
Clusterin interacts with Paclitaxel and confer Paclitaxel resistance in ovarian cancer
Upload
independentCategory
view
2download
0
Connexin 43 Interacts with Zona Occludens-1 and -2 Proteins ina Cell Cycle Stage-specific Manner*
Deepika Singh‡,¶,§, Joell L. Solan‡,¶, Steven M. Taffet||, Ronald Javier**, and Paul D.Lampe‡,¶,‡‡
‡Division of Public Health Sciences, Fred Hutchinson Cancer Research Center, Seattle,Washington 98109¶Department of Pathobiology, University of Washington, Seattle, Washington 98195||Department of Microbiology, SUNY Upstate Medical University, Syracuse, New York 13210**Department of Molecular Virology and Microbiology, Baylor College of Medicine, Houston,Texas 77030
AbstractGap junction channels play an important role in cell growth control, secretion and embryonicdevelopment. Gap junctional communication and channel assembly can be regulated by protein-protein interaction with kinases and phosphatases. We have utilized tandem mass spectrometry(MS/MS) sequence analysis as a screen to identify proteins from cell lysates that interact with theC-terminal cytoplasmic region of connexin 43 (Cx43). MS/MS analysis of tryptic fragmentsyielded several proteins including zona occludens-1 (ZO-1), a structural protein previouslyidentified to interact with Cx43, and ZO-2, a potential novel interacting partner. We confirmed theinteraction of ZO-2 with Cx43 by using a combination of fusion protein “pull down,” co-immunoprecipitation, and co-localization experiments. We show that the C-terminal region ofCx43 is necessary for interaction with the PDZ2 domain of ZO-2. Far Western analysis revealedthat ZO-2 can directly bind to Cx43 independent of other interacting partners.Immunofluorescence studies indicate that both ZO-1 and ZO-2 can co-localize with Cx43 withinthe plasma membrane at apparent gap junctional structures. We examined Cx43 interaction withZO-1 and ZO-2 at different stages of the cell cycle and found that Cx43 had a strong preferencefor interaction with ZO-1 during G0, whereas ZO-2 interaction occurred approximately equallyduring G0 and S phases. Since essentially all of the Cx43 in G0 cells is assembled into TritonX-100-resistant junctions, Cx43-ZO-1 interaction may contribute to their stability.
Gap junctions are tightly packed clusters of intercellular channels that directly connect thecytoplasms of adjacent cells. These pathways provide for the cell-to-cell diffusion of smallmolecules, including ions, amino acids, nucleotides, and secondary messengers (e.g. Ca2+,cAMP, cGMP, inositol 1,4,5-trisphosphate) and for the conductance of electrical impulses inexcitable cells. There are at least 20 gap junction protein or “connexin” family members inhumans, many of which have been cloned and characterized in mice (1, 2). Connexinexpression is tissue specific, and connexin 43 (Cx43)1 is the predominant connexin inepithelial and most other tissues. Recent studies utilizing transgenic mice with altered
*This work was supported by National Institutes of Health Grants GM55632 and AR47963.
© 2005 by The American Society for Biochemistry and Molecular Biology, Inc.‡‡To whom correspondence should be addressed: Fred Hutchinson Cancer Research Center, PO Box, 19024, Mailstop M5C800, 1100Fair-view Ave. N., Seattle, WA 98109. Tel.: 206-667-4123; [email protected].§Recipient of an International Training and Research in Emerging Infectious Diseases fellowship.
NIH Public AccessAuthor ManuscriptJ Biol Chem. Author manuscript; available in PMC 2012 November 20.
Published in final edited form as:J Biol Chem. 2005 August 26; 280(34): 30416–30421. doi:10.1074/jbc.M506799200.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

connexin genes and linkage of connexin gene alterations to human disease provide strongsupport for roles in cell growth control and embryonic development. For example, thehomozygous deletion of Cx37, Cx40, Cx43, and Cx45 causes neonatal or embryoniclethality in mice (1–3). In humans, mutations in Cx26, Cx30, Cx31.1, Cx32, Cx43, Cx46,and Cx50 are associated with deafness/hearing loss, skin disorders, Charcot-Marie-Toothdisease, developmental defects, and cataract formation (1, 3).
Recent studies have shown that Cx43 can interact with several different signaling andscaffolding proteins. Perhaps the most well known is the interaction of zona occludens-1(ZO-1) with the carboxyl terminus of Cx43 (4, 5). Originally identified as a component oftight junctions, ZO-1, ZO-2, and ZO-3 are members of the membrane-associated guanylatekinase family of proteins that each contain at least one PSD95/Dlg/ZO-1 (PDZ) domain, anSrc homology 3 domain, and an enzymatically inactive guanylate kinase domain (6–9). PDZdomains are ~90-amino acid protein-protein binding domains that recognize at least a 3-residue peptide motif in the COOH termini of their binding partners (10). PDZ domain-containing proteins like ZO-1 typically act as scaffolding proteins that organize membranereceptors and cytosolic proteins into multimeric signaling complexes often at the sites ofcell-cell contact (11, 12). Divergent roles have been proposed for the interaction of ZO-1and Cx43 including the control of gap junction formation and localization to gap junctionplaques (5), internalization and remodeling of Cx43 in response to intracellular changes (13,14), and targeting for endocytosis (15). Furthermore, c-Src interaction with Cx43 has beenshown to regulate Cx43-ZO-1 interaction (16–18).
Other proteins that have been shown to interact with Cx43 include several kinases andcytoskeletal proteins such as c- and v-Src kinase, protein kinase C, mitogen-activatedprotein kinase, casein kinase 1, cAMP-dependent protein kinase, receptor protein-tyrosinephosphatase μ, α/β tubulins, CCN3/NOV, and Drebrin (e.g. see Ref. 19 for review andRefs. 20–22). Other connexins, including Cx31.9, Cx45, Cx46, and Cx50, have also beenshown to interact with ZO-1 (19, 23–25). Although several potential functions have beenproposed for these protein-Cx43 interactions, the roles that they play in the regulation ofCx43 trafficking, assembly, gating, and turnover are not well understood. We believed that athorough analysis and classification of Cx43-interacting proteins might elucidate these roles,so we employed a proteomic approach to identify proteins that interact with Cx43 directly orvia a protein complex.
Our mass spectrometry-based approach yielded many potential binding partners (26), andhere we report that Cx43 directly binds to ZO-2. We have characterized the interaction ofZO-2 with Cx43 by using a combination of “pull down,” co-immunoprecipitation, farWestern, and immunofluorescence experiments. We present evidence that the C-terminaltail of Cx43 interacts directly with ZO-2 in vitro, in cultured cells and in cardiac tissue.Comparison of the binding of ZO-1 and ZO-2 shows that ZO-1 binds preferentially to Cx43during the G0 stage of the cell cycle. Our data suggest that ZO-1 and ZO-2 are keyscaffolding proteins that can differentially bind to Cx43 at the membrane junctional complexduring the cell cycle.
1The abbreviations used are: Cx43, connexin 43; NRK, normal rat kidney; CT, carboxyl-terminal; GST, glutathione S-transferase;PBS, phosphate-buffered saline; ZO-1, zona occludens-1; ZO-2, zona occludens-2; MS/MS, tandem mass spectrometry; PDZ,PSD-95/Discs-large/ ZO-1; HA, hemagglutinin.
Singh et al. Page 2
J Biol Chem. Author manuscript; available in PMC 2012 November 20.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

EXPERIMENTAL PROCEDURESPlasmids
N-terminal, HA-tagged wild type ZO-2 (HA-ZO-2) was cloned in the cytomegalovirusexpression vector GW1 as previously described to generate GW1HA-ZO-2 (27). ZO-2sequences encoding the PDZ1 (amino acids 4–113), PDZ2 (amino acids 290–371), andPDZ3 (amino acids 492–585) domains were cloned in the PGEX-2T vector to generateglutathione S-transferase (GST)-PDZ1, GST-PDZ2, and GST-PDZ3, respectively. TheCx43 sequence encoding for the carboxyl terminus (CT) amino acids 236–382 of Cx43 wascloned into pGEX-2T or pGEX-2TK vector to generate GST-Cx43CT. CT deletionconstructs Δ290 (missing amino acids 280–300), Δ330 (missing amino acids 321–340),Δ369 missing (amino acids 364–373), T-374 (missing amino acids 375–382), and T-379(missing amino acids 380–382) were all cloned into the pGEX-2T vector to generate therespective GST deletion constructs. GST constructs were transformed into DH5αEscherichia coli, and GST fusion proteins were expressed and purified as previouslydescribed (28). The recombinant proteins were quantified in gels by Coomassie BrilliantBlue staining using bovine serum albumin as a standard.
Cell Culture and Cell ExtractsNormal rat kidney (NRK) epithelial cells (NRK-E51; American Tissue Culture Collection,Manassas, VA) were grown in Dulbecco’s modified Eagle’s medium supplemented with10% fetal bovine serum (Atlanta Biologicals, Atlanta, GA) in a humidified chamber with5% CO2 at 37 °C. The cells were grown to confluence, washed in PBS, and harvested on icein 400 μl of radioimmune precipitation assay lysis buffer (25 mM Tris-HCl, 100 mM NaCl,10 mM EDTA, 50 mM NaF, 500μM Na3VO4, 0.25% Triton X-100, 0.02% NaN3, 1×Complete protease inhibitors (Roche Applied Science), and 2 mM phenylmethylsulfonylfluoride, pH 7.2). The lysate was cleared by centrifugation at 13,000 rpm for 2 min. Cellsupernatant from NRK cells was precleared with GST bound to glutathione beads or proteinA and used in pull down or immunoprecipitation experiments, respectively. Our protocol forthe cell cycle synchronization of NRK cells has been previously described (29). 293-T cellswere grown to 80% confluence and transiently transfected with 1 μg of GW1HA-Z0 –2using Fugene reagent (Roche Applied Science) as per the manufacturer’s instructions.Transfected cells were harvested in radioimmune precipitation assay buffer after 48 h.
Liquid Chromatography-MS/MS AnalysisOur protocol for analyzing Cx43CT-binding proteins via mass spectrometry has beendescribed earlier (26). Briefly, GST-Cx43CT bound on glutathione-coated beads wasincubated with NRK cell lysates. After washing, Cx43CT and any potential interactingproteins were eluted from the beads using either 6 M urea or cleavage at the thrombin siteengineered into the fusion protein. The eluted proteins were digested with trypsin andanalyzed by electrospray-MS/MS on a ThermoFinnegan LCQ spectrometer. Tandem massspectrometry data of individual peptides was analyzed with SEQUEST™, which identifiedpossible peptide sequence matches by searching recorded MS/MS spectra against a ratsequence data base (30, 31). High scoring peptides were manually inspected to ensurecorrect matches.
GST Pull Down, Immunoprecipitation, and ImmunoblottingFor GST pull down assays, 5 μg of the individual GST fusion proteins bound toglutathioneagarose beads (Pierce) were incubated at 4 °C for 1 h with 400 μl of cell lysate(prepared as described above). For immuno-precipitations, rabbit anti-ZO-1, rabbit anti-ZO-2 (Zymed Laboratories Inc., San Francisco, CA), rabbit anti-Cx43 (C6219; Sigma), or
Singh et al. Page 3
J Biol Chem. Author manuscript; available in PMC 2012 November 20.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

anti-Myc (Cell Signaling Technology, Beverly, MA) antibodies were incubated with celllysate for 3 h followed by immunoprecipitation with protein A-Sepharose beads for 1 h. Thebeads were washed extensively in PBS, and bound protein was eluted in Laemmli samplebuffer followed by separation on SDS-10% PAGE and immunoblotting. For immunoblotdetection, rabbit anti-ZO-1, rabbit anti-ZO-2, rabbit anti-Cx43 (C6219; Sigma), or mouseanti-Cx43NT (described in Ref. 32) were used as primary antibodies and then visualizedwith fluorescent dye-labeled secondary antibodies (AlexaFluor 680 goat anti-rabbit(Molecular Probes, Inc., Eugene, OR) and IRDye800-conjugated donkey anti-mouse IgG(Rockland Immunochemicals, Gilbertsville, PA); both extensively cross-reacted againstother species) and directly quantified using the Li-Cor Biosciences Odyssey infraredimaging system and associated software (inverted images are presented).
Confocal Microscopy and Co-localization AnalysisCultured cells were grown on glass coverslips, washed with PBS, and fixed in chilledmethanol/acetone (1:1) for 2 min. Heart tissue was harvested from a mouse; the heart wasbisected and frozen in Tissue-TEK OCT compound (Torrance, CA); and 5-μm sectionswere cut on a cryostat. The sections were fixed in ice-cold acetone and washed in PBS.After blocking with 1% bovine serum albumin in PBS, heart sections or cells were incubatedwith mouse anti-Cx43IF1 (IgG2a; described in Ref. 28), rabbit or mouse (IgG1) anti-ZO-1,and/or rabbit anti-ZO-2 antibody for 1 h. After four washes, the cells were incubated withAlexaFluor 488-conjugated goat anti-mouse IgG2a, AlexaFluor 633-conjugated goat anti-mouse IgG1, and AlexaFluor 546-conjugated goat anti-rabbit secondary antibodies(Molecular Probes) for 1 h. Coverslips were washed in PBS and mounted onto slides usingDABCO anti-fade medium (25 mg/ml of 1,4-diazabicyclo-(2,2,2)octane (Sigma) diluted inSpectroglycerol (East-man Kodak Co.) and 10% PBS, pH 8.6), and images were acquired ona Leica TCS Spectral Confocal Microscope or a Zeiss LSM 510 laser-scanning microscopewith a ×100 oil objective. For quantitative studies, acquisition parameters were keptidentical between the different experimental groups. Colocalization measurements wereobtained by three-dimensional analysis of images, using Imaris Coloc (Bitplane Inc., SaintPaul, MN), selecting only the plasma membrane. The values from several images wereaveraged and are expressed as the percentage of Cx43 co-localized with the ZO proteins.S.D. and standard t test (two-sided) values were calculated. Z-planes and line scan analysisof images were generated using Metamorph 4.6 (Universal Imaging Corp., Westchester,PA). Z-planes were projected from 0.3-μm optical slices, and line scans were 0.9 μm wide.
Far Western Overlay AssayThe far Western assay described by Margolis et al. (33) was employed with minormodifications. GST-Cx43CT (10 μg) bound to glutathioneagarose beads was suspended inkinase buffer (50 mM potassium phosphate, pH 7.2, 10 mM MgCl2, 5 mM NaF, and 5 mMdithiothreitol) to which activated MAP kinase (prepared as described in Ref. 34) and 50 μCiof [γ-32P]ATP (Blu002; PerkinElmer Life Sciences) were added. After washing the beadswith cold PBS, the phosphorylated protein was eluted with 500 μl of 100 mM Tris-HCl, pH8.0, containing 1 mM each of EDTA, dithiothreitol, phenylmethylsulfonyl fluoride, 0.005%Nonidet P-40 detergent, and 20 mM reduced glutathione. The protein was resuspended in farWestern buffer (20 mM HEPES, pH 7.5, 1 mM KCl, 5 mM MgCl2, 5 mM dithiothreitol,0.02% sodium azide, and 2% nonfat dry milk). Cell lysates were separated on an SDS-10%polyacrylamide gel and were transferred to nitrocellulose. The membrane was blocked in thefar Western buffer and incubated with the radiolabeled GST-Cx43CT probe overnight at 4°C. The membrane was washed three times with PBS containing 0.1% Triton X-100, air-dried, and exposed to x-ray film with an intensifying screen. Densitometry ofautoradiographs was performed with NIH Image (a public domain program developed at theNational Institutes of Health and available on the World Wide Web at rsb.info.nih.gov/nih-
Singh et al. Page 4
J Biol Chem. Author manuscript; available in PMC 2012 November 20.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

image/) on images collected with a Sharp JX-325 scanner. In some experiments, the sameblot was also probed with antibodies to ZO-1 and ZO-2 as described above and quantifiedusing the Li-Cor system.
RESULTSCx43 Associates with ZO-2
The C-terminal region of Cx43 contains multiple putative protein interaction motifs andseveral phosphorylation sites that have been shown to regulate protein function. Weemployed tandem mass spectrometry (MS/MS) as a screen to identify Cx43-interactingpartners in a pull down experiment. The C-terminal tail (amino acids 246–382) of Cx43fused to a GST tag (Cx43CT) was incubated with NRK cell lysates, and bound proteinswere precipitated, washed, eluted, trypsinized, and subjected to MS/MS. The mass spectrawere analyzed using SEQUEST, an algorithm that correlates the experimental massspectrum data with theoretical spectra generated from known protein sequences in a database and ranks the “fit” with X-correlation and Delta Cn values (30).
Our mass spectrometry-based approach identified numerous proteins that potentially interactwith the C-terminal tail of Cx43. In a preliminary report associated with the 2003International Gap Junction Conference, we identified 19 possible interacting proteins (26).Four have been reported to interact with Cx43, including protein kinase Cα, casein kinase 1,tubulin, and ZO-1 (4, 5, 16, 28, 35). Another ZO family member, ZO-2, was identified withthe highest degree of certainty of the group. Two distinct ZO-2 peptides were identified,STGDITA-AGVTEANKEPR (P1) and VVDTLYDGK (P2), with high X-correlation (3.53and 2.54) and Delta Cn values (0.58 and 0.47), respectively. These sequences are absent inother ZO proteins; the sequence of P1 is found in the interdomain region between PDZ2 andPDZ3, whereas P2 falls in the Src homology 3 domain region of ZO-2.
The PDZ2 Domain of ZO-2 Interacts with the C-terminal End of CT in GST Pull-downAssays
To assess the ZO-2 interaction with Cx43, we incubated Cx43CT bound to glutathionebeads with NRK cell lysates for 1 h. After extensive washing of the beads, the boundproteins were eluted in sample buffer, and the presence of ZO-2 was examined via Westernanalysis. The antibody for ZO-2 detected a strong band at the expected molecular size (160kDa) when GST-Cx43CT was incubated with NRK cell lysates but not with GST alone (Fig.1A).
To map the minimal domain of Cx43 that interacted with ZO-2, we prepared several mutantconstructs of Cx43CT, including a truncation lacking the last 3 and 8 residues (T379 andT374), a deletion of residues 364–373 (Δ369), a deletion of residues 280–300 (Δ290), and adeletion lacking 320–340 (Δ330). In Fig. 1A, we show that ZO-2 bound to full-lengthCx43CT, to Δ290, and to Δ330 but neither to Δ369, T379, nor T374. Thus, similar to ZO-1(18), ZO-2 appears to bind to the most C-terminal residues of Cx43 but in a mannerdependent on residues up to 20 from the C terminus. Other reports have shown thatsequences C-terminal to the 3-amino acid PDZ domain-binding motif can influence bindingto PDZ domains (10).
We next determined which region of ZO-2 bound to full-length Cx43. Lysates of NRK cellswere incubated with GST fusions of the three separate PDZ1,-2, and -3 regions of the ZO-2molecule bound to glutathione beads (Fig. 1B). After extensive washing of the beads, boundproteins were eluted in sample buffer, and Western analysis for Cx43 was performed.Connexin signal was only detected in the lane where the fusion protein containing PDZ2was used and not with PDZ1, PDZ3, or GST alone (Fig. 1B).
Singh et al. Page 5
J Biol Chem. Author manuscript; available in PMC 2012 November 20.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

Cellular Cx43 and ZO-2 InteractWe sought to determine whether cellular Cx43 complexes with endogenous ZO-2 in NRKcells. We immunoprecipitated ZO-2 from NRK cell lysates using an anti-ZO-2 antibody andimmunoblotted for Cx43 (Fig. 2A). Cellular Cx43 appears as several bands. The bands oflower electrophoretic mobility correspond to phosphorylated forms of Cx43 and are oftenlabeled as P1 and P2, whereas species that co-migrate with the nonphosphorylated bandhave the highest mobility (P0). Unidentified Cx43 phosphorylation events have beenassociated with the formation of gap junction plaques (36, 37). A strong Cx43 signal wasobserved in lanes representing whole cell lysate, ZO-1 immunoprecipitate (which served asa positive control) and the ZO-2 immunoprecipitate. No signal was observed when a controlantibody (to Myc) or protein A beads alone were incubated with the cell lysate. We alsoimmunoprecipitated Cx43 and immunoblotted for ZO-2 (Fig. 2B). ZO-2 was detected in thewhole cell lysate and when Cx43 antibody was used for immunoprecipitation but not whenthe Myc antibody or Protein A beads alone was used.
ZO-2 Co-localizes with Cx43 in NRK Cells and HeartTo show whether ZO-2 could interact with Cx43 prior to cellular lysis, we performed co-immunolabeling using confocal microscopy. As shown in Fig. 3, ZO-2 and Cx43 arepredominately localized to the plasma membrane in NRK cells and were frequently but notexclusively co-localized at cell-cell contacts in punctuate staining patterns reminiscent ofgap junctions (shown in all three dimensions by superimposing the signal for Cx43 andZO-2 as shown in the Overlay panel).
We also performed high resolution immunofluorescence to determine whether Cx43 andZO-2 can co-localize in the intercalated disk region of heart tissue (Fig. 3). Distinct co-localization of Cx43 and ZO-2 was frequently demonstrated in the x-y, x-z, and y-z planes.In both NRK cells and heart tissue, comparison of the individual channels (i.e. Cx43 andZO-2) indicates that they often coincidentally increase in intensity (i.e. signal strength andshape for each channel are related), indicating bona fide co-localization. However,fluorescence co-localization studies are limited by the resolution of light, so we cannot tellwhether the apparent interaction is direct or not.
ZO-2 Can Bind Directly to Cx43ZO-1 is known to form independent complexes with ZO-2 and ZO-3 at tight junctions (38),and thus, ZO-2 could be interacting indirectly with Cx43 via its interacting partner ZO-1. Toinvestigate the specificity of the ZO-Cx43 interaction further, we employed a far Westernanalysis. Western blots of lysates from GW1HA-ZO-2-transfected and untransfected 293-Tcells were incubated with a radiolabeled Cx43CT probe. The Cx43CT probe bound to a 160-kDa protein only in the lane of the 293-T cells transfected with HA-ZO-2 (Fig. 4, FW panel,+ lane). Western analysis of the same blot using ZO-2 antibodies indicated that this bandcorresponds to the exogenously expressed ZO-2 (IB panel, lower portion). Untransfected293-T cells contain only low levels of intrinsic ZO-1 (IB panel, upper portion) and ZO-2 (IBside, lower panel, − lane), so essentially no detectable radiolabeled probe was bound. Thus,Cx43 only bound to ZO-2 that was ectopically expressed in the 293-T cells. This confirmedthat the binding of Cx43 with ZO-2 was specific and direct.
ZO-1 and ZO-2 Bind Differentially to Cx43 during the Cell CycleSince both ZO-1 and ZO-2 can bind to the same region of Cx43, we compared their co-localization at different stages of the cell cycle to potentially elucidate distinct roles. Asshown in Fig. 5A, Cx43, ZO-1, and ZO-2 interact extensively during G0 (Cx43 (green) +ZO-1 (blue) + ZO-2 (red) = white in the overlay), whereas green (Cx43) and yellow (Cx43 +
Singh et al. Page 6
J Biol Chem. Author manuscript; available in PMC 2012 November 20.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

ZO-2) were more apparent and independent of ZO-1 in S phase (Fig. 5B). Calculation of thepercentage of co-localization (see “Experimental Procedures”) indicated that significantlymore (p < 0.03) Cx43 co-localized with ZO-1 in G0 phase (~60%) than in S phase (~30%)cells (Fig. 5C). Cx43 binding to ZO-2 appeared to be independent of cell cycle stage, with~45% co-localization at either stage (Fig. 5C). Higher magnification of the indicatedjunctional regions (marked with parallel white lines) in the G0 and S phase images (Fig. 5, Aand B) showed more apparent green (Cx43) and yellow (Cx43 + ZO-2) in the S phase (Fig.5E) compared with G0 (Fig. 5D) images consistent with the co-localization values. Line scananalysis of the indicated regions often showed very good concordance of all three proteins inG0 cells (Fig. 5D; regions with good overlay are indicated by an asterisk), but S phase cellsshowed less alignment of Cx43 and ZO-1, so Cx43-ZO-2 association was more apparent(Fig. 5E). Since co-localization studies are limited by the resolution of light and both Cx43and the ZO proteins are present in the plasma membrane, the percentage of co-localizationcalculated above may overestimate the true extent of interaction. However, these analysesconfirm that the signal from Cx43, ZO-1, and ZO-2 channels can increase proportionally instrength and shape. Therefore, bona fide co-localization can occur, and the relative levelschange during the cell cycle.
In order to get a more direct measurement of interaction, we also examined whether ZO-1bound directly to Cx43 in a cell cycle-dependent manner via far Western analysis. Westernblots containing cell lysates from G0, G1, S, and G2/M were analyzed (Fig. 6) via farWestern analysis with radiolabeled Cx43CT (Fig. 6A) followed by reprobing of the sameblot for ZO-1 (Fig. 6B), ZO-2 (Fig. 6C), and Cx43 (Fig. 6D) using multiple antibodies andthe Li-Cor imaging system. In G2/M cells, Cx43 migrated as multiple bands, including an Mphase-specific band (P3), which has reduced mobility compared with P1 and P2 (39).Comparison of the extent of Cx43CT binding with the amount of ZO-1 protein present (Fig.6E) showed that Cx43 bound significantly more extensively (p < 0.01) to ZO-1 present inG0 cells than in either G1, S, or G2/M cells. The level of binding of Cx43CT to ZO-2 waslower than ZO-1 and not significantly different at any of the cell cycle stages. Thus, cellcycle-dependent post-translational modification of ZO-1 may affect its interaction withCx43. However, one caveat with these experiments is that although the gel-fractionatedproteins are full-length and presumably appropriately post-translationally modified, theradiolabeled Cx43CT probe is not, so judging relative affinities of ZO-1 and ZO-2 is notpossible with this method. However, both the far Western and immunofluorescence studiesindicate that Cx43 interacts most extensively with ZO-1 when cells are quiescent and thatZO-2 may play a more predominant role during the cell cycle.
DISCUSSIONMultiple laboratories have used the C-terminal region of Cx43 in a pull-down approach toscreen for possible interacting proteins (20, 26, 40). Two of these studies utilized SDS-PAGE separation and sequencing to identify specific, prominent bands (20, 40). Tubulinwas detected in all three studies, and Drebrin interaction was detected in one (20). Our studyutilized direct detection by mass spectrometry and yielded many additional possibleinteracting proteins including HSP71, 40S ribosomal protein S7, serine/threonine proteinphosphatase 2A, POU domain class 3 transcription factor 4, SNIP, interleukin-12α,fibrillin-2, α3-tubulin, vimentin, γ-aminobutyric acid receptor, Lin-7, Discs-large, band4.1(L), casein kinase Iα, vascular protein-tyrosine phosphatase 1, BIG-2, cyclic nucleotide-gated channel, ZO-1, and protein kinase Cα (26). Although this technique is quite sensitive,false identifications are likely, and thus any identification has to be considered preliminaryuntil it is confirmed via other methods. However, four of these were known to be or haveturned out to be reported as Cx43-interacting proteins.
Singh et al. Page 7
J Biol Chem. Author manuscript; available in PMC 2012 November 20.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

As is the case for several highly regulated membrane proteins, Cx43 can interact withmultiple scaffolding and signaling molecules. However, in many cases, these interactionswere detected by immunofluorescence or co-precipitation studies performed in a mannerthat did not distinguish whether the interactions were direct or indirect. Interaction of ZO-1with Cx43 has been clearly documented by multiple laboratories (e.g. see Refs. 4 and 5).Here we present several lines of evidence that indicate that the PDZ2 domain of ZO-2interacts with Cx43: (a) a GST fusion containing the PDZ2 portion of ZO-2 can pull downCx43; (b) an intact C-terminal region of Cx43 is necessary for the interaction; (c) anantibody to ZO-2 can co-precipitate Cx43, and vice versa; (d) Cx43 and ZO-2 can co-localize in the plasma membrane of NRK cells and at intercalated disks in heart cells. As hasbeen previously reported for Cx43 interaction with ZO-1 in cardiac tissue, co-localization ofCx43 and ZO-2 was primarily at the plasma membrane and was extensive but not completein the intercalated disk region (13).
Since ZO-1 and ZO-2 have been shown to interact with each other, it was possible that Cx43might interact with ZO-2 indirectly via ZO-1. Furthermore, given the direct binding of ZO-1to ZO-2, the resolution of light microscopy and even immunoelectron microscopy orfluorescence recovery after photobleach could not definitively distinguish between directand indirect interaction between these molecules and Cx43. Here we present evidence thatZO-2 can interact directly with Cx43, including the following: (a) Cx43 binds to ZO-2 in afar Western overlay assay independent of ZO-1; (b) analysis of confocal images indicatesthat the level of interaction of Cx43 with ZO-1 and ZO-2 changes independently during thecell cycle.
The functional significance of ZO-1 or ZO-2 interaction with Cx43 is not as yet welldefined. PDZ-containing proteins often play an important role in the clustering oftransmembrane receptors (e.g. the spatial organization of ion channels) (11). Most proteinswith PDZ domains are membrane-associated and organize specialized membrane domains atsynapses, junctions, and apical basolateral interface regions (12). In mouse epithelial cellslacking any ZO-1 expression, there is retarded tight junction formation. This phenotype wasrescued by exogenous expression of ZO-1, but not by ZO-2, suggesting that these proteinsare not functionally redundant (41). Dissociation of myocytes from intact ventricle causedan increase in association of ZO-1 and Cx43 (13), and activation of endocytosis increasedCx43-ZO-1 interaction (15). However, more ZO-1-Cx43 interaction occurred as gapjunction assembly increased after a switch from 5 μM to 1.8 mM calcium (42). Furthermore,Src phosphorylation of Cx43 at Tyr265, which negatively regulates gap junctionalcommunication, also appears to negatively regulate the interaction between Cx43 and ZO-1(16, 17).
Previous immunofluorescence studies indicate that Cx43 is almost exclusively in the plasmamembrane during G0, and immunoblot analysis indicates that most of the Cx43 resides inthe Triton X-100-insoluble fraction and migrates as the P2 isoform (43). However, Cx43 isfound both at the plasma membrane and in numerous cytoplasmic vesicles and intracellularcompartments during S phase. The ratio of Triton X-100-insoluble to whole cellular Cx43decreases in S phase cells, and more Cx43 migrates in the P0 and P1 positions (43). Gapjunction assembly also is cell cycle-regulated, with G0 cells being about 50% more efficientat assembly than S phase cells (29). Given our results indicating that Cx43 binds primarilyto ZO-1 present in G0 cells, we propose that Cx43-ZO-1 interaction may enhance theassembly and/or stabilization (as defined by Triton X-100 resistance) of gap junctions. It isconceivable that the ratio of ZO-1 and ZO-2 interacting with Cx43 may affect the stability ofgap junctions and explain why different studies have come to different conclusions about therole of the interaction of Cx43 and ZO-1.
Singh et al. Page 8
J Biol Chem. Author manuscript; available in PMC 2012 November 20.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

AcknowledgmentsDr. Phil Gafken of the Fred Hutchinson Cancer Research Center Proteomics Shared Resource performed the massspectrometry.
References1. White T, Paul D. Annu Rev Physiol. 1999; 61:283–310. [PubMed: 10099690]
2. Sohl G, Willecke K. Cardiovasc Res. 2004; 62:228–232. [PubMed: 15094343]
3. Wei CJ, Xu X, Lo CW. Annu Rev Cell Dev Biol. 2004; 20:811–838. [PubMed: 15473861]
4. Giepmans BN, Moolenaar WH. Curr Biol. 1998; 8:931–934. [PubMed: 9707407]
5. Toyofuku T, Yabuki M, Otsu K, Kuzuya T, Hori M, Tada M. J Biol Chem. 1998; 273:12725–12731. [PubMed: 9582296]
6. Gumbiner B, Lowenkopf T, Apatira D. Proc Natl Acad Sci U S A. 1991; 88:3460–3464. [PubMed:2014265]
7. Jesaitis LA, Goodenough DA. J Cell Biol. 1994; 124:949–961. [PubMed: 8132716]
8. Haskins J, Gu L, Wittchen ES, Hibbard J, Stevenson BR. J Cell Biol. 1998; 141:199–208. [PubMed:9531559]
9. Anderson JM, Van Itallie CM. Am J Physiol. 1995; 269:G467–G475. [PubMed: 7485497]
10. Songyang Z, Fanning AS, Fu C, Xu J, Marfatia SM, Chishti AH, Crompton A, Chan AC,Anderson JM, Cantley LC. Science. 1997; 275:73–77. [PubMed: 8974395]
11. Craven SE, Bredt DS. Cell. 1998; 93:495–498. [PubMed: 9604925]
12. Fanning AS, Anderson JM. J Clin Invest. 1999; 103:767–772. [PubMed: 10079096]
13. Barker RJ, Price RL, Gourdie RG. Circ Res. 2002; 90:317–324. [PubMed: 11861421]
14. Seki A, Duffy HS, Coombs W, Spray DC, Taffet SM, Delmar M. Circ Res. 2004; 95:e22–28.[PubMed: 15284189]
15. Segretain D, Fiorini C, Decrouy X, Defamie N, Prat JR, Pointis G. Biochimie (Paris). 2004;86:241–244.
16. Giepmans BN, Hengeveld T, Postma FR, Moolenaar WH. J Biol Chem. 2001; 276:8544–8549.[PubMed: 11124251]
17. Toyofuku T, Akamatsu Y, Zhang H, Kuzuya T, Tada M, Hori M. J Biol Chem. 2001; 276:1780–1788. [PubMed: 11035005]
18. Sorgen PL, Duffy HS, Sahoo P, Coombs W, Delmar M, Spray DC. J Biol Chem. 2004;279:54695–54701. [PubMed: 15492000]
19. Giepmans BN. Cardiovasc Res. 2004; 62:233–245. [PubMed: 15094344]
20. Butkevich E, Hulsmann S, Wenzel D, Shirao T, Duden R, Majoul I. Curr Biol. 2004; 14:650–658.[PubMed: 15084279]
21. Fu CT, Bechberger JF, Ozog MA, Perbal B, Naus CC. J Biol Chem. 2004; 279:36943–36950.[PubMed: 15213231]
22. Gellhaus A, Dong X, Propson S, Maass K, Klein-Hitpass L, Kibschull M, Traub O, Willecke K,Perbal B, Lye SJ, Winterhager E. J Biol Chem. 2004; 279:36931–36942. [PubMed: 15181016]
23. Kausalya PJ, Reichert M, Hunziker W. FEBS Lett. 2001; 505:92–96. [PubMed: 11557048]
24. Laing JG, Manley-Markowski RN, Koval M, Civitelli R, Steinberg TH. J Biol Chem. 2001;276:23051–23055. [PubMed: 11313345]
25. Nielsen PA, Beahm DL, Giepmans BN, Baruch A, Hall JE, Kumar NM. J Biol Chem. 2002;277:38272–38283. [PubMed: 12154091]
26. Singh D, Lampe PD. Cell Commun Adhes. 2003; 10:215–220. [PubMed: 14681019]
27. Glaunsinger BA, Weiss RS, Lee SS, Javier R. EMBO J. 2001; 20:5578–5586. [PubMed:11598001]
28. Cooper CD, Lampe PD. J Biol Chem. 2002; 277:44962–44968. [PubMed: 12270943]
29. Solan JL, Fry MD, TenBroek EM, Lampe PD. J Cell Sci. 2003; 116:2203–2211. [PubMed:12697837]
Singh et al. Page 9
J Biol Chem. Author manuscript; available in PMC 2012 November 20.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

30. Yates JR III, Eng JK, McCormack AL, Schieltz D. Anal Chem. 1995; 67:1426–1436. [PubMed:7741214]
31. Link AJ, Eng J, Schieltz DM, Carmack E, Mize GJ, Morris DR, Garvik BM, Yates JR III. NatBiotechnol. 1999; 17:676–682. [PubMed: 10404161]
32. Goldberg GS, Moreno AP, Lampe PD. J Biol Chem. 2002; 277:36725–36730. [PubMed:12119284]
33. Kamberov E, Makarova O, Roh M, Liu A, Karnak D, Straight S, Margolis B. J Biol Chem. 2000;275:11425–11431. [PubMed: 10753959]
34. Khokhlatchev A, Xu S, English J, Wu P, Schaefer E, Cobb MH. J Biol Chem. 1997; 272:11057–11062. [PubMed: 9110999]
35. Bowling N, Huang X, Sandusky GE, Fouts RL, Mintze K, Esterman M, Allen PD, Maddi R,McCall E, Vlahos CJ. J Mol Cell Cardiol. 2001; 33:789–798. [PubMed: 11273731]
36. Musil LS, Cunningham BA, Edelman GM, Goodenough DA. J Cell Biol. 1990; 111:2077–2088.[PubMed: 2172261]
37. Musil LS, Goodenough DA. J Cell Biol. 1991; 115:1357–1374. [PubMed: 1659577]
38. Wittchen ES, Haskins J, Stevenson BR. J Biol Chem. 1999; 274:35179–35185. [PubMed:10575001]
39. Lampe PD, Kurata WE, Warn-Cramer BJ, Lau AF. J Cell Sci. 1998; 111:833–841. [PubMed:9472011]
40. Giepmans BN, Verlaan I, Moolenaar WH. Cell Commun Adhes. 2001; 8:219–223. [PubMed:12064592]
41. Umeda K, Matsui T, Nakayama M, Furuse K, Sasaki H, Furuse M, Tsukita S. J Biol Chem. 2004;279:44785–44794. [PubMed: 15292177]
42. Wu JC, Tsai RY, Chung TH. J Cell Biochem. 2003; 88:823–835. [PubMed: 12577316]
43. Solan JL, Lampe PD. Biochim Biophys Acta. 2005; 1711:154–163. [PubMed: 15955300]
Singh et al. Page 10
J Biol Chem. Author manuscript; available in PMC 2012 November 20.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

Fig. 1. Binding of the C-terminal region of Cx43 to ZO-2 and PDZ2 of ZO-2 to Cx43 in GST pulldown assaysA, immunoblot of NRK cell lysate proteins bound to the various GST-Cx43CT constructsprobed using a ZO-2 antibody. GST-Cx43 (residues 236–382 of Cx43) and other GST-Cx43constructs used include CT deletion constructs Δ290 (missing amino acids 280–300), Δ330(missing amino acids 320–340), Δ369 (missing amino acids 364–373), T374 (missingamino acids 375–382), and T379 (missing amino acids 380–382). Whole cell lysate (WC)was run to indicate the migration position of native ZO-2. Whereas ZO-2 was seen to bindthe wild type CT (amino acids 236–382), Δ290, and Δ330 constructs, binding was lost withΔ369, T374, and T379. B, Cx43 interacts with the PDZ2 domain of ZO-2. GST-ZO-2constructs representing the PDZ1, PDZ2, and PDZ3 were used to pull down endogenousCx43 from NRK cell lysates. Immunoblot of NRK cell lysate proteins bound to the variousGST-ZO-2 constructs were probed using a Cx43 antibody. Whole cell lysate was run toindicate native Cx43 migration. A schematic representation of the various domains of ZO-2(PDZ1–3, an Src homology 3 domain, a guanylate-like kinase, and a proline-rich domain) isshown.
Singh et al. Page 11
J Biol Chem. Author manuscript; available in PMC 2012 November 20.
$waterm
ark-text$w
atermark-text
$waterm
ark-text
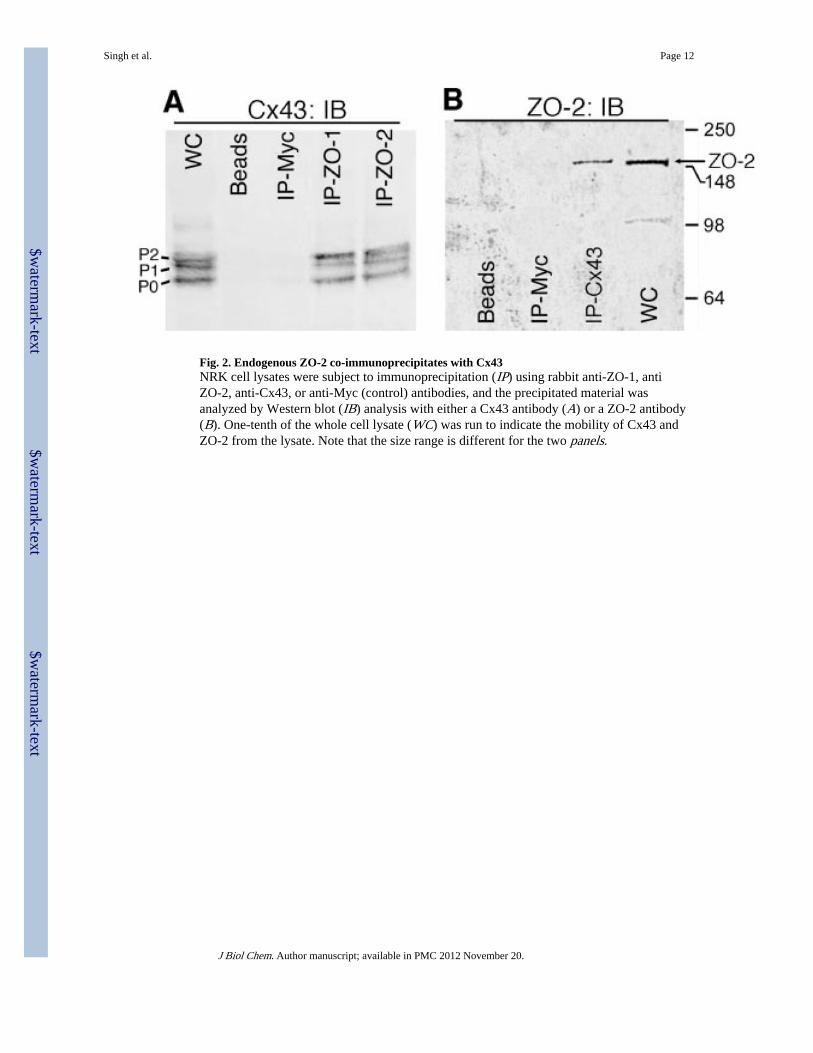
Fig. 2. Endogenous ZO-2 co-immunoprecipitates with Cx43NRK cell lysates were subject to immunoprecipitation (IP) using rabbit anti-ZO-1, antiZO-2, anti-Cx43, or anti-Myc (control) antibodies, and the precipitated material wasanalyzed by Western blot (IB) analysis with either a Cx43 antibody (A) or a ZO-2 antibody(B). One-tenth of the whole cell lysate (WC) was run to indicate the mobility of Cx43 andZO-2 from the lysate. Note that the size range is different for the two panels.
Singh et al. Page 12
J Biol Chem. Author manuscript; available in PMC 2012 November 20.
$waterm
ark-text$w
atermark-text
$waterm
ark-text
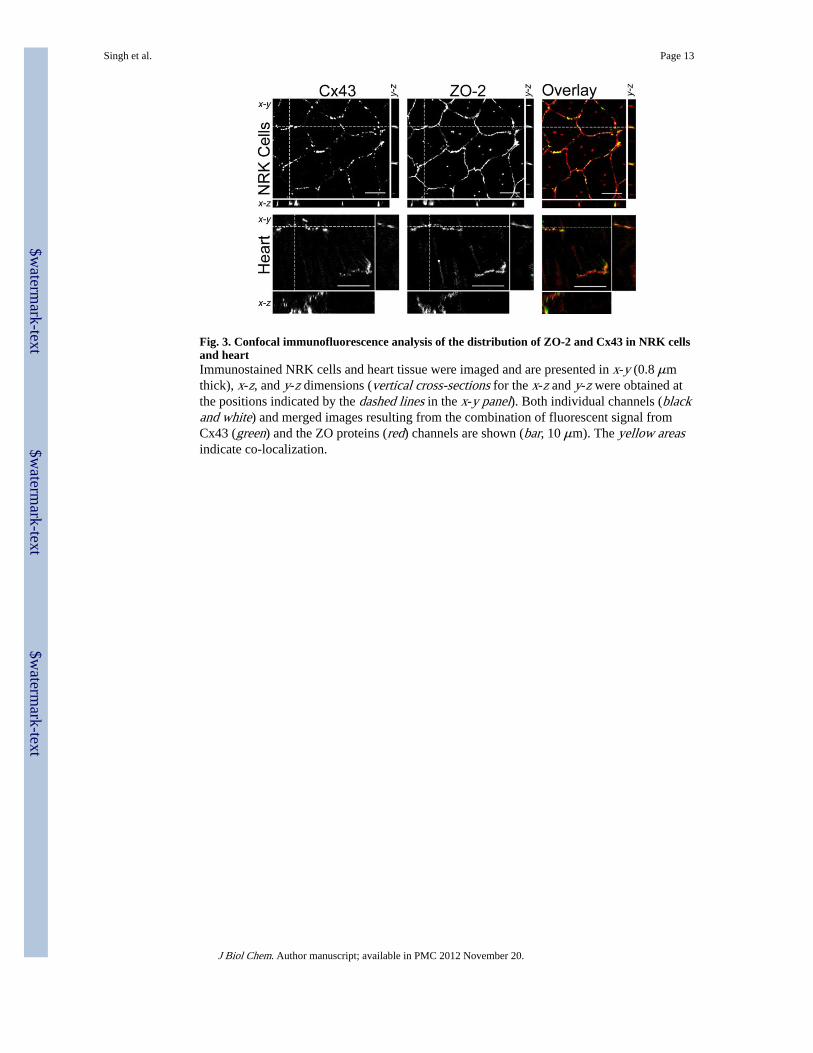
Fig. 3. Confocal immunofluorescence analysis of the distribution of ZO-2 and Cx43 in NRK cellsand heartImmunostained NRK cells and heart tissue were imaged and are presented in x-y (0.8 μmthick), x-z, and y-z dimensions (vertical cross-sections for the x-z and y-z were obtained atthe positions indicated by the dashed lines in the x-y panel). Both individual channels (blackand white) and merged images resulting from the combination of fluorescent signal fromCx43 (green) and the ZO proteins (red) channels are shown (bar, 10 μm). The yellow areasindicate co-localization.
Singh et al. Page 13
J Biol Chem. Author manuscript; available in PMC 2012 November 20.
$waterm
ark-text$w
atermark-text
$waterm
ark-text
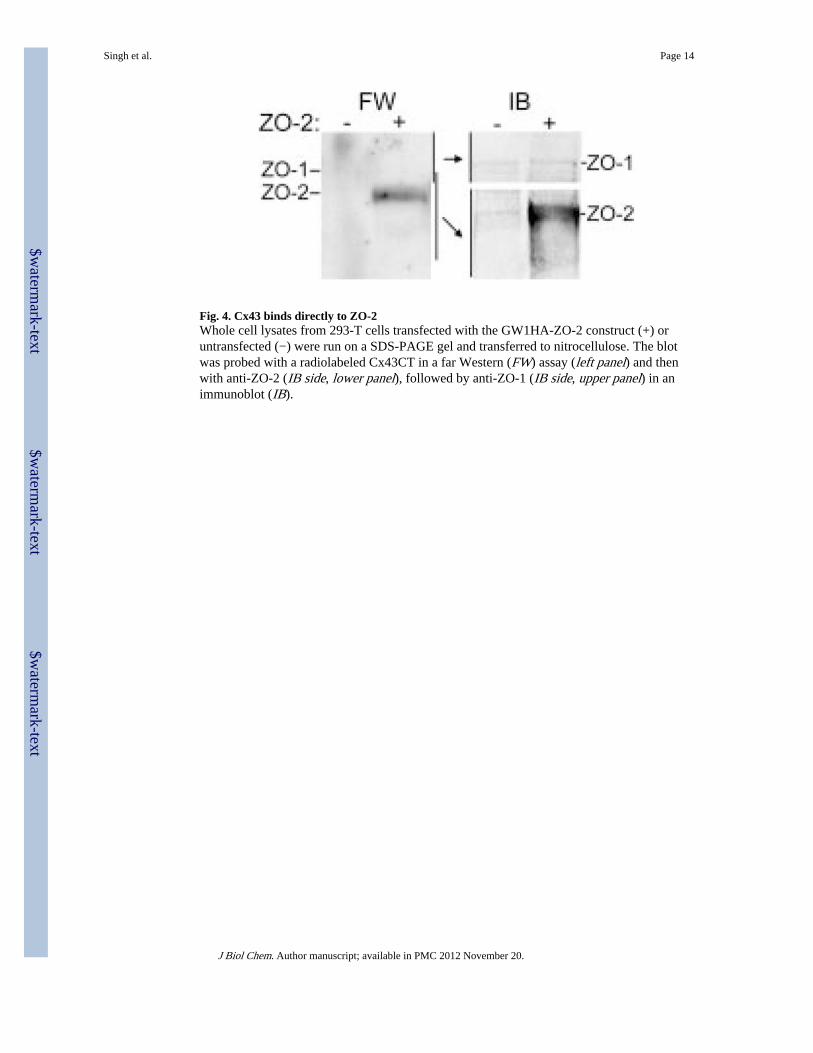
Fig. 4. Cx43 binds directly to ZO-2Whole cell lysates from 293-T cells transfected with the GW1HA-ZO-2 construct (+) oruntransfected (−) were run on a SDS-PAGE gel and transferred to nitrocellulose. The blotwas probed with a radiolabeled Cx43CT in a far Western (FW) assay (left panel) and thenwith anti-ZO-2 (IB side, lower panel), followed by anti-ZO-1 (IB side, upper panel) in animmunoblot (IB).
Singh et al. Page 14
J Biol Chem. Author manuscript; available in PMC 2012 November 20.
$waterm
ark-text$w
atermark-text
$waterm
ark-text
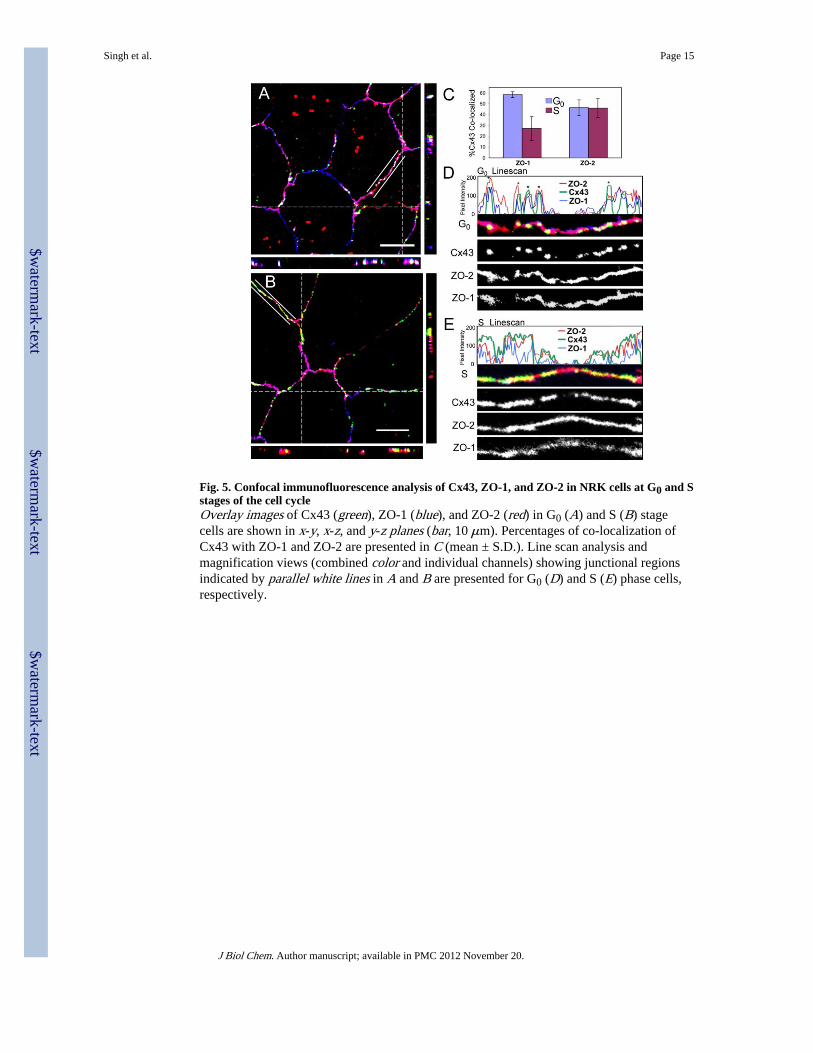
Fig. 5. Confocal immunofluorescence analysis of Cx43, ZO-1, and ZO-2 in NRK cells at G0 and Sstages of the cell cycleOverlay images of Cx43 (green), ZO-1 (blue), and ZO-2 (red) in G0 (A) and S (B) stagecells are shown in x-y, x-z, and y-z planes (bar, 10 μm). Percentages of co-localization ofCx43 with ZO-1 and ZO-2 are presented in C (mean ± S.D.). Line scan analysis andmagnification views (combined color and individual channels) showing junctional regionsindicated by parallel white lines in A and B are presented for G0 (D) and S (E) phase cells,respectively.
Singh et al. Page 15
J Biol Chem. Author manuscript; available in PMC 2012 November 20.
$waterm
ark-text$w
atermark-text
$waterm
ark-text
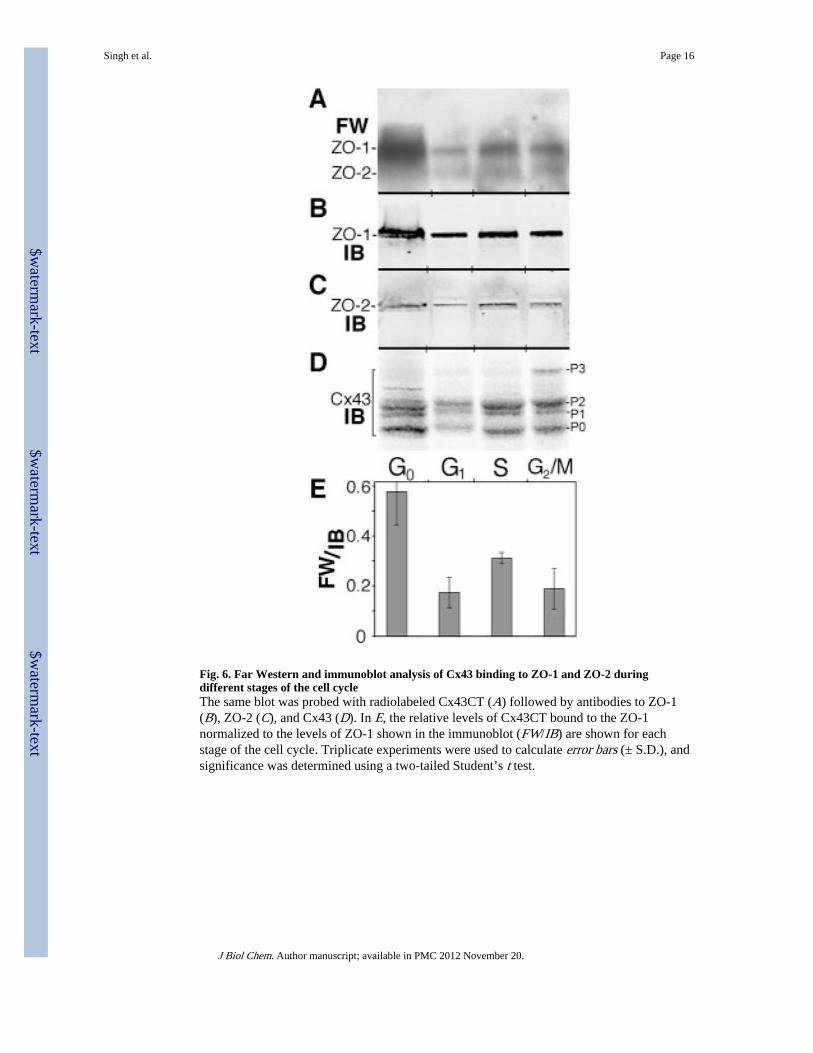
Fig. 6. Far Western and immunoblot analysis of Cx43 binding to ZO-1 and ZO-2 duringdifferent stages of the cell cycleThe same blot was probed with radiolabeled Cx43CT (A) followed by antibodies to ZO-1(B), ZO-2 (C), and Cx43 (D). In E, the relative levels of Cx43CT bound to the ZO-1normalized to the levels of ZO-1 shown in the immunoblot (FW/IB) are shown for eachstage of the cell cycle. Triplicate experiments were used to calculate error bars (± S.D.), andsignificance was determined using a two-tailed Student’s t test.
Singh et al. Page 16
J Biol Chem. Author manuscript; available in PMC 2012 November 20.
$waterm
ark-text$w
atermark-text
$waterm
ark-text
Copyright © 2022 FDOKUMEN




















