
Monitoring the structural consequences of Phe12→d-Phe and Leu15→Aib substitution in human/rat...
11
Monitoring the structural consequences of Phe12 fi D-Phe and Leu15 fi Aib substitution in human/rat corticotropin releasing hormone Implications for design of CRH antagonists Georgios A. Spyroulias, Spyridon Papazacharias, George Pairas and Paul Cordopatis Department of Pharmacy, University of Patras, Greece A new human/rat CRH analogue has been synthesized using the Fmoc/tBu solid-phase synthetic protocol. The sequence of the new peptide differs from the original in two positions, 12 and 15, at which the native amino acids L-phenylalanine 12 and L-leucine 15 have been replaced by the nonprotein amino acids D-phenylalanine and a-aminoisobutyric acid (Aib), respectively. The high resolution three-dimensional solution structure of [D-Phe12, Aib15]CRH has been determined by 688 dis- tance constraints (656 meaningful NOE and 32 H-bonds distance limits) and 21 angle constraints. A family of 40 energy-minimized conformers was obtained with average rmsd of 0.39 ± 0.16 A ˚ and 0.99 ± 0.13 A ˚ for backbone and heavy atoms, respectively, and distance penalty functions of 0.42 ± 0.03 A ˚ 2 . The NMR data acquired in a solvent system of water/trifluoroethanol (34%/66%, v/v) revealed that this 41-polypeptide adopts an almost linear helical structure in solution with helical content which reaches an 84% of the residues. Structural analysis con- firmed the existence of two helical peptide fragments. The first was comprised of residues Ile6–Arg16 and the second of residues Glu20–Ile40, forming an angle of 34.2Ŷ. The structural differences with respect to the native peptide have been identified in the region D-Phe12–Glu20 where double substitution at positions 12 and 15 seems to per- turb the elements of the native 35-residue helix. These structural rearrangements promote non-native intramo- lecular interactions in the region of the molecule between either the hydrophobic side-chains of D-Phe12, Aib15 and Leu18, or the charged groups of the residue pairs Arg16– Glu20 and His13–Glu17 being responsible for changes in hormonal functionality. This CRH analogue currently exhibits lack of any activity. Keywords: corticotropin releasing hormone; NMR; solid phase synthesis; solution structure; synthetic analogue. Corticotropin releasing hormone (CRH) is a 41-amino acid neuropeptide characterized by the presence of an isoleucine-amide in its C-terminal end (Scheme 1). It is produced mainly in the hypothalamus and acts as the principal neuroregulator of the secretion of adrenocor- ticotropic hormone (ACTH), b-endorphin and other proopiomelanocortin products of the anterior pituitary gland. CRH is considered to be the key hormone in the control of the hypothalamic–pituitary–adrenal axis func- tion and is responsible for the endocrine, autonomic, immunological and behavioural responses of mammalians to stress [1–3]. The hormone interacts at a nanomolar level with a transmembrane receptor belonging to the Gs-protein coupled family. CRH binds to membrane homogenates from the anterior pituitary, revealing a high-affinity component of 0.1 nM and a low-affinity binding site at 20 nM [4]. There is a wide distribution of CRH receptors throughout the human body, indicating that the hormone is probably implicated in a variety of actions, including inflammatory phenomena, the effects of various stressors, pregnancy and labour, food intake, thermoregulation, etc. [5–11]. The role of CRH has not yet been elucidated in every possible situation and new evidence is constantly appearing, establishing an even more complicated involvement in many biological func- tions. Moreover, inappropriate or disturbed CRH neur- onal activity manifests itself in various neuropsychiatric disorders such as affective disorders, anorexia nervosa and Alzheimer’s disease [12]. The great variety of functions mediated by CRH, or in which the hormone plays an active role, led to an effort to determine the structure–activity relationships which could clarify its action. Thus, an a-helical conformation of high amphiphilicity was found to be the major characteristic of the molecule’s tertiary structure. Residues 4–8 were found to be essential for receptor recognition, while residues 9–41 are responsible for binding and exertion of action [13]. The study of this molecule has led to the development of agonists Correspondence to P. Cordopatis, Department of Pharmacy, University of Patras, GR-265 04, Greece. Fax: + 30 2610997 714, Tel.: + 30 2610997 713, E-mail: [email protected] Abbreviations: CRH, corticotropin releasing hormone; Aib, a-aminoisobutyric acid; TFE, 2,2,2-trifluoroethanol; DQF-COSY, Double-quantum-filtered phase sensitive correlated spectroscopy; WATERGATE, water suppression by gradient-tailored excitation; REM, restrained energy minimization. Note: atomic coordinates of the 40 best and the mean energy mini- mized structures have been deposited in the Protein Data Bank (accession codes 1goe and 1go9, respectively). Note: a web site is available at http://www.pharmacy.upatras.gr (Received 22 July 2002, accepted 13 September 2002) Eur. J. Biochem. 269, 6009–6019 (2002) ȑ FEBS 2002 doi:10.1046/j.1432-1033.2002.03278.x
Transcript of Monitoring the structural consequences of Phe12→d-Phe and Leu15→Aib substitution in human/rat...

Monitoring the structural consequences of Phe12fi D-Pheand Leu15fiAib substitution in human/rat corticotropin releasinghormoneImplications for design of CRH antagonists
Georgios A. Spyroulias, Spyridon Papazacharias, George Pairas and Paul Cordopatis
Department of Pharmacy, University of Patras, Greece
A new human/rat CRH analogue has been synthesizedusing the Fmoc/tBu solid-phase synthetic protocol. Thesequence of the new peptide differs from the original intwo positions, 12 and 15, at which the native amino acidsL-phenylalanine 12 and L-leucine 15 have been replacedby the nonprotein amino acids D-phenylalanine anda-aminoisobutyric acid (Aib), respectively. The highresolution three-dimensional solution structure of[D-Phe12, Aib15]CRH has been determined by 688 dis-tance constraints (656 meaningful NOE and 32 H-bondsdistance limits) and 21 angle constraints. A family of 40energy-minimized conformers was obtained with averagermsd of 0.39 ± 0.16 A and 0.99 ± 0.13 A for backboneand heavy atoms, respectively, and distance penaltyfunctions of 0.42 ± 0.03 A2. The NMR data acquired ina solvent system of water/trifluoroethanol (34%/66%, v/v)revealed that this 41-polypeptide adopts an almost linearhelical structure in solution with helical content which
reaches an 84% of the residues. Structural analysis con-firmed the existence of two helical peptide fragments. Thefirst was comprised of residues Ile6–Arg16 and the secondof residues Glu20–Ile40, forming an angle of 34.2�. Thestructural differences with respect to the native peptidehave been identified in the region D-Phe12–Glu20 wheredouble substitution at positions 12 and 15 seems to per-turb the elements of the native 35-residue helix. Thesestructural rearrangements promote non-native intramo-lecular interactions in the region of the molecule betweeneither the hydrophobic side-chains of D-Phe12, Aib15 andLeu18, or the charged groups of the residue pairs Arg16–Glu20 and His13–Glu17 being responsible for changes inhormonal functionality. This CRH analogue currentlyexhibits lack of any activity.
Keywords: corticotropin releasing hormone; NMR; solidphase synthesis; solution structure; synthetic analogue.
Corticotropin releasing hormone (CRH) is a 41-aminoacid neuropeptide characterized by the presence of anisoleucine-amide in its C-terminal end (Scheme 1). It isproduced mainly in the hypothalamus and acts as theprincipal neuroregulator of the secretion of adrenocor-ticotropic hormone (ACTH), b-endorphin and otherproopiomelanocortin products of the anterior pituitarygland. CRH is considered to be the key hormone in thecontrol of the hypothalamic–pituitary–adrenal axis func-tion and is responsible for the endocrine, autonomic,immunological and behavioural responses of mammalians
to stress [1–3]. The hormone interacts at a nanomolarlevel with a transmembrane receptor belonging to theGs-protein coupled family. CRH binds to membranehomogenates from the anterior pituitary, revealing ahigh-affinity component of 0.1 nM and a low-affinitybinding site at 20 nM [4]. There is a wide distribution ofCRH receptors throughout the human body, indicatingthat the hormone is probably implicated in a variety ofactions, including inflammatory phenomena, the effectsof various stressors, pregnancy and labour, food intake,thermoregulation, etc. [5–11]. The role of CRH has notyet been elucidated in every possible situation and newevidence is constantly appearing, establishing an evenmore complicated involvement in many biological func-tions. Moreover, inappropriate or disturbed CRH neur-onal activity manifests itself in various neuropsychiatricdisorders such as affective disorders, anorexia nervosaand Alzheimer’s disease [12].
The great variety of functions mediated by CRH, or inwhich the hormone plays an active role, led to an effort todetermine the structure–activity relationships which couldclarify its action. Thus, an a-helical conformation of highamphiphilicity was found to be the major characteristic ofthe molecule’s tertiary structure. Residues 4–8 were found tobe essential for receptor recognition, while residues 9–41 areresponsible for binding and exertion of action [13]. Thestudy of this molecule has led to the development of agonists
Correspondence to P. Cordopatis, Department of Pharmacy,
University of Patras, GR-265 04, Greece.
Fax: + 30 2610997 714, Tel.: + 30 2610997 713,
E-mail: [email protected]
Abbreviations: CRH, corticotropin releasing hormone; Aib,
a-aminoisobutyric acid; TFE, 2,2,2-trifluoroethanol; DQF-COSY,
Double-quantum-filtered phase sensitive correlated spectroscopy;
WATERGATE, water suppression by gradient-tailored excitation;
REM, restrained energy minimization.
Note: atomic coordinates of the 40 best and the mean energy mini-
mized structures have been deposited in the Protein Data Bank
(accession codes 1goe and 1go9, respectively).
Note: a web site is available at http://www.pharmacy.upatras.gr
(Received 22 July 2002, accepted 13 September 2002)
Eur. J. Biochem. 269, 6009–6019 (2002) � FEBS 2002 doi:10.1046/j.1432-1033.2002.03278.x

and antagonists of CRH, which can be either peptidic ornonpeptidic. These analogues have the potential to bepharmacological tools for intervention in pathologicalconditions such as Cushing’s syndrome, inflammatory dis-eases, pre-eclampsia or preterm labour, anxiety and depres-sion and a potentially great number of other disorders.
We have previously described the synthesis and thebiological evaluation of a new series of CRH analogues,which showed that most analogues tested were deprived ofany significant agonist effects [14]. These findings promptedus to study the conformational features of the analogue[D-Phe12, Aib15]CRH through one- and two-dimensionalJ-correlated 1H NMR spectroscopy. Structure calculationswere performed using simulated annealing protocols andrestrained energy minimization (REM). Structural charac-teristics imposed by the introduction of the novel residuesin the sequence of the molecule and the differences betweenthe CRH analogue and the native structure [15] are alsodiscussed.
The major goal of our study is the solution structuredetermination of a CRH analogue devoid of any agonistactivity, a fact particularly relevant to the further design anddevelopment of CRH antagonists with wide-ranging clinicalapplications in psychiatric disorders.
M A T E R I A L S A N D M E T H O D S
Sample preparation
The CRH analogue was synthesized using Fmoc/tBuchemistry by solid-phase peptide synthesis and was purifiedand characterized via reversed phase HPLC and ESI-MS[14]. The CRH analogue was dissolved to a final concen-tration of 2–2.5 mM in H2O/2,2,2-trifluoroethanol (TFE)-d2(CORTEC, Paris, France; 34%/66%, v/v) to record one-and two-dimensional NMR spectra, and the pH wasadjusted to 3.8. For amide exchange experiments, thepeptide was lyophilized and dissolved in D2O/TFE-d3(34%/66%, v/v).
NMR spectroscopy
NMR spectra were recorded on Bruker’s 500 and 600-MHzspectrometers in 298K and 310K. One- and two-dimen-sional double-quantum-filtered-COSY [16], TOCSY [17,18]and time-proportional phase incrementation-NOESY[19,20] spectra were acquired in the 10–14 p.p.m. spectrumwindow using PRESATURATION [21] and WATER-GATE [22] pulse sequences for H2O suppression. Spectrawere calibrated according to the characteristic methyleneresonance of TFE-d2 at 3.926 p.p.m. TOCSY experimentswere carried out with a mixing time of 80–100 ms, whereasNOESY experiments were recorded using 100 and 200 msmixing times. All two-dimensional spectra were acquiredusing a spectral width of 6410.26 Hz (10.7 p.p.m) andconsisted of 2048 data points in the F2 dimension, 16–32
transients and 720–1024 complex increments in the F1dimension. Raw data were multiplied in both dimensions bya pure cosine-squared bell window function and Fourier-transformed to obtain 2048 · 1024 or 2048 · 2048 real datapoints. A polynomial base-line correction was applied inboth directions.
NMR data processing was performed using thestandard Bruker software package on a Silicon GraphicsO2 workstation. The two-dimensional maps were ana-lysed on Silicon Graphics O2 or PentiumIII PC-Linuxcomputers with the aid of the XEASY program (ETH,Zurich) [23].
NOE constraints, 3JHNHa and CaH chemical shift analysis
A total of 1868 NOESY cross-peaks were assigned in bothdimensions and their intensities were converted into upperlimits of interatomic distances [24], yielding 928 uniqueNOE constraints (23.1 per assigned residue).
3JHNHa were determined using the Protocol 1 ofD. S. Wishart et al. [25]. Twenty-one out of 28 measured3J were calculated in the range of 4.8–5.6 Hz or >8.0 Hz[26] and introduced to the calculations. Seven 3JHNHa werefound in the range 6–7 Hz and were not used in structuralcalculations [27].
Slowly exchanging amides were identified using thelyophilized original sample in D2O/TFE-d3 solution,through two-dimensional NOESY spectra over a timeperiod of 4–5 h.
Chemical shift analysis was performed according to theapproach established by Wishart et al. [28,29] and shiftdifferences between the Ha-protons of the human/rat CRHanalogue and their values for coil conformation wereextracted. No correction for the solvent was applied to theinitial CaH shifts values drawn from the TOCSY/NOESYmaps [30].
Structure calculations and refinement
Structure was calculated using the NMR data acquired at310 K, the temperature at which native human/rat CRHsolution structure was also determined. Of 928 constraints,656 were found meaningful (16.4 per assigned residue) inDYANA structural calculations [31]. Appropriate pseudo-atom corrections were applied to methylene and methylhydrogens that were not stereospecifically assigned [32].a-Aminoisobutyrilo (Aib) residue is derived from a valinewhere the Ca of the Aib corresponds to valine Ca andCb1/Cb2 of the Aib corresponds to valine Cc1/Cc2.
Sixteen hydrogen bonds involving nonexchangeableHN amide protons, with an occurrence of 87.5% (in 35out of 40 structures), were used as additional structuralconstraints in the final stage of DYANA calculation.Upper (2.40 A) distance limits between HN and O atomsinvolved in hydrogen bonds, together with upper(3.30 A) and lower (2.60 A) distance limits between thecorresponding N and O atoms, were set for each one ofthe 16 hydrogen bonds. Fourteen pairs of diastereotopicprotons (among them the two methyls of Aib15) werestereospecifically assigned through the GLOMSA program[24].
Each one of the 40 best structures (out of 400 calculated)in terms of DYANA target function (larger NOE constraint
Scheme 1. Primary structure of various natural CRH peptides.
6010 G. A. Spyroulias et al. (Eur. J. Biochem. 269) � FEBS 2002

violation 0.09 A) has been refined through REM (AMBER
5.0 [33], SANDER [34]). The whole procedure followedprotocols previously applied to the solution structuredetermination of larger biomolecules, such as metallopro-teins [35]. A force constant of 133.76 kJÆmol)1 A2 wasapplied for the distance constraints.
Structural calculations were performed on IBM RISC6000 or PentiumIII PC-Linux computers.
Quality assessment of the structure
PROCHECK [36] and PROCHECK-NMR [37] programs wereused to check the stereochemical quality of the peptidestructure and provide the elements of the secondarystructure, as well as other statistical parameters.
R E S U L T S
NMR spectra–proton assignment
The spin patterns of 39 out of 41 residues have beenidentified through analysis of the TOCSY spectrum (Fig. 1),whereas Ser1 and Leu27 proton resonances have not beenassigned at this stage. The latter have been identified inNOESY maps recorded at two different temperatures dueto their sequential dipolar connectivities while the formerhave remained undetectable in all two-dimensional mapsat both temperatures. Proton chemical shifts are given inTable S1.
Secondary structure
Fig. 2A shows the short and medium range NOEs observedfor the backbone and CaH protons. Sequential HN–HN
connectivities for stretches up to two amino acids wereobserved for residues 6–13, 14–19, 20–28, 29–31, and 31–40.HN–HN connectivities involving the HN resonances of Ser7/Thr11, Phe12/Leu14, Val18/Met21, Leu19/Glu20 andGln26/Ala28/Gln30, could not be detected due to theirdegenerated shifts. The same is true of some Ha–HN
medium-range connectivities, where the Ha resonances ofAla28/Gln29 and Arg35/Lys36 have degenerated chemicalshifts.
Eighteen 3JHNHa (Fig. 2B, upper panel) were found tobe £ 5.6 Hz while three (Glu2, Glu3 and Val18) werefound to be in the range 8.0–8.5 Hz. 3JHNHa for D-Phe12
was found 6.9 Hz and six (Leu10, Thr11, Leu19, Gln26,Ile39 and Ile40) exhibited values in the range 6.0–6.6 Hz.
Chemical shift difference analysis between the observedCaH shift values and the corresponding random coilvalues is presented in Fig. 2B (lower panel) and providesstrong evidence for the conformational preference of themajority of amino acids towards the helical configur-ation. The only tetrapeptide in which Ha values presentlarge positive differences from random coil values isthe N-terminal Glu2-Pro5/Ile6 region (Ile6 Dd Ha
0.06 p.p.m). A three-residue fragment, Ala31–Asn34,exhibited slightly positive shift differences (in the rangeDd Ha 0.009–0.045 p.p.m) while the remaining 32 res-idues in the region Ser7–Ile41 exhibited negative Ha shiftdifferences in the range 0.011–0.524 p.p.m. According todata reported previously the conformation of thatfragment seems to be different in ovine and humanCRH, and for this reason its structure has been discussedin the literature [38] (Scheme 1). However, despite thepositive Ha shift difference, Ala31–Asn34 Ha chemicalshift values compared with the averaged Ha shifts forhelical and b-sheet conformation [28] and they exhibiteda smaller deviation from a-helical rather than b-strandvalues (see Fig. 2C).
Tertiary structure
An overall evaluation of NOE, 3JHNHa and Ha chemicalshifts suggests a helical conformation for the peptidefragment Ile6–Ile40 and an extended structure for theN-terminal hexapeptide. Structure was calculated usingNMR data acquired at 310 K (as in native CRH [15]). Theresulting DYANA family of 40 structures has rmsd values(calculated for residues 6–41) of 0.56 ± 0.24 A and1.40 ± 0.20 A for backbone and heavy atoms, respectively.The target function lies in the range 0.10–0.12 A2. The finalREM family exhibits pair-wise rmsd values of0.56 ± 0.23 A (backbone), 1.42 ± 0.19 A (heavy atoms)for the 40 structures and 0.39 ± 0.16 A (backbone),0.99 ± 0.13 A (heavy atoms) for the mean structure. TheNOE distance penalty function for the family of 40structures is 0.42 ± 0.03 A2 and 0.44 A2 for the meanstructure. The corresponding values for H-bond and angle
constraints are 0.016 ± 0.01 A2 (40 structures), 0.012 A2
(mean) and 0.014 ± 0.006 A2 (40 structures), 0.005 A2
(mean), respectively (see also Table 1) and in both cases the
Fig. 1.1H-
1H two-dimensional TOCSY 600-
MHz NMR of the fingerprint region of Ha–HN
protons recorded on a 10.7-p.p.m. spectrum
width (H2O/TFE-d2 34%/66% v/v, at
pH = 3.8, T = 310 K). The number of the
amino acid in the CRH sequence, to which the
Ha–HN connectivity belongs, is noted.
� FEBS 2002 NMR structure of a CRH synthetic analogue in TFE/H2O (Eur. J. Biochem. 269) 6011

penalty function is around 30 times smaller than that ofNOEs. The NOE-derived distance constraints are reportedin Fig. 2D while rmsd per residue for the final family isgiven in Fig. S1. Fig. 3A shows the family of 40 structuresand Table 1 contains a summary of statistical data indica-ting the quality of the 40 models and the average energy-minimized structure.
The polypeptide chain of the human/rat CRH analogueseems to fold to an almost linear tertiary structure with themain features being the high degree of helical character anda small bend along the principal molecular axis. As far asthe ill-defined N terminus is concerned, all data (chemicalshift index, 3JHNHa, and u vs. w plots, Fig. S2) imply aconformational preference for an extended structure. Theunambiguous conformational preference for helical struc-ture for the C-terminal 10-peptide is manifested by numer-ous helix-diagnostic NOEs (see also Fig. 2A), and the3JHNHa values. Strong evidence for the Glu39–Ile41 tripep-tide’s compact structure was obtained from the intenseNOESY peaks of the two –CONH2 amide protons which,in turn, give rise to a network of HN–HN cross-peaksthroughout in this region. The numerous OiH
Niþ 4 type
H-bonds identified after the final DYANA calculation (Ala28O¢–HN His32, Gln29 O¢–HN Ser33, His32 O¢–HN Lys36,Ser33 O¢–HN Leu37, and Arg35 O¢–HN Glu39 in 40 out
of 40 DYANA models) further support the continuousC-terminal helix structure.
Monitoring the Phe12fiD-Phe and Leu15fiAibsubstitution
The NOE pattern of the CRH analogue involving HN, Ha
and Hb protons of D-Phe12, was constituted only bydNN(i,i + 2) and daN(i,i + 2) connectivities, while connec-tivities diagnostic for a-helix such as daN(i,i + 3),dab(i,i + 3) and daN(i,i + 2) were not observed (Fig. 2A).
The Aib15 residue possesses two diastereotopic methylgroups, with the pro-R methyl replacing the Ha proton atCa carbon of an L-valine and the pro-S group correspondingto the side chain (Cb of an L-valine). The protons of the twoCbH3 groups were found to be chemically and magneticallynonequivalent and to yield two different resonances in theregion 1.70–1.90 p.p.m. Moreover, the stereospecific assign-ment of these methyl groups was achieved through analysisof the observed spin couplings and the intensities ofdbN(i,i + 1), dbN(i,i + 2) and dbN(i,i + 3) NOEs. Thus,the Cb1 methyl group which resonates at 1.861 p.p.m. isassigned to the pro-S because it exhibits NOEs of lowintensity in respect with Cb2 (1.716 p.p.m) methyl, of thetype dbN(i,i + 2) and dbN(i,i + 3) [39].
Fig. 2. Analysis of NMR-derived data. (A)
Schematic representation of the sequential and
medium-range NOE connectivities involving
NH, a- and b-protons. The thickness of the
bar indicates the intensity of NOEs. The
average, over the 40 DYANA structures,
secondary structure elements are also
reported. (B) Measured values of 3JHNHa
coupling constants in Hz (upper panel) and
Ha chemical shift differences from random coil
values (lower panel). (C) Comparison of Ha
chemical shift values of [D-Phe12, Aib15]CRH
with the corresponding Ha chemical shifts for
helical ( ) and b-strand (m) configurations.
(D) Number of meaningful NOE constraints
per residue for [D-Phe12, Aib15]CRH used in
the structural calculations. White, grey, and
dark grey bars represent, respectively,
intraresidue, sequential, and medium-range
connectivities. Long-range connectivities were
not observed.
6012 G. A. Spyroulias et al. (Eur. J. Biochem. 269) � FEBS 2002

Distribution of electrostatic charges on the surfaceand along the principal axis of the molecule
In the synthetic CRH analogue, as in native peptide, thereare seven negatively charged (six Glu and one Asp) and fourpositively charged residues. Analysis of the distribution ofthe electrostatic potential on the surface of the moleculereveals the ampiphilic character of the helix (Fig. 4A).Schematic representation of the charge distribution alongand perpendicularly to the [D-Phe12, Aib15]CRH axis ispresented at Fig. 4B and C. Non-native interaction betweencharged groups in this synthetic analogue is discussedbelow.
D I S C U S S I O N
Analysis of the calculated structures performed by thePROCHECK program provides quality data for the calculatedCRH models (Table 1). Neither the 40 models nor the meanstructure possess a residue in disallowed regions of Rama-chandran plot (Table 1 and Fig. S2).
According to the PROCHECK-NMR [37] program, theaverage of the 40 models and the mean minimized structureof the synthetic analogue [D-Phe12, Aib15]CRH, contain upto 84% helical structure. This value is either somewhatlarger than that of the 76–78% regarding the helicity of thenative peptide in solution containing 66–100% of TFE
[15,40], or identical to the 84% for studies carried out in40% TFE in an H2O mixture [38]. According to studies ofCRH binding to single bilayer egg phosphatidylcholinevesicles, the development of the intramolecular interactionwith the cell membrane is accompanied by the induction ofa a-helical structure and this conformation characterizes thebiologically active form of the peptide [41]. TFE assistsamphipathic peptides with high helical propensity, such asCRH, to reach the maximum helical content at around 40%TFE [42], since it diminishes the exposure of CO and NHgroups in the solvent [43], favouring hydrogen bondformation [44]. This apparently occurred in the case of thecurrently studied CRH molecule and the same has beenreported earlier for ovine and human/rat-CRH [38,41].
In calculated models, the CRH a-helix consists of 34residues in regions 6–18 and 20–40 and this is fully consistentwith the diagram of sequential dipolar connectivities(Fig. 2A), the 3Js and OiH
Niþ 4 type H-bonds. The structure
of the Gln30–Ile40 fragment has been determined withhigh resolution and exhibits remarkably low rmsd values(0.28–0.54 A). These data: (a) suggest a conformationaldifference with the structureless C terminus of ovine CRH[38] and (b) contradict the helix-stop signal identified for thenative human/rat CRH in the residue fragment 31–34 andthe ill-defined terminal pentapeptide 37–41 [15,38].
Leu19 interrupts the helical structure in all structurescomposing the family of 40 models and also in the mean
Fig. 2. (Continued).
� FEBS 2002 NMR structure of a CRH synthetic analogue in TFE/H2O (Eur. J. Biochem. 269) 6013
structure. However, in various DYANA models, helicalstructure is also interrupted for two up to five residues inthe 6/9–13/15 region. Destabilization of the helix infragment 9–13 could be discussed in terms of theintroduction of a D-amino acid at position 12 [45].Destabilization caused by D-amino acid depends on thenature of its side chain (charged or not, bulky or not) andD-phenylalanine has been found to present one of thehighest a-helix disturbing propensities [46]. Loss of NOEsbetween D-Phe and sequential residues in the CRHanalogue was observed as in a similar kind of L-/D-aminoacid substitution [45,46], and suggests slight helix destabil-ization in the region of Leu10, Thr11 and D-Phe12. Forthis fragment, experimental coupling constant values aremeasured between 6 and 7 Hz (Fig. 2B), suggestingconformational averaging over the contributing conforma-tions [27].
On the other hand, the helix-promoter/stabilizer Aib,which replaced Leu15, increases the helix propensity of apeptide fragment towards either a- or 310-helix by restrictionof the backbone conformational freedom [47–49]. Thislimitation is imposed by the steric hindrance that arises fromits gem-dimethyl group. The torsion angles for the main-chain conformation of Aib15 in the family of 40 energyminimized structures are calculated as u¼ )59.6� ± 1.5and w¼ )31.9 ± 1.5, while for the mean energy minimizedstructure they are )56.5� and )31.0�, respectively, close tothose ofAib conformational energyminima (u ¼ ) 58� and
w¼ )34� [50]). According to these values, the Aib u torsionangle deviates less from a- than from 310-helix typical values()55.2� and )49.9�, respectively [48]) while the w torsionangle is practically the same as the ideal w-value ()34�) for310 helical configurations ()52.2� for a- helix [48]). As far asthe surrounding amino acids in the His13–Glu17 fragmentare concerned, His13 and Glu17 w angles are found slightlyout of the range of values for a-helix structure, while wangles for Leu14 and Arg16 are considerably close to therange of values for 310 helix. Additionally, Leu14 possess a utorsion angle almost typical for the 310 helix.
In this region of the polypeptide chain, the secondarystructure is characterized also by the OiH
Niþ 3 hydrogen
bonds, found in all 40 DYANA structures in His13–Arg16and Leu14–Glu17, which are indicative of a 310-, rather thana pure a-helical structure. Nevertheless, H-bonds OiH
Niþ 4,
typical in a-helices, are also observed in Leu14–Val18 andAib15–Leu19.
All of the above data imply that the His13–Leu19backbone structure is probably a conformational averagebearing features of a pure a-helix and 310-helix structure.This is in agreement with the �ambihelicity� behaviourpattern stated by Basu et al. [51,52] according to which thea- and 310-helical forms are almost equivalent energeticallywhen only one or two Aib residues replace other aminoacids in a peptide sequence.
Furthermore, a noticeable feature of the [D-Phe12,Aib15]CRH molecule is a characteristic bend occurring
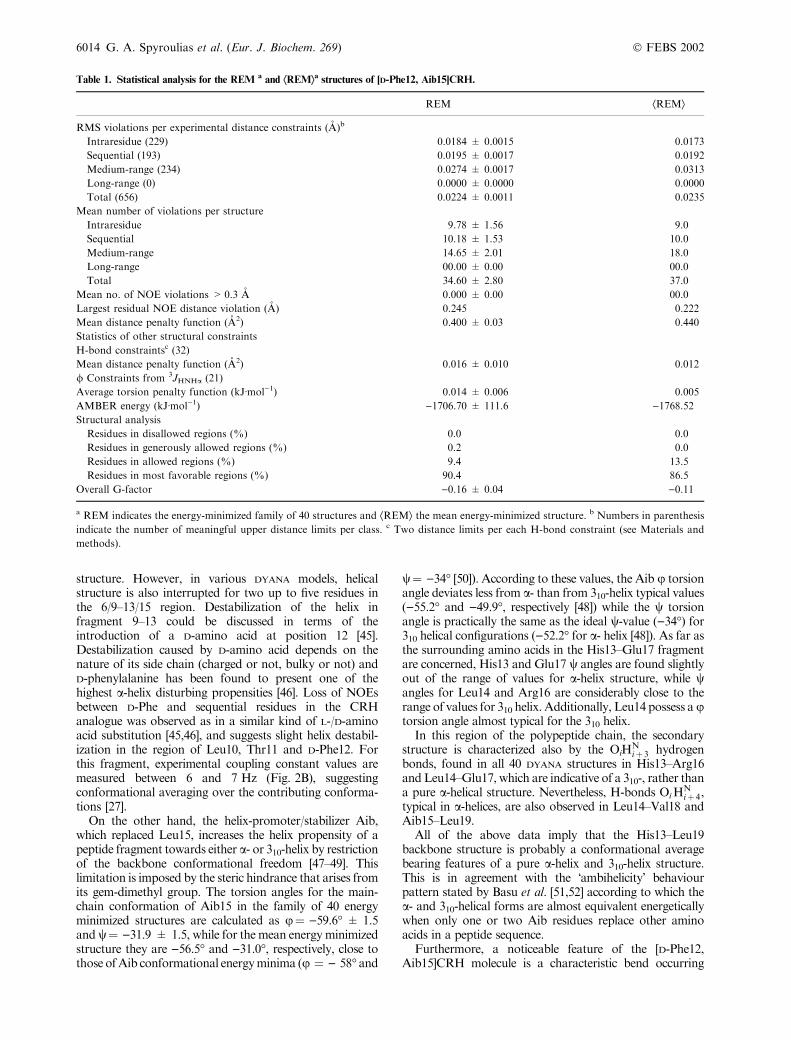
Table 1. Statistical analysis for the REM a and ÆREMæa structures of [D-Phe12, Aib15]CRH.
REM ÆREMæ
RMS violations per experimental distance constraints (A)b
Intraresidue (229) 0.0184 ± 0.0015 0.0173
Sequential (193) 0.0195 ± 0.0017 0.0192
Medium-range (234) 0.0274 ± 0.0017 0.0313
Long-range (0) 0.0000 ± 0.0000 0.0000
Total (656) 0.0224 ± 0.0011 0.0235
Mean number of violations per structure
Intraresidue 9.78 ± 1.56 9.0
Sequential 10.18 ± 1.53 10.0
Medium-range 14.65 ± 2.01 18.0
Long-range 00.00 ± 0.00 00.0
Total 34.60 ± 2.80 37.0
Mean no. of NOE violations > 0.3 A 0.000 ± 0.00 00.0
Largest residual NOE distance violation (A) 0.245 0.222
Mean distance penalty function (A2) 0.400 ± 0.03 0.440
Statistics of other structural constraints
H-bond constraintsc (32)
Mean distance penalty function (A2) 0.016 ± 0.010 0.012
/ Constraints from 3JHNHa (21)
Average torsion penalty function (kJÆmol)1) 0.014 ± 0.006 0.005
AMBER energy (kJÆmol)1) )1706.70 ± 111.6 )1768.52
Structural analysis
Residues in disallowed regions (%) 0.0 0.0
Residues in generously allowed regions (%) 0.2 0.0
Residues in allowed regions (%) 9.4 13.5
Residues in most favorable regions (%) 90.4 86.5
Overall G-factor )0.16 ± 0.04 )0.11
a REM indicates the energy-minimized family of 40 structures and ÆREMæ the mean energy-minimized structure. b Numbers in parenthesis
indicate the number of meaningful upper distance limits per class. c Two distance limits per each H-bond constraint (see Materials and
methods).
6014 G. A. Spyroulias et al. (Eur. J. Biochem. 269) � FEBS 2002

in the 12–16 region which is a striking difference from thenative CRH molecule. This feature is apparently related tothe contemporary incorporation of a D- and an a,a-dialkylo amino acid in close proximity within the CRHsequence. Besides its tendency to form constrained helices,Aib is also known for its ability to bend helical peptides[53]. Analogous helical structure perturbation has beenobserved where either a sequential double replacement ofD-amino acid [46] or a double incorporation of Aib residuein a tripeptide fragment [53] has been carried out. These
features in the CRH analogue are supported by numerous,well-defined NOE cross-peaks between the side-chains ofthe residues in the 12–22 region (see Fig. 3B). Ha, Hb andCc2 methyl protons of Thr11 give rise to NOE interactionswith Aib15 methyls while a NOE signal was also observedbetween Thr11 Cc2 methyl and Aib15 HN proton. Aib15Cb1 methyl (1.861 p.p.m) participates in a NOE networkinvolving the backbone or side-chain protons of theresidues D-Phe12 (Ha, phenyl ring Hd and He), His13(HN), Leu14 (HN) and Val18 (HN, Cc1 methyl). On the
Fig. 3. (A) The family of 40 structures calculated for the [D-Phe12, Aib15]CRH analogue and (B) the mean energy minimized [D-Phe12, Aib15]CRH
structure and conformation of selected residues in the site where double substitution has been performed. Figures were generated with the program
MOLMOL [55].
� FEBS 2002 NMR structure of a CRH synthetic analogue in TFE/H2O (Eur. J. Biochem. 269) 6015

6016 G. A. Spyroulias et al. (Eur. J. Biochem. 269) � FEBS 2002

other hand the Cb2 methyl (1.716 p.p.m) gives rise tointense NOE cross-peaks with practically all Val18 protons(HN, Ha, Hb, Cc1 and Cc2 methyls). Additionally, dipolarinteractions between HN, Ha, Cc1 and Cc2 Val18 and HN
and Cb Ala22 protons have been observed. This distortionof the linear structure of human/rat CRH [15] in the regionof Aib residue probably acts as a helix-breaking signal inthe region of Val18–Leu19 (3JHNHa 8.0 and 6.0 Hz,respectively) justifying the nonhelical backbone conforma-tion of Leu19 observed in all calculated models andaverage structures.
The question raised at this point is whether or not theseNOEs are consistent with an extended helix such as that ofthe native CRH. As no coordinates for the native CRHNMR structure were available, a direct comparison couldnot be performed. However, we executed an additionalstructural calculation excluding the Aib15 NOE-derivedconstraints. In the absence of Aib15 constraints, the frag-ment His13–Leu19 becomes linear and in some modelsthe fragment His13–Glu17 possesses a pure a-helicalconformation.
However, the extended CRH native, Ile6–Ile40 helix, isdivided in this synthetic analogue, into two independenthelical fragments, Ile6–Val18 and Glu20–Ile40, respectively.These two 13-residue N- and 21-residue C-terminal frag-ments form an angle of 34.2� (Fig. 3B). This bend, inconcert with the His13–Aib15 3- and Aib15–Leu19 4-resi-due turns, help the D-Phe12 aromatic ring to approachthe vicinal Aib methyl while Val18 main and side-chainapproach the other Aib methyl group (Fig. 3B). Theaverage distance between the vicinal Val18 and Aib15methyls is measured as 3.206 A in the mean structure, whilethe other methyl of Aib15 is found 3.817 and 3.903 A fromthe aliphatic Cb and the vicinal Hd aromatic proton ofD-Phe12, respectively.
Assisted by this bend, D-Phe12, Aib15 and Val18 form ahighly hydrophobic core in the middle of the CRHsequence, a region that is particularly important for receptorbinding and activation [41,54]. On the opposite side, the�external part� of this helical fragment, the extended sidechains of Arg16 and Glu20, as well as those of His13 andGlu17, are oriented towards the same direction and arerelatively close. The Oe1 and Oe2 atoms of Glu20 are 3.726and 1.983 A, respectively, from the imine He proton and3.644 and 5.121 A from the proximal guanidyl Hg11 andHg12 protons of Arg16 (Fig. 3B). On the other hand, theOe2 atom of Glu17 is found at 5.485 A, far from the Hd2
proton of the imidazole ring of His13.The above-mentioned side-chain interactions between
His13–Glu17 and Arg16–Glu20 are remarkably different inthe calculated models without Aib15 NOEs. When thefragment His13–Leu19 is linear, the interatomic distancesbetween Arg16 He, Hg11/Hg12 and Glu20 Oe1/Oe2 becomeon average larger than 5.0–6.0 A, whereas when the 310-helix conformation becomes a pure a-helix, the His13–
Glu17 atoms (Hd2 and Oe2, respectively) are found atdistances larger than 6.5–7.0 A. The above data couldprobably be indicative for a native-like linear CRHstructure and suggest a conformational difference betweenthe parent and the synthetic molecule.
C O N C L U D I N G R E M A R K S – B I O L O G I C A LI M P L I C A T I O N S
The NMR data acquired and analysed in 66% TFE sug-gest that the synthetic analogue [D-Phe12, Aib15]CRHadopts a rather helical structure (84% of residues), which isconsidered to be the active conformation of the hormone[38,40]. The changes engineered in the sequence of thepeptide, either through the nature of the side chain of thenew amino acids or through their non-native conforma-tional features, increase the peptide–CRH complex stabilitywith the following consequences: they (a) differentiate thehelical content and/or characteristics (a- or 310-helix); (b)alter the hydrophobicity of the molecule in the 16–20region which is considered the �interface� between the 6–19region, where the side-chains of the hydrophobic part ofthe helix play a key role in receptor binding and activa-tion [54], and the 20–41 region, where side-chains have aprominent role in structural conservation rather than infunction [54]; and (c) disturb the linearity of the molecule inthe region Ile6–Ile40.
Helix bending is a structural feature that has not beenobserved in the solution structure of the native peptidehuman/rat CRH [15] and has not been implied by studiesof other CRH sequences [38]. Although the peptide bondsin the Aib15–Glu20 region could be accommodated with-out severely perturbing the helical character of bothfragments, the helix breaking/bending effect is stronglycoupled with reorientation of the residue side-chains in theamphipathic region 12–20. Indeed, helix bending dimi-nishes the exposure of the hydrophobic core of Phe12,Aib15 and Val18 to the solvent, while establishing thestructural basis for the side-chain interaction of the chargedresidues Arg16 and Glu20.
The biological impact of the Aib15 and D-Phe12modifications on the synthetic CRH analogue is illustratedby the peptide binding affinity enhancement towards theCRH receptor and its modified biological response. Morespecifically, this CRH synthetic analogue seems to effect-ively bind to the receptor pocket while its biological activityis extinguished when its effect on the release of catechol-amines from the PC12 rat pheochromocytoma cell line iscompared with that of native CRH. Consequently, theseproperties imposed on CRH comprise the first and possiblythe most important achievement in engineering a pharma-cologically interesting compound with properties anta-gonistic to CRH receptors. Analysis of the non-nativestructural features provides valuable information towardsthe design and synthesis of new molecules with higherbinding affinity and enhanced stability against biologicaldegradation and possibly biological activity. The impact ofAib introduction in the position of residues with similar ordifferent hydrophobic character, in the amphipathic regionLeu14–Glu20, was studied in silico before the synthesis ofnew analogues. Conformational studies and biologicalactivity evaluation of new CRH analogues are in progressin our laboratory.
Fig. 4. (A) Surface electrostatic potentials of CRH analogue, displayed
with MOLMOL [55] and schematic representation of the charge resi-
dues’ distribution along (B) and (C) perpendicularly to the axis of the
molecule is also presented. The molecular surface is colour coded:
potential less than )10 kT are illustrated in red while potential larger
than 10 kT in blue and neutral (0 kT) potential are white.
� FEBS 2002 NMR structure of a CRH synthetic analogue in TFE/H2O (Eur. J. Biochem. 269) 6017

A C K N O W L E D G E M E N T S
General Secretariat of Research and Technology of Greece (P.C) and
Marie Curie Research Training Grant – Contract No. HPMF-CT-
1999-00344 (G.A.S) are acknowledged for financial support. EC’s
Access to Research Infrastructures Action of the Improving Human
Potential Program (PARABIO, Contract No. HPRI-CT-1999-00009)
and CERM (University of Florence) are also acknowledged for access
to NMR instrumentation and to powerful computational systems.
R E F E R E N C E S
1. Vale, W., Spiess, J., Rivier, C. & Rivier, J. (1981) Characteriza-
tion of a 41-residue ovine hypothalamic peptide that stimulates
secretion of corticotropin and beta-endorphin. Science 213, 1394–
1397.
2. Vale, W., Rivier, C., Brown, M.R., Spiess, J., Koob, G. &
Swanson, A. (1983) Chemical and biological characterization of
corticotropin releasing factor. Recent Prog. Horm. Res. 39, 245–
270.
3. Orth, D. (1992) Corticotropin-releasing hormone in humans.
Endocr. Rev. 13, 164–191.
4. De Souza, E.B., Insel, T.R., Perrin, M.H., Rivier, J., Vale, W.W. &
Kuhar, M.J. (1985) Differential regulation of corticotropin-
releasing factor receptors in anterior and intermediate lobes of
pituitary and in brain following adrenalectomy in rats. Neurosci.
Lett. 56, 121–128.
5. Perrin, M., Donaldson, C., Chen, R., Blount, A., Berggren, T.,
Bilezikjian, L., Sawchenko, P. & Vale, W. (1995) Identification of
a second corticotropin-releasing factor receptor gene and char-
acterization of a cDNA expressed in heart. Proc. Natl Acad. Sci.
USA 92, 2969–2973.
6. Kishimoto, T., Pearse, R.V. II, Lin, C.R. & Rosenfeld, M.G.
(1995) A sauvagine/corticotropin-releasing factor receptor
expressed in heart and skeletal muscle. Proc. Natl Acad. Sci. USA
92, 1108–1112.
7. Lovenberg, T.W., Liaw, C.W., Grigoriadis, D.E., Clevenger, W.,
Chalmers, D.T., De Souza, E.B. & Oltersdorf, T. (1995) Cloning
and characterization of a functionally distinct corticotropin-
releasing factor receptor subtype from rat brain. Proc. Natl Acad.
Sci. USA 92, 836–840.
8. Chen, R., Lewis, K.A., Perrin, M.H. & Vale, W.W. (1993)
Expression cloning of a human corticotropin-releasing-factor
receptor. Proc. Natl Acad. Sci. USA 90, 8967–8671.
9. Karalis, K., Sano, H., Redwine, J., Listwark, S., Wilder, R.L. &
Chrousos, G.P. (1991) Autocrine or paracrine inflammatory
actions of corticotropin-releasing hormone in vivo. Science 254,
421–423.
10. Makrigiannakis, A., Margioris, A.N., Le Goascogne, C.,
Zoumakis, E., Nikas, G., Stournaras, E., Psychoyos, A. &
Gravanis, A. (1995) Corticotropin-releasing hormone (CRH) is
expressed at the implantation sites of early pregnant rat uterus.
Life Sci. 57, 1869–1875.
11. Zoumakis, E., Makrigiannakis, A., Margioris, A.N., Stournaras,
E. & Gravanis, A. (1996) Corticotropin releasing hormone (CRH)
in normal and pregnant uterus: physiological implications. Front.
Biosci. 1, e1–8.
12. Owens, M.J. & Nemeroff, C.B. (1991) Physiology and pharma-
cology of corticotropin-releasing factor. Pharmacol. Rev. 43, 425–
473.
13. Gilligan, P.J., Robertson, D.W. & Zaczek, R. (2000) Cortico-
tropin releasing factor (CRF) receptor modulators: progress and
opportunities for new therapeutic agents. J. Med. Chem. 43, 1641–
1660.
14. Papazacharias, S., Pairas, G., Kouimtzoglou, E., Dermitzaki, E.,
Manessi-Zoupa, E., Gravanis, A. & Cordopatis, P. (2001)
Synthesis and biological evaluation of new h/rCRH peptide
analogs. In Peptides 2000 (Martinez, J. & Fehrentz, J.-A., eds), pp.
933–934. EDK, Paris, France.
15. Romier, C., Bernassau, J.-M., Cambillau, C. & Darbon, H. (1993)
Solution structure of human corticotropin releasing factor by 1H
NMR and distance geometry with restrained molecular dynamics.
Protein Eng. 6, 149–156.
16. Piantini, U., Sorensen, O.W. & Ernst, R.R. (1982) Multiple
quantum filters for elucidating NMR coupling networks. J. Am.
Chem. Soc. 104, 6800–6801.
17. Braunschweiler, L. & Ernst, R.R. (1983) Coherence transfer by
isotropic mixing: application to proton correlation spectroscopy.
J. Magn. Reson. 53, 521–528.
18. Bax, A. & Davis, D.G. (1985) MLEV-17-based two-dimensional
homonuclear magnetization transfer spectroscopy. J. Magn.
Reson. 65, 355–360.
19. Marion, D. & Wuthrich, K. (1983) Application of phase sensitive
two-dimensional correlated spectroscopy (COSY) for measure-
ments of 1H-1H spin-spin coupling constants in proteins. Biochem.
Biophys. Res. Commun. 113, 967–974.
20. Jeener, J., Meier, B.H., Bachmann, P. & Ernst, R.R. (1979)
Investigation of exchange processes by two-dimensional NMR
spectroscopy. J. Chem. Phys. 71, 4546–4553.
21. Macura, S., Wuthrich, K. & Ernst, R.R. (1982) The relevance of
J cross-peaks in two-dimensional NOE experiments of macro-
molecules. J. Magn. Reson. 47, 351–357.
22. Piotto, M., Saudek, V. & Sklenar, V. (1992) Gradient-tailored
excitation for single-quantum NMR spectroscopy of aqueous
solution. J. Biomol. NMR 2, 661–666.
23. Eccles, C., Guntert, P., Billeter, M. & Wuthrich, K. (1991) Effi-
cient analysis of protein 2D NMR spectra using the software
package EASY. J. Biomol. NMR 1, 111–130.
24. Guntert, P., Braun, W. & Wuthrich, K. (1991) Efficient compu-
tation of three-dimensional protein structures in solution from
nuclear magnetic resonance data using the program DIANA
and the supporting programs CALIBA, HABAS and GLOMSA.
J. Mol. Biol. 217, 517–530.
25. Wang, Y., Nip, A.M. & Wishart, D.S. (1997) A simple method to
quantitatively measure polypeptide JHNHa coupling constants
from TOCSY or NOESY spectra. J. Biomol. NMR 10, 373–382.
26. Smith, L.J., Bolin, K.A., Schwalbe, H., MacArthur, M.W.,
Thorton, J.M. & Dobson, C.M. (1996) Analysis of main chain
torsion angles in proteins: prediction of NMR coupling cons-
tants for native and random coil conformations. J. Mol. Biol. 255,
494–506.
27. Wuthrich, K. (1986) NMR of Proteins and Nucleic Acids. Wiley,
New York.
28. Wishart, D., Sykes, B. & Richards, F. (1991) Relationship between
nuclear magnetic resonance chemical shift and protein secondary
structure. J. Mol. Biol. 222, 311–333.
29. Wishart, D., Sykes, B. & Richards, F. (1992) The chemical shift
index: a fast and simple method for the assignment of protein
secondary structure through NMR spectroscopy.Biochemistry 31,
1647–1651.
30. Merutka, G., Dyson, H.J. & Wright, P.E. (1995) �Random coil� 1Hchemical shifts obtained as a function of temperature and tri-
fluoroethanol concentration for the peptide Series GGXGG.
J. Biomol. NMR 5, 14–24.
31. Guntert, P., Mumenthaler, C. & Wuthrich, K. (1997) Torsion
angle dynamics for NMR structure calculation with the new
program DYANA. J. Mol. Biol. 273, 283–298.
32. Wuthrich, K., Billeter, M. & Brown, W. (1995) Pseudo-structures
for the 20 common amino acids for use in studies of protein
conformations by measurements of intramolecular proton-proton
distance constraints with nuclear magnetic resonance. J.Mol. Biol.
169, 949–961.
33. Pearlman, D.A., Case, D.A., Caldwell, J.W., Ross, W.S., Cheat-
ham, T.E., Ferguson, D.M., Seibel, G.L., Singh, U.C., Weiner,
6018 G. A. Spyroulias et al. (Eur. J. Biochem. 269) � FEBS 2002

P.K. & Kollman, P.A. (1997) AMBER 5.0. University of Cali-
fornia, San Francisco.
34. Pearlman, D.A., Case, D.A., Caldwell, G.C., Siebel, G.L., Singh,
U.C., Weiner, P. & Kollman, P.A. (1991) AMBER 4.0.University
of California, San Francisco.
35. Banci, L., Bertini, I., Luchinat, C. & Turano, P. (2000) Solution
Structures of Hemoproteins. InThe Porphyrin Handbook (Kadish,
K.M., Smith, K.M. & R.Guilard, R., eds), pp. 323–350. Academic
Press, San Diego.
36. Laskowski, R.A., MacArthur, M.W., Moss, D.S. & Thornton,
J.M. (1993) PROCHECK – a program to check the stereo-
chemical quality of protein structures. J. Appl. Crystallogr. 26,
283–291.
37. Laskowski, R.A., Rullmann, J.A.C., MacArthur, M.W., Kaptein,
R. & Thornton, J.M. (1996) AQUA and PROCHECK-NMR:
programs for checking the quality of protein structures solved by
NMR. J. Biomol. NMR 8, 477–486.
38. Dathe, M., Fabian, H., Gast, K., Zirwer, D., Winter, R., Beyer-
mann, M., Schumann, M. & Bienert, M. (1996) Conformational
differences of ovine and human Corticotropin releasing hormone.
Int. J. Peptide Protein Res. 47, 383–393.
39. Pallai, P.V., Mabilia, M., Goodman, M., Vale, W. & Rivier, J.
(1983) Structural homology of corticotropin-releasing factor,
sauvagine, and urotensin I: circular dichroism and prediction
studies. Proc. Natl Acad. Sci. USA 80, 6770–6774.
40. Goodman, M. & Listowsky, I. (1962) Conformational aspects of
synthetic polypeptides. VI. Hypochromic spectral studies of oligo-
c-methyl-L-glutamate peptides. J. Am. Chem. Soc. 84, 3770–3771.
41. Lau, S.H., Rivier, J., Vale, W., Kaiser, E.T. & Kezdy, F.J. (1983)
Surface properties of an amphiphilic peptide hormone and of its
analog: corticotropin-releasing factor and sauvagine. Proc. Natl
Acad. Sci. USA 80, 7070–7074.
42. Luo, P. & Baldwin, R. (1997) Mechanism of helix induction by
trifluoroethanol: a framework for extrapolating the helix-forming
properties of peptides from trifluoroethanol/water mixtures back
to water. Biochemistry 36, 8413–8421.
43. Cammers-Goodwin, A., Allent, T.J., Oslick, S.L., McClure, K.F.,
Lee, J.H. & Kemp, D.S. (1996) Mechanism of stabilization of
helical conformations of polypeptides by water containing tri-
fluoroethanol. J. Am. Chem. Soc. 118, 3082–3090.
44. Jaravine, V.A., Alexandrescu, A. & Grzesiek, S. (2001) Observa-
tion of the closing of individual hydrogen bonds during TFE-
induced helix formation in a peptide. Protein Sci. 10, 943–950.
45. Krauser, E., Bienert, M., Schmieder, P. & Wenschuh, H. (2000)
The helix-destabilizing propensity scale of D-amino acids: The
influence of side chain steric effects. J. Am. Chem. Soc. 122, 4865–
4870.
46. Rothemund, S., Beyermann, M., Krause, E., Krause, G., Bienert,
M., Hodges, R.S., Sykes, B.D. & Sonnichsen, F.D. (1995) Struc-
ture effects of double D-amino acid replacements: a nuclear mag-
netic and circular dichroism study using amphipathic model
helices. Biochemistry 34, 12954–12962.
47. Karle, I.L. & Balaram, P. (1990) Structural characteristics of
alpha-helical peptide molecules containing Aib residues. Bio-
chemistry 29, 6747–6756.
48. Toniolo, C. & Benedetti, E. (1991) Structures of polypeptides from
a-amino acids disubstituted at the a-carbon. Macromolecules 24,
4004–4009.
49. Toniolo, C. & Benedetti, E. (1991) The polypeptide 310-helix.
Trends Biochem. Sci. 16, 350–353.
50. Burgess, A.W. (1994) Designing amino acids to determine the
local conformations of peptides. Proc. Natl Acad. Sci. USA 91,
2649–2653.
51. Basu, G., Bagghi, K. & Kuki, A. (1991) Conformational pre-
ference of oligopeptides rich in a-aminoisobutyric acid. I. Obser-
vation of a 310/a-helical transition upon sequence permutation.
Biopolymers 31, 1763–1774.
52. Basu, G. & Kuki, A. (1992) Conformational preference of oligo-
peptides rich in a-aminoisobutyric acid. II. A model for the 310/a-
helix transition with composition and sequence sensitivity.
Biopolymers 32, 61–71.
53. Gibbs, N., Sessions, R.B., Williams, P.B. & Dempsey, C.E. (1997)
Helix bending in alamethicin: molecular dynamics simulations
and amide hydrogen exchange in methanol. Biophys. J. 72, 2490–
2495.
54. Kornreich, W.D., Galyean, R., Hernandez, J.F., Craig, A.G.,
Donaldson, C.J., Yamamoto, G., Rivier, C., Vale, W. & Rivier, J.
(1992) Alanine series of ovine corticotropin releasing factor
(oCRF): a structure-activity relationship study. J. Med. Chem. 35,
1870–1876.
55. Koradi, R., Billeter, M. & Wuthrich, K. (1996) MOLMOL: a
program for display and analysis of macromolecular structures.
J. Mol. Graph. 14, 51–55.
S U P P L E M E N T A R Y M A T E R I A L
The following material is available from http://www.blackwell-science.com/products/journals/suppmat/EJB/EJB3278/ EJB3278sm.htmTable S1. Chemical shifts (p.p.m) of the protons of the
residues in the [D-Phe12, Aib15]CRH.
Table S2. Experimental NOESY cross-peak intensities and
corresponding upper distance limits.
Fig. S1. (A) Rmsd per residue for the 40 structures for
backbone (black bars) and heavy atoms (grey bars), and (B) of
the mean structure.
Fig. S2. Data extracted from PROCHECK analysis.
� FEBS 2002 NMR structure of a CRH synthetic analogue in TFE/H2O (Eur. J. Biochem. 269) 6019




















