
"NaV1.5 Na+ channels allosterically regulate the NHE-1 exchanger ...

Multiple Signals Regulate PLC beta 3 in Human Myometrial Cells
Miao Zhong#, Dilyara A. Murtazina#, Jennifer Phillips, Chun-Ying Ku, and Barbara M.Sanborn*Department of Biomedical Sciences, Colorado State University, Fort Collins, Colorado 80523
SummaryThe regulation of PLCB3-Serine1105 phosphorylation by both negative feedback and negativecrosstalk facilitates the integration of multiple signaling pathways in myometrial cells.
Phospholipase CB3 (PLCB3) Serine1105, a substrate for multiple protein kinases, represents apotential point of convergence of several signaling pathways in the myometrium. To explore thishypothesis, the regulation of PLCB3-Serine1105 phosphorylation (P-S1105) was studied inimmortalized and primary human myometrial cells. CPT-cAMP and calcitonin gene-related peptide(CALCA) transiently increased P-S1105. Relaxin also stimulated P-S1105; this effect was partiallyblocked by the protein kinase A (PRKA) inhibitor Rp-8-CPT-cAMPS. Oxytocin, which stimulatesGαq-mediated pathways, also rapidly increased P-S1105, as did PGF2α and ATP. Oxytocin-stimulated phosphorylation was blocked by the protein kinase C (PRKC) inhibitor Go6976 and bypretreatment overnight with a phorbol ester. Cypermethrin, a PP2B phosphatase inhibitor, but notokadaic acid, a PP1/PP2A inhibitor, prolonged the effect of CALCA on P-S1105, whereas the reversewas the case for the oxytocin-stimulated increase in P-S1105. PLCB3 was the predominant PLCisoform expressed in the myometrial cells and PLCB3 shRNA constructs significantly attenuatedoxytocin-stimulated increases in intracellular calcium. Oxytocin-induced phosphatidylinositol (PI)turnover was inhibited by CPT-cAMP and okadaic acid but enhanced by pretreatment with Go6976.CPT-cAMP inhibited oxytocin-stimulated PI turnover in the presence of overexpressed PLCB3, butnot overexpressed PLCB3-S1105A. These data demonstrate that both negative crosstalk from thecAMP/PRKA pathway and a negative feedback loop in the oxytocin/G protein/PLCB pathwayinvolving PRKC operate in myometrial cells and suggest that different protein phosphatasespredominate in mediating P-S1105 dephosphorylation in these pathways. The integration of multiplesignal components at the level of PLCB3 may be important to its function in the myometrium.
Keywordsmyometrium; phospholipase CB3; protein kinase A; protein kinase C; protein phosphatase;phosphorylation
INTRODUCTIONThe myometrium maintains relative quiescence during pregnancy, exhibits sporadiccontractures near term and gradually develops the coordinated, rhythmic contractions of labor.The biochemical changes that account for this transition reflect a shift in the relativepredominance of relaxant and contractant pathways, but are not completely understood at themolecular level [1-3]. The influence of cyclic AMP, a second messenger linked to myometrial
#Co-equal first authors.*Corresponding author and person to whom reprint requests should be made: Barbara M. Sanborn, Ph.D., Department of BiomedicalSciences, 102 Physiology Campus Delivery 1680, Colorado State University, Fort Collins, CO 80523, Phone: (970)-491-4263, Fax:(970)-491-7569, E-mail: [email protected]
NIH Public AccessAuthor ManuscriptBiol Reprod. Author manuscript; available in PMC 2009 June 1.
Published in final edited form as:Biol Reprod. 2008 June ; 78(6): 1007–1017.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

relaxation, diminishes near the end of pregnancy. In contrast, Gαq/phospholipase CB (PLCB)pathways mediating the signaling of a number of uterine contractants, including oxytocin(OXT), prostaglandins E2 and F2α and ATP, are enhanced near parturition.
Phosphatidylinositide-specific phospholipase C (PLC) enzymes catalyze the hydrolysis ofphosphatidylinositol 4,5-bisphosphate (PIP2) to produce inositol 1,4,5-trisphosphate (IP3) anddiacylglycerol (DAG). Six subfamilies of PLCs (B, G, D, E, Z and H) constitute part ofubiquitous signaling cascades that translate hormonal signals into intracellular events, leadingto alternations in cell function [4;5]. PLCB isoforms 1-4 are stimulated by G-protein activation(Gαq/11 and/or Gβγ) [4;5].
Several PLCB isoforms can be phosphorylated by protein kinases, and this may contribute tothe regulation of their activities [6]. Protein kinase C (PRKC) phosphorylates and negativelyregulates PLCB1 activity [7], but PLCB1 is not directly phosphorylated by protein kinase A(PRKA) [8]. PLCB2 is a substrate of PRKA; this phosphorylation accounts for the inhibitionby cAMP of Gβγ-induced PLCB2 activity in vitro [9]. PRKA phosphorylates PLCB3exclusively on Serine1105 (S1105) and inhibits both Gαq and Gβγ-stimulated activity intransfected cells [8;10]. S1105 is conserved between a number of species in a reasonableconsensus PRKA target sequence in PLCB3, but a comparable sequence is not present in theother PLCB isoforms [11]. Interestingly, PLCB3-S1105 is also phosphorylated by activation ofprotein kinase C (PRKC) and protein kinase G [10;12]. In the latter case, phosphorylation atanother residue (Serine26) was required for full inactivation [12] Hence, PLCB3 may beregulated by multiple protein kinases and control hormonal signaling. Regulation of PLCB4activity by protein kinases has not been reported to date.
In myometrial smooth muscle, the contractant hormone oxytocin signals through a G-protein-coupled receptor that primarily couples to Gαq/11 to activate PLCBs [13;14]. Cyclic AMPgenerally induces relaxation in smooth muscles [15]. The inhibition of oxytocin-inducedphosphatidylinositide (PI) turnover by increasing intracellular cAMP in myometrial cells isboth PRKA and A-Kinase Anchoring Protein (AKAP) dependent [16;17]. Activation of PRKAby 8-[4-chlorophenylthio]-cAMP (CPT-cAMP) inhibits GTPγS-stimulated PI turnover in ratmyometrial plasma membrane, indicating that the effect of PRKA is distal to oxytocin receptor/G protein coupling [18].
To examine the functional importance of specific PLCB3 phosphorylation sites in hormonesignaling in myometrial cells, we have examined the effect of stimulating PRKA and PRKCpathways on the phosphorylation of endogenous PLCB3-S1105. We demonstrate that PLCB3is the predominant PLCB isoform in the human myometrial cells used. The data support thepresence of negative crosstalk from relaxant-stimulated cAMP pathways, as well as negativefeedback from the oxytocin-induced PLCB activation pathway itself, targeted at PLCB3-S1105. In addition, the data suggest that protein phosphatase 2B regulates PRKA-mediated P-S1105 by CALCA whereas PP1 or PP2A regulate oxytocin-stimulated PRKC-dependent P-S1105 in PHM1 cells.
MATERIALS AND METHODSMaterials
[3H]-myoinositol (22.3 Ci/mmol) was obtained from Perkin-Elmer Life Sciences (Boston,MA) and bovine serum albumin (fatty acid free, fraction V) (BSA) from ICN Biomedicals Inc.(Irvine, CA). Calcitonin gene-related peptide (CALCA, rat) was obtained from TocrisBioscience (Ellisville, MO). Cypermethrin, Go6976, okadaic acid, oxytocin, and phosphataseinhibitor cocktail Set 1 were obtained from Calbiochem (La Jolla, CA). CPT-cAMP, 8-[4-chlorophenylthio]-cAMP-thioate, Rp-isomer (Rp-8-CPT-cAMPS), 3-isobutyl-1-
Zhong et al. Page 2
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

methylxanthine (IBMX), phorbol 12-myristate 13-acetate (PMA), ATP, PGF2α tris salt,Reactive Blue-2 and protease inhibitor cocktail (# P8340) were obtained from Sigma-Aldrich(St. Louis, MO). Protein kinase and protein phosphatase inhibitors were used at concentrationsapproximately 1000 times higher than their Ki values. Fura-2/acetoxymethylester (Fura-2-AM) was purchased from Invitrogen (Carlsbad, CA). Polyclonal antibodies against humanPLCB1, PLCB3, human PRKA catalytic subunit and horseradish peroxidase conjugateddonkey anti-rabbit secondary antibody (1:2000) were obtained from Santa Cruz BiotechnologyInc. (Santa Cruz, CA). Polyclonal antibody against phosphorylated PLCB3-S1105 wasproduced for us by Cell Signaling Technology (Beverly, MA). Enhanced chemiluminescence(ECL) reagent was obtained from Amersham Biosciences (Piscataway, NJ). Primers werepurchased from Integrated DNA Technologies, Inc. (Coralville, IA). iScript One-Step RT-PCRKit with SYBR Green and AG 1-X8 resin were purchased from Bio-Rad Laboratories, Inc.(Hercules, CA). Restriction enzymes were obtained from New England Biolabs Inc. (Beverly,MA) or Promega (Madison, WI). SureSilencing short hairpin RNA (shRNA) plasmids forhuman PLCB3 (# KH02817G) were obtained from SuperArray Bioscience Corporation(Frederick, MD). The cAMP ELISA kit was obtained from Assay Designs, Inc. (Ann Arbor,MI). AG 1-X8 resin was obtained from Bio-Rad Laboratories, Inc. (Hercules, CA).Nitrocellulose membranes were purchased from Whatman Schleicher & Schuell (FlorhamPark, NJ).
Primary uterine smooth muscle cells (UtSMC) were obtained from Cambrex (Walkersville,MD). AD293 cells were obtained from Stratagene (La Jolla, CA). Human myometrial pre-termlabor samples were obtained at the time of cesarean section under approved IRB protocols atthe University of Texas Medical School at Houston (Sanborn and Monga) and the Universityof Texas Southwestern Medical School (Sanborn and Word).
PLCB3 Adenoviral and psi-CHECK-2 Reporter ConstructioncDNAs encoding human PLCB3 and PLCB3-S1105A were excised from pCR3.1 vectors [8]and subcloned into the Not I/Xho I sites of the pAdTrack-CMV shuttle vector (Stratagene,Cedar Creek, TX). These vectors were used to produce adenoviral vectors expressing PLCB3or the S1105A mutant according to the manufacturer’s instructions. The multiplicity of infection(MOI) for each adenovirus was determined in AD293 cells as recommended by themanufacturer.
For use in shRNA silencing experiments, the PLCB3 sequence was subcloned into the SgfI/PmeI sites of the psi-CHECK-2 vector (Promega, Madison, WI), located downstream of thereporter gene (Renilla luciferase). The resulting PLCB3-psiCHECK-2 (psiPLCB3) constructwas used in reporter assays.
Cell CultureImmortalized PHM1-41 cells derived from pregnant human myometrial tissue (passages19-23) [19], UtSMC and AD293 cells were cultured in Dulbecco modified Eagle medium(DMEM) containing 10% fetal calf serum, 2 mM L-glutamine, 50 units/ml penicillin, and 50μg/ml streptomycin. Myometrial cells were trypsinized and plated (2 × 105 cells/well) into 6-well plates for PI turnover and cAMP assays or into 35-mm dishes (1-2 × 105 cells/dish) forthe PLCB3 phosphorylation assay.
Viral Infection and Plasmid ElectroporationIn overexpression studies, adenoviral constructs expressing PLCB3 or the S1105A mutant wereadded at an MOI of 1000 to PHM1 cells at the time of plating, and the cells were used 72 hlater. Under these conditions, the infection efficiency was ∼ 100%, estimated by expression
Zhong et al. Page 3
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

of green fluorescent protein (GFP) from a second promoter. Infected PHM1 cells exhibitedsimilar morphology and PI turnover in response to oxytocin as non-infected cells.
In RNA interference experiments, Suresilencing shRNA plasmid DNA (4 μg) was added to100 μl of Basic Nucleofector Solution (Basic Nuclefection kit, # VPI-1004, AmaxaBiosystems, Gaithersburg, MD) and was electroporated into 1 × 106 PHM1 cells. Cells werepulsed at 140 V/35 ms/2 mm cuvette with time constant protocol using the Genepulser Xcell(Bio-Rad, Hercules, CA). After electroporation, cells were plated into dishes and were culturedfor 72 h before measurement of intracellular calcium. Transfection efficiency was estimatedby fluorescence of modified GFP (mGFP) expressed in the vector from a second promoter.
In the experiments using psiCHECK-2 system, psiPLCB3 (0.5 μg) was co-transfected withPLCB3-shRNA plasmids (1.5 μg) into AD293 cells (1 × 105 cells/12 well plate) withGenePORTER 2 transfection reagent (Genlantis, San-Diego, CA) as recommended by themanufacturer. Samples were analyzed 72 hours after transfection using the Dual-luciferaseReporter Assay (Promega, Madison, WI). Firefly luciferase activity, driven by a secondpromoter, allowed normalization of Renilla-luciferase expression.
Measurement of Intracellular CalciumCells were loaded at room temperature with 5 μM of Fura-2 for 30 min in fluorescence buffer(145 mM NaCl, 5 mM KCl, 1 mM Na2HPO4, 0.5 mM MgCl2, 1mM CaCl2, 10 mM Hepes,and 5 mM glucose; pH 7.4). After loading, the cells were washed twice in the same buffer and,after 30 min, were placed in Ca2+-free buffer (substitution of 100 μM EGTA for 1 mMCaCl2 in fluorescence buffer). Changes in individual cell intracellular free Ca2+ concentrationafter the addition of 100 nM oxytocin were measured at 340-and 380-nm excitation and 510nm emission (Intracellular Imaging, Inc., Cincinnati, OH). The individual responses of 20-36cells in one dish were aligned and averaged using the CalciumComp program produced for usby an engineering consultant.
Membrane Preparation and Protein Phosphorylation AnalysisPHM1 cells and UtSMC were cultured [20] and harvested either as total cell lysates or used toprepare crude membranes (PHM1 cells). For whole cell lysates, PHM1 cells were harvestedin 200 μl lysis buffer (50 mM NaCl, 0.3% Triton X-100). To prepare membranes, cells werecollected in chilled hypotonic buffer (20 mM HEPES pH 7.4, 10 mM EDTA) containing 1.04mM 4-(2_Aminoethyl) benzenesulfonyl fluoride hydrochloride (AEBSF), 0.8 μM aprotinin,21 μM leupeptin, 36 μM bestatin, 15 μM pepstatin A, 14 μM N-(trans-Epoxysuccinyl)-L-leucine 4- guanidinobutylamide (E-64). The collected cells were lysed by passing through a0.5 inch needle (27 gauge). Cell debris was removed by centrifugation at 3,000 g for 15 minat 4° C. Crude membrane fractions were prepared by centrifugation at 100,000 g for 1 h at 4°C, resuspended in phosphate buffered saline (2.68 mM KCl, 1.47 mM KH2PO4, 136.9 mMNaCl, 8.1 mM Na2HPO4, pH 7.2) and stored at -80° C. Protein concentration was quantifiedby the Bradford method (Bio-Rad, Hercules, CA).
Total cell lysates (10-40 μl) or membrane proteins (10 μg) were separated on 6% SDS-PAGEgels and transferred onto nitrocellulose membranes. Phosphorylated PLCB3-S1105 wasdetected with anti-P-S1105 antibody (1:250-1000) using the ECL reagent. Following stripping,total PLCB3 was probed with anti-PLCB3 (1:1000) antibody on the same blot. Inoverexpression experiments, PLCB3 and PRKA catalytic subunits (1:2000) were detectedsequentially on the same blot with different antibodies. PRKA served as an internal controland did not change with treatment. ECL signals were detected using a Storm imager andquantitated using ImageQuant TL software (Amersham Biosciences, Piscataway, NJ).
Zhong et al. Page 4
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

In Vitro PLCB3 PhosphorylationPurified recombinant PLCB3(His)6 (1 μg) and S1105/A-PLCB3 (1 μg) [8] were incubated withor without PRKA-cat subunit (69 μM) in PRKA buffer (10 mM Tris-HCl, pH 7.0, 5 mMMgCl2, 0.1 mM ATP) for 30 min at 30°C, and the reactions were terminated by addition of4X SDS sample buffer (final concentration 62.5 mM Tris-HCl pH 6.8, 4M urea, 10% glycerol,2% SDS, 0.01% β-mercaptoethanol, 0.002% bromophenol blue) and boiled for 5 min. Proteinswere separated on SDS-PAGE (4-15%) and were transferred to nitrocellulose membranes. Theblots were probed with primary (1:500) and secondary antibodies, and the bands visualized byECL. Dephosphorylation of purified recombinant PLCB3(His)6 (1 μg) was carried out at 37 °C for 1 hour in 50 mM Tris-HCl, 10 mM MgCl2, 100 mM NaCl, 1 mM dithiothreitol, pH 7.9,using calf intestine alkaline phosphatase (CIP) (1 unit/1 μg protein) from New England BiolabsInc. (Beverly, MA).
PLCB Isoform RNA and Protein DeterminationRNA was isolated from PHM1 and UtSMC cells with the RNeasy kit (QIAGEN, Valencia,CA) and from human myometrium tissue using TRIZOL reagent (Invitrogen, Carlsbad, CA).Primers for PLCB1, PLCB2, PLCB3, PLCB4 and the RNA normalizer HMBS(hydroxymethylbilane synthase) are presented in Table 1. RT-PCR was performed using theiScript one-step RT-PCR kit with the SYBR Green protocol in a 25 μl reaction volume with500 ng RNA (denatured at 95 °C for 15 min and subjected to 35 cycles of denaturation at 94°C for 15 sec, annealing for 30 sec at 58 °C for PLCB1, PLCB2 and PLCB3 and 60 °C forPLCB4, and extension at 72 °C for 1 min) in an iCycler (Biorad, Hercules, CA). The identitiesof the products were confirmed by sequencing. Melting curves for PLCB1, PLCB2, PLCB3,PLCB4 and HMBS cDNA exhibited one peak. Abundance was determined by the ΔΔCtmethod in which data were normalized to the housekeeping gene HMBS in the same sampleand expressed relative to PLCB3
Quantitation of PLCB1 and PLCB3 protein was determined by Western blot analysis inreference to standard curves generated using known quantities of purified recombinant PLCB1(His)6 and PLCB3(His)6 [8].
PI TurnoverOne day prior to the experiments, PHM1 cells were washed twice with phosphate bufferedsaline (PBS) and cultured in 1 ml DMEM, 0.5% fetal calf serum, 0.4 μM [3H]-myoinositol at37 °C overnight. The labeled cells were washed twice with PBS and were incubated with HanksBalanced Salt Solution (HBSS, pH 7.4) containing 0.2% BSA, 10 mM LiCl and the indicatedinhibitors of protein kinases or phosphatases at 37 °C for 30 min. The cells were then stimulatedfor 30 min with 30 nM oxytocin. Total [3H]-inositol phosphates (IPs) were isolated by ionexchange chromatography and counted as described previously [21].
cAMP AssayPHM1 cells were washed twice with PBS and preincubated with HBSS for 1 h at 37 °C. Thecells were stimulated with 10 nM calcitonin gene related peptide (CALCA) for the intervalsindicated in the figure legends at 37 °C. The medium was removed and the reaction stoppedby the addition of 0.5 ml of 0.1 N HCl. cAMP content in the cell lysate was measured byimmunoassay following the protocol recommended by the manufacturer (Assay Designs, Inc.,Ann Arbor, MI).
Zhong et al. Page 5
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Data AnalysisDose-response curves were analyzed with a four-parameter logistics curve-fitting program (M.L. Jaffe, Silver Spring, MD). Where indicated, data are presented as mean ± SEM and wereanalyzed by ANOVA and Duncan’s modified multiple range test.
RESULTSPLCB3 is the Predominant Isoform in Human Myometrial Cells
To examine the relative expression of PLCB isoforms, we carried out both RNA (real timeQRT-PCR) and protein (Western blot) analysis. PLCB3 mRNA was expressed at 1.6 and 2-fold greater relative abundance than PLCB1 in PHM1 and UtSMC cells, respectively (Fig.1A,B). In contrast, PLCB2 and PLCB4 mRNAs were barely detectable in these samples.
Previous studies had detected PLCB1 and PLCB3, but not PLCB2, protein in PHM1 cells[22]. Since we had recombinant PLCB1 and PLCB3 available, we were able to quantitatePLCB1 and PLCB3 protein in reference to standard curves. PLCB3 was significantly moreabundant than PLCB1 in both PHM1 and UtSMC cell (Fig. 1C). The PLCB3/PLCB1 ratioswere 12.6 ± 2.0 and 21.2 ± 4.9 in PHM1 and UtSMC cells, respectively. In 3 pregnant lateterm human myometrial tissue homogenates, the PLCB3/PLCB1 concentration ratios were1.3, 2.0 and 16.0 (data not shown), suggesting some patient to patient variability.
shRNA interference was used to probe the functional importance of PLCB3 to oxytocinsignaling in myometrial cells. Prior experience has indicated that efficiencies for conventionaltransfection are very poor in these cells. Therefore, we used an electroporation protocol.Plasmids expressing a commercial PLCB3 shRNA (PLCB3sh1) or scrambled sequence wereexpressed at ∼40-50% efficiency in PHM1 cells following electroporation, as judged by mGFPfluorescence. There were no significant differences in calcium responses to oxytocin betweencells electroporated without adding DNA and cells electroporated with plasmid expressingscrambled shRNA (Fig. 2A). Individual PHM1 cells electroporated with PLCB3sh1 shRNAand selected by mGFP fluorescence showed a marked decrease in the ability of oxytocin toincrease intracellular free calcium compared to the scrambled control (43.1 ± 11.5% decreasein the initial rise in intracellular calcium, 39.5 ± 11.2 % decrease in the integrated calciumtransient) (Fig. 2A). Since the experiment was conducted in the absence of extracellularcalcium, primarily the Gq/PLC/IP3-mediated response was being measured. Because of therelatively low electroporation efficiency, it was not possible to accurately assess the amountof suppression of PLCB3 expression by conventional methods in PHM1 cells. However, weconstructed a PLCB3 psiCHECK-2 reporter assay in order to assess the amount of PLCB3mRNA suppression in AD293 cells. Fig. 2B shows that PLCB3sh1 suppressed PLCB3 reporterexpression by 49 ± 2.2% in transfected AD293 cells. We have observed >85% knockdown ofother reporters by their corresponding shRNA plasmids in this assay (A. Ulloa, D. Murtazina,unpublished observations), suggesting that PLCB3sh1 may not be an optimal sequence.Although the knockdown achieved with this commercial plasmid was only partial, the effectof PLCB3sh1 on oxytocin-stimulated calcium transients supports the contention that thePLCB3 isoform plays a significant role in oxytocin-stimulated signal transduction.
Activation of Either Endogenous PRKA or PRKC Results in PLCB3-S1105 PhosphorylationPLCB3- S1105 represents a potential site for regulation by both Gαq/11-stimulated contractantand cAMP-mediated relaxant pathways. To determine if regulation of PLCB3 phosphorylationby these signaling pathways is operable in intact myometrial cells, we arranged for a rabbitpolyclonal antibody against the PLCB3-P-S1105 sequence to be produced. Fig. 3A shows thatthis antibody reacts only weakly with recombinant PLCB3 but detects this protein after it isphosphorylated in vitro with PRKA. In contrast, the antibody does not recognize an epitope in
Zhong et al. Page 6
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

the recombinant S1105A mutant. The weak immunoreactivity observed in the absence of PRKAtreatment probably represents residual endogenous S1105 phosphorylation in the isolatedprotein, as it was reduced by >90% by treatment with alkaline phosphatase (data not shown).
Treatment of PHM1 cells with CPT-cAMP for 10 min induced a dose-dependent increase inP-S1105 in crude membranes (Fig. 3B); the EC50 of this response was 0.21 ± 0.02 mM. Todetermine if an increase in endogenous cAMP would induce a similar P-S1105 response, effectsof hormones that are able to increase intracellular myometrial cAMP, such as relaxin andcalcitonin gene related peptide (CALCA), were examined. Treatment with 1.6 nM relaxin for15 min induced P-S1105, and this was partially inhibited by the PRKA inhibitor Rp-8-CPT-cAMPS (Fig. 4A). Treatment with 10 nM CALCA induced a transient increase in intracellularcAMP and in P-S1105 (Fig. 4B) with an EC50 of 0.33 ± 0.14 nM (Fig. 4C). Pretreatment withthe PRKC inhibitor Go6976 lowered basal phosphorylation but did not alter the net responseto CALCA-induced P-S1105 (Fig. 4C). Stimulation of P-S1105 by 10 nM CALCA was alsoobserved in primary UtSMC cells (Fig. 4D).
Oxytocin also rapidly increased P-S1105, with an EC50 of 11.9 ± 6.9 nM (Fig. 5A, 5B).Pretreatment with the PRKC inhibitor Go6976 (1 μM) completely eliminated oxytocin-inducedphosphorylation (Fig. 5C). Consistent with an effect mediated by PRKC activation, treatmentovernight with 1 μM PMA to desensitize PRKC eliminated the effect of oxytocin on P-S1105
(Fig. 5D). Oxytocin also stimulated P-S1105 in primary uterine smooth muscle cells (Fig. 4D).
In addition to stimulation by oxytocin, P-S1105 was increased by stimulation of other G-proteincoupled receptors. Fig. 6A shows that treatment with PGF2α and ATP significantly increasedP-S1105 in PHM1 cells. The effect of ATP was inhibited >90% by the P2Y receptor antagonistReactive Blue-2 (100 μM) (data not shown), confirming that the effect of ATP was G-proteincoupled receptor-mediated.
CALCA- and Oxytocin-induced PLCB3-S1105 Phosphorylation are Sensitive to DifferentPhosphatase Inhibitors
To determine which type of phosphatase is responsible for reversing CALCA- and OXT-stimulated phosphorylation of PLCB3 (Fig. 6B), the effects of okadaic acid, an inhibitor withhigh affinity for PP1/PP2A phosphatases and cypermethrin, an inhibitor with high affinity forPP2B, were assessed at concentrations used by others to distinguish the effects of PP1/PP2Aand PP2B [23-26]. Since Go6976 pretreatment did not interfere with the net response toCALCA but reduced basal P-S1105 and therefore enhanced detection of stimulated changes inP-S1105, PHM1 cells were pretreated with this inhibitor for 1 h prior to these experiments. Asshown in Fig. 7A, P-S1105 levels in cells treated with CALCA (10 nM), with or without priorexposure to 2 μM okadaic acid, returned to basal levels after 10 min. In contrast, the responseto CALCA was relatively persistent for 15 min in cells pretreated with 4 nM cypermethrin. Onthe other hand, the decrease in oxytocin-stimulated PLCB3-S1105 was attenuated more byokadaic acid than it was by cypermethrin (Fig. 7B). These data are consistent with apredominant influence of PP2B on PRKA-mediated S1105 phosphorylation and a predominantinfluence of PP1/PP2A on oxytocin-mediated phosphorylation of PLCB3-S1105 in PHM1 cells.
Roles of PRKA, PRKC and Protein Phosphatases in the Regulation of Oxytocin-induced PITurnover
PLCB3 has several phosphorylation sites. To address the link between specific phosphorylationof S1105 and oxytocin-stimulated PLC activity, we constructed adenoviruses that expressPLCB3 or the S1105A mutant and infected PHM1 cells. As shown in the inset in Fig. 8A, bothPLCB3 and the S1105A mutant were overexpressed in total cell lysates from infected cells,compared to those infected with empty adenovirus, whereas PRKA catalytic subunit was
Zhong et al. Page 7
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

expressed at a similar concentration in all groups. In all groups, basal PLC activity, as reflectedin PI turnover, was comparable (data not shown). Oxytocin treatment significantly stimulatedPI turnover in cells infected with empty adenovirus and viruses expressing either PLCB3 orthe S1105 A mutant (Fig. 8A). While enhancement of oxytocin-stimulated PI turnover wasoccasionally observed with overexpression of the S1105A mutant, it is not clear why this wasneither consistent nor statistically significant overall. CPT-cAMP (1.5 mM) pretreatmentreduced oxytocin-stimulated PI turnover by approximately 50% in cells infected with eitherempty adenovirus or the adenovirus expressing PLCB3. Importantly, this inhibitory effect wasnot observed in cells overexpressing the S1105A mutant (Fig. 8A). These data suggest that asignificant proportion of the inhibition by PRKA of oxytocin-induced PI turnover in these cellscan be attributed to effects on PLCB3-S1105 (Fig. 6B).
We next attempted to determine if effects on P-S1105 were reflected in influences on oxytocin-stimulated PI turnover in PHM1 cells and assessed effects of phosphatase inhibition on thisparameter. Oxytocin induced a significant increase in PI turnover (Fig. 8B). Pretreatment withthe PRKC inhibitor Go6976 (1 μM) had no effect on basal PI turnover (data not shown), butsignificantly enhanced oxytocin-stimulated PI turnover, as might be expected if negativefeedback from oxytocin-stimulated PRKC activity were inhibited (Fig. 6B). In contrast toeffects on CALCA-stimulated P-S1105, okadaic acid (2 μM) significantly reduced oxytocin-stimulated PI turnover whereas cypermethrin (4 nM) had no effect (Fig.8B). These data areconsistent with the interpretation that negative feedback by PRKC has an impact on theoxytocin-PLC-PI turnover pathway in PHM1 cells and that the reversal of PRKC-mediated P-S1105, primarily by PP1/PP2A, partially attenuates this pathway (Fig. 6B).
DISCUSSIONControl of PLCB3 covalent modification by multiple pathways could play an important rolein the regulation of myometrial calcium dynamics. We demonstrate here that PLCB3 is thepredominant PLCB isoform in the human myometrial cells used and that suppression of thisprotein by shRNA significantly reduces the initial rise in intracellular calcium induced byoxytocin. We show that site-specific phosphorylation of PLCB3-S1105 occurs in intactmyometrial cells in response to diverse hormonal stimuli, and that specific proteinphosphatases are involved in reversing these effects.
cAMP is known to promote relaxation in smooth muscle [15]. We observed increases in P-S1105 in PHM1 cells in response to treatment with CPT-cAMP and with two hormones(CALCA and relaxin) that have receptors in myometrium and that increase cAMP [27-30].The effect of relaxin was partially inhibited by Rp-8-CPT-cAMPS. Taken together, these dataare consistent with an effect of G-protein coupled receptor/Gαs pathways on the PRKA-mediated increase in P-S1105 in PHM1 cells. Inhibition by cAMP of oxytocin-induced PIturnover has been observed in pregnant rat uterus but not at term, coincident with a partial lossof membrane- and AKAP- associated PRKA [16;17;31;32]. PRKA inhibited GTPγS-stimulated PI turnover in rat myometrial plasma membrane, indicating that this effect was distalto possible effects on the oxytocin receptor itself [18]. These data support the presence ofnegative crosstalk from the receptor/Gαs/adenylyl cyclase/PRKA pathway that contributes tothe inhibition of the oxytocin receptor/Gαq/PLCB3 pathway via phosphorylation of PLCB3-S1105(Fig. 6B).
CPT-cAMP pretreatment significantly reduced oxytocin-induced PI turnover in PHM1 cellsinfected with empty adenovirus, as well as in cells overexpressing PLCB3, but had no effecton PI turnover in cells overexpressing the S1105A mutant, consistent with the importance ofS1105 as a target for PRKA. Overexpression of either PLCB3 or the S1105A mutant did notenhance basal or oxytocin-stimulated PI turnover. These data are consistent with the lack of
Zhong et al. Page 8
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

constitutive activity of overexpressed PLCB3 [8] and support the concept that activated Gαqand Gβγ are probably limiting factors in oxytocin-stimulated PI turnover. Overexpression ofPLCB3 and the S1105A mutant should not have any impact on signal pathways upstream ofPLCB coupling and activation, such as the phosphorylation of regulator of G protein signaling4 (RGS4) and adrenergic beta receptor kinase 1 (ADRBK1). also known as G protein-coupledreceptor kinase 2 (GRK2) proteins [33].
In addition to negative signaling pathway crosstalk targeted at PLCB3-S1105, our data provideevidence for negative feedback by downstream components of the PLCB pathway itself inintact cells (Fig. 6B). Oxytocin stimulated phosphorylation of PLCB3-S1105, and this wasinhibited by the PRKC inhibitor Go6976 or by prior treatment with PMA to desensitize PRKC.Importantly, Go6976 pretreatment also enhanced oxytocin-induced PI turnover, which wouldbe expected if negative feedback elicited by PRKC were inhibited. This implies thatdownstream effects of oxytocin on calcium release and entry may be influenced by thephosphorylation status of PLCB3. Interestingly, the PRKC inhibitor Go6976 decreased basalP-S1105. These data suggest that PRKC activity also contributes to basal phosphorylation ofPLCB3-S1105. Basal PLCB3 phosphorylation was noted in 32P labeled cells in previous studiesbut the specific sites were not defined [8;12].
Conventional PRKC isozymes (A, B, G) are activated by both diacylglycerol and calcium,whereas novel PRKC isozymes (D, E, H, Q, M) are activated by diacylglycerol but areinsensitive to calcium, and atypical PRKC isozymes are not regulated by either of these secondmessengers [34]. Both PRKCA and PRKCD have been identified as downstream componentsof oxytocin receptor activation in pregnant human myometrium [35]. Downregulation of thePRKC effect by overnight treatment with PMA is consistent with involvement of conventionalor novel PRKC isozymes. According to data provided by the supplier, PRKCD is not inhibitedby the concentration of Go6976 used in this study, suggesting that the effects noted are mostlikely due to the action of conventional PRKC isozymes.
The dynamics of protein phosphorylation are controlled by both protein kinases and proteinphosphatases (PP), as well as by multiple protein-protein interactions that result in theformation of localized signaling complexes [36;37]. The inhibitory effect of cAMP on PIturnover in myometrial cells requires interaction with an AKAP PRKA anchoring protein.AKAP5 (AKAP79) is expressed in myometrial membranes [17]. PLCB3-S1105 is an in vitrosubstrate for PP2B [16]. PP2B also binds to AKAP5 and has been implicated in local controlof PRKA-stimulated phosphorylations [38]. In the present study, cypermethrin, which exhibitspreferential specificity for PP2B, but not okadaic acid, which shows preferential specificityfor PP1/PP2A, slowed the decay of PRKA-stimulated P-S1105. These data are consistent withthe presence of a functional signaling complex involving AKAP, PRKA and PP2B that exertsan influence on the phosphorylation status of PLCB3-S1105 when cAMP activates the PRKApathway.
In contrast, okadaic acid, but not cypermethrin, slowed the decay of oxytocin (PRKC)-stimulated PLCB3-S1105 phosphorylation, implicating a role for PP1/PP2A indephosphorylation in response to this stimulus. Consistent with these observations, okadaicacid inhibited oxytocin-induced PI turnover whereas cypermethrin had no effect. AlthoughPRKCs have multiple effects on the contractile apparatus as well as on the signaling pathways,the fact that okadaic acid does not affect basal contractile activity but significantly impairs theability of oxytocin to induce mouse myometrial contraction [39] is at least consistent with anegative effect of PP1/PP2A on oxytocin action. PP1 regulates many cell functions, withsubstrate targeting and specificity determined by its interaction with different regulatory units[40;41]. PRKCs can also be targeted by interactions with scaffolding proteins [42;43]. Thedata presented here suggest that PRKC-mediated PLCB3-S1105 phosphorylation may involve
Zhong et al. Page 9
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

the close physical proximity of PP1 or PP2A with one or more of the other signalingcomponents.
Three PLCB isoforms (1, 3, and 4) are expressed in detectable amounts in PHM1 cells(unpublished observations). On the basis of data presented here and previously [14;18], itappears that a significant proportion of oxytocin receptor/Gαq is coupled to PLCB3 in PHM1cells and primary human cells and rat myometrium. PLCB1 is not a substrate of PRKA andmay account for the PRKA-resistant PLC component of the activity elicited by oxytocin.PLCB4 has not been shown to be a target of PRKA or PRKC and lacks a potential targetsequence in the homologous region [11]. Although there is some evidence that receptorscoupling to Gαq can preferentially couple to a given PLCB subtype [44-46], there is no evidenceto date that oxytocin receptor exhibits this ability. The oxytocin receptor can signal both withinand outside membrane subdomains [47;48], whereas PLCB1 and PLCB3 form complexes withscaffolding proteins or other signal components that may restrict their location in cells [49;50].
There are numerous implications of these findings for uterine smooth muscle function. CALCAinhibits human myometrial contractility during pregnancy [51;52]. Because of upregulation ofrelaxant G-protein coupled receptors, the Gs/Gi ratio and adenylyl cyclase isoforms, therelaxant effects of cAMP-mediated pathways predominate during mid-pregnancy [16;53;54].The inhibitory effect of CPT-cAMP on oxytocin-stimulated PI turnover in pregnant ratmyometrium decreases shortly before the initiation of labor, in conjunction with a decrease inPRKA binding to AKAP5 (AKAP150) [16]. The PRKA-dependent phosphorylation ofPLCB3-S1105 could potentially contribute to the dampening by cAMP of both oxytocin- andPGF2α-stimulated and spontaneous contractions in pregnant rat myometrium [55;56]. Whileimportant, this is only one of several effects of PRKA that would favor relaxation [57]. On theother hand, at the end of pregnancy, contractant G-protein coupled receptors and Gq areupregulated, the phosphodiesterases that hydrolyze cAMP are also upregulated, and membranePRKA diminishes, thus reducing the effectiveness of this relaxant pathway [1;54;58;59]. Thedata presented here also indicate that PLCB3-S1105 phosphorylation occurs in response tooxytocin and PGF2α stimulation. Presumably this pathway would become more important nearthe end of pregnancy when contraction-related signaling components are upregulated [1;54;59]. The oxytocin-generated feedback pathway has effects on PI turnover and hence IP3generation. Oxytocin stimulates intracellular calcium oscillations in near-confluent PHM1myometrial cells [60] and fluctuations in IP3 have been implicated in calcium oscillations,along with complex feedback loops regulating calcium pump activity [61-63]. As a result ofdownstream signaling through PLCB, oxytocin also stimulates calcium entry through receptor-operated pathways [57]. Therefore, regulation of oxytocin-stimulated PLC activity by PRKA-and PRKC-mediated pathways may have important implications for the nature of uterinecontractile activity at different stages of pregnancy.
In summary, endogenous PLCB3 is phosphorylated on Serine1105 in intact PHM1 cells by bothPRKA- and PRKC-dependent pathways, representing negative signaling pathway crosstalkand a negative feedback loop, respectively. Since the relative predominance of these twosignaling pathways changes over gestation, their relative influence may be dependent ongestational stage.
ACKNOWLEDGEMENTS
We acknowledge the contribution of J. Jamison (supported by NIH-R25DK067017) in developing the PLCB PCRassay, A. Ulloa for advice on construction of the psiCHECK PLCB3 reporter, and Dr. M. Monga (University of TexasMedical School at Houston) and Dr. R.A. Word (Human Biological Tissues and Fluid Repository, University of TexasSouthwestern Medical Center at Dallas, supported by NIH HD1114) for the human tissue samples. This work wassupported in part by NIH-HD09618.
Zhong et al. Page 10
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

REFERENCES[1]. Sanborn BM. Hormones and calcium: mechanisms controlling uterine smooth muscle contractile
activity. The Litchfield Lecture. Exp. Physiol 2001;86:223–237. [PubMed: 11429639][2]. Lopez-Bernal A. Mechanisms of labour - biochemical aspects. BJOG 2003;110:39–45. [PubMed:
12763110][3]. Challis JR, Lye SJ, Dong XS. Transcriptional regulation of human myometrium and the onset of
labor. J. Soc. Gynecol. Investig 2005;12:65–66.[4]. Rhee S. Regulation of phosphoinositide-specific phospholipase C. Annu Rev Biochem 2001;70:281–
312. [PubMed: 11395409][5]. Nakahara M, Shimozawa M, Nakamura Y, Irino Y, Morita M, Kudo Y, Fukami K. A novel
phospholipase C, PLC(eta)2, is a neuron-specific isozyme. J. Biol. Chem 2005;280:29128–29134.[PubMed: 15899900]
[6]. Litosch I. Regulation of phospholipase C-beta activity by phosphatidic acid: isoform dependence,role of protein kinase C, and G protein subunits. Biochemistry 2003;42:1618–1623. [PubMed:12578375]
[7]. Xu A, Wang Y, Xu LY, Gilmour RS. Protein kinase C alpha -mediated negative feedback regulationis responsible for the termination of insulin-like growth factor I-induced activation of nuclearphospholipase C beta1 in Swiss 3T3 cells. J. Biol. Chem 2001;276:14980–14986. [PubMed:11278470]
[8]. Yue C, Dodge KL, Weber G, Sanborn BM. Phosphorylation of serine 1105 by protein kinase Ainhibits phospholipase Cbeta3 stimulation by Galphaq. J. Biol. Chem 1998;273:18023–18027.[PubMed: 9660757]
[9]. Liu M, Simon MI. Regulation by cAMP-dependent protein kinease of a G-protein-mediatedphospholipase C. Nature 1996;382:83–87. [PubMed: 8657310]
[10]. Yue C, Ku CY, Liu M, Simon MI, Sanborn BM. Molecular mechanism of the inhibition ofphospholipase C beta 3 by protein kinase C. J. Biol. Chem 2000;275:30220–30225. [PubMed:10893237]
[11]. Zhong M, Ku CY, Sanborn BM. Pathways used by relaxin to regulate myometrial phospholipaseC. Ann. N. Y. Acad. Sci 2005;1041:300–304. [PubMed: 15956722]
[12]. Xia C, Bao Z, Yue C, Sanborn BM, Liu M. Phosphorylation and regulation of G-protein-activatedphospholipase C-beta 3 by cGMP-dependent protein kinases. J. Biol. Chem 2001;276:19770–19777. [PubMed: 11278298]
[13]. Arnaudeau S, Lepretre N, Mironneau J. Oxytocin mobilizes calcium from a unique heparin-sensitiveand thapsigargin-sensitive store in single myometrial cells from pregnant rats. Pflugers Arch1994;428:51–59. [PubMed: 7971161]
[14]. Ku CY, Qian A, Wen Y, Anwer K, Sanborn BM. Oxytocin stimulates myometrial guanosinetriphosphatase and phospholipase-C activities via coupling to G alpha q/11. Endocrinology1995;136:1509–1515. [PubMed: 7895660]
[15]. Abdel-Latif AA. Cross talk between cyclic nucleotides and polyphosphoinositide hydrolysis,protein kinases, and contraction in smooth muscle. Exp. Biol. Med. (Maywood. ) 2001;226:153–163. [PubMed: 11361033]
[16]. Dodge KL, Carr DW, Yue C, Sanborn BM. A role for AKAP (A kinase anchoring protein)scaffolding in the loss of a cyclic adenosine 3′,5′-monophosphate inhibitory response in latepregnant rat myometrium. Mol. Endocrinol 1999;13:1977–1987. [PubMed: 10598575]
[17]. Dodge KL, Carr DW, Sanborn BM. Protein kinase A anchoring to the myometrial plasma membraneis required for cyclic adenosine 3′,5′-monophosphate regulation of phosphatidylinositide turnover.Endocrinology 1999;140:5165–5170. [PubMed: 10537145]
[18]. Wen Y, Anwer K, Singh SP, Sanborn BM. Protein kinase-A inhibits phospholipase-C activity andalters protein phosphorylation in rat myometrial plasma membranes. Endocrinology1992;131:1377–1382. [PubMed: 1324160]
[19]. Monga M, Ku CY, Dodge K, Sanborn BM. Oxytocin-stimulated responses in a pregnant humanimmortalized myometrial cell line. Biol. Reprod 1996;55:427–432. [PubMed: 8828850]
Zhong et al. Page 11
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

[20]. Zhong M, Yang M, Sanborn BM. Extracellular signal-regulated kinase 1/2 activation by myometrialoxytocin receptor involves Galpha(q)Gbetagamma and epidermal growth factor receptor tyrosinekinase activation. Endocrinology 2003;144:2947–2956. [PubMed: 12810550]
[21]. Yang M, Wang W, Zhong M, Philippi A, Lichtarge O, Sanborn BM. Lysine 270 in the thirdintracellular domain of the oxytocin receptor is an important determinant for G alpha(q) couplingspecificity. Mol. Endocrinol 2002;16:814–823. [PubMed: 11923477]
[22]. Dodge KL, Sanborn BM. Evidence for inhibition by protein kinase A of receptor/G alpha(q)/phospholipase C (PLC) coupling by a mechanism not involving PLCbeta2. Endocrinology1998;139:2265–2271. [PubMed: 9564832]
[23]. Mukhopadhyay S, Webster C, Anwer M. Role of protein phosphatases in cyclic AMP-mediatedstimulation of hepatic Na+/taurocholate cotransport. J Biol Chem 1998;273:30039–30045.[PubMed: 9792726]
[24]. Webster C, Blanch C, Anwer M. Role of PP2B in cAMP-induced dephosphorylation andtranslocation of NTCP. Am J Physiol Gastrointest Liver Physiol 2002;283:G44–G50. [PubMed:12065290]
[25]. Shin D, Wilkie M, Pamenter M, Buck L. Calcium and protein phosphatase 1/2A attenuate N-methyl-D-aspartate receptor activity in the anoxic turtle cortex. Comp Biochem Physiol A Mol IntegrPhysiol 2005;142:50–57. [PubMed: 16139540]
[26]. Haystead TA, Sim AT, Carling D, Honnor RC, Tsukitani Y, Cohen P, Hardie DG. Effects of thetumour promoter okadaic acid on intracellular protein phosphorylation and metabolism. Nature1989;337:78–81. [PubMed: 2562908]
[27]. Poyner DR, Sexton PM, Marshall I, Smith DM, Quirion R, Born W, Muff R, Fischer JA, FoordSM. International Union of Pharmacology. XXXII. The mammalian calcitonin gene-relatedpeptides, adrenomedullin, amylin, and calcitonin receptors. Pharmacol. Rev 2002;54:233–246.[PubMed: 12037140]
[28]. Hsu CJ, McCormack SM, Sanborn BM. The effect of relaxin on cyclic adenosine 3′,5′-monophosphate concentrations in rat myometrial cells in culture. Endocrinology 1985;116:2029–2035. [PubMed: 2985368]
[29]. Hsu SY, Nakabayashi K, Nishi S, Kumagai J, Kudo M, Sherwood OD, Hsueh AJ. Activation oforphan receptors by the hormone relaxin. Science 2002;295:671–674. [PubMed: 11809971]
[30]. Nguyen BT, Yang L, Sanborn BM, Dessauer CW. Phosphoinositide 3-kinase activity is requiredfor biphasic stimulation of cyclic adenosine 3′,5′-monophosphate by relaxin. Mol. Endocrinol2003;17:1075–1084. [PubMed: 12595573]
[31]. Anwer K, Hovington JA, Sanborn BM. Involvement of protein kinase A in the regulation ofintracellular free calcium and phosphoinositide turnover in rat myometrium. Biol. Reprod1990;43:851–859. [PubMed: 1963319]
[32]. Khac LD, Arnaudeau S, Lepretre N, Mironneau J, Harbon S. Beta adrenergic receptor activationattenuates the generation of inositol phosphates in the pregnant rat myometrium. Correlation withinhibition of Ca++ influx, a cAMP-independent mechanism. J. Pharmacol. Exp. Ther1996;276:130–136. [PubMed: 8558422]
[33]. Huang J, Zhou H, Mahavadi S, Sriwai W, Murthy KS. Inhibition of Galphaq-dependent PLC-beta1activity by PKG and PKA is mediated by phosphorylation of RGS4 and GRK2. Am. J. Physiol CellPhysiol 2007;292:C200–C208. [PubMed: 16885398]
[34]. Violin JD, Zhang J, Tsien RY, Newton AC. A genetically encoded fluorescent reporter revealsoscillatory phosphorylation by protein kinase C. J. Cell Biol 2003;161:899–909. [PubMed:12782683]
[35]. Bermeo ME, Fomin VP, Ventolini G, Gibbs SG, McKenna DS, Hurd WW. Magnesium sulfateinduces translocation of protein kinase C isoenzymes alpha and delta in myometrial cells frompregnant women. Am. J. Obstet. Gynecol 2004;190:522–527. [PubMed: 14981400]
[36]. Pawson T, Nash P. Assembly of cell regulatory systems through protein interaction domains.Science 2003;300:445–452. [PubMed: 12702867]
[37]. Pawson T. Dynamic control of signaling by modular adaptor proteins. Curr. Opin. Cell Biol2007;19:112–116. [PubMed: 17317137]
Zhong et al. Page 12
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

[38]. Hoshi N, Langeberg LK, Scott JD. Distinct enzyme combinations in AKAP signalling complexespermit functional diversity. Nat. Cell Biol 2005;7:1066–1073. [PubMed: 16228013]
[39]. Smith GD, Liu XT, Phillippe M. Divergence in murine myometrium spontaneous and oxytocin-stimulated contractile responses to serine/threonine protein phosphatase-1 inhibition. Biol. Reprod2000;63:781–788. [PubMed: 10952921]
[40]. Cohen PT. Protein phosphatase 1--targeted in many directions. J. Cell Sci 2002;115:241–256.[PubMed: 11839776]
[41]. Ceulemans H, Bollen M. Functional diversity of protein phosphatase-1, a cellular economizer andreset button. Physiol Rev 2004;84:1–39. [PubMed: 14715909]
[42]. Schechtman D, Mochly-Rosen D. Adaptor proteins in protein kinase C-mediated signaltransduction. Oncogene 2001;20:6339–6347. [PubMed: 11607837]
[43]. Osmanagic-Myers S, Wiche G. Plectin-RACK1 (receptor for activated C kinase 1) scaffolding: anovel mechanism to regulate protein kinase C activity. J. Biol. Chem 2004;279:18701–18710.[PubMed: 14966116]
[44]. Strassheim D, Williams CL. P2Y2 purinergic and M3 muscarinic acetylcholine receptors activatedifferent phospholipase C-beta isoforms that are uniquely susceptible to protein kinase C-dependentphosphorylation and inactivation. J. Biol. Chem 2000;275:39767–39772. [PubMed: 10995776]
[45]. Arthur JF, Matkovich SJ, Mitchell CJ, Biden TJ, Woodcock EA. Evidence for selective couplingof alpha 1-adrenergic receptors to phospholipase C-beta 1 in rat neonatal cardiomyocytes. J. Biol.Chem 2001;276:37341–37346. [PubMed: 11489909]
[46]. Oh YS, Jo NW, Choi JW, Kim HS, Seo SW, Kang KO, Hwang JI, Heo K, Kim SH, Kim YH, KimIH, Kim JH, Banno Y, Ryu SH, Suh PG. NHERF2 specifically interacts with LPA2 receptor anddefines the specificity and efficiency of receptor-mediated phospholipase C-beta3 activation. Mol.Cell Biol 2004;24:5069–5079. [PubMed: 15143197]
[47]. Gimpl G, Fahrenholz F. Human oxytocin receptors in cholesterol-rich vs. cholesterol-poormicrodomains of the plasma membrane. Eur J. Biochem 2000;267:2483–2497. [PubMed:10785367]
[48]. Reversi A, Rimoldi V, Brambillasca S, Chini B. Effects of cholesterol manipulation on the signalingof the human oxytocin receptor. Am J Physiol Requl Integr Comp Physiol 2006;291:R861–R869.
[49]. Ranganathan R, Ross EM. PDZ domain proteins: scaffolds for signaling complexes. Curr. Biol1997;7:R770–R773. [PubMed: 9382826]
[50]. Suh PG, Hwang JI, Ryu SH, Donowitz M, Kim JH. The roles of PDZ-containing proteins in PLC-beta-mediated signaling. Biochem. Biophys. Res. Commun 2001;288:1–7. [PubMed: 11594744]
[51]. Casey ML, Smith J, Alsabrook G, MacDonald PC. Activation of adenylyl cyclase in humanmyometrial smooth muscle cells by neuropeptides. J. Clin. Endocrinol. Metab 1997;82:3087–3092.[PubMed: 9284749]
[52]. Dong YL, Fang L, Kondapaka S, Gangula PR, Wimalawansa SJ, Yallampalli C. Involvement ofcalcitonin gene-related peptide in the modulation of human myometrial contractility duringpregnancy. J. Clin. Invest 1999;104:559–565. [PubMed: 10487770]
[53]. Charpigny G, Leroy MJ, Breuiller-Fouche M, Tanfin Z, Mhaouty-Kodja S, Robin P, Leiber D,Cohen-Tannoudji J, Cabrol D, Barberis C, Germain G. A functional genomic study to identifydifferential gene expression in the preterm and term human myometrium. Biol. Reprod2003;68:2289–2296. [PubMed: 12606369]
[54]. Lopez BA. The regulation of uterine relaxation. Semin. Cell Dev. Biol 2007;18:340–347. [PubMed:17582797]
[55]. Vedernikov YP, Syal AS, Okawa T, Jain V, Saade GR, Garfield RE. The role of cyclic nucleotidesin the spontaneous contractility and responsiveness to nitric oxide of the rat uterus at midgestationand term. Am J Obstet Gynecol 2000;182:612–619. [PubMed: 10739517]
[56]. Mhaouty-Kodja S, Houdeau E, Legrand C. Regulation of myometrial phospholipase C system anduterine contraction by beta-adrenergic receptors in midpregnant rat. Biol Reprod 2004;70:570–576.[PubMed: 14585815]
[57]. Sanborn BM, Ku CY, Shlykov S, Babich L. Molecular signaling through G-protein-coupledreceptors and the control of intracellular calcium in myometrium. J. Soc. Gynecol. Investig2005;12:479–487.
Zhong et al. Page 13
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

[58]. Ku CY, Word A, Sanborn BM. Differential expression of protein kinase A, AKAP79 and PP2B inpregnant human myometrial membranes prior to and during labor. J Soc Gynecol Investig2005;12:1509–1515.
[59]. Breuiller-Fouche M, Germain G. Gene and protein expression in the myometrium in pregnancy andlabor. Reproduction 2006;131:837–850. [PubMed: 16672349]
[60]. Burghardt RC, Barhoumi R, Sanborn BM, Andersen J. Oxytocin-induced Ca2+ responses in humanmyometrial cells. Biol. Reprod 1999;60:777–782. [PubMed: 10084948]
[61]. Malysz J, Donnelly G, Huizinga JD. Regulation of slow wave frequency by IP(3)-sensitive calciumrelease in the murine small intestine. Am. J. Physiol Gastrointest. Liver Physiol 2001;280:G439–G448. [PubMed: 11171626]
[62]. Bootman MD, Lipp P, Berridge MJ. The organisation and functions of local Ca(2+) signals. J. CellSci 2001;114:2213–2222. [PubMed: 11493661]
[63]. Young KW, Nash MS, Challiss RA, Nahorski SR. Role of Ca2+ feedback on single cell inositol1,4,5-trisphosphate oscillations mediated by G-protein-coupled receptors. J. Biol. Chem2003;278:20753–20760. [PubMed: 12670945]
Zhong et al. Page 14
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

FIG. 1.Comparison of the expression of PLCB isoforms in human myometrial cells. Relativeexpression of PLC mRNA in PHM1 (A) and UtSMC (B), as analyzed by QRT-PCR. Ct valueswere normalized to HMBS and expressed relative to PLCB3 using the ΔΔCt method (mean ±SEM, n=6 for PLCB1, PLCB2 and PLCB3 and n = 3 for PLCB4 in PHM1 cells; n = 4 for allPLC isoforms in UtSMC). (C) Expression of PLCB1 (open bars) and PLCB3 (black bars)protein in PHM1 and UtSMC cell lysates. Concentrations of PLCB1 and PLCB3 in PHM1cells were 1.6 ± 0.4 and 20.2 ± 7.8 ng/μg cell lysate protein respectively (n = 4) and in UtSMCwere 0.8 ± 0.1 and 17.2 ± 4.8 ng/μg of cell lysate protein, respectively (n=3). Representativestandard curves for UtSMC cell lysates are shown on the right. In (A) - (C). data were analyzed
Zhong et al. Page 15
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

by ANOVA; significant differences (p < 0.05) between groups are designated with differentlowercase letters.
Zhong et al. Page 16
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
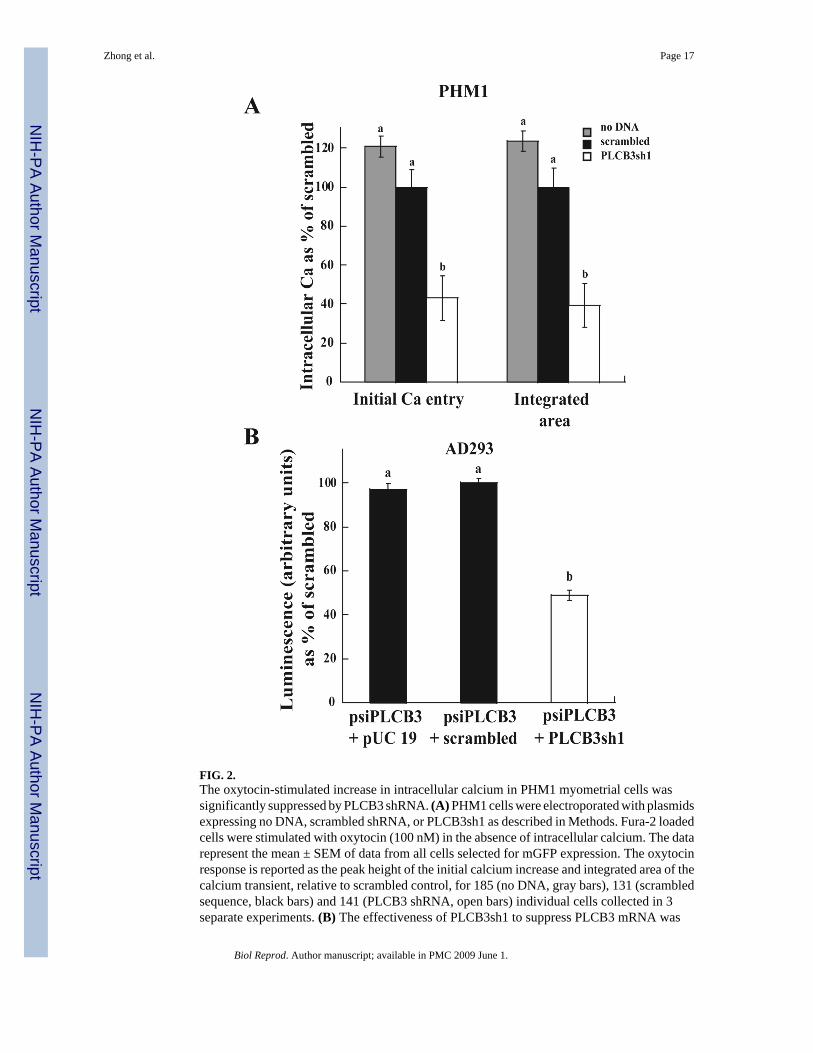
FIG. 2.The oxytocin-stimulated increase in intracellular calcium in PHM1 myometrial cells wassignificantly suppressed by PLCB3 shRNA. (A) PHM1 cells were electroporated with plasmidsexpressing no DNA, scrambled shRNA, or PLCB3sh1 as described in Methods. Fura-2 loadedcells were stimulated with oxytocin (100 nM) in the absence of intracellular calcium. The datarepresent the mean ± SEM of data from all cells selected for mGFP expression. The oxytocinresponse is reported as the peak height of the initial calcium increase and integrated area of thecalcium transient, relative to scrambled control, for 185 (no DNA, gray bars), 131 (scrambledsequence, black bars) and 141 (PLCB3 shRNA, open bars) individual cells collected in 3separate experiments. (B) The effectiveness of PLCB3sh1 to suppress PLCB3 mRNA was
Zhong et al. Page 17
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

assessed in AD293 cells using a PLCB3 psiCHECK-2 reporter luminescence. The reporter (0.5μg) and other plasmids (pUC19 plasmid DNA, plasmid expressing scrambled shRNA sequence(scrambled), or PLCB3sh1, 1.5 μg) were transfected into AD293 cells as described in Methods.The data represent the mean ± SEM for the ratio of luminescence of Renilla luciferase/Fireflyluciferase in 1 experiment (n=3). Data in (A) and (B) were analyzed by ANOVA; significantdifferences (p < 0.05) between groups are designated with different lowercase letters.
Zhong et al. Page 18
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

FIG. 3.PRKA activation by CPT-cAMP increases phosphorylation of endogenous PLCB3-S1105 inPHM1 cells. (A) The specificity of the anti-P-S1105 antibody for phosphorylated PLCB3-S1105. Recombinant PLCB3 and the S1105A mutant proteins were treated with PRKA catalyticsubunit as described in Methods and examined by immunoblot. The anti-P-S1105 antibodydetected increased phosphorylation of PLCB3 but not the S1105A mutant after PRKA treatment.The membrane was reprobed for detection of total PLCB3 (anti-PLCB3) after stripping. (B)Dose-dependence of phosphorylation of crude membrane PLCB3-S1105 following CPT-cAMPtreatment for 10 min (mean ± SEM, n = 4). Data were analyzed by ANOVA. * indicatessignificant difference from solvent control (p < 0.05).
Zhong et al. Page 19
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

FIG. 4.Phosphorylation of endogenous PLCB3-S1105 by hormones that increase endogenous cAMPin PHM1 cells. (A) Sensitivity of relaxin (RLN)-induced P-S1105 to Rp-CPT-cAMPS (1 mM).C= solvent control. Data are expressed as net relaxin-stimulated phosphorylation (n = 3) andwere analyzed by paired Student t-test. Significant difference compared to relaxin treatment(p < 0.05) is indicated by *. (B) Time-course of CALCA (10 nM)-induced P-S1105 and cAMPaccumulation (n=3). (C) Dose-dependence and lack of effect of PRKC inhibition by Go6976(1 μM) on net CALCA-induced P-S1105 (n = 4). C= solvent control. Data from B and C areexpressed as the mean ± SEM and were analyzed by ANOVA. * indicates significant differencefrom control (p < 0.05). (D) Increased P-S1105 in primary uterine smooth muscle cells (UtSMC)
Zhong et al. Page 20
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

in response to 10 nM CALCA and 30 nM oxytocin (OXT). Data represent mean ± SEM oftriplicate determinations in 1 of 4 (CALCA) or 3 (OXT) similar experiments and were analyzedby ANOVA. * indicates significant difference from solvent control (p < 0.05).
Zhong et al. Page 21
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

FIG. 5.Oxytocin receptor activation increases endogenous PLCB3-S1105 phosphorylation by a PRKC-dependent pathway in PHM1 cells. (A) Time-course of P-S1105 following treatment withoxytocin (100 nM) (n = 6). (B) Dose-dependent increase in P-S1105 following oxytocin (OXT)treatment for 5 min (n = 4). Data are expressed as oxytocin-stimulated phosphorylation relativeto solvent control. (C) Sensitivity of oxytocin-induced P-S1105 to Go6976 (1 μM) (n = 3).(D) Reduction in oxytocin-induced P-S1105 after desensitization of PRKC by exposure for 24h to PMA (1 μM) (n = 3). All data are expressed as mean ± SEM and were analyzed by ANOVA.(A, B): * indicates significant difference from the first point in each graph (p < 0.05). (C, D):
Zhong et al. Page 22
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

significant differences between groups (p < 0.05) are designated with different lowercaseletters.
Zhong et al. Page 23
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

FIG. 6.G-protein coupled receptor activation increases endogenous PLCB3-S1105 in PHM-1 cells.Cells were exposed to PGF2α (100 nM), ATP (100 μM) or oxytocin ( OXT, 30 nM) for 5 min.Data represent mean ± SEM of triplicate determinations in 1 experiment. The effects ofPGF2α and ATP were replicated in 1 and 2 other experiments, respectively. Significantdifferences between groups (p < 0.05) are designated with different lowercase letters. (B)Diagrammatic representation of negative crosstalk from the cAMP/PRKA pathway andnegative feedback from the PLC/PRKC pathway focused on PLCB3-S1105 in myometrial cells.According to this scheme, inhibition of PRKA or PRKC would enhance PLCB3 activity,whereas inhibition of protein phosphatases would reduce PLCB3 activity.
Zhong et al. Page 24
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
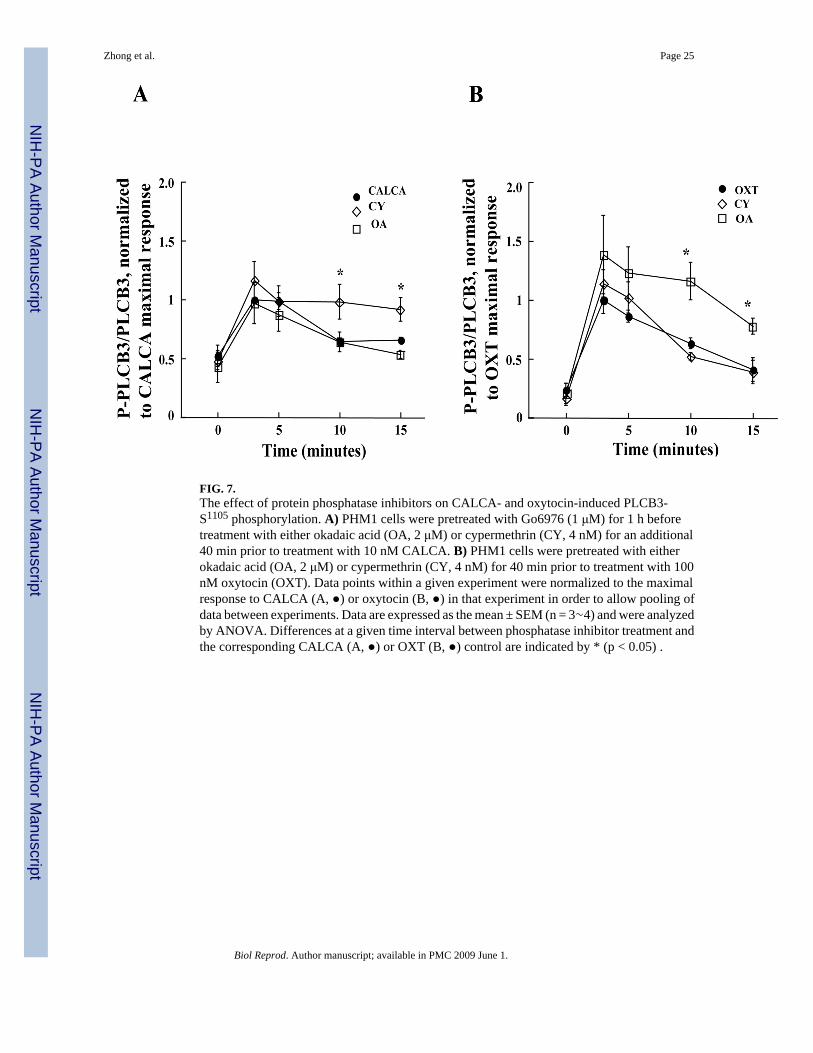
FIG. 7.The effect of protein phosphatase inhibitors on CALCA- and oxytocin-induced PLCB3-S1105 phosphorylation. A) PHM1 cells were pretreated with Go6976 (1 μM) for 1 h beforetreatment with either okadaic acid (OA, 2 μM) or cypermethrin (CY, 4 nM) for an additional40 min prior to treatment with 10 nM CALCA. B) PHM1 cells were pretreated with eitherokadaic acid (OA, 2 μM) or cypermethrin (CY, 4 nM) for 40 min prior to treatment with 100nM oxytocin (OXT). Data points within a given experiment were normalized to the maximalresponse to CALCA (A, ●) or oxytocin (B, ●) in that experiment in order to allow pooling ofdata between experiments. Data are expressed as the mean ± SEM (n = 3∼4) and were analyzedby ANOVA. Differences at a given time interval between phosphatase inhibitor treatment andthe corresponding CALCA (A, ●) or OXT (B, ●) control are indicated by * (p < 0.05) .
Zhong et al. Page 25
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

FIG. 8.Both PRKA-mediated negative crosstalk and PRKC-mediated negative feedback mechanismsregulate oxytocin-induced PI turnover in PHM1 cells. A) Phosphorylation of PLCB3-S1105
contributes to the inhibition by PRKA of net oxytocin-induced PI turnover. PHM1 cells werepretreated with 1.5 mM CPT-cAMP for 20 min before stimulation with 30 nM oxytocin. Theinset demonstrates overexpression of PLCB3 and the S1105A mutant in PHM1 cells, detectedby an antibody that recognizes both forms. PRKA catalytic subunit (PRKA) was analyzed inthe same blot as a loading control. B) Both PRKC and PP1/PP2A influence the amount ofoxytocin-stimulated PI turnover. PHM1 cells labeled with 3H-inositol were exposed to DMSO,Go6976 (1 μM), okadaic acid (OA, 2 μM), or cypermethrin (CY, 4 nM) in the presence of 10mM LiCl for 20 min prior to stimulation with 30 nM oxytocin (OXT) for 30 min and PI turnoverdetermined. Oxytocin-stimulated PI turnover is expressed as the mean ± SEM (n = 3∼4) anddata were analyzed by ANOVA. Significant differences between groups (p < 0.05) aredesignated with different lowercase letters.
Zhong et al. Page 26
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Zhong et al. Page 27
Table 1PCR Primers and shRNA Constructs Used
Gene Bank accession # Gene Symbol Primer or shRNA sequence ( 5′-3′) Nucleotide NumberNM_000190 HMBS Forward: CCAAAGATGAGAGTGATTCGCG
Reverse: GTCCGTCTGTATGCGAGCAAG203-224245-265
NM_015192NM_182734
PLCB1 Forward:AGCTCTCAGAACAAGCCTCCAACAReverse:ATCATCGTCGTCGTCACTTTCCGT
1445-14681528-1551
NM_004573 PLCB2 Forward:AAGGTGAAGGCCTATCTGAGCCAAReverse:CTTGGCAAACTTCCCAAAGCGAGT
335-358500-523
NM_000932 PLCB3 Forward: TATCTTCTTGGACCTGCTGACCGTReverse: TGTGCCCTCATCTGTAGTTGGCTT
1645-16651714-1737
NM_000933NM_182797
PLCB4 Forward: GCACAGCACACAAAGGAATGGTCAReverse:CGCATTTCCTTGCTTTCCCTGTCA
3188-32113343-3366
shRNA target PLCB3 Forward: GCCGTATCCTGGTGAAGAACA 1382-1402Scrambled shRNA PLCB3 Forward: GGAATCTCATTCGATGCATAC
Biol Reprod. Author manuscript; available in PMC 2009 June 1.
Copyright © 2022 FDOKUMEN




















