Expression and action of keratinocyte growth factor (KGF) in normal ovarian surface epithelium and...
11
Molecular and Cellular Endocrinology 167 (2000) 77 – 87 Expression and action of keratinocyte growth factor (KGF) in normal ovarian surface epithelium and ovarian cancer Jeff A. Parrott 1 , Grace Kim, Rachel Mosher, Michael K. Skinner * Center for Reproducti6e Biology, Department of Genetics and Cell Biology, Washington State Uni6ersity, Pullman, WA 99163 -4231, USA Received 27 March 2000; accepted 23 May 2000 Abstract The current study investigates the expression and action of keratinocyte growth factor (KGF) in normal ovarian surface epithelium (OSE) and ovarian cancer tissues. Ovarian tumors are primarily derived from the OSE. KGF is a mesenchymal cell-derived growth factor that mediates stromal cell-epithelial cell interactions in a variety of different tissues. Human ovarian tumors from borderline, stage I and stage III cases were found to express KGF protein in the epithelial cell component by immunocytochemical analysis. The stromal cell component of human ovarian tumors contained little or no KGF immunostaining. Normal bovine ovaries have similarities to human ovaries and are used as a model system to investigate normal OSE functions. KGF protein was detected in the OSE from normal human and bovine ovaries by immunocytochemistry. Ovarian stromal tissue contained light but positive KGF immunostaining. RNA was collected from normal bovine OSE and ovarian stromal cells to examine KGF gene expression. KGF transcripts were detected in cultured OSE and stromal cells by Northern blot analysis. In order to examine and quantitate KGF gene expression in freshly isolated versus cultured tissues, a sensitive quantitative RT-PCR assay for KGF was utilized. KGF gene expression was found to be high in freshly isolated OSE, but very low in freshly isolated stroma. Levels of KGF gene expression after culture of OSE and stromal cells increased. Observations indicate that normal OSE express high levels of KGF in vivo and in vitro. Expression of KGF by normal epithelial cells versus stromal cells was unexpected and suggests KGF may be an important autocrine stimulator of OSE. KGF actions on bovine OSE cells were investigated. KGF was found to stimulate the growth of normal OSE cells to the same level as epidermal growth factor. Two ovarian cancer cell lines, SKOV3 and OCC1, were also stimulated to proliferate in response to KGF. Current results demonstrate the production and action of KGF on normal OSE cells and ovarian cancer cells. Observations can be interpreted to suggest that KGF may in part be involved in the growth of ovarian tumors. This appears to be one of the first reports of KGF production by an epithelial cell. The autocrine stimulation of OSE growth by the local production and action of KGF provides insight into how the OSE may develop abnormal growth characteristics involved in the onset and progression of ovarian cancer. © 2000 Elsevier Science Ireland Ltd. All rights reserved. Keywords: Keratinocyte growth factor; Mesenchymal-epithelium; Ovary; Ovarian surface epithelium; Ovarian cancer www.elsevier.com/locate/mce 1. Introduction Ovarian cancer causes more deaths than any other cancer of the female reproductive system. It is estimated that there will be approximately 20 000 deaths this year from ovarian cancer in the US (Cancer Facts and Figures, 1998). Although a small number of ovarian cancers originate from cells associated with the ovarian follicle, greater than 95% of ovarian cancers originate in the epithelial cells on the surface of the ovary (Weiss et al., 1977; Piver et al., 1991). The epithelial cells that cover the surface of the ovary are most commonly referred to as the ovarian surface epithelium (OSE) which is a modified mesothelium. The OSE is a simple epithelium separated from underlying ovarian stromal tissue by a basal lamina (Nicosia and Nicosia, 1988; Auersperg et al., 1991). During normal ovarian function the OSE undergoes cyclic changes. It can release enzymes that contribute to the breakdown of the underlying stroma that is adjacent to the preovu- * Corresponding author. Tel.: +1-509-3351524; fax: +1-509- 3352176. E-mail address: [email protected] (M.K. Skinner). 1 Present address: Atairgin Technologies Inc., 4 Jenner, Suite 180, Irvine, CA 92618, USA. 0303-7207/00/$ - see front matter © 2000 Elsevier Science Ireland Ltd. All rights reserved. PII:S0303-7207(00)00284-7
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Expression and action of keratinocyte growth factor (KGF) in normal ovarian surface epithelium and...

Molecular and Cellular Endocrinology 167 (2000) 77–87
Expression and action of keratinocyte growth factor (KGF) innormal ovarian surface epithelium and ovarian cancer
Jeff A. Parrott 1, Grace Kim, Rachel Mosher, Michael K. Skinner *Center for Reproducti6e Biology, Department of Genetics and Cell Biology, Washington State Uni6ersity, Pullman, WA 99163-4231, USA
Received 27 March 2000; accepted 23 May 2000
Abstract
The current study investigates the expression and action of keratinocyte growth factor (KGF) in normal ovarian surfaceepithelium (OSE) and ovarian cancer tissues. Ovarian tumors are primarily derived from the OSE. KGF is a mesenchymalcell-derived growth factor that mediates stromal cell-epithelial cell interactions in a variety of different tissues. Human ovariantumors from borderline, stage I and stage III cases were found to express KGF protein in the epithelial cell component byimmunocytochemical analysis. The stromal cell component of human ovarian tumors contained little or no KGF immunostaining.Normal bovine ovaries have similarities to human ovaries and are used as a model system to investigate normal OSE functions.KGF protein was detected in the OSE from normal human and bovine ovaries by immunocytochemistry. Ovarian stromal tissuecontained light but positive KGF immunostaining. RNA was collected from normal bovine OSE and ovarian stromal cells toexamine KGF gene expression. KGF transcripts were detected in cultured OSE and stromal cells by Northern blot analysis. Inorder to examine and quantitate KGF gene expression in freshly isolated versus cultured tissues, a sensitive quantitative RT-PCRassay for KGF was utilized. KGF gene expression was found to be high in freshly isolated OSE, but very low in freshly isolatedstroma. Levels of KGF gene expression after culture of OSE and stromal cells increased. Observations indicate that normal OSEexpress high levels of KGF in vivo and in vitro. Expression of KGF by normal epithelial cells versus stromal cells was unexpectedand suggests KGF may be an important autocrine stimulator of OSE. KGF actions on bovine OSE cells were investigated. KGFwas found to stimulate the growth of normal OSE cells to the same level as epidermal growth factor. Two ovarian cancer celllines, SKOV3 and OCC1, were also stimulated to proliferate in response to KGF. Current results demonstrate the production andaction of KGF on normal OSE cells and ovarian cancer cells. Observations can be interpreted to suggest that KGF may in partbe involved in the growth of ovarian tumors. This appears to be one of the first reports of KGF production by an epithelial cell.The autocrine stimulation of OSE growth by the local production and action of KGF provides insight into how the OSE maydevelop abnormal growth characteristics involved in the onset and progression of ovarian cancer. © 2000 Elsevier Science IrelandLtd. All rights reserved.
Keywords: Keratinocyte growth factor; Mesenchymal-epithelium; Ovary; Ovarian surface epithelium; Ovarian cancer
www.elsevier.com/locate/mce
1. Introduction
Ovarian cancer causes more deaths than any othercancer of the female reproductive system. It is estimatedthat there will be approximately 20 000 deaths this yearfrom ovarian cancer in the US (Cancer Facts andFigures, 1998). Although a small number of ovarian
cancers originate from cells associated with the ovarianfollicle, greater than 95% of ovarian cancers originatein the epithelial cells on the surface of the ovary (Weisset al., 1977; Piver et al., 1991).
The epithelial cells that cover the surface of the ovaryare most commonly referred to as the ovarian surfaceepithelium (OSE) which is a modified mesothelium. TheOSE is a simple epithelium separated from underlyingovarian stromal tissue by a basal lamina (Nicosia andNicosia, 1988; Auersperg et al., 1991). During normalovarian function the OSE undergoes cyclic changes. Itcan release enzymes that contribute to the breakdownof the underlying stroma that is adjacent to the preovu-
* Corresponding author. Tel.: +1-509-3351524; fax: +1-509-3352176.
E-mail address: [email protected] (M.K. Skinner).1 Present address: Atairgin Technologies Inc., 4 Jenner, Suite 180,
Irvine, CA 92618, USA.
0303-7207/00/$ - see front matter © 2000 Elsevier Science Ireland Ltd. All rights reserved.PII: S 0 3 0 3 -7207 (00 )00284 -7

J.A. Parrott et al. / Molecular and Cellular Endocrinology 167 (2000) 77–8778
latory follicle (Woessner et al., 1989; Bjersing and Ca-jander, 1975). After ovulation, the OSE proliferates andcovers the area affected by follicular rupture (Oster-holze et al., 1985). The hypothesis has developed thatrepetitious ovulation contributes to the etiology ofovarian cancer through the altered cellular activity ofthe OSE at ovulation (Godwin et al., 1992; Hornby etal., 1992). Tumorigenic tissue derived from the OSEalso has close associations with stromal tissue. Tumorinvasion often requires an association with host stromaltissue and most ovarian tumors have a stromal-likecomponent (Scully, 1979; Kurman, 1987) that is re-cruited from the host site. Therefore, stromal-epithelialcell interactions appear to have a critical role in thefunction and growth of normal and tumorigenic OSE.
Studies involving OSE have been limited by the lackof an efficient experimental model. Normal OSE of therat, rabbit and human have been isolated and cultured(Adams et al., 1981; Piquette and Timms, 1990), but thesize and availability of tissue often limit the use of thesemodels. Bovine ovaries present a useful model for studyof OSE and ovarian stromal interactions. The bovineovary has essentially the same physiology and size asthe human ovary. The cow is a mono-ovulator and hasan ovarian cycle similar to that of the human. Bovineovarian cancer has been reported (Marchant, 1980)suggesting that bovine OSE has similar tumorigenicpotential as human OSE. Large numbers of freshbovine ovaries can be obtained and large numbers offresh OSE and stromal tissue can be isolated fromthem. Therefore, the bovine ovary provides a usefulmodel for examining the specific cell–cell interactionsinvolving normal OSE (Vigne et al., 1994).
Keratinocyte growth factor (KGF) is a 28-kilodalton(kDa) protein that is a member of the fibroblast growthfactor family (FGF-7) (Ledoux et al., 1992; Benharrochand Birnbaum, 1990). KGF is primarily produced bystromal- or mesenchymal-derived cells in many tissuesand acts as an epithelial cell-specific mitogen (Aaronsonet al., 1990; Rubin et al., 1995a,b). Before the currentstudy, KGF has not been previously shown to beexpressed by epithelial cells. The KGF receptor(KGFR) is a splice variant of the FGF receptor family(FGFR-2) that is primarily localized on epithelial cells(Rubin et al., 1995b; Bottaro et al., 1990; Werner et al.,1992; Miki et al., 1992). This KGFR mediates its signalthrough a dimerization event and phosphorylation cas-cade. KGF mediates mesenchymal–epithelial cell inter-actions in many tissues including the ovary (Parrott etal., 1994; Parrott and Skinner, 1998a,b), placenta(Izumi et al., 1996), endometrium (Koji et al., 1994),ventral prostate (Sugimura et al., 1996), and seminalvesicle (Alarid et al., 1994). Expression of KGF can beregulated by androgen, progesterone, and glucocorti-coids (Koji et al., 1994; Chedid et al., 1996; Fasciana etal., 1996). In the ovarian follicle, KGF mediates cell–
cell interactions between theca cells and granulosa cells(Parrott and Skinner, 1998a). Theca cell KGF expres-sion in the follicle can be regulated by estradiol andgonadotropins (Parrott and Skinner, 1998a). Thecacells and ovarian surface stromal cells are derived fromthe same ovarian interstitial cell population. Therefore,some similarities may exist between theca cell–granu-losa cell interactions and ovarian stromal–OSEinteractions.
The current study was designed to examine the localproduction and action of a specific stromal-epithelialcell factor, keratinocyte growth factor (KGF), in hu-man and bovine ovarian surface epithelium. The hy-pothesis examined in the current study is that KGF hasan important role in the normal biology of OSE, as wellas in ovarian cancer. Information regarding the expres-sion and action of KGF provides insight into thecell–cell interactions that regulate OSE and ovariantumor progression.
2. Materials and methods
2.1. Tissues
Fresh human adult ovarian tissues were obtainedfrom Dr Bethan Powell in the Department of Obstetricsand Gynecology at the University of California, SanFrancisco. The normal human tissues were collectedfrom salpingo-oophorectomy specimens removed forbenign diseases from women of child-bearing age. Hu-man cancer tissues were surgically collected fromwomen with borderline, stage I and stage III ovariancancer. Bovine ovaries were obtained from young non-pregnant cycling heifers less that 10 min after slaughter.The ovaries selected and used were in the follicularphase with no apparent corpus luteum as previouslydescribed (Parrott et al., 1994; Parrott and Skinner,1998a,b; Kruk et al., 1990). A minimum of three differ-ent tissue samples from different ovaries were obtainedfor analysis from both the normal ovaries and all stagesof tumors. When required, ovaries were fixed in 4%paraformaldehyde, embedded in paraffin, and cut into5 mm sections. Sections were stored at 4°C until im-munocytochemistry was performed.
2.2. Embedding, histology and immunocytochemistry forkeratinocyte growth factor (KGF)
Tissues were fixed in 4% paraformaldehyde and em-bedded in paraffin according to standard procedures(Parrott et al., 1994; Parrott and Skinner, 1998a,b;Kruk et al., 1990). Immunocytochemistry for KGF wasperformed according to standard procedures as previ-ously described (Parrott et al., 1994; Parrott and Skin-ner, 1998a,b; Kruk et al., 1990). Briefly, 5-mm sections

J.A. Parrott et al. / Molecular and Cellular Endocrinology 167 (2000) 77–87 79
were deparaffinized and rehydrated, quenched in 20%methanol/3% hydrogen peroxide, and blocked in 5%serum for several hours at room temperature beforeincubation with primary antibody, polyclonal rabbitanti-human KGF (Chemicon International Inc, Temec-ula, CA) at 1:500 dilution (approximately 10mg/ml)overnight at 4°C. Secondary antibody (biotinylatedgoat anti-rabbit IgG from Vector Laboratories,Burlingame, CA) was detected by using the Vectastainkit (Vector) and diaminobenzadine (DAB). Controlsincluded the use of non-immune serum and absence ofprimary antibody. Slides were counterstained lightlywith hematoxylin to visualize the tissue. KGF-positivecells stained brown to black.
2.3. OSE and stromal cell isolation and cell culture
OSE cells and ovarian stromal cells were isolatedfresh (for QRT-PCR studies) or cultured (for Northernand growth studies) (Vigne et al., 1994). OSE cells werescraped from the surface of the ovary with a rubberpoliceman as previously described (Kruk et al., 1990).Sheets of epithelial cells were suspended in Hanks’buffered salt solution and then pelleted and washedprior to suspension for plating. After the removal ofOSE cells, the ovarian surface stromal cells were mi-crodissected from the ovarian surface using areas of theovary devoid of follicles (Vigne et al., 1994). A sectionof surface stromal cells 1–2 mm wide by 5–8 mm longand 1-mm deep was collected. The tissue piece wasminced and digested with 1 mg/ml collagenase and 1mg/ml hyaluronidase for 2 h at 37°C or 18 h at 4°C.Cells were plated with an initial density of approxi-mately 106 cells/2 cm2, and were maintained at 37°C in5% CO2 atmosphere in Ham’s F-12 (Gibco Labs.,Grand Island, NY) supplemented with 10% calf serum.Medium was changed every 48–72 h. Once the cellshad grown to confluence they were trypsinized and splitinto appropriate plates. For isolation of RNA fromcultured cells, OSE and stromal cells were plated in 100mm large culture plates (Nunc) and maintained inHam’s F-12 supplemented with 10% calf serum. Forgrowth assays, OSE cells were plated in 24-well platesin DMEM (Gibco) supplemented with 10% calf serum.When cells achieved 50–70% confluency, cells werewashed in DMEM containing 0.1% calf serum forgrowth assays. The purity of OSE isolated by thisprocedure is greater than 98% by keratin staining withno detectable stromal contamination (Vigne et al.,1994; Kruk et al., 1990). The stromal cells isolated werealso greater than 98% as previously documented (Vigneet al., 1994). Human ovarian cancer cell lines, SKOV3and OCC1, were obtained from American Type CultureCollection (Rockville, MD).
2.4. RNA preparation
Total RNA was prepared from freshly isolated orcultured cells using Trizol reagent (Gibco) (Parrott etal., 1994; Parrott and Skinner, 1998a,b). Trizol wasadded directly to freshly isolated cells or to the cultureplate to prevent RNA degradation. Total RNA wasused to purify mRNA from with the FastTrack 2.0mRNA Isolation Kit (Invitrogen, Carlsbad, CA). RNAwas stored at −70°C until use.
2.5. Northern blot analysis
Total RNA and mRNA from OSE cells and ovarianstromal cells were isolated as described above. Approx-imately 6 mg of total RNA and 6 mg mRNA werefractionated on a 1.5% formaldehyde–agarose gel. Fol-lowing fractionation the RNA in the gel was trans-ferred to Nylon membrane (Hybond N+ , Amersham)in 1X MOPS buffer and UV cross linked. The mem-branes were then prehybridized (500 mM phosphatebuffer pH 7.2, 1 mM EDTA, 1% BSA, 7% SDS) for 2h at 60°C. The hybridization was carried out at 60°Covernight with 32P labeled KGF probe obtained byrandom primer extension (Prime-It II, Stratagene) of abovine KGF partial cDNA. The membrane waswashed in wash buffer (0.2× SSC, 0.1% SDS) at roomtemperature for 10 min and then at 60°C for 20 min.Membranes were exposed to X-ray film (X-OMAT,Kodak) overnight at −70°C using an intensifyingscreen. The membrane was subsequently stripped andrehybridized with bovine cyclophilin using a similarprocedure (Parrott and Skinner, 1998a,b).
2.6. Quantitati6e RT–PCR assays
Steady state levels of KGF and cyclophilin (i.e. 1B15)mRNA’s were analyzed using a specific quantitativereverse transcription-polymerase chain reaction (RT-PCR) assay for each gene. These quantitative RT-PCRassays have been described in detail (Parrott and Skin-ner, 1998a). Primers were selected that spanned anintron. The primers used in this quantitative analysis ofKGF and 1B15 were: KGF, 5%-ATA CTG ACA TGGATC CTG CCA AGT TTG CTC TAC AGA TCATGC TTC-3% (5% primer, 45-mer) and 5%-TCC AACTGC CAC GGT CCT GAT-3% (3% primer, 21-mer)which generated a specific 306-bp KGF PCR product;and 1B15, 5%-ACA CGC CAT AAT GGC ACT GGTGGC AAG TCC ATC-3% (5% primer, 33-mer) and 5%-ATT TGC CAT GGA CAA GAT GCC AGG ACCTGT ATG-3% (3% primer, 33-mer) which generated aspecific 105-bp product from all cell types demonstrat-ing the integrity of the RNA samples. Total RNA (1mg) was reverse transcribed for 1 h at 37°C using thefollowing conditions: 1 mg total RNA, 1 mM specific

J.A. Parrott et al. / Molecular and Cellular Endocrinology 167 (2000) 77–8780
3%-primers of interest (up to four different primersincluding 1B15), 0.1 mM dNTP’s, 10 mM DTT, 40units RNAse inhibitor (Promega, Madison, WI), and200 units M-MLV reverse transcriptase (Gibco BRL,Gaithersburg, MD) in 40 ml RT buffer (50 mM Tris–HCl pH 8.3, 75 mM KCl, 3 mM MgCl2). After 1 hsamples were heated to 95°C for 5 min to inactivate thereverse transcriptase enzyme. Immediately before am-plification each unknown sample was further diluted1:10 in order to improve the fidelity of the PCR reac-tion. Plasmid DNA’s (i.e. Bluescript) containing bovineKGF or 1B15 subclones were used to generate standardcurves from 1 attogram/ml (10−15 g/ml) to 10 pg/ml(10×10−9 g/ml) each containing 10 ng/ml Bluescriptcarrier DNA. Identical 10 ml aliquots of each sampleand standard were pipetted in duplicate into a 96 wellreaction plate (Marsh Biomedical Products, Rochester,NY) and sealed with adhesive film (Marsh BiomedicalProducts) for PCR amplification. Amplification wasperformed in a Perkin Elmer 9600 equipped with aheated lid using the following conditions: 0.4 mM eachprimer, 16 mM dNTP’s, and 1.25 units AmpliTaq poly-merase in 50 ml GeneAmp PCR buffer (containing 1.5mM MgCl2, Perkin Elmer). Each PCR amplificationconsisted of an initial denaturing reaction (5 min,95°C); 25–31 cycles of denaturing (30 s, 95°C), anneal-ing (1 min, 60°C), and elongation (2 min, 72°C) reac-tions; and a final elongation reaction (10 min, 72°C). Atleast 0.25 mCi of 32P-labeled dCTP (Redivue, Amer-sham Life Sciences, Arlington Heights, IL) was in-cluded in each sample during amplification fordetection purposes. Specific PCR products were quanti-tated by electrophoresing all samples on 4–5% poly-acrylamide gels, simultaneously exposing the gels to aphosphor screen for 8–24 h, followed by quantitatingthe specific bands on a Storm PhosphorImager (Molec-ular Dynamics, Sunnyvale, CA). Equivalent steadystate mRNA levels for each gene were determined bycomparing each sample to the appropriate standardcurve. All KGF data were normalized for 1B15. Aminimum of three different experiments each with dif-ferent tissue samples or cell preparations were analyzedand data combined.
Optimal cycle number for amplification was deter-mined for each assay in order to achieve maximumsensitivity while maintaining linearity (i.e. logarithmicphase of PCR reactions). KGF quantitative PCR prod-ucts were amplified for 28 cycles and 1B15 PCR prod-ucts were amplified for 25 cycles. The sensitivity of eachquantitative PCR assay was below 1 fg which corre-sponds to less than 125 fg target mRNA/mg total RNA.For each assay, all samples were simultaneously mea-sured in duplicate resulting in intra-assay variabilities of11.3% (KGF) and 6.5% (1B15).
2.7. Growth assays
Cell growth was analyzed by quantifying (3H)thymidine incorporation into newly synthesized DNA.OSE cells were plated (approximately 1 million cells/cm2 providing 50% confluence) in 0.5 ml DMEMmedium containing 0.1% calf serum. After 48 h, cellswere treated with either no growth factor (control), 50ng/ml keratinocyte growth factor (KGF), or 40 ng/mlepidermal growth factor (EGF). These concentrationshave been shown to be optimal as assessed with doseresponse curves (Parrott et al., 1994; Parrott and Skin-ner, 1998a,b). Cells were plated for 48 h and thentreated for 20 h. After treatment, 0.5 ml DMEM con-taining 2.5 mCi (3H) thymidine was added to each well,and the cells were incubated for 4 h at 37°C and thensonicated. The quantity of (3H) thymidine incorporatedinto DNA was determined, as previously described(Vigne et al., 1994). Data were normalized to totalDNA per well using an ethidium bromide procedurepreviously described (Parrott et al., 1994; Parrott andSkinner, 1998a,b; Vigne et al., 1994).
2.8. DNA assays
DNA was measured fluorometrically with ethidiumbromide as previously described (Vigne et al., 1994). Analiquot of the sonicated cell suspension was added to anequal volume of ethidium bromide solution (0.25 mMethidium bromide 100 U/ml heparin in ethidium bro-mide buffer (EBB: 20 mM sodium chloride, 5 mMethylene diamine tetracetate, 10 mM Tris, pH 7.8;Sigma), was diluted 1:2 with EBB, and was allowed toincubate at room temperature for 30 min. Fluorescentemission at 585 nm with 350 nm excitation was thenmonitored. A standard curve with calf thymus DNAwas used to quantify DNA levels in the culture wells.This assay has a sensitivity of approximately 0.1 mgDNA.
2.9. Statistical analysis
All data were analyzed by a JMP 3.1 statisticalanalysis program (SAS Institute Inc., Cary, NC). Ef-fects of growth factors on (3H)thymidine incorporationinto DNA and differences among fresh versus culturedOSE and stromal cell KGF expression were analyzedby a one-way analysis of variance (ANOVA). Observedsignificance probabilities of 0.05 (P\F) or less wereconsidered evidence that an ANOVA model fits thedata. Significant differences between treated cells andcontrol (untreated) cells were determined using theDunnett’s test which guards against the high alpha-size(Type I) error rate across the hypothesis (Dunnett,1955). Significant differences among fresh versus cul-tured OSE and stromal cell KGF expression was deter-

J.A. Parrott et al. / Molecular and Cellular Endocrinology 167 (2000) 77–87 81
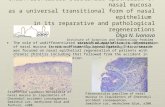
Fig. 1. KGF protein expression in human ovarian tumors. Human ovarian tumors were surgically removed and immediately fixed in 4%paraformaldehyde followed by paraffin embedding. Ovaries were obtained from patients diagnosed with borderline, stage I and stage III ovariancancer. Sections were cut at 5 mm thickness and stored on glass slides at 4°C. Immunocytochemistry was performed using a KGF antibody asdescribed in Section 2. Sections were visualized and photographed at approximately 400× magnification. Similar results were obtained in fourseparate experiments using four different ovarian tumors. (A) Borderline ovarian cancer; (B) Stage I ovarian cancer; (C and D), Stage III ovariancancers; (E) control section stained with non-immune IgG.
J.A. Parrott et al. / Molecular and Cellular Endocrinology 167 (2000) 77–8782
mined using the Tukey–Kramer HSD (honestly signifi-cant difference) test which protects the significance testsof all combinations of pairs (Tukey, 1953, 1991;Kramer, 1956). These tests are recommended for multi-ple comparisons (Hsu, 1996). A minimum of threedifferent experiments were performed in replicate ondifferent samples and tissue preparations.
3. Results
Expression of KGF protein was examined at differ-ent stages of ovarian cancer by immunocytochemistry(ICC). Tissues were examined from cases of borderline,stage I and stage III ovarian cancers. KGF protein wasdetected in the epithelial cell component of the diseasedtissue at all stages examined (Fig. 1). Light stainingcould also be detected in the stromal cells that borderthe epithelial cells, but the most intense staining was in
the epithelium. Control slides were analyzed using non-immune IgG and showed no staining (Fig. 1E). Resultsdemonstrate that the expression of KGF protein ispresent during different stages of ovarian cancer inhumans. Experiments were next designed to character-ize KGF expression in normal OSE and ovarian stro-mal cells.
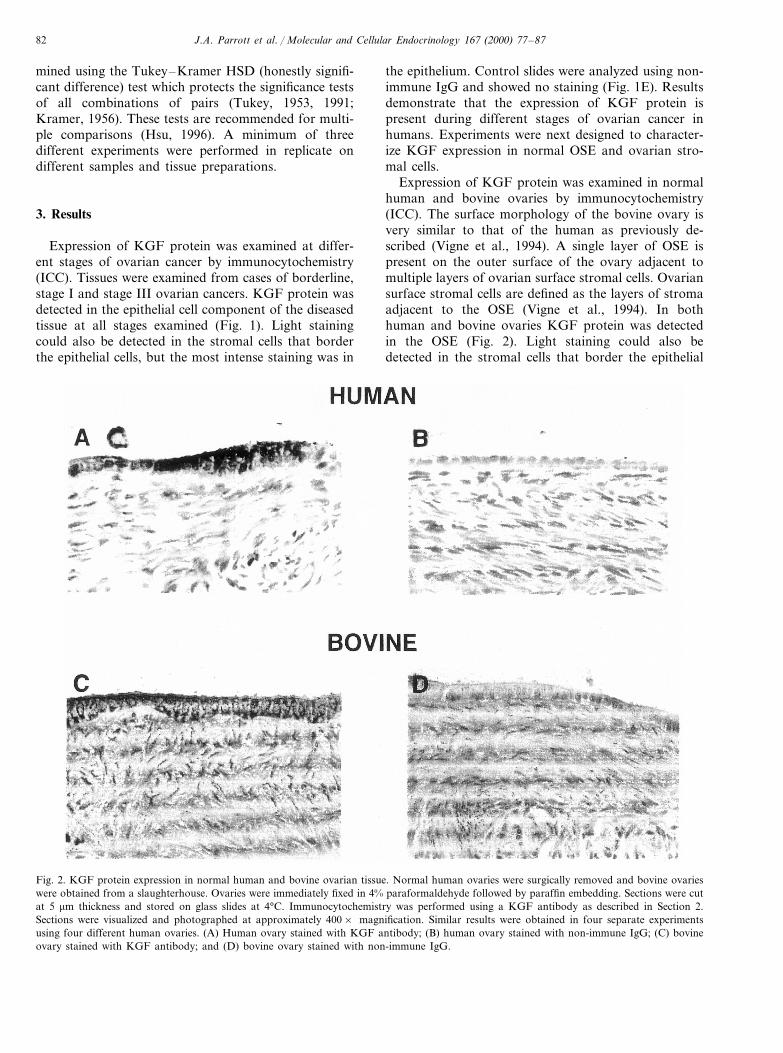
Expression of KGF protein was examined in normalhuman and bovine ovaries by immunocytochemistry(ICC). The surface morphology of the bovine ovary isvery similar to that of the human as previously de-scribed (Vigne et al., 1994). A single layer of OSE ispresent on the outer surface of the ovary adjacent tomultiple layers of ovarian surface stromal cells. Ovariansurface stromal cells are defined as the layers of stromaadjacent to the OSE (Vigne et al., 1994). In bothhuman and bovine ovaries KGF protein was detectedin the OSE (Fig. 2). Light staining could also bedetected in the stromal cells that border the epithelial
Fig. 2. KGF protein expression in normal human and bovine ovarian tissue. Normal human ovaries were surgically removed and bovine ovarieswere obtained from a slaughterhouse. Ovaries were immediately fixed in 4% paraformaldehyde followed by paraffin embedding. Sections were cutat 5 mm thickness and stored on glass slides at 4°C. Immunocytochemistry was performed using a KGF antibody as described in Section 2.Sections were visualized and photographed at approximately 400× magnification. Similar results were obtained in four separate experimentsusing four different human ovaries. (A) Human ovary stained with KGF antibody; (B) human ovary stained with non-immune IgG; (C) bovineovary stained with KGF antibody; and (D) bovine ovary stained with non-immune IgG.
J.A. Parrott et al. / Molecular and Cellular Endocrinology 167 (2000) 77–87 83
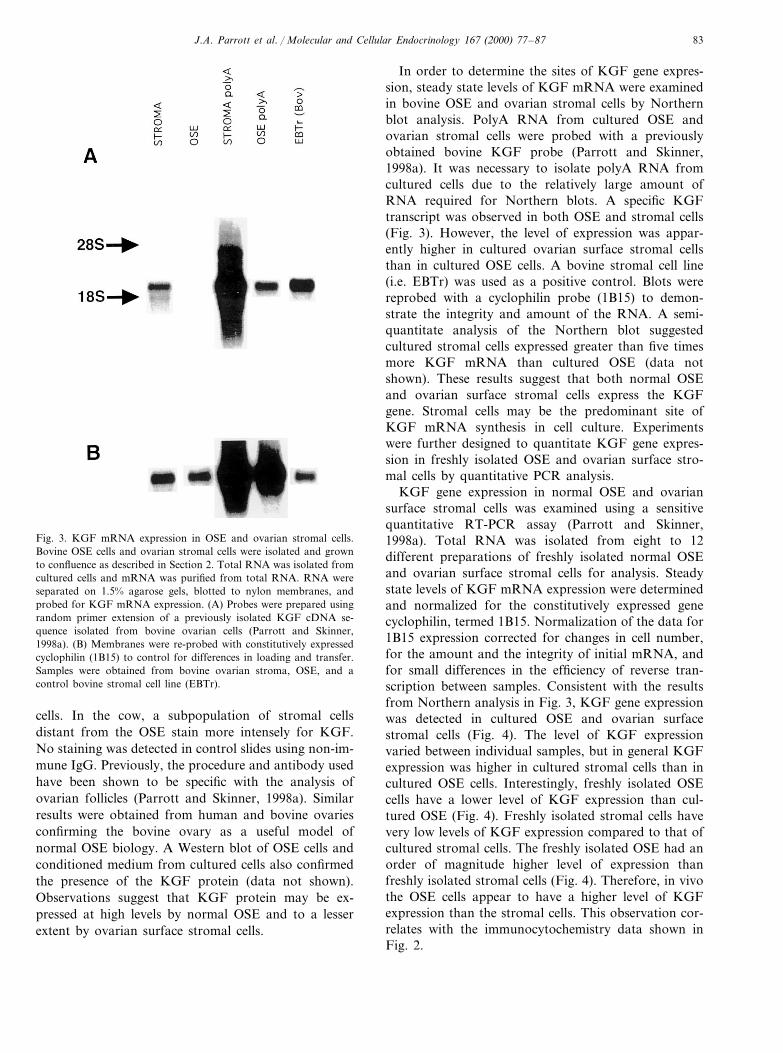
Fig. 3. KGF mRNA expression in OSE and ovarian stromal cells.Bovine OSE cells and ovarian stromal cells were isolated and grownto confluence as described in Section 2. Total RNA was isolated fromcultured cells and mRNA was purified from total RNA. RNA wereseparated on 1.5% agarose gels, blotted to nylon membranes, andprobed for KGF mRNA expression. (A) Probes were prepared usingrandom primer extension of a previously isolated KGF cDNA se-quence isolated from bovine ovarian cells (Parrott and Skinner,1998a). (B) Membranes were re-probed with constitutively expressedcyclophilin (1B15) to control for differences in loading and transfer.Samples were obtained from bovine ovarian stroma, OSE, and acontrol bovine stromal cell line (EBTr).
In order to determine the sites of KGF gene expres-sion, steady state levels of KGF mRNA were examinedin bovine OSE and ovarian stromal cells by Northernblot analysis. PolyA RNA from cultured OSE andovarian stromal cells were probed with a previouslyobtained bovine KGF probe (Parrott and Skinner,1998a). It was necessary to isolate polyA RNA fromcultured cells due to the relatively large amount ofRNA required for Northern blots. A specific KGFtranscript was observed in both OSE and stromal cells(Fig. 3). However, the level of expression was appar-ently higher in cultured ovarian surface stromal cellsthan in cultured OSE cells. A bovine stromal cell line(i.e. EBTr) was used as a positive control. Blots werereprobed with a cyclophilin probe (1B15) to demon-strate the integrity and amount of the RNA. A semi-quantitate analysis of the Northern blot suggestedcultured stromal cells expressed greater than five timesmore KGF mRNA than cultured OSE (data notshown). These results suggest that both normal OSEand ovarian surface stromal cells express the KGFgene. Stromal cells may be the predominant site ofKGF mRNA synthesis in cell culture. Experimentswere further designed to quantitate KGF gene expres-sion in freshly isolated OSE and ovarian surface stro-mal cells by quantitative PCR analysis.
KGF gene expression in normal OSE and ovariansurface stromal cells was examined using a sensitivequantitative RT-PCR assay (Parrott and Skinner,1998a). Total RNA was isolated from eight to 12different preparations of freshly isolated normal OSEand ovarian surface stromal cells for analysis. Steadystate levels of KGF mRNA expression were determinedand normalized for the constitutively expressed genecyclophilin, termed 1B15. Normalization of the data for1B15 expression corrected for changes in cell number,for the amount and the integrity of initial mRNA, andfor small differences in the efficiency of reverse tran-scription between samples. Consistent with the resultsfrom Northern analysis in Fig. 3, KGF gene expressionwas detected in cultured OSE and ovarian surfacestromal cells (Fig. 4). The level of KGF expressionvaried between individual samples, but in general KGFexpression was higher in cultured stromal cells than incultured OSE cells. Interestingly, freshly isolated OSEcells have a lower level of KGF expression than cul-tured OSE (Fig. 4). Freshly isolated stromal cells havevery low levels of KGF expression compared to that ofcultured stromal cells. The freshly isolated OSE had anorder of magnitude higher level of expression thanfreshly isolated stromal cells (Fig. 4). Therefore, in vivothe OSE cells appear to have a higher level of KGFexpression than the stromal cells. This observation cor-relates with the immunocytochemistry data shown inFig. 2.
cells. In the cow, a subpopulation of stromal cellsdistant from the OSE stain more intensely for KGF.No staining was detected in control slides using non-im-mune IgG. Previously, the procedure and antibody usedhave been shown to be specific with the analysis ofovarian follicles (Parrott and Skinner, 1998a). Similarresults were obtained from human and bovine ovariesconfirming the bovine ovary as a useful model ofnormal OSE biology. A Western blot of OSE cells andconditioned medium from cultured cells also confirmedthe presence of the KGF protein (data not shown).Observations suggest that KGF protein may be ex-pressed at high levels by normal OSE and to a lesserextent by ovarian surface stromal cells.
J.A. Parrott et al. / Molecular and Cellular Endocrinology 167 (2000) 77–8784
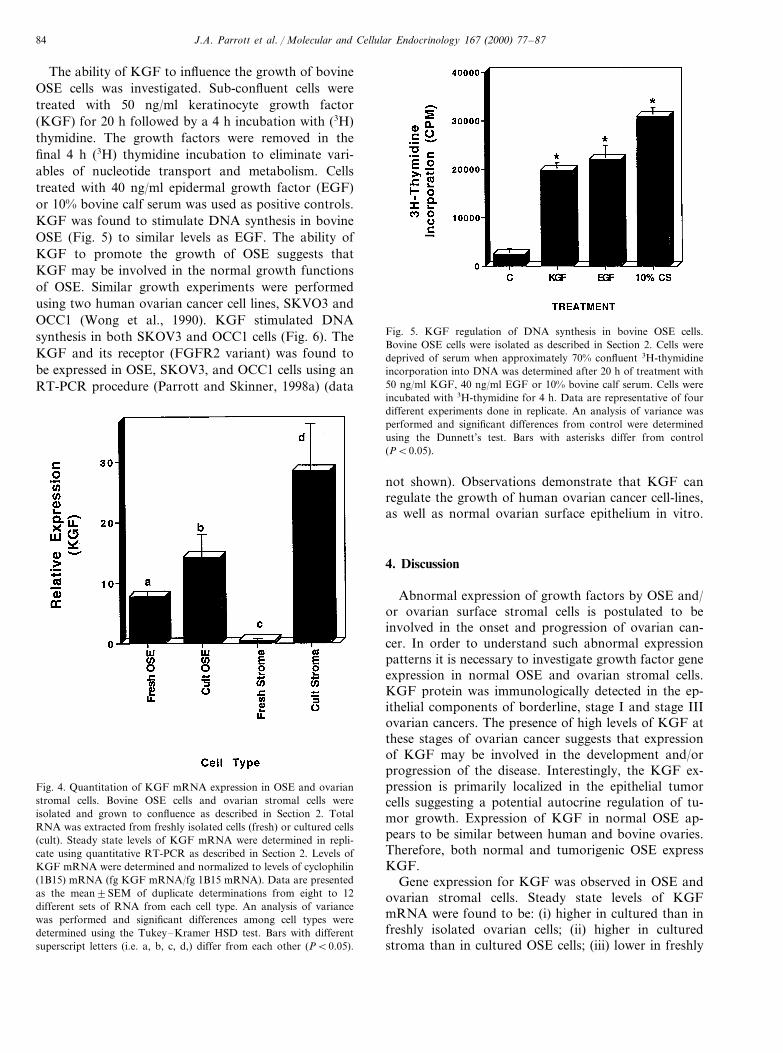
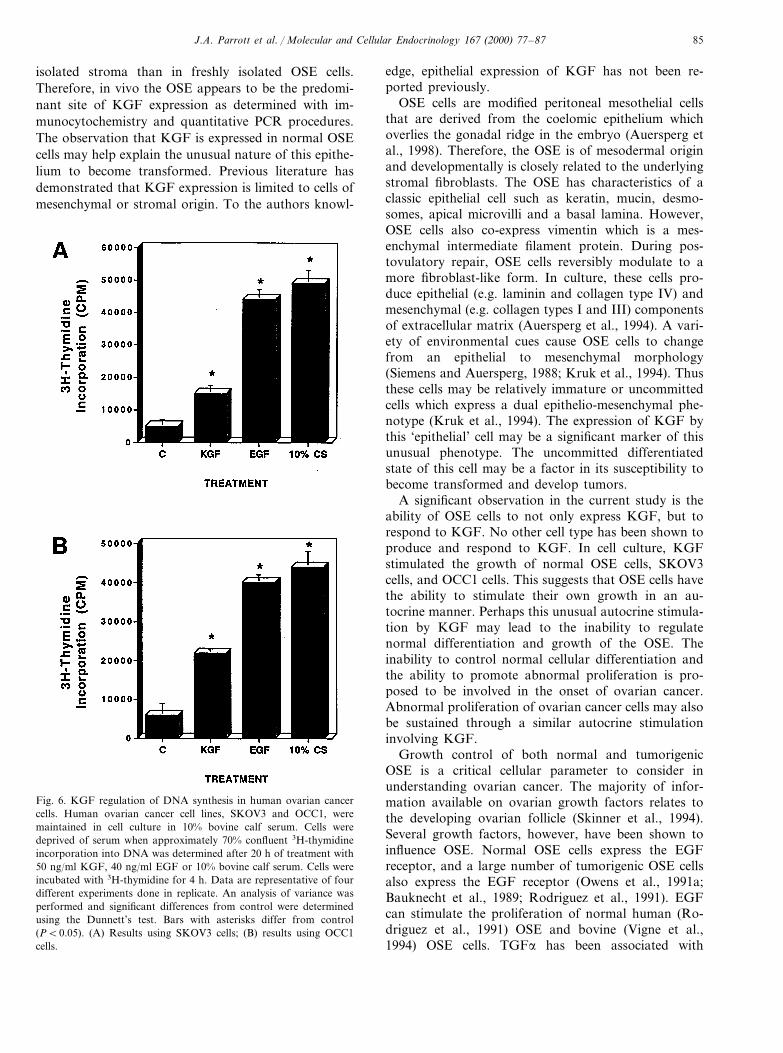
The ability of KGF to influence the growth of bovineOSE cells was investigated. Sub-confluent cells weretreated with 50 ng/ml keratinocyte growth factor(KGF) for 20 h followed by a 4 h incubation with (3H)thymidine. The growth factors were removed in thefinal 4 h (3H) thymidine incubation to eliminate vari-ables of nucleotide transport and metabolism. Cellstreated with 40 ng/ml epidermal growth factor (EGF)or 10% bovine calf serum was used as positive controls.KGF was found to stimulate DNA synthesis in bovineOSE (Fig. 5) to similar levels as EGF. The ability ofKGF to promote the growth of OSE suggests thatKGF may be involved in the normal growth functionsof OSE. Similar growth experiments were performedusing two human ovarian cancer cell lines, SKVO3 andOCC1 (Wong et al., 1990). KGF stimulated DNAsynthesis in both SKOV3 and OCC1 cells (Fig. 6). TheKGF and its receptor (FGFR2 variant) was found tobe expressed in OSE, SKOV3, and OCC1 cells using anRT-PCR procedure (Parrott and Skinner, 1998a) (data
Fig. 5. KGF regulation of DNA synthesis in bovine OSE cells.Bovine OSE cells were isolated as described in Section 2. Cells weredeprived of serum when approximately 70% confluent 3H-thymidineincorporation into DNA was determined after 20 h of treatment with50 ng/ml KGF, 40 ng/ml EGF or 10% bovine calf serum. Cells wereincubated with 3H-thymidine for 4 h. Data are representative of fourdifferent experiments done in replicate. An analysis of variance wasperformed and significant differences from control were determinedusing the Dunnett’s test. Bars with asterisks differ from control(PB0.05).
Fig. 4. Quantitation of KGF mRNA expression in OSE and ovarianstromal cells. Bovine OSE cells and ovarian stromal cells wereisolated and grown to confluence as described in Section 2. TotalRNA was extracted from freshly isolated cells (fresh) or cultured cells(cult). Steady state levels of KGF mRNA were determined in repli-cate using quantitative RT-PCR as described in Section 2. Levels ofKGF mRNA were determined and normalized to levels of cyclophilin(1B15) mRNA (fg KGF mRNA/fg 1B15 mRNA). Data are presentedas the mean9SEM of duplicate determinations from eight to 12different sets of RNA from each cell type. An analysis of variancewas performed and significant differences among cell types weredetermined using the Tukey–Kramer HSD test. Bars with differentsuperscript letters (i.e. a, b, c, d,) differ from each other (PB0.05).
not shown). Observations demonstrate that KGF canregulate the growth of human ovarian cancer cell-lines,as well as normal ovarian surface epithelium in vitro.
4. Discussion
Abnormal expression of growth factors by OSE and/or ovarian surface stromal cells is postulated to beinvolved in the onset and progression of ovarian can-cer. In order to understand such abnormal expressionpatterns it is necessary to investigate growth factor geneexpression in normal OSE and ovarian stromal cells.KGF protein was immunologically detected in the ep-ithelial components of borderline, stage I and stage IIIovarian cancers. The presence of high levels of KGF atthese stages of ovarian cancer suggests that expressionof KGF may be involved in the development and/orprogression of the disease. Interestingly, the KGF ex-pression is primarily localized in the epithelial tumorcells suggesting a potential autocrine regulation of tu-mor growth. Expression of KGF in normal OSE ap-pears to be similar between human and bovine ovaries.Therefore, both normal and tumorigenic OSE expressKGF.
Gene expression for KGF was observed in OSE andovarian stromal cells. Steady state levels of KGFmRNA were found to be: (i) higher in cultured than infreshly isolated ovarian cells; (ii) higher in culturedstroma than in cultured OSE cells; (iii) lower in freshly
J.A. Parrott et al. / Molecular and Cellular Endocrinology 167 (2000) 77–87 85
isolated stroma than in freshly isolated OSE cells.Therefore, in vivo the OSE appears to be the predomi-nant site of KGF expression as determined with im-munocytochemistry and quantitative PCR procedures.The observation that KGF is expressed in normal OSEcells may help explain the unusual nature of this epithe-lium to become transformed. Previous literature hasdemonstrated that KGF expression is limited to cells ofmesenchymal or stromal origin. To the authors knowl-
edge, epithelial expression of KGF has not been re-ported previously.
OSE cells are modified peritoneal mesothelial cellsthat are derived from the coelomic epithelium whichoverlies the gonadal ridge in the embryo (Auersperg etal., 1998). Therefore, the OSE is of mesodermal originand developmentally is closely related to the underlyingstromal fibroblasts. The OSE has characteristics of aclassic epithelial cell such as keratin, mucin, desmo-somes, apical microvilli and a basal lamina. However,OSE cells also co-express vimentin which is a mes-enchymal intermediate filament protein. During pos-tovulatory repair, OSE cells reversibly modulate to amore fibroblast-like form. In culture, these cells pro-duce epithelial (e.g. laminin and collagen type IV) andmesenchymal (e.g. collagen types I and III) componentsof extracellular matrix (Auersperg et al., 1994). A vari-ety of environmental cues cause OSE cells to changefrom an epithelial to mesenchymal morphology(Siemens and Auersperg, 1988; Kruk et al., 1994). Thusthese cells may be relatively immature or uncommittedcells which express a dual epithelio-mesenchymal phe-notype (Kruk et al., 1994). The expression of KGF bythis ‘epithelial’ cell may be a significant marker of thisunusual phenotype. The uncommitted differentiatedstate of this cell may be a factor in its susceptibility tobecome transformed and develop tumors.
A significant observation in the current study is theability of OSE cells to not only express KGF, but torespond to KGF. No other cell type has been shown toproduce and respond to KGF. In cell culture, KGFstimulated the growth of normal OSE cells, SKOV3cells, and OCC1 cells. This suggests that OSE cells havethe ability to stimulate their own growth in an au-tocrine manner. Perhaps this unusual autocrine stimula-tion by KGF may lead to the inability to regulatenormal differentiation and growth of the OSE. Theinability to control normal cellular differentiation andthe ability to promote abnormal proliferation is pro-posed to be involved in the onset of ovarian cancer.Abnormal proliferation of ovarian cancer cells may alsobe sustained through a similar autocrine stimulationinvolving KGF.
Growth control of both normal and tumorigenicOSE is a critical cellular parameter to consider inunderstanding ovarian cancer. The majority of infor-mation available on ovarian growth factors relates tothe developing ovarian follicle (Skinner et al., 1994).Several growth factors, however, have been shown toinfluence OSE. Normal OSE cells express the EGFreceptor, and a large number of tumorigenic OSE cellsalso express the EGF receptor (Owens et al., 1991a;Bauknecht et al., 1989; Rodriguez et al., 1991). EGFcan stimulate the proliferation of normal human (Ro-driguez et al., 1991) OSE and bovine (Vigne et al.,1994) OSE cells. TGFa has been associated with
Fig. 6. KGF regulation of DNA synthesis in human ovarian cancercells. Human ovarian cancer cell lines, SKOV3 and OCC1, weremaintained in cell culture in 10% bovine calf serum. Cells weredeprived of serum when approximately 70% confluent 3H-thymidineincorporation into DNA was determined after 20 h of treatment with50 ng/ml KGF, 40 ng/ml EGF or 10% bovine calf serum. Cells wereincubated with 3H-thymidine for 4 h. Data are representative of fourdifferent experiments done in replicate. An analysis of variance wasperformed and significant differences from control were determinedusing the Dunnett’s test. Bars with asterisks differ from control(PB0.05). (A) Results using SKOV3 cells; (B) results using OCC1cells.

J.A. Parrott et al. / Molecular and Cellular Endocrinology 167 (2000) 77–8786
ovarian cancer (Owens et al., 1991b; Yeh and Yeh,1989), and may act as an autocrine growth factor toinduce cell proliferation in both normal and tumori-genic OSE (Stromberg et al., 1992; Bast et al., 1992).Basic fibroblast growth factor (bFGF) and its receptorare expressed by human ovarian epithelial neoplasms(Di Blasio et al., 1993, 1995) suggesting that bFGF mayalso regulate ovarian cancer proliferation through anautocrine mechanism. Several ovarian cancer cell linesproliferate in response to bFGF (Berchuck et al., 1990).TGFb is a multifunctional protein that has a major rolein inhibiting the actions of growth stimulators such asEGF/TGFa, bFGF, and KGF. TGFb has been shownto be produced by OSE (Berchuck et al., 1992), andTGFb can inhibit the growth of normal OSE cells andsome tumorigenic OSE cells (Berchuck et al., 1990,1992; Grunt et al., 1992). It is postulated that a numberof different growth factors, including KGF, will beinvolved in the normal and abnormal growth control ofOSE.
In the current study, the role that KGF may have inmediating cell-cell interactions involving OSE was in-vestigated. Human and bovine OSE cells and ovariancancers were shown to express KGF mRNA andprotein. These observations justify future studies de-signed to determine the mechanistic role of KGF innormal OSE biology and the etiology of OSE-derivedcancers. Further analysis of the actions of KGF inovarian cancer requires considering the combined ef-fects with other growth factors. KGF is primarily astromal cell-derived growth factor in other tissues. Ex-pression of KGF by OSE cells may indicate that thisepithelial cell is normally in an altered differentiationstate that is susceptible to transformation.
Acknowledgements
We thank Gene Herrington for technical assistance.We also thank Susan Cobb and Jaideep Chaudhary forassistance in preparation of the manuscript. This workwas supported by an Ovarian Cancer Program Projectgrant from the National Institutes of Health (NIH).
References
Aaronson, S.A., Rubin, J.S., Finch, P.W., et al., 1990. Growthfactor-regulated pathways in epithelial cell proliferation. Am.Rev. Respir. Dis. 142, S7–S10.
Adams, A.T., Auersperg, N., 1981. Transformation of cultured ratovarian surface epithelial cells by Kirsten murine sarcoma virus.Cancer Res. 41, 2063–2072.
Alarid, E.T., Rubin, J.S., Young, P., et al., 1994. Keratinocytegrowth factor functions in epithelial induction during seminalvesicle development. Proc. Natl. Acad. Sci. USA 91, 1074–1078.
Auersperg, N., Maclaren, I.A., Kruk, P.A., 1991. Ovarian surfaceepithelium: autonomous production of connective tissue-type ex-tracellular matrix. Biol. Reprod. 44, 717–724.
Auersperg, N., Maines-Bandiera, S.L., Dyck, H.G., Kruk, A., 1994.Characterization of cultured human ovarian surface epithelialcells: phenotypic plasticity and premalignant changes. Lab. Invest.71, 510–518.
Auersperg, N., Edelson, M.I., Mok, S.C., Johnson, S.W., Hamilton,T.C., 1998. The biology of ovarian cancer. Semin. Oncol. 25,281–304.
Bast, R.C. Jr., Jacobs, I., Berchuck, A., 1992. Malignant transforma-tion of ovarian epithelium. J. Natl. Cancer Inst. 84, 556–558.
Bauknecht, T., Kohler, M., Janz, I., Pfleiderer, A., 1989. The occur-rence of epidermal growth factor receptors and the characteriza-tion of EGF-like factors in human ovarian, endometrial, cervicaland breast cancer. EGF receptors and factors in gynecologicalcarcinomas. J. Cancer Res. Clin. Oncol. 115, 193–199.
Benharroch, D., Birnbaum, D., 1990. Biology of the fibroblastgrowth factor gene family. Isr. J. Med. Sci. 26, 212–219.
Berchuck, A., Olt, G.J., Everitt, L., Soisson, A.P., Bast, R.C. Jr.,Boyer, C.M., 1990. The role of peptide growth factors in epithelialovarian cancer. Obstet. Gynecol. 75, 255–262.
Berchuck, A., Rodriguez, G., Olt, G., Whitaker, R., Boente, M.P.,Arrick, B.A., Clarke-Pearson, D.L., Bast, R.C. Jr., 1992. Regula-tion of growth of normal ovarian epithelial cells and ovariancancer cell lines by transforming growth factor-beta. Am. J.Obstet. Gynecol. 166, 676–684.
Bjersing, L., Cajander, S., 1975. Ovulation and the role of the ovariansurface epithelium. Experientia 31, 605–608.
Bottaro, D.P., Rubin, J.S., Ron, D., Finch, P.W., Florio, C.,Aaronson, S.A., 1990. Characterization of the receptor for kerati-nocyte growth factor. Evidence for multiple fibroblast growthfactor receptors. J. Biol. Chem. 265, 12767–12770.
ACS Cancer Facts and Figures, 1998. American Cancer Society,Atlanta, GA.
Chedid, M., Hoyle, J.R., Csaky, K.G., Rubin, J.S., 1996. Glucocorti-coids inhibit keratinocyte growth factor production in primarydermal fibroblasts. Endocrinology 137, 2232–2237.
Di Blasio, A.M., Cremonesi, L., Vigano, P., Ferrari, M., Gospo-darowicz, D., Vignali, M., Jaffe, R.B., 1993. Basic fibroblastgrowth factor and its receptor messenger ribonucleic acids areexpressed in human ovarian epithelial neoplasms. Am. J. Obstet.Gynecol. 169, 1517–1523.
Di Blasio, A.M., Carniti, C., Vigano, P., Vignali, M., 1995. Basicfibroblast growth factor and ovarian cancer. J. Steroid Biochem.Mol. Biol. 53, 375–379.
Dunnett, C.W., 1955. A multiple comparison procedure for compar-ing several treatments with control. J. Am. Stat. Assoc. 50,1096–1121.
Fasciana, C., van der Made, A.C., Faber, P.W., Trapman, J., 1996.Androgen regulation of the rat keratinocyte growth factor (KGF/FGF7) promoter. Biochem. Biophys. Res. Commun. 220, 858–863.
Godwin, A.K., Testa, J.R., Handel, L.M., et al., 1992. Spontaneoustransformation of rat ovarian surface epithelial cells: associationwith cytogenetic changes and implications of repeated ovulationin the etiology of ovarian cancer. J. Natl. Cancer Inst. 84,592–601.
Grunt, T.W., Somay, C., Oeller, H., Dittrich, E., Dittrich, C., 1992.Comparative analysis of the effects of dimethyl sulfoxide andretinoic acid on the antigenic pattern of human ovarian adenocar-cinoma cells. J. Cell Sci. 103, 501–509.
Hornby, A.E., Pan, J., Auersperg, N., 1992. Intermediate filaments inrat ovarian surface epithelial cells: changes with neoplastic pro-gression in culture. Biochem. Cell Biol. 70, 16–25.
Hsu, J.C., 1996. Multiple Comparisons: Theory and Methods. Chap-man & Hall, New York, pp. 1–277.

J.A. Parrott et al. / Molecular and Cellular Endocrinology 167 (2000) 77–87 87
Izumi, S., Slayden, O.D., Rubin, J.S., Brenner, R.M., 1996. Kerati-nocyte growth factor and its receptor in the rhesus macaqueplacenta during the course of gestation. Placenta 17, 123–135.
Koji, T., Chedid, M., Rubin, J.S., et al., 1994. Progesterone-depen-dent expression of keratinocyte growth factor mRNA in stromalcells of the primate endometrium: keratinocyte growth factor as aprogestomedin. J. Cell Biol. 125, 393–401.
Kramer, C.Y., 1956. Extension of multiple range tests to groupmeans with unequal numbers of replications. Biometrics 12, 309–310.
Kruk, P.A., Maines–Bandiera, S.L., Auersperg, N.A., 1990. Sim-plified method to culture human ovarian surface epithelium. Lab.Invest. 63, 132–136.
Kruk, P.A., Uitto, V.J., Firth, J.D., Dedhar, S., Auersperg, N., 1994.Reciprocal interactions between human ovarian surface epithelialcells and adjacent extracellular matrix. Exp. Cell Res. 215, 97–108.
Kurman, R.K., 1987. Bleustein’s Pathology of the Female GenitalTract, 3rd edn. Springer-Verlag, New York, pp. 571–597.
Ledoux, D., Gannoun-Zaki, L., Barritault, D., 1992. Interactions ofFGFs with target cells. Prog. Growth Factor Res. 4, 107–120.
Marchant, J., 1980. Animal models for tumors of the ovary. In:Murphy, E.D., Beamer, W.G. (Eds.), Biology of Ovarian Neo-plasia, vol. 50. UICC Tech Rep, pp. 50–65.
Miki, T., Bottaro, D.P., Fleming, T.P., et al., 1992. Determination ofligand-binding specificity by alternative splicing: two distinctgrowth factor receptors encoded by a single gene. Proc. Natl.Acad. Sci. USA 89, 246–250.
Nicosia, S.V., Nicosia, R.F., 1988. Pathology of Human Neoplasms.Raven Press, New York, pp. 435–486.
Osterholze, H.O., Johnson, J.H., Nicosia, S.V., 1985. An autoradio-graphic study of rabbit ovarian surface epithelium before andafter ovulation. Biol. Reprod. 33, 729–738.
Owens, O.J., Stewart, C., Brown, I., Leake, R.E., 1991a. Epidermalgrowth factor receptors (EGFR) in human ovarian cancer. Br. J.Cancer 64, 907–910.
Owens, O.J., Stewart, C., Leake, R.E., 1991b. Growth factors inovarian cancer [published erratum appears in Br J Cancer 1992,65 (4), 361]. Br J Cancer 64, 1177–1181
Parrott, J.A., Skinner, M.K., 1998a. Developmental and hormonalregulation of keratinocyte growth factor expression and action inthe ovarian follicle. Endocrinology 139, 228–235.
Parrott, J.A., Skinner, M.K., 1998b. Thecal cell-granulosa cell inter-actions involve a positive feedback loop among keratinocytegrowth factor, hepatocyte growth factor, and Kit ligand duringovarian follicular development. Endocrinology 139, 2240–2245.
Parrott, J.A., Vigne, J.L., Chu, B.Z., Skinner, M.K., 1994. Mesenchy-mal-epithelial interactions in the ovarian follicle involve kerati-nocyte and hepatocyte growth factor production by thecal cellsand their action on granulosa cells. Endocrinology 135, 569–575.
Piquette, G.N., Timms, B.G., 1990. Isolation and characterization ofrabbit ovarian surface epithelium, granulosa cells, and peritonealmesothelium in primary culture. In Vitro Cell Dev. Biol. 26,471–481.
Piver, M.S., Baker, T.R., Piedmonte, M., Sandecki, A.M., 1991.Epidemiology and etiology of ovarian cancer. Semin. Oncol. 18,177–185.
Rodriguez, G.C., Berchuck, A., Whitaker, R.S., Schlossman, D.,Clarke-Pearson, D.L., Bast, R.C. Jr., 1991. Epidermal growthfactor receptor expression in normal ovarian epithelium andovarian cancer. II. Relationship between receptor expression andresponse to epidermal growth factor. Am. J. Obstet. Gynecol.164, 745–750.
Rubin, J.S., Bottaro, D.P., Chedid, M., et al., 1995a. Keratinocytegrowth factor. Cell Biol. Int. 19, 399–411.
Rubin, J.S., Bottaro, D.P., Chedid, M., et al., 1995b. Keratinocytegrowth factor as a cytokine that mediates mesenchymal–epithelialinteraction. Exs. 74, 191–214.
Scully, R.E., 1979. Tumors of the ovary and maldeveloped gonads.Atlas of Tumor Pathology. Armed Forces Institute of Pathology,Washington, DC, pp. 353–363.
Siemens, C.H., Auersperg, N., 1988. Serial propagation of humanovarian surface epithelium in tissue culture. J. Cell Physiol. 134,347–356.
Skinner, M.K., Parrott, J.A., 1994. Growth factor mediated cell–cellinteractions in the ovary. In: Findlay, J.K. (Ed.), MolecularBiology of the Female Reproductive System. Academic Press,New York, pp. 67–81.
Stromberg, K., Collins, T.J.T., Gordon, A.W., Jackson, C.L., John-son, G.R., 1992. Transforming growth factor-alpha acts as anautocrine growth factor in ovarian carcinoma cell lines. CancerRes. 52, 341–347.
Sugimura, Y., Foster, B.A., Hom, Y.K., et al., 1996. Keratinocytegrowth factor (KGF) can replace testosterone in the ductalbranching morphogenesis of the rat ventral prostate. Int. J. Dev.Biol. 40, 941–951.
Tukey, J.A., 1953. Problem of multiple comparisons. Princeton Uni-versity, Princeton, NJ.
Tukey, J., 1991. The philosophy of multiple comparisons. Stat. Sci. 6,100–116.
Vigne, J.L., Halburnt, L.L., Skinner, M.K., 1994. Characterization ofbovine ovarian surface epithelium and stromal cells: identificationof secreted proteins. Biol. Reprod. 51, 1213–1221.
Weiss, N.S., Homonchuk, T., Young, J.L. Jr., 1977. 1969–1971,Incidence of the histologic types of ovarian cancer: the US ThirdNational Cancer Survey. Gynecol. Oncol. 5, 161–167.
Werner, S., Duan, D.S., de Vries, C., Peters, K.G., Johnson, D.E.,Williams, L.T., 1992. Differential splicing in the extracellularregion of fibroblast growth factor receptor 1 generates receptorvariants with different ligand-binding specificities. Mol. Cell Biol.12, 82–88.
Woessner, J.F. Jr., Morioka, N., Zhu, C., Mukaida, T., Butler, T.,LeMaire, W.J., 1989. Connective tissue breakdown in ovulation.Steroids 54, 491–499.
Wong, W.S., Wong, Y.F., Ng, Y.T., et al., 1990. Establishment andcharacterization of a new human cell line derived from ovarianclear cell carcinoma. Gynecol. Oncol. 38, 37–45.
Yeh, J., Yeh, Y.C., 1989. Transforming growth factor-alpha andhuman cancer. Biomed. Pharmacother. 4, 651–659.
.