summary
The MADS-box transcription factor-encoding genes are expressed mainly during plant reproductive develop-
ment, where they play important roles in controlling floral organ initiation and identity. Few previous reports
have investigated the functions of MADS-box transcription factors expressed in vegetative tissues. Here we
describe the functional characterization of a rice AG-like MADS-box protein, OsMDP1 (Oryza sativa MADS-
domain-containing protein 1). A partial cDNA encoding a MADS-box domain was identified via high-
throughput screening of rice brassinolide-regulated genes, and the full-length cDNA was subsequently
isolated via screening of a cDNA library constructed from rice materials at tillering stage. Expression pattern
analyses indicated that OsMDP1 is transcribed mainly in vegetative tissues, including the mature leaf,
coleoptile, root-elongation zone, culm internode, and especially the joint region between the leaf blade and
sheath. Further studies revealed that transcription of OsMDP1 is stimulated by darkness and suppressed by
brassinolide treatment. OsMDP1 deficiency resulted in shortened primary roots, elongated coleoptiles and
enhanced lamina joint inclinations. Moreover, transgenic plants showed hypersensitivities to exogenous
brassinolide in terms of lamina joint inclination and coleoptile elongation. OsMDP1 deficiency resulted in
enhanced expression of OsXTR1, which encodes xyloglucan endotransglycosylase, the cell-wall loosening
enzyme necessary for cell elongation, and modulated expressions of multiple genes involved in cell signalling
and gene transcription, indicating the key negative regulatory role of OsMDP1 in BR signalling.
Keywords: OsMDP1, MADS-box protein, BR signalling.
Introduction
Proper plant growth and development is the result of co-
ordinated gene expression regulated by integrated internal
and external signal transduction networks that are centrally
controlled by transcription factors. The steroid phytohor-
mone brassinosteroid (BR) is an essential regulator of plant
growth, organogenesis, cell expansion, vascular differenti-
ation, etiolation, reproductive development, embryogenic
tissue initiation, and stress responses (Bishop and Koncz,
2002; Bishop and Yokota, 2001; Nakashita et al., 2003; Pull-
man et al., 2003). Although fewer studies have investigated
BR responses in monocots, some have examined the BR
effects in rice growth and development. One well-known
response is the BR-mediated stimulation of lamina inclina-
tion (Maeda, 1965), which is generally accepted as a sensi-
tive bioassay for BR (Wada et al., 1981). In addition, higher
concentrations (ranging from 0.001 to 1 lM) of 24-epibras-
sinolide (24-eBL), one of the most active BRs, can promin-
ently increase coleoptile length and decrease root length in
rice (Yamamuro et al., 2000). Study of a rice dwarf mutant
that is defective in the BR biosynthesis enzyme, BR-6-oxid-
ase, indicates that BR is involved in many rice morphological
and physiological processes, including the elongation and
ª 2006 The Authors 519Journal compilation ª 2006 Blackwell Publishing Ltd
The Plant Journal (2006) 47, 519–531 doi: 10.1111/j.1365-313X.2006.02804.x

unrolling of leaves, development of tillers, photomorpho-
genesis, root differentiation and reproductive growth (Hong
et al., 2002; Mori et al., 2002). Recent studies on BR-defici-
ency dwarf mutants further reveal the roles of the BR-bio-
synthesis network in regulating rice growth and bending
(Hong et al., 2005; Tanabe et al., 2005). In brd2, the single
base deletion of the coding region of Dim/dwf1 resulted in
an activated biosynthesis pathway of the 24-methylene BR
and an uncommon BR, dolichosterone (DS), which could
enhance the bending angle of the rice lamina joint, rescue
the brd1 dwarf phenotype, and inhibit root elongation (Hong
et al., 2005).
BR signalling and the related functional mechanisms are
emerging as a hot topic in the field of plant hormone
research. Previous studies have shown that multiple genes
are regulated (induced or repressed) by BR and play critical
roles in BR effects. For example, multiple genes encoding
xyloglucan endotransglycosylases (XTHs), the cell wall-
loosening enzymes necessary for cell elongation, have been
shown to be regulated by BR and to function in BR-promoted
growth responses (Kauschmann et al., 1996; Koka et al.,
2000; Muessig and Altmann, 2001). In rice, researchers
found that the XTH-encoding genes OsXTR1 and OsXTR3,
which were preferentially transcribed in the elongating
zones of internodes, could be upregulated by BR and
gibberellic acid (GA), and were critical for internodal elon-
gation (Uozu et al., 2000). Other studies revealed that
OsBLE2 (a BR-enhanced gene), identified by cDNA microar-
ray expression profiling analysis of 1265 rice genes in BR-
treated rice lamina joints, might be involved in BR-induced
cell expansion of lamina joints and leaf sheaths (Yang et al.,
2003). Furthermore, recent proteomics analyses identified
more than 500 proteins having physiological functions
during 24-eBL-induced rice lamina inclination and root
elongation (Konishi and Komatsu, 2003). BR biosynthesis
in rice involves BR-6-oxidase, the coding gene of which is
preferentially transcribed in the leaf sheath and negatively
regulated by 24-eBL treatment (Hong et al., 2002), and also a
novel cytochrome P450, CYP90D2, which was recently isola-
ted and found to be required for catalyzing the late steps in
BR biosynthesis (Hong et al., 2003).
BR signalling in Arabidopsis thaliana and rice – dicoty-
ledonous and monocotyledonous models, respectively – is
mediated by the receptor-like kinases BRI1 and OsBRI1. BRI1
transduces the BR signal into the nucleus through a phos-
phorylation cascade that induces differential expressions of
target genes, such as those controlling cell expansions
(Friedrichsen et al., 2000; reviewed by Muessig and Alt-
mann, 2001). BR signalling through cell-surface receptor
kinases and intracellular components leads to dephospho-
rylation and accumulation of the nuclear protein BZR1, a
transcriptional repressor that has a previously unknown
DNA-binding domain, and binds directly to the promoters of
feedback-regulated BR biosynthetic genes (He et al., 2005;
Wang et al., 2002). Another positive regulator, BES1, defines
a new class of plant-specific transcription factors that co-
operate with transcription factors such as BIM1 to regulate
BR-induced genes (Yin et al., 2005). Additionally, three
closely related bHLH transcription factors, BEE1–3, are
functionally redundant positive regulators of BR signalling
(Friedrichsen et al., 2002). Although it remains unknown
how BR signalling regulates gene expression during plant
growth, all these studies provide cues that transcriptional
regulators or transcription factors co-ordinate BR home-
ostasis and signalling through regulating BR biosynthesis
and further downstream responses.
In rice, OsBRI1 functions in various growth and develop-
mental processes, including internode elongation, lamina
joint bending and skotomorphogenesis (Yamamuro et al.,
2000). Using excised rice lamina, studies further suggest that
Ca2þ -dependent protein kinase (CDPK) and Ca2þ -dependent
protein phosphorylation are involved in BR-induced rice
lamina inclination (Yang and Komatsu, 2000). BR was shown
to activate the membrane-bound mitogen-activated protein
kinase (MAPK) and cytosolic CDPK. However, the BR-regu-
lated tendencies of these two kinase genes were reversed in
OsBRI1-deficient plants, suggesting that the signalling
pathways downstream of BR-mediated activation of MAPK
and CDPK are discrete but parallel, and probably involve
other receptors in addition to BRI1 for BR response (Sharma
et al., 2001). Although amounts of BR-associated genes have
been characterized, the detailed functional mechanisms,
especially regulatory mechanisms, are still unclear.
MADS-box transcription factors play diverse, important
roles in floral organ development (Jack, 2001; Ng and
Yanofsky, 2001) in reproductive and vegetative develop-
ment, and in signal transduction (Becker and Theissen,
2003; De Bodt et al., 2003; Messenguy and Dubois, 2003).
Analysis of the complete Arabidopsis genome reveals the
presence of 107 genes encoding MADS-box proteins, 84%
of which have unknown functions (Parenicova et al., 2003).
To date, a total of 89 rice MADS-box proteins coding
cDNAs have been reported (National Center for Biotech-
nology Information, NCBI database, 16 May 2006), 24 of
which were isolated from floral tissues at different devel-
opmental stages, and only eight of which were isolated
from vegetative tissues, including callus (BP185002), GA-
treated callus (AB003322), NaCl-treated seedlings
(CA758412) and roots (CA755986), untreated roots
(AB003326), green shoots (AB003327 and AB003328) and
leaves (AJ293816). There is no previous report on the
detailed functional characterization of any vegetative tis-
sue-expressed rice MADS-box gene.
Here we report the isolation and functional characteriza-
tion of a vegetative tissue-expressed rice MADS-box gene,
OsMDP1. A comprehensive analysis of the expression
pattern and physiological roles of this gene revealed that
OsMDP1 negatively regulates BR-mediated rice lamina joint
520 Ke Duan et al.
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 519–531

inclination and is also involved in BR-mediated coleoptile
elongation.
Results
Isolation and structural analysis of OsMDP1
Our previous studies of rice BR-responsive genes (Z-Q. Chu
et al., unpublished data) identified an approximately 500-bp
BR-suppressed cDNA fragment sharing homology with the
plant MADS-box genes. Specific primers were designed
from this fragment and used to screen a cDNA library con-
structed from rice materials at tillering stage. A full-length
rice cDNA clone was isolated and designated as OsMDP1
(Oryza sativa MADS domain protein 1), which has a length of
1045 bp and encodes a 248-aa peptide. OsMDP1 is located
on rice chromosome 3 (clone AC125471). Comparison of the
cDNA and genomic sequences reveals the presence of eight
exons ranging in size from 42 to 312 bp, and seven introns
ranging in size from 85 to 3804 bp (Figure 1a). Comparative
analysis on the corresponding genomic region indicates that
the predicted presence of two spliced forms is incorrect, but
that there is a single gene at this locus. In addition, the actual
cDNA does not contain the10-bp exon that was theoretically
predicted; the final exon is 53 bp longer, consistent with the
previous analysis in which the OsMDP1 was identified as
OsMADS47 through a systematic reverse genetic screening
of T-DNA target gene in rice (accession number AY345221;
Lee et al., 2003).
OsMDP1 shares high homology with the other plant
MADS-box proteins, having 80% identity with HvMADS1–2
(barley, AJ249142) and HvMADS1 (barley, AJ249141); 38.7%
with OsMADS7 (rice, U78891); and 34% with AtAG (Arabid-
opsis, X53579). In addition, OsMDP1 possesses a modular
structure shared by all plant AG-like MADS proteins. This
structure (N-M-I-K-C) sequentially consists of the N-termi-
nus; the MADS domain that is essential for DNA-binding and
dimerization; the I region between the MADS and K
domains; the K-domain that is postulated to be important
for protein–protein interactions (Jack, 2001); and the C-
terminus (Figure 1b). The N, I and C regions are less well
conserved in OsMDP1, whereas the MADS and K domains
are highly conserved; the 60-aa MADS domain shares 81.7%
identity with the Arabidopsis AG protein, and only 1 aa out
of 60 differs from the HvMADS1 and HvMADS1–2 MADS
domain sequences (Figure 1c). Only AG-like MADS proteins
vary in the length of their N-termini prior to the MADS
domain (Johansen et al., 2002), providing further evidence
that OsMDP1, which has a 25-aa N-terminus prior to the
MADS-domain, is likely to be an AG-like MADS domain
transcription factor.
OsMDP1 is expressed during various rice developmental
stages and is regulated by plant hormones and light
conditions
Northern blotting was originally used to study the expres-
sion patterns of OsMDP1, but the signals were nearly un-
detectable in RNA samples from roots, seedlings, culms,
flowers and seeds, and very weak in leaf samples, suggest-
ing that OsMDP1 is expressed at very low level and/or in very
specific tissues (data not shown). To examine these possi-
bilities more fully, reverse-transcription PCR (RT–PCR),
quantitative real-time RT–PCR, and promoter-b-glucuroni-
Figure 1. Gene structure and domain organiza-
tion of the OsMDP1 gene.
(a) Schematic diagram of the exons (boxes) and
introns (lines) of OsMDP1. Numbers indicate the
size of exons/introns.
(b) Structural organization of OsMDP1. Con-
served domains including amino-terminal (N),
MADS domain (M), interval region between
MADS and K domain (I), K domain (K) and
carbon terminal (C) are marked, also for other
MADS proteins. Accession numbers: OsMDP1
(Oryza sativa, AJ536158), HvMADS1 (Hordeum
vulgare, AJ249141), HvMADS1–2 (H. vulgare,
AJ249142), OsMADS7 (O. sativa, U78891), AtAG
(Arabidopsis thaliana, X53579). Numbers indi-
cate the positions of the various domains.
(c) Alignment of the predicted MADS-domain of
OsMDP1 to other MADS proteins. Identical
amino acids are marked.
OsMDP1 in brassinosteroid signalling 521
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 519–531

dase (GUS) reporter gene-fusion studies were performed.
RT–PCR analysis revealed the expression of OsMDP1 in the
roots and shoots of 2-week-old seedlings, mature stems and
leaves of 45-day-old rice, flowering panicles, developing
seeds at 4, 8, 12, 16, 20, 24 days after anthesis (DAA), and
mature seeds (Figure 2a). Consistent with previous reports
regarding other AG-type MADS-box genes, OsMDP1 was
found to be expressed in a wide range of organs, but the
expression varied markedly among the different organs.
Relatively high expression of OsMDP1 was detected in
young seedlings and mature leaves; moderate expression
was observed in culms and immature seeds (4–16 DAA), and
negligible expression was detected in flowers and mature
seeds.
Promoter–reporter gene fusion studies were used for a
more detailed investigation of the expression of OsMDP1 in
various organs. The 1.5-kb promoter region of OsMDP1 was
PCR amplified and fused to the GUS reporter gene. Histo-
chemical analysis of rice transformants showed strong GUS
activities in rice leaves, preferentially in the lamina joints of
both etiolated seedlings grown in the dark (Figure 2b,1) and
normal green seedlings (Figure 2b,2), as well as in the joint
region between the mature leaf blade and the sheath
(Figure 2b,3), and the mature blade (Figure 2b,4) and sheath
(Figure 2b,5). GUS activities were also detected in the leaf
joint regions of a yellow, senescent leaf (data not shown).
However, detailed microscope studies of OsMDP1 promoter
activity in the leaf joint region did not reveal polarity-specific
activities – GUS activities were detected similarly in both
adaxial and abaxial cells.
During seed germination, GUS activities were found in the
coleoptile (Figure 2b,6,7) and radicle (Figure 2b,8), while in
young seedlings expression was detected in the lamina joint
as well as in primary and lateral roots (Figure 2b,9–12).
Interestingly, in roots the OsMDP1 promoter was highly
active in the elongation zone, but only weakly active in the
division and mature zones (Figure 2b,9,11), suggesting it
may play a role in cell elongation during root growth.
In culm tissue, GUS activity was detected in the upper-
most two internodes directly prior to the rapid elongation of
the culm (Figure 2b,13). During the booting stage, GUS
activity in the elongating culm was confined to the lower
part of the elongated third internode, presumably corres-
ponding to the division and elongating zones (Figure 2b,15–
17), but was no longer detected in the uppermost two
internodes. GUS activity later disappeared from the culm,
and was observed only in the peduncle of immature panicle
(Figure 2b,14).
We further tested the effects of exogenously applied plant
hormones on OsMDP1 transcription through quantitative
real-time RT–PCR. As shown in Figure 2(c), expression of
OsMDP1 was significantly suppressed following treatments
with IAA for 4 h or 24-eBL for 2 h under light-grown
conditions (Figure 2c, upper panel). In contrast, application
of GA and abscisic acid (ABA) showed no obvious effects.
Interestingly, OsMDP1 was transcribed at a relatively higher
level in dark-grown etiolated seedlings than in those grown
in the light (Figure 2c, lower panel).
Pleiotropic phenotypic changes in OsMDP1-deficient plants:
abnormal elongation of primary roots and coleoptiles, and
enhanced lamina joint inclination
The pA-OsMDP1 binary vector, which harboured the Os-
MDP1 coding region in an antisense orientation, was trans-
formed into rice. More than 20 independent resistant T0 lines
were obtained, and transformants confirmed as OsMDP1-
deficient (Figure 3a) were further cultivated for an additional
two generations to obtain homozygous lines for analysis.
As expression of OsMDP1 is dark-inducible, we first
focused the observations on dark-grown seedlings. As
shown in Figure 3(b), coleoptile and primary root develop-
ments were inversely modified in dark-grown transgenic
seedlings compared with control seedlings: transgenic
seedlings developed longer coleoptiles, but shorter primary
roots. Three lines, A18, A19 and A1, showed weak, moderate
and strong phenotypes, respectively, consistent with the
deficiency degree of OsMDP1 in each transgenic line, and
were used for detailed statistical analyses. The coleoptiles of
lines A1, A19 and A18 were 53%, 38% and 29% longer than
those of control plants, while the primary roots were 30%,
17% and 16% shorter, respectively (Figure 3b–d, left panel).
Concretely, the average length of control coleoptiles was
1.29 � 0.17 cm (1 week after germination in darkness), while
that in line A1 was 1.98 � 0.50 cm (Figure 3c, left panel); the
average primary root length in control plants was
15.4 � 1.4 cm, while that in line A1 was 10.8 � 1.0 cm
(Figure 3d, left panel), and analyses of these differences
were statistically significant. Although no perceptible differ-
ence in coleoptile length was observed in light-grown
seedlings, the light-grown transgenic plants did show some
inhibition of primary root elongation as well (data not
shown).
A previous study showed that relatively high concentra-
tions (e.g. 0.1 lM) of 24-epibrassinolide (24-eBL) could
obviously increase coleoptile length and decrease root
length in rice seedlings (Yamamuro et al., 2000). Based on
the observation that exogenous brassinolide suppressed the
expression of OsMDP1, we next investigated whether the
effects of exogenous 24-eBL on rice growth were altered.
Under treatment of 0.1 lM 24-eBL, BR-induced coleoptile
elongation in darkness was greatly magnified in OsMDP1-
deficiency plants (Figure 3b). The average length of coleop-
tiles in A1 plants is 4.12 � 0.93 cm, while that in control
plants is 1.9 � 0.32 cm. Statistical analysis showed that the
relative coleoptile length increase in A1 plants under 24-BL
treatment (91.6–108%) was substantially higher than that in
control plants (58.3%; Figure 3c, right panel). However,
522 Ke Duan et al.
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 519–531
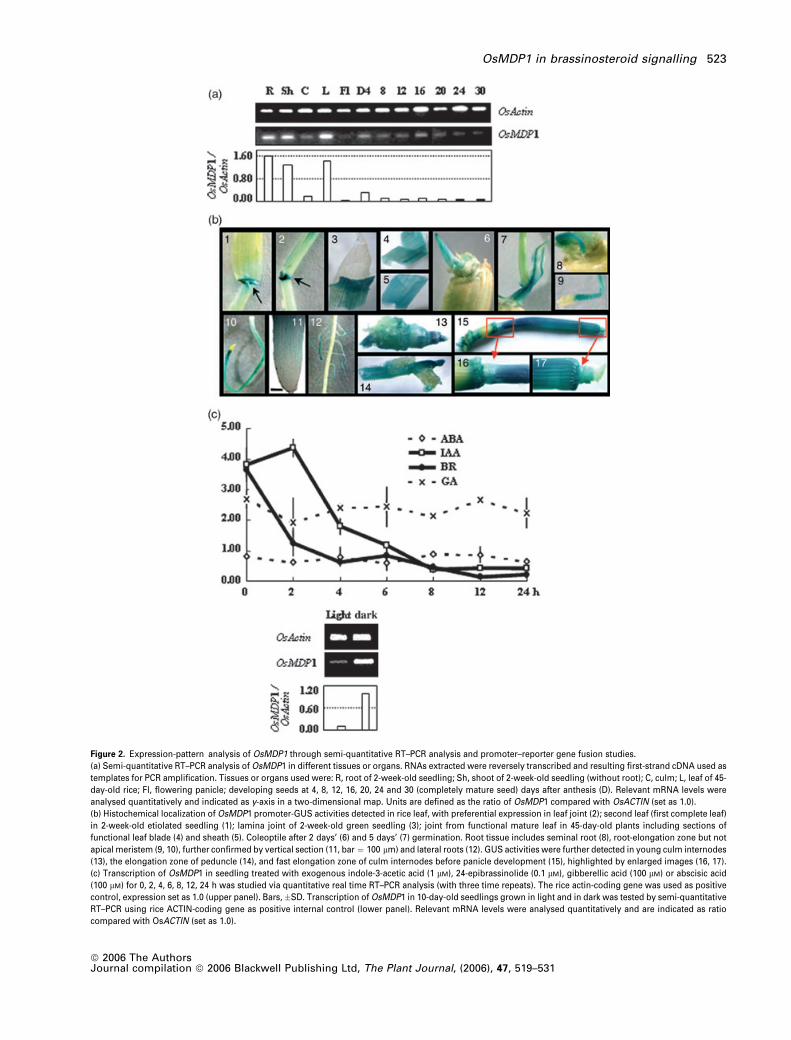
Figure 2. Expression-pattern analysis of OsMDP1 through semi-quantitative RT–PCR analysis and promoter–reporter gene fusion studies.
(a) Semi-quantitative RT–PCR analysis of OsMDP1 in different tissues or organs. RNAs extracted were reversely transcribed and resulting first-strand cDNA used as
templates for PCR amplification. Tissues or organs used were: R, root of 2-week-old seedling; Sh, shoot of 2-week-old seedling (without root); C, culm; L, leaf of 45-
day-old rice; Fl, flowering panicle; developing seeds at 4, 8, 12, 16, 20, 24 and 30 (completely mature seed) days after anthesis (D). Relevant mRNA levels were
analysed quantitatively and indicated as y-axis in a two-dimensional map. Units are defined as the ratio of OsMDP1 compared with OsACTIN (set as 1.0).
(b) Histochemical localization of OsMDP1 promoter-GUS activities detected in rice leaf, with preferential expression in leaf joint (2); second leaf (first complete leaf)
in 2-week-old etiolated seedling (1); lamina joint of 2-week-old green seedling (3); joint from functional mature leaf in 45-day-old plants including sections of
functional leaf blade (4) and sheath (5). Coleoptile after 2 days’ (6) and 5 days’ (7) germination. Root tissue includes seminal root (8), root-elongation zone but not
apical meristem (9, 10), further confirmed by vertical section (11, bar ¼ 100 lm) and lateral roots (12). GUS activities were further detected in young culm internodes
(13), the elongation zone of peduncle (14), and fast elongation zone of culm internodes before panicle development (15), highlighted by enlarged images (16, 17).
(c) Transcription of OsMDP1 in seedling treated with exogenous indole-3-acetic acid (1 lM), 24-epibrassinolide (0.1 lM), gibberellic acid (100 lM) or abscisic acid
(100 lM) for 0, 2, 4, 6, 8, 12, 24 h was studied via quantitative real time RT–PCR analysis (with three time repeats). The rice actin-coding gene was used as positive
control, expression set as 1.0 (upper panel). Bars, �SD. Transcription of OsMDP1 in 10-day-old seedlings grown in light and in dark was tested by semi-quantitative
RT–PCR using rice ACTIN-coding gene as positive internal control (lower panel). Relevant mRNA levels were analysed quantitatively and are indicated as ratio
compared with OsACTIN (set as 1.0).
OsMDP1 in brassinosteroid signalling 523
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 519–531
although 24-eBL treatment results in the decrease of the root
length in A1 plants by 48.5–53%, similar effects (49%) were
detected in control plants (Figure 3d, right panel), indicating
the elevated sensitivity of OsMDP1-deficiency seedling to
exogenous 24-eBL in terms of coleoptile elongation, but
normal response sensitivity in term of primary root growth.
The bending degree between the leaf blade and sheath is
known to be sensitive to exogenous 24-eBL and IAA.
However, only 24-eBL can promote leaf bending at a
concentration as low as 0.1 ng ml)1 (approximately
0.2 nM). This phenomenon, which has been used as a
bioassay for BR activity (Wada et al., 1984), is due to BR-
induced stimulation of the rapid division and elongation of
adaxial cells at the joint region (Cao and Chen, 1995). As
expression-pattern analysis indicated that OsMDP1 was
suppressed by 24-eBL, and promoter analysis showed that
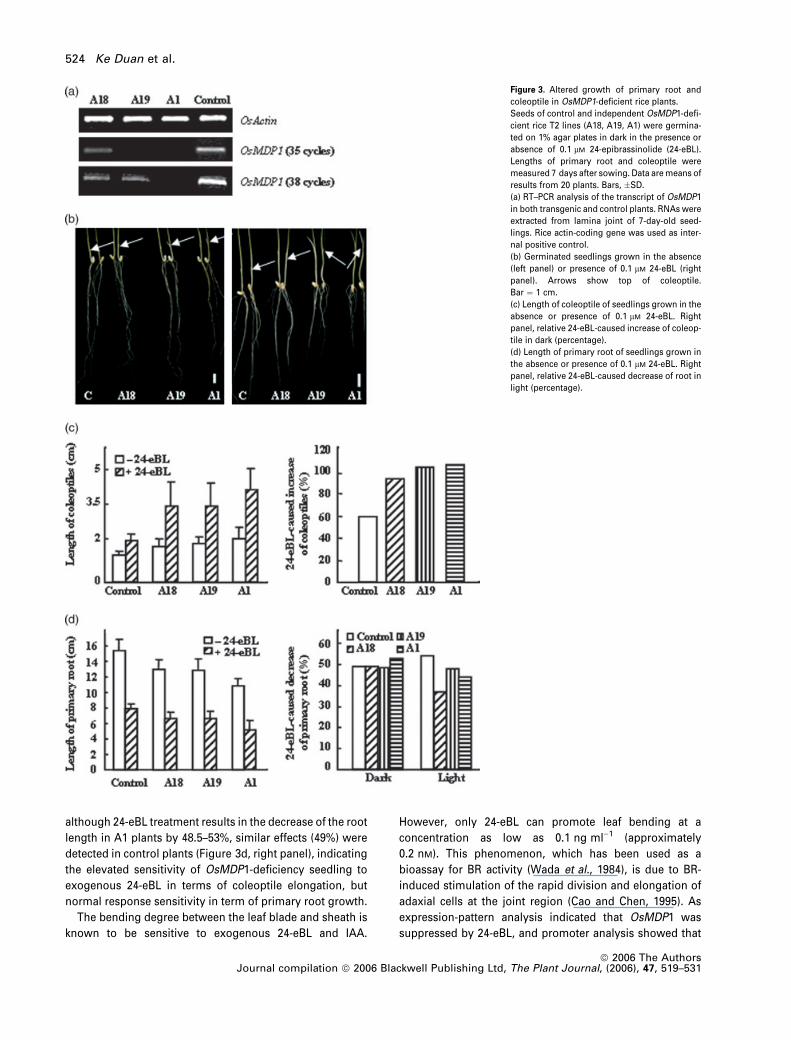
Figure 3. Altered growth of primary root and
coleoptile in OsMDP1-deficient rice plants.
Seeds of control and independent OsMDP1-defi-
cient rice T2 lines (A18, A19, A1) were germina-
ted on 1% agar plates in dark in the presence or
absence of 0.1 lM 24-epibrassinolide (24-eBL).
Lengths of primary root and coleoptile were
measured 7 days after sowing. Data are means of
results from 20 plants. Bars, �SD.
(a) RT–PCR analysis of the transcript of OsMDP1
in both transgenic and control plants. RNAs were
extracted from lamina joint of 7-day-old seed-
lings. Rice actin-coding gene was used as inter-
nal positive control.
(b) Germinated seedlings grown in the absence
(left panel) or presence of 0.1 lM 24-eBL (right
panel). Arrows show top of coleoptile.
Bar ¼ 1 cm.
(c) Length of coleoptile of seedlings grown in the
absence or presence of 0.1 lM 24-eBL. Right
panel, relative 24-eBL-caused increase of coleop-
tile in dark (percentage).
(d) Length of primary root of seedlings grown in
the absence or presence of 0.1 lM 24-eBL. Right
panel, relative 24-eBL-caused decrease of root in
light (percentage).
524 Ke Duan et al.
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 519–531

OsMDP1 was specifically expressed at the joint region
between the leaf blade and sheath, we hypothesized that
OsMDP1 may modulate lamina joint inclination via tran-
scriptional regulation of relevant genes. To test this possi-
bility, lamina joint assays were performed on both control
and transgenic plants, and the results showed that the leaf
blades bent more sharply in the presence of 24-eBL, in a
dose-dependent manner (Figure 4a,b). This effect was
observed in both control and transgenic plants, although
the bending of transgenic rice laminas was more severe
under the same conditions. Indeed, the joints of some A1
plants even bent beyond 180� in the presence of 5.0 ng ml)1
(approximately 10 nM) 24-eBL (Figure 4b). However, the
differences in lamina bending between the control and
transgenic plants were more obvious at the lower concen-
trations of 24-eBL (Figure 4b,c). At a concentration of
0.2 ng ml)1 (approximately 0.4 nM) 24-eBL, no prominent
promotion of inclination was observed in control plants,
whereas enhanced effects were detected in all transgenic
plants (Figure 4a,c); the bending angle in line A1 was
137 � 3�, compared with 113 � 1� in control plants.
Lamina bending was also tested in light-grown seedlings
in vivo following the method introduced by Fujioka et al.
(1998), in which the lamina joint was not excised from the
seedling. As above, we observed larger exterior angles
between the lamina and sheaths in OsMDP1-deficient plants
as compared with controls in the absence of 24-eBL, and line
A1 showed more severe inclinations than the other lines
under the same conditions. In experiments examining the
effects of 24-eBL, similar results as in dark-grown seedlings
were obtained, although a relatively higher concentration of
24-eBL was required for obvious promotion of lamina
bending.
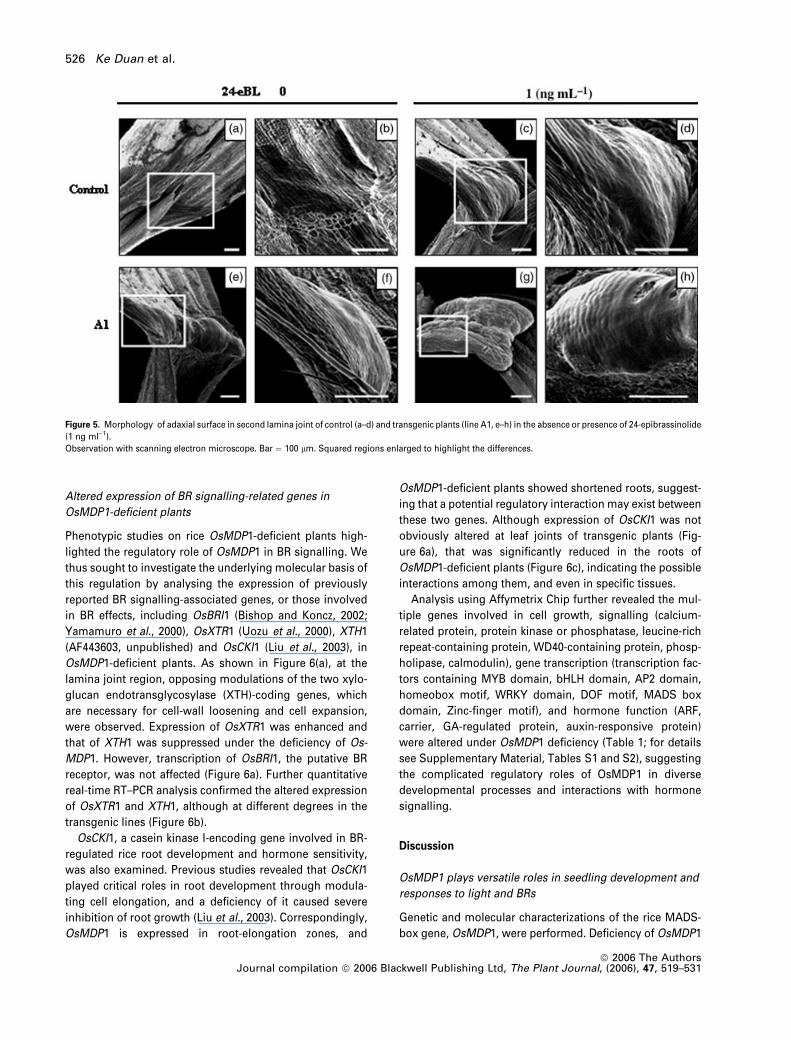
OsMDP1-deficient plants showed altered expansion of
adaxial cells at the leaf joint
In previous work, the cause of lamina joint bending was
shown to be the BR-stimulated rapid division and elongation
of adaxial cells at the joint (Cao and Chen, 1995). Thus we
questioned whether abnormal adaxial cell growth occurs in
transgenic plants, or whether such abnormal growth re-
quires stimulation by exogenously applied 24-eBL. SEM
analysis was performed on the lamina joints of dark-grown
seedlings, and the adaxial surface of the lower part of the
leaf blade opposite the ligule was observed. The results re-
vealed that, in the absence of 24-eBL, the adaxial surface of
the second lamina joint of control plants was smooth and
lacked obvious cell bumps (Figure 5a,b), whereas in the
presence of 24-eBL, the adaxial cells formed bumps that
gradually enlarged and shifted downwards over time (Fig-
ure 5c,d). As the adaxial cell bumps neared the bottom of the
lamina, the joint bend became more severe. In plants of
transgenic line A1, even in the absence of 24-eBL, the cell
bump was present on the adaxial lamina surface, on the
upper side of the joint region between the blade and sheath
(Figure 5e,f). In the presence of 24-eBL, the adaxial cells at
the bottom of the lamina were enlarged and bumped,
causing the lamina to be bent at an even sharper angle
(Figure 5g,h), indicating that abnormal adaxial cell growth
occurred in OsMDP1-deficient plants, and the downward
shift of bumped adaxial cells may be the direct cause of the
observed leaf-blade drooping.
Figure 4. OsMDP1 deficiency results in the enhanced inclination of etiolated
leaf lamina joint following 24-epibrassinolide (24-eBL) treatments.
After germinating for 8 days on 1% agar in darkness, second leaf joints
(1 þ 1 cm long) were floated in distilled water for 1 day, then treated with 24-
eBL (ng ml)1) for 2.5 days. The angle was measured by protractor.
Bar ¼ 1 cm.
(a) Response of second lamina joints of control plants, transgenic plants (lines
A18, A19 and A1) to 24-eBL (0.2 ng ml)1).
(b) Dose response to 24-eBL (ng ml)1) of second leaf lamina joint of control
and transgenic plants (line A1).
(c) Dose response to 24-eBL of the bending angles of control plants and
transgenic plants (lines A18, A19 and A1); y-axis, angle; x-axis, 24-eBL
concentration (ng ml)1). Data presented are means of results from 10 plants.
Bars, �SD.
OsMDP1 in brassinosteroid signalling 525
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 519–531
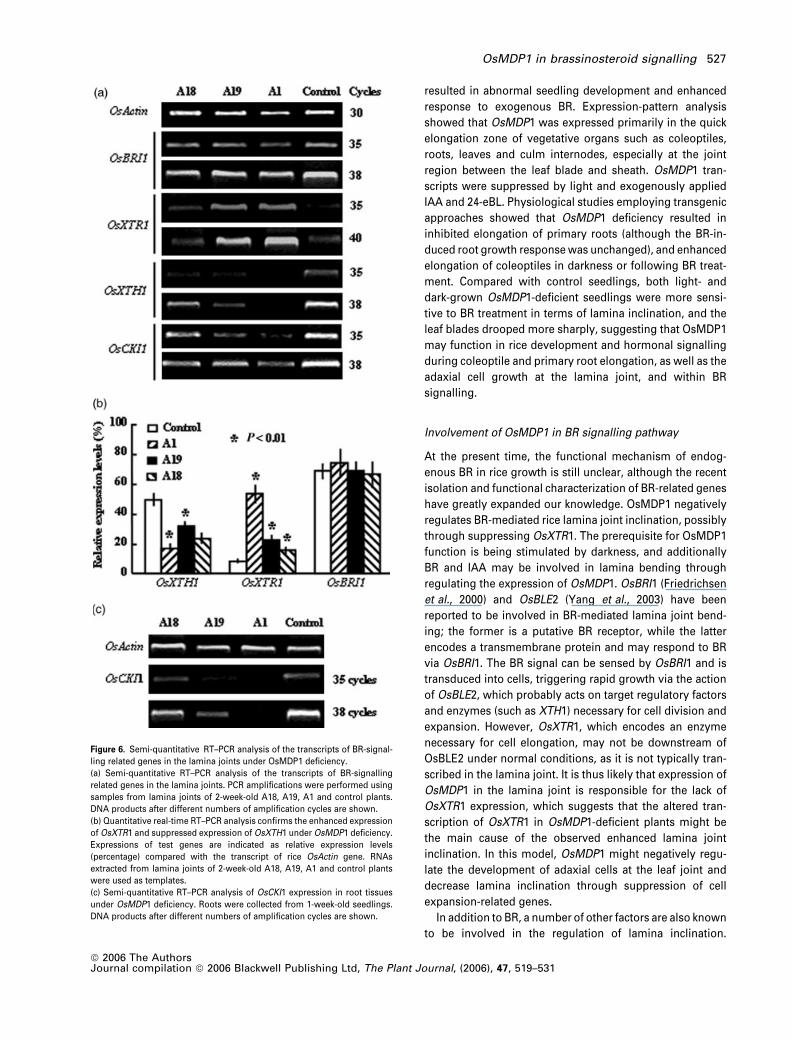
Altered expression of BR signalling-related genes in
OsMDP1-deficient plants
Phenotypic studies on rice OsMDP1-deficient plants high-
lighted the regulatory role of OsMDP1 in BR signalling. We
thus sought to investigate the underlying molecular basis of
this regulation by analysing the expression of previously
reported BR signalling-associated genes, or those involved
in BR effects, including OsBRI1 (Bishop and Koncz, 2002;
Yamamuro et al., 2000), OsXTR1 (Uozu et al., 2000), XTH1
(AF443603, unpublished) and OsCKI1 (Liu et al., 2003), in
OsMDP1-deficient plants. As shown in Figure 6(a), at the
lamina joint region, opposing modulations of the two xylo-
glucan endotransglycosylase (XTH)-coding genes, which
are necessary for cell-wall loosening and cell expansion,
were observed. Expression of OsXTR1 was enhanced and
that of XTH1 was suppressed under the deficiency of Os-
MDP1. However, transcription of OsBRI1, the putative BR
receptor, was not affected (Figure 6a). Further quantitative
real-time RT–PCR analysis confirmed the altered expression
of OsXTR1 and XTH1, although at different degrees in the
transgenic lines (Figure 6b).
OsCKI1, a casein kinase I-encoding gene involved in BR-
regulated rice root development and hormone sensitivity,
was also examined. Previous studies revealed that OsCKI1
played critical roles in root development through modula-
ting cell elongation, and a deficiency of it caused severe
inhibition of root growth (Liu et al., 2003). Correspondingly,
OsMDP1 is expressed in root-elongation zones, and
OsMDP1-deficient plants showed shortened roots, suggest-
ing that a potential regulatory interaction may exist between
these two genes. Although expression of OsCKI1 was not
obviously altered at leaf joints of transgenic plants (Fig-
ure 6a), that was significantly reduced in the roots of
OsMDP1-deficient plants (Figure 6c), indicating the possible
interactions among them, and even in specific tissues.
Analysis using Affymetrix Chip further revealed the mul-
tiple genes involved in cell growth, signalling (calcium-
related protein, protein kinase or phosphatase, leucine-rich
repeat-containing protein, WD40-containing protein, phosp-
holipase, calmodulin), gene transcription (transcription fac-
tors containing MYB domain, bHLH domain, AP2 domain,
homeobox motif, WRKY domain, DOF motif, MADS box
domain, Zinc-finger motif), and hormone function (ARF,
carrier, GA-regulated protein, auxin-responsive protein)
were altered under OsMDP1 deficiency (Table 1; for details
see Supplementary Material, Tables S1 and S2), suggesting
the complicated regulatory roles of OsMDP1 in diverse
developmental processes and interactions with hormone
signalling.
Discussion
OsMDP1 plays versatile roles in seedling development and
responses to light and BRs
Genetic and molecular characterizations of the rice MADS-
box gene, OsMDP1, were performed. Deficiency of OsMDP1
Figure 5. Morphology of adaxial surface in second lamina joint of control (a–d) and transgenic plants (line A1, e–h) in the absence or presence of 24-epibrassinolide
(1 ng ml)1).
Observation with scanning electron microscope. Bar ¼ 100 lm. Squared regions enlarged to highlight the differences.
526 Ke Duan et al.
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 519–531
resulted in abnormal seedling development and enhanced
response to exogenous BR. Expression-pattern analysis
showed that OsMDP1 was expressed primarily in the quick
elongation zone of vegetative organs such as coleoptiles,
roots, leaves and culm internodes, especially at the joint
region between the leaf blade and sheath. OsMDP1 tran-
scripts were suppressed by light and exogenously applied
IAA and 24-eBL. Physiological studies employing transgenic
approaches showed that OsMDP1 deficiency resulted in
inhibited elongation of primary roots (although the BR-in-
duced root growth response was unchanged), and enhanced
elongation of coleoptiles in darkness or following BR treat-
ment. Compared with control seedlings, both light- and
dark-grown OsMDP1-deficient seedlings were more sensi-
tive to BR treatment in terms of lamina inclination, and the
leaf blades drooped more sharply, suggesting that OsMDP1
may function in rice development and hormonal signalling
during coleoptile and primary root elongation, as well as the
adaxial cell growth at the lamina joint, and within BR
signalling.
Involvement of OsMDP1 in BR signalling pathway
At the present time, the functional mechanism of endog-
enous BR in rice growth is still unclear, although the recent
isolation and functional characterization of BR-related genes
have greatly expanded our knowledge. OsMDP1 negatively
regulates BR-mediated rice lamina joint inclination, possibly
through suppressing OsXTR1. The prerequisite for OsMDP1
function is being stimulated by darkness, and additionally
BR and IAA may be involved in lamina bending through
regulating the expression of OsMDP1. OsBRI1 (Friedrichsen
et al., 2000) and OsBLE2 (Yang et al., 2003) have been
reported to be involved in BR-mediated lamina joint bend-
ing; the former is a putative BR receptor, while the latter
encodes a transmembrane protein and may respond to BR
via OsBRI1. The BR signal can be sensed by OsBRI1 and is
transduced into cells, triggering rapid growth via the action
of OsBLE2, which probably acts on target regulatory factors
and enzymes (such as XTH1) necessary for cell division and
expansion. However, OsXTR1, which encodes an enzyme
necessary for cell elongation, may not be downstream of
OsBLE2 under normal conditions, as it is not typically tran-
scribed in the lamina joint. It is thus likely that expression of
OsMDP1 in the lamina joint is responsible for the lack of
OsXTR1 expression, which suggests that the altered tran-
scription of OsXTR1 in OsMDP1-deficient plants might be
the main cause of the observed enhanced lamina joint
inclination. In this model, OsMDP1 might negatively regu-
late the development of adaxial cells at the leaf joint and
decrease lamina inclination through suppression of cell
expansion-related genes.
In addition to BR, a number of other factors are also known
to be involved in the regulation of lamina inclination.
Figure 6. Semi-quantitative RT–PCR analysis of the transcripts of BR-signal-
ling related genes in the lamina joints under OsMDP1 deficiency.
(a) Semi-quantitative RT–PCR analysis of the transcripts of BR-signalling
related genes in the lamina joints. PCR amplifications were performed using
samples from lamina joints of 2-week-old A18, A19, A1 and control plants.
DNA products after different numbers of amplification cycles are shown.
(b) Quantitative real-time RT–PCR analysis confirms the enhanced expression
of OsXTR1 and suppressed expression of OsXTH1 under OsMDP1 deficiency.
Expressions of test genes are indicated as relative expression levels
(percentage) compared with the transcript of rice OsActin gene. RNAs
extracted from lamina joints of 2-week-old A18, A19, A1 and control plants
were used as templates.
(c) Semi-quantitative RT–PCR analysis of OsCKI1 expression in root tissues
under OsMDP1 deficiency. Roots were collected from 1-week-old seedlings.
DNA products after different numbers of amplification cycles are shown.
OsMDP1 in brassinosteroid signalling 527
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 519–531

Previous studies showed that OsBLE2 was co-ordinately
regulated by BR, IAA and GA (Yang et al., 2003), and theXTH-
related gene OsXTR1 was upregulated by BR and GA (Uozu
et al., 2000). IAA-enhanced lamina bending might be due to
enhanced expression of OsBLE2 and suppressed expression
of OsMDP1 in a manner consistent with that postulated for
BR. Similarly, GA might regulate lamina bending by stimu-
lating the expression ofOsBLE2 andXTHs. BR is an important
regulator of photomorphogenesis, and transcription of many
BR-related genes can be induced by darkness, including that
ofOsBRI1 (Friedrichsen et al., 2000).OsMDP1 is repressed by
the lamina bending-promoting factors BR and IAA, and is
induced by the lamina bending-promoting condition of
darkness, suggesting that it plays a negative regulatory role
in BR signalling during lamina bending. In this case, the dark-
inducible expression of OsMDP1 is a prerequisite for its
physiological function in vivo, providing insight into the
mechanisms of BR signalling in rice. However, as OsBRI1
may not be the only receptor for BR (Sharma et al., 2001), it is
interesting to speculate whether OsMDP1 is involved in the
signalling pathway by which BR is sensed by OsBRI1. The
altered multiple genes involved in signalling and gene
transcription, revealed by microarray analysis using Affyme-
trix Chip, may help in further studies.
In conclusion, OsMDP1 plays a negative regulatory role
in BR-mediated rice lamina joint inclination through neg-
ative regulation of cell expansion-related genes. The study
suggests the existence of other BR-responsive positive
downstream regulators in the rice lamina joint, which will
be a target for future studies. Going forward, it will be
valuable to identify OsMDP1-interacting proteins that may
reveal new regulatory mechanisms, and to determine
whether OsMDP1 exerts a similar role in culm internode
elongation to its role in bending of the lamina joint.
Together, our novel data shed light on the molecular
mechanisms underlying BR signalling, and may allow
researchers to explore further the biological functions of
endogenous BR in rice.
Table 1 Altered genes under OsMDP1 deficiency
Regulation Protein
Cell growth and structureI (4.2) Cyclin-like F-boxI (4.2) Cyclin-like F-boxI (2.9) Actin/actin-likeI (2.1) Tubby/Cyclin-like F-boxI (1.8) Cyclin-like F-boxI (1.8) ChitinaseI (1.8) Putative chitinaseI (1.3) Wall-associated protein kinaseI (1.3) Cyclin-like F-boxI (1) Cyclin-like F-boxI (1) SKP1 component of cyclinS ()1) Actin/actin-likeS ()1) MAP65/ASE1S ()1) Tubulin/FtsZ, GTPaseS ()1.2) Cyclin, N-terminal domainS ()1.3) MAP65/ASE1 (microtubule associated)S ()1.3) Cyclin, N-terminal domainS ()1.6) Cyclin-like F-boxS ()1.7) Expansin Os-EXPA2S ()2) Cyclin, C-terminalS ()2) Cyclin, C-terminalS ()2.1) Cyclin-like F-boxS ()4) Cyclin, C-terminalS ()4.3) Cyclin-like F-box
Gene transcriptionI (1.3) Myb DNA-binding domainI (1.3) Helix-turn-helix motifI (1.3) Basic helix-loop-helixI (1.3) Dimerization domain bHLHI (1.2) Myb DNA-binding domainI (1.2) Basic helix-loop-helix dimerization
domain bHLHI (1.1) Zn-finger, RINGI (1) Myb DNA-binding domainI (1) Zn-finger, B-boxI (1) Sterol desaturaseS ()1) Myb DNA-binding domainS ()1) Zn-finger, RINGS ()1.1) Myb DNA-binding domainS ()1.3) DNA-binding WRKYS ()1.3) E2F/dimerisation partner (TDP)S ()1.4) Zn-finger, RINGS ()1.4) Zn-finger, Dof typeS ()1.5) GRAS family transcription factorS ()1.7) Basic helix-loop-helix dimerization
domain bHLHS ()2.2) Myb DNA-binding domainS ()2.6) MADS-boxS ()3.7) Zn-finger, CCHC typeS ()3.8) CBF/NF-Y/archaeal histone
Signal transductionI (3.6) Putative MAP kinaseI (3.1) Ras GTPaseI (1.1) Calcium-binding EF-handI (1.1) G-protein beta WD-40 repeatI (1) Cadmium-induced protein-likeI (1) Calcium-binding EF-handS ()1) Calcium-binding EF-handS ()1) Ras GTPase superfamily
Table 1 (Continued.)
Regulation Protein
S ()1.2) Putative calmodulinS ()4.2) G-protein beta WD-40 repeat
Hormone functionI (2.8) Putative ethylene-forming enzymeI (1.3) Auxin responsive SAUR proteinI (1.2) Auxin efflux carrierS ()1.1) Transcriptional factor B3/Auxin response factorS ()1.8) Putative auxin response transcription factorS ()4) Auxin responsive SAUR protein
RNAs extracted from transgenic line A1 were used for hybridization.Regulatory ratios are indicated as log 2.I, induced; S, suppressed.
528 Ke Duan et al.
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 519–531

Experimental procedures
Plant material and growth conditions
Rice plants (Oryza sativa, japonica cultivar Zhonghua 11) werecultivated for 3 months in a phytotron with a light/dark cycle of12 h 28�C/12 h 22�C. For hormone treatments, 2-week-old seed-lings were treated in a final concentration of 1 lM IAA (indole-3-acetic acid, I2886, Sigma), 0.1 lM 24-eBL (24-epibrassinolide,E1641, Sigma, St Louis, MO, USA), 100 lM GA3 (gibberellic acid,G7645, Sigma) and 100 lM ABA (A1049, Sigma), respectively.Treated materials were harvested at different time points for RNAextraction. For dark treatment, seeds were germinated and grownfor 10 days in darkness.
Isolation of full-length OsMDP1 cDNA
A 500-bp rice cDNA clone, S110, identified through fluorescencedifferential display PCR analysis of rice 24-epibrassinolide-regula-ted genes, shared a high homology to the plant MADS-box genes(Z-Q. Chu et al., unpublished data). Specific primers P11 (5¢-CTG-TTCAAGAAGGCCGAGGA-3¢) and P0 (5¢-AGTCCATCGATCTCATCC-AGA-3¢) were designed based on the cDNA sequence of S110 andused for PCR-based screening of a rice cDNA library constructedfrom rice materials at tillering stage (Alfandari and Darribere, 1994).The obtained positive clone was sequenced by Genecore (GenecoreBio Technologies Co., Ltd., Shanghai, China). Computational ana-lysis was performed with the help of Wisconsin Genetics ComputerGroup software (GCG ver. 10.1). The BLASTsearch programs providedthrough the NCBI web interface (http://www.ncbi.nlm.nih.gov/BLAST) were used for sequence comparisons involving the Gen-Bank, EMBL, dbEST and SwissProt databases. Sequence align-ments were performed using the BESTFITand PILEUPprograms.
Semi-quantitative RT–PCR analysis
RT–PCR was used to study the expression pattern of OsMDP1. TotalRNAs were extracted from 2-week-old rice roots and seedlings,mature culms and leaves (from 45-day-old rice), flowering panicles,developing seeds at 4, 8, 12, 16, 20 and 24 days after anthesis (DAA)and mature seeds with Trizol solution (Huashun, Shanghai, China).The total RNAs were reverse-transcribed into first-strand cDNA asinstructed (RT–PCR kit, TaKaRa Shuzo, Dalian, China), and thecDNAs obtained were used as templates for PCR amplification withspecific primers P11 and P0. The rice actin coding gene wasamplified using primers OsActin1 (5¢-GAACTGGTATGGTCAAG-GCTG-3¢) and OsActin2 (5¢-ACACGGAGCTCGTTGTAGAAG-3¢) andused as an internal positive control. All utilized primers were locatedin separated exons to exclude the possibility of DNA contamination.
For further studies of the expression of BR signalling-relatedgenes in OsMDP1-deficient plants, total RNAs were extracted from2-cm-long leaf joints (consisting of blade and sheath equal inlength) of 2-week-old green seedlings. The primers used are asfollows: OsMDP1 (P11 and KMA, 5¢-TGTAATCGGTTGCTTT-CAGCTGG-3¢); OsBRI1 (Yamamuro et al., 2000; OsBRI1-S, 5¢-CCGACGATGCTGAAGGTGA-3¢ and OsBRI1-A, 5¢-GCGATTGCTAA-GCTTGTGTGG-3¢); two xyloglucan endotransglycosylase-codinggenes OsXTR1 (Uozu et al., 2000; OsXTR1-S, 5¢-TACCCGTTCAACC-AGCCGA-3¢ and OsXTR1-A, 5¢-GCCAGCAAGCATCGATCACAG-3¢);XTH1 (AF443603, XTH1-S, 5¢-CCTCGCAGCACATCCTTGT-3¢ andXTH1-A, 5¢-TCGAGTCCGCGTTCATCAAAG-3¢); and OsCKI1(AJ487966, OsCKI1–1, 5¢-TACGGAGAGCACACAAAGCACAG-3¢ andOsCKI1–2, 5¢-TCCTCAAAGTGGGATCATCATGG-3¢).
Promoter–reporter gene fusion studies
For further analysis of OsMDP1 expression, the 1.5-kb promoterregion of the gene was PCR amplified with primers KP1 (5¢-TCAAATGCTCCCTCCGTCAG-3¢) and KP2 (5¢-TCAGATCACACAC-CAATTCAC-3¢), then subcloned into vector pCambia1301 (Cambia,Canberra, Australia) resulting in fusion of the promoter to theb-glucuronidase (GUS) reporter gene. The resulting construct wasconfirmed by sequencing, then transformed into the rice genome byAgrobacterium tumefaciens (strain EHA105)-mediated transforma-tion with immature embryo as materials (Hiei et al., 1994). DNAfrom resistant plants was PCR amplified with primers KP1 andGUS-R (5¢-AGACTTCGCGCTGATACCAG-3¢) to confirm integrationof the T-DNA into the rice genome. The progeny of individualconfirmed transformants were harvested and further selected bygermination on culture medium supplemented with 40 lg ml)1
hygromycin. GUS activities were detected histochemically accord-ing to the methods of Jefferson et al. (1987).
Real-time quantitative RT–PCR (RT–qPCR) analysis
RT–qPCR analyses were performed to study both the expressionpattern ofOsMDP1 under diverse hormone treatments (BR, IAA, ABAand GA), and altered transcripts of selected genes (XTH1, XTR1 andBRI1) in the transgenic plants. Total RNAs were extracted from theon-ground part of 2-week-old rice plants with Trizol solution (Hua-shun). Reverse transcription was performed with 1 lg total RNA withan RT–PCR kit (TaKaRa Shuzo) and quantitative analyses were car-ried out on a RotorGene 3000 (Corbett Research, Sydney, Australia)using a SYBR green detection protocol according to the manufac-turer’s instructions (SYBR Premix Ex Taq system, TaKaRa). Theamount of product was determined at the end of each cycle by theROTORGENE software. Expressions of the diverse samples after dif-ferent cycles during the linear amplification phase were used toexamine the differential gene expression, which was indicated asrelative expression levels (percentage) compared with the transcriptof rice OsActin gene. Primers used for PCR amplification were: Os-Actin1 (forward: 5¢-GAACTGGTATGGTCAAGGCTG-3¢ and reverse:5¢-ACACGGAGCTCGTTGTAGAAG-3¢), OsMDP1 (forward: 5¢-TTA-TTGACCGGTACAACTCGCA-3¢ and reverse: 5¢-TCCAGTCCATC-GATCTCATCC-3¢), OsXTH1 (forward: 5¢-GATCCGGGAGATGAAGAACCA-3¢ and reverse: 5¢-TCCATCTGCTGGTCGTACCC-3¢); Os-XTR1 (forward: 5¢-GGAGCCGTACATCCTGCAGA-3¢ and reverse 5¢-AGGCTGGAGTAGAGCTTCATCG-3¢); and Os BRI1 (forward: 5¢-CT-TGGTTCCCTTGCCACATTT-3¢ and reverse 5¢-CCTCATTCTTCAGCC-TCCGC-3¢).
Quantification of each cDNA sample was examined twice and theaveraged data analysed statistically.
Generation of OsMDP1-deficient rice plants through
antisense strategy
The physiological roles of OsMDP1 were studied through antisenseknock-down. A cDNA fragment (814 bp) was PCR-amplified usingprimers P11 and P4 (5¢-CAGGTAACATCCAAGAACAG-3¢) with PfuDNA polymerase (SC0014, Sangon, Shanghai) and subcloned intovector p35S-1301 (modified from pCambia1301 by insertion of aCaMV 35S promoter and a NOS terminator into the Multiple CloningSite). The resulting construct, harbouring the OsMDP1-codingregion in an antisense orientation under control of the CaMV35Spromoter, was used for rice transformation. To confirm the trans-genic lines, resistant plants were analysed via two strategies: (1)genomic DNAs were used as templates for PCR analysis by primers
OsMDP1 in brassinosteroid signalling 529
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 519–531

35Sp (forward: 5¢-ATGCCATCATTGCGATAAAGG -3¢) and P11 toconfirm the integration of the T-DNA into the rice genome; and (2)lamina joint RNAs were extracted and RT–PCR was used to test forsuppression of OsMDP1 expression.
After the primary transformation, >20 independent resistant T0
lines were obtained and confirmed as transformants. The progenyof the individual transformants were harvested and further selectedby germination on culture medium supplemented with 40 lg ml)1
hygromycin. Single-copy integration of the T-DNA was confirmedby a germination segregation ratio analysis (a 3:1 ratio of germina-ted to ungerminated seeds was expected). Eight independenthomozygous lines were obtained after two generations and usedfor phenotypic observation.
Lamina joint inclination bioassay and BR treatments
Rice seeds were surface-sterilized, soaked in distilled water andimbibed for 48 h in the dark at 30�C. Synchronously germinatedseeds were selected and cultivated in 1% agar in a dark chamber for8 days, and the rice lamina joint inclination bioassay was performedas described by Wada et al. (1981).
To test the effects of BR on coleoptiles and roots, >20 seeds ofeach independent transgenic line and control plants were sowed invertically oriented agar plates supplemented with or without 0.1 lM
24-eBL. After 7 days’ growth in darkness, the lengths of thecoleoptiles and primary roots were measured and the seedlingswere photographed.
Preparation of sections for SEM analysis
The second lamina joint (0.5 cm long) was excised from dark-growntransgenic and control seedlings with or without 24-eBL treatment,then fixed in formalin:glacial acetic acid:50% ethanol (2:1:17, v:v:v).After dehydration in a graded ethanol series, the sections were driedfor 2 h (Hitachi critical point dryer, HCP-2; Hitachi Koki Co., Ltd.,Tokyo, Japan), surface-sprayed with gold powder, and examinedunder a scanning electron microscope (Hitachi S-450).
Gene-expression profiles under OsMDP1 deficiency via
cDNA microarray analysis
Seeds of wild-type and transgenic plants (line A1) were surface-sterilized and germinated in the dark. Sections of second laminajoint (2 cm long) were excised from the seedlings of 1-week-oldwild-type (control) and transgenic plants, respectively, and frozen inliquid N2 for RNA isolation. Total RNA was isolated using theRNAiso Reagent (TaKaRa) and further cRNA was prepared followingthe manufacturer’s instructions (Affymetrix, Santa Clara, CA, USA:http://www.affymetrix.com/support/technical/manual).
The Affymetrix arrays (900599 GeneChip Rice Genome Array)contain 54 168 probe sets representing a single array. Labelling andhybridization (one sample per chip) were performed by Gene TechBiotechnology (Shanghai) and the probe arrays were scanned andanalysed with MICROARRAY SUITE ver. 5.0 (Affymetrix). A coefficientof log 2 was used as the selection criterion for differential geneexpression between control and transgenic plants.
Acknowledgements
The studies were supported by programs 2005CB120803 and2002AA2Z1003. We greatly thank Xiao-Yan Gao for help with SEMobservations.
Supplementary Material
The following supplementary material is available for this articleonline:Table S1 Functional description of induced genes under OSMDP1deficiencyTable S2 Functional description of suppressed genes underOSMDP1 deficiencyThis material is available as part of the online article from http://www.blackwell-synergy.com
References
Alfandari, D. and Darribere, T. (1994) A simple PCR method forscreening cDNA libraries. PCR Methods Appl. 4, 46–49.
Becker, A. and Theissen, G. (2003) The major clades of MADS-boxgenes and their role in the development and evolution offlowering plants. Mol. Phylogenet. Evol. 29, 464–489.
Bishop, G.J. and Koncz, C. (2002) Brassinosteroids and plant steroidhormone signaling. Plant Cell, 14, S97–S110.
Bishop, G.J. and Yokota, T. (2001) Plant steroid hormones, brassi-nosteroids: current highlights of molecular aspects on their syn-thesis/metabolism, transport, perception and response. Plant CellPhysiol. 42, 114–120.
Cao, H.P. and Chen, S.K. (1995) Brassinosteroid-induced rice laminajoint inclination and its relation to indole-3-acetic acid and ethy-lene. Plant Growth Regul. 16, 189–196.
De Bodt, S., Raes, J., Van de Peer, Y. and Theissen, G. (2003) Andthen there were many: MADS goes genomic. Trends Plant Sci. 8,475–483.
Friedrichsen, D.M., Joazeiro, C.A., Li, J., Hunter, T. and Chory, J.
(2000) Brassinosteroid-insensitive-1 is a ubiquitously expressedleucine-rich repeat receptor serine/threonine kinase. Plant Phys-iol. 123, 1247–1256.
Friedrichsen, D.M., Nemhauser, J., Muramitsu, T., Maloof, J.N.,
Alonso, J., Ecker, J.R., Furuya, M. and Chory, J. (2002) Threeredundant brassinosteroid early response genes encode putativebHLH transcription factors required for normal growth. Genetics,162, 1445–1456.
Fujioka, S., Noguchi, T., Takatsuto, S. and Yoshida, S. (1998)Activity of brassinosteroids in the dwarf rice lamina inclinationbioassay. Phytochemistry, 49, 1841–1848.
He, J.X., Gendron, J.M., Sun, Y., Campala, S.S., Gendron, N., Sun,
C.Q. and Wang, Z.Y. (2005) BZR1 is a transcriptional repressorwith dual roles in brassinosteroid homeostasis and growth re-sponses. Science, 307, 1634–1638.
Hiei, Y., Ohta, S., Komari, T. and Kumashiro, T. (1994) Efficienttransformation of rice (Oryza sativa L.) mediated by Agrobacte-rium and sequence analysis of the boundaries of the T-DNA. PlantJ. 6, 271–282.
Hong, Z., Ueguchi-Tanaka, M., Shimizu-Sato, S. et al. (2002)Loss-of-function of a rice brassinosteroid biosynthetic enzyme,C-6 oxidase, prevents the organized arrangement and polarelongation of cells in the leaves and stem. Plant J. 32, 495–508.
Hong, Z., Ueguchi-Tanaka, M., Umemura, K., Uozu, S., Fujioka, S.,
Takatsuto, S., Yoshida, S., Ashikari, M., Kitano, H. and Matsuoka,
M. (2003) A rice brassinosteroid-deficient mutant, ebisu dwarf(d2), is caused by a loss of function of a new member of cyto-chrome P450. Plant Cell, 15, 2900–2910.
Hong, Z., Ueguchi-Tanaka, M., Fujioka, S., Takatsuto, S., Hasegawa,
Y., Ashikari, M., Kitano, H. and Matsuoka, M. (2005) The ricebrassinosteroid-deficient dwarf2 mutant, defective in the ricehomolog of Arabidopsis DIMINUTO/DWAERF1, is rescued by the
530 Ke Duan et al.
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 519–531

endogenously accumulated alternative bioactive brassinosteroid,dolichosterone. Plant Cell, 17, 2243–2254.
Jack, T. (2001) Plant development going MADS. Plant Mol. Biol. 46,515–520.
Jefferson, R.A., Kavanagh, T.A. and Bevan, M.W. (1987) GUS fu-sions: beta-glucuronidase as a sensitive and versatile gene fusionmarker in higher plants. EMBO J. 6, 3901–3907.
Johansen, B., Pedersen, L.B., Skipper, M. and Frederiksen, S. (2002)MADS-box gene evolution – structure and transcription patterns.Mol. Phylogenet. Evol. 23, 458–480.
Kauschmann, A., Jessop, A., Koncz, C., Szekeres, M., Willmitzer,
L. and Altmann, T. (1996) Genetic evidence for an essentialrole of brassinosteroids in plant development. Plant J. 9, 701–713.
Koka, C.V., Cerny, R.E., Gardner, R.G., Noguchi, T., Fujioka, S.,
Takatsuto, S., Yoshida, S. and Clouse, S.D. (2000) A putative rolefor the tomato genes DUMPY and CURL-3 in brassinosteroidbiosynthesis and response. Plant Physiol. 122, 85–98.
Konishi, H. and Komatsu, S. (2003) A proteomics approach toinvestigating promotive effects of brassinolide on lamina incli-nation and root growth in rice seedlings. Biol. Pharm. Bull. 26,401–408.
Lee, S., Kim, J., Son, J.S. et al. (2003) Systematic reverse geneticscreening of T-DNA tagged genes in rice for functional genomicanalyses: MADS-box genes as a test case. Plant Cell Physiol. 44,1403–1411.
Liu, W., Xu, Z.H., Luo, D. and Xue, H.W. (2003) Roles of OsCKI1, a ricecasein kinase I, in root development and plant hormone sensi-tivity. Plant J. 36, 189–202.
Maeda, E. (1965) Rate of lamina inclination in excised rice leaves.Physiol Plant. 18, 813–827.
Messenguy, F. and Dubois, E. (2003) Role of MADS box proteins andtheir cofactors in combinatorial control of gene expression andcell development. Gene, 316, 1–21.
Mori, M., Nomura, T., Ooka, H. et al. (2002) Isolation and charac-terization of a rice dwarf mutant with a defect in brassinosteroidbiosynthesis. Plant Physiol. 130, 1152–1161.
Muessig, C. and Altmann, T. (2001) Brassinosteroid signaling inplants. Trends Endocrinol. Metab. 12, 398–402.
Nakashita, H., Yasuda, M., Nitta, T., Asami, T., Fujioka, S., Arai, Y.,
Sekimata, K., Takatsuto, S., Yamaguchi, I. and Yoshida, S. (2003)Brassinosteroid functions in a broad range of disease resistancein tobacco and rice. Plant J. 33, 887–898.
Ng, M. and Yanofsky, M.F. (2001) Function and evolution of theplant MADS-box gene family. Nat. Rev. Genet. 2, 186–195.
Parenicova, L., de Folter, S., Kieffer, M. et al. (2003) Molecular andphylogenetic analyses of the complete MADS-box transcriptionfactor family in Arabidopsis: new openings to the MADS world.Plant Cell, 15, 1538–1551.
Pullman, G.S., Zhang, Y. and Phan, B.H. (2003) Brassinolide im-proves embryogenic tissue initiation in conifers and rice. PlantCell Rep. 22, 96–104.
Sharma, A., Matsuoka, M., Tanaka, H. and Komatsu, S. (2001)Antisense inhibition of a BRI1 receptor reveals additional proteinkinase signaling components downstream to the perception ofbrassinosteroids in rice. FEBS Lett. 507, 346–350.
Tanabe, S., Ashikari, M., Fujioka, S. et al. (2005) A novel cytochromeP450 is implicated in brassinosteroid biosynthesis via the char-acterization of a rice dwarf mutant, dwarf11, with reduced seedlength. Plant Cell, 17, 776–790.
Uozu, S., Tanaka-Ueguchi, M., Kitano, H., Hattori, K. and Matsuoka,
M. (2000) Characterization of XET-related genes of rice. PlantPhysiol. 122, 853–859.
Wada, K., Marumo, S., Ikekawa, N., Morisaki, M. and Mori, K. (1981)Brassinolide and homobrassinolide promotion of lamina incli-nation of rice seedlings. Plant Cell Physiol. 22, 323–325.
Wada, K., Marumo, S., Abe, H., Morishita, T., Nakamura, K.,
Uchiyama, M. and Mori, K. (1984) A rice lamina inclination test – amicro-quantitative bioassay for brassinosteroids. Agric. Biol.Chem. 48, 719–726.
Wang, Z.Y., Nakano, T., Gendron, J., He, J., Chen, M., Vafeados, D.,
Yang, Y., Fujioka, S., Yoshida, S. and Asami, T. (2002) Nuclear-localized BZR1mediates brassinosteroid-induced growth andfeedback suppression of brassinosteroid biosynthesis. Dev. Cell2, 505–513.
Yamamuro, C., Ihara, Y., Wu, X., Noguchi, T., Fujioka, S., Takatsuto,
S., Ashikari, M., Kitano, H. and Matsuoka, M. (2000) Loss offunction of a rice brassinosteroid insensitive1 homolog preventsinternode elongation and bending of the lamina joint. Plant Cell,12, 1591–1606.
Yang, G. and Komatsu, S. (2000) Involvement of calcium-dependentprotein kinase in rice (Oryza sativa L.) lamina inclination causedby brassinolide. Plant Cell Physiol. 41, 1243–1250.
Yang, G., Matsuoka, M., Iwasaki, Y. and Komatsu, S. (2003) A novelbrassinolide-enhanced gene identified by cDNA microarray isinvolved in the growth of rice. Plant Mol. Biol. 52, 843–854.
Yin, Y., Vafeados, D., Tao, Y., Yoshida, S., Asami, T. and Chory, J.
(2005) A new class of transcription factors mediates brassino-steroid-regulated gene expression in Arabidopsis. Cell, 120, 249–259.
The nucleotide sequences reported in this paper have been submitted to the GenBank/EMBL Data Bank under accession number
AJ536158.
OsMDP1 in brassinosteroid signalling 531
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 519–531
Copyright © 2022 FDOKUMEN




















