Connexin-dependent signaling in neuro-hormonal systems
18
Review Connexin-dependent signaling in neuro-hormonal systems ☆ Ilaria Potolicchio a , Valentina Cigliola a , Silvia Velazquez-Garcia a , Philippe Klee a , Amina Valjevac b , Dina Kapic c , Esad Cosovic c , Orhan Lepara b , Almira Hadzovic-Dzuvo b , Zakira Mornjacovic c , Paolo Meda a, ⁎ a Department of Cell Physiology and Metabolism, University of Geneva Medical School, Switzerland b Department of Physiology and Biochemistry, University of Sarajevo Medical School, Bosnia and Herzegovina c Department of Histology and Embryology, University of Sarajevo Medical School, Bosnia and Herzegovina abstract article info Article history: Received 16 June 2011 Received in revised form 14 September 2011 Accepted 23 September 2011 Available online 1 October 2011 Keywords: Endocrine gland Hormone Insulin Renin Cx36, Cx40, diabetes, hypertension The advent of multicellular organisms was accompanied by the development of short- and long-range chem- ical signalling systems, including those provided by the nervous and endocrine systems. In turn, the cells of these two systems have developed mechanisms for interacting with both adjacent and distant cells. With evolution, such mechanisms have diversified to become integrated in a complex regulatory network, where- by individual endocrine and neuro-endocrine cells sense the state of activity of their neighbors and, accord- ingly, regulate their own level of functioning. A consistent feature of this network is the expression of connexin-made channels between the (neuro)hormone-producing cells of all endocrine glands and secretory regions of the central nervous system so far investigated in vertebrates. This review summarizes the distribu- tion of connexins in the mammalian (neuro)endocrine systems, and what we know about the participation of these proteins on hormone secretion, the life of the producing cells, and the action of (neuro)hormones on specific targets. The data gathered since the last reviews on the topic are summarized, with particular empha- sis on the roles of Cx36 in the function of the insulin-producing beta cells of the endocrine pancreas, and of Cx40 in that of the renin-producing juxta-glomerular epithelioid cells of the kidney cortex. This article is part of a Special Issue entitled: The Communicating junctions, composition, structure and characteristics. © 2011 Elsevier B.V. All rights reserved. Contents 1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1920 2. Connexins and hormone-producing cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1920 3. Cells producing peptide hormones . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1921 3.1. GnRH-producing neurons . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1921 3.2. Dopamine-producing neurons . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1921 3.3. Vasopressin- and oxytocin-producing neurons . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1921 3.4. Pineal gland . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1921 3.5. Anterior pituitary . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1921 3.6. Pancreatic islets . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1922 3.6.1. The insulin-producing beta cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1922 3.6.2. The glucagon-producing alpha cells and the somatostatin-producing delta cells . . . . . . . . . . . . . . . . . . . . . . . . 1923 4. Cells producing glycoprotein hormones . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1923 4.1. Thyroid . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1923 4.2. Placenta . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1924 4.3. The renin-producing myo-epithelioid cells of kidney . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1924 5. Cells producing steroid hormones . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1925 5.1. Adrenal gland . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1925 5.2. Testis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1925 5.2.1. Inhibin- and antimullerian hormone-producing Sertoli cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1925 5.2.2. Androgen-producing Leydig cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1925 Biochimica et Biophysica Acta 1818 (2012) 1919–1936 ☆ This article is part of a special issue entitled: The Communicating junctions, composition, structure and characteristics. ⁎ Corresponding author at: Department of Cell Physiology and Metabolism, University of Geneva, C.M.U., 1 rue Michel Servet, 1211 Geneva 4, Switzerland. Tel.: +41 22 702 52 10; fax: +41 22 702 52 60. E-mail address: [email protected] (P. Meda). 0005-2736/$ – see front matter © 2011 Elsevier B.V. All rights reserved. doi:10.1016/j.bbamem.2011.09.022 Contents lists available at SciVerse ScienceDirect Biochimica et Biophysica Acta journal homepage: www.elsevier.com/locate/bbamem
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Connexin-dependent signaling in neuro-hormonal systems

Biochimica et Biophysica Acta 1818 (2012) 1919–1936
Contents lists available at SciVerse ScienceDirect
Biochimica et Biophysica Acta
j ourna l homepage: www.e lsev ie r .com/ locate /bbamem
Review
Connexin-dependent signaling in neuro-hormonal systems☆
Ilaria Potolicchio a, Valentina Cigliola a, Silvia Velazquez-Garcia a, Philippe Klee a, Amina Valjevac b,Dina Kapic c, Esad Cosovic c, Orhan Lepara b, Almira Hadzovic-Dzuvo b, Zakira Mornjacovic c, Paolo Meda a,⁎a Department of Cell Physiology and Metabolism, University of Geneva Medical School, Switzerlandb Department of Physiology and Biochemistry, University of Sarajevo Medical School, Bosnia and Herzegovinac Department of Histology and Embryology, University of Sarajevo Medical School, Bosnia and Herzegovina
☆ This article is part of a special issue entitled: The Co⁎ Corresponding author at: Department of Cell Physiolo
+41 22 702 52 60.E-mail address: [email protected] (P. Meda).
0005-2736/$ – see front matter © 2011 Elsevier B.V. Alldoi:10.1016/j.bbamem.2011.09.022
a b s t r a c t
a r t i c l e i n f oArticle history:Received 16 June 2011Received in revised form 14 September 2011Accepted 23 September 2011Available online 1 October 2011
Keywords:Endocrine glandHormoneInsulinReninCx36, Cx40, diabetes, hypertension
The advent of multicellular organisms was accompanied by the development of short- and long-range chem-ical signalling systems, including those provided by the nervous and endocrine systems. In turn, the cells ofthese two systems have developed mechanisms for interacting with both adjacent and distant cells. Withevolution, such mechanisms have diversified to become integrated in a complex regulatory network, where-by individual endocrine and neuro-endocrine cells sense the state of activity of their neighbors and, accord-ingly, regulate their own level of functioning. A consistent feature of this network is the expression ofconnexin-made channels between the (neuro)hormone-producing cells of all endocrine glands and secretoryregions of the central nervous system so far investigated in vertebrates. This review summarizes the distribu-tion of connexins in the mammalian (neuro)endocrine systems, and what we know about the participation ofthese proteins on hormone secretion, the life of the producing cells, and the action of (neuro)hormones onspecific targets. The data gathered since the last reviews on the topic are summarized, with particular empha-sis on the roles of Cx36 in the function of the insulin-producing beta cells of the endocrine pancreas, and ofCx40 in that of the renin-producing juxta-glomerular epithelioid cells of the kidney cortex. This article is partof a Special Issue entitled: The Communicating junctions, composition, structure and characteristics.
mmunicating junctions, composition, structure and chagy andMetabolism, University of Geneva, C.M.U., 1 rueMic
rights reserved.
© 2011 Elsevier B.V. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19202. Connexins and hormone-producing cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19203. Cells producing peptide hormones . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1921
3.1. GnRH-producing neurons . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19213.2. Dopamine-producing neurons . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19213.3. Vasopressin- and oxytocin-producing neurons . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19213.4. Pineal gland. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19213.5. Anterior pituitary . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19213.6. Pancreatic islets . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1922
3.6.1. The insulin-producing beta cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19223.6.2. The glucagon-producing alpha cells and the somatostatin-producing delta cells . . . . . . . . . . . . . . . . . . . . . . . . 1923
4. Cells producing glycoprotein hormones . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19234.1. Thyroid . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19234.2. Placenta . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19244.3. The renin-producing myo-epithelioid cells of kidney . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1924
5. Cells producing steroid hormones . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19255.1. Adrenal gland . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19255.2. Testis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1925
5.2.1. Inhibin- and antimullerian hormone-producing Sertoli cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19255.2.2. Androgen-producing Leydig cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1925
racteristics.hel Servet, 1211 Geneva 4, Switzerland. Tel.:+41 22 702 52 10; fax:

1920 I. Potolicchio et al. / Biochimica et Biophysica Acta 1818 (2012) 1919–1936
5.3. Ovary . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19265.3.1. Oestrogens-producing granulosa and theca cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19265.3.2. Progesterone-producing luteal cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1926
6. Cells producing catecholamine hormones. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19267. Cells producing pheromones . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19278. Connexins contribution to the peripheral action of hormones . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19279. Hormonal control of connexin expression . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1927
9.1. Peptides, second messengers and glycosides . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19279.2. Steroids . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19279.3. Gonadotrophins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19279.4. Thyroid hormones . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1928
10. Connexins and endocrine disruptors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 192811. Connexins and human endocrine disorders . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1928
11.1. Diabetes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 192811.2. Hypertension . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 192811.3. Metabolic syndrome and diabetic complications . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 192911.4. Endocrine tumors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1929
12. Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1930Acknowledgment . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1930References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1930
1. Introduction
The appearance of multicellular systems was associated to the de-velopment of the nervous and endocrine systems, that ensure chem-ical signaling between adjacent and distant cells [1–5]. In turn, propersignalling by these systems to the multicellular organism has im-posed the development of signaling units sizably larger than the uni-cellular exocrine systems, which are considered their primordialprecursors [5–9]. Thus, most of the vital endocrine functions of ani-mals are now dependent on the proper functioning of multicellularassemblies of secretory cells, whether in endocrine glands or special-ized nervous regions, which implies the existence of systems coordi-nating the function of the hormone-producing cells. With evolution,such coordination systems have developed in a complex network, inwhich signals diffusing in the intercellular spaces interplay with thesignalling cascades dependent on the surface proteins that concen-trate at cell contacts [10–12]. Thus, individual hormone-producingcells sense the state of activity of their neighbors and regulate, ac-cordingly, their own level of functioning. In vertebrates, a consistentfeature of this network are the cell-to-cell channels made of connexinproteins [10–17]. As compared to other forms of cell-to-cell commu-nication, connexin-dependent cell-to-cell coupling achieves a rapidintercellular equilibration of electrochemical gradients of both cyto-plasmic ions and molecules, which may be crucial for hormone secre-tion [10–19]. In turn, this equilibration ensures the recruitment andcoordination of cells which, taken individually, would otherwisefunction asynchronously [10–19]. Although secretory cells of a giventype are generally considered to be similar, ample evidence indicatethat they actually differ in several respects, including the ability tobiosynthesize, store and release hormones [10–23], still function ashomogeneous populations when integrated by coupling. Thus, alarge body of circumstantial evidence has accumulated, indicating thatgap junctions are required for the fine regulation of the biosynthesis,storage and release of various hormones, as well as for their proper ac-tion on target cells [10–19]. Conversely, several hormones have nowbeen documented to contribute to the control of connexin expressionand function. More intriguingly, recent work implies that connexinsignaling may also be involved between selected endocrine cells, andtheir cognate, closely associated target cells, a hitherto unsuspectedmechanism for the two-way interaction between signaling and effectorcells.
Here, we have reviewed this body of evidence, with particular regardto the studies which have investigated the connexin-dependent signal-ling of vertebrate endocrine cells. The goal was not to provide an in-
depth, comprehensive review of the abundant information gathered onthe topic in all endocrine glands, rather to stress on recent developments,specifically when in vivo evidence is available. Readers are referred toprevious reviews [10–19], for earlier publications on the topic.
2. Connexins and hormone-producing cells
The hormone-producing cells of most endocrine glands, of secretorycentral nervous system nuclei, and of their putative precursor phero-mone glands, usually express one or more connexins of either the αor the γ group (Table 1). Cx43 is the most widespread isoform. A fewendocrine cells, including most neurosecretory neurons, have selectedCx36, whereas Cx40 is so far restricted to the renin-producing cells ofkidney (Table 1). With the notable exception of the thyroid, which em-bryologically develops as an exocrine gland and maintains throughoutthe life an architectural organization reminiscent of this primordial exo-crine function [5,24], no endocrine cell has yet been shown to expressCx32, eventhough a few of these cells have selected Cx26, another con-nexin of the β group (Table 1). Conversely, the parenchymal cells ofexocrine glands express various combinations of β connexins, typicallyCx32 [5]. This different connexin selection is under transcriptional con-trol. Thus, the 5' regulatory region of the human Gjd2, contains a regionsufficient to restrict the expression of Cx36 to insulin-secreting cells andneurons. Within this region, a conserved neuron-restrictive silencer el-ement binds the NRSF/REST factor, which functions as a potent repres-sor of neuronal genes. NRSF/REST iswidely expressed inmost cell types,but insulin-secreting cells and neurons, and its ectopic expression re-duces that of Cx36 in insulin-producing cells [25]. Also, mice lackingthe basic helix-loop-helix transcription factor Mist1, which is normallyexpressed in most exocrine cell types, feature pancreatic acini with al-tered architecture and function, due, at least in part, to a transcriptionaldown regulation of Gjb1 and Cx32 [26].
The reason why different connexin species have been selected bydifferent types of endocrine cells cannot be directly related to eitherthe embryological origin, the developmental time or the architectureof the adult glands. It cannot be related either to the biochemical na-ture of the hormones produced by each gland, the rate of their secre-tion and the control of their release and biosynthesis (Table 1).However, since different connexins form channels with distinct con-ductance, permeability and regulatory characteristics [10,18,27–30],it is plausible that, during evolution, (neuro)endocrine cells selectedthe sets of channels that, by favoring the intercellular exchange ofspecific signals while preventing the diffusion of others, better fittedthe requirements of their own secretory machinery.

Table 1Connexins of vertebrate (neuro)endocrine glands.
Type of secretion Gland Cell Hormone Connexin isoform Species Reference
Endocrine Hypothalamus Neurons GnRH Cx43, Cx32Cx36
RatMouse
[41,42][37]
Vasopressin Cx43 Mouse [40]Oxytocin Cx32 Rat [59]
Pineal Pinealocytes Melatonin, serotonin Cx26, Cx32 Rat [60]Pituitary Acidophil GH, prolactin, Cx43, Cx26 Rat [24,65]Thyroid Follicular T3, T4 Cx43, Cx32, Cx26 Rat [24,133]
C Calcitonin Cx43 Rat [24]Parathyroid Principal Parathormone Cx43 Rat [24]Pancreas Beta Insulin Cx36
Cx36Rat, mouseHuman
[24,85,335][75]
Kidney Myoepithelial Renin Cx40, Cx37, Cx45Cx40, Cx37
Mouse, ratHuman
[154,164][155]
Adrenal Medullary EpinephrineNorepinephrine
Cx36, Cx43Cx36Cx43, Cx50
RatMouseHuman
[24][325,336][334]
Cortical Corticoids, mineralocorticoidsAndrogens
Cx43Cx43
RatMouse
[24,181][182]
Testis Leydig Testosterone Cx43Cx43, Cx30.2
RatMouse
[337][339]
Sertoli InhibinAntimullerian hormone
Cx43 Rat [337,338]
Luteal Progesterone Cx43Cx43, Cx32, Cx26Cx43
RatSheepHuman, baboon
[223][211][224]
Ovary Granulosa and theca Estradiol Cx43Cx43, Cx37, Cx26Cx43, Cx26, Cx30.3, Cx32Cx43, Cx37, Cx26, Cx32Cx43
Mouse, ratSheepPorcBeefHuman
[198,201][211][341][342][340]
Placenta Throphoblast Chorionic gonadotrophin RatHuman
[144,145][146,147]
Pheromone Skin Sebaceous Several in sebum Cx43 Human [239]Preputial glands Parenchymal Several farnesenes Cx43 Rat [24]
1921I. Potolicchio et al. / Biochimica et Biophysica Acta 1818 (2012) 1919–1936
3. Cells producing peptide hormones
3.1. GnRH-producing neurons
Several of the two magnocellular and five parvicellular neurontypes which produce hormones in the paraventricular nucleus ofthe hypothalamus are coupled in vertebrates [31–37]. Most of thiscoupling is presumably mediated by Cx36 [36,38], the only connexinwhich has so far been convincingly shown to be expressed at neuro-nal gap junction plaques [39]. Other neurosecretory neurons may alsoexpress Cx43 [40,41]. Eventually, GnRH neurons appear not coupledin the hypothalamus [42–44], but are coupled in the neuromodula-tory, extrahypothalamic nerve GnRH3 subpopulation [45]. Electro-physiological studies testing gap junction blockers have indicatedthat coupling is implicated in the control of the pulsatile release ofseveral neuropeptides, including GHRH [46], GnRH [47,48] andLHRH [49], presumably via the intercellular synchronization of Ca2+
waves [50]. siRNA targeting Cx43 abolished the pulsatile release ofGnRH in cultures of immortalized GT1–7 neuronal cells [41], andcell-specific deletion of Cx36 in either prenatal or adult mice inducedalterations in estrous cyclicity, indicating that preoptic kisspeptinneurons require Cx36 to properly control the GnRH network [43]. Im-mortalized GnRH-producing neurons of the GT1–7 line, express Cx26and are coupled [51–53] by channels that appear up-regulated bycAMP [35]. The primary neurons that intermittently deliver luteiniz-ing hormone-releasing hormone (GnRH) to the anterior pituitaryare also coupled in vivo [55].
3.2. Dopamine-producing neurons
The secretion of prolactin by the anterior pituitary is suppressed,under non lactating conditions, by tuberoinfundibular dopamine neu-rons of the hypothalamus [54]. These neurons oscillate rhythmically
via a mechanism that persists during blockade of chemical synaptictransmission, and it is abolished by gap junction antagonists. The thyro-tropin-releasing hormone forces these neurons to enter a tonic, not os-cillatory firing, during which their dopamine release is decreased,leading to a potent stimulation of prolactin from the pituitary [55].
3.3. Vasopressin- and oxytocin-producing neurons
The magnocellular neurons of the paraventricular and supraopticnuclei, which produce vasopressin and oxytocin, are electrically- anddye-coupled [56–58]. Towards the end of pregnancy, lactation becomesregulated by a synchronized and high-frequency firing of oxytocinergicneurons, coupled by Cx32 [59]. The expression of this connexin is low inthe supraoptic nucleus of virgin females, and increases during late preg-nancy and lactation [59].
3.4. Pineal gland
Pinealocytes have been reported to be coupled by Cx26 channels,that are up-regulated in vitro by norepinephrine, possibly to improvemelatonin secretion [60].
3.5. Anterior pituitary
The different types of endocrine cells of the anterior pituitary arecoupled [61–64] via channels made predominantly of Cx43 and Cx26[24,65]. This coupling increases the intercellular synchronization ofCa2+ transients [51,66]. Ca2+ waves are also propagated throughout theentire gland via the network of the non endocrine folliculo-stellate cells,which are extensively coupled by Cx43 channels [63,66,67,68]. In thehorse, the gap junctions of folliculo-stellate appear modulated by theseason and/or photoperiod [69]. Seasonal changes in the expression of pi-tuitary Cx43 have been associated to changes in prolactin secretion [70].

Fig. 2. Beta cell gap junctions increase after glibenclamide treatment. A–B) Transmis-sion electron microscopy shows the abundant number of insulin-containing granules(round clear structures with a dense core) in the beta cells (bc) of a control rat islet(A), and the marked loss of these organelles after an in vivo treatment with glibencla-mide (B). This sulphonylurea chronically stimulates insulin secretion. C–D) The corre-sponding freeze-fracture electron microscopy reveals that control rat beta cells areconnected by small gap junction plaques (C), whereas the glibenclamide-treated, insu-lin-depleted beta cells feature sizably larger connexon plaques (D). In both cases, mostgap junctions are associated to short tight junction fibrils. Bar, 800 nm in A and B, and125 nm in C and D.
1922 I. Potolicchio et al. / Biochimica et Biophysica Acta 1818 (2012) 1919–1936
3.6. Pancreatic islets
The pancreatic islets of Langerhans are minute assemblies (50–600 μm diameter) of 50–3000 endocrine cells, intermixed with vascu-lar, connective and immune cells. In humans, about a million of such is-lets are dispersed in the pancreas, representing in total about 1% volumeof the adult gland, and 1 gwet weight tissue [71]. The insulin-producingbeta cells account for 60–80% of the endocrine population. The remain-ing endocrine cells are glucagon-producing alpha cells, somatostatin-producing delta cells, pancreatic polypeptide-producing PP-cells, andghrelin-producing epsilon cells [71].
3.6.1. The insulin-producing beta cellsPancreatic beta cells of rodents and humans are coupled only by
Cx36 (Fig. 1) channels [72–76], which allow for their ionic couplingthroughout each pancreatic islet [77–83], and for a more spatially re-stricted dye and metabolic coupling [23,84–93], specifically for nega-tively charged molecules [86]. This dye coupling is increased in theislets of transgenic mice whose β-cells are forced to ectopically ex-press Cx32 or Cx43 in addition to the native Cx36 [23,85,86,94,95],and also increases in control islets after in vivo treatment with gliben-clamide, a sulphonylurea which chronically stimulates insulin secre-tion [86,88,90] (Fig. 2).
To assess the functional role of this coupling, initial experimentsinvestigated single beta cells, which cannot communicate via gapjunctions. These cells showed increased basal release of insulin(i.e. the secretion of the hormone observed in the absence of anystimulus, and which cumulatively accounts for most of the daily insu-lin production), poor responsiveness to glucose, decreased basal ex-pression of the 2 insulin genes, decreased proinsulin biosynthesis,and poor or no elevation in free cytosolic Ca2+ after glucose stimula-tion [21,22,96–105]. Reestablishment of contacts between beta
Fig. 1. The insulin-producing beta cells of pancreas are connected by Cx36-made gapjunctions. A) Immunofluorescence detects Cx36 (red) within an endocrine pancreaticislet (eni) from a control mouse, but not in the adjacent exocrine acini (exa). B) Co-immunostaining for E-cadherin (green) reveals the spotted distribution of Cx36 atthe membrane of islet cells. C) Transmission electron microscopy at the periphery oftwo adjacent beta cells (bc) from a control mouse, identified by typical, dense core in-sulin-containing granules, shows the apposition of their membranes at the site of a gapjunction (red arrowheads). Bar, 20 μm in A and B, and 50 nm in C.
cells, rapidly restores glucose-induced insulin release, (pro)insulin biosynthesis, and the elevations in free cytosolic Ca2+
[21,22,90,91,97,98,106]. Beta cell contact also promotes the recruit-ment of secretory and biosynthetically active beta cells [21,22,96–99,103]. Beta cells are structurally, biochemically and functionallyheterogeneous, including in terms of the biosynthesis, storage and re-lease of insulin [21,22,96,100,102,103,108–112]. Under such condi-tions, a mechanism dependent on cell contact allows for therecruitment of increasing numbers of secretory cells with both cellaggregation and the degree of stimulation [21–23,97,98,113]. Lackof coupling prevents the sharing by β-cells of sparse ion channelsthat are critical for proper activation of the stimulus-secretion cou-pling [114]. Comparable conclusions were reached by exposing isolat-ed pancreatic islets [115] or whole pancreas [116] to drugs blockinggap junction channels [117]. Further experiments on insulin-produc-ing cell lines, showed that the lines which do not release insulin in re-sponse to a physiological stimulation by glucose, do not expressconnexins and are uncoupled [84,107,118,119], whereas cell linesretaining at least some glucose responsiveness express Cx36 and arecoupled [107]. Remarkably, Cx36 is detected in developing rodentsat the time the first insulin-containing beta cells are detected, andits expression increases around birth, coincidentally with the acquisi-tion by beta cells of a normal sensitivity to glucose [93].
With the availability of Cx36 null and transgenic mice, the role ofCx36, and other connexins that are not expressed in native islets,could be investigated in vivo. Loss of Cx36 [76,120], results in theuncoupling of beta cells lacking gap junction plaques, and in the asyn-chrony of the oscillations in free cytosolic Ca2+, which are normallyinduced during stimulation by the natural secretagogue glucose[76]. Cx36-null mice have morphologically normal islets which, how-ever, do not release insulin in a normal, pulsatile fashion, show an in-creased basal release of insulin and no significant increase in

1923I. Potolicchio et al. / Biochimica et Biophysica Acta 1818 (2012) 1919–1936
hormonal output when the pancreas was stimulated by glucose[76,120]. These islets also cannot be inhibited as rapidly as control is-lets when the glucose stimulation ends, due to lack of intercellulardiffusion of hyperpolarizing, inhibitory electrotonic currents [83].These defects are pancreatic autonomous, inasmuch as similar obser-vations were made after a beta-cell specific deletion of Cx36 [120],and in clones of MIN6 cells that were depleted in Cx36 by an anti-sense approach [107,118,119,122]. These cells allowed to further es-tablish that the secretion control achieved by the Cx36 signaling(Fig. 3) cannot be mediated by other connexins, nor by E-cadherin[84,118,121], and that excess Cx36 is as deleterious for insulin secre-tion as the lack of the connexin [84,119,122]. In this perspective, trans-genic mice forced to express Cx32 in beta cells, show a quite largeincrease in the dye coupling of beta cells, and are glucose intolerant,due to decreased insulin secretory response to supra-physiological glu-cose stimulation [85].
Cx36-dependent signaling appears also implicated in the life ofbeta cells (Fig. 3). Thus, loss of Cx36 sensitizes beta cells to
Fig. 3. Schematic view of the interactions of beta and alpha cells within pancreatic is-lets. The scheme depicts 3 beta cells (bc) contacting each other and an adjacentalpha cell (ac). Experimental data show that most beta cells are coupled (blue coloredcells) via Cx36 channels, which allow for the intercellular synchronization of glucose-induced Ca2+ transients, the exchange of depolarizing and hyperpolarizing currents,second messengers and metabolites. These intercellular communications promotethe recruitment of secretory cells as a function of stimulation by secretagogues, and po-tentiate their secretory output. Cx36-dependent coupling also increases the resistanceof beta cells to cytotoxic conditions. Loss of coupling connections, as a result of eitherabsence of Cx36, or blockade of the channels this protein makes, results in increasedbasal insulin secretion, and beta cell apoptosis. In parallel, the stimulated secretion ofinsulin, and the expression of the cognate genes are decreased, as is the recruitmentof secretory cells during exposure to secretagogues. Also, several signaling pathways,including those dependent of E-cadherin and ephrins, are perturbed. The phenotypicresults of these changes are alterations in both insulin secretion, and beta cell resis-tance, that are reminiscent of those observed in (pre-)diabetic states.
pharmacological and immunological aggressions [95,123,124], in-cluding the cytokines which induce beta cell apoptosis at the onsetof the auto-immune attack of pancreatic islets that leads to type 1 di-abetes [95]. Conversely, transgenic mice overexpressing Cx36, or ec-topically expressing in beta cells Cx32 [85] or Cx43, featured betacells which were protected in vitro and in vivo against the same ag-gressions, possibly because of enlarged coupling [95]. Native Cx36does not alter islet size or insulin content, whereas the Cx43 isoformincreases both parameters, and Cx32 has a similar effect only whencombined with growth hormone [85,94,125].
3.6.2. The glucagon-producing alpha cells and the somatostatin-producingdelta cells
Pancreatic islets are coordinated assemblies of multiple types ofendocrine cells, which functionally interact with each other. Thus,the insulin released by beta cells acts on receptors on the closeby,sometimes adjacent alpha, delta, epsilon and pancreatic polypep-tide-producing-cells, and, vice versa, glucagon and somatostatin acton receptors of beta cells [126,127]. There has been an early reportthat at least some of these different cell types may be joined by gapjunctions [128]. However, this evidence is indirect, and not supportedby functional coupling data [78,92,129,130]. Thus, whereas secreta-gogue-induced Ca2+ oscillations are fully synchronized by Cx36channels among beta cells [76,107], they are asynchronous in alphaand delta cells of rodent and human islets, both with respect toeach other cell type, and to beta cells [129,130]. The data are consis-tent with the view that, if the latter cell types were to also expressCx36, this protein was not involved in the intercellular synchroniza-tion of Ca2+ transients, as it does in beta cells. The potential couplingof different islet cell types should be tightly controlled, if not very se-lective, inasmuch as beta cell function usually parallels that of deltacells, but is antagonist to that of alpha cells [126,127,131]. Thus, glu-cose stimulates in parallel insulin and somatostatin release, whichfeature synchronous oscillations [129], most likely due to regularCa2+ transients [76,107]. Conversely, the nadirs of glucagon oscilla-tions are antisynchronous to the coincident pulses of insulin and so-matostatin [129], suggesting lack of simultaneous Cx36-dependentcoupling between these 3 cell types. Still, the finding that human is-lets express several connexin transcripts [75], calls for a careful eval-uation of alternative possibilities. Since all secretory cell types have sofar been found to express at least one connexin species, it is not clearwhy alpha and delta pancreatic cells would be an exception. The reg-ular oscillations they do display in terms of secretory output [131],rather suggests that these cells may also coupled by some, still elusiveconnexin isoform.
In summary, while there is ample evidence that Cx36 plays a sig-nificant role in the life and secretion of the pancreatic beta cells, littleis known about whether a connexin-dependent signaling is also im-plicated in the intra-islet integration of the other types of cells, thatcollectively form the endocrine pancreas.
4. Cells producing glycoprotein hormones
4.1. Thyroid
The studies implicating gap junctions and connexins in the func-tioning of the thyroid gland have been reviewed in detail [17,132].The follicular cells of thyroid express Cx32, Cx26 and Cx43[24,133,134]. The involvement of connexin signaling in thyroid secre-tion is supported by the observation that TSH stimulation increasesthe coupling of thyrocytes in a time- and concentration-dependentmanner [135], whereas loss of coupling, due to a Cx32 mutation, re-duces the release of thyroxin [136]. Furthermore, transfection of celllines for Cx32 also resulted in increased expression of the thyroglob-ulin gene [137]. Cx32 and coupling are lost with passages in culture, to-gether with the ability of the cells to form follicular structures [136,138],
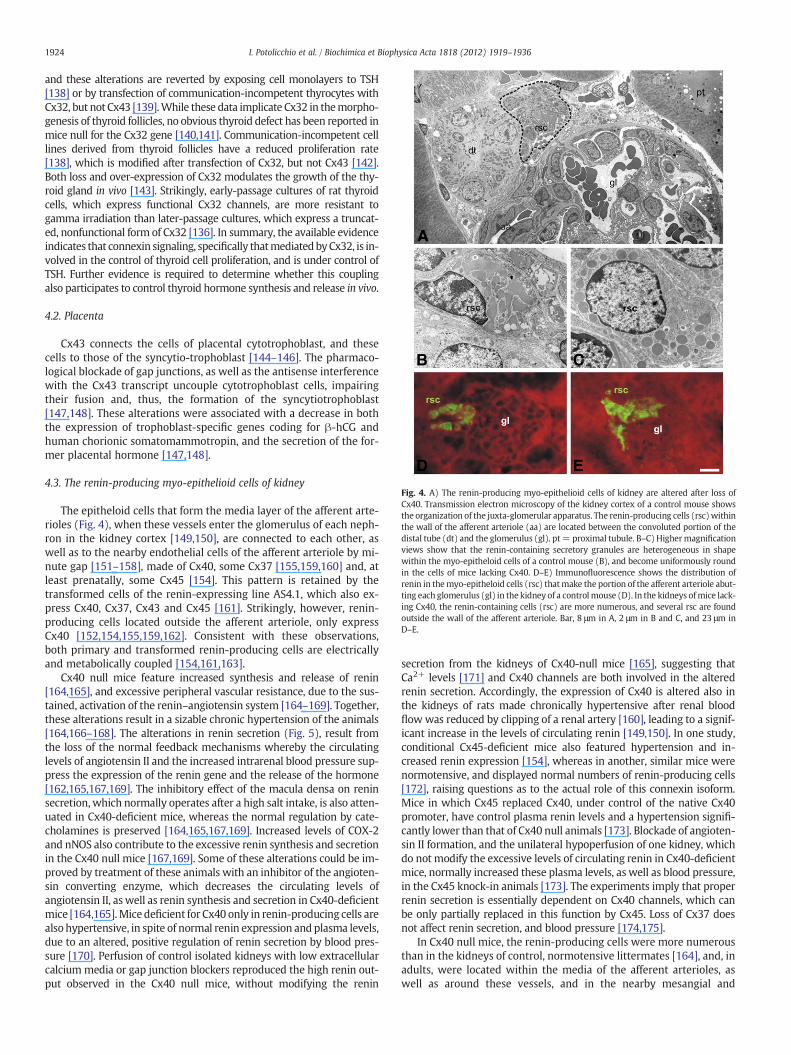
Fig. 4. A) The renin-producing myo-epithelioid cells of kidney are altered after loss ofCx40. Transmission electron microscopy of the kidney cortex of a control mouse showsthe organization of the juxta-glomerular apparatus. The renin-producing cells (rsc)withinthe wall of the afferent arteriole (aa) are located between the convoluted portion of thedistal tube (dt) and the glomerulus (gl). pt= proximal tubule. B–C) Highermagnificationviews show that the renin-containing secretory granules are heterogeneous in shapewithin the myo-epitheloid cells of a control mouse (B), and become uniformously roundin the cells of mice lacking Cx40. D–E) Immunofluorescence shows the distribution ofrenin in themyo-epitheloid cells (rsc) thatmake the portion of the afferent arteriole abut-ting each glomerulus (gl) in the kidney of a controlmouse (D). In the kidneys ofmice lack-ing Cx40, the renin-containing cells (rsc) are more numerous, and several rsc are foundoutside the wall of the afferent arteriole. Bar, 8 μm in A, 2 μm in B and C, and 23 μm inD–E.
1924 I. Potolicchio et al. / Biochimica et Biophysica Acta 1818 (2012) 1919–1936
and these alterations are reverted by exposing cell monolayers to TSH[138] or by transfection of communication-incompetent thyrocytes withCx32, but not Cx43 [139].While these data implicate Cx32 in themorpho-genesis of thyroid follicles, no obvious thyroid defect has been reported inmice null for the Cx32 gene [140,141]. Communication-incompetent celllines derived from thyroid follicles have a reduced proliferation rate[138], which is modified after transfection of Cx32, but not Cx43 [142].Both loss and over-expression of Cx32 modulates the growth of the thy-roid gland in vivo [143]. Strikingly, early-passage cultures of rat thyroidcells, which express functional Cx32 channels, are more resistant togamma irradiation than later-passage cultures, which express a truncat-ed, nonfunctional form of Cx32 [136]. In summary, the available evidenceindicates that connexin signaling, specifically thatmediatedbyCx32, is in-volved in the control of thyroid cell proliferation, and is under control ofTSH. Further evidence is required to determine whether this couplingalso participates to control thyroid hormone synthesis and release in vivo.
4.2. Placenta
Cx43 connects the cells of placental cytotrophoblast, and thesecells to those of the syncytio-trophoblast [144–146]. The pharmaco-logical blockade of gap junctions, as well as the antisense interferencewith the Cx43 transcript uncouple cytotrophoblast cells, impairingtheir fusion and, thus, the formation of the syncytiotrophoblast[147,148]. These alterations were associated with a decrease in boththe expression of trophoblast-specific genes coding for β-hCG andhuman chorionic somatomammotropin, and the secretion of the for-mer placental hormone [147,148].
4.3. The renin-producing myo-epithelioid cells of kidney
The epitheloid cells that form the media layer of the afferent arte-rioles (Fig. 4), when these vessels enter the glomerulus of each neph-ron in the kidney cortex [149,150], are connected to each other, aswell as to the nearby endothelial cells of the afferent arteriole by mi-nute gap [151–158], made of Cx40, some Cx37 [155,159,160] and, atleast prenatally, some Cx45 [154]. This pattern is retained by thetransformed cells of the renin-expressing line AS4.1, which also ex-press Cx40, Cx37, Cx43 and Cx45 [161]. Strikingly, however, renin-producing cells located outside the afferent arteriole, only expressCx40 [152,154,155,159,162]. Consistent with these observations,both primary and transformed renin-producing cells are electricallyand metabolically coupled [154,161,163].
Cx40 null mice feature increased synthesis and release of renin[164,165], and excessive peripheral vascular resistance, due to the sus-tained, activation of the renin–angiotensin system [164–169]. Together,these alterations result in a sizable chronic hypertension of the animals[164,166–168]. The alterations in renin secretion (Fig. 5), result fromthe loss of the normal feedback mechanisms whereby the circulatinglevels of angiotensin II and the increased intrarenal blood pressure sup-press the expression of the renin gene and the release of the hormone[162,165,167,169]. The inhibitory effect of the macula densa on reninsecretion, which normally operates after a high salt intake, is also atten-uated in Cx40-deficient mice, whereas the normal regulation by cate-cholamines is preserved [164,165,167,169]. Increased levels of COX-2and nNOS also contribute to the excessive renin synthesis and secretionin the Cx40 null mice [167,169]. Some of these alterations could be im-proved by treatment of these animals with an inhibitor of the angioten-sin converting enzyme, which decreases the circulating levels ofangiotensin II, as well as renin synthesis and secretion in Cx40-deficientmice [164,165].Mice deficient for Cx40 only in renin-producing cells arealso hypertensive, in spite of normal renin expression and plasma levels,due to an altered, positive regulation of renin secretion by blood pres-sure [170]. Perfusion of control isolated kidneys with low extracellularcalcium media or gap junction blockers reproduced the high renin out-put observed in the Cx40 null mice, without modifying the renin
secretion from the kidneys of Cx40-null mice [165], suggesting thatCa2+ levels [171] and Cx40 channels are both involved in the alteredrenin secretion. Accordingly, the expression of Cx40 is altered also inthe kidneys of rats made chronically hypertensive after renal bloodflowwas reduced by clipping of a renal artery [160], leading to a signif-icant increase in the levels of circulating renin [149,150]. In one study,conditional Cx45-deficient mice also featured hypertension and in-creased renin expression [154], whereas in another, similar mice werenormotensive, and displayed normal numbers of renin-producing cells[172], raising questions as to the actual role of this connexin isoform.Mice in which Cx45 replaced Cx40, under control of the native Cx40promoter, have control plasma renin levels and a hypertension signifi-cantly lower than that of Cx40 null animals [173]. Blockade of angioten-sin II formation, and the unilateral hypoperfusion of one kidney, whichdo not modify the excessive levels of circulating renin in Cx40-deficientmice, normally increased these plasma levels, as well as blood pressure,in the Cx45 knock-in animals [173]. The experiments imply that properrenin secretion is essentially dependent on Cx40 channels, which canbe only partially replaced in this function by Cx45. Loss of Cx37 doesnot affect renin secretion, and blood pressure [174,175].
In Cx40 null mice, the renin-producing cells were more numerousthan in the kidneys of control, normotensive littermates [164], and, inadults, were located within the media of the afferent arterioles, aswell as around these vessels, and in the nearby mesangial and

Fig. 5. Schematic view of the interactions of renin-producing cells within the wall ofthe renal afferent arterioles. The scheme depicts 3 renin-producing cells (rsc) contact-ing each other and adjacent endothelial (ec) and smooth muscle cells (smc). Experi-mental data show that most renin-producing cells are coupled (blue colored cells) toeach other, as well as to nearby endothelial and smooth muscle cells via Cx40 channels(blue on the drawing), which allow for the intercellular exchange of currents, and ex-ogenous tracers permeating gap junctions. These intercellular communications modu-late the regulated secretion of renin, and the positional imformation that maintains rscwithin the vessel wall. Loss of Cx40 coupling results in a large increase in renin secre-tion, which associates with the appearance of rsc within the juxtaglomerular mesan-gium. These changes are Cx40 specific, inasmuch as they are not observed after lossof either Cx37 or Cx45, 2 other connexin isoforms expressed by rsc. Of note, the endo-thelial and smooth muscle cells of the afferent arteriole are further coupled to compan-ion cells by Cx37 and Cx43 (ec) and Cx43 (smc; violet on the drawing) The phenotypicresults of these changes are alterations in both renin secretion and vascular resistance,that result in chronic hypertension.
1925I. Potolicchio et al. / Biochimica et Biophysica Acta 1818 (2012) 1919–1936
interstitial regions of the juxtaglomerular apparatus [162] (Fig. 5).Comparable observations were made in mice featuring a renin-specif-ic loss of Cx40 [170]. This anomalous positioning suggests that Cx40,but not Cx45 [172], is also required for the correct positional informa-tion of renin-producing cells [170], which typically increase in num-ber during hypertension [176,177]. The renin-producing cells alsofeatured smaller and more regular secretory granules than controls[164] (Fig. 4), further suggesting a shift in their differentiation pat-tern, as usually observed when cells of the renin lineage are not inthe native, afferent arteriole niche [178]. These experiments haveprovided clear, and substantial evidence that Cx40, but not Cx37and Cx45, play a critical role in the proper control of renin secretion.Together with the altered expression of Cx43 in the wall of resistancearteries, the loss of Cx40 results in a sustained and chronic hyperten-sion, which is reminiscent of the human pathology.
5. Cells producing steroid hormones
5.1. Adrenal gland
The role of gap junctions and connexins in the function of the adre-nal gland has been repeatedly reviewed [19,179,180]. The endocrinecells of the adrenal cortex are coupled by Cx43 channels [24,181], par-ticularly in the glucocorticoid- and the androgen-producing regions[177]. In vitro experiments testing gap junction blockers, have docu-mented an impaired secretion of cortisol from clusters of adrenal cells,but not single cells, stimulated by ACTH, but not cAMP [183]. In agree-ment with these findings, transfection of adrenal cells with a Cx43 anti-sense construct also resulted in the inhibition of ACTH-induced steroidsecretion [184]. These data are consistent with the observation thatACTH increases the expression of Cx43 both in vivo and in vitro [185].However, and unexpectedly, another gap junction blockerwas reportedto stimulate steroid production, via the activation of both an extracellu-lar signal-related kinase and a calcium/calmodulin-dependent kinase,i.e. by two pathways distinct from the protein kinase A-dependentpathway which normally controls steroidogenesis of adrenal cells[186]. Adrenocortical cells normally respond to ACTH stimulation by in-creasing steroid production and Cx43 expression, and by decreasingtheir growth [185]. Pharmacological inhibition of their gap junctionsproduces the opposite effect, i.e. increased cell growth and decreasedACTH-stimulated steroidogenesis, without altering the levels of Cx43[187]. Furthermore, exposure of adrenal cells to a Cx43 antisensecDNA,which reduced these levels, also resulted in a decreased steroido-genic response, and in an increase in the cell growth rate [182]. Thus,Cx43 modulates both the steroidogenic activity and the proliferationof adrenal cells.
5.2. Testis
The involvement of connexins in the function of the main endo-crine cell types of testis has been recently reviewed in detail [188].
5.2.1. Inhibin- and antimullerian hormone-producing Sertoli cellsCx43-made gap junctions connect Sertoli cells to both companion
cells, and to selected germ cells [183]. Cx43 channels contribute to thefunctional synchronization of Sertoli cells, and to the reciprocal cou-pling interactions these cells have with germ cells [189] to ensureproper spermatogenesis [188]. Accordingly, mice with a Sertoli cell-specific loss of Cx43 feature interrupted spermatogeneis at the sper-matogonia stage, and a proliferation of Sertoli cells [190]. The lattercells were partly undifferentiated, as revealed by the expression ofthe anti-Muellerian hormone, whose pubertal disappearance wasdelayed [190]. However, when testes of mice lacking Cx43 weregrafted under the kidney capsules of adult males (to allow for invivo maturation) and then stimulated in vitro with cAMP or LH, theandrogen production was similar to that of controls, suggesting that,in this system, Cx43 was dispensable for steroidogenesis [191]. Still,Cx43 null mutants show a marked germ cell deficiency [191]. siRNAknockdown of Cx43 in Sertoli cells also slows down the resealing ofthe blood-testis barrier after exposure to either low extracellularCa2+ or bisphenol A in vitro [192]. In vivo, FSH treatment induces Ser-toli cells to proliferate and differentiate, irrespective of the presenceof Cx43-dependent coupling, whereas T3, in the presence or absenceof FSH, reduced this proliferation and induced the formation of lumenin the seminiferous testis tubules, as a function of the maturation ofCx43 gap junctions [193].
5.2.2. Androgen-producing Leydig cellsCells derived from the testosterone-producing Leydig cells of tes-
tis express Cx43 [194,195], and are coupled in vitro up to confluence,under which conditions Cx43 expression and coupling decrease, pre-sumably as a result of activation of pathways dependent on protein

1926 I. Potolicchio et al. / Biochimica et Biophysica Acta 1818 (2012) 1919–1936
kinases A and C [195,196]. A comparable decrease in coupling was ob-served during LH-induced stimulation of testosterone secretion, sug-gesting a tonic inhibitory influence of connexin signaling on theoutput of the steroid hormone [196]. Mice null for Cx43 feature Ley-dig cell which, upon grafting in castrated male hosts, produced con-trol serum levels of androgens, with increased proportions of17betaHSD and 5alphaR activities, indicating that Cx43 is dispensablefor androgen production [197]. In the absence of Cx43, Leydig cellsare still coupled, suggesting that other connexins contribute to theirgap junctions [197], and/or were induced to compensate the loss ofCx43. Together, the available evidence indicates that connexin signal-ing plays an important role, together with the cell-to-cell communi-cations involving paracrine/autocrine signaling, to ensure propercontrol of Sertoli cell synchronization and differentiation and, ulti-mately, of spermatogenesis. However, whether this signaling is es-sential for androgenesis remains to be fully elucidated.
5.3. Ovary
The role and function of gap junctions and connexins in the ovarianfollicle has been the subject of a large amount of work, which has beenrecently reviewed in a number of excellent reviews [198–203].
5.3.1. Oestrogens-producing granulosa and theca cellsThe theca cells derived from the endocrine differentiation of the
granulosa cells, which make the walls of the ovarian follicles, are con-nected by Cx43 [205,206]. The expression of this connexin varies withthe stage of follicle development, consistent with an involvement ofconnexins in the endocrine function of thecal cells [207–209].
In dogs, Cx43 expression changes with the estrus stage in secondaryfollicles, but not in primary and antral follicles, indicating that the signal-ing dependent on this connexin isoform is not involved in the prolongedfollicle and oocyte development which are characteristics of canines[210]. In the sheep, expression of theca Cx43 decreases 24 h after hCGtreatment [211]. The expression of Cx43 is up-regulated by FSH, anddown-regulated by LH, by PKA- and MAPK-dependent effects on Cx43translation [212]. In fetal sheep ovaries, Cx26 and Cx32were immunolo-calized in oocytes, granulosa and theca cells, whereas Cx37 and Cx43were between oocyte and cumulus cells, and Cx43 was most abundantin granulosa and theca cells [213]. A maternal diet enriched in seleniumdecreased Cx26 and increased Cx43 expression [213]. Cx26 and Cx43transcripts are altered during the peri-ovulatory period in sheeps, sug-gesting that the 2 connexins play a role in follicle remodeling [214].After grafting under the kidney capsule of immunodeficient females,chimeric ovaries containing either connexin (Cx43 or Cx37) null mutantoocytes and wild-type granulosa cells, or null granulosa cells and wildtype oocyteswere compared [215]. Ovariesmadewith Cx43 null oocytesandwild-type granulosa cells featured antral follicles enclosing normallydeveloping oocytes, as did ovaries made of wild type oocytes and Cx37null granulosa cells. In contrast, ovaries made with Cx43 null granulosacells or Cx37 null oocytes featured defective follicles and oocyte develop-ment, indicating that lack of Cx43 (in granulosa cells) or Cx37 (in oo-cytes) is sufficient to largely compromise follicle development andoogenesis [215]. Patch clamp and dye injection experiments indicatethat folliculogenesis is impaired in the absence of Cx43-dependentcoupling between granulosa cells [216,217]. Gene expression analysisfurther supports the view that impaired folliculogenesis in mice lackingCx43 is due at least in part to reduced responsiveness of granulosa cellsto oocyte-derived GDF9, stressing an interaction between gap junctionaland paracrine intercellular communication [218].
Cx43 also couples the granulosa cells of cumulus to the oocyte[202,203,204]. Mice of the Zp3Cre;Gja1(lox/lox) line, which featureoocytes lacking Cx43, also feature decreased expression of the con-nexin in cumulus granulosa cell, ovulated normally and form func-tional corpora lutea [219]. Rat follicle exposed to the gap junctionblocker carbenoxolone result in the meiotic maturation of oocytes,
associated to decreased intraoocyte concentrations of cAMP, mimick-ing the mechanism of LH-induced oocyte maturation [220]. WhenCx43 was ectopically expressed in mouse oocytes lacking Cx37, whichhave been shownnot tomaturemeiotically, oocyte growth andmatura-tion were restored, suggesting that Cx43 can replace Cx37 to coupleoocytes and cumulus cells [221]. Human cumulus cells express Cx43plaques, as well as cytoplasmic Cx26, Cx30, Cx30.3, Cx32 and Cx40,and are coupled [222].
5.3.2. Progesterone-producing luteal cellsCells of corporea lutea are connected by Cx43 [224,225]. Pharma-
cological treatment with agonists and antagonists of cAMP, proteinkinase C and calcium showed that drugs enhancing the coupling of lu-teal cells increase progesterone secretion, whereas drugs uncouplingluteal cells decrease steroid release [225]. Consistent with a relation-ship between connexin signaling and steroid production, the experi-mental inhibition of Cx43 expression decreases the LH-inducedsteroid secretion of luteal cells [226]. Mice lacking Cx37, one of theconnexins that forms gap junctions between oocytes and granulosacells, develop numerous abnormal corpora lutea [227]. In vitro, thelevels of Cx43 expression and coupling of luteal cells, which varywith the day of the estrous cycle and culture conditions, correlatewith progesterone secretion [228]. Furthermore, downregulation ofCx43 expression by 2 siRNAs, also gradedly decreases progesteroneproduction [228]. Expression of Cx26, but not Cx32 or Cx43 decreasesduring PGF-induced regression of the sheep corpus luteum, suggest-ing a role in its growth, differentiation, and/or regression [229].Also, in cows injected with the PGF2alpha analog Cloprostenol,Cx26, which is not expressed in luteal cells, increases rapidly duringthe late luteal phase and after luteal regression [230]. Subsequently,Cx43 expressionwas distinctly decreased after luteal regression [230].In sheep, Cx37 is expressed in corporea lutea, at levels dependent onluteal development, and on PGF2alpha-induced luteal regression, in-dicating that Cx37 contributes to luteal tissue growth and differenti-ation [231].
In summary, a large body of experimental evidence supports acentral role of different connexin isoforms, mostly Cx43 and Cx37,in the in vitro and in vivo control of ovarian folliculogenesis, oocytematuration and corpus luteum function, in many of the animal spe-cies which have so far been investigated.
6. Cells producing catecholamine hormones
The chromaffin cells of the adrenal medulla are joined by Cx36-made gap junctions in all animal species so far investigated, as wellas by Cx43 and Cx50 in some species [232–235]. Cx36 accounts forthe cell-to-cell spreading of the [Ca2+]i transients that are driven byaction potentials [232]. This spreading is enhanced after exposure ofa few cells to nicotine, which also triggers the release of catechol-amines, indicating that connexin signaling can amplify the secretionof epinephrine and norepinephrine induced by the synaptic activa-tion of individual cells [232,233]. Interestingly, both the pharmacolog-ical blockade of synaptic transmission, as well as the surgicaldenervation of the adrenal glands resulted in increased coupling ofchromaffin cells, an effect which was also observed in newborn rats ata time when the synaptic transmission of the adrenal gland had notyet fully matured [233].These results indicate that the Cx36-dependentsignaling of chromaffin cells is tonically inhibited by cholinergic synap-tic inputs [233]. During cold stress, the expression of Cx36 and Cx43, aswell as the coupling of chromaffin cells increase, allowing for the inter-cellular transmission of action potentials and Ca2+ signaling [235–237].Electrical stimulation of slices of murine adrenal glands increases thesynchronous secretion of epinephrine and, to a lesser extent of norepi-nephrine, frommultiple cells. This release is modulated by the signalingdependent on nicotinic acetylcholine receptors, but not on muscarinicreceptors or gap junctions [238].

1927I. Potolicchio et al. / Biochimica et Biophysica Acta 1818 (2012) 1919–1936
7. Cells producing pheromones
Pheromone-producing glands are considered the phylogeneticprecursors of vertebrate multicellular endocrine systems, combiningthe ancestral exocrine mode of secretion with the evolved chemicalsignaling, via the vascular or the paracrine route, provided by hor-mones [5,6]. Whether Cx43, the only connexin so far described inthe pheromone-producing cells of rodents [24] and of their humansebaceous glands, which form one of their known derivatives [239],plays any role in the secretion of these volatile chemical messageshas not yet been investigated. However, when viewed in the back-ground of the short half-life of connexins, the presence of an unusual-ly high expression of Cx43 between sebocytes [239] suggests someessential physiological role throughout life time.
8. Connexins contribution to the peripheral action of hormones
Most targets of hormones are at a sizable distance from the glandthat produces the endocrine products, usually preventing the estab-lishment of connexin-dependent signaling between the secretorycells and the cognate effector cells. However, connexins may be in-volved in the cross-talk between these two cell types when theseare in contact. This is the case, for example, of the juxtaglomerular ap-paratus, the endocrine regions of the renal nephrons, which collec-tively play a central role in the control of blood pressure [149,150].The renin-producing cells of the afferent arteriole, which are centralplayers of this apparatus, share Cx40 junctions with both adjacentcompanion cells, and closeby endothelial cells [159]. These endotheli-al cells are also connected by gap junctions to the smooth muscle cellsof afferent arterioles [159]. Rats made hypertensive after clipping ofone renal artery, show a selective increase in kidney renin andCx40, indicating that the connexin is implicated in the control ofrenin secretion, and/or in the vasomotor control of renal vessels[159], a model which also results in changes of aortic Cx43 [152]. Intransgenic mice in which the coding region of Cx43 was replaced bythat of Cx32 [240], wild type and heterozygous mice, which show anormal distribution of Cx43, become hypertensive as a result of in-creased plasma renin levels. In contrast, homozygous littermates, inwhich Cx32 had fully replaced Cx43, retain a normal blood pressure,and control levels of circulating renin [241]. The data provide thefirst support for a mechanism, whereby altered connexin signaling be-tween endothelial cells modifies the functioning of the renin-secretingcells (Fig. 5). The case of the juxtaglomerular apparatus may not beunique. Thus, in the pituitary, the non endocrine folliculo-stellate cellscross-talk with distant endocrine cells by establishing gap junctions,that allow for a rapid and extensive propagation of waves of cytosoliccalcium [63,64,68]. This arrangement provides for an efficient mecha-nism to orchestrate the function of endocrine cell types scatteredthroughout the gland.
9. Hormonal control of connexin expression
Many hormones have been reported in a variety of models to af-fect the transcription, mRNA stability, translation and cytoplasmictrafficking of connexins, as well as the gating and regulation of thecell-to-cell channels formed by these proteins. The effects observedin different studies are variable, and sometimes opposite, reflectingthe essential influence of different cell types, physiological states atthe time of the experiment and different connexin patterns. Sincethe previous reviews, which provided a comprehensive coverage ofthis topic [10–19], several observations have been made.
9.1. Peptides, second messengers and glycosides
Cells of a GnRH-secreting cell line, which express Cx26, increasethe expression of this connexin and dye coupling after treatment
with agents increasing cAMP [35]. Throughout the estrous cycle, theCx43 channels of luteal cells are modulated by agonists and antago-nists of cytosolic calcium, cAMP and protein kinase C [225]. cAMPalso increases gap junction plaques made of Cx43 in adrenal cells[266]. Dibutyril cAMP increases gap junctions of dispersed lutealcells, throughout the estrous cycle [232].
ACTH increases the coupling of thymic epithelial cells [242]. Infu-sion of this hormone, but not dexamethasone, increases Cx43 expres-sion, Cx43 gap junction plaques and cortex size in adrenal glands[183,184,203].
CRH upregulates the expression of Cx43 and the coupling ofIMR32 neuroblastoma cells and primary astrocytes in organotypichippocampal slice cultures, by a mechanism which involves the CRHreceptor, MAPKs and protein kinase A-cAMP response element bind-ing protein. The extent of oxidative stress-induced protein carbonyla-tion and cell death inversely correlate with Cx43 levels, and Cx43contributes to the neuroprotective effects of CRH [243].
Ouabain enhances the expression of Cx32 and the coupling ofMDCK cells [244].
9.2. Steroids
Steroids suppress the expression of Cx26 in the uterine epitheli-um, and Cx43 in the uterine myometrium and stroma, during the re-ceptive phase of implantation. Connexin expression then locallyrecovers under control of factors secreted by the growing thropho-blast [245]. During preimplantation, this induction is due to estro-gens, via the activation of a pathway initiated at estrogen receptorsalpha. During the subsequent embryo implantation and decidualiza-tion, endometrial connexins are upregulated by an estrogen recep-tor-independent pathway, possibly implicating catechol estrogen,prostaglandin F(2α) and interleukin-1β [246].
Hydrocortisone delays gap junction formation in the folliculo-stellatecells of the anterior pituitary [247]. Treatment of castrated rats with ste-roids differently changes the number of gap junctions of these cells,depending on the type of steroid tested and the sex of the animals[248]. The pancreatic islets of rats made insulin resistant by daily injec-tions of dexamethasone, feature an increase in the expression of Cx36and in glucose-stimulated insulin secretion [249],
Ovariectomized rats treated with different ratios of 17beta-estradioland progesterone showa dose- and time-dependent regulation of the ex-pression of uterine Cx26 and Cx43, the prominent connexin of uterinestroma and myometrium, but not of Cx32 [250]. Estradiol and testoster-one modulate the expression of Cx43 in the testis of Rana esculenta,both in vivo and in vitro [251]. 17beta-estradiol alters the phosphorylationof Cx43 in cardiomyocytes, an effectwhich is inhibited by a specific estro-gen receptor antagonist [252]. Progesterone increases Cx43 gap junctionsin luteal cells [253].
9.3. Gonadotrophins
LH and FSHmodulate the coupling of ovarian thecal cells, by regulat-ing Cx43 expression at the transcriptional, translational and post-transla-tional levels [254]. The effects of the two gonadotrophins vary as afunction of the developmental stage of the ovarian follicles [255]. LH in-duces germinal vesicle breakdown in bothmature and immature oocytesof Bufo arenarum, as long as these cells are coupled to follicle cells, by in-ducing the cell-to-cell transfer of IP3 or Ca2+ [256]. LH determines thematuration competence of fish oocytes by favoring the coupling-dependent transfer of gonadotrophin-elicited second messengers fromgranulosa cells to the oocyte [257]. Short-term exposure to FSH, cAMPand various steroids alter Cx43 distribution and coupling in fragmentsof seminiferous tubules and in a Sertoli cell line [258]. Infusion ofchorionic gonadotrophin does not prevent the excessive expression ofCx26 in the stroma of baboon uterus with endometriotic lesions, andthe loss of this connexin in the epithelium [259]. FSH uncouples

1928 I. Potolicchio et al. / Biochimica et Biophysica Acta 1818 (2012) 1919–1936
porcine cumulus-oocyte complexes in vitro [260]. In ewes, the effects ofFSH and LH on the coupling of cumulus cells depends on the stage offollicular development and on the presence of the oocyte [261]. FSHincreases coupling in the granulosa cells of small, but not large follicles,whereas LH and LH+FSH have no effect [261]. LH increases thecoupling of thecal cells of large follicles, whereas FSH or LH+FSH haveno effects [262].
9.4. Thyroid hormones
T3 inhibits the growth of Sertoli cells and increases the levels ofCx43 and Sertoli cell coupling in the testis, by an actin cytoskeleton-dependent pathway [263]. Cardiomyocytes of mice over expressing adominant negative mutant of thyroid hormone receptor-beta developa cardiac hypothyroid phenotype, and feature a reduction in the atrialexpression of Cx40, but not of Cx43, and on atrial depolarization veloc-ity. Hyperthyroid animals show an increase in both the expression ofCx40, and the depolarization velocity [264]. Triiodo-L-thyronine andcAMP, but not 11-ketotestosterone increases the expression of Cx43in the trout testis in vitro [265].
In summary, most hormones can affect connexin expression, gapjunction formation and functional coupling in a variety of cell types,even though a direct effect of a hormone on connexins is establishedin only rare cases. Rather, the observation that a given hormone mayhave different effects in different cell types and/or under differentphysiological conditions, suggests that the hormonal effects on con-nexin signaling may be the result of the pleiotropic cellular eventswhich hormones usually elicit upon binding to their cognatereceptors.
10. Connexins and endocrine disruptors
Endocrine disruptors are chemicals that, in most species, interferewith several endocrine systems of, leading to a variety of biological al-terations andmedical conditions [267,268]. Recent evidence indicatesthat these chemicals also affect connexin expression and function.Thus, the pesticide ioxynil, and its derivative ioxynil octanoate, de-crease the expression of Cx43, its phosphorylation and the functionof the cell-to-cell channels this connexin makes [269]. It is yet notestablished whether these effects account for the effects of these en-docrine disruptors on tumorigenesis and thyroid alterations.
Bisphenol A decreases the fertility of male rats, increases the post-implantation loss of embryos in females, and reduces the size of thenewborn litters. The chemical also reduces spermatogenesis and the ex-pression of Cx43 in the Sertoli cells of testis [270]. Bisphenol-A also re-duces coupling, but not Cx43 expression, and cell viability in cultures ofepithelial rat BICR-M1Rk cells [271]. Low doses of 17a-ethynylestradiol,medroxyprogesterone acetate and levonorgestrel, 3 contraceptive ste-roids discarded in the environment, alter Cx43 trafficking and impaircoupling of testis tubules and a Sertoli cell line in vitro, by a mechanismwhich does not involve estrogen receptors [272]. Neonatal exposure tothe anti-androgen flutamide decreases Cx43 gene expression and de-lays the morphological and functional maturation of adult pig testes,leading to spermatogenic arrest and Leydig cell hyperplasia[273].These observations add endocrine disruptors to the long list ofchemicals that, by a large variety of mechanisms, eventually impacton connexin signaling and coupling [274].
11. Connexins and human endocrine disorders
The abundant information we have summarized provides compel-ling evidence for an obligatory, if not essential role of connexins in theproper function of most of endocrine glands, including those (pancre-atic beta cells and kidney renin cells) that are central to the regulationof blood glucose and pressure. In turn, this consideration implies that,by altering the coupling or other forms of interactions between key
cells, whether of endocrine, vascular or target nature, some connexindefect may contribute to the abnormal multi-organ interactions thatcharacterize endocrine disorders. While there is still little direct evi-dence for such a scenario in humans, a number of considerationsbased on experimental and genetic findings support a likely pathoge-netic role of connexins.
11.1. Diabetes
Correlative observations suggest that Cx36-dependent signalingmaybe significantly involved in the early pathogenesis of type 1 diabetes,specifically in the autoimmune destruction of beta cells by apoptosiswhich is thought to bemediated by increased levels of pro-inflammatorycytokines in the islet environment [275]. Thus, Cx36 null mice are sensi-tized to pharmacological and immune pro-apoptotic conditions, rapidlyloose most of their beta cells and become hyperglycemic as observed intype 1 diabetes [9,94,95]. Conversely, transgenic mice whose beta cellsspecifically over-express Cx36, or other ectopic connexin isoforms, areprotected against these effects [94,95]. Themechanismof this protectionremains to be fully unraveled. Available data show that it depends oncell-to-cell contact, and is not due to loss of activity of the pro-apoptoticcytokines in the islet environment [95]. Rather, a combination of IL-1β,IFN-γ and TNF-α decreases the expression of the Cx36 protein, bydown-regulating the transcription of the cognate gene [95]. Consistentwith these experimental observations, decreased expression of theCx36 transcript has been detected in genome wide scans of type 1 dia-betes models [276,277].
The alterations in insulin secretion observed in Cx36-null mice[76] are similar to those observed in prediabetic states (loss of regularglucose-induced insulin oscillations) and type 2 diabetics (increasedbasal release of insulin, no response to stimulation by post-prandiallevels of glucose) [278,279,289], suggesting that Cx36 may be a rele-vant pathogenetic factor in this disease, which accounts for the vastmajority of the diabetes cases. By necessity, the evidence in supportof this possibility is presently rather indirect, starting with the findingthat human pancreatic beta cells are functionally connected by Cx36[75], as observed in rodents. The human GJD2 gene coding for Cx36,is located on the 14q region of chromosome 15 [280], which is a sus-ceptibility locus for type 2 diabetes, and the diabetic syndrome [281–284]. Strikingly, a single nucleotide polymorphism in the coding se-quence of human GJD2 is pathogenic for a hereditary form of epilepsiathat has a complex pattern of inheritance [285,286], and the samepolymorphism appears unusually frequent in those type 2 diabeticsthat show the more reduced insulin secretion. Eventually, the plasmaparameters which are most affected in type 2 diabetes, i.e. elevatedglucose and free fatty acid levels, may further reduce the expressionof the native, residual Cx36. Thus, chronic exposure to high glucoselevels reduces the expression of Cx36 in insulin-producing cell linesand rat sislets, in a dose- and time-dependent manner, due to a tran-scriptional down-regulation mediated by the highly conserved cAMP-responsive CRE element, in the promoter of the Gjd2 gene [124]. CREis also repressed by high concentrations of palmitate, in vitro andin vivo [123]. Eventually, a sulphonylurea which efficiently stimu-lates insulin release from beta cells of type 2 diabetics, also pro-motes the assembly of Cx36 channels and beta cell coupling[86,106,287,288].
11.2. Hypertension
Cx40-null mice display a chronic hypertension [164–168], whichmimics several characteristics of the most common forms of thehuman disease [149,150], raising the possibility that altered Cx40 sig-naling be implicated in human hypertension. Several circumstantialconsiderations support this possibility. Thus, the similar connexin dis-tribution in the human and the rodent kidneys [155] is consistent withthe possibility that the sustained activation of the renin–angiotensin

1929I. Potolicchio et al. / Biochimica et Biophysica Acta 1818 (2012) 1919–1936
system,which results from the Cx40-dependent alterations in the num-ber, localization and function of the renin secreting cells, may also occurin humans lacking normal Cx40 signaling. Strikingly, a single nucleotidepolymorphism (SNP) of the promoter of the Cx40 gene has been associ-ated with hypertension in selected populations of patients [289–291].This pathogenic SNP is located close to a Ets-1/NK-box/T-box/SP1 anda GATA site [291], 2 elements thought to control basal and inducedCx40 expression [292–295].
The Cx40 null mice also feature alterations in the renal expressionof Cx43 and Cx37 [164], two connexins which are expressed by thesmooth muscle and renin cells of the afferent arteriole, respectively[296]. However, neither Cx37 null mice nor mice in which Cx43 wasconditionally deleted in the vascular tree, feature increased bloodpressure [172–174], demonstrating that Cx40 predominates in thepathogenesis of hypertension. Still, because of their distribution andability to interact with Cx40, both Cx37 and Cx43 may finely tunethe cross-talk between the multiple cell types of the juxta-glomerularapparatus, which sense Na+ concentration, as well as blood flow andpressure. Accordingly, the in vivo replacement of Cx43 by Cx32 [240],a connexin isoform which cannot form channels with Cx40, leads to adown-regulation of renin secretion, and protects the transgenic miceagainst a form of experimental, renin-dependent hypertension [241].Rats made hypertensive after clipping one renal artery, show a selec-tive increase in kidney renin, as well as in the Cx40 and Cx43 levelswithin elastic arteries, implying that these 2 connexins are also impli-cated in the vasomotor control of vessels, which is perturbed duringhypertension [152,159]. Cx40 and Cx43 are the main connexin iso-forms of endothelial and smooth muscle cells, and are altered inboth renin-dependent and renin-independent hypertension. Thesealterations are expected to impair the proper reactivity of the vesselsto perturbations of the hemodynamic conditions, such as those pro-vided by increased blood pressure, inasmuch as experimental evi-dence shows the requirement of junctional channels made of Cx40in the spreading of hyperpolarization along the arterial bed [297–302]. This spreading involves communications between adjacent en-dothelial cells, probably via Cx40–Cx40 channels, as well as betweenendothelial and smooth muscle cells, possibly via Cx40–Cx43 chan-nels [152–159]. Thus, the hypertension observed when Cx40 signal-ing is impaired cannot be solely accounted for by the perturbedrenin production. Rather, it should also take into account the segmen-tal vasoconstriction and altered vasomotion of small, resistance arte-rioles [168,298], which is induced by the sustained activation of theangiotensin production, itself resulting from the excessive reninlevels. Indeed, blockade of either the angiotensin converting enzymeor the angiotensin II AT1 receptors reduce, but does not normalize theblood pressure of Cx40-deficient mice [166]. The necessary participa-tion of peripheral vascular factors is also supported by the observa-tion that Cx40 knock-in Cx45 mice exhibit a modest hypertension,in spite of normal levels of plasma renin [173]. In both cases, the vas-cular component of the hypertension triggered by the absence ofCx40, is probably due to an interruption of the cell-to-cell communi-cation mechanism that normally controls the vasomotor tone[168,298].
11.3. Metabolic syndrome and diabetic complications
In the context of the so-called metabolic syndrome, type 2 diabe-tes is associated to lipid disorders and hypertension [303,304]. Thecombination of long term hyperglycemia, altered levels of free fattyacids and hypertension leads to devastating complications that, inmost cases, result from inadequate vascularization of the target or-gans. Given the widespread distribution of connexins in most tissuesand vessels, and their regulation by several of the metabolic parame-ters which are altered in the metabolic syndrome, their role should beevaluated in the development and/or maintenance of this condition,and of diabetic complications. In vitro studies have shown that a
chronic increase in the levels of environmental glucose reduces thecommunication between both the endothelial and the smoothmusclecells of large vessels, via a PKC-dependent phosphorylation of Cx43[305]. Similar conditions also reduce the expression of Cx43 and thecoupling of microvascular endothelial cells in several tissues, includ-ing the kidney and the retina that are prominent targets of hypergly-cemic complications [306,313]. Also, loss of Cx37 accelerates thedevelopment of intimal lesions in the large vessels of apolipoproteinE-deficient mice, which are prone to develop atherosclerosis [307].Eventually, connexin alterations are consistent markers of coronaryartery disease [152,308–312] and diabetic nephropathy in humans[303,313]. These observations suggest that connexins may contributeto the development and/or maintenance of both micro- and macro-angiopathies. Also, in rats made hyperglycemic by treatment withstreptozotocin, a drug which acutely kills pancreatic beta cells, aPKCε-mediated increase in the phosphorylation of Cx43 sensitizesthis connexin to proteolytic degradation, hence reducing its levels be-tween cardiac myocytes, which results in improper heart contraction[312,314,315]. A similar alteration was also observed in vitro in thepresence of high glucose levels [316]. This set of experiments suggeststhat altered connexin signaling may contribute to the developmentand/or maintenance of the remodeled sequence of ventricular con-traction, which sets the basis for arrhythmias in the diabetic heart.Eventually, alterations in the expression of Cx43 and/or in Cx43-me-diated coupling have been observed in a number of peripheral organsof hyperglycemic, streptozotocin-treated mice, which are frequentsites of diabetic complications, including lens [317], skin [318] andcorpus cavernosum [319]. The latter study provided the first evidencethat selective alterations in the permeability of connexin channelsmay also contribute to dysfunction of peripheral organs in chronicallyhyperglycemic animals [319]. Zucker fatty rats, which represent amodel of type 2 diabetes, feature increased phosphorylation of renalCx43, and decreased expression of Cx37 in renin-secreting cells, com-pared to age- and strain-matched lean rats [320]. Long term diabetesis also frequently complicated by peripheral neuropathies, via a vari-ety of mechanisms which are triggered by the sustained hyperglyce-mia, and which target central and peripheral neurons, as well astheir supportive glial and Schwann cells [321]. There is now abundantexperimental evidence supporting a physio-pathological role of dif-ferent connexins in the function of neurons and glial cells in both cen-tral and peripheral nervous system [322]. The relevance of thesefunctions in humans is exemplified by the finding that multiple muta-tions of Cx32 cause the X-linked form of the Charcot-Marie-Tooth dis-ease [322], and that a SNP of Cx36, which is the connexin accountingfor electrical synapses [322], is pathogenic in monoclonal juvenileepilepsia, a form of epileptic disease that shares with type 2 diabetesa complex, polygenic inheritance [285,286]. Whether connexins aredirectly involved in the development, maintenance and evolution ofdiabetic neuropathy is beginning to be investigated [313]. Earlychanges in the expression of Cx26, Cx32 and Cx43 parallel alter-ations of autonomous innervation in a model of diabetes-relatedbladder dysfunction [323]. In another model, decreased levels ofCx26 and Cx32 have been described in the perineurium of the pe-nile nerve of hyperglycemic rats [324].
11.4. Endocrine tumors
As summarized in several sections of this review, increasing evidenceindicates that connexin signaling significantly influences the differentia-tion, growth and death of different types of endocrine cells, opening thepossibility that abnormal connexin signaling could contribute to the stillobscure pathogenesis of endocrine cancers. So far, only circumstantial,correlative evidence supports this possibility. A statistically significantassociation is observed between the expression of Cx26, tumor sizeand lymph node metastases in human papillary thyroid cancer and fol-licular thyroid cancer [325]. Also, the intra-glandular dissemination of

1930 I. Potolicchio et al. / Biochimica et Biophysica Acta 1818 (2012) 1919–1936
tumor cells is more frequent in Cx26-positive than Cx26-negativehuman papillary carcinomas [325].
Contrasting with the catecholamine-producing cells of the humanadrenal medulla, which normally express Cx50 and some Cx43, thecatecholamine-producing cells of benign pheochromocytomas pre-dominantly express the former connexin isoform, and those of malig-nant adrenal pheochromocytomas express Cx26 and Cx32 [235,326].Mice deficient for Cx32 are sensitized to radiation-induced tumori-genesis and develop pheochromocytomas more frequently [327]. Inbenign adrenocortical adenomas, the number of Cx43-made gapjunctions is reduced in the zona fasciculata, and this number is fur-ther reduced in the malignant forms of the tumors [328]. Normalhuman follicular cells express large amounts of Cx43 in situ and invitro, and this expression is markedly reduced, together with cou-pling, in 4 human ovarian adenocarcinoma cell lines, and in samplesof human serous cystadenocarcinomas [329]. Cx43 expression isalso reduced, if not abolished in human testis infiltrated with in situcarcinomas and in seminomas [185,330,331], an alteration which isusually associated to increased Cx26 expression [331]. The prolifera-tion of the tumoral cells of the JKT-1 seminoma line is reduced byCx43 over expression [332]. In this case, as in a line derived from aLeydig tumor, most of Cx43 is cytoplasmic [332,333].
12. Conclusion
With evolution, the production of hormones by endocrine glandshas come under control of a complex signaling network that inte-grates multiple regulatory mechanisms. The studies summarizedabove provide compelling evidence that, in this network, the cell-to-cell coupling mediated by connexins plays a physiologically rele-vant role in the proper regulation of hormone gene expression, bio-synthesis and release, as well as in the maintenance of adequatenumbers of hormone-producing cells. The experimental observationson a variety of in vitro and in vivo models have unraveled the multi-faceted signaling that is established by different connexins in mostneuro-endocrine systems, which participate to control a vertebrateorganism. The recent observations made in the basic science laborato-ry strongly support the view that connexins significantly contributeto proper regulation of several endocrine secretions, and suggeststhat a dysregulation of connexin signaling could contribute to someof the most frequent endocrine disorders of the human clinic. Thispossibility calls for further investigations on the molecular organiza-tion of the underlying signaling, the cellular mechanisms it controls,and the relationships between the connexin-dependent signalingand other pathways that concur to regulate the function of endocrineand target cells. This knowledge is a prerequisite to evaluate thepatho-physiological role of connexins in several endocrine disorders.In turn, this knowledge will determine the usefulness of a connexin-targeted approach in the development of innovative therapies ofthese diseases.
With regard to the molecular mechanism whereby connexins af-fect the functions of endocrine cells, future studies should identifythe endogenous signals which permeate connexin channels and actas second messengers. The task is complicated by the fact that manyof the endogenous signal molecules and metabolites which permeateconnexin channels are also important players for most types of endo-crine secretions. Therefore, these experiments should also test whichof the permeant ions and molecules is actually dispensable for properendocrine cell function. As connexins may signal cells in a variety ofways, future experiments should further investigate whether connex-ins may contribute to the proper function of endocrine cells by amechanism independent of junctional coupling and, if so, whetherthis implies any role of connexin “hemi-channels”, an interaction ofconnexins with other proteins and specific transcription factors, ortheir effects on the expression of selected genes [10]. It is certainly
conceivable that connexins may not need to assemble into functionalgap junction channels to exert a biological effect. However, much ad-ditional data is still required for the validation of the still putative al-ternative mechanisms. With regard to the cellular mechanisms thatare affected by connexin signaling, we need to understand which sub-populations of endocrine cells, and possibly target cells, cross-talkwith companion cells and/or with other cell types via connexins.Also, the hierarchical position of the connexin-dependent pathway,relative to the other signaling pathways which allow individual cellsto become integrated in a functionally coherent tissue, remains tobe elucidated. With regard to the pathogenesis of endocrine disor-ders, studies aimed at determining whether qualitative and/or quan-titative alterations in connexin expression/function participate in acausal or consequential manner to altered endocrine physiology arenow required in a variety of animal models. This search should notbe limited to genetically encoded disorders, but extend to acquiredand environmental conditions, that have been largely neglected sofar. Obviously, these studies should be extended as much as possibleto patients, at least by taking advantage of the genetic and molecularbiology approaches that are applicable to humans.
These studies should provide important clues as to whethermeans to specifically interact with connexin signaling represent anavenue worth further exploration for correcting structural and func-tional alterations of endocrine glands and secretions. This may in-volve both genetic approaches to correct defects in the expressionof the proteins, and the development of a connexin-targeted pharma-cology. In turn, the latter development requires automatic methodsfor the high-throughput screening of candidate molecules [288]. Theunavailability of such molecules is presently a major limitation inthe connexin field. Eventually, it may be worth considering whetherwe could take advantage of connexins to foster the implementationof a cell therapy of some endocrine disorders, e.g. of diabetes. Thistherapy implies that the surrogate, insulin-producing cells thatare transplanted in the patients, be able to functionally interactwith the native, resident companion cells, in order to ensure an in-sulin secretion commensurate with the requirements of the host. Inview of the data so far gathered in laboratory rodents, proper con-nexin signaling appears as an obligatory requirement to achievesuch a goal.
Given the variety of connexins and connexin-dependent mecha-nisms and functions, the large number of hormones with exquisitesensitivity and specific effects that interact to ensure homeostasis,still are modulated by the ever changing environmental conditions,these challenges are exciting, if formidable. They certainly are nowtimely, and necessary to foster the development of innovative thera-pies of several endocrine disorders, which nowadays show an explod-ing prevalence, worldwide.
Acknowledgment
Our work is supported by grants from the Swiss National ScienceFoundation (310000–122423, 310000–109402, CR32I3_129987), theJuvenileDiabetes Research Foundation (1-2007-158), and the EuropeanUnion (BETAIMAGE 222980; IMIDIA, C2008-T7).
References
[1] E.J. Barrington, The phylogeny of the endocrine system, Experientia 42 (1986)775–781.
[2] K.I. Becker, E.S. Nylen, R.H. Snider, Endocrinology and the endocrine patient, in:K.I. Becker (Ed.), Principles and Practice of Endocrinology and Metabolism, J.B.Lippincott Company, Philadelphia, 1990, pp. 2–13.
[3] R.K. Campbell, N. Satoh, B.M. Degnan, Piecing together evolution of the verte-brate endocrine system, Trends Genet. 20 (2004) 359–366.
[4] A. Gorbman, W.D. Dickoff, S.R. Vigna, N.B. Clark, C.L. Ralph, Comparative Endo-crinology, John Wiley & Sons, Hoboken, USA, 1983, p. 592.
[5] A.M. Stoka, Phylogeny and evolution of chemical communication: an endocrineapproach, J. Mol. Endocrinol. 22 (1999) 207–225.

1931I. Potolicchio et al. / Biochimica et Biophysica Acta 1818 (2012) 1919–1936
[6] D. LeRoith, Are all cells “endocrine”? in: K.L. Becker, et al., (Eds.), Principles andPractice of Endocrinology and Metabolism, JB Lippincott Company, Philadelphia,1990, pp. 10–13.
[7] J.E. Blalock, A molecular basis for bidirectional communication between the im-mune and neuroendocrine systems, Physiol. Rev. 69 (1989) 1–32.
[8] D. LeRoith, G. Delahunty, G.L. Wilson, C.T. Roberts, J. Sheme, C. Hart, M.A. Lesniak, J.Shiloach, J. Roth, Evolutionary aspects of the endocrine and nervous systems, Re-cent Prog. Horm. Res. 42 (1986) 549–587.
[9] J. Roth, D. LeRoith, E.S. Collier, N.R. Weaver, A. Watkinson, C.F. Cleland, S.M.Glick, Evolutionary origins of neuropeptides, hormones, and receptors: possibleapplications to immunology, J. Immunol. 135 (Suppl 2) (1985) 816s–819s.
[10] D. Bosco, J.A. Haefliger J-A, P. Meda, Connexins: key mediators of endocrinefunction, Physiol Rev. 91 (4) (2011) 1393–1445.
[11] P. Meda, D. Bosco, Communication of islet cells: molecules, mechanisms, func-tions, in: J.F. Habener, M. Hussein (Eds.), Molecular Basis of Endocrine PancreasDevelopment and Functions, 2001, pp. 138–159.
[12] V. Serre-Beinier, C. Mas, A. Calabrese, D. Caton, J. Bauquis, D. Caille, A. Charollais,V. Cirulli, P. Meda, Connexins and secretion, Biol. Cell 94 (2002) 477–492.
[13] L. Michon, R. Nlend Nlend, S. Bavamian, L. Bischoff, N. Boucard, D. Caille, J. Cancela,A. Charollais, E. Charpantier, P. Klee, M. Peyrou, C. Populaire, L. Zulianello, P. Meda,Involvement of gap junctional communication in secretion, Biochim. Biophys. Acta1719 (2005) 82–101.
[14] P. Meda, A. Perrelet, L. Orci, Gap junctions and cell-to-cell coupling in endocrineglands, in: B.H. Satir (Ed.), Modern Cell Biology, vol. 3, Alan Liss, New York, 1984,pp. 131–196.
[15] P. Meda, The role of gap junction membrane channels in secretion and hormonalaction, J. Bioenerg. Biomembr. 28 (1996) 369–377.
[16] O.H. Petersen, The Electrophysiology of Gland Cells, Academic Press, London,1980, pp. 2–244.
[17] Y. Munari-Silem, B. Rousset, Gap junction-mediated cell-to-cell communicationin endocrine glands—molecular and functional aspects: a review, Eur. J. Endocri-nol. 135 (1996) 251–264.
[18] P. Meda, D.C. Spray, Gap junction function, Adv. Mol. Cell Biol. 30 (2000) 263–322.[19] S.A. Murray, K. Davis, V. Gay, ACTH and adrenocortical gap junctions, Microsc.
Res. Tech. 61 (2003) 240–246.[20] D. Salomon, P. Meda, Heterogeneity and contact-dependent regulation of hor-
mone secretion by individual B cells, Exp. Cell Res. 162 (1986) 507–520.[21] D. Bosco, L. Orci, P. Meda, Homologous but not heterologous contact increases the
insulin secretion of individual pancreatic B-cells, Exp. Cell Res. 184 (1989) 72–80.[22] D. Bosco, P. Meda, Actively synthesizing beta-cells secrete preferentially after
glucose stimulation, Endocrinology 129 (1991) 3157–3166.[23] S. Bavamian, P. Klee, A. Britan, C. Populaire, D. Caille, J. Cancela, A. Charollais, P.
Meda, Islet-cell-to-cell communication as basis for normal insulin secretion, Di-abetes Obes. Metab. 9 (Suppl 2) (2007) 118–132.
[24] P. Meda, M.S. Pepper, O. Traub, K. Willecke, D. Gros, E. Beyer, B. Nicholson, D.Paul, L. Orci, Differential expression of gap junction connexins in endocrineand exocrine glands, Endocrinology 133 (1993) 2371–2378.
[25] D. Martin, T. Tawadros, L. Meylan, A. Abderrahmani, D.F. Condorelli, G. Waeber,J.A. Haefliger, Critical role of the transcriptional repressor neuron-restrictive si-lencer factor in the specific control of connexin36 in insulin-producing cell lines,J. Biol. Chem. 278 (2003) 53082–53089.
[26] J.M. Rukstalis, A. Kowalik, L. Zhu, D. Lidington, C.L. Pin, S.F. Konieczny, Exocrinespecific expression of Connexin32 is dependent on the basic helix–loop–helixtranscription factor Mist1, J. Cell Sci. 116 (2003) 3315–3325.
[27] R.D. Veenstra, D.A. Beblo, H.Z. Wang, in: R. Werner (Ed.), Gap Junctions, IOSPress, Amsterdam, 1998, pp. 40–44.
[28] G.S. Goldberg, P.D. Lampe, B.J. Nicholson, Selective transfer of endogenous me-tabolites through gap junctions composed of different connexins, Nat. CellBiol. 1 (1999) 457–459.
[29] H. Niessen, H. Harz, P. Bedner, K. Kramer, K.Willecke, Selective permeability of dif-ferent connexin channels to the second messenger inositol 1,4,5-trisphosphate, J.Cell Sci. 113 (2000) 1365–1372.
[30] A.L. Harris, Emerging issues of connexin channels: biophysics fills the gap, Q.Rev. Biophys. 34 (2001) 325–472.
[31] C.P. Taylor, F.E. Dudek, A physiological test for electrotonic coupling betweenCA1 pyramidal cells in rat hippocampal slices, Brain Res. 235 (1982) 351–357.
[32] D.F. Matesic, J.A. Germak, E. Dupont, B.V. Madhukar, Immortalized hypothalamicluteinizing hormone-releasing hormone neurons express a connexin 26-likeprotein and display functional gap junction coupling assayed by fluorescence re-covery after photobleaching, Neuroendocrinology 58 (1993) 485–492.
[33] D. Matesic, B. Attardi, T. Dellovade, D. Pfaff, J. Germak, Differential LHRH secre-tion, dye coupling, and protein expression in two morphologically distinct celltypes identified in GT1–7 cultures, J. Neuroendocrinol. 9 (1997) 467–478.
[34] D.M. Simmons, L.W. Swanson, Comparison of the spatial distribution of seventypes of neuroendocrine neurons in the rat paraventricular nucleus: toward aglobal 3D model, J. Comp. Neurol. 516 (2009) 423–441.
[35] D.F. Matesic, T. Hayashi, J.E. Trosko, J.A. Germak, Upregulation of gap junctionalintercellular communication in immortalized gonadotropin-releasing hormoneneurons by stimulation of the cyclic AMP pathway, Neuroendocrinology 64(1996) 286–297.
[36] T. Funabashi, K. Suyama, T. Uemura, M. Hirose, F. Hirahara, F. Kimura, Immortal-ized gonadotropin-releasing hormone neurons (GT1–7 cells) exhibit synchro-nous bursts of action potentials, Neuroendocrinology 73 (2001) 157–165.
[37] L. Westberg, E. Sawa, A.Y. Wang, L.A. Gunaydin, A.C. Ribeiro, D.W. Pfaff, Coloca-lization of connexin 36 and corticotropin-releasing hormone in the mouse brain,BMC Neurosci. 30 (10) (2009) 41.
[38] Z. Li, G.I. Hatton, Oscillatory bursting of phasically firing rat supraoptic neuronesin low-Ca2+ medium: Na+ influx, cytosolic Ca2+ and gap junctions, J. Physiol.496 (1996) 379–394.
[39] J.I. Nagy, F.E. Dudek, J.E. Rash, Update on connexins and gap junctions in neuronsand glia in the mammalian nervous system, Brain Res. Brain Res. Rev. 47 (2004)191–215.
[40] I. Eisner, J.A. Colombo, Detection of a novel pattern of connexin 43 immunoreac-tivity responsive to dehydration in rat hypothalamic magnocellular nuclei, Exp.Neurol. 177 (2002) 321–325.
[41] S. Bose, G.M. Leclerc, R. Vasquez-Martinez, F.R. Boockfor, Administration of con-nexin43 siRNA abolishes secretory pulse synchronization in GnRH clonal cellpopulations, Mol. Cell. Endocrinol. 314 (2010) 75–83.
[42] S. Hosny, L. Jennes, Identification of gap junctional connexin-32 mRNA and pro-tein in gonadotropin-releasing hormone neurons of the female rat, Neuroendo-crinology 67 (1998) 101–108.
[43] R.E. Campbell, E. Ducret, R. Porteous, X. Liu, M.K. Herde, K. Wellerhaus, S. Sonntag,K. Willecke, A.E. Herbison, Gap junctions between neuronal inputs but not gonad-otropin-releasing hormone neurons control estrous cycles in the mouse, Endocri-nology 152 (2011) 2290–2301.
[44] K.J. Suter, J.P. Wuarin, B.N. Smith, F.E. Dudek, S.M. Moenter, Whole-cell recordingsfrom preoptic/hypothalamic slices reveal burst firing in gonadotropin-releasinghormone neurons identifiedwith green fluorescent protein in transgenic mice, En-docrinology 141 (2000) 3731–3736.
[45] Oka Y. Electrophysiological, characteristics of gonadotrophin-releasing hormone1–3 neurones: insights from a study of fish brains, J. Neuroendocrinol. 22 (2010)659–663.
[46] K. Shinohara, T. Funabashi, T.J. Nakamura, F. Kimura, Effects of estrogen and pro-gesterone on the expression of connexin-36 mRNA in the suprachiasmatic nu-cleus of female rats, Neurosci. Lett. 309 (2001) 37–40.
[47] L. Hu, A.J. Olson, R.I. Weiner, P.C. Goldsmith, Connexin 26 expression and exten-sive gap junctional coupling in cultures of GT1–7 cells secreting gonadotropin-releasing hormone, Neuroendocrinology 70 (1999) 221–227.
[48] R. Vazquez-Martinez, S.L. Shorte, W.J. Faught, D.C. Leaumont, L.S. Frawley, F.R.Boockfor, Pulsatile exocytosis is functionally associatedwithGnRHgene expressionin immortalized GnRH-expressing cells, Endocrinology 142 (2001) 5364–5370.
[49] E. Terasawa, Luteinizing hormone-releasing hormone (LHRH) neurons: mecha-nism of pulsatile LHRH release, Vitam. Horm. 63 (2001) 91–129.
[50] A.C. Charles, S.K. Kodali, R.F. Tyndale, Intercellular calcium waves in neurons,Mol. Cell. Neurosci. 7 (1996) 337–353.
[51] D.F. Matesic, J.A. Germak, E. Dupont, B.V. Madhukar, Immortalized hypothalamicluteinizing hormone-releasing hormone neurons express a connexin 26-likeprotein and display functional gap junction coupling assayed by fluorescence re-covery after photobleaching, Neuroendocrinology 58 (1993) 485–492.
[52] D. Matesic, B. Attardi, T. Dellovade, D. Pfaff, J. Germak, J. Differential, LHRH secre-tion, dye coupling, and protein expression in two morphologically distinct celltypes identified in GT1–7 cultures, J. Neuroendocrinol. 9 (1997) 467–478.
[53] W.C. Wetsel, M.M. Valença, I. Merchenthaler, Z. Liposits, F.J. López, R.I. Weiner,P.L. Mellon, A. Negro-Vilar, Intrinsic pulsatile secretory activity of immortalizedluteinizing hormone-releasing hormone-secreting neurons, Proc. Natl. Acad. Sci.U. S. A. 89 (1992) 4149–4153.
[54] S.S. Stojilkovic, J. Tabak, R. Bertram, Ion channels and signaling in the pituitarygland, Endocr. Rev. 31 (2010) 845–915.
[55] D.J. Lyons, E. Horjales-Araujo, C. Broberger, Synchronized network oscillations inrat tuberoinfundibular dopamine neurons: switch to tonic discharge by thyro-tropin-releasing hormone, Neuron 65 (2010) 217–229.
[56] G.I. Hatton, Z. Li, Intrinsic controls of intracellular calcium and intercellular com-munication in the regulation of neuroendocrine cell activity, Cell. Mol. Neuro-biol. 18 (1998) 13–28.
[57] R.D. Andrew, B.A. MacVicar, F.E. Dudek FE, G.I. Hatton, Dye transfer through gapjunctions between neuroendocrine cells of rat hypothalamus, Science 211(1981) 1187–1189.
[58] F.E. Dudek, R.D. Andrew, B.A. MacVicar, G.I. Hatton, Intracellular electrophysiol-ogy of mammalian peptidergic neurons in rat hypothalamic slices, Fed. Proc. 41(1982) 2953–2958.
[59] P.E. Micevych, P. Popper, G.I. Hatton, Connexin 32 mRNA levels in the rat supra-optic nucleus: up-regulation prior to parturition and during lactation, Neuroen-docrinology 63 (1996) 39–45.
[60] J.C. Saez, V.M. Berthoud, R. Kadle, O. Traub, B.J. Nicholson,M.V. Bennett, R. Dermietzel,Pinealocytes in rats: connexin identification and increase in coupling caused by nor-epinephrine, Brain Res. 568 (1991) 265–275.
[61] N.C. Guérineau, R.A. McKinney, D. Debanne, P. Mollard, B.H. Gähwiler, Organoty-pic cultures of the rat anterior pituitary: morphology, physiology and cell-to-cellcommunication, J. Neurosci. Methods 73 (1997) 169–176.
[62] I. Morand, P. Fonlupt, A. Guerrier, J. Trouillas, A. Calle, C. Remy, B. Rousset, Y.Munari-Silem, Cell-to-cell communication in the anterior pituitary: evidencefor gap junction-mediated exchanges between endocrine cells and folliculostel-late cells, Endocrinology 137 (1996) 3356–3367.
[63] T. Fauquier, N.C. Guerineau, R.A. McKinney, K. Bauer, P. Mollard, Folliculostellatecell network: a route for long-distance communication in the anterior pituitary,Proc. Natl. Acad. Sci. 98 (2001) 8891–8896.
[64] C. Sánchez-Cárdenas, A. Hernández-Cruz, GnRH-induced [Ca2+]i-signallingpatterns in mouse gonadotrophs recorded from acute pituitary slices in vitro,Neuroendocrinology 91 (2010) 239–255.
[65] T. Yamamoto, M.Z. Hossain, E.L. Hertzberg, H. Uemura, L.J. Murphy, J. Nagy, Con-nexin43 in rat pituitary: localization at pituicyte and stellate cell gap junctionsand within gonadotrophs, Histochemistry 100 (1993) 53–64.

1932 I. Potolicchio et al. / Biochimica et Biophysica Acta 1818 (2012) 1919–1936
[66] N.C. Guerineau, X. Bonnefont, L. Stoeckel, P. Mollard, Synchronized spontaneousCa2+ transients in acute anterior pituitary slices, J. Biol. Chem. 273 (1998)10389–10395.
[67] N. Shirasawa, Y.Mabuchi, E. Sakuma,O.Horiuchi, T. Yashiro,M. Kikuchi, Y.Hashimoto,Y. Tsuruo, D.C. Herbert, T. Soji, Intercellular communicationwithin the rat anterior pi-tuitary gland: X. Immunohistocytochemistry of S-100 and connexin 43 of folliculo-stellate cells in the rat anterior pituitary gland, Anat. Rec. A Discov. Mol. Cell. Evol.Biol. 278 (2004) 462–473.
[68] B. Levavi-Sivan, C.L. Bloch, M.J. Gutnick, I.A. Fleidervish, Electrotonic coupling inthe anterior pituitary of a teleost fish, Endocrinology 146 (2005) 1048–1052.
[69] D.J. Hodson, J. Townsend, S.J. Gregory, C. Walters, D.J. Tortonese, Role of prolac-tin in the gonadotroph responsiveness to gonadotrophin-releasing hormoneduring the equine annual reproductive cycle, J. Neuroendocrinol. 22 (2010)509–517.
[70] M.L. Vitale, J. Cardin, N.B. Gilula, M.E. Carbajal, R.M. Pelletier, Dynamics of con-nexin 43 levels and distribution in the mink (Mustela vison) anterior pituitaryare associated with seasonal changes in anterior pituitary prolactin content,Biol. Reprod. 64 (2001) 625–633.
[71] M. Brissova, A.C. Powers, Architecture of pancreatic islets, in: S. Seino, G.I. Bell (Eds.),Pancreatic Beta Cell in Health and Disease, Springer, Germany, 2008, pp. 3–11.
[72] V. Serre-Beinier, S. Le Gurun, N. Belluardo, A. Trovato-Salinaro, A. Charollais, J.A.Haefliger, D.F. Condorelli, P. Meda, Cx36 preferentially connects beta-cells with-in pancreatic islets, Diabetes 49 (2000) 727–734.
[73] J. Degen, C. Meier, R.S. Van Der Giessen, G. Sohl, E. Petrasch-Parwez, S. Urschel, R.Dermietzel, K. Schilling, C.I. De Zeeuw, K. Willecke, Expression pattern of lacZ re-porter gene representing connexin36 in transgenic mice, J. Comp. Neurol. 473(2004) 511–525.
[74] M. Theis, C. Mas, B. Doring, J. Degen, C. Brink, D. Caille, A. Charollais, O. Kruger, A.Plum, V. Nepote, P. Herrera, P. Meda, K. Willecke, Replacement by a lacZ reportergene assigns mouse connexin36, 45 and 43 to distinct cell types in pancreatic is-lets, Exp. Cell Res. 294 (2004) 18–29.
[75] V. Serre-Beinier, D. Bosco, L. Zulianello, A. Charollais, D. Caille, E. Charpantier,B.R. Gauthier, G.R. Diaferia, B.N. Giepmans, R. Lupi, P. Marchetti, S. Deng, L. Buh-ler, T. Berney, V. Cirulli, P. Meda, Cx36 makes channels coupling human pancre-atic beta-cells, and correlates with insulin expression, Hum. Mol. Genet. 18(2009) 428–443.
[76] M.A. Ravier,M. Guldenagel, A. Charollais, A. Gjinovci, D. Caille, G. Söhl, C.B.Wollheim,K.Willecke, J.C.Henquin, P.Meda, Loss of connexin36 channels altersβ-cell coupling,islet synchronization of glucose-induced Ca2+ and insulin oscillations, and basal in-sulin release, Diabetes 54 (2005) 1798–1807.
[77] G.T. Eddlestone, A. Goncalves, J.A. Bangham, E. Rojas, Electrical coupling be-tween cells in islets of Langerhans from mouse, J. Membr. Biol. 77 (1984) 1–14.
[78] T. Kanno, S.O. Gopel, P. Rorsman, M. Wakui, Cellular function in multicellular systemfor hormone-secretion: electrophysiological aspect of studies on alpha-, beta- anddelta-cells of the pancreatic islet, Neurosci. Res. 42 (2002) 79–90.
[79] D. Mears, N.F. Sheppard, I. Atwater, E. Rojas, Magnitude and modulation of pan-creatic beta-cell gap junction electrical conductance in situ, J. Membr. Biol. 146(1995) 163–176.
[80] P. Meda, I. Atwater, A. Goncalves, J.A. Bangham, L. Orci, E. Rojas, The topographyof electrical synchrony among beta-cells in the mouse islet of Langerhans, Q. J.Exp. Physiol. 69 (1984) 719–735.
[81] H.P. Meissner. Electrophysiological evidence for coupling between beta cells ofpancreatic islets. Nature 262(1976) 502–504.
[82] A.P. Moreno, V.M. Berthoud, G. Perez-Palacios, E.M. Perez-Armendariz, Biophys-ical evidence that connexin-36 forms functional gap junction channels betweenpancreatic mouse beta-cells, Am. J. Physiol. Endocrinol. Metab. 288 (2005)E948–E956.
[83] S. Speier, A. Gjinovci, A. Charollais, P. Meda, M. Rupnik, Cx36 - mediated cou-pling reduces beta-cell heterogeneity, confines the stimulating glucose con-centration range and affects insulin release kinetics, Diabetes 56 (2007)1078–1086.
[84] D. Caton, A. Calabrese, C.Mas, V. Serre-Beinier, A. Charollais, D. Caille, R. Zufferey, D.Trono, P. Meda, Lentivirus-mediated transduction of connexin cDNAs shows level-and isoform-specific alterations in insulin secretion of primary pancreatic beta-cells, J. Cell Sci. 116 (2003) 2285–2294.
[85] A. Charollais, G. Gjinovci, J. Huarte, J. Bauquis, A. Nadal, F. Martin, E. Andreu, J.V.Sanchez-Andres, A. Calabrese, D. Bosco, B. Soria, C.B. Wollheim, P.L. Herrera, P.Meda, Junctional communication of pancreatic beta cells contributes to the con-trol of insulin secretion and glucose tolerance, J. Clin. Invest. 106 (2000)235–243.
[86] E. Charpantier, J. Cancela, P. Meda, Beta cells preferentially exchange cationic mol-ecules via connexin 36 gap junction channels, Diabetologia 50 (2007) 2332–2341.
[87] E. Kohen, C. Kohen, B. Thorell, D.H. Mintz, A. Rabinovitch, Intercellular commu-nication in pancreatic islet monolayer cultures: a microfluorometric study, Sci-ence 204 (1979) 862–865.
[88] P. Meda, M. Amherdt, A. Perrelet, L. Orci, Metabolic coupling between culturedpancreatic β-cells, Exp. Cell Res. 133 (1981) 421–430.
[89] P. Meda, I. Atwater, A. Goncalves, J.A. Bangham, L. Orci, E. Rojas, The topographyof electrical synchrony among beta-cells in the mouse islet of Langerhans, Q. J.Exp. Physiol. 69 (1984) 719–735.
[90] P. Meda, R.L. Michaels, P.A. Halban, L. Orci, J.D. Sheridan, In vivo modulation ofgap junctions and dye coupling between B-cells of the intact pancreatic islet, Di-abetes 32 (1983) 858–868.
[91] P. Meda, R.M. Santos, I. Atwater, Direct identification of electrophysiologicallymonitored cells within intact mouse islets of Langerhans, Diabetes 35 (1986)232–236.
[92] I. Quesada, E. Fuentes, E. Andreu, P. Meda, A. Nada, B. Soria, On-line analysis ofgap junctions reveals more efficient electrical than dye coupling between isletcells, Am. J. Physiol. Endocrinol. Metab. 284 (2003) E980–E987.
[93] C.P. Carvalho, H.C. Barbosa, A. Britan, J.C. Santos-Silva, A.C. Boschero, P. Meda, C.B.Collares-Buzato, Beta cell coupling and connexin expression change during thefunctional maturation of rat pancreatic islets, Diabetologia 53 (2010) 1428–1437.
[94] P. Klee, S. Lamprianou, A. Charollais, D. Caille, R. Sarro, M. Cederroth, J.A. Haefliger,P.Meda, Connexin implication in the control ofmurine beta-cell mass, Pediatr. Res.70 (2011) 142–147.
[95] P. Klee, F. Allagnat, H. Pontes, M. Cederroth, A. Charollais, D. Caille, A. Britan, J.A.Haefliger, P. Meda, Connexins protect pancreatic β-cells against apoptosis, J. ClinInvest. 121 (12) (2011) 4870–4879.
[96] E. Giordano, D. Bosco, V. Cirulli, P. Meda, Repeated glucose stimulation revealsdistinct and lasting secretion patterns of individual pancreatic B-cells, J. Clin. In-vest. 87 (1991) 2178–2185.
[97] F.C. Jonkers, J.C. Jonas, P. Gilon, J.C. Henquin, Influence of cell number on thecharacteristics and synchrony of Ca2+ oscillations in clusters of mouse pancre-atic islet cells, J. Physiol. 520 (1999) 839–849.
[98] F.C. Jonkers, J.C. Henquin, Measurements of cytoplasmic Ca2+ in islet cell clus-ters show that glucose rapidly recruits beta-cells and gradually increases the in-dividual cell response, Diabetes 50 (2001) 540–550.
[99] A. Lernmark, The preparation of, and studies on, free cell suspensions frommouse pancreatic islets, Diabetologia 10 (1974) 43143–43148.
[100] J. Philippe, E. Giordano, A. Gjinovci, P. Meda, Cyclic adenosine monophosphateprevents the glucocorticoid-mediated inhibition of insulin gene expression inrodent islet cells, J. Clin. Invest. 90 (1992) 2228–2233.
[101] D. Pipeleers, P.I. in't Veld, E. Maes, M. Van De Winkel, Glucose-induced insulinrelease depends on functional cooperation between islet cells, Proc. Natl. Acad.Sci. U. S. A. 79 (1982) 7322–7325.
[102] D. Salomon, P. Meda, Heterogeneity and contact-dependent regulation of hor-mone secretion by individual B cells, Exp. Cell Res. 162 (1986) 507–520.
[103] D.G. Pipeleers, Heterogeneity in pancreatic beta-cell population, Diabetes 41(1992) 777–781.
[104] F.C. Schuit, P.I. In't Veld, D.G. Pipeleers, Glucose stimulates proinsulin biosynthe-sis by a dose-dependent recruitment of pancreatic beta cells, Proc. Natl. Acad.Sci. U. S. A. 85 (1988) 3865–3869.
[105] F.C. Schuit, D.G. Pipeleers, Regulation of adenosine 3′,5′-monophosphate levelsin the pancreatic B cell, Endocrinology 117 (1985) 834–840.
[106] P. Meda, A. Perrelet, L. Orci, Increase of gap junctions between pancreatic B-cellsduring stimulation of insulin secretion, J. Cell Biol. 82 (1979) 441–448.
[107] A. Calabrese, V. Serre-Beinier, M. Zhang, D. Caton, C. Mas, L.S. Satin, P. Meda,Connexin 36 controls synchronization of Ca2+ oscillations and insulin secretionof MIN6 cells, Diabetes 52 (2003) 417–424.
[108] H. Heimberg, A. De Vos, A. Vandercammen, E. Van Schaftingen, D. Pipeleers, F.Schuit, Heterogeneity in glucose sensitivity among pancreatic beta-cells is cor-related to differences in glucose phosphorylation rather than glucose transport,EMBO J. 12 (1993) 2873–2879.
[109] T.L. Jetton, M.A. Magnuson, Heterogeneous expression of glucokinase amongpancreatic beta cells, Proc. Natl. Acad. Sci. U. S. A. 89 (1992) 2619–2623.
[110] R. Kiekens, P. in ’t Veld, T. Mahler, F. Schuit, M. Van De Winkel, D. Pipeleers, Dif-ferences in glucose recognition by individual rat pancreatic B cells are associatedwith intercellular differences in glucose-induced biosynthetic activity, J. Clin. In-vest. 89 (1992) 117–125.
[111] B. Soria, M. Chanson, E. Giordano, D. Bosco, P. Meda, Ion channels of glucose-responsive and -unresponsive beta-cells, Diabetes 40 (1991) 1069–1078.
[112] Y. Stefan, P. Meda, M. Neufeld, L. Orci, Stimulation of insulin secretion revealsheterogeneity of pancreatic B cells in vivo, J. Clin. Invest. 80 (1987) 175–183.
[113] T.L. Jetton, G. Ying, M.A. Magnuson, D.W. Piston, Quantitative subcellular imag-ing of glucose metabolism within intact pancreatic islets, J. Biol. Chem. 271(1996) 3647–3651.
[114] J.V. Rocheleau, M.S. Remedi, B. Granada, W.S. Head, J.C. Koster, C.G. Nichols, D.W.Piston, Critical role of gap junction coupled K(ATP)channel activity for regulatedinsulin secretion, PLoS Biol. 4 (2006) e26.
[115] P. Meda, D. Bosco, M. Chanson, E. Giordano, L. Vallar, C. Wollheim, L. Orci, Rapidand reversible secretion changes during uncoupling of rat insulin-producingcells, J. Clin. Invest. 86 (1990) 759–768.
[116] R. Bruzzone, P. Meda, The gap junction: a channel for multiple functions? Eur. J.Clin. Invest. 18 (1988) 444–453.
[117] P. Meda, R. Bruzzone, M. Chanson, D. Bosco, L. Orci, Gap junctional couplingmodulates secretion of exocrine pancreas, Proc. Natl. Acad. Sci. U. S. A. 84(1987) 4901–4904.
[118] A. Calabrese, D. Caton, P. Meda, Differentiating the effects of Cx36 and E-cadherinfor proper insulin secretion of MIN6 cells, Exp. Cell Res. 294 (2004) 379–391.
[119] C. Vozzi, S. Ullrich, A. Charollais, J. Philippe, L. Orci, P. Meda, Adequate connexin-mediated coupling is required for proper insulin production, J. Cell Biol. 131(1995) 1561–1572.
[120] K. Wellershaus, J. Degen, J. Deuchars, M. Theis, A. Charollais, D. Caille, B. Gauthier, U.Janssen-Bienhold, S. Sonntag, P. Herrera, P. Meda, K. Willecke, A new conditionalmouse mutant reveals specific expression and functions of connexin36 in neuronsand pancreatic beta-cells, Exp. Cell Res. 314 (2008) 997–1012.
[121] F. Jaques, H. Jousset, A. Tomas, A.L. Prost, C.B. Wollheim, J.C. Irminger, N.Demaurex, P.A. Halban, Dual effect of cell–cell contact disruption on cytosoliccalcium and insulin secretion, Endocrinology 149 (2008) 2494–2505.
[122] S. Le Gurun, D. Martin, A. Formenton, P. Maechler, D. Caille, G. Waeber, P. Meda,J.A. Haefliger, Connexin-36 contributes to control function of insulin-producingcells, J. Biol. Chem. 278 (2003) 37690–37697.

1933I. Potolicchio et al. / Biochimica et Biophysica Acta 1818 (2012) 1919–1936
[123] F. Allagnat, F. Alonso, D. Martin, A. Abderrahmani, G. Waeber, J.A. Haefliger,ICER-1gamma overexpression drives palmitate-mediated connexin36 down-regulation in insulin-secreting cells, J. Biol. Chem. 283 (2008) 5226–5234.
[124] F. Allagnat, D. Martin, D.F. Condorelli, G. Waeber, J.A. Haefliger, Glucose re-presses connexin36 in insulin-secreting cells, J. Cell Sci. 118 (2005)5335–5344.
[125] D.L. Eizirik, M.L. Colli, F. Ortis, The role of inflammation in insulitis and beta-cellloss in type 1 diabetes, Nat. Rev. Endocrinol. 5 (2009) 219–226.
[126] K. Kawai, E. Ipp, L. Orci, A. Perrelet, R.H. Unger, Circulating somatostatin acts onthe islets of Langerhans by way of a somatostatin-poor compartment, Science218 (1982) 477–478.
[127] Y.C. Patel, M. Amherdt, L. Orci, Quantitative electron microscopic autoradiogra-phy of insulin, glucagon, and somatostatin binding sites on islets, Science 217(1982) 1155–1156.
[128] L. Orci, F. Malaisse-Lagae, M. Ravazzola, D. Rouiller, A.E. Renold, A. Perrelet, R.Unger, A morphological basis for intercellular communication betweenalpha- and beta-cells in the endocrine pancreas, J. Clin. Invest. 56 (1975)1066–1070.
[129] A. Nadal, I. Quesada, B. Soria, Homologous and heterologous asynchronicity be-tween identified alpha-, beta- and delta-cells within intact islets of Langerhansin the mouse, J. Physiol. 517 (1999) 85–93.
[130] I. Quesada, M.G. Todorova, P. Alonso-Magdalena, M. Beltrá, E.M. Carneiro, F.Martin, A. Nadal, B. Soria, Glucose induces opposite intracellular Ca2+ concen-tration oscillatory patterns in identified alpha- and beta-cells within intacthuman islets of Langerhans, Diabetes 55 (2006) 2463–2469.
[131] B. Hellman, A. Salehi, E. Gylfe, H. Dansk, E. Grapengiesser, Glucose generates co-incident insulin and somatostatin pulses and antisynchronous glucagon pulsesfrom human pancreatic islets, Endocrinology 150 (2009) 5334–5340.
[132] B. Rousset, Y. Munari, R. Mornex, Iodine metabolism in isolated human thyroidcells, Ann. Endocrinol. 36 (1975) 163–164.
[133] A. Guerrier, P. Fonlupt, I. Morand, R. Rabilloud, C. Audebet, V. Krutovskikh, D.Gros, B. Rousset, Y. Munari-Silem, Gap junctions and cell polarity: connexin32and connexin43 expressed in polarized thyroid epithelial cells assemble intoseparate gap junctions, which are located in distinct regions of the lateral plas-ma membrane domain, J. Cell Sci. 108 (1995) 2609–2617.
[134] L.M. Green, M. LaBue, J.P. Lazarus, J.C. Jennings, Reduced cell–cell communica-tion in experimentally induced autoimmune thyroid disease, Endocrinology137 (1996) 2823–2832 (Full Text via CrossRef | View Record in Scopus | CitedBy in Scopus (14)).
[135] Y. Munari-Silem, C. Audebet, B. Rousset, Hormonal control of cell to cell commu-nication: regulation by thyrotropin of the gap junction-mediated dye transferbetween thyroid cells, Endocrinology 128 (1991) 3299–3309.
[136] L.M. Green, D.K. Murray, D.T. Tran, G.A. Nelson, M.M. Shah, R.A. Luben, A sponta-neously arising mutation in connexin32 with repeated passage of FRTL-5 cellscoincides with increased growth rate and reduced thyroxine release, J. Mol.Endocrinol. 27 (2001) 145–163 (Full Text via CrossRef).
[137] M. Statuto, C. Audebet, H. Tonoli, S. Selmi-Ruby, B. Rousset, Y. Munari-Silem,Restoration of cell-to-cell communication in thyroid cell lines by transfectionwith and stable expression of the connexin-32 gene. Impact on cell proliferationand tissue-specific gene expression, J. Biol. Chem. 272 (1997) 24710–24716.
[138] Y. Munari-Silem, A. Guerrier, C. Fromaget, R. Rabilloud, D. Gros, B. Rousset, Dif-ferential control of connexin-32 and connexin-43 expression in thyroid epithe-lial cells: evidence for a direct relationship between connexin-32 expression andhistiotypic morphogenesis, Endocrinology 135 (1994) 724–734.
[139] H. Tonoli, V. Flachon, C. Audebet, A. Calle, T. Jarry-Guichard, M. Statuto, B.Rousset, Y. Munari-Silem, Formation of three-dimensional thyroid follicle-likestructures by polarized FRT cells made communication competent by transfec-tion and stable expression of the connexin-32 gene, Endocrinology 141 (2000)1403–1413.
[140] E. Nelles, C. Butzler, D. Jung, A. Temme, H.D. Gabriel, U. Dahl, O. Traub, F.Stumpel, K. Jungermann, J. Zielasek, K.V. Toyka, R. Dermietzel, K. Willecke,Defective propagation of signals generated by sympathetic nerve stimulationin the liver of connexin32-deficient mice, Proc. Natl. Acad. Sci. U. S. A. 93(1996) 9565–9570.
[141] F.D. Houghton, E. Thonnissen, G.M. Kidder, C.C. Naus, K. Willecke, E. Winterhager,Doubly mutant mice, deficient in connexin32 and -43, show normal prenatal de-velopment of organs where the two gap junction proteins are expressed in thesame cells, Dev. Genet. 24 (1999) 5–12.
[142] V. Flachon, H. Tonoli, S. Selmi-Ruby, C. Durand, R. Rabilloud, B. Rousset, Y.Munari-Silem, Thyroid cell proliferation in response to forced expression ofgap junction proteins, Eur. J. Cell Biol. 81 (2002) 243–252.
[143] G. Prost, F. Bernier-Valentin, Y. Munari-Silem, S. Selmi-Ruby, B. Rousset, Connexin-32 acts as a downregulator of growth of thyroid gland, Am. J. Physiol. Endocrinol.Metab. 294 (2008) E291–E299.
[144] E. Winterhager, R. Stutenkemper, O. Traub, E. Beyer, K. Willecke, Expression ofdifferent connexin genes in rat uterus during decidualization and at term, Eur.J. Cell Biol. 55 (1991) 133–142.
[145] B. Risek, N.B. Gilula, Spatiotemporal expression of three gap junction gene prod-ucts involved in fetomaternal communication during rat pregnancy, Develop-ment 113 (1991) 165–181.
[146] L. Cronier, B. Bastide, J.C. Herve, J. Deleze, A. Malassine, Gap junctional commu-nication during human trophoblast differentiation: influence of human chorion-ic gonadotropin, Endocrinology 135 (1994) 402–408.
[147] L. Cronier, J.L. Frendo, N. Defamie, G. Pidoux, G. Bertin, J. Guibourdenche, G. Pointis,A. Malassine, Requirement of gap junctional intercellular communication forhuman villous trophoblast differentiation, Biol. Reprod. 69 (2003) 1472–1480.
[148] J.L. Frendo, L. Cronier, G. Bertin, J. Guibourdenche, M. Vidaud, D. Evain-Brion, A.Malassine, Involvement of connexin 43 in human trophoblast cell fusion and dif-ferentiation, J. Cell Sci. 116 (2003) 3413–3421.
[149] H. Castrop, K. Höcherl, A. Kurtz, F. Schweda, V. Todorov, C. Wagner, Physiology ofkidney renin, Physiol. Rev. 90 (2010) 607–673.
[150] M. Paul, A. Poyan Mehr, R. Kreutz, Physiology of local renin–angiotensin sys-tems, Physiol. Rev. 86 (2006) 747–803.
[151] W.G. Forssmann, R. Taugner, Studies on the juxtaglomerular apparatus. V. Thejuxtaglomerular apparatus in Tupaia with special reference to intercellular con-tacts, Cell Tissue Res. 177 (1977) 291–305.
[152] J.A. Haefliger, P. Nicod, P. Meda, Contribution of connexins to the function of thevascular wall, Cardiovasc. Res. 62 (2004) 345–356.
[153] F. Hanner, C.M. Sorensen, N.H. Holstein-Rathlou, J. Peti-Peterdi, Connexins and thekidney, Am. J. Physiol. Regul. Integr. Comp. Physiol. 298 (2010) R1143–R1155.
[154] F. Hanner, J. von Maltzahn, S. Maxeiner, I. Toma, A. Sipos, O. Krüger, K. Willecke,J. Peti-Peterdi, Connexin45 is expressed in the juxtaglomerular apparatus and isinvolved in the regulation of renin secretion and blood pressure, Am. J. Physiol.Regul. Integr. Comp. Physiol. 295 (2008) R371–R380.
[155] L. Kurtz, K. Madsen, B. Kurt, B.L. Jensen, S. Walter, B. Banas, C. Wagner, A. Kurtz,High-level connexin expression in the human juxtaglomerular apparatus, Neph-ron Physiol. 116 (2010) 1–8.
[156] R. Taugner, H. Kirchheim, W.G. Forssmann, Myoendothelial contacts in glomerulararterioles and in renal interlobular arteries of rat, mouse and Tupaia belangeri, CellTissue Res. 235 (1984) 319–325.
[157] C. Wagner, Function of connexins in the renal circulation, Kidney Int. 73 (2008)547–555.
[158] J. Yao, T. Oite, M. Kitamura, Gap junctional intercellular communication in thejuxtaglomerular apparatus, Am. J. Physiol. Renal Physiol. 296 (2009) F939–F946.
[159] J.A. Haefliger, S. Demotz, O. Braissant, E. Suter, B. Waeber, P. Nicod, P. Meda, Con-nexins 40 and 43 are differentially regulated within the kidneys of rats with re-novascular hypertension, Kidney Int. 60 (2001) 190–201.
[160] N.M. Rummery, C.E. Hill, Vascular gap junctions and implications for hyperten-sion, Clin. Exp. Pharmacol. Physiol. 31 (2004) 659–667.
[161] M.J. Ryan, B. Liu, M.T. Herbowy, K.W. Gross, G. Hajduczok, Intercellular commu-nication between renin expressing As4.1 cells, endothelial cells and smoothmuscle cells, Life Sci. 72 (2003) 1289–1301.
[162] L. Kurtz, F. Schweda, C. de Wit, W. Kriz, R. Witzgall, R. Warth, A. Sauter, A. Kurtz,C. Wagner, Lack of connexin 40 causes displacement of renin-producing cellsfrom afferent arterioles to the extraglomerular mesangium, J. Am. Soc. Nephrol.18 (2007) 1103–1111.
[163] C.P. Bührle, R. Nobiling, E. Mannek, D. Schneider, E. Hackenthal, R. Taugner, Theafferent glomerular arteriole: immunocytochemical and electrophysiological in-vestigations, J. Cardiovasc. Pharmacol. 6 (Suppl 2) (1984) S383–S393.
[164] N. Krattinger, A. Capponi, L. Mazzolai, J.F. Aubert, D. Caille, P. Nicod, G. Waeber, P.Meda, J.A. Haefliger, Connexin40 regulates renin production and blood pressure,Kidney Int. 72 (2007) 814–822.
[165] C. Wagner, C. deWit, L. Kurtz, C. Grünberger, A. Kurtz, F. Schweda, Connexin40 isessential for the pressure control of renin synthesis and secretion, Circ. Res. 100(2007) 556–563.
[166] F. Alonso, N. Krattinger, L. Mazzolai, A. Simon, G. Waeber, P. Meda, J.A. Haefliger,An angiotensin II- and NF-kappaB-dependent mechanism increases connexin 43in murine arteries targeted by renin-dependent hypertension, Cardiovasc. Res.87 (2010) 166–176.
[167] N. Krattinger, F. Alonso, A. Capponi, L. Mazzolai, P. Nicod, P. Meda, J.A. Haefliger,Increased expression of renal cyclooxygenase-2 and neuronal nitric oxidesynthase in hypertensive Cx40-deficient mice, J. Vasc. Res. 46 (2009) 188–198.
[168] A.M. Simon, D.A. Goodenough, D.L. Paul, Mice lacking connexin40 have cardiacconduction abnormalities characteristic of atrioventricular block and bundlebranch block, Curr. Biol. 8 (1988) 295–298.
[169] A.J. Wagner, N.H. Holstein-Rathlou, D.J. Internephron, coupling by conducted va-somotor responses in normotensive and spontaneously hypertensive rats, Am. J.Physiol. 272 (1997) F372–F379.
[170] C. Wagner, A. Jobs, F. Schweda, L. Kurtz, M.L. Lopez, R.A. Gomez, T.A. van Veen, C.de Wit, A. Kurtz, Selective deletion of Connexin 40 in renin-producing cells im-pairs renal baroreceptor function and is associated with arterial hypertension,Kidney Int. 78 (2010) 762–768.
[171] W.H. Beierwaltes, The role of calcium in the regulation of renin secretion, Am. J.Physiol. Renal Physiol. 298 (2010) F1–F11.
[172] B. Kurt, L. Kurtz, M.L. Sequeira-Lopez, R.A. Gomez, K.Willecke, C. Wagner, A. Kurtz,Reciprocal expression of connexin 40 and 45 during phenotypical changes inrenin-secreting cells, Am. J. Physiol. Renal Physiol. 300 (2011) F743–F748.
[173] F. Schweda, L. Kurtz, C. de Wit, U. Janssen-Bienhold, A. Kurtz, C. Wagner, Substi-tution of connexin40 with connexin45 prevents hyperreninemia and attenuateshypertension, Kidney Int. 75 (2009) 482–489.
[174] A.M. Simon, A.R. McWhorter, Decreased intercellular dye-transfer and downre-gulation of non-ablated connexins in aortic endothelium deficient in con-nexin37 or connexin40, J. Cell Sci. 116 (2003) 2223–2236.
[175] C. Wagner, L. Kurtz, F. Schweda, A.M. Simon, A. Kurtz, Connexin 37 is dispens-able for the control of the renin system and for positioning of renin-producingcells in the kidney, Pflugers Arch. 459 (2009) 151–158.
[176] M. Kihara, S. Umemura, T. Sugaya, Y. Toya, M. Yabana, S. Kobayash, K. Tamura, T.Kadota, R. Kishida, K. Murakami, A. Fukamizu, M. Ishii, Expression of neuronaltype nitric oxide synthase and renin in the juxtaglomerular apparatus of angio-tensin type-1a receptor gene-knockout mice, Kidney Int. 53 (1998) 1585–1593.
[177] M.L. Lopez, R.A. Gomez, The renin phenotype: roles and regulation in the kidney,Curr. Opin. Nephrol. Hypertens. 19 (2010) 366–371.

1934 I. Potolicchio et al. / Biochimica et Biophysica Acta 1818 (2012) 1919–1936
[178] M.L. Sequeira López, E.S. Pentz, T. Nomasa, O. Smithies, R.A. Gomez, Renin cellsare precursors for multiple cell types that switch to the renin phenotype whenhomeostasis is threatened, Dev. Cell 6 (2004) 719–728.
[179] S.A. Murray, B.M. Nickel, V.L. Gay, Gap junctions as modulators of adrenal corti-cal cell proliferation and steroidogenesis, Mol. Cell. Endocrinol. 300 (2009)51–56.
[180] S.A. Murray, K. Davis, V. Gay, ACTH and adrenocortical gap junctions, Microsc.Res. Tech. 61 (2003) 240–246.
[181] S.A. Murray, A. Oyoyo, Y. Pharrams, N.M. Kumar, N.B. Gilula, Characterization ofgap junction expression in the adrenal gland, Endocr. Res. 21 (1995) 221–229.
[182] K.T. Davis, N. Prentice, V.L. Gay, S.A. Murray, Gap junction proteins and cell–cellcommunication in the three functional zones of the adrenal gland, J. Endocrinol.173 (2002) 13–21.
[183] Y. Munari-Silem, M.C. Lebrethon, I. Morand, B. Rousset, J.M. Saez, Gap junction-mediated cell-to-cell communication in bovine and human adrenal cells. A pro-cess whereby cells increase their responsiveness to physiological corticotropinconcentrations, J. Clin. Invest. 95 (1995) 1429–1439.
[184] U.A. Oyoyo, U.S. Shah, S.A. Murray, The role of alpha1 (connexin-43) gap junctionexpression in adrenal cortical cell function, Endocrinology 138 (1997) 5385–5397.
[185] S.A. Murray, U.S. Shah, Modulation of adrenal gap junction expression, Horm.Metab. Res. 30 (1998) 426–431.
[186] S.H. Huang, J.C.Wu,R.D. Hwang, H.L. Yeo, S.M.Wang, Effects of 18beta-glycyrrhetinicacid on the junctional complex and steroidogenesis in rat adrenocortical cells, J. Cell.Biochem. 90 (2003) 33–41.
[187] U.S. Shah, S.A. Murray, Bimodal inhibition of connexin 43 gap junctions de-creases ACTH-induced steroidogenesis and increases bovine adrenal cell popu-lation growth, J. Endocrinol. 171 (2001) 199–208.
[188] G. Pointis, J. Gilleron, D. Carette, D. Segretain, Physiological and physiopatholog-ical aspects of connexins and communicating gap junctions in spermatogenesis,Philos. Trans. R. Soc. Lond. B Biol. Sci. 365 (2010) 1607–1620.
[189] X. Decrouy, J.M. Gasc, G. Pointis, D. Segretain, Functional characterization ofCx43 based gap junctions during spermatogenesis, J. Cell. Physiol. 200 (2004)146–154.
[190] K. Weider, M. Bergmann, S. Giese, F. Guillou, K. Failing, R. Brehm. Altered differ-entiation and clustering of Sertoli cells in transgenic mice showing a Sertoli cellspecific knockout of the connexin 43 gene. Differentiation 82 (2011) 38–49.
[191] W.A. Roscoe, K.J. Barr, A.A. Mhawi, D.K. Pomerantz, G.M. Kidder, Failure of sper-matogenesis in mice lacking connexin43, Biol. Reprod. 65 (2001) 829–838.
[192] M.W. Li, D.D. Mruk, W.M. Lee, C.Y. Cheng, Connexin 43 is critical to maintain thehomeostasis of the blood–testis barrier via its effects on tight junction reassem-bly, Proc. Natl. Acad. Sci. U. S. A. 107 (2010) 17998–18003.
[193] K. Marchlewska, K. Kula, R. Walczak-Jedrzejowska, E. Oszukowska, E. Filipiak, J.Slowikowska-Hilczer, Role of FSH and triiodothyronine in Sertoli cell develop-ment expressed by formation of connexin 43-based gap junctions, J. Exp. Zool.A Ecol. Genet. Physiol. 315 (2011) 329–336.
[194] E.M. Perez-Armendariz, M.C. Romano, J. Luna, C. Miranda, M.V. Bennett, A.P.Moreno, Characterization of gap junctions between pairs of Leydig cells frommouse testis, Am. J. Physiol. 267 (1994) C570–C580.
[195] S. You, W. Li, T. Lin, Expression and regulation of connexin43 in rat Leydig cells, J.Endocrinol. 166 (2000) 447–453.
[196] R.C. Goldenberg, F.S. Fortes, J.M. Cristancho, M.M.Morales, C.R. Franci, W.A. Varanda,A.C. Campos deCarvalho,Modulationof gap junctionmediated intercellular commu-nication in TM3 Leydig cells, J. Endocrinol. 177 (2003) 327–335.
[197] C.N. Kahiri, M.W. Khalil, F. Tekpetey, G.M. Kidder, Leydig cell function in micelacking connexin43, Reproduction 132 (2006) 607–616.
[198] G.M. Kidder, A.A. Mhawi, Gap junctions and ovarian folliculogenesis, Reproduc-tion 123 (2002) 613–620.
[199] R.J. Scaramuzzi, D.T. Baird, B.K. Campbell, M.A. Driancourt, J. Dupont, J.E. Fortune,R.B. Gilchrist, G.B. Martin, K.P. McNatty, A.S. McNeilly, P. Monget, D. Monniaux,C. Viñoles, R. Webb, Regulation of folliculogenesis and the determination of ovu-lation rate in ruminants, Reprod. Fertil. Dev. 23 (2011) 444–467.
[200] E. Gershon, V. Plaks, N. Dekel, Gap junctions in the ovary: expression, localiza-tion and function, Mol. Cell. Endocrinol. 282 (2008) 18–25.
[201] I. Granot, N. Dekel, The ovarian gap junction protein connexin43: regulation bygonadotropins, Trends Endocrinol. Metab. 13 (2002) 310–313.
[202] R.B. Gilchrist, Recent insights into oocyte–follicle cell interactions provide op-portunities for the development of new approaches to in vitro maturation,Reprod. Fertil. Dev. 23 (2011) 23–31.
[203] G.M. Kidder, B.C. Vanderhyden, Bidirectional communication between oocytesand follicle cells: ensuring oocyte developmental competence, Can. J. Physiol.Pharmacol. 88 (2010) 399–413.
[204] Y. Yamamoto, G. Yoshizaki, Heterologous gap junctions between granulosa cellsand oocytes in ayu (Plecoglossus altivelis): formation and role during luteinizinghormone-dependent acquisition of oocyte maturational competence, J. Reprod.Dev. 54 (2008) 1–5.
[205] B. Risek, S. Guthrie, N. Kumar, N.B. Gilula,Modulation of gap junction transcript andprotein expression during pregnancy in the rat, J. Cell Biol. 110 (1990) 269–282.
[206] A.M. Simon, D.A. Goodenough, E. Li, D.L. Paul, Female infertility in mice lackingconnexin 37, Nature 385 (1997) 525–529.
[207] G.M. Kidder, A.A. Mhawi, Gap junctions and ovarian folliculogenesis, Reproduc-tion 123 (2002) 613–620.
[208] F.G. Klinger, M. De Felici, In vitro development of growing oocytes from fetalmouse oocytes: stage-specific regulation by stem cell factor and granulosacells, Dev. Biol. 244 (2002) 85–95.
[209] M.L. Johnson, D.A. Redmer, L.P. Reynolds, J.J. Bilski, A.T. Grazul-Bilska, Gap junc-tional intercellular communication of bovine granulosa and thecal cells from
antral follicles: effects of luteinizing hormone and follicle-stimulating hormone,Endocrine 18 (2002) 261–270.
[210] L.A. Willingham-Rocky, M.C. Golding, J.M. Wright, D.C. Kraemer, M.E. Westhusin,R.C. Burghardt, Cloning of GJA1 (connexin43) and its expression in canine ovarianfollicles throughout the estrous cycle, Gene Expr. Patterns 7 (2007) 66–71.
[211] E. Borowczyk, M.L. Johnson, J.J. Bilski, P.P. Borowicz, D.A. Redmer, L.P. Reynolds,A.T. Grazul-Bilska, Expression of gap junctional connexins 26, 32, and 43 mRNAin ovarian preovulatory follicles and corpora lutea in sheep, Can. J. Physiol. Phar-macol. 84 (2006) 1011–1020.
[212] Y. Kalma, I. Granot, D. Galiani, A. Barash, N. Dekel, Luteinizing hormone-inducedconnexin 43 down-regulation: inhibition of translation, Endocrinology 145(2004) 1617–1624.
[213] A.T. Grazul-Bilska, K.A. Vonnahme, J.J. Bilski, E. Borowczyk, D. Soni, B. Mikkelson,M.L. Johnson, L.P. Reynolds, D.A. Redmer, J.S. Caton, Expression of gap junctionalconnexin proteins in ovine fetal ovaries: Effects of maternal diet, Domest. Anim.Endocrinol. 41 (2011) 185-194.
[214] E. Borowczyk, M.L. Johnson, J.J. Bilski, P.P. Borowicz, D.A. Redmer, L.P. Reynolds,A.T. Grazul-Bilska, Expression of gap junctional connexins 26, 32, and 43 mRNAin ovarian preovulatory follicles and corpora lutea in sheep, Can. J. Physiol. Phar-macol. 84 (2006) 1011–1020.
[215] J.E. Gittens, G.M. Kidder, Differential contributions of connexin37 and con-nexin43 to oogenesis revealed in chimeric reaggregated mouse ovaries, J. CellSci. 118 (2005) 5071–5078.
[216] D. Tong, J.E. Gittens, G.M. Kidder, D. Bai, Patch-clamp study reveals that the im-portance of connexin43-mediated gap junctional communication for ovarianfolliculogenesis is strain specific in the mouse, Am. J. Physiol. Cell Physiol. 290(2006) C290–C297.
[217] J.E. Gittens, A.A. Mhawi, D. Lidington, Y. Ouellette, G.M. Kidder, Functional anal-ysis of gap junctions in ovarian granulosa cells: distinct role for connexin43 inearly stages of folliculogenesis, Am. J. Physiol. Cell Physiol. 284 (2003)C880–C887.
[218] J.E. Gittens, K.J. Barr, B.C. Vanderhyden, G.M. Kidder, Interplay between para-crine signaling and gap junctional communication in ovarian follicles, J. CellSci. 118 (2005) 113–122.
[219] E. Gershon, V. Plaks, I. Aharon, D. Galiani, Y. Reizel, S. Sela-Abramovich, I. Granot, E.Winterhager, N. Dekel, Oocyte-directed depletion of connexin43 using the Cre-LoxP system leads to subfertility in female mice, Dev. Biol. 313 (2008) 1–12.
[220] S. Sela-Abramovich, I. Edry, D. Galiani, N. Nevo, N. Dekel, Disruption of gap junc-tional communication within the ovarian follicle induces oocyte maturation, En-docrinology 147 (2006) 2280–2286.
[221] T.Y. Li, D. Colley, K.J. Barr, S.P. Yee, G.M. Kidder, Rescue of oogenesis in Cx37-nullmutant mice by oocyte-specific replacement with Cx43, J. Cell Sci. 120 (2007)4117–4125.
[222] H.X. Wang, D. Tong, F. El-Gehani, F.R. Tekpetey, G.M. Kidder, Connexin expres-sion and gap junctional coupling in human cumulus cells: contribution to em-bryo quality, J. Cell. Mol. Med. 13 (2009) 972–984.
[223] A. Mayerhofer, R.E. Garfield, Immunocytochemical analysis of the expression ofgap junction protein connexin 43 in the rat ovary, Mol. Reprod. Dev. 41 (1995)331–338.
[224] F.S. Khan-Dawood, J. Yang, M.Y. Dawood, Expression of gap junction proteinconnexin-43 in the human and baboon (Papio anubis) corpus luteum, J. Clin.Endocrinol. Metab. 81 (1996) 835–842.
[225] A.T. Grazul-Bilska, L.P. Reynolds, J.J. Bilski, D.A. Redmer, Effects of second mes-sengers on gap junctional intercellular communication of ovine luteal cellsthroughout the estrous cycle, Biol. Reprod. 65 (2001) 777–783.
[226] F.S. Khan-Dawood, J. Yang, M.Y. Dawood, Hormonal regulation of connexin-43in baboon corpora lutea, J. Endocrinol. 157 (1998) 405–414.
[227] A.M. Simon, D.A. Goodenough, E. Li, D.L. Paul, Female infertility in mice lackingconnexin 37, Nature 385 (1997) 525–529.
[228] E. Borowczyk, M.L. Johnson, J.J. Bilski, M.A. Bilska, D.A. Redmer, L.P. Reynolds,A.T. Grazul-Bilska, Role of gap junctions in regulation of progesterone secretionby ovine luteal cells in vitro, Reproduction 133 (2007) 641–651.
[229] S.V. Seekallu, B.M. Toosi, A.T. Grazul-Bilska, N.C. Rawlings, Markers of ovarianantral follicular development in sheep: comparison of follicles destined to ovu-late from the final or penultimate follicular wave of the estrous cycle, Reproduc-tion 140 (2010) 559–568.
[230] B. Berisha, P. Bridger, A. Toth, H.H. Meyer, D. Schams, C. Pfarrer, Expression andlocalization of gap junctional connexins 26 and 43 in bovine periovulatory folli-cles and in corpus luteum during different functional stages of oestrous cycleand pregnancy, Reprod. Domest. Anim. 44 (2009) 295–302.
[231] I. Granot, E. Bechor, A. Barash, N. Dekel, Connexin43 in rat oocytes: developmen-tal modulation of its phosphorylation, Biol. Reprod. 66 (2002) 568–573.
[232] A.O. Martin, M.N. Mathieu, C. Chevillard, N.C. Guerineau, Gap junctions mediateelectrical signaling and ensuing cytosolic Ca2+ increases between chromaffincells in adrenal slices: a role in catecholamine release, J. Neurosci. 21 (2001)5397–5405.
[233] A.O. Martin, M.N. Mathieu, N.C. Guerineau, Evidence for long-lasting cholinergiccontrol of gap junctional communication between adrenal chromaffin cells, J.Neurosci. 23 (2003) 3669–3678.
[234] J. Degen, C. Meier, R.S. Van Der Giessen, G. Sohl, E. Petrasch-Parwez, S. Urschel, R.Dermietzel, K. Schilling, C.I. De Zeeuw, K. Willecke, Expression pattern of lacZ re-porter gene representing connexin36 in transgenic mice, J. Comp. Neurol. 473(2004) 511–525.
[235] C. Colomer, M.G. Desarménien, N.C. Guérineau, Revisiting the stimulus-secretioncoupling in the adrenal medulla: role of gap junction-mediated intercellularcommunication, Mol. Neurobiol. 40 (2009) 87–100.

1935I. Potolicchio et al. / Biochimica et Biophysica Acta 1818 (2012) 1919–1936
[236] C. Colomer, L.A. Olivos Ore, N. Coutry, M.N. Mathieu, S. Arthaud, P. Fontanaud, I.Iankova, F. Macari, E. Thouënnon, L. Yon, Y. Anouar, N.C. Guérineau, Functionalremodeling of gap junction-mediated electrical communication between adre-nal chromaffin cells in stressed rats, J. Neurosci. 28 (2008) 6616–6626.
[237] N.C. Guérineau, M.G. Desarménie, Developmental and stress-induced remodel-ing of cell–cell communication in the adrenal medullary tissue, Cell. Mol. Neuro-biol. 30 (2010) 1425–1431.
[238] J. Petrovic, P.L. Walsh, K.T. Thornley, C.E. Miller, R.M. Wightman, Real-time mon-itoring of chemical transmission in slices of the murine adrenal gland, Endocri-nology 151 (2010) 1773–1783.
[239] D. Salomon, E. Masgrau, S. Vischer, S. Ullrich, E. Dupont, P. Sappino, J.H. Saurat, P.Meda, Topography of mammalian connexins in human skin, J. Invest. Dermatol.103 (1994) 240–247.
[240] A. Plum, G.Hallas, T.Magin, F. Dombrowski, A. Hagendorff, B. Schumacher, C.Wolpert,J. Kim,W.H. Lamers, M. Evert, P. Meda, O. Traub, K.Willecke, Unique and shared func-tions of different connexins in mice, Curr. Biol. 10 (2000) 1083–1091.
[241] J.A. Haefliger, N. Krattinger, D. Martin, T. Pedrazzini, A. Capponi, B. Doring, A.Plum, A. Charollais, K. Willecke, P. Meda, Connexin43-dependent mechanismmodulates renin secretion and hypertension, J. Clin. Invest. 116 (2006) 405–413.
[242] W. Savino, E. Arzt, M. Dardenne, Immunoneuroendocrine connectivity: the par-adigm of the thymus-hypothalamus/pituitary axis, Neuroimmunomodulation 6(1999) 126–136.
[243] R. Hanstein, J. Trotter, C. Behl, A.B. Clement, Increased connexin 43 expression as apotential mediator of the neuroprotective activity of the corticotropin-releasinghormone, Mol. Endocrinol. 23 (2009) 1479–1493.
[244] I. Larre, A. Ponce, R. Fiorentino, L. Shoshani, R.G. Contreras, M. Cereijido, Contactsand cooperation between cells depend on the hormone ouabain, Proc. Natl.Acad. Sci. U. S. A. 103 (2006) 10911–10916.
[245] R. Grummer, E. Winterhager, Regulation of gap junction connexins in the endo-metrium during early pregnancy, Cell Tissue Res. 293 (1998) 189–194.
[246] R. Grummer, S.W. Hewitt, O. Traub, K.S. Korach, E. Winterhager, Different regulatorypathways of endometrial connexin expression: preimplantation hormonal-mediatedpathway versus embryo implantation-initiated pathway, Biol. Reprod. 71 (2004)273–281.
[247] E. Sakuma, D.C. Herbert, T. Soji, The effects of sex steroids on the formation gapjunctions between folliculo-stellate cells; a study in castrated male rats andovariectomized female rats, Arch. Histol. Cytol. 66 (2003) 229–238.
[248] E. Sakuma, T. Soji, D.C. Herbert, Effects of hydrocortisone on the formation of gapjunctions and the abnormal growth of cilia within the rat anterior pituitarygland: possible role of gap junctions on the regulation of cell development,Anat. Rec. 262 (2001) 169–175.
[249] A. Rafacho, L.P. Roma, S.R. Taboga, A.C. Boschero, J.R. Bosqueiro, Dexametha-sone-induced insulin resistance is associated with increased connexin 36mRNA and protein expression in pancreatic rat islets, Can. J. Physiol. Pharmacol.85 (2007) 536–545.
[250] R. Grummer, O. Traub, E. Winterhager, Gap junction connexin genes cx26 andcx43 are differentially regulated by ovarian steroid hormones in rat endometri-um, Endocrinology 140 (1999) 2509–2516.
[251] G. Izzo, D. Ferrara, G. Delrio, M. d'Istria, S. Minucci, Connexin43 expression in thetestis of frog Rana esculenta: sex hormonal regulation, Ann. N. Y. Acad. Sci. 1163(2009) 425–427.
[252] T.H. Chung, S.M. Wang, J.Y. Liang, S.H. Yang, J.C. Wu, The interaction of estrogenreceptor alpha and caveolin-3 regulates connexin43 phosphorylation in meta-bolic inhibition-treated rat cardiomyocytes, Int. J. Biochem. Cell Biol. 41 (2009)2323–2333.
[253] E. Borowczyk, M.L. Johnson, J.J. Bilski, M.A. Bilska, D.A. Redmer, L.P. Reynolds,A.T. Grazul-Bilska, Role of gap junctions in regulation of progesterone secretionby ovine luteal cells in vitro, Reproduction 133 (2007) 641–651.
[254] I. Granot, N. Dekel, The ovarian gap junction protein connexin43: regulation bygonadotropins, Trends Endocrinol. Metab. 13 (2001) 310–313.
[255] M.L. Johnson, D.A. Redmer, L.P. Reynolds, J.J. Bilski, A.T. Grazul-Bilska, Gap junc-tional intercellular communication of bovine granulosa and thecal cells from an-tral follicles: effects of luteinizing hormone and follicle-stimulating hormone,Endocrine 18 (2001) 261–270.
[256] G.S. Toranzo, J. Oterino, L. Zelarayán, F. Bonilla, M.I. Bühler, Spontaneous and LH-induced maturation in Bufo arenarum oocytes: importance of gap junctions, Zy-gote 15 (2007) 65–80.
[257] Y. Yamamoto, G. Yoshizaki, Heterologous gap junctions between granulosa cellsand oocytes in ayu (Plecoglossus altivelis): formation and role during luteinizinghormone-dependent acquisition of oocyte maturational competence, J. Reprod.Dev. 54 (2008) 1–5.
[258] J. Gilleron, D. Carette, F. Carpentier, D. Segretain, G. Pointis, Three-dimensionalanalysis of connexin 43 gap junction in the ex vivo rat seminiferous tubules:short-term effects of hormonal effectors, Microsc. Res. Tech. 72 (2009)845–855.
[259] E. Winterhager, R. Grümmer, P.A. Mavrogianis, C.J. Jones, J.M. Hastings, A.T.Fazleabas, Connexin expression pattern in the endometrium of baboons is influ-enced by hormonal changes and the presence of endometriotic lesions, Mol.Hum. Reprod. 15 (2009) 645–652.
[260] M. Ozawa, T. Nagai, T. Somfai, M. Nakai, N. Maedomari, M. Fahrudin, N.W. Karja,H. Kaneko, J. Noguchi, K. Ohnuma, N. Yoshimi, H. Miyazaki, K. Kikuchi, Compar-ison between effects of 3-isobutyl-1-methylxanthine and FSH on gap junctionalcommunication, LH-receptor expression, and meiotic maturation of cumulus-oocyte complexes in pigs, Mol. Reprod. Dev. 75 (2008) 857–866.
[261] D. Pant, L.P. Reynolds, J.S. Luther, P.P. Borowicz, T.M. Stenbak, J.J. Bilski, R.M. Weigl,F. Lopes, K. Petry, M.L. Johnson, D.A. Redmer, A.T. Grazul-Bilska, Expression of
connexin 43 and gap junctional intercellular communication in the cumulus–oo-cyte complex in sheep, Reproduction 129 (2005) 191–200.
[262] M.L. Johnson, D.A. Redmer, L.P. Reynolds, J.J. Bilski, A.T. Grazul-Bilska, Gap junc-tional intercellular communication of bovine granulosa and thecal cells from an-tral follicles: effects of luteinizing hormone and follicle-stimulating hormone,Endocrine 18 (2002) 261–270.
[263] J. Gilleron, M. Nebout, L. Scarabelli, F. Senegas-Balas, S. Palmero, D. Segretain, G.Pointis, A potential novel mechanism involving connexin 43 gap junction forcontrol of Sertoli cell proliferation by thyroid hormones, J. Cell. Physiol. 209(2006) 153–161.
[264] N.A. Almeida, A. Cordeiro, D.S. Machado, L.L. Souza, T.M. Ortiga-Carvalho, A.C.Campos-de-Carvalho, F.E. Wondisford, C.C. Pazos-Moura, Connexin40 messen-ger ribonucleic acid is positively regulated by thyroid hormone (TH) acting incardiac atria via the TH receptor, Endocrinology 150 (2009) 546–554.
[265] B. de Montgolfier, A. Faye, C. Audet, D.G. Cyr, Seasonal variations in testicularconnexin levels and their regulation in the brook trout, Salvelinus fontinalis,Gen. Comp. Endocrinol. 162 (2009) 276–285.
[266] J. Wynn, U. Shah, S.A. Murray, Redistribution of connexin 43 by cAMP: a mech-anism for growth control in adrenal cells, Endocr. Res. 28 (2002) 663–668.
[267] A.M. Soto, C. Sonnenschein, Environmental causes of cancer: endocrine disrup-tors as carcinogens, Nat. Rev. Endocrinol. 6 (2010) 363–370.
[268] E. Diamanti-Kandarakis, J.P. Bourguignon, L.C. Giudice, R. Hauser, G.S. Prins, A.M.Soto, R.T. Zoeller, A.C. Gore, Endocrine-disrupting chemicals: an Endocrine Soci-ety scientific statement, Endocr. Rev. 30 (2009) 293–342.
[269] E. Leithe, A. Kjenseth, J. Bruun, S. Sirnes, E. Rivedal, Inhibition of connexin 43 gapjunction channels by the endocrine disruptor ioxynil, Toxicol. Appl. Pharmacol.247 (2010) 10–17.
[270] S. Salian, T. Dosh, G. Vanage, Neonatal exposure of male rats to Bisphenol A im-pairs fertility and expression of sertoli cell junctional proteins in the testis, Tox-icology 265 (2009) 56–67.
[271] I.K. Lee, S.K. Rhee, Inhibitory effect of bisphenol A on gap junctional intercellularcommunication in an epithelial cell line of rat mammary tissue, Arch. Pharm.Res. 30 (2007) 337–343.
[272] M. Tramoni, J. Gilleron, K. Tahiri, D. Carette, M.T. Corvol, D. Segretain, G. Pointis,J.F. Savouret, Contraceptive steroids from pharmaceutical waste perturbatejunctional communication in Sertoli cells, Biochimie 91 (2009) 1366–1375.
[273] I. Kopera, M. Durlej, A. Hejmej, K. Knapczyk-Stwora, M. Duda, M. Slomczynska, B.Bilinska B. Differential expression of connexin 43 in adult pig testes during normalspermatogenic cycle and after flutamide treatment, Reprod. Domest. Anim. 46(2011) 34–46.
[274] J.E. Trosko, B.L. Upham, A paradigm shift is required for the risk assessment of po-tential human health after exposure to low level chemical exposures: a response tothe toxicity testing in the 21st century report, Int. J. Toxicol. 29 (2010) 344–357.
[275] D.L. Eizirik, M.L. Colli, F. Ortis, The role of inflammation in insulitis and beta-cellloss in type 1 diabetes, Nat. Rev. Endocrinol. 5 (2009) 219–226.
[276] B. Kutlu, A.K. Cardozo, M.I. Darville, M. Kruhoffer, N. Magnusson, T. Orntoft, D.L.Eizirik, Discovery of gene networks regulating cytokine-induced dysfunctionand apoptosis in insulin-producing INS-1 cells, Diabetes 52 (2003) 2701–2719.
[277] J. Rasschaert, D. Liu, B. Kutlu, A.K. Cardozo, M. Kruhoffer, D.L. Eizirik, Global pro-filing of double stranded RNA- and IFN-gamma-induced genes in rat pancreaticbeta cells (2003), Diabetologia 46 (2003) 1641–1657.
[278] S. O'Rahilly, R.C. Turner, D.R. Matthews, Impaired pulsatile secretion of insulin inrelatives of patients with non-insulin-dependent diabetes, N. Engl. J. Med. 318(1988) 1225–1230.
[279] M. Prentki, C.J. Nolan, Islet beta cell failure in type 2 diabetes, J. Clin. Invest. 116(2006) 1802–1812.
[280] N. Belluardo, A. Trovato-Salinaro, G. Mudo, Y.L. Hurd, D.F. Condorelli, Structure,chromosomal localization, and brain expression of human Cx36 gene, J. Neu-rosci. Res. 57 (1999) 740–752.
[281] Y. Bosse, J.P. Despres, Y.C. Chagnon, T. Rice, D.C. Rao, C. Bouchard, L. Perusse, M.C.Voh, l Quantitative trait locus on 15q for a metabolic syndrome variable derivedfrom factor analysis, Obesity 15 (2007) 544–550.
[282] N.J. Cox, M. Frigge, D.L. Nicolae, P. Concannon, C.L. Hanis, G.I. Bell, A. Kong, Locion chromosomes 2 (NIDDM1) and 15 interact to increase susceptibility to dia-betes in Mexican Americans, Nat. Genet. 21 (1999) 213–215.
[283] D. Meyre, C. Lecoeur, J. Delplanque, S. Francke, V. Vatin, E. Durand, J. Weill, C.Dina, P. Froguel, A genome-wide scan for childhood obesity-associated traitsin French families shows significant linkage on chromosome 6q22.31-q23.2, Di-abetes 53 (2004) 803–811.
[284] Y. Mori, S. Otabe, C. Dina, K. Yasuda, C. Populaire, C. Lecoeur, V. Vatin, E. Durand,K. Hara, T. Okada, K. Tobe, P. Boutin, T. Kadowaki, P. Froguel, Genome-widesearch for type 2 diabetes in Japanese affected sib-pairs confirms susceptibilitygenes on 3q, 15q, and 20q and identifies two new candidate Loci on 7p and11p, Diabetes 51 (2002) 1247–1255.
[285] C. Mas, N. Taske, S. Deutsch, M. Guipponi, P. Thomas, A. Covanis, M. Friis, M.J.Kjeldsen, G.P. Pizzolato, J.G. Villemure, C. Buresi, M. Rees, A. Malafosse, M.Gardiner, S.E. Antonarakis, P. Meda, Association of the connexin36 genewith juvenile myoclonic epilepsy, J. Med. Genet. 41 (2004) e93.
[286] A. Hempelmann, A. Heils, T. Sander, Confirmatory evidence for an association ofthe connexin-36 gene with juvenile myoclonic epilepsy, Epilepsy Res. 71 (2006)223–228.
[287] P.Meda, P.A. Halban, A. Perrelet, A.E. Renold, L. Orci, Gap junction development is cor-related with insulin content in the pancreatic B cell, Science 209 (1980) 1026–1028.
[288] S. Bavamain, H. Pontes, J. Cancela, A. Charollais, S. Startchicl, D. Van de Ville, P.Meda, The intercellular synchronization of Ca2+ oscillations evaluates couplingof pancreatic beta cells, Biophys. J. (submitted for publication).

1936 I. Potolicchio et al. / Biochimica et Biophysica Acta 1818 (2012) 1919–1936
[289] M. Firouzi, B. Kok, W. Spiering, A. Busjahn, C.R. Bezzina, J.M. Ruijter, B.P. Koeleman,M. Schipper, W.A. Groenewegen, H.J. Jongsma, P.W. de Leeuw, Polymorphisms inhuman connexin40 gene promoter are associated with increased risk of hyperten-sion in men, J. Hypertens. 24 (2006) 325–330.
[290] M. Firouzi, H. Ramanna, B. Kok, H.J. Jongsma, B.P. Koeleman, P.A. Doevendans,W.A. Groenewegen, R.N. Hauer, Association of human connexin40 gene poly-morphisms with atrial vulnerability as a risk factor for idiopathic atrial fibrilla-tion, Circ. Res. 95 (2004) e29–e33.
[291] L. Dupays, D. Mazurais, C. Rucker-Martin, T. Calmels, D. Bernot, L. Cronier, A.Malassine, D. Gros, M. Theveniau-Ruissy, Genomic organization and alternativetranscripts of the human Connexin40 gene, Gene 305 (2003) 79–90.
[292] B.G. Bruneau, G. Nemer, J.P. Schmitt, F. Charron, L. Robitaille, S. Caron, D.A. Conner,M. Gessler, M. Nemer, C.E. Seidman, J.C. Seidman, A murine model of Holt-Oramsyndrome defines roles of the T-box transcription factor Tbx5 in cardiogenesisand disease, Cell 106 (2001) 709–721.
[293] F.A. Stennard, M.W. Costa, D.A. Elliott, S. Rankin, S.J. Haast, D. Lai, L.P. McDonald,K. Niederreither, P. Dolle, B.G. Bruneau, A.M. Zorn, R.P. Harvey, Cardiac T-boxfactor Tbx20 directly interacts with Nkx2-5, GATA4, and GATA5 in regulationof gene expression in the developing heart, Dev. Biol. 262 (2003) 206–224.
[294] V.M. Christoffels, W.M. Hoogaars, A. Tessari, D.E. Clout, A.F. Moorman,M. Campione,T-box transcription factor Tbx2 represses differentiation and formation of the cardiacchambers, Dev. Dyn. 229 (2004) 763–770.
[295] W.M. Hoogaars, A. Tessari, A.F. Moorman, P.A. de Boer, J. Hagoort, A.T. Soufan, M.Campione, V.M. Christoffels, The transcriptional repressor Tbx3 delineates thedeveloping central conduction system of the heart, Cardiovasc. Res. 62 (2004)489–499.
[296] J. Zhang, C.E. Hill, Differential connexin expression in preglomerular and post-glomerular vasculature: accentuation during diabetes, Kidney Int. 68 (2005)1171–1185.
[297] N.M. Rummery, C.E. Hill, Vascular gap junctions and implications for hyperten-sion, Clin. Exp. Pharmacol. Physiol. 31 (2004) 659–667.
[298] C. deWit, F. Roos, S.S. Bolz, U. Pohl, Lack of vascular connexin 40 is associatedwith hy-pertension and irregular arteriolar vasomotion, Physiol. Gen. 13 (2003) 169–177.
[299] X.F. Figueroa, D.L. Paul, A.M. Simon, D.A. Goodenough, K.H. Day, D.N. Damon, B.R.Duling, Central role of connexin40 in the propagation of electrically activated vaso-dilation in mouse cremasteric arterioles in vivo, Circ. Res. 92 (2003) 793–800.
[300] S.L. Sandow, M. Tare, H.A. Coleman, C.E. Hill, H.C. Parkington, Involvement ofmyoendothelial gap junctions in the actions of endothelium-derived hyperpo-larizing factor, Circ. Res. 90 (2002) 1108–1113.
[301] S.S. Segal, B.R. Duling, Conduction of vasomotor responses in arterioles: a rolefor cell-to-cell coupling? Am. J. Physiol. 256 (1989) H838–H845.
[302] S.S. Segal, D.G. Welsh, D.T. Kurjiaka, Spread of vasodilatation and vasoconstric-tion along feed arteries and arterioles of hamster skeletal muscle, J. Physiol.516 (1999) 283–291.
[303] S.M. Grundy, Does the metabolic syndrome exist? Diabetes Care 29 (2006)1689–1692.
[304] C.M. Alexander, P.B. Landsman, S.M. Grundy, Metabolic syndrome and hypergly-cemia: congruence and divergence, Am. J. Cardiol. 98 (2006) 982–985.
[305] T. Kuroki, T. Inoguchi, F. Umeda, F. Ueda, H. Nawata, High glucose induces alter-ation of gap junction permeability and phosphorylation of connexin-43 in cul-tured aortic smooth muscle cells, Diabetes 47 (1998) 931–936.
[306] T. Sato, R. Haimovici, R. Kao, A.F. Li, S. Roy, Downregulation of connexin 43 ex-pression by high glucose reduces gap junction activity in microvascular endo-thelial cells, Diabetes 51 (2002) 1565–1571.
[307] C.W. Wong, T. Christen, I. Roth, C.E. Chadjichristos, J.P. Derouette, B.F. Foglia, M.Chanson, D.A. Goodenough, B.R. Kwak, Connexin37 protects against atheroscle-rosis by regulating monocyte adhesion, Nat. Med. 12 (2006) 950–954.
[308] C.E. Chadjichristos, J.P. Derouette, B.R. Kwak, Connexins in atherosclerosis, Adv.Cardiol. 42 (2006) 255–267.
[309] A.S. Wilkins, Gap junction-mediated intercellular signalling in health and dis-ease, Bioessays 20 (1998) 686–688.
[310] A. Hirashiki, Y. Yamada, Y. Murase, Y. Suzuki, H. Kataoka, Y. Morimoto, T. Tajika,T. Murohara, M. Yokota, Association of gene polymorphisms with coronary ar-tery disease in low- or high-risk subjects defined by conventional risk factors,J. Am. Coll. Cardiol. 42 (2003) 1429–1437.
[311] Y. Yamada, S. Ichihara, H. Izawa, M. Tanaka, M. Yokota, Genetic risk for coronaryartery disease in individuals with or without type 2 diabetes, Mol. Genet. Metab.81 (2004) 282–290.
[312] H. Lin, K. Ogawa, I. Imanaga, N. Tribulova, Remodeling of connexin 43 in the di-abetic rat heart, Mol. Cell. Biochem. 290 (2006) 69–78.
[313] K. Sawai, M. Mukoyama, K. Mori, H. Yokoi, M. Koshikawa, T. Yoshioka, R. Takeda,A. Sugawara, T. Kuwahara, M.A. Saleem, O. Ogawa, K. Nakao, Redistribution ofconnexin43 expression in glomerular podocytes predicts poor renal prognosisin patients with type 2 diabetes and overt nephropathy, Nephrol. Dial. Trans-plant. 21 (2006) 2472–2477.
[314] H. Lin, K. Ogawa, I. Imanaga, N. Tribulova, Alterations of connexin 43 in the dia-betic rat heart, Adv. Cardiol. 42 (2006) 243–254.
[315] J.J. Sheu, L.T. Chang, C.H. Chiang, C.K. Sun, N.K. Chang, A.A. Youssef, C.J. Wu, F.Y.Lee, H.K. Yip, Impact of diabetes on cardiomyocyte apoptosis and connexin43gap junction integrity: role of pharmacological modulation, Int. Heart J. 48(2007) 233–245.
[316] T. Inoguchi, H.Y. Yu, M. Imamura, M. Kakimoto, T. Kuroki, T. Maruyama, H.Nawata, Altered gap junction activity in cardiovascular tissues of diabetes,Med. Electron Microsc. 34 (2001) 86–91.
[317] D. Lin, R. Harris, R. Stutzman, G.A. Zampighi, H. Davidson, D.J. Takemoto, Proteinkinase C-gamma activation in the early streptozotocin diabetic rat lens, Curr.Eye Res. 32 (2007) 523–532.
[318] C.M. Wang, J. Lincoln, J.E. Cook, D.L. Becker, Abnormal connexin expression un-derlies delayed wound healing in diabetic skin, Diabetes 56 (2007) 2809–2817.
[319] P.R. Brink, V. Valiunas, H.Z. Wang, W. Zhao, K. Davies, G.J. Christ, Experimentaldiabetes alters connexin43 derived gap junction permeability in short-term cul-tures of rat corporeal vascular smooth muscle cells, J. Urol. 175 (2006) 381–386.
[320] T. Takenaka, T. Inoue, H. Okada, Y. Ohno, T. Miyazaki, D.J. Chaston, C.E. Hill, H.Suzuki H. Altered gap junctional communication and renal haemodynamics inZucker fatty rat model of type 2 diabetes, Diabetologia 54 (2011) 2192–2201.
[321] D.R. Tomlinson, N.J. Gardiner, Glucose neurotoxicity, Nat. Rev. Neurosci. 9(2008) 36–45.
[322] G. Söhl, B. Odermatt, S. Maxeiner, J. Degen, K. Willecke, New insights into the ex-pression and function of neural connexins with transgenic mouse mutants,Brain Res. Brain Res. Rev. 47 (2004) 245–259.
[323] D.P. Poladia, B. Schanbacher, L.J. Wallace, J.A. Bauer, Innervation and connexinisoform expression during diabetes-related bladder dysfunction: early structu-ral vs. neuronal remodelling, Acta Diabetol. 42 (2005) 147–152.
[324] D.A. Pitre, J.L. Seifert, J.A. Bauer, J.A. Perineurium, inflammation and altered con-nexin isoform expression in a rat model of diabetes related peripheral neuropa-thy, Neurosci. Lett. 303 (2001) 67–71.
[325] Y. Naoi, Y. Miyoshi, T. Taguchi, S.J. Kim, T. Arai, N. Maruyama, Y. Tamaki, S. Nogu-chi, Connexin26 expression is associated with aggressive phenotype in humanpapillary and follicular thyroid cancers, Cancer Lett. 262 (2008) 248–256.
[326] H.S. Willenberg, M. Schott, W. Saeger, A. Tries, W.A. Scherbaum, S.R. Bornstein,Expression of connexins in chromaffin cells of normal human adrenals and inbenign and malignant pheochromocytomas, Ann. N. Y. Acad. Sci. 1073 (2006)578–583.
[327] T.J. King, P.D. Lampe, Mice deficient for the gap junction protein Connexin32 ex-hibit increased radiation-induced tumorigenesis associated with elevated mito-gen-activated protein kinase (p44/Erk1, p42/Erk2) activation, Carcinogenesis 25(2004) 669–680.
[328] S.A. Murray, K. Davis, L.M. Fishman, S.R. Bornstein, Alpha1 connexin 43 gap junc-tions are decreased in human adrenocortical tumors, J. Clin. Endocrinol. Metab.85 (2000) 890–895.
[329] E.A. Hanna, S. Umhauer, S.L. Roshong, M.P. Piechocki, M.J. Fernstrom, J.D. Fanning,R.J. Ruch, Gap junctional intercellular communication and connexin43 expressionin human ovarian surface epithelial cells and ovarian carcinomas in vivo and invitro, Carcinogenesis 20 (1999) 369–373.
[330] R. Brehm, C. Rüttinger, P. Fischer, I. Gashaw, E. Winterhager, S. Kliesch, R.M.Bohle, K. Steger, M. Bergmann, Transition from preinvasive carcinoma in situto seminoma is accompanied by a reduction of connexin 43 expression in Sertolicells and germ cells, Neoplasia 8 (2006) 499–509.
[331] R. Brehm, A. Marks, R. Rey, S. Kliesch, M. Bergmann, K. Steger, Altered expressionof connexins 26 and 43 in Sertoli cells in seminiferous tubules infiltrated withcarcinoma-in-situ or seminoma, J. Pathol. 197 (2002) 647–653.
[332] C. Roger, B. Mograb, D. Chevallier, J.F. Michiels, H. Tanaka, D. Segretain, G. Pointis, P.Fenichel, Disrupted traffic of connexin 43 in human testicular seminoma cells:overexpression of Cx43 inducesmembrane location and cell proliferation decrease,J. Pathol. 202 (2004) 241–246.
[333] D. Segretain, X. Decrouy, J. Dompierre, D. Escalier, N. Rahman, C. Fiorini, B.Mograbi, J.P. Siffroi, I. Huhtaniemi, P. Fenichel, G. Pointis, Sequestration of con-nexin43 in the early endosomes: an early event of Leydig cell tumor progres-sion, Mol. Carcinog. 38 (2003) 179–187.
[334] H.S. Willenberg, M. Schott, W. Saeger, A. Tries, W.A. Scherbaum, S.R. Bornstein, Ex-pression of connexins in chromaffin cells of normal human adrenals and in benignand malignant pheochromocytomas, Ann. N. Y. Acad. Sci. 1073 (2006) 578–583.
[335] X. Li, C. Olson, S. Lu, J.I. Nagy, Association of connexin36 with zonula occludens-1in HeLa cells, betaTC-3 cells, pancreas, and adrenal gland, Histochem. Cell Biol.122 (2004) 485–498.
[336] J. Degen, C. Meier, R.S. Van Der Giessen, G. Söhl, E. Petrasch-Parwez, S. Urschel, R.Dermietzel, K. Schilling, C.I. De Zeeuw, K. Willecke, Expression pattern of lacZ re-porter gene representing connexin36 in transgenic mice, J. Comp. Neurol. 473(2004) 511–525.
[337] M.S. Risley, I.P. Tan, C. Roy, J.C. Saez, Cell-, age- and stage-dependent distributionof connexin43 gap junctions in testes, J. Cell Sci. 103 (1992) 81–96.
[338] I.P. Tan, C. Roy, J.C. Saez, C.G. Saez, D.L. Paul, M.S. Risley, Regulated assembly ofconnexin33 and connexin43 into rat Sertoli cell gap junctions, Biol. Reprod. 54(1996) 1300–1310.
[339] P.A. Nielsen, N.M. Kumar, Differences in expression patterns between mouseconnexin-30.2 (Cx30.2) and its putative human orthologue, connexin-31.9,FEBS Lett. 540 (2003) 151–156.
[340] E.A. Hanna, S. Umhauer, S.L. Roshong, M.P. Piechocki, M.J. Fernstrom, J.D. Fanning,R.J. Ruch, Gap junctional intercellular communication and connexin43 expressionin human ovarian surface epithelial cells and ovarian carcinomas in vivo and invitro, Carcinogenesis 20 (1999) 1369–1373.
[341] K. Itahana, Y. Morikazu, T. Takeya, Differential expression of four connexingenes, Cx-26, Cx-30.3, Cx-32, and Cx-43, in the porcine ovarian follicle, Endocri-nology 137 (1996) 5036–5044.
[342] M.L. Johnson, D.A. Redmer, L.P. Reynolds, J.J. Bilski, A.T. Grazul-Bilska, Gap junc-tional intercellular communication of bovine granulosa and thecal cells from an-tral follicles: effects of luteinizing hormone and follicle-stimulating hormone,Endocrine 18 (2002) 261–270.