
Phosphatidylcholine-Specific Phospholipase C Activation in Epithelial Ovarian Cancer Cells
10
Phosphatidylcholine-Specific Phospholipase C Activation in Epithelial Ovarian Cancer Cells Francesca Spadaro, 1 Carlo Ramoni, 1 Delia Mezzanzanica, 2 Silvia Miotti, 2 Paola Alberti, 2 Serena Cecchetti, 1 Egidio Iorio, 1 Vincenza Dolo, 3 Silvana Canevari, 2 and Franca Podo 1 1 Section of Molecular and Cellular Imaging, Department of Cell Biology and Neurosciences, Istituto Superiore di Sanita `, Rome, Italy; 2 Unit of Molecular Therapies, Department of Experimental Oncology and Laboratories, Fondazione IRCCS Istituto Nazionale dei Tumori, Milan, Italy; and 3 Department of Experimental Medicine, University of L’Aquila, L’Aquila, Italy Abstract Elucidation of the mechanisms responsible for aberrant phosphatidylcholine (PC) metabolism in cancer cells may allow identification of novel biomarkers of tumor progression and design of new targeted anticancer therapies. We recently reported up-regulation of PC-specific phospholipases in epi- thelial ovarian cancer cells (EOC) compared with nontumoral (normal or immortalized) counterparts (EONT). In the pre- sent study, we focused, in the same cell systems, on levels, subcellular localization, and activity of PC-specific phospho- lipase C (PC-PLC), for which a key role in cell prolifera- tion, differentiation, and apoptosis has been shown in several mammalian cells. A 66-kDa PC-PLC isoform, detected in nuclear and cytoplasmic compartments of both EOC and EONT cells, accumulated on the external plasma membrane of cancer cells only, where it colocalized with B1 integrin, in nonraft membrane domains. PC-PLC activity was 3-fold higher in total cell lysates and 5-fold higher in membrane- enriched fractions of EOC compared with EONT cells. Serum deprivation induced in EOC, but not in EONT, cells a 3-fold decrease in PC-PLC activity, associated with a 40% drop in S-phase fraction. The recovery of both variables to their original levels in serum-restimulated (or lysophosphatidic acid–restimulated) EOC cells was strongly delayed, for at least 24 h, in the presence of the PC-PLC inhibitor tricyclo- decan-9-yl-potassium xanthate (D609). The S-phase of serum- restimulated EONT cells was not sensitive to D609. These findings warrant further investigations on the role of PC-PLC and on the effects of its inhibition on the pathways respon- sible for constitutive EOC cell stimulation and cell proliferation. [Cancer Res 2008;68(16):6541–9] Introduction Epithelial ovarian cancer (EOC), the most common cause of death from gynecologic malignancy in the Western world, still presents a high case-fatality ratio with an overall 5-year survival lower than 45%, mainly due to asymptomatic disease progression at early stage, high recurrence rate, and frequent onset of chemo- resistance after first line treatment (1, 2). The existing gap in the knowledge of molecular mechanisms responsible for EOC progres- sion leads to current limitations of therapy regimens, still mainly restricted to surgery followed by chemotherapy with cytotoxic drugs. Only limited attention has been, thus far, devoted to altera- tions of metabolic fluxes and enzyme activities associated in these tumors with gene-driven deregulation of cell signaling pathways responsible for uncontrolled cell proliferation, cell invasiveness, and escape from default apoptotic programs. An aberrant phosphatidylcholine (PC) metabolism is reported in cancer cells, in which substantial changes in the steady-state levels of PC cycle derivatives are commonly detected by in vitro and in vivo nuclear magnetic resonance signals (3–5). Among these, an increase in phosphocholine (PCho) content is a common feature of different cancer cells, beyond their otherwise wide phenotypic variability (5). Elucidation of the underlying molecular mechanisms may characterize novel biomarkers of tumor progression (6–8) and allow identification of metabolic targets for newly designed selective antitumor therapies (9–11). We recently reported (8) a 3-fold to 8-fold increase in the PCho content in EOC cell lines compared with epithelial ovarian nontumoral (normal or immortalized) cells (EONT). Enzymatic assays showed that PCho accumulation in EOC cells was associated with activation of choline kinase, a biosynthetic enzyme well known for its implication in human carcinogenesis (12–14), and PC-specific phospholipases of the C and D type (PC-PLC and PC-PLD). These findings prompted our interest on investigating the role of PC-PLC in ovary tumor progression, also in view of our previous reports on ectopic localization and/or activation of this enzyme in oncogene-transformed and mitogen-stimulated fibro- blasts (15, 16). A growing body of evidence has pointed, since the 1990s, to the implication of PC-PLC in proliferation, differentiation, and apop- tosis of mammalian cells. In fact, this enzyme has been implicated in cellular processes crucial for cell signaling, such as ( a ) mitogen- driven and oncogene-driven extracellular signal-regulated kinase phosphorylation and activation of gene transcription factors in fibroblasts (16, 17), (b) agonist stimulation of cells transfected with receptor proteins (18), (c) neuronal and endothelial cell differen- tiation (19–21), (d ) programmed cell death (22, 23), and ( e ) acti- vation of cells of the immune system (24–30). Currently, the mammalian PC-PLC has not been cloned and its sequence is unknown. Only scanty knowledge has been accumu- lated on expression, subcellular distribution, and activity of PC-PLC in cancer cells. Nevertheless, indirect evidence on increased PC- PLC activity was reported in breast tumor cells (31). Moreover, the present unavailability of specific monoclonal antibodies (mAb) can be partly compensated by the use of rabbit polyclonal antibodies raised against bacterial (Bacillus cereus ) PC-PLC, possessing proved Note: Supplementary data for this article are available at Cancer Research Online (http://cancerres.aacrjournals.org/). F. Spadaro and C. Ramoni contributed equally to this work. Requests for reprints: Franca Podo, Department of Cell Biology and Neurosciences, Istituto Superiore di Sanita `, Viale Regina Elena 299, 00161 Rome, Italy. Phone: 39-06-49902686; Fax: 39-06-49387144; E-mail: [email protected] or Silvana Canevari, Department of Experimental Oncology, Fondazione IRCCS Istituto Nazionale dei Tumori, Via Venezian 1, 20133 Milan, Italy. Phone: 39-02-23902567; Fax: 39-02-23903073; E-mail: [email protected]. I2008 American Association for Cancer Research. doi:10.1158/0008-5472.CAN-07-6763 www.aacrjournals.org 6541 Cancer Res 2008; 68: (16). August 15, 2008 Research Article Research. on June 8, 2015. © 2008 American Association for Cancer cancerres.aacrjournals.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Phosphatidylcholine-Specific Phospholipase C Activation in Epithelial Ovarian Cancer Cells

Phosphatidylcholine-Specific Phospholipase C Activation in
Epithelial Ovarian Cancer Cells
Francesca Spadaro,1Carlo Ramoni,
1Delia Mezzanzanica,
2Silvia Miotti,
2Paola Alberti,
2
Serena Cecchetti,1Egidio Iorio,
1Vincenza Dolo,
3Silvana Canevari,
2and Franca Podo
1
1Section of Molecular and Cellular Imaging, Department of Cell Biology and Neurosciences, Istituto Superiore di Sanita, Rome, Italy;2Unit of Molecular Therapies, Department of Experimental Oncology and Laboratories, Fondazione IRCCS Istituto Nazionaledei Tumori, Milan, Italy; and 3Department of Experimental Medicine, University of L’Aquila, L’Aquila, Italy
Abstract
Elucidation of the mechanisms responsible for aberrantphosphatidylcholine (PC) metabolism in cancer cells mayallow identification of novel biomarkers of tumor progressionand design of new targeted anticancer therapies. We recentlyreported up-regulation of PC-specific phospholipases in epi-thelial ovarian cancer cells (EOC) compared with nontumoral(normal or immortalized) counterparts (EONT). In the pre-sent study, we focused, in the same cell systems, on levels,subcellular localization, and activity of PC-specific phospho-lipase C (PC-PLC), for which a key role in cell prolifera-tion, differentiation, and apoptosis has been shown in severalmammalian cells. A 66-kDa PC-PLC isoform, detected innuclear and cytoplasmic compartments of both EOC andEONT cells, accumulated on the external plasma membraneof cancer cells only, where it colocalized with B1 integrin,in nonraft membrane domains. PC-PLC activity was 3-foldhigher in total cell lysates and 5-fold higher in membrane-enriched fractions of EOC compared with EONT cells. Serumdeprivation induced in EOC, but not in EONT, cells a 3-folddecrease in PC-PLC activity, associated with a 40% drop inS-phase fraction. The recovery of both variables to theiroriginal levels in serum-restimulated (or lysophosphatidicacid–restimulated) EOC cells was strongly delayed, for atleast 24 h, in the presence of the PC-PLC inhibitor tricyclo-decan-9-yl-potassium xanthate (D609). The S-phase of serum-restimulated EONT cells was not sensitive to D609. Thesefindings warrant further investigations on the role of PC-PLCand on the effects of its inhibition on the pathways respon-sible for constitutive EOC cell stimulation and cell proliferation.[Cancer Res 2008;68(16):6541–9]
Introduction
Epithelial ovarian cancer (EOC), the most common cause ofdeath from gynecologic malignancy in the Western world, stillpresents a high case-fatality ratio with an overall 5-year survivallower than 45%, mainly due to asymptomatic disease progressionat early stage, high recurrence rate, and frequent onset of chemo-
resistance after first line treatment (1, 2). The existing gap in theknowledge of molecular mechanisms responsible for EOC progres-sion leads to current limitations of therapy regimens, still mainlyrestricted to surgery followed by chemotherapy with cytotoxicdrugs. Only limited attention has been, thus far, devoted to altera-tions of metabolic fluxes and enzyme activities associated in thesetumors with gene-driven deregulation of cell signaling pathwaysresponsible for uncontrolled cell proliferation, cell invasiveness,and escape from default apoptotic programs.An aberrant phosphatidylcholine (PC) metabolism is reported in
cancer cells, in which substantial changes in the steady-state levelsof PC cycle derivatives are commonly detected by in vitro andin vivo nuclear magnetic resonance signals (3–5). Among these, anincrease in phosphocholine (PCho) content is a common feature ofdifferent cancer cells, beyond their otherwise wide phenotypicvariability (5). Elucidation of the underlying molecular mechanismsmay characterize novel biomarkers of tumor progression (6–8) andallow identification of metabolic targets for newly designedselective antitumor therapies (9–11).We recently reported (8) a 3-fold to 8-fold increase in the PCho
content in EOC cell lines compared with epithelial ovariannontumoral (normal or immortalized) cells (EONT). Enzymaticassays showed that PCho accumulation in EOC cells wasassociated with activation of choline kinase, a biosynthetic enzymewell known for its implication in human carcinogenesis (12–14),and PC-specific phospholipases of the C and D type (PC-PLC andPC-PLD). These findings prompted our interest on investigatingthe role of PC-PLC in ovary tumor progression, also in view of ourprevious reports on ectopic localization and/or activation of thisenzyme in oncogene-transformed and mitogen-stimulated fibro-blasts (15, 16).A growing body of evidence has pointed, since the 1990s, to the
implication of PC-PLC in proliferation, differentiation, and apop-tosis of mammalian cells. In fact, this enzyme has been implicatedin cellular processes crucial for cell signaling, such as (a) mitogen-driven and oncogene-driven extracellular signal-regulated kinasephosphorylation and activation of gene transcription factors infibroblasts (16, 17), (b) agonist stimulation of cells transfected withreceptor proteins (18), (c) neuronal and endothelial cell differen-tiation (19–21), (d) programmed cell death (22, 23), and (e) acti-vation of cells of the immune system (24–30).Currently, the mammalian PC-PLC has not been cloned and its
sequence is unknown. Only scanty knowledge has been accumu-lated on expression, subcellular distribution, and activity of PC-PLCin cancer cells. Nevertheless, indirect evidence on increased PC-PLC activity was reported in breast tumor cells (31). Moreover, thepresent unavailability of specific monoclonal antibodies (mAb) canbe partly compensated by the use of rabbit polyclonal antibodiesraised against bacterial (Bacillus cereus) PC-PLC, possessing proved
Note: Supplementary data for this article are available at Cancer Research Online(http://cancerres.aacrjournals.org/).
F. Spadaro and C. Ramoni contributed equally to this work.Requests for reprints: Franca Podo, Department of Cell Biology and
Neurosciences, Istituto Superiore di Sanita, Viale Regina Elena 299, 00161 Rome,Italy. Phone: 39-06-49902686; Fax: 39-06-49387144; E-mail: [email protected] or SilvanaCanevari, Department of Experimental Oncology, Fondazione IRCCS IstitutoNazionale dei Tumori, Via Venezian 1, 20133 Milan, Italy. Phone: 39-02-23902567;Fax: 39-02-23903073; E-mail: [email protected].
I2008 American Association for Cancer Research.doi:10.1158/0008-5472.CAN-07-6763
www.aacrjournals.org 6541 Cancer Res 2008; 68: (16). August 15, 2008
Research Article
Research. on June 8, 2015. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

selective cross-reactivity against mammalian PC-PLC (32). Thisapproach allowed identification of PC-PLC isoforms in variousnormal and tumor cells of different origin (15, 16, 25–27, 32, 33).This article reports evidence on subcellular localization and acti-
vity of PC-PLC in EOC cell lines compared with epithelial ovariannormal cells or immortalized cell variants. The possible role of thisenzyme in cell proliferation was investigated by measuring PC-PLCactivity in relation to alterations in the S-phase fraction induced inEOC cells by serum deprivation and subsequent restimulation bygrowth factors, in the presence or absence of a PC-PLC inhibitor.
Materials and Methods
Antibodies and reagents. Rabbit polyclonal anti–PC-PLC antibodyraised against bacterial (B. cereus) PC-PLC and selectively cross-reacting
with mammalian PC-PLC was obtained according to the procedure de-
scribed by Clark and colleagues (32) and modified as reported in ref. 15.Immunochemical characterization and specificity of these antibodies for
proteins with PC-PLC activity have already been described (15, 16, 25). In
particular, specific binding inhibition tests showed that this antibody was
directed against the catalytic site of a mammalian 66-kDa PC-PLC isoform(16). Anti–h-actin mAb was supplied by Sigma-Aldrich. Anti–folate receptor(FR) MOV18 and MOV19 (34), available through Alexis Life Sciences, Inc.,
and anti–h1 integrin MAR4 (35) mAbs were produced at the Fondazione
IRCCS Istituto Nazionale dei Tumori, Milan. Rabbit anti–caveolin-1 (Cav-1)antibody was from Santa Cruz Biotechnology. Alexa Fluor-488 and Alexa
Fluor-594 F(ab)2 fragment of goat anti-rabbit IgG (H + L) and Alexa Fluor-
594 F(ab)2 fragment of goat anti-mouse IgG (H + L), purchased from
Molecular Probes, Inc., were used as secondary antibodies.Epidermal growth factor (EGF) was supplied by Calbiochem. Lysophos-
phatidic acid (LPA) was purchased from Avanti Polar Lipids, Inc. Triton
X-100, propidium iodide, tricyclodecan-9-yl-potassium xanthate (D609),5-bromo-2¶-deoxyuridine (BrdUrd), and all other chemicals and bioche-
micals were from Sigma-Aldrich.
Normal ovarian epithelial cells, immortalized cell variants, andcancer cells. Normal ovarian surface epithelial (OSE) cells (six independentpreparations), stably immortalized OSE cell variants obtained by OSE
transfection with SV40 large T antigen (IOSE) and by further IOSE
transfection with cDNA of the catalytic subunit of human telomerase
reverse transcriptase (hTERT), and EOC cell lines (IGROV1, OVCAR3,SKOV3, and CABA I) were obtained and cultured as reported (8). Ascitic
fluids were collected during surgical procedures from four advanced stage
ovarian cancer patients undergoing debulking surgery at the FondazioneIRCCS Istituto Nazionale dei Tumori at the first diagnosis and not
previously treated with chemotherapeutic regimens. Histopathologic
diagnosis for all patients was poorly differentiated serous papillary ovarian
cancer. All clinical specimens used for this study were obtained withInstitutional Review Board approval from patients who gave informed
consent to use leftover biological material for investigative purposes.
Tumor cells were isolated by centrifugation on a discontinuous 75% to 100%
Ficoll gradient followed by separation from adherent cells, essentially asdescribed (36).
Confocal laser scanning microscopy and flow cytometry analyses.PC-PLC subcellular distribution in either unfixed or fixed and permeabilizedcells was characterized by indirect immunofluorescence and confocal laser
scanning microscopy (CLSM) analysis, as previously described (16, 25–27).
CLSM observations were performed on a Leica TCS SP2 AOBS apparatus,
using excitation spectral laser lines at 405, 488, and 594 nm and using theLeica Confocal Software (Leica Lasertechnik GmbH) and Adobe Photoshop
software programs (Adobe Systems).
Signals from different fluorescent probes were taken in parallel, and
colocalization was detected in yellow. Different fields of view (>200 cells)were analyzed on the microscope for each labeling condition, and
representative results are shown.
Flow cytometry analyses were performed on cells suspended in 0.2%
PBS-EDTA and stained as described (16).
Western blot analyses. Total cell lysates were obtained suspending cellpellets in ice-cold lysis buffer [150 mmol/L NaCl, 100 mmol/L Tris-HCl
(pH 8), 1% Triton X-100, 1 mmol/L MgCl2, and protease inhibitor cocktail]
for 20 min. Lysates were cleared of detergent insoluble materials by 10 min
centrifugation at 20,000 � g .Membrane fractions were prepared, and their purification was controlled
as described (37).
Protein concentration was determined by Bio-Rad protein assay
according to the manufacturer’s protocol (Bio-Rad Laboratories, Inc.) andtotal cell lysates or purified membranes (30 Ag proteins) were analyzed byWestern blotting, as previously described (27). Densitometry analyses of
protein bands were performed with a Bio-Rad apparatus (Bio-Rad
Laboratories Srl) using the Quantity One software.In vitro PC-PLC and PLD activity assays. Relative changes of PC-PLC
and PLD activities were determined in whole-cell lysates and enriched mem-
brane fractions using the Amplex Red assay kit (Molecular Probe, Inc.), asdescribed by the manufacturer and adapted by Spadaro and colleagues (26).
Separation of lipid rafts by sucrose gradient. Cells grown in T75 flaskswere washed in cold PBS, lysed in 1% Triton X-100 containing buffer
[25 mmol/L MES (pH 6.5), and 150 mmol/L NaCl], and the lysate was mixed1:1 with 80% sucrose solution. Gradient fractionation (5–30% sucrose) and
protein separation were carried out as previously described (38, 39). After
ultracentrifugation at 200,000 � g for 17 h in TH641 Sorvall rotor, twelve
1-mL fractions were collected from the top of the gradient, and the samevolume (90 AL) was taken from each fraction for analysis in SDS-PAGE in
parallel with that of total cell lysate (35 Ag total protein). To concentrateraft membranes, top fractions 3 to 6 were diluted 1:2 with 25 mmol/L MES(pH 6.5) and 150 mmol/L NaCl, and raft membranes were pelleted by
centrifugation at 13,000 rpm for 30 min. Pellet was resuspended in loading
buffer for SDS-PAGE analysis. Distribution of relevant proteins separated
by 10% SDS-PAGE was analyzed by Western blotting.Glycosyl-phosphatidyl inositol–anchored FR and Cav-1 were used as
markers of lipid rafts, whereas h1 integrin was considered as a nonraft
marker. To analyze FR and h1 integrin reactivity, samples were analyzed
under nonreducing conditions.Immunohistochemical detection of PC-PLC in EOC tissues. Analyses
were performed on a commercial tissue array slide constructed with
paraffin sections from 50 ovarian cancer specimens of different histo-types (SuperBioChips Laboratories). Antigen retrieval was performed in
10 mmol/L citrate buffer (pH 6), at 120jC for 2 min; endogenous peroxidasewas quenched with 3% H2O2 for 10 min, and slides were saturated in
blocking solution (30 min) and then incubated with rabbit anti–PC-PLCantibodies (1 h at room temperature) and revealed with biotin-conjugated
antirabbit secondary antibody followed by avidin-biotin complex (Vectas-
tain Universal Elite ABC kit, Vector Laboratories, Inc.). Peroxidase reaction
was developed with 3,3-diaminobenzidine, and sections were counter-stained with hematoxylin. Slides incubated with the secondary antibodies
alone provided negative controls; a control with hematoxylin/eosin was also
prepared. Staining was evaluated considering both staining intensity and
the proportion of cells showing positive reaction.Statistical analysis. Data were analyzed using GraphPad software
version 3.03. Statistical significance of differences was determined by one-
way ANOVA or by Student’s t test (as specified). Differences were consideredsignificant at P < 0.05.
Results
Increased levels of PC-PLC localization and activity on themembrane of EOC compared with EONT cells. CLSM analyseson unfixed cells showed massive (although different) levels of PC-PLC localization on the outer membrane surface of a number ofEOC cell lines, as well as on that of cancer cells isolated fromascitic exudates (examples in Fig. 1A). This characteristic PC-PLCdistribution was much less evident in hTERT and only barelydetectable in OSE and IOSE cells. Flow cytometry analyses onunfixed cells showed a 5.0 F 0.9 (SD)–fold increase (ANOVA
Cancer Research
Cancer Res 2008; 68: (16). August 15, 2008 6542 www.aacrjournals.org
Research. on June 8, 2015. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from
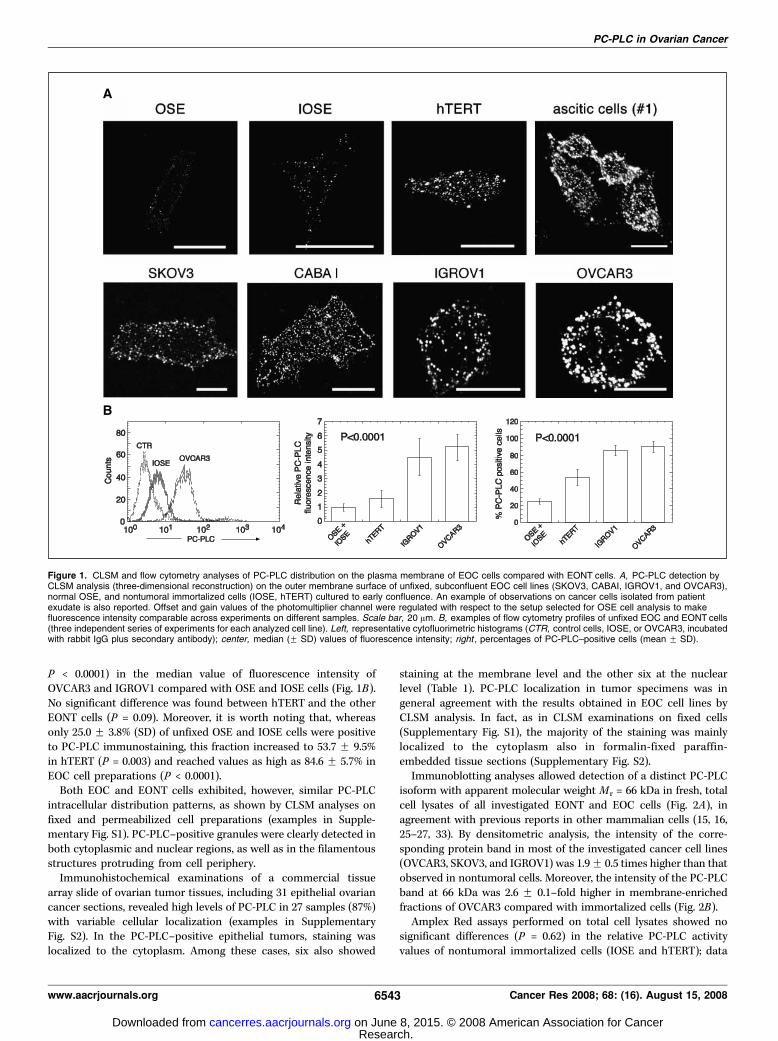
P < 0.0001) in the median value of fluorescence intensity ofOVCAR3 and IGROV1 compared with OSE and IOSE cells (Fig. 1B).No significant difference was found between hTERT and the otherEONT cells (P = 0.09). Moreover, it is worth noting that, whereasonly 25.0 F 3.8% (SD) of unfixed OSE and IOSE cells were positiveto PC-PLC immunostaining, this fraction increased to 53.7 F 9.5%in hTERT (P = 0.003) and reached values as high as 84.6 F 5.7% inEOC cell preparations (P < 0.0001).Both EOC and EONT cells exhibited, however, similar PC-PLC
intracellular distribution patterns, as shown by CLSM analyses onfixed and permeabilized cell preparations (examples in Supple-mentary Fig. S1). PC-PLC–positive granules were clearly detected inboth cytoplasmic and nuclear regions, as well as in the filamentousstructures protruding from cell periphery.Immunohistochemical examinations of a commercial tissue
array slide of ovarian tumor tissues, including 31 epithelial ovariancancer sections, revealed high levels of PC-PLC in 27 samples (87%)with variable cellular localization (examples in SupplementaryFig. S2). In the PC-PLC–positive epithelial tumors, staining waslocalized to the cytoplasm. Among these cases, six also showed
staining at the membrane level and the other six at the nuclearlevel (Table 1). PC-PLC localization in tumor specimens was ingeneral agreement with the results obtained in EOC cell lines byCLSM analysis. In fact, as in CLSM examinations on fixed cells(Supplementary Fig. S1), the majority of the staining was mainlylocalized to the cytoplasm also in formalin-fixed paraffin-embedded tissue sections (Supplementary Fig. S2).Immunoblotting analyses allowed detection of a distinct PC-PLC
isoform with apparent molecular weight Mr = 66 kDa in fresh, totalcell lysates of all investigated EONT and EOC cells (Fig. 2A), inagreement with previous reports in other mammalian cells (15, 16,25–27, 33). By densitometric analysis, the intensity of the corre-sponding protein band in most of the investigated cancer cell lines(OVCAR3, SKOV3, and IGROV1) was 1.9F 0.5 times higher than thatobserved in nontumoral cells. Moreover, the intensity of the PC-PLCband at 66 kDa was 2.6 F 0.1–fold higher in membrane-enrichedfractions of OVCAR3 compared with immortalized cells (Fig. 2B).Amplex Red assays performed on total cell lysates showed no
significant differences (P = 0.62) in the relative PC-PLC activityvalues of nontumoral immortalized cells (IOSE and hTERT); data
Figure 1. CLSM and flow cytometry analyses of PC-PLC distribution on the plasma membrane of EOC cells compared with EONT cells. A, PC-PLC detection byCLSM analysis (three-dimensional reconstruction) on the outer membrane surface of unfixed, subconfluent EOC cell lines (SKOV3, CABAI, IGROV1, and OVCAR3),normal OSE, and nontumoral immortalized cells (IOSE, hTERT) cultured to early confluence. An example of observations on cancer cells isolated from patientexudate is also reported. Offset and gain values of the photomultiplier channel were regulated with respect to the setup selected for OSE cell analysis to makefluorescence intensity comparable across experiments on different samples. Scale bar, 20 Am. B, examples of flow cytometry profiles of unfixed EOC and EONT cells(three independent series of experiments for each analyzed cell line). Left, representative cytofluorimetric histograms (CTR, control cells, IOSE, or OVCAR3, incubatedwith rabbit IgG plus secondary antibody); center, median (F SD) values of fluorescence intensity; right , percentages of PC-PLC–positive cells (mean F SD).
PC-PLC in Ovarian Cancer
www.aacrjournals.org 6543 Cancer Res 2008; 68: (16). August 15, 2008
Research. on June 8, 2015. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

were therefore pooled together, and their mean value wasnormalized to 1.0. In this arbitrary unit scale, the PC-PLC activityof EOC cell lines (Fig. 2C) in repeated experiments was 2.9 F 0.7(FSD)–fold higher than that of normal cells (ANOVA P < 0.0001).Assays on enriched membrane fractions (Fig. 2D) showed that the
PC-PLC activity of OVCAR3 cells was 5-fold higher than that ofEONT cells, and a similar result was obtained from ascitic cancercells isolated from patient exudate.These results altogether indicated that EOC differed from EONT
cells in a substantial increase in the PC-PLC level on outer cell
Table 1. Immunohistochimical detection of PC-PLC in an ovarian cancer tissue array slide
Samples No. sections No/weak staining* Immunohistochemical staining
Strong staining* localized to
Cytoplasm Cytoplasm/membrane Cytoplasm/nucleus
Epithelial tumors: 31 4 15 6 6Mucinous 1 1
Serous 17 2 11 1 3
Mullerian 2 2
Endometrioid 5 1 2 2Clear cell 5 1 2 1 1
Brenner tumor 1 1
*Immunohistochemical score was obtained by adding the value of staining intensity (0, no staining; 1, faint staining; 2, moderate staining; 3, intensestaining) to the estimated percentage of positive cells (0, no reactive cells; 1, 1–10% reacting cells; 2, 11–25% reacting cells; 3, 26–50% reacting cells;
4, over 50% reacting cells). Weak staining, score V 3.
Figure 2. Expression and activity ofPC-PLC in cell lysates of nontumoral (OSE,IOSE, hTERT) and EOC cell lines at earlyconfluence. Western blotting of total celllysates (A ) and membrane fractions (B).h-Actin and h1 integrin were used as loadingcontrol for densitometric analyses in Aand B , respectively. Representative setsof experiments [of three (A ) or two (B )performed] are reported. PC-PLC activity(C and D ) was measured from the relativeincrease in Resorufin fluorescence in AmplexRed assay in cancer cells compared withIOSE and hTERT cells. C, relative increasein PC-PLC activity in whole-cell lysates(mean F SD of five experiments; P < 0.0001by ANOVA in EOC with respect to IOSE plushTERT cell samples); D , relative increase inPC-PLC activity in cell membrane preparations(mean F SD of three experiments; P < 0.0002by Student’s t test). The PC-PLC activity ofascitic cells from peritoneal exudates wasmeasured in triplicate (variability F 10%).
Cancer Research
Cancer Res 2008; 68: (16). August 15, 2008 6544 www.aacrjournals.org
Research. on June 8, 2015. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from
membrane, in a higher activity of this enzyme in total cell lysates,including membrane fragments and in an even higher activity inmembrane-enriched fractions.
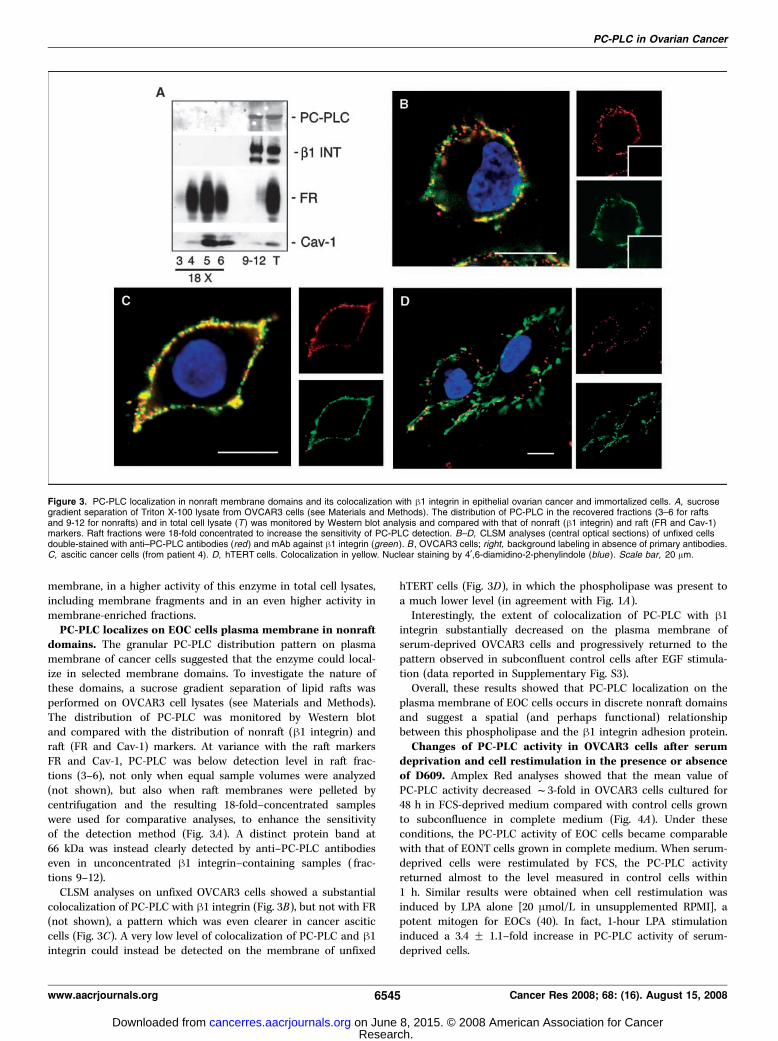
PC-PLC localizes on EOC cells plasma membrane in nonraftdomains. The granular PC-PLC distribution pattern on plasmamembrane of cancer cells suggested that the enzyme could local-ize in selected membrane domains. To investigate the nature ofthese domains, a sucrose gradient separation of lipid rafts wasperformed on OVCAR3 cell lysates (see Materials and Methods).The distribution of PC-PLC was monitored by Western blotand compared with the distribution of nonraft (h1 integrin) andraft (FR and Cav-1) markers. At variance with the raft markersFR and Cav-1, PC-PLC was below detection level in raft frac-tions (3–6), not only when equal sample volumes were analyzed(not shown), but also when raft membranes were pelleted bycentrifugation and the resulting 18-fold–concentrated sampleswere used for comparative analyses, to enhance the sensitivityof the detection method (Fig. 3A). A distinct protein band at66 kDa was instead clearly detected by anti–PC-PLC antibodieseven in unconcentrated h1 integrin–containing samples ( frac-tions 9–12).CLSM analyses on unfixed OVCAR3 cells showed a substantial
colocalization of PC-PLC with h1 integrin (Fig. 3B), but not with FR(not shown), a pattern which was even clearer in cancer asciticcells (Fig. 3C). A very low level of colocalization of PC-PLC and h1integrin could instead be detected on the membrane of unfixed
hTERT cells (Fig. 3D), in which the phospholipase was present toa much lower level (in agreement with Fig. 1A).Interestingly, the extent of colocalization of PC-PLC with h1
integrin substantially decreased on the plasma membrane ofserum-deprived OVCAR3 cells and progressively returned to thepattern observed in subconfluent control cells after EGF stimula-tion (data reported in Supplementary Fig. S3).Overall, these results showed that PC-PLC localization on the
plasma membrane of EOC cells occurs in discrete nonraft domainsand suggest a spatial (and perhaps functional) relationshipbetween this phospholipase and the h1 integrin adhesion protein.
Changes of PC-PLC activity in OVCAR3 cells after serumdeprivation and cell restimulation in the presence or absenceof D609. Amplex Red analyses showed that the mean value ofPC-PLC activity decreased f3-fold in OVCAR3 cells cultured for48 h in FCS-deprived medium compared with control cells grownto subconfluence in complete medium (Fig. 4A). Under theseconditions, the PC-PLC activity of EOC cells became comparablewith that of EONT cells grown in complete medium. When serum-deprived cells were restimulated by FCS, the PC-PLC activityreturned almost to the level measured in control cells within1 h. Similar results were obtained when cell restimulation wasinduced by LPA alone [20 Amol/L in unsupplemented RPMI], apotent mitogen for EOCs (40). In fact, 1-hour LPA stimulationinduced a 3.4 F 1.1–fold increase in PC-PLC activity of serum-deprived cells.
Figure 3. PC-PLC localization in nonraft membrane domains and its colocalization with h1 integrin in epithelial ovarian cancer and immortalized cells. A, sucrosegradient separation of Triton X-100 lysate from OVCAR3 cells (see Materials and Methods). The distribution of PC-PLC in the recovered fractions (3–6 for raftsand 9-12 for nonrafts) and in total cell lysate (T ) was monitored by Western blot analysis and compared with that of nonraft (h1 integrin) and raft (FR and Cav-1)markers. Raft fractions were 18-fold concentrated to increase the sensitivity of PC-PLC detection. B–D, CLSM analyses (central optical sections) of unfixed cellsdouble-stained with anti–PC-PLC antibodies (red) and mAb against h1 integrin (green ). B , OVCAR3 cells; right, background labeling in absence of primary antibodies.C, ascitic cancer cells (from patient 4). D, hTERT cells. Colocalization in yellow. Nuclear staining by 4¶,6-diamidino-2-phenylindole (blue ). Scale bar, 20 Am.
PC-PLC in Ovarian Cancer
www.aacrjournals.org 6545 Cancer Res 2008; 68: (16). August 15, 2008
Research. on June 8, 2015. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from
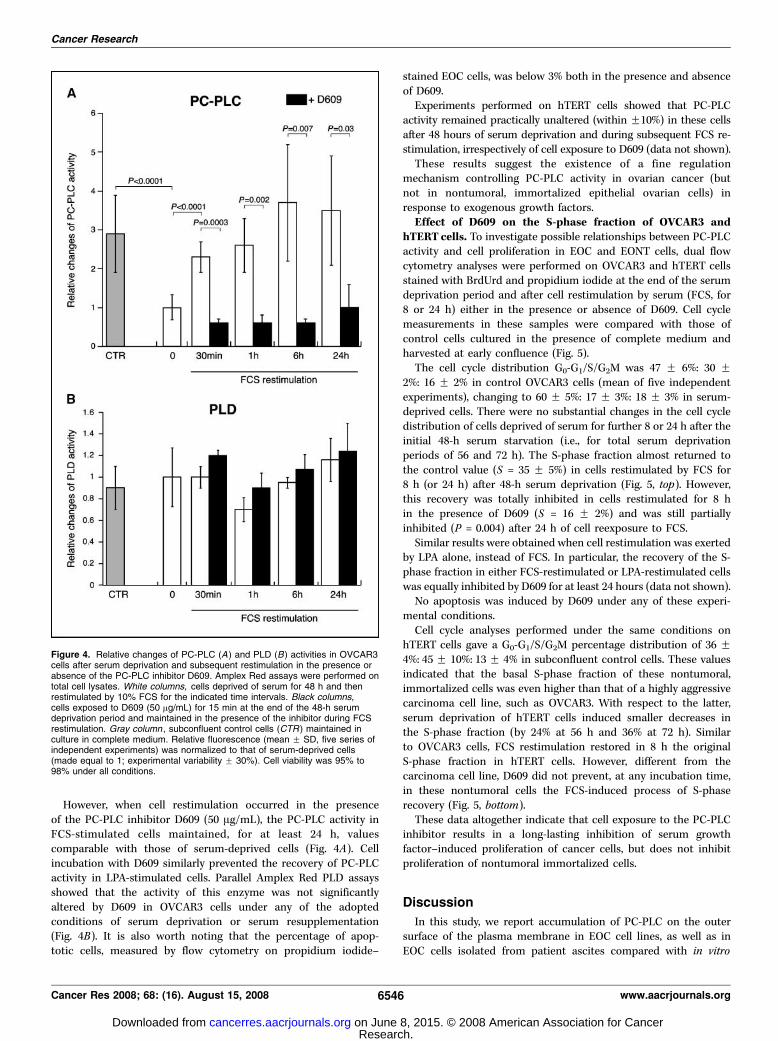
However, when cell restimulation occurred in the presenceof the PC-PLC inhibitor D609 (50 Ag/mL), the PC-PLC activity inFCS-stimulated cells maintained, for at least 24 h, valuescomparable with those of serum-deprived cells (Fig. 4A). Cellincubation with D609 similarly prevented the recovery of PC-PLCactivity in LPA-stimulated cells. Parallel Amplex Red PLD assaysshowed that the activity of this enzyme was not significantlyaltered by D609 in OVCAR3 cells under any of the adoptedconditions of serum deprivation or serum resupplementation(Fig. 4B). It is also worth noting that the percentage of apop-totic cells, measured by flow cytometry on propidium iodide–
stained EOC cells, was below 3% both in the presence and absenceof D609.Experiments performed on hTERT cells showed that PC-PLC
activity remained practically unaltered (within F10%) in these cellsafter 48 hours of serum deprivation and during subsequent FCS re-stimulation, irrespectively of cell exposure to D609 (data not shown).These results suggest the existence of a fine regulation
mechanism controlling PC-PLC activity in ovarian cancer (butnot in nontumoral, immortalized epithelial ovarian cells) inresponse to exogenous growth factors.
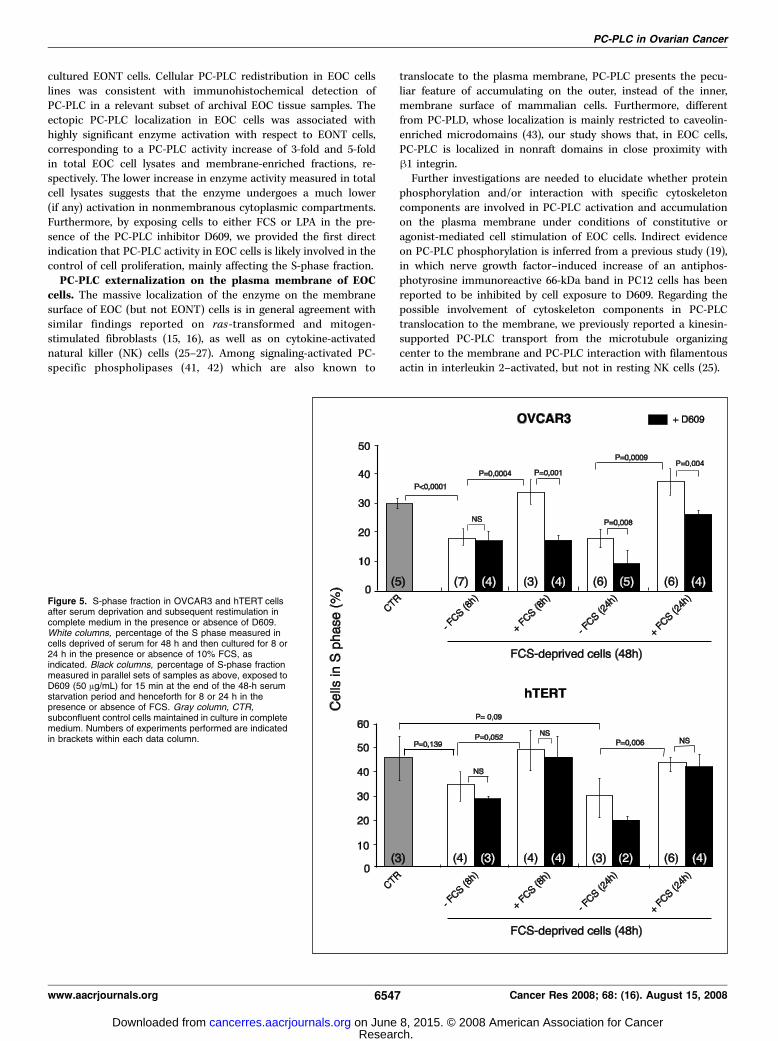
Effect of D609 on the S-phase fraction of OVCAR3 andhTERT cells. To investigate possible relationships between PC-PLCactivity and cell proliferation in EOC and EONT cells, dual flowcytometry analyses were performed on OVCAR3 and hTERT cellsstained with BrdUrd and propidium iodide at the end of the serumdeprivation period and after cell restimulation by serum (FCS, for8 or 24 h) either in the presence or absence of D609. Cell cyclemeasurements in these samples were compared with those ofcontrol cells cultured in the presence of complete medium andharvested at early confluence (Fig. 5).The cell cycle distribution G0-G1/S/G2M was 47 F 6%: 30 F
2%: 16 F 2% in control OVCAR3 cells (mean of five independentexperiments), changing to 60 F 5%: 17 F 3%: 18 F 3% in serum-deprived cells. There were no substantial changes in the cell cycledistribution of cells deprived of serum for further 8 or 24 h after theinitial 48-h serum starvation (i.e., for total serum deprivationperiods of 56 and 72 h). The S-phase fraction almost returned tothe control value (S = 35 F 5%) in cells restimulated by FCS for8 h (or 24 h) after 48-h serum deprivation (Fig. 5, top). However,this recovery was totally inhibited in cells restimulated for 8 hin the presence of D609 (S = 16 F 2%) and was still partiallyinhibited (P = 0.004) after 24 h of cell reexposure to FCS.Similar results were obtained when cell restimulation was exerted
by LPA alone, instead of FCS. In particular, the recovery of the S-phase fraction in either FCS-restimulated or LPA-restimulated cellswas equally inhibited by D609 for at least 24 hours (data not shown).No apoptosis was induced by D609 under any of these experi-
mental conditions.Cell cycle analyses performed under the same conditions on
hTERT cells gave a G0-G1/S/G2M percentage distribution of 36 F4%: 45 F 10%: 13 F 4% in subconfluent control cells. These valuesindicated that the basal S-phase fraction of these nontumoral,immortalized cells was even higher than that of a highly aggressivecarcinoma cell line, such as OVCAR3. With respect to the latter,serum deprivation of hTERT cells induced smaller decreases inthe S-phase fraction (by 24% at 56 h and 36% at 72 h). Similarto OVCAR3 cells, FCS restimulation restored in 8 h the originalS-phase fraction in hTERT cells. However, different from thecarcinoma cell line, D609 did not prevent, at any incubation time,in these nontumoral cells the FCS-induced process of S-phaserecovery (Fig. 5, bottom).These data altogether indicate that cell exposure to the PC-PLC
inhibitor results in a long-lasting inhibition of serum growthfactor–induced proliferation of cancer cells, but does not inhibitproliferation of nontumoral immortalized cells.
Discussion
In this study, we report accumulation of PC-PLC on the outersurface of the plasma membrane in EOC cell lines, as well as inEOC cells isolated from patient ascites compared with in vitro
Figure 4. Relative changes of PC-PLC (A ) and PLD (B) activities in OVCAR3cells after serum deprivation and subsequent restimulation in the presence orabsence of the PC-PLC inhibitor D609. Amplex Red assays were performed ontotal cell lysates. White columns, cells deprived of serum for 48 h and thenrestimulated by 10% FCS for the indicated time intervals. Black columns,cells exposed to D609 (50 Ag/mL) for 15 min at the end of the 48-h serumdeprivation period and maintained in the presence of the inhibitor during FCSrestimulation. Gray column , subconfluent control cells (CTR ) maintained inculture in complete medium. Relative fluorescence (mean F SD, five series ofindependent experiments) was normalized to that of serum-deprived cells(made equal to 1; experimental variability F 30%). Cell viability was 95% to98% under all conditions.
Cancer Research
Cancer Res 2008; 68: (16). August 15, 2008 6546 www.aacrjournals.org
Research. on June 8, 2015. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from
cultured EONT cells. Cellular PC-PLC redistribution in EOC cellslines was consistent with immunohistochemical detection ofPC-PLC in a relevant subset of archival EOC tissue samples. Theectopic PC-PLC localization in EOC cells was associated withhighly significant enzyme activation with respect to EONT cells,corresponding to a PC-PLC activity increase of 3-fold and 5-foldin total EOC cell lysates and membrane-enriched fractions, re-spectively. The lower increase in enzyme activity measured in totalcell lysates suggests that the enzyme undergoes a much lower(if any) activation in nonmembranous cytoplasmic compartments.Furthermore, by exposing cells to either FCS or LPA in the pre-sence of the PC-PLC inhibitor D609, we provided the first directindication that PC-PLC activity in EOC cells is likely involved in thecontrol of cell proliferation, mainly affecting the S-phase fraction.
PC-PLC externalization on the plasma membrane of EOCcells. The massive localization of the enzyme on the membranesurface of EOC (but not EONT) cells is in general agreement withsimilar findings reported on ras -transformed and mitogen-stimulated fibroblasts (15, 16), as well as on cytokine-activatednatural killer (NK) cells (25–27). Among signaling-activated PC-specific phospholipases (41, 42) which are also known to
translocate to the plasma membrane, PC-PLC presents the pecu-liar feature of accumulating on the outer, instead of the inner,membrane surface of mammalian cells. Furthermore, differentfrom PC-PLD, whose localization is mainly restricted to caveolin-enriched microdomains (43), our study shows that, in EOC cells,PC-PLC is localized in nonraft domains in close proximity withh1 integrin.Further investigations are needed to elucidate whether protein
phosphorylation and/or interaction with specific cytoskeletoncomponents are involved in PC-PLC activation and accumulationon the plasma membrane under conditions of constitutive oragonist-mediated cell stimulation of EOC cells. Indirect evidenceon PC-PLC phosphorylation is inferred from a previous study (19),in which nerve growth factor–induced increase of an antiphos-photyrosine immunoreactive 66-kDa band in PC12 cells has beenreported to be inhibited by cell exposure to D609. Regarding thepossible involvement of cytoskeleton components in PC-PLCtranslocation to the membrane, we previously reported a kinesin-supported PC-PLC transport from the microtubule organizingcenter to the membrane and PC-PLC interaction with filamentousactin in interleukin 2–activated, but not in resting NK cells (25).
Figure 5. S-phase fraction in OVCAR3 and hTERT cellsafter serum deprivation and subsequent restimulation incomplete medium in the presence or absence of D609.White columns, percentage of the S phase measured incells deprived of serum for 48 h and then cultured for 8 or24 h in the presence or absence of 10% FCS, asindicated. Black columns, percentage of S-phase fractionmeasured in parallel sets of samples as above, exposed toD609 (50 Ag/mL) for 15 min at the end of the 48-h serumstarvation period and henceforth for 8 or 24 h in thepresence or absence of FCS. Gray column, CTR,subconfluent control cells maintained in culture in completemedium. Numbers of experiments performed are indicatedin brackets within each data column.
PC-PLC in Ovarian Cancer
www.aacrjournals.org 6547 Cancer Res 2008; 68: (16). August 15, 2008
Research. on June 8, 2015. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

Regarding functional aspects, externalization of PC-PLC on thecell membrane may confer to this activated enzyme the capabilityof participating in membrane receptor-mediated cell signaling, byproducing diacyl glycerol (DG) second messengers from hydrolysisof PC, the major phospholipid component of the outer layer ofeukaryotic cells. In fact, a persistent production of PC-derived DGsis reputed to contribute to long-lasting (PKC-mediated or PKC-independent) signaling events ending with gene transcription andDNA synthesis (43, 44). In this context, it is interesting to note thateven exogenously added bacterial PC-PLC has been shown to elicita potent mitogenic response in fibroblasts by promoting DNAsynthesis and cell proliferation (45).Furthermore, the ectopic localization of activated PC-PLC on
cancer cell membrane may allow this enzyme to modulate theexpression of some membrane receptors. In fact, we recentlyreported evidence on the crucial role of ectopically localizedactivated PC-PLC in selectively regulating the expression of CD16,the FcIgG receptor on the membrane of human NK cells, withstriking effects on CD16-mediated cytotoxicity (27). The changesreported herein of PC-PLC and h1 integrin colocalization on themembrane of serum-deprived and EGF-restimulated OVCAR3 cells(Supplementary Fig. S3) may point to a similar role of PC-PLC inregulating the expression and signaling of this adhesion protein inEOC cells. A previous report on the mutual relationships betweenPC-PLC and h1 integrin in regulating cell-substratum adhesion invascular endothelial cells (46) seems to reinforce this hypothesis.Studies are in progress to further investigate and quantify possibleinteractions of PC-PLC with this adhesion molecule under differentconditions of EGF stimulation.
Relationships between PC-PLC activity and S-phase fractionin EOC cells. Serum deprivation caused a 3-fold decrease in thePC-PLC activity of OVCAR3 cells, inducing a drop in the activityof this enzyme down to the level steadily measured in EONT cells.PC-PLC down-regulation was associated in EOC cells with a 40%decrease in the S-phase fraction of serum-deprived OVCAR3 cells.PC-PLC reactivation in FCS-stimulated or LPA-stimulated OVCAR3cells preceded a full recovery of the S-phase fraction to the valuemeasured before serum deprivation. Similarly, although quantita-tively different, changes in PC-PLC activity (5-fold decrease) andS-phase fraction (f100% drop) were reported in serum-deprivedfibroblasts, where subsequent cell restimulation by platelet-derivedgrowth factor (PDGF) promptly restored PC-PLC activity and theoriginal proliferative status (16). The higher residual percentage ofPC-PLC activity maintained in serum-deprived EOC cells comparedwith the residual PC-PLC activity reported in serum-deprivednormal fibroblasts may be attributed to the well-known reduceddependence of cancer cells upon exogenous growth factors fortheir survival and proliferation (47). The long-lasting ( for at least24 h) inhibition on the recovery of PC-PLC activity induced byD609 in serum-restimulated or LPA-restimulated OVCAR3 cells isin general agreement with a similar effect observed in PDGF-stimulated fibroblasts (16), suggesting that altered PC-PLC activity
and changes in the S-phase fraction may both be linked to commonmitogen-dependent pathways. Further investigations are requiredto elucidate the nature of the underlying molecular mechanisms.The present study also shows that the suggested relationships
between PC-PLC activity and S-phase fraction in OVCAR3 do nothold true for the nontumoral immortalized hTERT cells, in whichsubcellular PC-PLC localization was mainly confined to innercellular compartments. In spite of their higher S-phase fraction atsubconfluence (probably due to telomerase transfection), thesecells maintained a 3-fold lower PC-PLC activity compared withOVCAR3 cells and no substantial change was observed in PC-PLCactivity in serum-deprived and FCS-restimulated cells, either in thepresence or in the absence of D609. Noteworthy, only scantyalterations were observed in the S-phase fraction of hTERT cellsexposed to D609.Further studies are needed to elucidate whether other enzymes,
besides PC-PLC, may also be directly or indirectly involved in D609-induced effects on EOC cell proliferation. In fact, it has been shownthat PC-PLC inhibition by D609 may also interfere with cellsignaling, by blocking, in some cell systems, mitogen-inducedprotein kinase activation (48, 49) or by inhibiting other enzymes,such as sphingomyelin synthase (50).In overall, the results reported in the present work support the
view that PC-PLC is directly involved in the complex machinery ofpathways responsible for EOC cell proliferation. Further evidenceon these mechanisms is expected from PC-PLC sequencing andcloning, which would allow a better elucidation of the role of thisenzyme in the multiple pathways controlling the development ofaberrant phenotypes typical of these (and probably other)malignant cells, together with the possible identification of noveltargets for selective antitumor therapies. At the present stage, thedifferent involvement of PC-PLC in EONT and EOC cellproliferation suggests that the use of a PC-PLC inhibitor (possiblyone even more effective and specific than D609) might provide apowerful method to selectively interfere, likely in combination withantitumor drugs, with some of the aberrant pathways responsiblefor EOC proliferation and invasiveness.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
Received 12/20/2007; revised 5/28/2008; accepted 6/6/2008.Grant support: Cassa di Risparmio delle Provincie Lombarde Foundation
CARIPLO grant 2003.1740/10.4890 (S. Canevari and F. Podo) and Associazione Italianaper la Ricerca sul Cancro grant 2007 (F. Podo).
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
We thank Dr. C. Pini and Dr. B. Barletta, Istituto Superiore di Sanita, for preparationand purification of anti–PC-PLC antibodies, the clinical staffs of the Gynecological andPathologic Units of Fondazione IRCCS Istituto Nazionale dei Tumori for providingpatients’ materials, and Marco Sabatini for excellent graphical assistance.
References
1. Jemal A, Siegel R, Ward E, Murray T, Xu J, Thun MJ.Cancer statistics, 2007. CA Cancer J Clin 2007;55:43–66.
2. Ozols RF, Bookman MA, Connolly DC, et al. Focus onepithelial ovarian cancer. Cancer Cell 2004;5:19–24.
3. Negendank WG. Studies of human tumors by MRS: areview. NMR Biomed 1992;5:303–24.
4. Podo F. Tumour phospholipid metabolism. Reviewarticle. NMR Biomed 1999;12:413–39.
5. Podo F, Sardanelli F, Iorio E, et al. Abnormal cholinephospholipid metabolism in breast and ovary cancer:
molecular bases for noninvasive imaging approaches.Curr Mol Imag Rev 2007;3:123–37.
6. Aboagye EO, Bhujwalla ZM. Malignant transformationalters membrane choline phospholipid metabolism ofhuman mammary epithelial cells. Cancer Res 1999;59:80–4.
Cancer Research
Cancer Res 2008; 68: (16). August 15, 2008 6548 www.aacrjournals.org
Research. on June 8, 2015. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

PC-PLC in Ovarian Cancer
www.aacrjournals.org 6549 Cancer Res 2008; 68: (16). August 15, 2008
7. Ackerstaff E, Pflug BR, Nelson JB, Bhujwalla ZM.Detection of increased choline compounds with protonnuclear magnetic resonance spectroscopy subsequent tomalignant transformation of human prostatic epithelialcells. Cancer Res 2001;61:3599–603.
8. Iorio E, Mezzanzanica D, Alberti P, et al. Alterations ofcholine phospholipid metabolism in ovarian tumorprogression. Cancer Res 2005;65:9369–76.
9. Ackerstaff E, Glunde K, Bhujwalla ZW. Cholinephospholipid metabolism: a target in cancer cells?J Cell Biochem 2003;90:525–33.
10. Glunde K, Ackerstaff E, Mori N, Jacobs MA, BhujwallaZM. Choline phospholipid metabolism in cancer:consequences for molecular pharmaceutical interven-tions. Mol Pharm 2006;3:496–506.
11. Glunde K, Serkova NJ. Therapeutic targets andbiomarkers identified in cancer choline phospholipidmetabolism. Pharmacogenomics 2006;7:1109–23.
12. Ramirez de Molina A, Rodriguez-Gonzalez A,Gutierrez R, et al. Overexpression of choline kinase isa frequent feature in human tumor-derived cell linesand in lung, prostate, and colorectal human cancers.Biochem Biophys Res Commun 2002;296:580–3.
13. Glunde K, Raman V, Mori N, Bhujwalla ZM. RNAinterference-mediated choline kinase suppression inbreast cancer cells induces differentiation and reducesproliferation. Cancer Res 2005;65:11034–43.
14. Al-Saffar NM, Troy H, Ramirez de Molina A, et al.Noninvasive magnetic resonance spectroscopic phar-macodynamic markers of the choline kinase inhibitorMN58b in human carcinoma models. Cancer Res 2006;66:427–34.
15. Podo F, Ferretti A, Knijn A, et al. Detection ofphosphatidylcholine-specific phospholipase C in NIH-3T3 fibroblasts and their H-ras transformants: NMRand immunochemical studies. Anticancer Res 1996;16:1399–412.
16. Ramoni C, Spadaro F, Barletta B, Dupuis ML andPodo F. Phosphatidylcholine-specific phospholipases Cin mitogen-stimulated fibroblasts. Exp Cell Res 2004;299:370–82.
17. Van Dijk MC, Muriana FJ, de Widt J, Hilkmann H, vanBlittezswijk WJ. Involvement of phosphatidylcholine-specific phospholipases C in platelet-derived growthfactor-induced activation of the mitogen-activated pro-tein kinase pathway in Rat-1 fibroblasts. J Biol Chem 1997;272:11011–6.
18. Li F, Wu N, Su R-B, et al. Involvement ofphosphatidylcholine-selective phospholipases C in acti-vation of mitogen-activated protein kinase pathways inimidazoline receptor antisera-selected protein. J CellBiochem 2006;98:1615–28.
19. Kahle PJ, Shooter EM, Johnson RM, Verity AN.Phosphatidylcholine-specific phospholipases inhibitorD609 differentially affects MAP kinases and immedi-ate-early genes in PC12 cells. Cell Signal 1998;10:321–30.
20. Wang N, Xie K, Huo S, Zhao J, Zhang S, Miao J.Suppressing phosphatidylcholine-specific phospholi-pases C and elevating ROS level, NADPH oxidaseactivity and Rb level induced neuronal differentiationin mesenchimal stem cells. J Cell Biochem 2007;100:1548–57.
21. Zhao J, Zhao B, Wang W, Huang B, Zhang S, Miao J.Phosphatidylcholine-specific phospholipases C and ROSwere involved in chicken blastodisc differentiation tovascular endothelial cell. J Cell Biochem 2007;102:421–8.
22. Cifone MG, Rocaioli P, De Maria R, et al. Multiplepathways originated at the Fas/Apo-1 (CD95) receptor:sequential involvement of phosphatidylcholine-specificphospholipase C and acidic sphingomyelinase in thepropagation of the apoptotic signal. EMBO J 1995;14:5859–68.
23. Liu X, Zhao Q, Araki S, Zhang S, Miao J. Contrastingeffects of phosphatidylcholine-specific phospholipases Con apoptosis in cultured endothelial cells. Endothelium2006;13:205–11.
24. von Knethen A, Brune B. PKC a depletion inRAW264.7 macrophages following microbial/IFNg stim-ulation is PC-PLC-mediated. Antioxid Redox Signal2005;7:1217–22.
25. Ramoni C, Spadaro F, Menegon M, Podo F. Cellularlocalization and functional role of phosphatidylcholine-specific phospholipases C in NK cells. J Immunol 2001;167:2642–50.
26. Spadaro F, Cecchetti S, Sanchez M, Ausiello CM,Podo F, Ramoni C. Expression and role of phosphati-dylcholine-specific phospholipase C in human NK andT-lymphocyte subsets. Eur J Immunol 2006;36:3277–87.
27. Cecchetti S, Spadaro F, Lugini L, Podo F, Ramoni C.Functional role of phosphatidylcholine-specific phos-pholipase C in regulating CD16 membrane expression innatural killer cells. Eur J Immunol 2007;37:2912–22.
28. Moreno-Garcia ME, Lopez-Bojorques LN, Zentella A,Humphries LA, Rawlings DJ, Santos-Argumedo L. CD38signaling regulates B lymphocyte activation via aphospholipases C (PLC)-g2-independent, protein kinaseC, phosphatidylcholine-PLC, and phospholipase D-dependent signalling cascade. J Immunol 2005;174:2687–95.
29. Andrei C, Margiocco P, Poggi A, Lotti LV, TorrisiMR, Rubartelli A. Phospholipases C and A2 controllysosome-mediated IL-h secretion: implications forinflammatory processes. Proc Natl Acad Sci U S A2004;101:9745–50.
30. Luft T, Rodionova E, Maraskovsky E, et al. Adaptivefunctional differentiation of dendritic cells: integratingthe network of extra- and intracellular signals. Blood2006;107:4763–9.
31. Glunde K, Jie C, Bhujwalla ZM. Molecular causes ofthe aberrant choline phospholipid metabolism in breastcancer. Cancer Res 2004;64:4270–6.
32. Clark MA, Shorr RGL, Bomalaski JS. Antibodiesprepared to Bacillus cereus phospholipase C cross reactwith a phosphatidylcholine preferring phospholipase Cin mammalian cells. Biochim Biophys Res Commun1986;140:114–9.
33. Mateas MV, Uranga RM, Salvador GA, Giusto NM.Coexistence of phosphatidylcholine-specific phospholi-pase C and phospholipase D activities in rat cerebralcortex synaptosomes. Lipids 2006;41:273–80.
34. Miotti S, Canevari S, Menard S, et al. Characterizationof human ovarian carcinoma-associated antigens de-fined by novel monoclonal antibodies with tumor-restricted specificity. Int J Cancer 1987;39:297–303.
35. Pellegrini R, Bazzini P, Tosi E, et al. Production andcharacterization of two monoclonal antibodies directedagainst the integrin h1 chain. Tumori 1992;78:1–4.
36. Bagnoli M, Balladore E, Luison E, et al. Sensitizationof p53-mutated epithelial ovarian cancer to CD95-mediated apoptosis is synergistically induced by cis-platin pretreatment. Mol Cancer Ther 2007;6:762–72.
37. Vachon L, Costa T, Hertz A. GTPase and adenylatecyclase desensitize at different rates in NG108–15 cells.Mol Pharmacol 1987;31:159–68.
38. Miotti S, Bagnoli M, Tomassetti A, Colnaghi MI,Canevari S. Interaction of folate receptor with signalingmolecules lyn and G(a)(i-3) in detergent-resistantcomplexes from the ovary carcinoma cell line IGROV1.J Cell Sci 2000;113:349–57.
39. Sargiacomo M, Sudol M, Tang Z, Lisanti MP. Signaltransducing molecules and glycosyl-phosphatidylinositol-linked proteins form a caveolin-rich insoluble complex inMDCK cells. J Cell Biol 1993;122:789–807.
40. Lee Z, Swaby RF, Liang Y, et al. Lysophosphatidic acidis a major regulator of growth-regulator oncogene a inovarian cancer. Cancer Res 2006;66:2740–8.
41. McDermott M, Wakelam MJO, Morris AJ. Phospho-lipase D. Biochem Cell Biol 2004;82:225–53.
42. Chakraborti S. Phospholipase A2 isoforms: a perspec-tive. Cell Signal 2003;15:637–65.
43. Xu L, Shen Y, Joseph T, et al. Mitogenic phospholipaseD activity is restricted to caveolin-enriched membranemicrodomains. Biochem Biophys Res Commun 2000;273:77–83.
44. Liskovitch M. Cross talk among multiple signal-activated phospholipases. Trends Biochem Sci 1992;17:393–9.
45. Larrodera P, Cornet ME, Diaz-Meco MJ, et al.Phospholipase C-mediated hydrolysis of phosphatidyl-choline is an important step in PDGF-stimulated DNAsynthesis. Cell 1990;61:1113–20.
46. Miao J-H, Araki S, Hayashi H. Relationships betweenphosphatidylcholine-specific phospholipase C andintegrins in cell-substratum adhesion and apoptosis invascular endothelial cells. Endothelium 1997;5:287–305.
47. Fang X, Schummer M, Mao M, et al. Lysophospha-tidic acid is a bioactive mediator in ovarian cancer.Biochim Biophys Acta 2002;1582:257–64.
48. Singh ATK, Radeff JM, Kunnel JG, Stern PH.Phosphatidylcholine-specific phospholipase C inhibitor,tricyclodecan-9-yl xanthogenate (D609), increases phos-pholipase D-mediated phosphatidylcholine hydrolysis inUMR-106 osteoblastic osteosarcoma cells. BiochimBiophys Acta 2000;1487:201–8.
49. Van Dijk MC, Muriana FJ, de Vidt J, Hilkmann H,van Blitterswjk WJ. Involvement of phosphatidylcholine-specific phospholipase C in platelet-derived growthfactor-induced activation of the mitogen-activatedprotein kinase pathway in Rat-1 fibroblasts. J Biol Chem1997;272:11011–6.
50. Luberto C, Hannun YA. Shingomyelin synthase, apotential regulator of intracellular levels of ceramideand diacylglycerol during SV40 transformation: doessphingomyelin synthase account for the putativephosphatidylcholine specific phospholipase C? J BiolChem 1998;273:14550–9.
Research. on June 8, 2015. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from
2008;68:6541-6549. Cancer Res Francesca Spadaro, Carlo Ramoni, Delia Mezzanzanica, et al. Epithelial Ovarian Cancer CellsPhosphatidylcholine-Specific Phospholipase C Activation in
Updated version
http://cancerres.aacrjournals.org/content/68/16/6541
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2008/08/11/68.16.6541.DC1.html
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/68/16/6541.full.html#ref-list-1
This article cites 50 articles, 18 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/68/16/6541.full.html#related-urls
This article has been cited by 4 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on June 8, 2015. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from




















