
Multiple Signals Regulate Phospholipase CBeta3 in Human Myometrial Cells

Molecular Human Reproduction vol.5 no.8 pp. 757–766, 1999
Localization of regulatory protein binding sites in the proximalregion of human myometrial connexin 43 gene
Clement O.Echetebu, Mariam Ali, Michael G.Izban, Lynnete MacKay and Robert E. Garfield1
Department of Obstetrics and Gynecology, The University of Texas Medical Branch, Division of Reproductive Sciences,Galveston, TX 77555, USA1To whom correspondence should be addressed
Parturition is preceded by a large increase in gap junctions between myometrial smooth muscle cells.
Connexin 43 is the major structural protein of myometrial gap junctions. To explore transcriptional regulation
of the myometrial Cx43 gene, we used DNase I footprinting, electrophoretic mobility shift and transient
transfection assays to examine a 312 bp promoter region (–164 to 1148) of the gene, utilizing human
myometrial cell cultures and nuclear extracts. The DNase I studies showed four regions of nucleoprotein
interactions. Protection of region 1 (–80 to –31) encompassed an Activator Protein 1 (AP1) (–44 to –36) and
two Specificity Protein 1 (Sp1) (–77 to –69 and –59 to –48) consensus sequences. Regions 2 to 4 included the
transcription initiation site (–10 to 125), an Ets/NF-kB consensus sequence (147 to 174) and a TA-rich region
(181 to 1101) respectively. Gel mobility shift and supershift assays demonstrated c-Jun and Sp1 binding at
the AP1 and Sp1 sites respectively. Promoter mutagenesis and transient transfection analyses combined with
Sp1 and c-Jun/c-Fos over-expression studies indicate that both Sp1 and c-Jun are required for maximal
promoter activity and, therefore, may positively regulate transcription of myometrial Cx43 during the initiation
of labour.
Key words: AP1/connexin-43/myometrium/promoter/Sp1
Introduction
Gap junctions are specialized channels that connect adjacentcells and enable metabolic and electrical co-ordinationbetween them (Pitts and Simms, 1977; Bennet andGoodenough, 1978). The major structural protein of gapjunctions is connexin. There are many different kinds ofconnexins encoded by a multigene family dispersed on numer-ous chromosomes (Fishmanet al., 1991). Connexin 43 (Cx43)is one of the most widely expressed members of this genefamily, and is found in numerous tissues such as the uterus,heart, lens epithelium, kidney, mammary gland, ovary, placentaand intestine (Beyeret al., 1987; Risek and Gilula, 1991; Leeet al., 1992; Li et al., 1993). The onset of labour is precededby a dramatic increase of gap junctions in uterine smoothmuscle, the myometrium (Garfieldet al., 1977; Miller et al.,1989; Hendrixet al., 1992; Chow and Lye, 1994). This increaseis believed to enable the development of co-ordinated andintense uterine contractions necessary for the delivery of thefetus, whereas its suppression before term is believed to aiduterine quiescence. Evidence to support this concept comesfrom many functional studies showing that propagation ofelectrical activity of the myometrium is increased significantlyduring labour when gap junctions/Cx43 protein concentrationsare increased (Garfieldet al., 1977, 1988; Simset al., 1982;Cole et al., 1985; Miller et al., 1989; Miyoshiet al., 1996).
Cx43mRNA and protein concentrations in the myometriumare low during pregnancy, but rise markedly at term, remainhigh throughout labour, then decline rapidly following
© European Society of Human Reproduction and Embryology 757
delivery in rat (Riseket al., 1990; Hendrixet al., 1992;Tabbet al., 1992; Lyeet al., 1993; Winterhageret al., 1993),sheep (McNuttet al., 1994) and human (Garfield and Hayashi,1981). While the expression ofCx43 is up-regulated byoestrogens (Garfieldet al., 1977; MacKenzie and Garfield,1985) and down-regulated by progesterone (Garfieldet al.,1977; MacKenzie and Garfield, 1985; Petrocelli and Lye,1993; Zhaoet al., 1996) the molecular events that regulatetranscriptional activation remain unclear.
Rat (Yuet al., 1994), mouse (Chenet al., 1995), and human(Geimonenet al., 1996) Cx43 promoters have been clonedand sequenced. In these promoters, extensive homology existsbetween –175 to1143, relative to the start site of transcription(Chen et al., 1995). Transient expression studies (DeLeonet al., 1994; Chenet al., 1995; Geimonenet al., 1996)indicated that this region is important for regulatingCx43promoter activity. It is likely, therefore, that similar regulatoryelements may be operational within this promoter region inthese species.
In the mouse, genetic and biochemical studies have demon-strated both ‘repressor’ (–100 to –91) and ‘activator’(–68 to –59) promoter elements (Chenet al., 1995). Theseregions are.90% conserved in the humanCx43 promoter.Transient transfection experiments in human cardiacmyocytes have revealed the presence of negative regulatoryelement(s) between –2400 and –360 and a positive elementbetween –175 and –50 ofCx43 gene (DeLeonet al., 1994).Transcription of the humanCx43 gene can be induced inprimary human myometrial cell cultures after activation of

C.O.Echetebu et al.
protein kinase C by tetradecanoyl phorbol acetate (TPA)treatment (Geimonenet al., 1996). HumanCx43 promoterregion from –697 to1170 has been used in mutation analysisand transient expression studies to demonstrate that TPA-induced activation required a consensus Activator Protein 1(AP1) site located between positions –44 to –36. The AP1 siteis conserved between rodent and human promoters but distinctfrom the mouse ‘activator’ region (Chenet al., 1995). Inaddition, these studies demonstrated that TPA induced adramatic rise in the protein concentrations of both c-Junand c-Fos, further supporting the notion that transcriptionalactivation occurred through the promoter proximal AP1 site.However, over-expression of c-Jun and or c-Fos in the absenceof TPA treatment had no effect on promoter activity, suggestingthat TPA functioned by increasing and activating c-Jun/c-Fos.
In this report, we used a combination of DNase I footprinting,electrophoresis mobility shift (EMSA) and transient transfec-tion assays to begin a systematic study of the humanCx43promoter. We confirm previous studies (Geimonenet al., 1996)and extend them by showing that the Ap1 site functionallyinteracts with c-Jun. In addition, we demonstrate that theCx43promoter contains tandem Specificity Protein 1 (Sp1) bindingsites and that the proximal site, which is encompassed withina promoter segment homologous to the mouse ‘activator’element functionally activates transcription. Furthermore, weshow that over-expression of c-Jun and Sp1 either alone orin combination up-regulatesCx43 transcription in untreatedprimary cells in culture. Thus, both Sp1 and c-Jun are capableof and necessary for positively regulating human myometrialCx43 transcription.
Materials and methods
Cell culture
Approval for the use of human tissue was obtained from the Universityof Texas Medical Branch Committee on Research Involving Humansubjects. To obtain human myometrial factors that bind theCx43promoter at term when the levels ofCx43mRNA and gap junctionsare high, nuclear extracts should be made from myometrium obtainedfrom biopsies or Caesarean section at these times. The quantity oftissue obtained from biopsies is insufficient for making amounts ofnuclear proteins needed for our experiments. Moreover, it is difficultto homogenize human myometrium to liberate intact nuclei withoutfirst freezing the tissue or using harsh disintegration procedures thatalso break the nuclei and freezing is not recommended for preparingfactors active in transcription (M.G.Izban, personal communication).Crude nuclear extracts were prepared from primary cultures of humanmyometrial cells initiated and grown as follows: the tissue fromCaesarian sections or biopsies (from patients, gestational age, 37–39weeks and not in labour) was cleaned of blood vessels, fat andendometrium. It was washed several times in wash buffer, Hank’sbalanced salt solution (HBSS; Gibco Life Technologies, Grand Island,NY, USA), supplemented with 100 IU/ml penicillin G; 100µg/mlstreptomycin sulphate and 25µg/ml amphotericin B as fungizone(Gibco); then minced and rinsed three times in the same buffer. Itwas then digested for ~75 min at 37°C with three times its volumeof dispersion solution (HBSS, containing 1 mmol/l Ca21, 1 mmol/lMg21, 25 mmol/l HEPES, pH 7.4; Gibco), antibiotics and fungizoneas above; 300 IU/ml Collagenase Type IV (Worthington, Freehold,NJ, USA) and 22.5 IU/ml DNase I (Worthington). The dispersion
758
process included trituration every 3 min with decreasing bore-sizepipettes, collection of supernatants by centrifugation (20g for 5 min) atroom temperature every 10 min and addition of fresh dispersionsolution. The pooled supernatants were centrifuged at 430g for 10min. The separated cells were washed twice with the wash bufferand once with the growth medium (Minimal Essential Medium;Gibco) supplemented with 10% (v/v) fetal calf serum, 2 mmol/lL-glutamine, 1 mmol/l sodium pyruvate; antibiotics and fungizone asabove. They were then resuspended in adequate volume of growthmedium, plated and grown at 37°C in a 5% CO2/95% humidified airincubator. Medium was changed every 3 days (Andersenet al., 1993;Zhaoet al., 1996).
Preparation of nuclear extractTo obtain sufficient quantity of nuclear proteins, the cultures werepassaged four times. A total of 50 culture dishes (150315 mm) ofcells (~33109 cells) were used per experiment. After change of media(Geimonenet al., 1996), the cells were treated for 6–8 h with phorbolester, 12-O-tetradecanoyl-phorbol-13-acetate (TPA), (Calbiochem-Novobiochem Corporation, La Jolla, CA, USA) at 100 ng/ml,harvested and nuclear extract made by a modified method (Shapiroet al., 1988) using a hand-held Dounce homogenizer. The modifica-tion included the addition of protease inhibitors: leupeptin andaprotinin, at 2µg/ml; pepstatin-A (1µg/ml), and phosphatase inhibitor(β-glycerophosphate) (10 mmol/l) to all buffers except the dialysisbuffer which contained onlyβ-glycerophosphate as an addition.Extracts were divided into small volume aliquots, flash-frozen inliquid nitrogen and stored at –70°C till used.
Plasmid constructs
A 312 bp fragment (–164 to1148) was amplified using the polymerasechain reaction (PCR) from a 6 kbEcoRI humanCx43gene 59-regionthat was cloned into pGF10 and kindly supplied by Dr G.I.Fishman(Fishmanet al., 1991). The PCR primers containedKpnI and XbaIsites, which were used to insert the 312 bp fragment into a pUC19polycloning site to create pCx312. The primers were:Sense: 59-GCGCGGTACCGATCCCACTGCTCTTTG-39Antisense: 59-GCGCTCTAGACCTTGGAGGATGAAGTAAAATG-39.
A 171 bp fragment (–164 to17), was excised from the 312 bpfragment and inserted intoKpnI/HindIII sites of pUC19 and ishereafter referred to as pCx171. The sequences of the fragments wereconfirmed by the dideoxy chain termination method.
In-vitro mutagenesis
Seven independent 10 bp replacement mutants (seriatim linker scanspanning –99 to –30 of the promoter) were made using PCR frompCx312 (Valletteet al., 1989; Vanden Heuvelet al., 1993). A 10 bpsequence 59-ATCTCGAGAT-39 bearing a uniqueXhoI site was usedfor each replacement. The mutants∆–99/–90,∆–89/–80,∆–79/–70,∆–69/–60,∆–59/–50,∆–49/–40,∆–39/–30 were inserted into pGL3basic vector in theKpnI/BglII cloning site. Specific confirmatory 2or 3 bp replacement mutants∆–93/–91 [CCC→TTT; ∆–74/–72,(CTC→TGT); ∆–55/–53, (CTC→TGT); and∆ –59/–58 (GC→TT)]were also constructed as additional checks for the∆–99/–90,∆–79/–70 and∆–59/–50 mutations. Briefly, the 312 bp fragment was excisedfrom pCx312 and inserted into PGL3 basic vector in theKpnI/BglIIcloning site and mutagenesis was performed directly in this vectorusing transformer site-directed mutagenesis kit (Clontech Labs Inc,Palo Alto, CA, USA) and the following primers:
for ∆–93/–91: 59-CAAAGGGAAAGGCTAAAAGGAGGTAG-GGAG-39,
∆–74/–72: 59-GCTAGAAGAAAGGGACAGGCAAAGGGAAA-GGC-39;

AP1 and Sp1 regulate human myometrial Cx43 gene expression
∆–59/–58: 59-CAACTGGGAGGAGGGAATAGAAGAAAGGGG-AGG-39,
∆–55/–53: 59-CTGACTCAACTGGGAGACAGGGCTAGAAGA-AAGGGG-39,and a selection primer, 59-GCCGGCCGCCCCGACTCGCGAATTA-CACGGCGATC-39 were used to changeXbaI (TCTAGA) to NruI(TCGCGA ) site. Sequences were confirmed by the ABI PRISMAutomatic Sequencer Model 373, Version 3.0.
Labelling of DNA fragments:
The 312 and 171 bp fragments used in DNaseI protection assayswere 39-end labelled atEcoRI, XbaI or HindIII sites of the plasmidspCx312 and pCx171 with [α-32P]dNTPs (specific activity, 3000 Ci/mmol; Amersham Life Science, Arlington Heights, IL, USA) and theKlenow fragment of DNA Polymerase I (Promega, Madison, WI,USA). High specific activity was achieved by employing doublelabels [α-32P]dATP and [α-32P]dTTP forEcoRI andHindIII 39-endsand [α-32P]dCTP and [α-32P]dTTP forXbaI 39-end. Labelled insertswere separated from the vector by restriction with appropriate enzymesand purified by 5% non-denaturing polyacrylamide gel electrophoresis(PAGE), ion exchange chromatography (NACS column; Gibco LifeTechnologies) and ethanol precipitation. Double-stranded oligo-nucleotides used in EMSA were 59-end labelled with [γ-32P]ATP(Amersham) and T4 Polynucleotide kinase, then purified by gelfiltration (Pharmacia, Piscataway, NJ, USA) using G-50 spin columnsor PAGE. They include oligonucleotides for AP1, AP2 and Sp1,purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA)and custom oligonucleotides made by Genosys Biotechnologies (TheWoodlands, TX, USA).
DNase I footprinting
DNase I footprint assay (Angelet al., 1987) was performed in a25 µl reaction mixture containing (final concentration); 10 mmol/lHEPES (pH 8.0), 1.6 mmol/l dithiothreitol (DTT), 5 mmol/l MgCl,0.1 mmol/l EDTA, 25 mmol/l KCl, 1µg [poly (dI-dC). poly (dI-dC)], (Pharmacia), 12% glycerol, 5µg bovine serum albumin (BSA),nuclear proteins (0.0 to 60µg) and 13105 cpm (0.1 pmoles) oflabelled DNA. After incubation on ice for 60 min, samples wereequilibrated for 3 min at room temperature and digested for 1 min.with 0.5–6 µl of DNase I (Worthington) diluted to 6.25µg/ml inbuffer (10 mmol/l HEPES, pH 7.6, and 25 mmol/l CaCl2). Thereaction was terminated with 200µl of stop buffer (10 mmol/l Tris–HCl, pH 7.6, 20 mmol/l EDTA, 3µg/ml sheared calf thymus DNA,0.2 mg/ml proteinase K and 0.2% sodium dodecyl sulphate), thenincubated for 45 min at 37°C. After phenol–chloroform extraction,the DNA was ethanol-precipitated, resuspended in formamide dyemixture (95% formamide, 0.05% Bromophenol Blue and 0.05%Xylene Cyanol FF) and fractionated on 8% polyacrylamide sequencinggel along with a G1A chemical sequencing reaction lane (Maxamand Gilbert, 1980) and visualized by autoradiography.
Electrophoresis motility shift assay (EMSA)
Standard EMSA were performed with [γ-32P]ATP-labelled double-stranded oligonucleotides. Binding reactions (15µl) contained10 mM Tris–HCl (pH 7.6), 50 mmol/l KCl, 5 mmol/l DTT, 0.5 mmol/l EDTA, 1 mmol/l MgCl2 and varying amounts of nuclear extractand poly (dI-dC) as indicated in the figure legends. The mixture wasincubated on ice for 10 min, then 5.0–7.53104 cpm of label wasadded and the incubation was continued for 25 min at 25°C. Sampleswere then fractionated on 4% non-denaturing polyacrylamide gelwith 0.53 Tris/Borate/EDTA buffer. Protein–DNA bands were visual-ized by autoradiography. For supershift and competition assays,antibodies to specific proteins (Santa Cruz Biotech Santa Cruz, CA,
759
USA; and Oncogene Research Products, Cambridge, MA, USA) orunlabelled mutant or non-specific oligonucleotides were added to theincubation mixtures. Incubations involving Sp1 antibody containedadditional 1.6 mmol/l ZnSO4. Double-stranded oligonucleotides used(showing only the coding strand sequences) contain either AP1(double underline), AP2 consensus (bold-faced), ‘CCTCC’ elements(single underline) or Ets/NF-kB consensus (dotted underline) areas follows:
AP1 (–52/–29): 59-CTCCCAGTTGAGTCAGTGGCTTGA-39.CCTCC/ proximal AP2: (–100/–80): 59-ACCTCCTCCCAG-CCTTTCCCT-39.AP2 distal (–139/–119):59-TCTCCCCCCAGCCCTTCCACA-39.–70/–48: 59-CCTTTCTTCTAGCCCCTCCTCCCA-39–100/–50: 59-ACCTCCTCCCAGCCTTTCCCTTTGCCCTTT-CTTCTAGCCCCTCCTTCCCCC-39–83/–65: 59-CCTTTGCCCTCCCCTTTC-39.150/173: 59-GGCGTGAGGAAAGTACCAAACAGC-39. (Ets/reversed NF-kB motif)–32/–9: 59-TTGAAACTTTTAAAAGCTCTGTGC-39bA-4: 59-TCTTCGCATCCCTCCTCCAGT-39, motif found inchicken adultβ-globin gene (Emersonet al., 1985).
Transient transfections
Human myometrial primary culture cells between second and fourthpassages were plated to ~50–60% confluence in 60 mm diameterculture dishes. They were transfected with 20µg total DNA (15.0µgof test plasmids, 1.0µg of internal control pCMVlacZ plasmid and4.0µg of Salmon sperm DNA) by the calcium phosphate procedure. At4–6 h post-transfection, the cells were glycerol-shocked for 1 min,washed with phosphate-buffered saline (PBS) and supplemented withfresh medium. Cells were harvested 48 h later, lysed and assayed forluciferase using the Promega Luciferase Assay System (Promega).β-Galactosidase activity was measured by the standard colorimetricmethod and used for normalizing luciferase activity.
Co-transfection with c-Jun, c-Fos and Sp1 expression
plasmids
The following plasmids were used: pCMV Sp1, which over-expressesSp1, was generously provided by R.Tjian; pCMV c-Jun and pCMVc-Fos, which over-express c-Jun and c-Fos respectively, were giftsfrom A.Kopland. Wild-typeCx43 (5 µg; 312 bp) in pGL3 basic-lucreporter was mixed with either 1µg of each expression plasmid ortheir combinations containing 1µg of each plasmid (see legend toFigure 7 for details) and 1µg of pCMVlacZ. The total DNA wasadjusted to 20µg with salmon sperm DNA. Transfections wereperformed as described above.
Statistical analysis
The transfection assays were performed four times. Samples in eachgroup of experiments were in duplicates or triplicates. All data wereexpressed as mean6 SE. One-factor analysis of variance (ANOVA)was used to test the overall hypothesis of no group differences,followed by two-samplet-tests for pairwise comparisons where theoverall hypothesis was rejected.P , 0.05 was considered to bestatistically significant.
Results
Transcription factor binding sites within the humanCx43 promoter
It has been demonstrated that TPA treatment of primarymyometrial cells in culture leads to stimulation ofCx43

C.O.Echetebu et al.
Figure 1. (A) DNase I footprint of 312 bp promoter region of humanCx43gene. DNase I protection analysis of the coding strand oflabelled DNA probe (–164 to1148). Amounts of human myometrial crude nuclear extract proteins used are indicated at the tops of lanes(3, 4 and 5). Protected regions one through four are indicated on the right of the autoradiograph and sequence positions relative totranscription start site are on the left and right margins. G1A (lane 1) indicates Maxam & Gilbert chemical sequence ladder. Free DNA(lane 2) contains no proteins and shows natural sensitivity to DNase I. (B) DNaseI footprint ofCx43gene region (–164 to17). The 171 bpSau3AI/HindIII Cx43promoter (region –164 to17), labelled at theHindIII ends was used as a probe. Amounts of extract proteins used areat the tops of lanes. Protected regions are indicated by lines along the right side of the autoradiograph. Positions relative to transcriptionstart site are shown on the left side. The positions of Activator Protein (AP)1, Sp1 and AP2 consensus sequences are indicated. Free DNA(lanes 1 and 3) contain no proteins. Lane 2 shows chemical sequence ladder.
promoter (Geimonenet al., 1996). To define the regions withinthe promoter of the humanCx43 gene capable of interactingwith transcription factors in similarly treated cells we carriedout DNase I footprinting analysis. The results using a promoterfragment bearing (relative to the start site of transcription)from –164 to1148 (Figure 1A) and from –164 to17 (Figure1B) demonstrated several protected regions separated by sitesof enhanced DNase I sensitivity. Protection on the coding(Figure 1) and non-coding (data not shown) strands is indicatedin Figure 2. Region 1 extends from –80 to –31 and encompassesa consensus AP1 site (–44 to –36) and a segment (–80to –45) that envelops other potential protein binding sitessummarized in Figure 2. The promoter sequence (–65 to –48)homologous to the mouse ‘activator’ element (Chenet al.,1995) is also located in this region. Three other regionsconsidered further only in the discussion include the site ofinitiation, Region 2 (–5 to115), Region 3 (147 to 170)which is coincident with an Ets/NFkB consensus site andRegion 4 (180 to 1 100) that spans a TA-rich site (181 to
760
196). Note that transcription factor binding was not observedeither within the CCTCC/AP2 sequence (–93 to –87) whichcorresponds to the mouse ‘repressor’ site (Chenet al., 1995),or at the AP2 consensus (–134 to –127).
c-Jun binds to the AP1 consensus site of the Cx43
promoter
Previous studies (Geimonenet al., 1996) have suggested thatCx43 promoter activation through the proximal AP1 sitefollowing TPA treatment of primary myometrial cells relieson the concomitant increase in c-Jun and c-Fos protein concen-trations. To test this directly, we performed mobility shift andsupershift assays using a double-stranded oligonucleotide thatencompassed the AP1 consensus site (–52 to –29). Incubationof the radiolabelledCx43 AP1 motif with nuclear extractsderived from TPA-treated primary myometrial cell cultures(Figure 3 A,B, lane 2) or, as a control, from actively growingHeLa cell spinner cultures (Figure 3B, lane 6) resulted in asingle shifted band. Formation of this nucleoprotein complex

AP1 and Sp1 regulate human myometrial Cx43 gene expression
Figure 2. Summary of human Cx43 promoter DNaseI footprint in the presence of human myometrial nuclear extract proteins. Lines aboveand below the sequences, marked off by numbers, indicate regions 1–4, protected from DNase1 digestion. The start site of transcription is at11. Brackets indicatecis-elements and arrow above Ets shows direction of the incomplete Ets element.
was competed by inclusion of non-labelled self (Figure 3A,lane 8) or by a commercially available oligonucleotide con-taining the consensus AP1 binding motif (Figure 3A, lane 9;Figure 3B, lane 10). Competition was not observed with amutated AP1 binding motif that does not support AP1 binding(Figure 3B, lane 11). These data support the notion that thisregion within theCx43promoter is an AP1-binding site.
AP1 is a family of dimeric transcription factors comprisedof either c-Jun homodimers or heterodimers of certain Jun/Fos family members. In order to identity the Jun/Fos familymembers within the band-shifted nucleoprotein complex,we initially performed supershift analysis using a monoclonalc-Jun antibody specific for phosphorylated c-Jun (c-JunP,Figure 3A, lane 4,), and polyclonal antibodies raised againstthe aminoterminal regions of either the c-Jun (c-Jun, Figure3A, lane 3) or c-Fos (c-Fos, Figure 3A, lane 5) proteins. InFigure 3A, a supershift for phosphorylated c-Jun was observed(lanes 4 and 7). However, supershifts were not observed withc-Jun (lanes 3 and 6) or c-Fos (lanes 5 and 6) either alone(lanes 3 and 5) or in combination (lane 6). It was surprisingthat c-Jun antibody had no effect on band mobility since thisantibody should have recognized both the phosphorylated andnon- phosphorylated forms of c-Jun. Therefore, we repeatedthese experiments using a polyclonal antibody (pan Jun) thatrecognizes a highly conserved region of the DNA bindingdomain of Jun family members c-Jun, Jun B and Jun D. Asexpected, this antibody disrupted Jun binding when eithermyometrial (Figure 3B, lane 3) or HeLa cells (Figure 3B,lane 7) were used as a protein source. The supershift assayperformed with the c-JunP antibody demonstrated that phos-phorylated c-Jun was present within the myometrial-derivedcomplex (Figure 3B, lane 5) but not within the nucleoproteincomplex formed using the HeLa cell nuclear extract (Figure3B, lane 9). This latter result was not entirely unexpectedsince the HeLa cell extracts were prepared using buffers thatlacked phosphatase inhibitors. In contrast, preincubation ofmyometrial (Figure 3B, lane 4) or HeLa cell nuclear extracts(lane 8) with pan Fos (which recognizes the amino terminusof Fos family members c-Fos, Fos B, Fra-1 and Fra-2) priorto addition of the labelled oligonucleotide had no effect on
761
the mobility shift. These data suggest that the AP1 motifwithin the Cx43 promoter was recognized either by a Junhomodimer or a heterodimer comprised of Jun and either anundefined Fos family member or the CREB/ATF family thatoften heterodimerize with Jun.
Identification of Sp1 binding sites in Cx43 promoter
We next sought to identify the factor(s) that bind to the restof Region 1 encompassing from –80 to –45. Based on theextent of DNase I protection, this region probably containstandem transcription factor binding sites. A portion of thisregion (–65 and –48) is homologous to the mouse ‘activator’element (Chenet al., 1995). This element overlaps a sequencemotif (–59 to –48) that resembles the ‘CCTCC’ elements,reported to bind the ubiquitous transcriptional activator proteinSp1 and found within the promoters of rat growth hormone(Schaufeleet al., 1990), low density lipoprotein receptorpromotors (Dawsonet al., 1988) and CDC11c promotors (Notiet al., 1996). The upstream portion of Region 1 (–77 to –69)contains another CCTCC motif. Electrophoretic mobility shiftassay was performed using a double-stranded oligonucleotidespanning theCx43promoter from –100 to –50 (Figure 4). Weincluded the DNase I sensitive promoter sequence from (–100to –81) because the sequence (–93 to –87) corresponds to themouse ‘repressor’ element (Chenet al., 1995). Note alsothat the sequence from –99 to –89 (59-CCTCCTCCCAG) ishomologous to a portion of the CCTCC element within thedownstream portion of Region 1 (–56 to –46).
The results in Figure 4 show two major (a and c) and aminor (b) complex formed using extracts from the TPA-treatedprimary myometrial cell cultures (lane 2). The complexes arecompeted out at 200-fold excess of unlabelled oligonucleotidesfor self –100/–50 (lane 3) indicating site specificity. We alsoperformed competition analysis using selected subregions ofthe –100/–50 segment. All three bands were efficiently com-peted by addition of an unlabelled oligonucleotide derivedfrom Region 1 containing either the distal CCTCC motif(–83/–65, lanes 11 and 12) or the proximal CCTCC motif andoverlapping mouse activator element (–70/–48 lanes 5 and 6).Competition was also demonstrated (data not shown) with a

C.O.Echetebu et al.
Figure 3. (A). c-Jun binds to the Activator Protein 1 (AP1) site inhumanCx43promoter. [γ-32]P 59-end-labelled promoter-sequencespecific AP1 double-stranded oligonucleotide –52/–29 wasincubated without (lane 1) or with 5µg of tetradecanoyl phorbolacetate (TPA)-treated myometrial extract protein (lanes 2–9).Antibodies (10µg each) were added as indicated. Lanes 3 and 6contained polyclonal c-Jun antibody (Sc-1694X); lanes (4 and 7)contained c-Jun monoclonal antibody specific for phosphorylatedc-Jun (Sc-822X); lanes (5, 6 and 7) contained polyclonal antibodyto c-Fos (Sc-52X). Lanes 8 and 9 contained 100-fold excessunlabelled AP1 oligonucleotides. Arrows indicate c-Jun and itsphosphorylated supershifted complex. (B) Binding of c-Jun proteinsfrom myometrial and HeLa extracts to theCx43AP1 site.Radiolabelled oligonucleotide –52/–29 was incubated as describedin (A) without added protein (lane 1), or with 5µg of myometrialextract (lanes 2–5), or HeLa extract (lanes 6–11). Polyclonalpan-Jun antibody (Sc-44X) was added to lanes 3 and 7; plyclonalpan Fos (Sc-413X) was added to lanes 4 and 8 and monoclonalantiphosphorylated c-Jun (Sc-822X) was added to lanes 5 and 9.Lanes 10 and 11 contained 100-fold excess AP1 consensus andmutant oligonucleotides respectively.
consensus SP1 oligonucleotide and an oligonucleotide thatcontained the CCTCC motif found within the chicken adultβ-globin promoter (Emersonet al., 1985). Competition wasnot observed using oligonucleotide (–139/–119) that containeda consensus AP2 site, (lanes 9 and 10) or the non-relatedRegion 4 sequence (150/173, lanes 13, 14). The –80/–100oligonucleotide (which encompasses the homologous mouse‘repressor’ region and contains an overlapping AP2 consensusand CCTCC-like elements) partially competed for complexformation (lanes 7 and 8). We attribute this to the CCTCCmotif within this sequence. These data indicate that all threenucleoprotein complexes contain factors bound at the CC-TCC motifs.
The CCTCC motif has been reported to bind the ubiquitous
762
Figure 4. Competition gel shift assay of region –100/–50 of humanCx43promoter. Binding of myometrial nuclear factors to a labelledprobe spanning the –100 to –50 region (lane 2) was challengedwith excess 100-fold (odd-numbered lanes) and 200-fold (even-numbered lanes) of specific competitors for Activator Protein 2(AP2) (lanes 9–10) or other CCTCC-containing oligonucleotides(lanes 3–8, 11 and 12) and are non site specific oligonucleotide(lanes 13 and 14); a, b, c indicate the complexes.
Figure 5. Sp1 binds to the ‘CCTCC’ motifs of the region–100/–50. Labelled oligonucleotides –100/–50 was incubated withmyometrial extract and probed with polyclonal antibodies or Etsconsensus oligonucleotide as indicated. Only Sp1 is supershifted(lane 6). a, b, c indicate three different complexes binding to theoligonucleotide.
transcriptional activator protein Sp1 (Dawsonet al., 1988;Noti et al., 1996). To test this prediction we performedsupershift analysis using an anti Sp1 antibody (Figure 5, lane6). We conclude from these and other similarly performedexperiments (data not shown) that the nucleoprotein complexesa and b contain Sp1. The identity of the protein(s) withincomplex c remains to be determined. It is possible that thisnucleoprotein complex contains the ‘activator’ protein thatappears not to be Sp1 (Chenet al., 1995). In this regard, recallthat all three complexes were effectively competed for by eitherthe –70/–48 or–83/–64 oligonucleotides. These sequences alsocontain the motif 59-TCC-39 identified as the core bindingbases for most Ets family of transcription factors (Batcheloret al., 1998; Graves, 1998). The Ets family of transcriptionfactors comprises more than 20 distinct proteins found indifferent organisms. Preliminary studies, however, indicatedthat complex c was unaffected by competition with a consensusoligonucleotide recognized by a subset of Ets family members(lane 7). Additionally, no effect was observed when supershiftassays were performed with antibodies that recognize the Etsfamily members PU1 (lane 3), Ets1/Ets2 (lane 4) and GABPα/β (lane 5). The identity of the protein(s) within complex cremains unresolved.
AP1 and Sp1 regulate human myometrial Cx43 gene expression
Figure 6. Effect of replacement mutations in the region –100/–30on Cx43promoter activity. (A) 10 bp replacement mutantconstructs of pCx312, and the wild-type (WT) pCx312 fused topGL3 basic luciferase reporter. Luciferase activity was normalizedagainst a co-transfected pCMV-β-galactosidase plasmid. The meanluciferase activities from triplicate experiments6 SE (error bars)are indicated. (B) 2 or 3 bp replacement mutant replacement mutantconstructs of pCx312, and WT fused to luciferase reporter.*P , 0.05 compared with wild-type.
AP1 and Sp1 binding sites are necessary for Cx43
promoter activity
To determine the functional importance of the defined elementson Cx43promoter activity, we performed transient expressionassays using human myometrial primary cell cultures andCx43 10 bp replacement mutant promoter constructs. Theresults (Figure 6A) show that systematic replacement mutationsin the region –99 to –60 did not adversely affect promoteractivity. However, sequential replacement mutations in theregion –59 to –39, resulted in a dramatic reduction in promoteractivity. In particular, a five-fold reduction in promoter activitywas observed when either the proximal Sp1 (∆–59/–50, lane4) or AP1 (∆–49/–40 and∆–39/–30, lanes 5 and 6) sites were
763
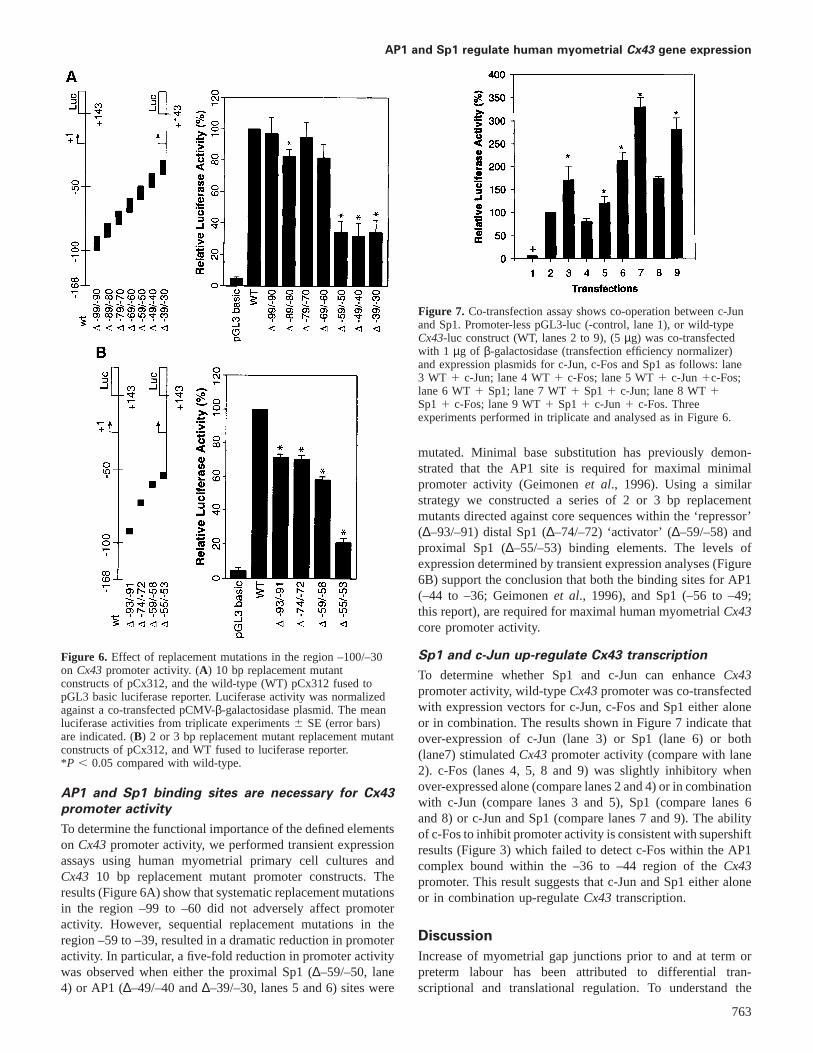
Figure 7. Co-transfection assay shows co-operation between c-Junand Sp1. Promoter-less pGL3-luc (-control, lane 1), or wild-typeCx43-luc construct (WT, lanes 2 to 9), (5µg) was co-transfectedwith 1 µg of β-galactosidase (transfection efficiency normalizer)and expression plasmids for c-Jun, c-Fos and Sp1 as follows: lane3 WT 1 c-Jun; lane 4 WT1 c-Fos; lane 5 WT1 c-Jun1c-Fos;lane 6 WT1 Sp1; lane 7 WT1 Sp11 c-Jun; lane 8 WT1Sp11 c-Fos; lane 9 WT1 Sp11 c-Jun1 c-Fos. Threeexperiments performed in triplicate and analysed as in Figure 6.
mutated. Minimal base substitution has previously demon-strated that the AP1 site is required for maximal minimalpromoter activity (Geimonenet al., 1996). Using a similarstrategy we constructed a series of 2 or 3 bp replacementmutants directed against core sequences within the ‘repressor’(∆–93/–91) distal Sp1 (∆–74/–72) ‘activator’ (∆–59/–58) andproximal Sp1 (∆–55/–53) binding elements. The levels ofexpression determined by transient expression analyses (Figure6B) support the conclusion that both the binding sites for AP1(–44 to –36; Geimonenet al., 1996), and Sp1 (–56 to –49;this report), are required for maximal human myometrialCx43core promoter activity.
Sp1 and c-Jun up-regulate Cx43 transcription
To determine whether Sp1 and c-Jun can enhanceCx43promoter activity, wild-typeCx43promoter was co-transfectedwith expression vectors for c-Jun, c-Fos and Sp1 either aloneor in combination. The results shown in Figure 7 indicate thatover-expression of c-Jun (lane 3) or Sp1 (lane 6) or both(lane7) stimulatedCx43promoter activity (compare with lane2). c-Fos (lanes 4, 5, 8 and 9) was slightly inhibitory whenover-expressed alone (compare lanes 2 and 4) or in combinationwith c-Jun (compare lanes 3 and 5), Sp1 (compare lanes 6and 8) or c-Jun and Sp1 (compare lanes 7 and 9). The abilityof c-Fos to inhibit promoter activity is consistent with supershiftresults (Figure 3) which failed to detect c-Fos within the AP1complex bound within the –36 to –44 region of theCx43promoter. This result suggests that c-Jun and Sp1 either aloneor in combination up-regulateCx43 transcription.
Discussion
Increase of myometrial gap junctions prior to and at term orpreterm labour has been attributed to differential tran-scriptional and translational regulation. To understand the

C.O.Echetebu et al.
transcriptional control, we began systematic studies to identifythe cis-elements in the humanCx43 promoter that interactwith transcription factors from human myometrium. We usedin-vitro DNase I footprinting to define potential regulatoryelements within the promoter region (–164 to1148) summar-ized in Figure 2. We identified four regions of protection:Region 1 encompassed ~50 bp of promoter sequence upstreamof the RNA start site; Region 2 includes the start site; Regions3 and 4 were localized to downstream sequences (Figure 1A).We then focused on Region 1 (–80 to –31) which contains anAP1 element (–44 to –36) and a region homologous to the‘activator’ region found in the mouseCx43 promoter (Chenet al., 1995). We have demonstrated by EMSA/supershiftassays that c-Jun binds at the AP1 site (–44 to – 36) and thatSp1 binds at tandem sites (–59 to –48 and –77 to –69).Mutational and transient transfection studies indicated that theAP1 and the proximal Sp1 sites are important for maximalpromoter function. Over-expression studies indicated that Sp1and c-Jun either alone or in combination stimulate transcription.We were unable to demonstrate c-Fos binding to the AP1 siteand over-expression of c-Fos either alone or in combinationwith c-Jun and or c-Fos slightly inhibited promoter activity.
TPA treatment of primary myometrial cells in cultureincreasesCx43expression and requires a functional AP1 site(Geimonenet al., 1996). TPA treatment of cells increases theirlevels of c-Jun and c-Fos proteins. Activation however, wasnot observed when c-Jun and or c-Fos were over-expressed inthe absence of TPA-treatment. Thus it was postulated that TPAfunctioned by activating c-Jun/c-Fos via phosphorylation. Incontrast, we performed similar over-expression experiments incells not treated with TPA. The results (Figure 7) showed thatover-expression of c-Jun stimulates promoter activity whileover-expression of c-Fos either alone or in combination withc-Jun is slightly inhibitory. Figure 7 also showed that over-expression of Sp1 either alone or in combination with c-Junstimulates promoter activity. Over-expression of c-Fos eitheralone or in combination with c-Jun and or Sp1 always resultedin slightly inhibited transient expression. The inhibitory effectof c-Fos and the inability to detect the Fos family in gel shifts(Figure 3A,B) lend support to the notion that the AP1 site isbound by c-Jun homodimers or heterodimers of c-Jun andeither unidentified Fos family members or other factors knownto interact with c-Jun (such as ATF/CREB).
The Sp1 family of proteins includes Sp1, Sp2, Sp3 andSp4 which bind to the canonical GC-box, (CCCGCCC;Kadonagaet al., 1986, 1988) and to the non-canonical CCTCCelement found in several gene promoters: rat growth hormone(Schaufeleet al., 1990), low density lipoprotein receptor(Dawsonet al., 1988), HIV-1 LTR (Joneset al., 1986) CDC11c(Noti et al., 1996) and KDR (Hataet al., 1998). Our data(Figures 4 and 5) strongly indicates that Region 1 containstandem Sp1 binding sites upstream of the AP1 element. Inaddition, this region promotes nucleoprotein complex forma-tion (c) of unknown composition. Based on studies of theKDR promoter which contains an Sp1/Sp3 binding elementidentical to that found within theCx43 promoter Region 1 –60 to –51 (59-AGCCCCTCCTCC-39), this complex could
764
contain Sp3. This study also determined which residues wereimportant (underline) or critical (bold-faced) for binding.
Our data from promoter mutagenesis and transfectionanalyses demonstrated that only the proximal Sp1 site (–59to –48) is functionally important and could correspond to themouse ‘activator’ element. The mouse ‘activator’ was definedusing an oligonucleotide (59-TTCTCCTAGCCCCTCCTT-39)that overlaps the human consensus Sp1 –67 to –50 (59-TTCTTCTAGCCCCTCCTC-39and contains the critical Sp1binding motif (bold). Mutations in the important region of themouse activator (GC→TT, AGCCCCTCCTT) had a modesteffect on activator protein binding. Paradoxically, this mutantalso abolished promoter activity in transient expression studies.In contrast, our studies indicate that an identical mutation hadessentially no effect on promoter activity. These discrepanciescould be due to differences in promoter constructs used in theassays. The mouse study used a 59-deletion construct thatremoved the distal Sp1 binding motif whereas our strategywas to perform mutational analysis within the context of theentire minimal promoter sequence. Thus, it is possible that theGC→TT mutation in the human sequence may be recompensedby the distal Sp1 binding site which is absent in the mousedeletion construct (Chenet al., 1995).
The mouse promoter also contains a ‘repressor’ elementcorresponding to a GC-box type (CCCGCC, –96 to–91) Sp1site (Chen et al., 1995). These studies demonstrated thatmutation∆–96/–94 (CCC→TTT) in the ‘repressor’ abolishedboth protein binding and repressive ability. However suscepti-bility of this region in the human promoter to DNase I digestionin our foot print experiment (Figure 1A) and the minimaleffect of identical mutational analysis on promoter activity(Figure 6A,B) strongly suggest that this region of theCx43promoter has diverged from the mouse promoter. Lack ofrepressor function is consistent with the A→C transversionwithin this region of the humanCX43promoter (CCCAGCC)which would be expected to abolish binding of Sp familymembers.
High levels of transcriptional activation by Sp1 requiremultiple Sp1 binding sites close to the transcription start site(Courey and Tjian, 1988), although a single binding siteappears to be sufficient for a promoter to be stimulated bySp1 (Kadonagaet al., 1987).Cx43promoter appears to containtandem Sp1 binding sites (Figure 2). Our data however indicatethat only the proximal Sp1 and the adjacent AP1 sites arerequired for maximal promoter activity. Sp1 recognitionsequences are often found near binding sites for other transcrip-tion factors, CTF/NF-1 (Joneset al., 1985) and AP1 (Leeet al., 1987a; Angelet al., 1987). Moreover, synergy has beenobserved for adjacent AP1 and Sp1 sites in other promoters(Lee et al., 1987b; Notiet al., 1996; Yeet al., 1996). It is notclear how AP1 and Sp1 interact to regulateCx43 promoteractivity, but Sp1 has been shown to interact physically with acomponent, dTAFII110 at the transcription factor IID (TFIID)that binds to the TATA-box element (Gillet al., 1994).Cx43promoter has a properly positioned TATA-like element (–25to –19) that is not protected from DNase I digestion, yettranscription, initiates accurately at the start site (Yuet al.,1994; DeLeonet al., 1994; Chenet al., 1995; Geimonen

AP1 and Sp1 regulate human myometrial Cx43 gene expression
et al., 1996). Sp1 interaction with dTAFII110 could stimulatetranscription by facilitating TFIID recruitment into the preiniti-ation complex. However, other elements might be used inestablishing the level of promoter activity.
Such other elements within the context of this study arethe down-stream protected regions (2–4) which include thetranscription initiation site, an Ets/NF-kB site and a TA-rich segment. The initiation site has the ascribed functionof binding the RNA polymeraseII and other factor(s) suchas the initiator factor (Zenzie-Gregoryet al., 1992) requiredfor accurate transcription initiation. The NF-kB family ofproteins is induced as defensive response to stress, injury,viral as well as bacterial pathogens (Siebenlistet al., 1994).It is up-regulated by tumour necrosis factor (TNF)-α,lipopolysaccharides, interleukins and TPA, and itself regulatesa number of stress as well as defensive genes. Labour is astress process for both mother and fetus, and may requirethe intervention of NF-kB activity. The roles that theseelements may play in uterineCx43 transcriptional regulationare, as yet, unknown.
In summary, our studies are the first to demonstrate byin-vitro DNase I foot printing potential regulatory proteinbinding sites in the region –164 to1148 of the humanmyometrial Cx43 promoter. We show that c-Jun and Sp1proteins bind to the promoter and independently up-regulateits activity but are both required for maximal promoter function.These data suggest that Sp1 and c-Jun may positively regulatetranscription of myometrialCx43 during the initiation oflabour.
ReferencesAndersen, J., Grime, E.A., Eng, C.L.-Y.et al. (1993) Expression of connexin
43 in human myometrium and leiomyomas.Am. J. Obstet. Gynecol., 169,1226–1276.
Angel, P., Imagawa, M., Chiu, R.et al. (1987) Phorbol ester-inducible genescontain a common cis-element recognized by a TPA-modulated trans-actingfactor. Cell, 49, 729–739.
Batchelor, A.H., Piper, D.E., de la Brousse, F.C.et al. (1998) The structureof GABPα/β: an Ets domain-ankyrin repeat heterodimer bound to DNA.Science, 279, 1037–1041.
Bennet, M.L.V and Goodenough, D.A. (1978) Gap junctions, electroniccoupling and intercellular communication.Neurosci. Res. Bull., 16, 373–386.
Beyer, E.C., Paul, D.L. and Goodenough, D.A (1987) Connexin 43: a proteinfrom rat heart homologous to a gap junction protein from liver.J. CellBiol., 105, 2621–2629.
Chen, Z.Q., Lefebvre, D., Bai, X.H.et al. (1995). Identification of tworegulatory elements within the promoter region of the mouse connexin 43gene.J. Biol. Chem., 270, 3863–3868.
Chow, L. and Lye, S.J. (1994) Expression of the gap junction protein connexin43 is increased in the human myometrium toward term and with the onsetof labor.Am. J. Obstet. Gynecol., 170, 788–795.
Cole, W.C., Garfield, R.E. and Kirkaldy, J.S. (1985) Gap junctions and directintercellular communication between rat uterine smooth muscle cells. Am.J. Physiol., 249, C20–C31.
Courey, A.J. and Tjian, R (1988) Analysis of Sp1in vivo reveals multipletranscriptional domains, including a novel glutamine-rich activation motif.Cell, 55, 887–898.
Dawson, P.A., Hofmann, S.l., van der Westhuyzen, D.R.et al. (1988)Sterol-dependent repression of Low Density Lipoprotein receptor promotermediated by 16-basepair sequence adjacent to binding site for transcriptionfactor SP1.J. Biol. Chem., 263, 3372–3379.
765
DeLeon, J.R., Buttrick, P.M and Fishman G.I. (1994) Functional analysis ofconnexin 43 gene promoterin vivo and in vitro. J. Mol. Cardiol., 26,379–389.
Emerson, B.M., Lewis, C.D. and Felsenfeld, G. (1985) Interaction of specificnuclear factors with the nuclease-hypersensitive region of the chicken adultβ-globin gene: nature of the binding domain. Cell, 41, 21–30.
Fishman, G.I., Eddy, R.L., Shows, T.B.et al. (1991) The human connexingene family of gap junctions: Distinct chromosomal locations but similarstructure.Genomics, 10, 250–256.
Garfield, R. E., Sims, S. and Daniel, E.E. (1977) Gap junctions: their presenceand necessity in myometrium during gestation. Science, 198, 958–959.
Garfield, R.E. and Hayashi, R.H. (1981) Appearance of gap junctions in themyometrium of women during labor.Am. J. Obstet. Gynecol., 140, 254–260
Garfield, R.E., Blennerhassett, M.G. and Miller, S.M. (1988a) Control ofmyometrial contractility: Role and regulation of gap junctions.Oxford Rev.Reprod. Biol., 10, 436–490.
Geimonen, E., Wei, J., Ali, M.et al. (1996) Activation of protein kinase C inhuman uterine smooth muscle induces connexin-43 gene transcriptionthrough an AP-1 site in the promoter sequence.J. Biol. Chem., 27,23667–23674.
Gill, G., Pascal, E., Tseng, Z.H. and Tjian, R. (1994) A glutamine-richhydrophobic patch in transcription factor Sp1 contacts the dTAFII110component of theDrosophilaTFIID complex and mediates transcriptionalactivation.Proc. Natl. Acad. Sci. USA, 91, 92–196.
Graves, B.J. (1998) Inner workings of a transcription factor partnership.Science, 279, 1000–1002.
Hata, Y., Duh, E., Zhang, K.et al. (1998) Transcription factors Sp1 and Sp3alter vascular endothelial growth factor receptor expression through a novelrecognition sequence.J. Biol. Chem., 273, 19294–19303.
Hendrix, E.M., Mao, S.J.T., Everson, W. and Larsen, W.J. (1992) Myometrialconnexin 43 trafficking and gap junction assembly at term and pretermlabor.Mol. Reprod. Dev., 33, 27–38.
Jones, K.A. Yamamoto, K.R. and Tjian, R. (1985) Two distinct transcriptionfactors bind to the HSV thymidine kinase promoterin vitro. Cell, 42,559–572.
Jones, K.A., Kadonaga, J.T., Luciw, P.A and Tjian, R. (1986) Activation ofthe AIDS retrovirus promoter by cellular transcription factor Sp1. Science,232,755–759.
Kadonaga, J.T., Jones, K.A. and Tjian, R. (1986) Promoter-specific activationof RNA polymerase II transcription by Sp1.Trends Biol. Sci., 11, 20–23
Kadonaga, T.J., Carner, K.R., Marsiarz, F.R. and Tjian, R. (1987) Isolation ofcDNA encoding transcription factor Sp1 and functional analysis of theDNA binding domain. Cell, 51, 1079–1090.
Kadonaga, J.T., Courey, A.J., Ladika, J. and Tjian, R. (1988) Distinct regionsof SP1 modulate DNA binding and transcriptional activation. Science, 242,1566–1570.
Lee, W., Haslinger, A., Karin, M. and Tjian, R. (1987a) Activation oftranscription by two factors that bind promoter and enhancer sequences ofthe human metallothionein gene and SV40. Nature, 325, 368–372.
Lee, W., Mitchell, P. and Tjian, R (1987b) Purified transcription factor AP-1interacts with TPA-inducible enhancer elements. Cell, 49, 741–752.
Lee, S.W., Tomasetto, C., Paul, D.et al. (1992) Transcriptional down-regulation of gap junction proteins blocks communication during ratpregnancy.J. Cell. Biol., 118, 1213–1221.
Li, Z., Zhao, Z. and Daniel, E.E. (1993) Expression of gap junction connexin43 mRNA in different tissues of intestine in dog.Am. J. Physiol., 265,G911–G916.
Lye, S.J, Nicholson, B.J., Mascarenhas, M.et al. (1993) Increased expressionof connexinn-43 in the rat myometrium during labor is associated with anincrease in the plasma estrogen. Endocrinology, 132, 2380–2396.
MacKenzie, L.J. and Garfield, R.E. (1985) Hormonal control of gap junctionsin the myometrium.Am. J. Physiol., 248, C297–C300.
Maxam, A.M. and Gilbert, W. (1980) Sequencing end-labeled DNA withbase-specific chemical cleavages.Methods Enzymol., 65, 499–560.
McNutt, C.M., Nicholson, B.J. and Lye, S.J. (1994) ACTH-induced pretermlabor in the ewe is associated with increased mRNA and protein levels ofmyometrial gap junction protein, connexin 43.J. Endocrinol., 141, 195–202.
Miller, S.M., Garfield, R.E. and Daniel, E.E. (1989) Improved propagation inmyometrium associated with gap junction during parturition.Am. J. Physiol.,256, C130–C141.
Miyoshi, M., Boyle, M.B., MacKay, L.B. and Garfield, R.E. (1996) Voltage-clamp studies of gap junctions between uterine muscle cells during termand preterm labor.Biophys J., 71, 1324–1334.

C.O.Echetebu et al.
Noti, J.D., Reinemann, B.C. and Petrus, M.N. (1996) Sp1 binds two sites inthe CD11c promoterin vivo specifically in myeloid cells and cooperateswith AP1 to activate transcription.Mol. Cell. Biol., 16, 2940–2950.
Petrocelli, T. and Lye, S.J. (1993) Regulation of transcripts encoding themyometrial gap junction protein, connexin 43, by estrogen and progesterone.Endocrinology, 133, 284–290.
Pitts, J.D and Simms, J.W. (1977) Permeability of junctions between animalcells; intercellular transfer of nucleotides but not of macromolecules.Exp.Cell, Res., 104, 153–163.
Risek, B., Guthrie, S., Kumar, N. and Gilula, N.B. (1990) Modulation of gapjunction transcript and protein expression during pregnancy in the rat.J.Cell. Biol., 110, 269–282.
Risek, B. and Gilula, N.B (1991) Spatio–temporal expression of three gapjunction gene products involved in fetomaternal communication during ratpregnancy.Development, 113, 165–181.
Schaufele, F. West, B.L. and Reudelhuber, T. (1990) Somatotroph- andlactotroph-specific interactions with the homeobox protein binding sites inthe rat growth hormone gene promoter.Nucleic Acids Res., 18, 5235–5243
Shapiro, D.J., Sharp, P.A. Wahli, W. and Keller, M.J. (1988) A high efficiencyHeLa cell nuclear transcription extract.DNA, 7, 47–55.
Siebenlist, U. Franzoso, G. and Brown, K. (1994) Structure, regulation andfunction of NF-kB.Ann. Rev. Cell Biol., 10, 405–455.
Sims, S.M., Daniel, E.E. and Garfield, R.E. (1982) Improved electricalcoupling in uterine smooth muscle is associated with increased numbers ofgap junctions at parturition.J. Gen. Physiol., 80, 353–375.
Tabb, T., Thilander, G., Grover, A. Hertzberg, E. and Garfield, R.E. (1992)An immunochemical and immunocytological study of the increase inmyometrial gap junctions (and connexin 43) in rats and humans duringpregnancy.Am. J. Obstet. Gynecol., 167, 559–567.
Vallette, F., Mege, E., Reiss, A. and Adesnik, M. (1989) Construction ofmutant and chimeric genes using the polymerase chain reaction.NucleicAcids Res., 2, 723–733.
Vanden Heuvel, J.P., Tyson, F.L. and Bell, D.A. (1993) Construction ofrecombinant RNA templates for use as Internal Standards in QuantitativeRT–PCR.Biotechniques, 14, 395–398.
Winterhager, E., Stutenkemper, R. Traub, O.et al. (1993) Expression ofdifferent connexin genes in rat uterus during decidualization and at term.Eur. J. Cell. Biol., 55, 133–142.
Ye, J., Zhang, X. and Dong, Z. (1996) Characterization of human granulocyte–macrophage colony stimulating factor gene promoter: an AP1 complex andan Sp1-related complex transactivate the promoter activity that is suppressedby a YY1 complex.Mol. Cell. Biol., 16, 157–167.
Yu, W., Dahl, G. and Werner, R. (194) The connexin 43 gene is responsiveto oestrogen.Proc. R. Soc. Lond. B, 255, 125–132.
Zenzie-Gregory, B., O’Shea-Greenfield, A. and Smale, S.T. (1992) Similarmechanisms for transcriptional initiation mediated through a TATA box oran initiator element.J. Biol. Chem., 267, 2823–2830.
Zhao, K, Kuperman, L., Geimonen, E. and Andersen, J. (1996) Progestinrepresses human connexin 43 gene similarly in primary cultures ofmyometrial and uterine leiomyomacells.J. Biol. Reprod., 54, 607–615.
Received on October 19, 1998; accepted on April 16, 1999
766
Copyright © 2022 FDOKUMEN




















