
Epidural steroids in the management of chronic spinal pain: a systematic review

The role of nitric oxide in the effects of ovarian steroids on
spontaneous myometrial contractility in rats
A. Bulbul a,*, A. Yagcı b, K. Altunbas b, A. Sevimli c, H.A. Celik d,A. Karadeniz e, E. Akdag f
a Department of Physiology, Faculty of Veterinary Medicine, Afyon Kocatepe University, Afyon, Turkeyb Department of Histology & Embryology, Faculty of Veterinary Medicine, Afyon Kocatepe University, Afyon, Turkey
c Department of Pathology, Faculty of Veterinary Medicine, Afyon Kocatepe University, Afyon, Turkeyd Department of Obstetrics and Gynaecology, Faculty of Veterinary Medicine, Afyon Kocatepe University, Afyon, Turkey
e Department of Physiology, Faculty of Veterinary Medicine, Ataturk University, Erzurum, Turkeyf Department of Laboratory Animals, Gulhane Military Medical Academy Research Center, Ankara, Turkey
Received 12 December 2006; received in revised form 26 July 2007; accepted 2 August 2007
www.theriojournal.com
Theriogenology 68 (2007) 1156–1168
Abstract
Forty ovariectomized rats were apportioned into one control and three experimental groups (n = 10 each) to evaluate the role
of nitric oxide in the effects of ovarian steroids on spontaneous myometrial contractility in rats. The control group (group Ov)
received sesame oil once daily for 10 days, whereas rats in the experimental groups were treated with progesterone (2 mg/
(rat day); group P), 17b-estradiol (10 mg/(rat day); group E2), or progesterone and 17b-estradiol together (group E2 + P). The
functionality of the arginine–nitric oxide synthase (NOS)–nitric oxide (NO) pathway in the uterine horns of sacrificed rats was
evaluated in an isolated organ bath. L-Arginine, sodium nitroprusside (SNP) and 8-Br-cGMP decreased uterine contractile
tension induced by electric field stimulation (EFS) in the Ov, P, and E2 + P groups, but not in the E2 group. In addition, L-
arginine was ineffective when applied together with a NOS inhibitor, L-nitro-N-arginine (L-NNA). The percentage of contractile
inhibition was higher in the Ov and P groups compared to the E2 + P group. Immunohistochemical evaluation revealed that
expression of neuronal NOS (nNOS), inducible NOS (iNOS), and endothelial NOS (eNOS) in smooth muscles and nerve cells
did not differ among the groups. Expression of nNOS and eNOS was strongly evident in the E2 and E2 + P groups at both
surface and glandular epithelium of the endometrium. iNOS expression was increased in surface epithelium of the E2 and
E2 + P groups. However, iNOS expression was only increased in glandular epithelial cells of the E2 + P group. In conclusion,
the L-arginine–NOS–NO pathway inhibits myometrial contractions via cGMP-dependent and -independent mechanisms, and
while progesterone maintains the nitric oxide effects, estrogen prevents them. These results suggest that NOS does not mediate
the effects of estrogen.
# 2007 Elsevier Inc. All rights reserved.
Keywords: Nitric oxide; Myometrial contractility; Organ bath; Ovarian steroids; Nitric oxide synthase
* Corresponding author. Tel.: +90 2722281312;
fax: +90 2722281349.
E-mail address: [email protected] (A. Bulbul).
0093-691X/$ – see front matter # 2007 Elsevier Inc. All rights reserved.
doi:10.1016/j.theriogenology.2007.08.014
1. Introduction
Ovarian steroids are one of the most important
factors affecting uterine morphology and motility. The
characteristics of spontaneous myometrial contractility
change due to direct actions of these hormones. Uterine

A. Bulbul et al. / Theriogenology 68 (2007) 1156–1168 1157
function and activity changes depending on the stage of
the estrus cycle or stage of pregnancy [1]. During the
follicular period, when estrogen levels are high, the
amplitude of uterine contraction increases in humans
[2,3]. Moreover, increased contractile tension following
17b-estradiol treatment has been demonstrated in vitro
in ovariectomized rats [4]. On the other hand,
progesterone has been reported to downregulate
prostaglandins, oxytocin receptors, and calcium chan-
nels, thus decreasing the occurrence and tension of
contraction [5,6].
In recent years, nitric oxide has been reported to
mediate the effects of sex hormones on uterine motility
[7]. Endogenous nitric oxide is derived from the amino
acid L-arginine by NOS [8]. Similarly, an exogenous
nitric oxide effect can be derived from nitrate-
containing compounds such as sodium nitroprusside
(SNP) [9]. One function of nitric oxide in reproductive
physiology is to regulate spontaneous uterine con-
tractility [10]. The level of nitric oxide synthesized via
NOS varies with different physiologic and pathologic
circumstances, thereby causing the patterns of sponta-
neous uterine contractility to change [11–13]. It is
known from in vitro studies that nitric oxide decreases
[8] or inhibits [9] uterine contractility depending on the
stages of menstrual period or gestation. Nitric oxide
inhibits the contractility of smooth muscles via
activating guanylate cyclase (GC), which is diffused
into smooth muscle cells [10]. Activated GC increases
cGMP levels, and consequently relaxes smooth
muscles [14].
NOS has inducible (iNOS) and constitutive (cNOS,
which includes nNOS and eNOS) isoforms, which are
present at various levels in different uterine tissues
[11,13]. eNOS levels in uterine vessels of humans and
rats increase during pregnancy [15]. Similarly, eNOS
activity in uterine vessels is higher during the
follicular phase of the estrus cycle than the luteal
phase [16]. Because estrogen increases nitric oxide
levels in genital organs and in circulating blood, it has
been suggested that estrogen could be responsible for
the increase in nitric oxide levels during pregnancy
and the follicular phase of the estrus cycle [15]. It has
been reported that the distribution of NOS and
changes in its activity in the female reproductive
system may play a role in the regulation of smooth
muscle contractility [17]. Therefore, the aim of this
study was to evaluate in vitro the role of nitric oxide in
the effects of ovarian steroids on spontaneous
myometrial contractility, and determine the effects
of ovarian steroids on localization of NOS protein in
vivo in the rat uterus.
2. Materials and methods
2.1. Animals
Female, Sprague–Dawley rats were obtained from
Gulhane Military Medical Academy Research Center,
Department of Laboratory Animals. Rats in all groups
were fed ‘‘ad libitum’’ with the same commercial rat
diet. The ethics committee of Afyonkocatepe Uni-
versity approved all procedures.
The ovariectomy procedure was performed at 12
weeks of age (n = 40). Rats were anesthetized by an
intra-peritoneal ketamine (21.2 mg/kg) and xylazine
(4.2 mg/kg) combination. Small bilateral incisions
were made on the dorsum to expose the ovaries
retroperitoneally. The ovarian vessels were then
clamped and the ovaries removed. Afterwards, the
uterine tubes were ligated and the muscles and skin
were sutured.
Two weeks after operation, the ovariectomized rats
were randomly assigned to four groups of 10 rats each.
Rats in the Ov group received sesame oil once daily for
10 days, whereas rats in the experimental groups were
treated with progesterone 2 mg/(rat day) (group P),
17b-estradiol 10 mg/(rat day) (group E2), and a
combination of progesterone and 17b-estradiol at the
same respective dosages (group E2 + P).
The hormone treatments were continued for 10 days,
after which the rats were killed by cervical dislocation.
Immediately after death, the abdominal cavities of the
rats were opened and the uterine horns were removed.
Each individual horn was placed in Krebs solution
(118 mM NaCl, 4.7 mM KCl, 2.5 mM CaCl2, 1 mM
MgSO4, 1 mM KH2PO4, 25 mM NaHCO3, and 11 mM
glucose). Subsequently, the surrounding mesentery and
fat tissues were carefully removed from the uterus, and
a strip-shaped tissue sample of the uterus with
dimensions of 0.1–0.3 cm � 1.0 cm was obtained.
One edge of each uterine tissue preparation was fixed
to platinum ring electrodes. The opposite edge of the
tissue was connected to a force–displacement transdu-
cer (model 10-A; MAY, Commat, Ankara, Turkey).
Isolated uterine tissues were placed in organ baths filled
with 20 mL Krebs solution, which were continuously
ventilated with gas mixture (95% O2–5% CO2) at 37 8C.
The isometric smooth muscle activity of the uterine
samples were monitored and recorded by computer via
the force transducer and an acquisition system (model
MP30 WSW with Biopac Student Lab PRO Software,
Biopac Systems). An EFS device (model ISO 150-C,
MAY, Commat, Ankara, Turkey) was used for electrical
stimulation.

A. Bulbul et al. / Theriogenology 68 (2007) 1156–11681158
Table 1
Experimental procedures applied to isolated uterine tissues in organ baths
Treatment Treatment procedure Purpose
Incubation Incubated in Krebs solution for 1 h Adaptation
Treatment I Electric stimulation (EFS) Determination of submaximal contraction
Treatment II L-Arginine (10�5 M, 10�4 M, 10�3 M) + EFS Determination of endogenous NO effect
L-NNA (10�3 M) + L-arginine (10�3 M) + EFS Determination of endogenous NO effect on enzymatic pathway
MB (10�5 M) + L-arginine (10�3 M) + EFS Inhibition of GC pathway
Treatment III SNP (10�8 M, 10�7 M, 10�6 M, 10�5 M) + EFS Determination of exogenous NO effect
Treatment IV 8-Br-cGMP (10�8 M, 10�7 M, 10�6 M) + EFS Determination of cGMP effect
2.2. Recording of isometric uterine contractility
Uterus samples in organ baths were kept in Krebs
solution for at least 1 h prior to the recordings to enable
the tissues to adapt to the environment, and the solution
was refreshed in 15-min intervals.Theappropriate resting
tension for the strips was determined in initial experi-
ments. Strips were placed under progressive increments
of tension, and contractile responses to KCl (90 mM in
Krebs solution) were measured under the various resting
tension conditions. Optimal tension relationships were
achieved with resting tensions of 1 g for the uterus strips.
Therefore, a resting tension of 1 g was applied to the
tissues. EFS was then used to elicit neurogenic responses
[18], carried out using single square wave pulses
(durations 0.25, 0.5 and 1 ms) at various frequencies
(2, 4, 8, 32 and 64 Hz). The voltage giving maximal
response was determined individually for each strip.
The experimental procedure applied to the tissue
samples is summarized in Table 1. In order to determine
endogenous nitric oxide activity, L-arginine solution
(1 mM to 1 mM) was added to the Krebs solution. SNP
(0.1 mM to 10 nM) was used for evaluation of the
exogenous nitric oxide pathway. 0.1 mM to 10 nM doses
of 8-Br-cGMP (an analogue for determining the effects of
cGMP) were added in logarithmically increasing
quantities every 10 min. L-Arginine (1 mM) was added
following 100 mM of L-NNA (nitro-N-arginine) treat-
ment in order to determine the effect of endogenous NO
on the enzymatic pathway. To evaluate the effect on the
guanyl cyclase pathway, L-arginine (1 mM) was applied
to the samples after incubation in methylene blue (MB)
for 30 min. All applications were performed on the same
uterus.
2.3. Determination of distribution of NOS in
uterine tissues
Immunohistochemistry for eNOS, iNOS, and nNOS
was performed using Universal LSAB Kits (Zymed
Histostain Plus Broad Spectrum, South San Francisco,
CA, USA) according to the manufacturer’s protocol.
Briefly, paraffin sections (5 mm) were treated with
primary antibodies for eNOS, iNOS, nNOS (Santa Cruz
Biotechnology, Santa Cruz, CA, USA) at 1:300
dilutions for 30 min at 37 8C. Afterwards, paraffin
sections were washed three times in phosphate buffered
saline (PBS) and incubated with biotinylated secondary
antibody for 30 min at room temperature (RT). Samples
were then washed three times in PBS and incubated in
streptavidin-HRP for 30 min at RT. Following rinsing in
PBS for 3 � 5 min, the sections were rinsed in distilled
water and incubated in 3,30-diaminobenzidine (DAB)
(Zymed Laboratories, San Francisco, CA, USA) for
5 min to reveal positive signals. Counterstaining was
carried out with haematoxylin. Sections were cleared in
xylene and mounted with Entellan. Negative controls
were performed using antibody dilutent solution instead
of the primary antibody. Relative immunoreactivity of
antibody staining was evaluated by two independent
observers in a blinded fashion and scored as follows: 0,
no immune reactivity; 1, weak immune reactivity; 2,
moderate immune reactivity; 3, strong immune reac-
tivity; 4, very strong immune reactivity.
2.4. Statistics
All values are presented as mean � S.D. For
statistical evaluation of the data, one-way ANOVA
was used. To compare the individual means of treatment
groups, the Tukey test was performed. Group differ-
ences were declared significant at P < 0.05.
3. Results
3.1. Uterine contractility
Electrically induced uterine smooth muscle con-
tractility of treatment groups in the presence of different
substances at various concentrations [L-arginine

A. Bulbul et al. / Theriogenology 68 (2007) 1156–1168 1159
Fig. 1. Recordings following L-arginine (10�5 M, 10�4 M, 10�3 M), SNP (10�8 M, 10�7 M, 10�6 M, 10�5 M) and 8-Br-cGMP (10�8 M, 10�7 M,
10�6 M) treatments and inhibition of EFS-induced uterine contractions (A: group Ov; B: group P; C: group E2; D: group E2 + P).
Fig. 2. Amplitude values of contractions in groups induced by EFS in
the presence of various concentrations of L-arginine, NNA + L-arginine,
and MB + L-arginine. Letters (a, b, c) indicate significant differences
between them in each column. P-value was <0.001 in groups Ov and
E2 + P, and <0.05 in group P, as determined by ANOVA.
(10�5 M, 10�4 M, 10�3 M), SNP (10�8 M, 10�7 M,
10�6 M, 10�5 M), and cGMP (10�8 M, 10�7 M,
10�6 M)] was recorded. Amplitude values and dose
responses are shown in Fig. 1.
The most effective dose of L-arginine in decreasing
uterus contractility was 10�3 M. Uterine contractile
tension due to L-arginine application significantly
decreased among the groups Ov (P < 0.01), P
(P < 0.05), and E2 + P (P < 0.01). None of the L-
arginine doses were effective at decreasing uterine
contractility in the E2 group. L-NNA (10�3 M) + L-
argininine (10�3 M) treatment decreased contractile
tension in all groups when compared to standard
contractile tension, though this difference was not
significant. Even though L-argininine (10�3 M) treat-
ment decreased the contractile tension, MB incubation
followed by L-argininine (10�3 M) treatment did not
alter the contractile tension in groups Ov, P, and E2 + P
(Fig. 2).
SNP application resulted in relaxation of uterine
contractile tension in the Ov (P < 0.001), P
(P < 0.001), and E2 + P (P < 0.01) groups. However,
none of the SNP doses were effective at decreasing
uterine contractility in the E2 group (Fig. 3). Similarly,
none of the cGMP doses were effective in the E2 group,
whereas cGMP caused a significant relaxation in the Ov,
P, and E2 + P groups (P < 0.05). The most effective
cGMP dose was 10�6 M (Fig. 4).
The percentage inhibition of contractility (when
contraction obtained by electrical field stimulation was
set as 100%) in the treatment groups is presented in
Table 2 and Figs. 5–7. L-Arginine (10�3 M), 8-Br-cGMP
(10�6 M), and all doses of SNP inhibited the contractile
tension of uterine smooth muscle (P < 0.001). Contrac-
tile tension was inhibited to a greater extent in the Ov and
P groups, followed by the E2 + P group. The least
affected treatment group was E2.
L-Arginine (10�3 M) applied to uterine tissue in the
presence of L-NNA did not cause any difference among
the groups. There was no difference between the P and
E2 + P groups following L-arginine treatment after
30 min of MB incubation, while MB inhibited the
contractility more in the Ov group than in the E2 and
E2 + P groups (P < 0.001).
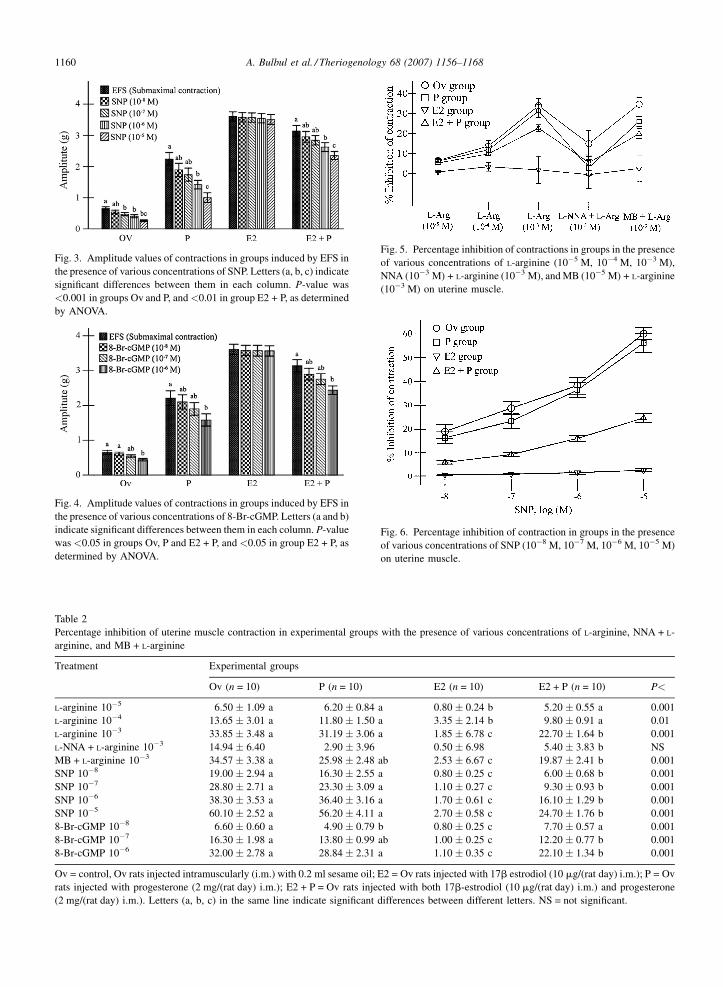
A. Bulbul et al. / Theriogenology 68 (2007) 1156–11681160
Fig. 3. Amplitude values of contractions in groups induced by EFS in
the presence of various concentrations of SNP. Letters (a, b, c) indicate
significant differences between them in each column. P-value was
<0.001 in groups Ov and P, and<0.01 in group E2 + P, as determined
by ANOVA.
Fig. 4. Amplitude values of contractions in groups induced by EFS in
the presence of various concentrations of 8-Br-cGMP. Letters (a and b)
indicate significant differences between them in each column. P-value
was <0.05 in groups Ov, P and E2 + P, and <0.05 in group E2 + P, as
determined by ANOVA.
Table 2
Percentage inhibition of uterine muscle contraction in experimental groups with the presence of various concentrations of L-arginine, NNA + L-
arginine, and MB + L-arginine
Treatment Experimental groups
Ov (n = 10) P (n = 10) E2 (n = 10) E2 + P (n = 10) P<
L-arginine 10�5 6.50 � 1.09 a 6.20 � 0.84 a 0.80 � 0.24 b 5.20 � 0.55 a 0.001
L-arginine 10�4 13.65 � 3.01 a 11.80 � 1.50 a 3.35 � 2.14 b 9.80 � 0.91 a 0.01
L-arginine 10�3 33.85 � 3.48 a 31.19 � 3.06 a 1.85 � 6.78 c 22.70 � 1.64 b 0.001
L-NNA + L-arginine 10�3 14.94 � 6.40 2.90 � 3.96 0.50 � 6.98 5.40 � 3.83 b NS
MB + L-arginine 10�3 34.57 � 3.38 a 25.98 � 2.48 ab 2.53 � 6.67 c 19.87 � 2.41 b 0.001
SNP 10�8 19.00 � 2.94 a 16.30 � 2.55 a 0.80 � 0.25 c 6.00 � 0.68 b 0.001
SNP 10�7 28.80 � 2.71 a 23.30 � 3.09 a 1.10 � 0.27 c 9.30 � 0.93 b 0.001
SNP 10�6 38.30 � 3.53 a 36.40 � 3.16 a 1.70 � 0.61 c 16.10 � 1.29 b 0.001
SNP 10�5 60.10 � 2.52 a 56.20 � 4.11 a 2.70 � 0.58 c 24.70 � 1.76 b 0.001
8-Br-cGMP 10�8 6.60 � 0.60 a 4.90 � 0.79 b 0.80 � 0.25 c 7.70 � 0.57 a 0.001
8-Br-cGMP 10�7 16.30 � 1.98 a 13.80 � 0.99 ab 1.00 � 0.25 c 12.20 � 0.77 b 0.001
8-Br-cGMP 10�6 32.00 � 2.78 a 28.84 � 2.31 a 1.10 � 0.35 c 22.10 � 1.34 b 0.001
Ov = control, Ov rats injected intramuscularly (i.m.) with 0.2 ml sesame oil; E2 = Ov rats injected with 17b estrodiol (10 mg/(rat day) i.m.); P = Ov
rats injected with progesterone (2 mg/(rat day) i.m.); E2 + P = Ov rats injected with both 17b-estrodiol (10 mg/(rat day) i.m.) and progesterone
(2 mg/(rat day) i.m.). Letters (a, b, c) in the same line indicate significant differences between different letters. NS = not significant.
Fig. 5. Percentage inhibition of contractions in groups in the presence
of various concentrations of L-arginine (10�5 M, 10�4 M, 10�3 M),
NNA (10�3 M) + L-arginine (10�3 M), and MB (10�5 M) + L-arginine
(10�3 M) on uterine muscle.
Fig. 6. Percentage inhibition of contraction in groups in the presence
of various concentrations of SNP (10�8 M, 10�7 M, 10�6 M, 10�5 M)
on uterine muscle.
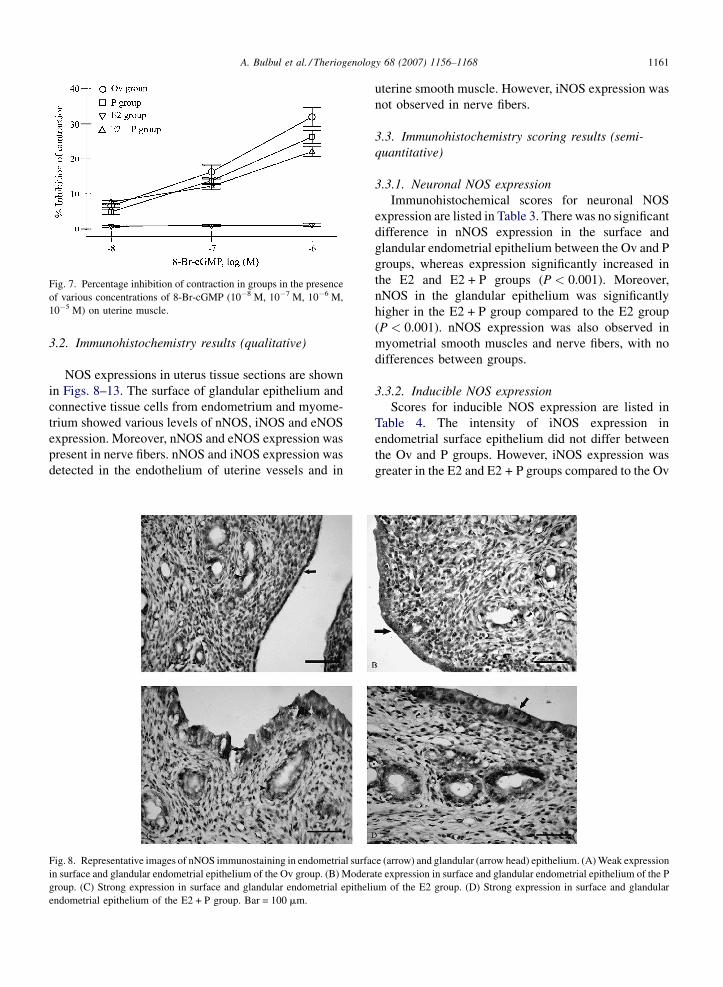
A. Bulbul et al. / Theriogenology 68 (2007) 1156–1168 1161
Fig. 7. Percentage inhibition of contraction in groups in the presence
of various concentrations of 8-Br-cGMP (10�8 M, 10�7 M, 10�6 M,
10�5 M) on uterine muscle.
3.2. Immunohistochemistry results (qualitative)
NOS expressions in uterus tissue sections are shown
in Figs. 8–13. The surface of glandular epithelium and
connective tissue cells from endometrium and myome-
trium showed various levels of nNOS, iNOS and eNOS
expression. Moreover, nNOS and eNOS expression was
present in nerve fibers. nNOS and iNOS expression was
detected in the endothelium of uterine vessels and in
Fig. 8. Representative images of nNOS immunostaining in endometrial surfa
in surface and glandular endometrial epithelium of the Ov group. (B) Modera
group. (C) Strong expression in surface and glandular endometrial epitheli
endometrial epithelium of the E2 + P group. Bar = 100 mm.
uterine smooth muscle. However, iNOS expression was
not observed in nerve fibers.
3.3. Immunohistochemistry scoring results (semi-
quantitative)
3.3.1. Neuronal NOS expression
Immunohistochemical scores for neuronal NOS
expression are listed in Table 3. There was no significant
difference in nNOS expression in the surface and
glandular endometrial epithelium between the Ov and P
groups, whereas expression significantly increased in
the E2 and E2 + P groups (P < 0.001). Moreover,
nNOS in the glandular epithelium was significantly
higher in the E2 + P group compared to the E2 group
(P < 0.001). nNOS expression was also observed in
myometrial smooth muscles and nerve fibers, with no
differences between groups.
3.3.2. Inducible NOS expression
Scores for inducible NOS expression are listed in
Table 4. The intensity of iNOS expression in
endometrial surface epithelium did not differ between
the Ov and P groups. However, iNOS expression was
greater in the E2 and E2 + P groups compared to the Ov
ce (arrow) and glandular (arrow head) epithelium. (A) Weak expression
te expression in surface and glandular endometrial epithelium of the P
um of the E2 group. (D) Strong expression in surface and glandular
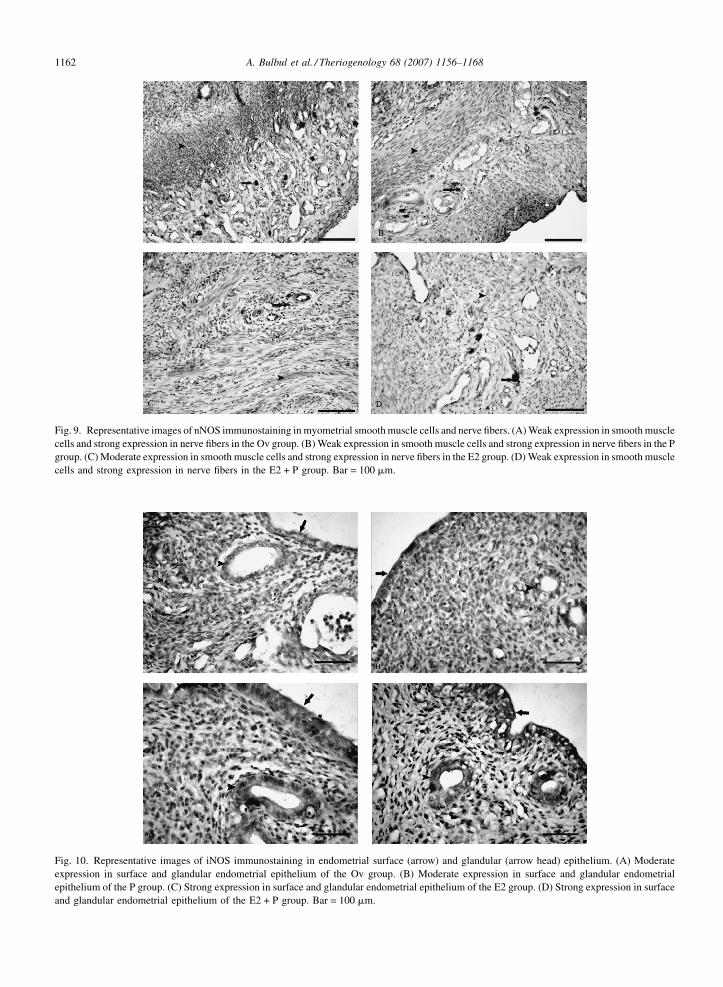
A. Bulbul et al. / Theriogenology 68 (2007) 1156–11681162
Fig. 9. Representative images of nNOS immunostaining in myometrial smooth muscle cells and nerve fibers. (A) Weak expression in smooth muscle
cells and strong expression in nerve fibers in the Ov group. (B) Weak expression in smooth muscle cells and strong expression in nerve fibers in the P
group. (C) Moderate expression in smooth muscle cells and strong expression in nerve fibers in the E2 group. (D) Weak expression in smooth muscle
cells and strong expression in nerve fibers in the E2 + P group. Bar = 100 mm.
Fig. 10. Representative images of iNOS immunostaining in endometrial surface (arrow) and glandular (arrow head) epithelium. (A) Moderate
expression in surface and glandular endometrial epithelium of the Ov group. (B) Moderate expression in surface and glandular endometrial
epithelium of the P group. (C) Strong expression in surface and glandular endometrial epithelium of the E2 group. (D) Strong expression in surface
and glandular endometrial epithelium of the E2 + P group. Bar = 100 mm.

A. Bulbul et al. / Theriogenology 68 (2007) 1156–1168 1163
Fig. 11. Representative images of iNOS immunostaining in myometrial smooth muscle cells. (A) Moderate expression in myometrial smooth
muscle cells of the Ov group. (B) Moderate expression in myometrial smooth muscle cells of the P group. (C) Moderate expression in myometrial
smooth muscle cells of the E2 group. (D) Moderate expression in myometrial smooth muscle cells of the E2 + P group. Bar = 100 mm.
Fig. 12. Representative images of eNOS immunostaining in endometrial surface (arrow) and glandular (arrow head) epithelium. (A) Moderate
expression in surface and glandular endometrial epithelium of the Ov group. (B) Moderate expression in surface and glandular endometrial
epithelium of the P group. (C) Strong expression in surface and glandular endometrial epithelium of the E2 group. (D) Strong expression in surface
and glandular endometrial epithelium of the E2 + P group. Bar = 100 mm.
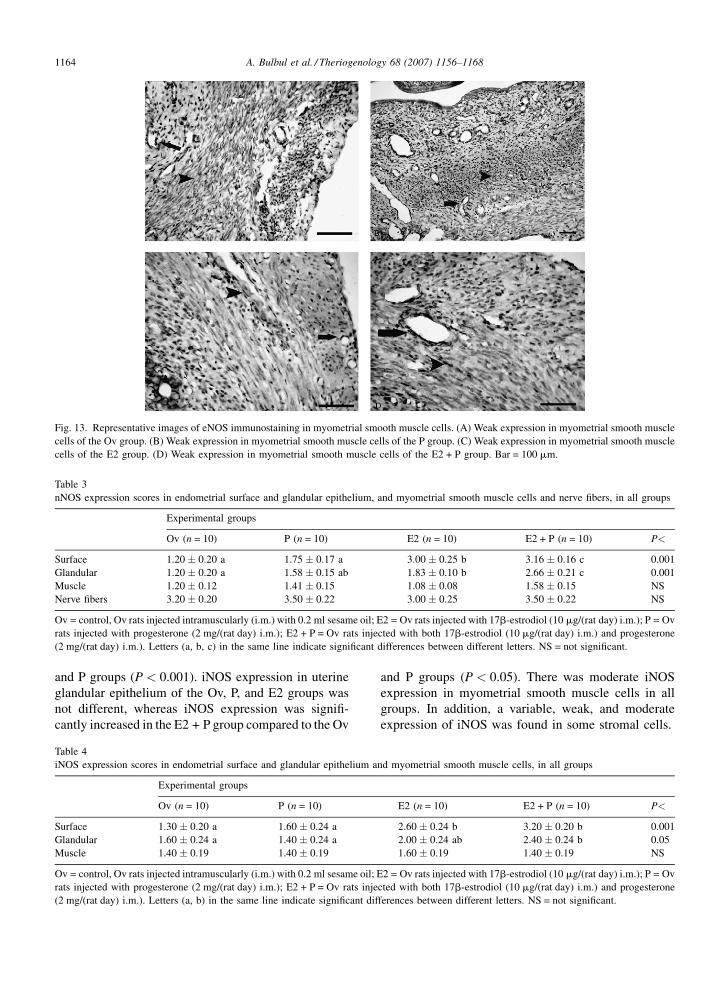
A. Bulbul et al. / Theriogenology 68 (2007) 1156–11681164
Fig. 13. Representative images of eNOS immunostaining in myometrial smooth muscle cells. (A) Weak expression in myometrial smooth muscle
cells of the Ov group. (B) Weak expression in myometrial smooth muscle cells of the P group. (C) Weak expression in myometrial smooth muscle
cells of the E2 group. (D) Weak expression in myometrial smooth muscle cells of the E2 + P group. Bar = 100 mm.
Table 3
nNOS expression scores in endometrial surface and glandular epithelium, and myometrial smooth muscle cells and nerve fibers, in all groups
Experimental groups
Ov (n = 10) P (n = 10) E2 (n = 10) E2 + P (n = 10) P<
Surface 1.20 � 0.20 a 1.75 � 0.17 a 3.00 � 0.25 b 3.16 � 0.16 c 0.001
Glandular 1.20 � 0.20 a 1.58 � 0.15 ab 1.83 � 0.10 b 2.66 � 0.21 c 0.001
Muscle 1.20 � 0.12 1.41 � 0.15 1.08 � 0.08 1.58 � 0.15 NS
Nerve fibers 3.20 � 0.20 3.50 � 0.22 3.00 � 0.25 3.50 � 0.22 NS
Ov = control, Ov rats injected intramuscularly (i.m.) with 0.2 ml sesame oil; E2 = Ov rats injected with 17b-estrodiol (10 mg/(rat day) i.m.); P = Ov
rats injected with progesterone (2 mg/(rat day) i.m.); E2 + P = Ov rats injected with both 17b-estrodiol (10 mg/(rat day) i.m.) and progesterone
(2 mg/(rat day) i.m.). Letters (a, b, c) in the same line indicate significant differences between different letters. NS = not significant.
and P groups (P < 0.001). iNOS expression in uterine
glandular epithelium of the Ov, P, and E2 groups was
not different, whereas iNOS expression was signifi-
cantly increased in the E2 + P group compared to the Ov
Table 4
iNOS expression scores in endometrial surface and glandular epithelium a
Experimental groups
Ov (n = 10) P (n = 10)
Surface 1.30 � 0.20 a 1.60 � 0.24 a
Glandular 1.60 � 0.24 a 1.40 � 0.24 a
Muscle 1.40 � 0.19 1.40 � 0.19
Ov = control, Ov rats injected intramuscularly (i.m.) with 0.2 ml sesame oil;
rats injected with progesterone (2 mg/(rat day) i.m.); E2 + P = Ov rats inje
(2 mg/(rat day) i.m.). Letters (a, b) in the same line indicate significant dif
and P groups (P < 0.05). There was moderate iNOS
expression in myometrial smooth muscle cells in all
groups. In addition, a variable, weak, and moderate
expression of iNOS was found in some stromal cells.
nd myometrial smooth muscle cells, in all groups
E2 (n = 10) E2 + P (n = 10) P<
2.60 � 0.24 b 3.20 � 0.20 b 0.001
2.00 � 0.24 ab 2.40 � 0.24 b 0.05
1.60 � 0.19 1.40 � 0.19 NS
E2 = Ov rats injected with 17b-estrodiol (10 mg/(rat day) i.m.); P = Ov
cted with both 17b-estrodiol (10 mg/(rat day) i.m.) and progesterone
ferences between different letters. NS = not significant.

A. Bulbul et al. / Theriogenology 68 (2007) 1156–1168 1165
Table 5
eNOS expression scores in endometrial surface and glandular epithelium, and myometrial smooth muscle cells and nerve fibers, in all groups
Experimental groups
Ov (n = 10) P (n = 10) E2 (n = 10) E2 + P (n = 10) P<
Surface 1.24 � 0.19 a 2.60 � 0.25 b 3.60 � 0.25 c 3.80 � 0.20 c 0.001
Glandular 1.36 � 0.37 a 2.60 � 0.25 b 3.40 � 0.25 c 3.80 � 0.20 c 0.001
Muscle 1.00 � 0.00 1.00 � 0.00 1.20 � 0.20 1.00 � 0.00 NS
Nerve fibers 3.00 � 0.00 3.00 � 0.00 2.900 � 0.10 3.00 � 0.00 NS
Vessel 0.40 � 0.25 0.60 � 0.25 0.60 � 0.25 0.80 � 0.20 NS
Ov = control, Ov rats injected intramuscularly (i.m.) with 0.2 ml sesame oil; E2 = Ov rats injected with 17b-estrodiol (10 mg/(rat day) i.m.); P = Ov
rats injected with progesterone (2 mg/(rat day) i.m.); E2 + P = Ov rats injected with both 17b-estrodiol (10 mg/(rat day) i.m.) and progesterone
(2 mg/(rat day) i.m.). Letters (a, b, c) in the same line indicate significant differences between different letters. NS = not significant.
3.3.3. Endothelial NOS expression
Immunohistochemical scores for endothelial NOS
expression are listed in Table 5. The intensity of eNOS
expression in endometrial surface and glandular
epithelium was greater in the E2 and E2 + P groups.
Moreover, eNOS expression was significantly higher in
group P than group Ov (P < 0.001). There was weak
expression in smooth muscle cells and vessel endothe-
lium layers in the myometrium in all groups.
4. Discussion
In rat serum, the physiological concentration of L-
arginine ranges from 0.2 to 3 mM [19], and it has been
observed that L-arginine completely inhibits in vitro
smooth muscle contractility at doses of 0.01–1 mM [9].
Moreover, L-argininine has been reported to decrease
myometrial contractile tension in pregnant humans and
rats [8] and in non-pregnant human myometrium in a
dose-dependent manner [20]. The relaxation effect of L-
arginine in the uterus decreases during labor, is further
diminished by postpartum day 1, and is completely
absent by postpartum day 2 [9]. Similarly, L-arginine
decreased uterine contractile tension in the Ov, P, and
E2 + P groups in the current study. However, none of the
L-arginine doses were effective in decreasing uterine
contractile tension in the E2 group. In addition, the
percentage inhibition of uterine contractile tension was
higher in the Ov and P groups than the E2 group with the
most efficient dose of L-arginine (10�3 M). Thus, the
results of the current study suggest that the inhibition of
uterine contractility by L-arginine was prevented by
estrogen, and that progesterone does not alter L-
arginine-modulated contractile tension.
In the present study, the effect of L-arginine on the
contractile tension of uterine smooth muscle at its most
effective dose (10�3 M) disappeared when it was
applied together with L-NNA. It has been previously
reported that the dose-dependent inhibition of smooth
muscle contractility by L-arginine is linked to nitric
oxide formation by NOS [8,9,20]. As L-NNA is a NOS
inhibitor, the above result of the current study suggests
that the relaxation of uterine smooth muscles caused by
L-arginine is mediated by nitric oxide.
SNP is an exogenous nitric oxide source, which
completely blocks spontaneous uterine contractility in
pregnant rats [9] and in non-pregnant humans [20].
Similarly, SNP completely removes spontaneous
myometrial contractility during pregnancy, but it is
not effective together with KCl [8]. Word and Cornwell
[21] have stated that SNP doses lower than 10�4 M are
not effective at inhibiting oxytocin-stimulated uterus
contractility, and higher doses cause irreversible
inhibition due to toxicity. In the current study, SNP
treatment at doses of 10�8 to 10�5 M inhibited
contractile tension in all groups except for the E2
group, with decreased inhibition in the E2 + P group.
Hence, it may be suggested that estrogen (but not
progesterone) prevents SNP-mediated uterine relaxa-
tion by SNP.
Nitric oxide conjugates and frees cyclic GC (cGC),
and hence increases cGMP levels and consequently
decreases uterine contractile tension [22]. Buhimschi
et al. [10] reported that the NO-cGMP pathway is
responsible for quiescence of the uterus in pregnant
women. Yallampali et al. [23] concluded that 17b-
estradiol and 17b-estradiol + progesterone treatment
decreases uterine cGMP and NO levels when compared
to control and progesterone-treated groups, and that
there is a positive correlation between NO formation
and cGMP levels. Krumenacker et al. [24] concluded
that 17b-estradiol inhibits soluble GC mRNA release in
uterine tissue. However, cGMP analogues do not inhibit
uterine contractility in non-pregnant monkey [25],
human [10,20] or mice [26]. Furthermore, nitric oxide-
driven inhibition of uterine contractility occurs inde-
pendently from the GC-cGMP pathway [10,25,26]. In
the current study, the most effective dose of cGMP was

A. Bulbul et al. / Theriogenology 68 (2007) 1156–11681166
10�6 M, and cGMP treatment of uterine tissues caused
relaxation in the Ov, P, and E2 + P groups. However, no
dose of cGMP was effective in the E2 group. This
suggests that progesterone had no effects on the cGMP
pathway. Estrogen, however, completely inhibited the
efficacy of cGMP-induced relaxation in uterine smooth
muscles.
It has been reported that MB inhibits cGC and
increases the contractile tension of smooth muscles
[27,28], while relaxation of spontaneous or induced
contractions mediated by the nitric oxide donor S-
nitroso L-cysteine (cys-NO) [29] and SNP [27,28] is
unaffected by prior addition of MB. In our study, we
showed that L-arginine-induced uterine contractility in
the Ov, P, and E2 + P groups decreased in presence of
MB. This is consistent with previous studies [20,29],
which suggested that NO-induced relaxation of the
myometrium occurs via a different pathway from the
cGC-cGMP pathway. It has also been suggested that
NO and/or cGMP may cause relaxation by directly
activating K+ channels in smooth muscles [30,31].
Modzelewska et al. [32] showed that diethylamine/NO
causes concentration-dependent decreases in amplitude
and frequency of myometrial contraction in non-
pregnant women, but that blockers of large-conduc-
tance and voltage-dependent Ca(2+)-activated K(+)
channels, including iberiotoxin and charybdotoxin,
reduce the diethylamine-nitric oxide-induced inhibition
of spontaneous activity of the myometrium.
NOS can moderate the motility effects of estrogen
and progesterone in the uterus. Progesterone regulates
iNOS expression in pregnancy, while estrogen receptors
regulate cNOS expression in the menstrual cycle [33].
In the current study, there was an intense nNOS
expression in nerve fibers between smooth muscle
layers of myometrium. Nevertheless, differences in
nNOS expression in nerve fibers in the uterus was not
significant among the groups. Similarly, Papka et al.
[34] reported nNOS expression in nerve fibers in the rat
uterus and suggested that nitric oxide-synthesizing
nerves in the rat myometrium might have a role in
controlling uterine vascularization. In addition, nNOS
expression in nerve fibers in smooth muscle layers has
been reported in the rabbit uterus, and that steroid
hormones had no effect on nNOS expression in nerve
fibers [35]. The results of the current study show that
there is weak nNOS expression in surface and glandular
epithelial cells and smooth muscle cells of the rat uterus,
except in myometrial nerve fibers of Ov rats. 17b-
Estradiol and particularly 17b-estradiol + progesterone
increased nNOS expression in the surface and glandular
epithelium of rat uteri. Similarly, Ogando et al. [36]
reported that there was no nNOS expression by western
blot analysis in uterine extracts of Ov rats, while nNOS
expression was observed in animals treated with
estrogen or progesterone.
In the present study, E2, P, and E2 + P treatment of
Ov rats significantly increased the intensity of eNOS
expression in surface and glandular epithelial cells,
while no significant difference was observed in smooth
muscle cells, nerve fibers, or endothelial cells. Some
studies have reported that estrogen [37,38] or proges-
terone [36,39] solely increase the intensity of eNOS
expression in uterine endometrial cells and smooth
muscle. In the current study, separate or combined
treatment of steroid hormones increased eNOS expres-
sion in surface and glandular epithelial cells, but only
weak expression was observed in all treatment groups.
Furthermore, estrogen has been reported to increase
eNOS expression in smooth muscle cells [40,41], and
Gangula et al. [42] reported that progesterone increases
in vitro eNOS in myometrial smooth muscle cells.
Contrary to the previous reports, our results suggest that
steroid hormones do not alter eNOS expression in
smooth muscle cells.
Huang et al. [43] reported weak iNOS expression
only in endometrial surface epithelial cells and mast
cells of Ov mice, and that progesterone treatment
increased iNOS expression in surface epithelial cells.
Yallampalli and Dong [41] have suggested that estradiol
reduce iNOS mRNA and protein levels in uteri of
pregnant and LPS-treated rats. However, in the current
study, there was a weak iNOS expression in endometrial
surface and glandular epithelium and myometrial
smooth muscle cells of rats. 17b-Estradiol increased
iNOS expression in surface epithelial cells of endome-
trium, and 17b-estradiol + progesterone treatment
increased iNOS expression in both surface epithelial
cells and glandular epithelial cells. Similarly, Saxena
et al. [13] reported that estrogen induces iNOS
expression, with greater expression when estrogen
and progesterone are applied together. Ogando et al.
[36] concluded that estrogen increases iNOS protein
expression in uteri of Ov rats. In addition, Huang et al.
[43] reported that undeterminable iNOS expression in
epithelial cells of Ov rats following estrogen treatment
becomes detectable after estrogen + progesterone treat-
ment.
In conclusion, the L-arginine–NOS–NO pathway
inhibits myometrial contractions via cGMP-dependent
and -independent mechanisms, and while progesterone
maintains the effects of nitric oxide, estrogen prevents
them. Furthermore, estrogen increases the expression of
nNOS, iNOS and eNOS enzymes in uterine epithelium

A. Bulbul et al. / Theriogenology 68 (2007) 1156–1168 1167
and glands. However, estrogen did not affect the
expression of the same enzymes in smooth muscles or
nerve cells and had no effect on uterine motility.
Acknowledgments
Financial assistance for this research was provided
by the Scientific Research Council of Turkey (project
numbered VHAG-2097). We are grateful to Dr Victor
M. S for his contributions.
References
[1] Gard PR. Drug action on the gut, bladder and uterus. In: Taylor,
Francis, editors. Human pharmacology. CRP Press LLC; 2001.
p. 107–10.
[2] Weiss G. Endocrinology of parturition. J Clin Endocrinol Metab
2000;12:4421–5.
[3] Oike K, Ishihara K, Kikuchi S. A study on the endometrial
movement and serum hormonal level in connection with uterine
contraction. Nippon Sanka Fujinka Gakkai Zasshi 1990;42:86–92.
[4] Vedernikov YP, Hartke JR, Long MA, Saade GR, Garfield RE.
Sex hormone effects in non-pregnant rat and human myome-
trium. Eur J Obstet Gynecol Reprod Biol 2003;108:59–66.
[5] Thorburn GD, Challis JR. Endocrine control of parturition.
Physiol Rev 1979;59:863–918.
[6] Abel MH, Baird DT. The effect of 17 beta-estradiol and pro-
gesterone on prostaglandin production by human endometrium
maintained in organ culture. Endocrinology 1980;106:
1599–606.
[7] Al Hijji J, Larsson I, Batra S. Effect of ovarian steroids on nitric
oxide synthase in the rat uterus, cervix and vagina. Life Sci
2001;69:1133–42.
[8] Izumi H, Yallampalli C, Garfield RE. Gestational changes in L-
arginine-induced relaxation of pregnant rat and human myome-
trial smooth muscle. Am J Obstet Gynecol 1993;169:1327–37.
[9] Yallampalli C, Garfield RE, Byam-Smith M. Nitric oxide inhi-
bits uterine contractility during pregnancy but not during deliv-
ery. Endocrinology 1993;133:1899–902.
[10] Buhimschi I, Yallampalli C, Dong YL, Garfield RE. Involvement
of a nitric oxide-cyclic guanosine monophosphate pathway in
control of human uterine contractility during pregnancy. Am J
Obstet Gynecol 1995;172:1577–84.
[11] Rosselli M, Keller PJ, Dubey RK. Role of nitric oxide in the
biology, physiology and pathophysiology of reproduction. Hum
Reprod 1998;4:3–24.
[12] Jablonka-Shariff A, Ravi S, Beltsos AN, Murphy LL, Olson LM.
Abnormal estrous cyclicity after disruption of endothelial and
inducible nitric oxide synthase in mice. Biol Reprod 1999;61:
171–7.
[13] Saxena D, Purohit SB, Kumar PG, Laloraya M. Increased
appearance of inducible nitric oxide synthase in the uterus
and embryo at implantation. Nitric Oxide 2000;4:384–91.
[14] Anggard EE. Endogenous and exogenous nitrates. Acta Anaes-
thesiol Scand Suppl 1992;97:7–10.
[15] Nelson SH, Steinsland OS, Wang Y, Yallampalli C, Dong YL,
Sanchez JM. Increased nitric oxide synthase activity and expres-
sion in the human uterine artery during pregnancy. Circ Res
2000;87:406–11.
[16] Taguchi M, Alfer J, Chwalisz K, Beier HM, Classen-Linke I.
Endothelial nitric oxide synthase is differently expressed in
human endometrial vessels during the menstrual cycle. Mol
Hum Reprod 2000;6:185–90.
[17] Dong YL, Gangula PR, Fang L, Yallampalli C. Nitric oxide
reverses prostaglandin-induced inhibition in ovarian progester-
one secretion in rats. Mol Hum Reprod 1999;14(1):27–32.
[18] Kim NN, Min K, Pessina MA, Munarriz R, Goldstein I, Traish
AM. Effects of ovariectomy and steroid hormones on vaginal
smooth muscle contractility. Int J Impot Res 2004;16:43–50.
[19] Langrehr JM, Dull KE, Ochoa JB, Billiar TR, Ildstad ST, Schraut
WH, Simmons RL, Hoffman RA. Evidence that nitric oxide
production by in vivo allosensitized cells inhibit the development
of allospecific CTL. Transplantation 1992;53:632–40.
[20] Hoffmann P, Stanke-Labesque F, Fanchin R, Dilai N, Pons JC,
Ayoubi JM. Effects of L-arginine and sodium nitroprusside on the
spontaneous contractility of human non-pregnant uterus. Hum
Reprod 2003;18:148–51.
[21] Word RA, Cornwell TL. Regulation of cGMP-induced relaxa-
tion and cGMP-dependent protein kinase in rat myometrium
during pregnancy. Am J Physiol 1998;274:C748–56.
[22] Francis SH, Corbin JD. Cyclic nucleotide-dependent protein
kinases: intracellular receptors for cAMP and cGMP action.
Crit Rev Clin Lab Sci 1999;36:275–328.
[23] Yallampali C, Izumi H, Byam-Smith M, Garfield RE. An L-
arginine-nitric oxide-cyclic guanosine monophosphate system
exists in the uterus and inhibits contractility during pregnancy.
Am J Obstet Gynecol 1994;170:175–85.
[24] Krumenacker JS, Hyder SM, Murad F. Estradiol rapidly inhibits
soluble guanylyl cyclase expression in rat uterus. Proc Natl Acad
Sci USA 2001;98:717–22.
[25] Kuenzli KA, Buxton IL, Bradley ME. Nitric oxide regulation of
monkey myometrial contractility. Br J Pharmacol 1998;124:63–8.
[26] Kuenzli KA, Bradley ME, Buxton IL. Cyclic GMP-independent
effects of nitric oxide on guinea-pig uterine contractility. Br J
Pharmacol 1996;119:737–43.
[27] Mayer B, Brunner F, Schmidt K. Inhibition of nitric oxide synth-
esis by methylene blue. Biochem Pharmacol 1993;45:367–74.
[28] Mayer B, Brunner F, Schmidt K. Novel actions of methylene
blue. Eur Heart J 1993;14:22–6.
[29] Buxton IL, Kaiser RA, Malmquist NA, Tichenor S. NO-induced
relaxation of labouring and non-labouring human myometrium
is not mediated by cyclic GMP. Br J Pharmacol 2001;134:
206–14.
[30] Carrier GO, Fuchs LC, Winecoff AP, Giulumian AD, White RE.
Nitrovasodilators relax mesenteric microvessels by cGMP-
induced stimulation of Ca-activated K channels. Am J Physiol
1997;273:H76–84.
[31] Koh SD, Dick GM, Sanders KM. Small-conductance Ca(2+)-
dependent K+ channels activated by ATP in murine colonic
smooth muscle. Am J Physiol 1997;273:C2010–21.
[32] Modzelewska B, Kostrzewska A, Sipowicz M, Kleszczewski T,
Batra S. Apamin inhibits NO-induced relaxation of the sponta-
neous contractile activity of the myometrium from non-pregnant
women. Reprod Biol Endocrinol 2003;1–8.
[33] Salhab WA, Shaul PW, Cox BE, Rosenfeld CR. Regulation of
types I and III NOS in ovine uterine arteries by daily and acute
estrogen exposure. Am J Physiol Heart Circ Physiol 2000;278:
H2134–42.
[34] Papka RE, McNeill DL, Thompson D, Schmidt HH. Nitric oxide
nerves in uterus are parasympathetic sensory and contain neu-
ropeptides. Cell Tissue Res 1995;279:339–49.

A. Bulbul et al. / Theriogenology 68 (2007) 1156–11681168
[35] Al-Hijji J, Larsson B, Batra S. Nitric oxide synthase in the rabbit
uterus and vagina: hormonal regulation and functional signifi-
cance. Biol Reprod 2000;62:1387–92.
[36] Ogando D, Farina M, Ribeiro ML, Perez Martinez S, Cella M,
Rettori V, Franchi A. Steroids hormones augment nitric oxide
synthase activity and expression in rat uterus. Reprod Fertil Dev
2003;15:269–74.
[37] Vagnoni KE, Shaw CE, Phernetton TM, Meglin BM, Bird IM,
Magness RR. Endothelial vasodilator production by uterine and
systemic arteries. III. Ovarian and estrogen effects on NO
synthase. Am J Physiol 1998;275:H1845–56.
[38] Yallampalli C, Dong YL, Gangula PR, Fong L. Role and
regulation of nitric oxide in the uterus during pregnancy and
parturition. J Soc Gynecol Investig 1998;5:58–67.
[39] Farina M, Ribeiro ML, Franchi A. Nitric oxide synthase in
pregnant rat uterus. Reproduction 2001;121:403–7.
[40] Figueroa JP, Massmann GA. Estrogen increases nitric oxide
synthase activity in the uterus of nonpregnant sheep. Am J
Obstet Gynecol 1995;173:1539–45.
[41] Yallampalli C, Dong YL. Estradiol-17beta inhibits nitric oxide
synthase (NOS)-II and stimulates NOS-III gene expression in the
rat uterus. Biol Reprod 2000;63:34–41.
[42] Gangula PR, Dong YL, Yallampalli C. Rat myometrial smooth
muscle cells express nitric oxide synthase. Hum Reprod
1997;12:561–8.
[43] Huang J, Roby KF, Pace JL, Russell SW, Hunt JS. Cellular
localization and hormonal regulation of inducible nitric oxide
synthase in cycling mouse uterus. J Leukoc Biol 1995;57:27–35.
Copyright © 2022 FDOKUMEN




















