
Cyanobacterial Neurotoxin β-N-Methylamino-L-alanine (BMAA) in Shark Fins
Upload
independentCategory
view
1download
0
Interaction network in cyanobacterial nitrogenregulation: PipX, a protein that interacts in a2-oxoglutarate dependent manner with PII and NtcA
Javier Espinosa,1,2 Karl Forchhammer,2
Sergio Burillo1 and Asunción Contreras1*1División de Genética, Universidad de Alicante,Apartado 99, E-03080 Alicante, Spain.2Institut für Mikrobiologie und Molekularbiologie,Universität Giessen, Heinrich-Buff-Ring 26-32, D 35392Giessen, Germany.
Summary
Cyanobacteria perceive nitrogen status by sensingintracellular 2-oxoglutarate levels. The global nitro-gen transcription factor NtcA and the signal transduc-tion protein PII are both involved in 2-oxoglutaratesensing. PII proteins, probably the most conservedsignal transduction proteins in nature, are remarkablefor their ability to interact with very diverse proteintargets in different systems. Despite widespreadefforts to understand nitrogen signalling in cyanobac-teria, the involvement of PII in the regulation of tran-scription activation by NtcA remains enigmatic. Herewe show that PipX, a protein only present in cyano-bacteria, interacts with both PII and NtcA and pro-vides a mechanistic link between these two factors. Avariety of in vivo and in vitro approaches were used tostudy PipX and its interactions with PII and NtcA.2-Oxoglutarate favours complex formation betweenPipX and NtcA, but impairs binding to PII, suggestingthat partner swapping between these nitrogen regu-lators is driven by the 2-oxoglutarate concentration.PipX is required for NtcA-dependent transcriptionalactivation in vivo, thus implying that PipX may func-tion as a prokaryotic transcriptional coactivator.
Introduction
Cyanobacteria are phototrophic organisms that performoxygenic photosynthesis. Autotrophic growth requires theconstant assimilation of ammonium via the GS-GOGATcycle (Muro-Pastor et al., 2005), resulting in consumptionof the carbon-skeleton of 2-oxoglutarate to yield
glutamate. Due to the lack of 2-oxoglutarate dehydroge-nase in cyanobacteria, synthesis of 2-oxoglutarate repre-sents the final step in the oxidative branch of the TCAcycle and directly links 2-oxoglutarate levels to nitrogenassimilation. Therefore, the cellular 2-oxoglutarate con-centration is an excellent indicator of the cell carbon tonitrogen balance.
The PII signal transduction protein, a trimeric protein, isone of the most conserved and widespread nitrogensignal transduction proteins (Arcondeguy et al., 2001;Ninfa and Jiang, 2005). The involvement of the cyanobac-terial PII protein, encoded by the glnB gene, in the controlof the short-term ammonium inhibition of the nitrateuptake in Synechococcus sp. PCC 7942 (hereafter calledSynechococcus) has been known for some time (Leeet al., 2000), although direct protein–protein interactionsbetween transport components and PII have not beenreported yet. In fact, PII targets remained elusive until theN-acetyl-glutamate kinase (NAGK), which catalyses thecommitted step of the cyclic arginine synthesis pathway,was identified in Synechococcus by yeast two-hybrid(Y2H) screening (Burillo et al., 2004; Heinrich et al.,2004). Conservation of the PII–NAGK interaction inorganisms performing oxygenic photosynthesis, predictedby Y2H analysis (Burillo et al., 2004), has been confirmedin plants (Sugiyama et al., 2004; Chen et al., 2006).
Cyanobacterial PII proteins binds 2-oxoglutarate andATP synergistically (Forchhammer and Hedler, 1997) andin most species PII is phosphorylated at a seryl residue(S49) located at the apex of the solvent-exposed T-loop(Forchhammer and Tandeau de Marsac, 1995). Of themodifying/demodifying enzymes only the PII phosphatasePphA, a protein phosphatase of the PP2C family, hasbeen identified in Synechocystis PCC 6803. Importantly,the phosphorylation status of PII correlates with the2-oxoglutarate levels, both being maximal during nitrogenstarvation. PII functions concerning the inhibition of nitratetransport and interaction with NAGK are regulated byeffector binding (Heinrich et al., 2004; Maheswaran et al.,2004; Kobayashi et al., 2005). ATP in concert withelevated 2-oxoglutarate levels or ADP relieves complexformation with NAGK. PII phosphorylation also inhibitsbinding to NAGK. So far, there are not reports of PIItargets requiring the phosphorylated form of PII.
Accepted 8 May, 2006. *For correspondence. E-mail [email protected]; Tel. (+34) 96 590 3957; Fax (+34) 96 590 9569.
Molecular Microbiology (2006) 61(2), 457–469 doi:10.1111/j.1365-2958.2006.05231.xFirst published online 7 June 2006
© 2006 Blackwell Publishing LtdNo claim to original Spanish government works

The transcriptional activator NtcA belongs to the CAP/CRP (the catabolite activator protein or cyclic AMP recep-tor protein) family. NtcA plays a key role in cyanobacterialnitrogen assimilation, being required for the expression ofmultiple genes subjected to ammonium repression(Herrero et al., 2001). Two-oxoglutarate stimulates bindingof NtcA to target sites and transcription activation in vitro(Tanigawa et al., 2002; Vazquez-Bermudez et al., 2002).
There are complex regulatory interactions betweenNtcA and PII. In Synechococcus, NtcA regulates PII at thetranscriptional and post-translational levels (Lee et al.,1999; Sauer et al., 1999). In addition, PII is required tostimulate NtcA activity under conditions of nitrogen depri-vation (Aldehni et al., 2003; Paz-Yepes et al., 2003) andhas an inhibitory role when cells are grown with nitrate asnitrogen source (Aldehni et al., 2003; Aldehni and Forch-hammer, 2006). However, the molecular basis of the PIIregulation of NtcA dependent genes remains elusive.
To contribute to the understanding of nitrogen signallingin cyanobacteria, two-hybrid approaches were used toidentify additional components of the nitrogen interactionregulatory network in Synechococcus and to uncovernovel interacting partners of nitrogen regulators. We showhere that PipX links the key nitrogen regulators PII andNtcA in a protein interaction network in which partnerbinding depends on the 2-oxoglutarate levels. In addition,we provide evidence for a positive regulatory role for PipXin NtcA-dependent activation of transcription.
Results and discussion
PipX interacts specifically with nitrogen regulators PIIand NtcA: two-hybrid analyses
In a previous work, we constructed Y2H libraries fromSynechococcus DNA and screened them withGAL4BD:PII as bait (Burillo et al., 2004). In addition toargB clones, the Sau3A1 libraries produced prey clonesthat were named PipX (PII Interaction Protein X). pipXwas predicted to encode an 89 amino acid protein with apI of 8.97 and no homology to characterized proteindomains. Interestingly, sequences with high homology topipX were found in all cyanobacterial genomes availableand not outside this phylum. An alignment of Synechoc-occus PipX with orthologous proteins is shown in theFig. S1.
To provide an independent assay for PipX–PII interac-tions, we used another in vivo approach, the bacterialadenylate cyclase two-hybrid (BACTH) system, which isbased on the interaction-mediated reconstitution of acyclic AMP signalling cascade in an Escherichia coli cyastrain (Karimova et al., 1998). We generated fusion pro-teins of each of the two fragments (T25 and T18) of thecatalytic domain of Bortedella pertussis adenylate cyclase
to PII and PipX proteins from Synechococcus and PII(GlnB) protein from E. coli, and analysed their ability tocomplement the Cya– phenotype. Both pairs of fusionproteins with PipX and PII from Synechococcus comple-mented the cya strain, thus indicating that PII and PipXretain their affinity in the E. coli system (Fig. S2). Thisinteraction was specific, because none of the PipX fusionproteins interacted with E. coli GlnB. As expected, PIIproteins from both Synechococcus and E. coli interactedwith themselves and to each other, indicating that PIIproteins behave in the BACTH system very much like inthe Y2H system, already shown to reflect relevant inter-actions of PII proteins (Burillo et al., 2004).
Yeast mating of two strains, each one containing one ofthe GAL4 domains fused to a set of proteins of interest,facilitates systematic interaction analyses of relativelylarge number of proteins in all possible combinations,including each protein with itself. Positive clones in theseassays provide direct evidence that each of the two fusionproteins of the diploid is appropriately expressed in yeast,which is important to interpret negative results. With this inmind, we performed Y2H analysis with PipX, heterologousPII proteins and additional Synechococcus proteins fromour laboratory collection. The idea was to provide addi-tional evidence of the specificity of the PII–PipX interac-tion and to use available fusion proteins, constructed withdifferent purposes, to explore additional connections ofPipX with Synechococcus proteins. In all cases, expres-sion of the three reporters (HIS3, ADE2 and lacZ) inY187/PJ696 diploids containing pairs of fusion proteinswas determined and classified according to the strengthof the signals, exactly as described (Burillo et al., 2004).For simplicity, only proteins giving signals with Synecho-coccus PII and/or with PipX have been included inTable 1.
Control Y2H assays with PII fusions from E. coli (GlnBand GlnK) and A. thaliana confirmed that the interactionbetween Synechococcus PII and PipX was specific.Given that PII proteins from plants and cyanobacteriashare common features (see Burillo et al., 2004 for anextended discussion), PII proteins from A. thaliana pro-vided a good control for specificity of binding. The inabilityof PipX to interact in the Y2H system with any of theheterologous PII proteins tested indicated that onlycyanobacterial PII proteins contain determinants for inter-actions with PipX.
Interestingly, the two PipX constructs also interactedwith one of the proteins assayed, the transcriptional regu-lator NtcA. Consistent with this, two-hybrid screening ofSynechococcus libraries using NtcA as bait also identifiedPipX as prey (data not shown). Taken together, the two-hybrid analyses performed here indicate that PipX is not a‘sticky’ protein and that contacts between PipX and thenitrogen regulators PII and NtcA are specific.
458 J. Espinosa, K. Forchhammer, S. Burillo and A. Contreras
© 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 457–469No claim to original Spanish government works

Y2H and BACTH assays both rendered negative resultsfor self interaction of PipX, suggesting that PipX does notoligomerize. It is worth noting that these assays are morelikely to produce false negative results when the interac-tion determinants are localized at the N-terminal part ofthe protein assayed. The lack of signals between NtcAconstructs in the Y2H assay (Table 1) may thus be due toocclusion, by GAL4 domains, of important dimerizationdeterminants from the N-terminus of NtcA.
In vitro properties of PipX and PipX–PII complexes:effect of ATP and 2-oxoglutarate on complex formation
Subsequent biochemical analysis of complex formationbetween PipX and PII and its response to the PII effectormolecules was performed using purified Strep-tagged PII(PII-ST) and H6-PipX protein. To provide in vitro evidencefor the monomeric structure of PipX, suggested by YTHand BACTH analysis, we performed fast protein liquidchromatography (FPLC) gel filtration analysis with purifiedH6-PipX protein (His-tagged PipX). H6-PipX eluted from acalibrated Superdex 200 column in a single peak near thetotal volume, indicating a size of less than 10 kDa, whichis in agreement with the assumed monomeric structure ofthe protein (Fig. 1A and B). Chromatographic mobility ofPipX in the presence of PII revealed a significant shifttowards increased molecular size, which partially over-lapped with the elution of PII (Fig. 1C). Moreover, theelution of PII chromatographed together with PipX wasslightly shifted towards increased size, as compared withthe chromatorgraphic profile of PII alone (Fig. 1A). Theelution shift of PipX towards increased molecular sizeimplies complex formation between PipX and PII, and thepartial overlap is an indication of slow complex dissocia-tion during chromatography.
1000
2
5)
a
Ve/ Vo
100
1, 6
28 30 32 34 36 38 40 4218 20 22 24 26
A
1
10
1,4 1, 1,8 2 2,2 2,4 2,6 2,8 3
1)
2)
3)4)
bc
fraction nr.
Mole
cula
r W
eig
ht (K
Da)
38 39 40 41 42 43 44 45
H6-PipX
B
fraction nr.
C 28 403938373635343332313029PII-STH6-PipX
Fig. 1. Gel filtration analysis of PipX and PipX–PII complexes.A. Determination of the apparent molecular weight of free PipX andPII and of PipX–PII complexes. The Superdex 200 HR 10/30 gelfiltration column was calibrated with the molecular mass standardsfrom Bio-Rad: (1) Thyroglobulin, 670 kDa; (2) Bovinegamma-globulin, 158 kDa; (3) Chicken ovalbumin, 44 kDa; (4)Equine myoglobin, 17 kDa; (5) Vitamin B12, 1,35 kDa. The elutionvolume (Ve) is shown as a fraction of the void volume (Vo). Below,the fraction numbers are given, corresponding to fraction numbersshown in B and C. (a) Elution peak of free PipX cromatographed inabsence of PII; (b) elution peak of PII chromatographed in absenceof PipX; (c) elution peak of PII from a PII/PipX mixture.B and C. Coomassie-stained SDS-polyacrylamide gel of theindicated fractions collected after gel filtration of H6-PipX (B) andSilver stained SDS-polyacrilamide gel of the indicated fractionscollected after gel filtration of the H6-PipX/PII-ST mixture (C).
Table 1. Yeast two-hybrid interactions involving PII and PipX proteins from Synechococcus.
Synechococcus E. coli A. thaliana
PII PipX NAGK NtcA GlnB GlnK PII NP
SynechococcusPII ++ +++ + – + + + –PipX + – – +* – – – –NAGK ++ – + – – – –NtcA – + – – – – – –
E. coliGlnB + – – – + + + –GlnK + – – – + + + –
A. thalianaPII + – – – + + + –NP – – – – – – – –
The GAL4AD and GAL4BD fusion proteins carried by diploids are indicated on left and top of panels respectively. The origin of proteins is indicatedin each case. NP refers to absence of proteins fused to GAL4 domains. Signs indicate levels of expression from GAL1:HIS3, GAL2:ADE2 andGAL1:lacZ reporters in PJ696/Y187 diploids, decreasing from +++ to –, according to previously described conventions (Burillo et al., 2004).Asterisks indicate very weak but still significant signals.
Nitrogen interaction regulatory network 459
© 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 457–469No claim to original Spanish government works
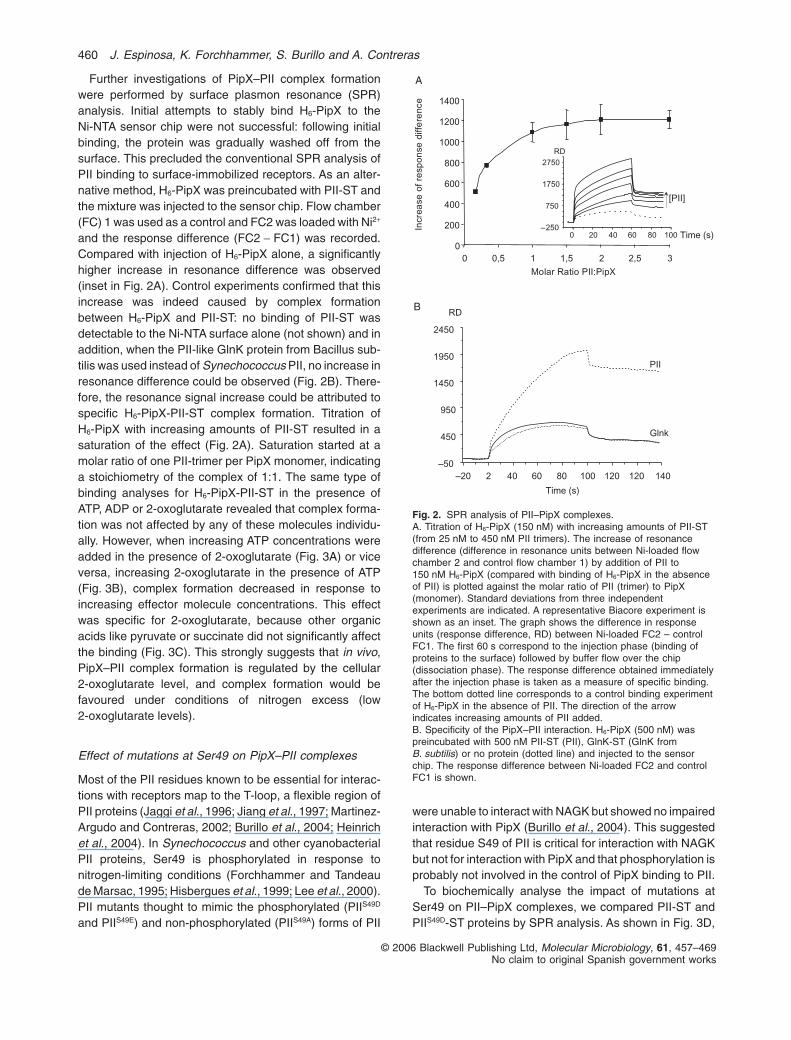
Further investigations of PipX–PII complex formationwere performed by surface plasmon resonance (SPR)analysis. Initial attempts to stably bind H6-PipX to theNi-NTA sensor chip were not successful: following initialbinding, the protein was gradually washed off from thesurface. This precluded the conventional SPR analysis ofPII binding to surface-immobilized receptors. As an alter-native method, H6-PipX was preincubated with PII-ST andthe mixture was injected to the sensor chip. Flow chamber(FC) 1 was used as a control and FC2 was loaded with Ni2+
and the response difference (FC2 - FC1) was recorded.Compared with injection of H6-PipX alone, a significantlyhigher increase in resonance difference was observed(inset in Fig. 2A). Control experiments confirmed that thisincrease was indeed caused by complex formationbetween H6-PipX and PII-ST: no binding of PII-ST wasdetectable to the Ni-NTA surface alone (not shown) and inaddition, when the PII-like GlnK protein from Bacillus sub-tilis was used instead of Synechococcus PII, no increase inresonance difference could be observed (Fig. 2B). There-fore, the resonance signal increase could be attributed tospecific H6-PipX-PII-ST complex formation. Titration ofH6-PipX with increasing amounts of PII-ST resulted in asaturation of the effect (Fig. 2A). Saturation started at amolar ratio of one PII-trimer per PipX monomer, indicatinga stoichiometry of the complex of 1:1. The same type ofbinding analyses for H6-PipX-PII-ST in the presence ofATP, ADP or 2-oxoglutarate revealed that complex forma-tion was not affected by any of these molecules individu-ally. However, when increasing ATP concentrations wereadded in the presence of 2-oxoglutarate (Fig. 3A) or viceversa, increasing 2-oxoglutarate in the presence of ATP(Fig. 3B), complex formation decreased in response toincreasing effector molecule concentrations. This effectwas specific for 2-oxoglutarate, because other organicacids like pyruvate or succinate did not significantly affectthe binding (Fig. 3C). This strongly suggests that in vivo,PipX–PII complex formation is regulated by the cellular2-oxoglutarate level, and complex formation would befavoured under conditions of nitrogen excess (low2-oxoglutarate levels).
Effect of mutations at Ser49 on PipX–PII complexes
Most of the PII residues known to be essential for interac-tions with receptors map to the T-loop, a flexible region ofPII proteins (Jaggi et al., 1996; Jiang et al., 1997; Martinez-Argudo and Contreras, 2002; Burillo et al., 2004; Heinrichet al., 2004). In Synechococcus and other cyanobacterialPII proteins, Ser49 is phosphorylated in response tonitrogen-limiting conditions (Forchhammer and Tandeaude Marsac, 1995; Hisbergues et al., 1999; Lee et al., 2000).PII mutants thought to mimic the phosphorylated (PIIS49D
and PIIS49E) and non-phosphorylated (PIIS49A) forms of PII
were unable to interact with NAGK but showed no impairedinteraction with PipX (Burillo et al., 2004). This suggestedthat residue S49 of PII is critical for interaction with NAGKbut not for interaction with PipX and that phosphorylation isprobably not involved in the control of PipX binding to PII.
To biochemically analyse the impact of mutations atSer49 on PII–PipX complexes, we compared PII-ST andPIIS49D-ST proteins by SPR analysis. As shown in Fig. 3D,
A
[PII]
PII
Glnk
–250
750
1750
2750
0 20 40 60 80 100
RD
Time (s)
Molar Ratio PII:PipX
Incre
ase
of
resp
on
se
diffe
ren
ce
0
200
400
600
800
1000
1200
1400
0 0,5 1 1,5 2 2,5 3
–50
450
950
1450
1950
2450
–20 2 40 60 80 100 120 120 140
RDB
Time (s)
Fig. 2. SPR analysis of PII–PipX complexes.A. Titration of H6-PipX (150 nM) with increasing amounts of PII-ST(from 25 nM to 450 nM PII trimers). The increase of resonancedifference (difference in resonance units between Ni-loaded flowchamber 2 and control flow chamber 1) by addition of PII to150 nM H6-PipX (compared with binding of H6-PipX in the absenceof PII) is plotted against the molar ratio of PII (trimer) to PipX(monomer). Standard deviations from three independentexperiments are indicated. A representative Biacore experiment isshown as an inset. The graph shows the difference in responseunits (response difference, RD) between Ni-loaded FC2 – controlFC1. The first 60 s correspond to the injection phase (binding ofproteins to the surface) followed by buffer flow over the chip(dissociation phase). The response difference obtained immediatelyafter the injection phase is taken as a measure of specific binding.The bottom dotted line corresponds to a control binding experimentof H6-PipX in the absence of PII. The direction of the arrowindicates increasing amounts of PII added.B. Specificity of the PipX–PII interaction. H6-PipX (500 nM) waspreincubated with 500 nM PII-ST (PII), GlnK-ST (GlnK fromB. subtilis) or no protein (dotted line) and injected to the sensorchip. The response difference between Ni-loaded FC2 and controlFC1 is shown.
460 J. Espinosa, K. Forchhammer, S. Burillo and A. Contreras
© 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 457–469No claim to original Spanish government works

no significant differences were observed between the twoPII variants in binding to H6-PipX and in their sensitivity toATP and 2-oxoglutarate. Therefore, the interactionbetween PipX and PIIS49D is also regulated by2-oxoglutarate and ATP. If the PIIS49D proteins can mimicthe phosphorylated state of PII, then our Y2H and SPRdata indicate that interactions with PipX would not dependon the phosphorylation status of PII.
Interactions of cyanobacterial PII proteins with NAGK(Maheswaran et al., 2004) and PamA (Osanai et al.,2005), a protein not conserved within cyanobacteria, arealso controlled by 2-oxoglutarate levels. The ability ofPIIS49A and PIIS49E proteins to bind effectors in vitro (Leeet al., 2000) and the similar behaviour of PII and PIIS49A,PIIS49D and PIIS49E constructs on interactions with PipX(Fig. 3D and Burillo et al., 2004) further support the impor-tance of 2-oxoglutarate for PII functions.
Recent reports suggest that the interactions of PII pro-teins with receptors may involve regions outside theT-loops and rather larger surfaces of PII proteins (Zhanget al., 2004; Zhu et al., 2006). In this context, the smallsize of PipX, the inferred stoichiometry of one PipXmonomer per PII trimer (Fig. 2A) and the insignificantimpact of substitutions at Ser49 on PipX–PII complexes(Fig. 3D and Burillo et al., 2004), suggest that the maindeterminants for interactions are at the main core of thePII trimer, rather than at the T-loops.
In vitro properties of PipX–NtcA complexes: effect ofATP and 2-oxoglutarate on complex formation
To obtain in vitro evidence of the interaction between NtcAand PipX, complex formation was analysed between puri-fied proteins using SPR analysis. H6-NtcA could be stably
–250
750
1750
2750
3750
4750
5750
–10 10 30 50 70 90 110 130 150
RD
Time (s)
A
0
2 [ATP]
–10 10 30 50 70 90 110 130 150
Time (s)
2-OG
Pyruvate
Succinate
–50
450
950
1450
1950
RD
C
–10 10 30 50 70 90 110 130 150
Time (s)
[2-OG]
B
–250
750
1750
2750
3750
4750
RD
5750
0
2
–10 10 30 50 70 90 110 130 150
Time (s)
–50
450
900
1350
1800
RD
PIIS49D
PIIS49D + effectors
D
Fig. 3. SPR analysis of the effect of ATP, organic acids and S49D substitution on PipX–PII complex formation. The sensorgrams show theresponse difference (RD) between Ni-loaded flow chamber and control chamber. The dashed (top) and dotted (bottom) lines correspond,respectively, to controls without effector molecules or without PII-ST protein.A. Effect of various ATP concentrations on the interaction of PII-ST (500 nM) with H6-PipX (500 nM). The direction of the arrow indicatesincreasing ATP concentrations (0, 0.1, 0.5, 1 and 2 mM) in the presence of 1 mM 2-oxoglutarate.B. Effect of different concentrations of 2-oxoglutarate (0, 0.1, 0.5, 1 and 2 mM) in the presence of 1 mM ATP.C. Effect of 1 mM of the indicated organic acids.D. Interaction of PipX–PIIS49D in the presence of 1 mM 2-OG and 1 mM ATP (+ effectors) or in its absence. Details in B–D are as in A.
Nitrogen interaction regulatory network 461
© 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 457–469No claim to original Spanish government works

bound to the Ni-NTA sensor chip and PipX-ST was usedas analyte. Injection of PipX-ST led to a H6-NtcA depen-dent and specific increase in difference resonance units(H6-NtcA covered FC2 – control FC1) during the injectionphase, followed by a rapid dissociation of the complex.Interestingly, the effect of 2-oxoglutarate on NtcA–PipXcomplex formation is opposite to that observed for thePipX–PII complex: 2-oxoglutarate significantly stimulatesNtcA–PipX complex formation (Fig. 4A). In contrast toPipX–PII binding, ATP was not required for the2-oxoglutarate effect (Fig. 4B). Further confirmation of the2-oxoglutarate requirement for PipX–NtcA complex for-mation was obtained by pull-down experiments.Co-elution of PipX-ST and H6-NtcA from Strep-Tactincolumns could only be achieved when 2-oxoglutarate wasincluded in all buffers used for affinity chromatography(Fig. 4C).
The basic properties of PipX–PII and PipX–NtcA com-plexes indicate that binding of PipX to these regulators isinversely affected by 2-oxoglutarate. In particular, theeffectors 2-oxoglutarate and ATP impaired the binding ofPipX to PII, while the binding of PipX to NtcA could only beobserved in the presence of 2-oxoglutarate. The depen-dence of 2-oxoglutarate for alternative binding of PipX toNtcA and PII is very significant, given the importance of2-oxoglutarate as a signal of the carbon/nitrogen balancein cyanobacteria. Concentrations disrupting or stimulat-ing, respectively, PipX–PII and PipX–NtcA complexes(Figs 3 and 4) were similar to those reported to affectPII–NAGK and PII–PamA complexes (Maheswaran et al.,2004; Osanai et al., 2005) or to maximally activate tran-scription initiation by NtcA (Tanigawa et al., 2002) andtherefore the effects of 2-oxoglutarate we observe onPipX complexes are likely to be of physiological signifi-cance, strongly suggesting that PipX plays a role in thecontext of global nitrogen regulation. The estimated2-oxoglutarate concentrations in cyanobacterial cells arein the 0.06–0.44 mM range (Muro-Pastor et al., 2001;Tanigawa et al., 2002), somehow lower than the concen-trations used in most in vitro studies of PII and NtcAproteins.
The Y2H, BACTH and SPR approaches used heredetected interactions of PipX with Synechococcus PII butnot with control PII proteins from other organisms(A. thaliana, E. coli or B. subtilis), indicating that PII deter-minants for interactions with PipX are not conserved inany of the heterologous proteins tested. Consistent withthe idea of coevolution of proteins working on the samepathways (Pellegrini et al., 1999), NtcA and PipX showidentical phylogenetic profiles, that is, both proteins areencoded only by cyanobacterial genomes. While theexclusive and constant presence of pipX and ntcA in allcyanobacterial genomes suggest their adaptative impor-tance, the specific interactions of PipX with nitrogen regu-
lators PII and NtcA strongly suggest that PipX is alsoinvolved in nitrogen signal transduction.
To study the role of PipX in transcriptional regulation,NtcA activity was analysed in null (PipX-) and constitu-
A
–50
0
50
100
150
200
10 50 90 130 170 210 250
RD
[2–OG]
Time (s)
–50
0
50
100
150
200
10 50 90 130 170 210 250
RD
Time (s)
[ATP]
B
C
14.418.425.035.045.0
F WI II
EI II III IV
F WI II
EI II III IV
H6-NtcA
PipX-ST
D
Fig. 4. SPR analysis of PipX–NtcA complex formation.A and B. BIAcore sensorgrams showing the response difference(FC2 FC1) from binding of PipX-ST to H6-NtcA immobilized surface.(A) 2-Oxoglutarate-dependence of PipX binding to NtcA. Thedirection of the arrow indicates increasing 2-oxoglutarateconcentrations (0, 0.5, 2 and 4 mM 2-oxoglutarate) in the presenceof 4 mM ATP. (B) Effect of ATP on the PipX-ST H6-NtcA interaction.Increasing concentrations of ATP (0, 0.5, 2 and 4 mM) were addedin the presence of 4 mM 2-oxoglutarate. The dotted bottom lineshows PipX binding in the absence of effector molecules.C. Co-elution of PipX-ST and H6-NtcA from a gravity flowStrep-Tactin Superflow mini column and separation of samples in15% SDS-PAGE gels stained with silver nitrate. Lane F,flow-through; lane W, wash fractions I–II; lane E, elution fractionsI-IV.D. As in C, except that 2-oxoglutarate was present in all buffers at10 mM. A protein molecular weight marker (Fermentas) is shown atthe left panel.
462 J. Espinosa, K. Forchhammer, S. Burillo and A. Contreras
© 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 457–469No claim to original Spanish government works

tively expressed (PipXc) mutants of pipX containing pro-moter derivatives glnB::luxAB and glnN::luxAB. Thesepromoters are strictly dependent of NtcA but exhibit dif-ferent induction profiles (Aldehni et al., 2003; Aldehni andForchhammer, 2006). Each of the two promoter fusionscarries a single NtcA dependent promoter, which in thecase of the glnN promoter contains an ‘imperfect’ NtcA-binding site that determines a rather weak and slowinduction. Reporter expression was determined by biolu-minescence measurements from cultures grown to mid-exponential phase in either the presence of ammonium ornitrate or after cultures were shifted from ammonium-containing to nitrogen-depleted medium.
As shown in Fig. 5, the nitrogen starvation responsewas severely impaired in PipX- and enhanced in PipXc
strains, indicating that PipX plays a positive regulatoryrole in transcription of the two NtcA dependent promoters
analysed. It is worth noting that despite the drastic effectof pipX inactivation on promoter activity, PipX- culturessubjected to nitrogen starvation showed very low but stillsignificant expression of both promoters. Taking intoaccount that in the NtcA- strain there is no residual acti-vation of glnB::luxAB and glnN::luxAB promoters (Aldehniet al., 2003; Aldehni and Forchhammer, 2006), the resultsindicate that PipX is required for physiological regulationof NtcA promoters, but not essential for transcriptionalactivation.
Interestingly, overexpression of PipX did not increasepromoter activity when the cells were cultured withammonium. In nitrate grown cultures of PipXc, someinduction was observed with glnB but not with the loweraffinity glnN promoter (insets in Fig. 5). The finding thatoverexpression of PipX or NtcA (Luque et al., 2004)cannot override the requirement of ammonium depriva-tion emphasizes the importance of the 2-oxoglutaratesignal in transcriptional regulation of the NtcA regulon.
In summary, under conditions of nitrogen deficiency,corresponding to high intracellular levels of2-oxoglutarate, PipX functions as an activator of NtcA-dependent promoters. Given the 2-oxoglutarate require-ment for both NtcA–PipX complex formation (Fig. 4) andin vitro transcriptional activation by NtcA (Tanigawa et al.,2002), it is tempting to propose that the PipX–NtcAcomplex can activate transcription of NtcA-dependentpromoters.
A model for nitrogen control in cyanobacteria:2-oxoglutarate, PipX and partner swapping
2-Oxoglutarate, the signal of carbon/nitrogen status, playsa key role on regulation of both PII and NtcA proteins,although in the case of NtcA, no direct binding of2-oxoglutarate has yet been shown. Evidence of regula-tory connections between NtcA and PII include the NtcAregulation of PII at the transcriptional and post-translational levels (Lee et al., 1999), the stimulatory roleof PII on NtcA activity in nitrogen starved cells (Aldehniet al., 2003; Paz-Yepes et al., 2003) and its inhibitory rolewhen cells are grown with nitrate (Aldehni et al., 2003;Aldehni and Forchhammer, 2006). Although NtcA and2-oxoglutarate can activate transcription of a target pro-moter in vitro (Tanigawa et al., 2002), a classical inducer-activator complex does not seem enough to explainregulation of NtcA activity in vivo. Importantly, PipX isclearly involved in the induction of NtcA genes by nitrogenstarvation (Fig. 5). To reconcile all these observations, wepropose a regulatory model in which the PipX–NtcAcomplex is part of the mechanism ensuring 2-oxoglutaratedependency for transcriptional activation by NtcA. Thefunction of PipX–PII complexes would be to assist regu-lation of NtcA–PipX interactions.
Lig
ht E
mis
sio
n (
mV
)glnB::luxAB
Time (h)
0.12± 0.0111.4± 5.87
PipX-
0.10± 0.011.1± 0.29
WT 0.12± 0.01.35± 0.12
PipXc
NH4+NO3
-
A
0
200
400
600
800
1000
0 20 40 60
Time (h)
0.4±0.181.7±0.92
PipX-
WT 0.13±0.021.3±0.48
PipXc
NH4+NO3
-
0.2±0.01.13±0.58
B
020
60
100
140
180
Lig
ht E
mis
sio
n (
mV
)
glnB::luxAB
0 2 0 40 60
Fig. 5. Effect of genetic background on glnB::luxAB andglnN::luxAB expression. The time course induction of light emission(mV) of cultures grown with ammonium and shifted to mediumwithout added nitrogen over a time period of 48 h is shown for WT,PipX- and PipXc strains. In each case, the data correspond to arepresentative time course out of four independent experimentsyielding similar results. Wild type, circles; PipX-, squares; PipXc,triangles. Values in the inset tables correspond to the mean valuesfrom four independent experiments (with standard deviation) of thesame three strains grown with ammonium or nitrate tomid-exponential phase. Values in the ammonium columncorrespond to time 0 in the graph.A. glnB::luxAB.B. glnN::luxAB.
Nitrogen interaction regulatory network 463
© 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 457–469No claim to original Spanish government works

According to the model schematically represented inFig. 6, when cells are grown with ammonium the2-oxoglutarate concentration favours binding of PipX toPII. In the absence of 2-oxoglutarate, PipX would not bindto NtcA, that would then be inactive. On the contrary, highlevels of 2-oxoglutarate favours (ternary) complex forma-tion between PipX and NtcA, bringing about transcrip-tional activation of target genes. At intermediate levels of2-OG, PipX could form complexes with either of the twoprotein partners. The inhibitory effect of PII on NtcA acti-vated genes when cells are grown with nitrate wouldreflect competition for PipX binding.
As a working hypothesis, we propose that the functionof PipX–PII complexes is to fine tune regulation of NtcA–PipX interactions. Indirect support for this idea has beenobtained while trying to find evidences of a role of PipX inmodulating PII function. For instance, an up to sixfoldexcess of PipX over PII trimers did not affect PII stimula-tion of NAGK activity (data not shown), indicating thatPipX is probably not involved in regulation of the PIIdependent NAGK activity. Moreover, PipX does not seemto modulate PII accumulation or phosphorylation (data notshown) and the PII–PipX interactions are not affected bysubstitutions at the site of phosphorylation (Fig. 3D;Burillo et al., 2004). The basis of the stimulatory role of PIIon NtcA-activated genes under nitrogen deficiency is stillunknown. One possibility, currently under investigation,could be that PII deficient mutants accumulate less PipX.
Reported examples of bacterial proteins functioning ascoactivators of DNA-binding activators are scarce andthey include the phage T4 protein AsiA, which functions inconcert with the MotA activator to change promoter speci-ficity by ‘s appropriation’ (Browning and Busby, 2004;Hinton et al., 2005), the IpgC chaperone (Mavris et al.,2002) and the dihydroxyacetone kinase subunit DhaL(Bachler et al., 2005). A remarkable feature of the NtcAsystem is that PipX appears to signal the carbon to nitro-gen balance, just as 2-oxoglutarate does. This apparentredundancy may nevertheless be important in vivo toavoid non-appropriated induction of the very large NtcAregulon and/or to efficiently amplify the nitrogen starvationsignal.
Concluding remarks
This work places a previously unknown protein, PipX, atthe centre of the nitrogen signalling network of cyanobac-teria, providing a new element to understand previouslyreported genetic connections between glnB and ntcA. Wehave shown that (i) the PipX protein can form complexeswith the key nitrogen regulators PII and NtcA, (ii) the2-oxoglutarate concentration differentially affects PipX–PII and PipX–NtcA complex formation, (iii) the pipX geneencodes a positive regulator of the NtcA-dependent pro-moters glnB and glnN, and (iv) the regulatory activity ofPipX is also dependent of the 2-oxoglutarate signal. Theability of PII to bind to PipX under non-inducing conditionsand the sensitivity of PII–PipX complexes to2-oxogulatarate may also play a role in the regulation ofNtcA activity by PipX.
Experimental procedures
Strains, plasmids and growth conditions
Strains and plasmids used in this work are listed in Tables 2and 3 respectively. Oligonucleotides used in this work arelisted in Table S1. Cloning procedures were carried out withE. coli DH5a, using standard techniques. Yeast proceduresand Y2H assays were as described (Burillo et al., 2004).BACTH assays were as described (Espinosa et al., 2006).Synechococcus growth conditions and luciferase assayswere as described (Aldehni et al., 2003), except that lightemission by bioluminescence was recorded in a LKB Wallac1250 luminometer as mV (milli-Volts) units.
Construction of plasmids
To construct pUAGC5 and pUAGC7 ntcA sequences wereamplified with primers NTCA-1F and NTCA-1R, cut withEcoRI and SalI and cloned into pGAD424(+2) andpGBT9(+2) respectively. To obtain pUAGC73, pipX was poly-merase chain reaction (PCR)-amplified with primers PipX-pTRC-1F and PipX-pTRC-1R using pUAGC71 as a template
Transcription OFF
Transcription ON
[2-OG]
Transcription ON/OFF
NH4+
N03-
-N
Fig. 6. Schematic model of the proposed mode of action of PipX.Under low 2-oxoglutarate levels (i.e. ammonium sufficiency) NtcA(white square pairs) is inactive and PipX (black octagons) is boundto PII (trimeric white spheres). In nitrate-grown cultures, part of thePII pool is phosphorylated (black dots) and the moderated levels ofintracellular 2-oxoglutarate allow PipX swapping between PII andNtcA, leading to NtcA activation (grey square pairs). In theseconditions different conformations of NtcA (active/inactive) and ofPII (phosphorylated/non-phosphorylated) coexist. High2-oxoglutarate levels (i.e. nitrogen starvation) further increasesPipX binding to NtcA, which is now fully activated.
464 J. Espinosa, K. Forchhammer, S. Burillo and A. Contreras
© 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 457–469No claim to original Spanish government works

and the product was cloned into NcoI and BamHI sites ofpTRC99A. To construct plasmid pUAGC470, pipX sequenceswere PCR-amplified with primers PipX-ST-For and PipX-ST-Rev, cut with BsaI and cloned into pASK-IBA3 vector. Toconstruct plasmid pIBA-PIIS49D, glnB sequence was PCR-amplified from pPM316 using primers glnBStrep-for andglnBStrep-rev, cut with BsaI and cloned into pASK-IBA3. Toobtain plasmids pUAGC441 and pUAGC442, glnBsequences were PCR-amplified with primers PII-BTH-F andPII-BTH-R, cut with BamHI and KpnI and cloned into pT25and pUT18c respectively. To obtain plasmids pUAGC443 andpUAGC444, pipX sequences were amplified with primersPipX-BTH-F and PipX-BTH-R, cut with BamHI and KpnI andcloned into pT25 and pUT18c respectively. glnB sequencefrom E. coli was PCR-amplified with primers GlnB-BTH-F andGlnB-BTH-R, cut with BamHI and KpnI and cloned into pT25and pUT18c giving plasmids pUAG653 and pUAG652respectively. To obtain pET15b-NtcA, ntcA sequences werePCR-amplified with NtcA-ET15F and NtcA-ET15R, the result-ing fragment was cloned into pGEM-T. From the resultingplasmid pGEM-TNtcA, the XhoI–NdeI fragment containingntcA was cloned into the NdeI–XhoI site of pET-15b, givingpET15b-NtcA.
Cloning genomic sequences adjacent to pipX
To clone genomic sequences upstream of pipX, Synechoc-occus DNA was digested separately with EcoRV, RsaI, ScaIand SspI, ligated to a 23 bp linker formed by oligonucleotidesTRANSPO-1 and TRANSPO-2 linker, cut with SalI, and theneach of the four fragment collections were amplified withTRANSPO-1 and PIP2-2R. The longest PCR product(1.7 kb), obtained from the RsaI digest was sequenced,assembled with the pipX sequence, confirmed by PCR usingoligonucleotides PIPX-126-F and PIP2-2R and thensequenced with PIPX-126-F. This sequence was sent toGeneBank with Accession number AY301618. To clonesequences downstream of pipX, the same strategy was usedwith enzymes EcoRV, RsaI, SmaI, SspI, HincII and PvuII andprimers PIPX-4F and TRANSPO-1. RsaI and HincII gavefragments of approximately 400 bp that were assembled tosequences already available in GenBank. Assembledsequences were confirmed by PCR with oligonucleotidesLCY-PIPX-1F and PIPX-5R and sequenced with LCY-PIPX-1F.
Generation of mutant and reporter strainsof Synechococcus
After amplification with primers LCY-PIPX-1F and PIPX-5R,the 2.1 kb genomic fragment expanding SynechococcuspipX was digested with XhoI and HindIII, and the resultingfragment cloned into pBluescript SK(+), giving plasmidpUAGC59. The 1.3 kb kanamycin-resistance cassette CK1was extracted from pRL161 with HincII and cloned intoplasmid pUAGC59 previously purified from E. coli GM119,cut with ClaI and SalI and klenow-treated. The resultingplasmid, pUAGC59.1, carries the CK1 cassette in antisenseto pipX with an internal in frame deletion. As Northernanalyses suggest that pipX is monocistronic (data notTa
ble
2.S
trai
nsus
edin
this
wor
k.
Str
ain
Gen
otyp
eor
rele
vant
char
acte
ristic
sS
ourc
eor
refe
renc
e
E.c
oliD
H5a
F–
f80
dlac
ZDM
15D(
lacZ
YA
-arg
F)U
169
endA
1re
cA1
hsdR
17(r
K–
mK
+ )de
oRth
i-1su
pE44
gyrA
96re
lA1
l–H
anah
an(1
985)
E.c
oliH
B10
1F
–D(
gpt-
proA
)62
leuB
6gl
nV44
ara-
14ga
lK2
lacY
1D(
mcr
C-m
rr)
rpsL
20(S
trr )
xyl-5
mtl-
1re
cA1
Sam
broo
ket
al.
(198
9)E
.col
iDH
M1
F– g
lnV
44(A
S)
recA
1en
dAgy
rA46
thi-1
hsdR
17sp
oT1
frbD
1cy
a-85
4K
arim
ova
etal
.(2
005)
E.c
oliB
L21
F–,
ompT
,hs
dS(r
B– ,
mB
– ),
gal,
dcm
Phi
llips
etal
.(1
984)
E.c
oliX
L1B
lueR
MF
�en
dA1,
gyrA
96,
hsdS
20(r
K–
mK
+ ),
lac,
recA
I,re
lAI,
supE
44,
thi-1
,F
�[pr
oAB
,la
cIq Z
DM15
,T
n10]
Str
atag
ene
S.c
erev
isia
eY
187
MA
Ta
ura3
-52
his3
-200
ade2
-101
trp1
-901
leu2
-3,
112
gal4
Dm
et– g
al80
DU
RA
::GA
L1U
AS-G
AL1
TATA
-lacZ
Har
per
etal
.(1
993)
S.c
erev
isia
eP
J696
MA
Taad
e2D
trp1
-901
leu2
-311
2ur
a3-5
2hi
s3-2
00cy
hRca
nRga
l4D
gal8
0Dm
et2– G
AL2
::AD
E2
GA
L1::H
IS3
GA
L7:la
cZJa
mes
etal
.(1
996)
Syn
echo
cocc
ussp
.P
CC
7942
Pas
teur
cultu
reco
llect
ion
Syn
echo
cocc
usS
A59
1K
mR
deriv
ativ
eof
stra
inP
CC
7942
;ka
nam
ycin
resi
stan
ceca
rtrid
gein
sert
edin
toth
epi
pXge
neT
his
wor
kS
ynec
hoco
ccus
SA
410
Km
Rde
rivat
ive
ofst
rain
PC
C79
42;
kana
myc
inre
sist
ance
cart
ridge
inse
rted
upst
ream
ofth
epi
pXge
neT
his
wor
kW
T-FA
M2
Cm
Rde
rivat
ive
ofst
rain
PC
C79
42;
glnB
::lux
AB
inse
rted
into
chro
mos
ome
neut
rals
iteT
his
wor
kW
T-FA
M84
WC
mR
deriv
ativ
eof
stra
inP
CC
7942
;gl
nN::l
uxA
Bin
sert
edin
toch
rom
osom
ene
utra
lsite
Thi
sw
ork
SA
591-
FAM
2C
mR
deriv
ativ
eof
stra
inS
A59
1;gl
nB::l
uxA
Bin
sert
edin
toch
rom
osom
ene
utra
lsite
Thi
sw
ork
SA
591-
FAM
84W
Cm
Rde
rivat
ive
ofst
rain
SA
591;
glnN
::lux
AB
inse
rted
into
chro
mos
ome
neut
rals
iteT
his
wor
kS
A41
0-FA
M2
Cm
Rde
rivat
ive
ofst
rain
SA
410;
glnB
::lux
AB
inse
rted
into
chro
mos
ome
neut
rals
iteT
his
wor
kS
A41
0-FA
M84
WC
mR
deriv
ativ
eof
stra
inS
A41
0;gl
nN::l
uxA
Bin
sert
edin
toch
rom
osom
ene
utra
lsite
Thi
sw
ork
Nitrogen interaction regulatory network 465
© 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 457–469No claim to original Spanish government works

shown), polar effects are not expected. In the same way,the CK1 cassette was extracted from pRL161 with HincIIand cloned into the AfeI site of pUAGC59, located 72 bpupstream of the pipX reading frame, giving plasmidpUAGC410. In this construct the pipX gene would be con-trolled from the strong CK1 promoter. Transformation ofSynechococcus cells with plasmids pUAGC59.1 orpUAGC410 generated strains SA591 and SA410 respec-tively. Verification of the constructs was carried out by PCRamplification of the engineered regions (Fig. S3). pRK2013(helper) and pRL443 (conjugative) plasmids were used totransfer pFAM2 and pFAM84W into Synechococcus deriva-
tives, generating strains SA591-FAM2, SA591-FAM84W,SA410-FAM2 and SA490-FAM84W.
Gel filtration experiments
One hundred and fifty micrograms of H6-PipX protein or100 mg of PII-ST were incubated in 100 ml of gel filtrationbuffer (10 mM sodium phosphate, pH 7.0, 5 mM MgCl2,0.5 mM EDTA, 1 mM DTT and 400 mM NaCl) for 10 min andpassed through a Superdex 200 HR 10/30 column. A Bio-Logic HR chromatography system (Bio-Rad Laboratories)
Table 3. Plasmids used in this work.
Plasmid Relevant characteristics Source or reference
pGAD424 Ampr, LEU2, GAL4(768-881) AD Bartel et al. (1993)pGAD424(+2) As pGAD424 with a different frame (+2) Roder et al. (1996)pGBT9 Ampr, TRP1, GAL4(1-147) BD Chien et al. (1991)pGBT9(+2) As pGBT9 with a different frame (+2) Roder et al. (1996)pUAG161 GAL4AD:GlnB (E. coli) Martinez-Argudo and Contreras (2002)pUAG162 GAL4BD:GlnB (E. coli) Martinez-Argudo and Contreras (2002)pUAG181 GAL4AD:GlnK (E. coli) Salinas and Contreras (2003)pUAG182 GAL4BD:GlnK (E. coli) Salinas and Contreras (2003)pUAGC11 GAL4AD:PII Burillo et al. (2004)pUAGC12 GAL4BD:PII Burillo et al. (2004)pUAGC61 GAL4AD:NAGK Burillo et al. (2004)pUAGC62 GAL4BD:NAGK Burillo et al. (2004)pUAGC119 GAL4BD:PII (A. thaliana) Burillo et al. (2004)pUAGC120 GAL4BD:PII (A. thaliana) Burillo et al. (2004)pUAGC71 GAL4AD:PipX Burillo et al. (2004)pUAGC72 GAL4BD:PipX Burillo et al. (2004)pT25 Cmr, T25 fragment of Bordetella pertussis CyaA (1-224) Karimova et al. (1998)pUT18C Ampr, T18 fragment of Bordetella pertussis CyaA (225-399) Karimova et al. (1998)pRL161 Ampr Kmr containing CK1 cassette Elhai and Wolk (1988)pAM1580 Ampr Cmr derivative of pAM1573 plasmid carrying luxAB reporter genes. Andersson et al. (2000)pFAM2 pAM1580 derivative with glnB::luxAB fusion Aldehni et al. (2003)pFAM1 pAM1580 derivative with glnB::luxAB fusion Aldehni et al. (2003)pFAM84W pAM1580 derivative with glnN::luxAB fusion Aldehni and Forchhammer (2006)pRK2013 Kmr Helper plasmid Figurski and Helinski (1979)pRL443 Ampr Conjugal plasmid Elhai et al. (1997)pPM316 pBR322 derivative containing mutated Synechococcus glnB gene coding for
PIIS49D proteinLee et al. (2000)
pIBA-GlnK pASK-IBA3:GlnK (B. subtilis) Henrich et al. (manuscript in preparation)pIBA-PII1 pASK-IBA3:PII Heinrich et al. (2004)pASK-IBA3 Ampr Streptag cloning vector IBA GmbHpGEM-T Ampr cloning vector PromegapET15b Ampr Histag cloning vector NovagenpTrc99A Ampr cloning vector StratagenepBluescriptSK (+) Ampr cloning vector StratagenepUAGC5 GAL4AD:NtcA This workpUAGC7 GAL4BD:NtcA This workpUAGC59 pBluescriptSK (+) derivative containing the 1,8 Kb Synechococcus
pipX genomic regionThis work
pUAGC59.1 pUAGC59 derivative with CK1 into pipX This workpUAGC410 pUAGC59 derivative with CK1 inserted upstream pipX This workpUAGC470 pASK-IBA3:PipX This workpUAGC441 T25:PII This workpUAGC442 T18:PII This workpUAGC443 T25:PipX This workpUAGC444 T18:PipX This workpUAGC73 pTrc99A:H6-PipX This workpUAG653 T25:GlnB (E. coli) This workpUAG652 T18:GlnB (E. coli) This workpGEM-TNtcA pGEM-T derivative carrying ntcA This workpET15b-NtcA pET15b derivative with H6-NtcA This workpIBA-PIIS49D pASK-IBA3:PIIS49D This work
466 J. Espinosa, K. Forchhammer, S. Burillo and A. Contreras
© 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 457–469No claim to original Spanish government works

was used to develop the column at a flow rate of 0.5 ml min-1,and 0.5 ml fractions were collected. For analysis of PipX–PIIcomplexes, 50 mg of H6-PipX and 100 mg of PII-ST proteinswere incubated in 100 ml of gel filtration buffer and after10 min applied to the column as described above.
Biacore SPR detection
Surface plasmon resonance experiments were performedusing a BIAcore X biosensor system (Biacore AB, Uppsala,Sweden). To load Ni2+ on the NTA surface of FC, 10 ml of a5 mM NiSO4 solution were injected. For regeneration of thesurface, first 50 ml 0.4 M EDTA was injected, followed bywashing with HBS buffer (10 mM HEPES, 150 mM NaCl,0.005% Nonidet P-40, pH 7.5) and finally Ni2+ was loadedagain as described above. To immobilize H6-NtcA on theNTA-biosensor surface, 125 nM H6-NtcA dimers in HBSbuffer was injected into the Ni2+ loaded FC2 in a volume of60 ml, resulting in an increase of resonance units of 1000. Noprotein was injected into Ni2+ loaded FC1. Experiments wereperformed at 25°C at a flow rate of 15 ml min-1. To analyse thebinding of PipX-ST on H6-NtcA loaded surface, PipX-ST(500 nM) was injected as analyte in a volume of 60 ml anddifference resonance spectra (FC2 – FC1) were recorded.Subsequent injections of analyte were carried out after com-plete dissociation of PipX-ST from the H6-NtcA surface. Tostudy the effect of metabolites on PipX–NtcA interaction,PipX-ST was incubated for 5 min on ice with effector mol-ecules as indicated, and then injected to the H6-NtcA loadedsurface. Injection of PipX-ST to empty Ni2+ NTA surface didnot increase resonance units. To analyse the binding ofPII-ST protein to His6-PipX, the protocol was modified,because H6-PipX did not stably bind to the Ni-NTA surface.PII-ST and H6-PipX were mixed in a final volume of 40 ml ofHBS buffer, incubated for 5 min on ice and finally injected tothe sensor chip, in which FC2 was Ni2+-loaded whereas FC1was not. Difference resonance spectra (FC2 – FC1) wererecorded. Experiments were performed at 25°C at a flow rateof 15 ml min-1. To analyse the effect of small molecules onPII-ST binding to H6-PipX, the two proteins were incubated for5 min on ice with the various effector molecules as indicatedand then injected to the sensor chip. To analyse the effect ofATP, equimolar mixtures of ATP and MgCl2 were used. Allexperiments, performed with Ni2+-loaded FC2, taking as ref-erence FC1, were repeated with Ni2+-loaded FC1 taking asreference FC2, yielding identical results.
Streptag pulldown assays of PipX–NtcA complexes
PipX-ST protein (25 mg) was added to H6-NtcA (10 mg) in100 ml of equilibration buffer (100 mM Tris-HCl, pH 7.8,150 mM NaCl, 1 mM EDTA, 1 mM MgCl2. After 10 min on ice,the mixture was loaded on a gravity flow Strep-Tactin Super-flow mini column (0.2 ml bed volume; IBA, Göttingen). Theflowthrough was collected and unbound protein discarded byfive consecutive washes with equilibration buffer. Elution ofStrep-Tactin-bound material was performed by the addition of6 ¥ 0.1 ml of elution buffer containing equilibration buffer with2.5 mM desthiobiotin. To analyse the effect of 2-oxoglutaratein complex formation, the metabolite was included in allbuffers to a final concentration of 10 mM.
Protein purification
PII-ST, PIIS49D-ST and PipX-ST proteins were purified as pre-viously described (Heinrich et al., 2004). BL21 (DE3) carryingpUAGC73 plasmid was grown to an absorbance (OD600) ofabout 0.8. Overexpression of H6-PipX was initiated by theaddition of 1 mM isopropyl 1-thio-b-D-galactopyranoside(IPTG). After 4 h of incubation at 26°C cells were harvestedby centrifugation, washed with Tris-Cl, pH 7.4, 100 mM NaCland resuspended in lysis buffer containing 20 mM Tris-Cl,pH 7.4, 50 mM KCl, 400 mM NaCl, 5 mM MgCl2, 0.5 mMEDTA, 1 mM DTT, 1 mM benzamidine, and 0.2 mM phenyl-methylsulphonyl fluoride. Cells were disrupted by sonicationand unbroken cells and cell debris removed by two consecu-tive centrifugations (10 min at 10 000 g and 30 min at30 000 g) at 4°C. The supernatant was loaded on a 5 ml HisSelect Cartridge (Sigma H-6662). The column was washedwith 20 mM Tris-C,l pH 8.1, 300 mM NaCl, 20 mM imidazole,and the bound His-tagged protein was then eluted withelution buffer (20 mM Tris-Cl, pH 8.1, 400 mM NaCl, 250 mMimidazole). The elution was collected in 2 ml fractions, andthe purity of the samples checked by SDS-PAGE. Fractionscontaining pure PipX were pooled and dialysed against abuffer consisting of 20 mM Tris-Cl, pH 7.4, 400 mM NaCl,5 mM MgCl2, 0.5 mM EDTA, and 50% (v/v) glycerol andstored at -20°C until use. The same steps were used to purifyH6-NtcA from E. coli BL21 carrying plasmid pET15b-NtcA.The concentrations of the purified proteins were calculatedfrom their absorbance at 280 nm using extinction coefficients,according to their amino acid composition, of 5120 M-1 cm-1
for H6-PipX, 8250 M-1 cm-1 for PII-ST proteins and11523 M-1 cm-1 for PipX-ST.
Acknowledgements
We thank M.L. Cayuela, I. Fuentes, A. Heinrich, I. Luque, M.Maheswaran and P. Salinas for contributions to this work andD. Ladant for BACTH strains and plasmids. We are grateful tothe Ministerio de Educación y Ciencia for Grant BMC2002-01156 and a predoctoral fellowship to J.E., to the GeneralitatValenciana for Grant GV04B-525 and a predoctoral fellow-ship to S.B., to the Deutsche Forschungsgemeinschaft forGrant Fo195/4 and to the Fonds der Chemischen Industriefor providing the BiaCore facilities.
References
Aldehni, M.F., and Forchhammer, K. (2006) Analysis of anon-canonical NtcA-dependent promoter in Synechococ-cus elongatus and its regulation by NtcA and PII. ArchMicrobiol 184: 378–386.
Aldehni, M.F., Sauer, J., Spielhaupter, C., Schmid, R., andForchhammer, K. (2003) Signal transduction protein P(II) isrequired for NtcA-regulated gene expression during nitro-gen deprivation in the cyanobacterium Synechococcuselongatus strain PCC 7942. J Bacteriol 185: 2582–2591.
Andersson, C.R., Tsinoremas, N.F., Shelton, J., Lebedeva,N.V., Yarrow, J., Min, H., and Golden, S.S. (2000) Appli-cation of bioluminescence to the study of circadian rhythmsin cyanobacteria. Methods Enzymol 305: 527–542.
Nitrogen interaction regulatory network 467
© 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 457–469No claim to original Spanish government works

Arcondeguy, T., Jack, R., and Merrick, M. (2001) P(II) signaltransduction proteins, pivotal players in microbial nitrogencontrol. Microbiol Mol Biol Rev 65: 80–105.
Bachler, C., Schneider, P., Bahler, P., Lustig, A., and Erni, B.(2005) Escherichia coli dihydroxyacetone kinase controlsgene expression by binding to transcription factor DhaR.EMBO J 24: 283–293.
Bartel, P.C.T., Chien, R., Sternglanz and Fields (1993) Usingthe two-hybrid system to detect protein–proteininteractions. In Cellular Interactions in Development: APractical Approach. Hartley D.A. (ed.). Oxford: Oxford Uni-versity Press, pp. 153–179.
Browning, D.F., and Busby, S.J. (2004) The regulation ofbacterial transcription initiation. Nat Rev Microbiol 2:57–65.
Burillo, S., Luque, I., Fuentes, I., and Contreras, A. (2004)Interactions between the nitrogen signal transductionprotein PII and N-acetyl glutamate kinase in organisms thatperform oxygenic photosynthesis. J Bacteriol 186: 3346–3354.
Chen, Y.M., Ferrar, T.S., Lohmeir-Vogel, E., Morrice, N.,Mizuno, Y., Berenger, B., et al. (2006) The PII signal trans-duction protein of Arabidopsis thaliana forms an arginine-regulated complex with plastid N-acetyl glutamate kinase.J Biol Chem 281: 5726–5733.
Chien, C.T., Bartel, P.L., Sternglanz, R., and Fields, S. (1991)The two-hybrid system: a method to identify and clonegenes for proteins that interact with a protein of interest.Proc Natl Acad Sci USA 88: 9578–9582.
Elhai, J., and Wolk, C.P. (1988) A versatile class of positive-selection vectors based on the nonviability of palindrome-containing plasmids that allows cloning into longpolylinkers. Gene 68: 119–138.
Elhai, J., Vepritskiy, A., Muro-Pastor, A.M., Flores, E., andWolk, C.P. (1997) Reduction of conjugal transfer efficiencyby three restriction activities of Anabaena sp. strain PCC7120. J Bacteriol 179: 1998–2005.
Espinosa, J., Fuentes, I., Burillo, S., Rodriguez-Mateos, F.,and Contreras, A. (2006) SipA, a novel type of protein fromSynechococcus sp. PCC 7942, binds to the kinase domainof NblS. FEMS Microbiol Lett 254: 41–47.
Figurski, D.H., and Helinski, D.R. (1979) Replication of anorigin-containing derivative of plasmid RK2 dependent on aplasmid function provided in trans. Proc Natl Acad Sci USA76: 1648–1652.
Forchhammer, K., and Hedler, A. (1997) Phosphoprotein PIIfrom cyanobacteria – analysis of functional conservationwith the PII signal-transduction protein from Escherichiacoli. Eur J Biochem 244: 869–875.
Forchhammer, K., and Tandeau de Marsac, N. (1995) Phos-phorylation of the PII protein (glnB gene product) in thecyanobacterium Synechococcus sp. strain PCC 7942:analysis of in vitro kinase activity. J Bacteriol 177: 5812–5817.
Hanahan, D. (1985) Techniques for transformation ofEscherichia coli. In DNA Cloning. Glover D.E. (ed.). Oxford:IRL Press, pp. 109–135.
Harper, J.W., Adami, G.R., Wei, N., Keyomarsi, K., andElledge, S.J. (1993) The p21 Cdk-interacting protein Cip1is a potent inhibitor of G1 cyclin-dependent kinases. Cell75: 805–816.
Heinrich, A., Maheswaran, M., Ruppert, U., and Forchham-mer, K. (2004) The Synechococcus elongatus P signaltransduction protein controls arginine synthesis bycomplex formation with N-acetyl-L-glutamate kinase. MolMicrobiol 52: 1303–1314.
Herrero, A., Muro-Pastor, A.M., and Flores, E. (2001) Nitro-gen control in cyanobacteria. J Bacteriol 183: 411–425.
Hinton, D.M., Pande, S., Wais, N., Johnson, X.B., Vuthoori,M., Makela, A., and Hook-Barnard, I. (2005) Transcriptionaltakeover by sigma appropriation: remodelling of thesigma70 subunit of Escherichia coli RNA polymerase bythe bacteriophage T4 activator MotA and co-activator AsiA.Microbiology 151: 1729–1740.
Hisbergues, M., Jeanjean, R., Joset, F., Tandeau de Marsac,N., and Bedu, S. (1999) Protein PII regulates both inor-ganic carbon and nitrate uptake and is modified by a redoxsignal in synechocystis PCC 6803. FEBS Lett 463: 216–220.
Jaggi, R., Ybarlucea, W., Cheah, E., Carr, P.D., Edwards,K.J., Ollis, D.L., and Vasudevan, S.G. (1996) The role ofthe T-loop of the signal transducing protein PII fromEscherichia coli. FEBS Lett 391: 223–228.
James, P., Halladay, J., and Craig, E.A. (1996) Genomiclibraries and a host strain designed for highly efficient two-hybrid selection in yeast. Genetics 144: 1425–1436.
Jiang, P., Zucker, P., Atkinson, M.R., Kamberov, E.S., Tira-sophon, W., Chandran, P., et al. (1997) Structure/functionanalysis of the PII signal transduction protein of Escheri-chia coli: genetic separation of interactions with proteinreceptors. J Bacteriol 179: 4342–4353.
Karimova, G., Pidoux, J., Ullmann, A., and Ladant, D. (1998)A bacterial two-hybrid system based on a reconstitutedsignal transduction pathway. Proc Natl Acad Sci USA 95:5752–5756.
Karimova, G., Dautin, N., and Ladant, D. (2005) Interactionnetwork among Escherichia coli membrane proteinsinvolved in cell division as revealed by bacterial two-hybridanalysis. J Bacteriol 187: 2233–2243.
Kobayashi, M., Takatani, N., Tanigawa, M., and Omata, T.(2005) Posttranslational regulation of nitrate assimilation inthe cyanobacterium Synechocystis sp. strain PCC 6803. JBacteriol 187: 498–506.
Lee, H.M., Vazquez-Bermudez, M.F., and de Marsac, N.T.(1999) The global nitrogen regulator NtcA regulates tran-scription of the signal transducer PII (GlnB) and influencesits phosphorylation level in response to nitrogen andcarbon supplies in the cyanobacterium Synechococcus sp.strain PCC 7942. J Bacteriol 181: 2697–2702.
Lee, H.M., Flores, E., Forchhammer, K., Herrero, A., andTandeau De Marsac, N. (2000) Phosphorylation of thesignal transducer PII protein and an additional effector arerequired for the PII-mediated regulation of nitrate and nitriteuptake in the cyanobacterium Synechococcus sp. PCC7942. Eur J Biochem 267: 591–600.
Luque, I., Vazquez-Bermudez, M.F., Paz-Yepes, J., Flores,E., and Herrero, A. (2004) In vivo activity of the nitrogencontrol transcription factor NtcA is subjected to metabolicregulation in Synechococcus sp. strain PCC 7942. FEMSMicrobiol Lett 236: 47–52.
Maheswaran, M., Urbanke, C., and Forchhammer, K. (2004)Complex formation and catalytic activation by the PII sig-
468 J. Espinosa, K. Forchhammer, S. Burillo and A. Contreras
© 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 457–469No claim to original Spanish government works

naling protein of N-acetyl-L-glutamate kinase from Syn-echococcus elongatus strain PCC 7942. J Biol Chem 279:55202–55210.
Martinez-Argudo, I., and Contreras, A. (2002) PII T-loopmutations affecting signal transduction to NtrB also abolishyeast two-hybrid interactions. J Bacteriol 184: 3746–3748.
Mavris, M., Page, A.L., Tournebize, R., Demers, B., San-sonetti, P., and Parsot, C. (2002) Regulation of transcrip-tion by the activity of the Shigella flexneri type III secretionapparatus. Mol Microbiol 43: 1543–1553.
Muro-Pastor, M.I., Reyes, J.C., and Florencio, F.J. (2001)Cyanobacteria perceive nitrogen status by sensing intrac-ellular 2-oxoglutarate levels. J Biol Chem 276: 38320–38328.
Muro-Pastor, M.I., Reyes, J.C., and Florencio, F.J. (2005)Ammonium assimilation in cyanobacteria. Photosynth Res83: 135–150.
Ninfa, A.J., and Jiang, P. (2005) PII signal transduction pro-teins: sensors of alpha-ketoglutarate that regulate nitrogenmetabolism. Curr Opin Microbiol 8: 168–173.
Osanai, T., Sato, S., Tabata, S., and Tanaka, K. (2005) Iden-tification of PamA as a PII-binding membrane proteinimportant in nitrogen-related and sugar-catabolic geneexpression in Synechocystis sp. PCC 6803. J Biol Chem280: 34684–34690.
Paz-Yepes, J., Flores, E., and Herrero, A. (2003) Transcrip-tional effects of the signal transduction protein P(II) (glnBgene product) on NtcA-dependent genes in Synechococ-cus sp. PCC 7942. FEBS Lett 543: 42–46.
Pellegrini, M., Marcotte, E.M., Thompson, M.J., Eisenberg,D., and Yeates, T.O. (1999) Assigning protein functions bycomparative genome analysis: protein phylogeneticprofiles. Proc Natl Acad Sci USA 96: 4285–4288.
Phillips, T.A., VanBogelen, R.A., and Neidhardt, F.C. (1984)lon gene product of Escherichia coli is a heat-shockprotein. J Bacteriol 159: 283–287.
Roder, K.H., Wolf, S.S., and Schweizer, M. (1996) Refine-ment of vectors for use in the yeast two-hybrid system.Anal Biochem 241: 260–262.
Salinas, P., and Contreras, A. (2003) Identification and analy-sis of Escherichia coli proteins that interact with the histi-dine kinase NtrB in a yeast two-hybrid system. Mol GenetGenomics 269: 574–581.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989) Molecu-lar Cloning: A Laboratory Manual. Cold Spring Harbor, NewYork: Cold Spring Harbor Laboratory Press.
Sauer, J., Gorl, M., and Forchhammer, K. (1999) Nitrogenstarvation in synechococcus PCC 7942: involvement ofglutamine synthetase and NtcA in phycobiliprotein degra-dation and survival. Arch Microbiol 172: 247–255.
Sugiyama, K., Hayakawa, T., Kudo, T., Ito, T., and Yamaya,T. (2004) Interaction of N-acetylglutamate kinase witha PII-like protein in rice. Plant Cell Physiol 45: 1768–1778.
Tanigawa, R., Shirokane, M., Maeda Si, S., Omata, T.,Tanaka, K., and Takahashi, H. (2002) Transcriptional acti-vation of NtcA-dependent promoters of Synechococcus sp.PCC 7942 by 2-oxoglutarate in vitro. Proc Natl Acad SciUSA 99: 4251–4255.
Vazquez-Bermudez, M.F., Flores, E., and Herrero, A. (2002)Analysis of binding sites for the nitrogen-control transcrip-tion factor NtcA in the promoters of Synechococcusnitrogen-regulated genes. Biochim Biophys Acta 1578:95–98.
Zhang, Y., Pohlmann, E.L., and Roberts, G.P. (2004) Identi-fication of critical residues in GlnB for its activation of NifAactivity in the photosynthetic bacterium Rhodospirillumrubrum. Proc Natl Acad Sci USA 101: 2782–2787.
Zhu, Y., Conrad, M.C., Zhang, Y., and Roberts, G.P. (2006)Identification of Rhodospirillum rubrum GlnB variants thatare altered in their ability to interact with different targets inresponse to nitrogen status signals. J Bacteriol 188: 1866–1874.
Supplementary material
The following supplementary material is available for thisarticle online:Fig. S1. Alignment of deduced amino acid sequences forPipX homologues from the following cyanobacteria:Fig. S2. PipX interactions in the BACTH system.Fig. S3. Insertional mutations in the pipX genomic region.Table S1. Oligonucleotides used in this work.
This material is available as part of the online article fromhttp://www.blackwell-synergy.com
Nitrogen interaction regulatory network 469
© 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 457–469No claim to original Spanish government works
Copyright © 2022 FDOKUMEN




















