
1 1 1 ( )( )( )( ) a b c a b b c c a a b c a b c = − − − + +
Upload
independentCategory
view
1download
0
R
Tci
La
b
c
d
a
ARRAA
KTCLcEO
1
tatmtip
iltT
of
0d
Behavioural Brain Research 201 (2009) 198–206
Contents lists available at ScienceDirect
Behavioural Brain Research
journa l homepage: www.e lsev ier .com/ locate /bbr
esearch report
eneurin C-terminal associated peptide (TCAP)-1 attenuatesorticotropin-releasing factor (CRF)-induced c-Fos expressionn the limbic system and modulates anxiety behavior in male Wistar rats
aura A. Tana,c,∗, Karen Xua, Franco J. Vaccarinoa,b,c, David A. Lovejoyc, Susan Rotzingera,b,d
Department of Psychology, University of Toronto, Toronto, CanadaDepartment of Psychiatry, University of Toronto, Toronto, CanadaDepartment of Cell and Systems Biology, University of Toronto, Toronto, CanadaInstitute of Medical Science, University of Toronto, Toronto, Canada
r t i c l e i n f o
rticle history:eceived 9 October 2008eceived in revised form 4 February 2009ccepted 10 February 2009vailable online 21 February 2009
eywords:eneurinorticotropin-releasing factor
a b s t r a c t
The teneurin C-terminal associated peptides (TCAPs) are a novel family of four endogenous peptides thathave previously shown bioactive properties both in vitro and in vivo. Previously we have shown thatrepeated intracerebral injections of synthetic TCAP-1 modulate anxiety-like behaviors in three tests ofanxiety, although the neural substrates responsible for these effects were previously unknown. In thecurrent study, we examined both c-Fos induction and behavioral responses in the elevated plus mazeand open field tests after a single intracerebroventricular dose of TCAP-1 followed by an intracerebroven-tricular injection of CRF in male Wistar rats. The results indicate that TCAP-1 injection attenuated theCRF-induced increase in c-Fos expression in the limbic system and many of the areas associated with the
imbic system-Foslevated plus mazepen field
behavioral responses to stress, including the hippocampus, central and basolateral nuclei of the amygdala,medial prefrontal cortex, and dorsal raphe nucleus. Other areas, such as the paraventricular nucleus of thehypothalamus, bed nucleus of the stria terminalis, medial nucleus of the amygdala, and locus coeruleus,displayed CRF-induced c-Fos levels that were unaffected by TCAP-1. Furthermore, TCAP-1 administrationincreased stretched-attend postures, a type of risk-assessment behavior, on the elevated plus maze. Theseresults indicate that TCAP-1 may play a potential role in the regulation of stress by blocking CRF-mediated
sensi
activity in specific stress-. Introduction
Corticotropin-releasing factor (CRF) is a 41-amino acid peptidehat plays an integral role in activating the hypothalamic–pituitary–drenal (HPA) axis [31]. CRF is responsible for behavioral responseso stress, where CRF acts as a neurotransmitter/neuromodulator in
any areas of the brain associated with stress and anxiety, such ashe limbic system and the hypothalamus. Intracerebrally admin-stered CRF has been used to elicit many of the behavioral andhysiological responses associated with stress [4,15,16].
Recently, a new neuropeptide family with sequence similar-
ty to CRF was discovered [23]. These endogenous peptides areocated on the C-terminus of each of the type-II transmembraneeneurin proteins that are found in the central nervous system.hese novel peptides are 40 or 41 amino acids long and were named∗ Corresponding author at: Department of Cell and Systems Biology, Universityf Toronto, 25 St George Street, Toronto, ON M5S 3G5, Canada. Tel.: +1 416 978 5652ax: +1 416 971 3190.
E-mail address: [email protected] (L.A. Tan).
166-4328/$ – see front matter © 2009 Elsevier B.V. All rights reserved.oi:10.1016/j.bbr.2009.02.013
tive areas of the brain.© 2009 Elsevier B.V. All rights reserved.
the teneurin C-terminal associated peptides (TCAPs). The familyconsists of four members (TCAP 1–4) each with a cleavage andamidation motif, characteristics of a bioactive peptide [23]. TCAPis strongly conserved between members and shares about a 20%amino acid identity with CRF. TCAP mRNA is widely distributed inthe adult rat brain [33], including in the hippocampus, amygdala,ventromedial nucleus of the hypothalamus, and cerebellum.
Synthetic TCAP-1 induces a number of bioactive effects. TCAP-1increases neurite outgrowth and upregulates cytoskeletal proteinsin vitro in both immortalized neuron and primary hippocampal cul-tures [1]. This peptide exhibits a modulatory response on cAMPaccumulation and cell proliferation, increasing both at low concen-trations and decreasing both at high concentrations [33]. TCAP-1has modulatory effects on anxiety-related behaviors, as acute TCAP-1 administration into the basolateral amygdala in male Wistar ratsinduced changes in the acoustic startle response, such that animals
that displayed high emotionality prior to testing had a reduced star-tle response, whereas animals that displayed low emotionality priorto testing had an increased startle response [33]. Repeated intrac-erebroventricular (ICV) injections of TCAP-1 affect CRF-inducedanxiety behaviors, as TCAP-1 pre-treatment increased the anxiety
rain R
rd[fiWTC
botrctsstbwo[pTwesmt
2
2
Lutwatf
2
1UfsAr−scnd
2
2
n(hti
2
tIe3
T
Rats were injected as described above and returned to the colony after injections.
L.A. Tan et al. / Behavioural B
esponse to CRF in the elevated plus maze and open field tests, butecreased the anxiety response to CRF in the acoustic startle test30]. This difference may be due to the differing brain areas utilizedor each test, as the hippocampus and amygdala are differentiallynvolved in tasks of exploratory behavior and reflexive fear [17,18].
e have further shown that repeated intravenous injections ofCAP-1 attenuate the anxiety-inducing effects of ICV-administeredRF injections [2].
The mechanism by which TCAP-1 regulates CRF-mediatedehavior is not known. Therefore, we sought to determine the areasf the brain that are sensitive to TCAP-1, and to examine poten-ial interactions between TCAP-1 and CRF in the brain. The ICVoute of administration was used in this study as CRF does notross the blood–brain barrier. ICV-administered TCAP-1 was usedo simulate an acute increase in brain TCAP-1 during periods oftress. c-Fos is particularly useful because it has low basal expres-ion but is highly inducible and maximally expressed 2 h afterhe given stimuli, returning to basal levels after 4 h [3]. c-Fos haseen used to determine areas of the brain responsive to CRF [3,13],hich shows similar expression patterns in response to other types
f stress, including restraint, forced swim, and immune challenge5,6,25]. ICV-administered CRF was used in the current study as aositive control, and also to assess the effects of ICV-administeredCAP-1 on CRF-induced c-Fos expression. Therefore, we treated ratsith TCAP-1 and CRF to determine if TCAP-1 could modulate the
xpected CRF-induced c-Fos expression. Furthermore, we used theame treatment paradigm in two tests of anxiety, the elevated plusaze and the open field test, to determine if acute TCAP-1 can alter
he behavioral response to CRF.
. Methods
.1. Animals
Male Wistar rats (n = 40) weighing 250–275 g were obtained from Charles Riveraboratories (Montreal, Canada). Rats were singly housed in Plexiglas shoebox cagesnder standard laboratory conditions (12:12 h light:dark cycle, lights on at 0700 h,emperature 21 ± 1 ◦C) with food and water available ad libitum. Rats were given oneeek to acclimatize to laboratory conditions before surgery. All procedures were
pproved by the University of Toronto Animal Care Committee in accordance withhe Canadian Council on Animal Care. Additional male Wistar rats (n = 72) were usedor behavioral testing, and were not used in the immunohistochemistry experiments.
.2. Surgery
All rats were anesthetized with isoflurane (3% induction, 2–3% maintenance in00% O2) and fit into a stereotaxic apparatus (David Kopf Instruments, Tujunga, CA,SA). Analgesics (0.5 mg/kg buprenorphine for c-Fos animals; 2.5 mg/kg ketaprofen
or behavioral test animals) were administered subcutaneously at the beginning ofurgery. A midline incision was made along the top of the head, exposing bregma.22-gauge guide cannula (Plastics One, Roanoke, VA, USA) was implanted into the
ight lateral ventricle using the following flat-skull coordinates: AP −1.0, ML −1.4, DV2.7 from dura [22]. The guide cannula was secured to the skull using four jewellers’
crews and dental cement. The opening of the cannula was covered by a removableannula dummy. Animals were kept under warm lamps until regaining conscious-ess and were given one week to recover from surgery. Rats were thoroughly handledaily to acclimatize individuals to the injection procedure and human contact.
.3. C-Fos immunohistochemistry
.3.1. DrugsSynthetic mouse TCAP-1 was synthesized by American Peptide Company (Sun-
yvale, CA). TCAP-1 stocks were prepared to a concentration of 2.1 × 10−4 Mequivalent to 1 �g/�l TCAP-1) by treating lyophilized TCAP-1 with ammoniumydroxide vapour, then dissolving in sterile saline. The TCAP-1 dose was based onhe molar equivalent of 3 �g of CRF. CRF (Sigma–Aldrich, Oakville, ON) was dissolvedn sterile saline at a concentration of 1 �g/�l.
.3.2. InjectionsInjections were given between 0800 h and 1000 h. Intracerebroventricular injec-
ions were delivered at a rate of 2 �l/min by a syringe pump (Razel Scientificnstrument Inc., Stamford, USA) connected to a 28-gauge stainless steel injector thatxtended 1 mm below the cannula guide tip. Animals were injected first with either�l of sterile saline (vehicle) or 3 �g of TCAP-1. The injector was left in place for
esearch 201 (2009) 198–206 199
60 s following infusion to prevent backflow up the cannula. Afterwards, the injectorwas removed, and a second injector was inserted into the cannula. An infusion ofeither 1 �l of saline or 1 �l of 1 �g/�l CRF was injected. There were four treatmentgroups, each with a total volume of 4 �l: saline + saline (“SAL”), saline + CRF (“CRF”),TCAP-1 + saline (“TCAP”), and TCAP-1 + CRF (“TCAP + CRF”).
2.3.3. Tissue processingAfter injections, rats were returned to their home cages and the colony for 2 h.
After that period, rats were removed from the colony and overdosed with isofluraneand decapitated. The brains were removed within 90 s, rinsed in PBS, and snap-frozen in −70 ◦C isopentane. The brains were then stored at −80 ◦C until they couldbe processed. Brains were sectioned in a 1-in-6 series on a cryostat at a thicknessof 25 �m. Sections were thaw-mounted onto Superfrost slides (Fisher Scientific,Canada) and allowed to dry overnight before being stored at −80 ◦C until processing.
2.3.4. ImmunohistochemistryImmunohistochemical protocols were adapted from Sundquist and Nisenbaum
[29]. All wash steps were performed with Tris-buffered saline with Tween-20 (TBS-; 50 mM Tris–HCl, 150 mM NaCl, 0.05% Tween-20, pH 7.6). Antibodies were diluted
in TBS-T with 1% weight by volume IgG-free bovine serum albumen fraction V(Jackson ImmunoResearch, Burlington, ON). Slides were immersed in 4% phosphate-buffered paraformaldehyde for 10 min. Endogenous peroxidases were quenchedwith a 1% hydrogen peroxide/methanol solution for 15 min. Endogenous proteinswere blocked with 10% normal goat serum (Vector Laboratories). The sections wereincubated with rabbit anti-c-Fos IgG (1:2000; sc-52, Santa Cruz Biotechnology, SantaCruz, CA, USA) for 2 h in a standard humid chamber, then incubated in biotiny-lated goat anti-rabbit IgG (1:200; BA-1000, Vector Laboratories) for 30 min. Thesections were reacted with an avidin–biotin–peroxidase complex (Vectastain ABCElite Kit, Vector Laboratories) for 30 min according to kit instructions. Subsequently,the sections were visualized using diaminobenzidine chromagen (DAB Substrate Kit,Vector Laboratories). The sections were dehydrated using graded ethanols, clearedin xylene, and coverslipped with Permount (Fisher Scientific). An adjoining series ofsections were stained using cresyl violet for reference purposes.
2.3.5. AnalysisAreas of the brain that are associated with the stress response and demonstrate
high c-Fos activity in response to CRF were investigated. This included componentsof the limbic system (amygdala and hippocampus) and other areas linked to boththe limbic system and the regulation of stress, including the medial prefrontal cortex(infralimbic and prelimbic cortices), paraventricular nucleus of the hypothalamus,septum (medial and lateral), bed nucleus of the stria terminalis, locus coeruleus, andthe dorsal raphe nucleus.
Sections were scanned for c-Fos activity using a Nikon Optiphot microscope(Nikon Canada, Mississauga, ON). c-Fos-positive cells were manually counted foreach area using NIS Elements Software Basic v2.10 (Nikon Canada, Mississauga, ON).Cumulative counts through the entire series for each region of interest were obtainedfor each brain. The observer was blind to the study.
Statistical analysis was performed using GraphPad Prism 4.0. Data was ana-lyzed using a two-way ANOVA with “TCAP-1” and “CRF” as factors, followed byBonferonni’s post hoc test.
2.4. Behavioral tests
2.4.1. DrugsSynthetic mouse TCAP-1 (1 �g/�l) was synthesized and prepared for injection
as previously described above. The 1 �g CRF dose (Sigma–Aldrich, Oakville, ON) wasdissolved in sterile saline at a concentration of 0.33 �g/�l, while the 3 �g dose wasdissolved in sterile saline at a concentration of 1 �g/�l.
2.4.2. InjectionsInjections were administered as outlined above and were given between 0900 h
and 1300 h. Animals were injected first with either 3 �l of sterile saline (vehicle) or3 �g of TCAP-1. The injector was left in place for 60 s following infusion to pre-vent backflow up the cannula. The injector was removed, and a second injectorwas inserted into the cannula. An infusion of either 3 �l of saline (vehicle), 3 �lof 0.33 �g/�l CRF (for 1 �g CRF dose) or 3 �l of 1 �g/�l CRF (for 3 �g CRF dose)was injected. There were 6 treatment groups, each with a total injection volumeof 6 �l: saline + saline (“SAL”), saline + CRF (1 �g) (“CRF (1 �g)”), saline + CRF (3 �g)(“CRF (3 �g)”), TCAP-1 + saline (“TCAP”), TCAP-1 + CRF (1 �g) (“TCAP + CRF (1 �g)”),and TCAP-1 + CRF (3 �g) (“TCAP + CRF (3 �g)”).
2.4.3. Elevated plus maze
Two hours later, rats were tested on the elevated plus maze. The maze consistedof four 50 cm × 10 cm arms extending from a 10 cm × 10 cm centre platform. Twoclosed arms situated 180◦ from each other were enclosed on three sides by 40 cmhigh black Plexiglas walls with the openings facing the centre platform. The twoother open arms were not enclosed. The maze was elevated 65 cm off the floor by asingle post extending from the centre platform. The entire maze was painted black.

2 rain R
tmr(–t
2
fitcbwTt“Ot
2
tth
3
3
Tw
F(TdaC
00 L.A. Tan et al. / Behavioural B
Rats were placed on the maze facing one of the open arms, and were allowedo explore for 5 min under low-light conditions (two 15-W white light bulbs). The
aze was cleaned between each test with mild soap and water. Movement of theats was recorded from above and quantified using Ethovision proprietary softwareNoldus Information Technology, Utrecht, Netherlands). Other behavioral measuresrearing and stretched-attend postures – were manually scored by observers blind
o the study.
.4.4. Open field testThree days after testing on the elevated plus maze, rats were tested in the open
eld. Rats were injected as described above and returned to the colony after injec-ions. Two hours later, rats were tested in the open field test. This test apparatusonsisted of a 50 cm × 50 cm arena with walls 40 cm high made of black particleoard. Tests were performed under lower light conditions (two 100-W bulbs). Ratsere gently placed in the middle of the arena and allowed to explore for 10 min.
he maze was cleaned between each test with mild soap and water. Movement ofhe rats was recorded from above and quantified using Ethovision software, and theCentre” of the maze was defined as a 30 cm × 30 cm square in the centre of the maze.ne other behavioral measure, rearing, was manually scored by observers blind to
he study.
.4.5. AnalysisRaw data from Ethovision was transferred to Microsoft Excel for sorting. Statis-
ical analysis of data was performed using Graphpad Prism 4.0, and analyzed using awo-way ANOVA with “TCAP-1” and “CRF” as factors, followed by Bonferonni’s postoc test.
. Results
.1. Immunohistochemistry
Data from the saline (n = 9), CRF (n = 9), TCAP (n = 9), andCAP + CRF (n = 7) groups were analyzed using a two-way ANOVAith Bonferroni’s post hoc test. Saline produced a relatively low c-
ig. 1. Representative images of 25 �m coronal rat brain sections from they hippocampA–D), CA2 (E–H), CA3 (I–L) and the dentate gyrus (DG; M–P). Rats were cannulated intoCAP-1 and saline (TCAP; B, F, J, and N), saline and 1 �g of CRF (CRF; C, G, K, and O), or 3ifference in c-Fos immunoreactivity between TCAP-1-only treated rats and saline treatll regions of the hippocampus that was measured (p < 0.001). Rats treated with the comRF-only treated animals in the CA1 (p < 0.01), CA2 (p < 0.05), CA3 (p < 0.001), and the DG
esearch 201 (2009) 198–206
Fos activity, whereas CRF elicited high c-Fos immunoreactivity inareas associated with the stress response, including the hippocam-pus (Fig. 1), amygdala, medial prefrontal cortex (Fig. 2), dorsalraphe nuclei and paraventricular nucleus of the hypothalamus (notshown).
In the hippocampus, the two-way ANOVA revealed that therewas a significant main effect of CRF on c-Fos expression in theCA1 (Fig. 3A; F(1) = 17.03, p = 0.0003), CA2 (Fig. 3B; F(1) = 19.82,p = 0.0001), and CA3 (Fig. 3C; F(1) = 25.00, p < 0.0001) subfields, aswell as in the dentate gyrus (Fig. 3D; F(1) = 33.52, p < 0.0001). Therewas a significant main effect of TCAP-1 on c-Fos expression in theCA1 (F(1) = 4.655, p = 0.0391) and CA3 (F(1) = 9.943, p = 0.0037) sub-fields as well as in the dentate gyrus (F(1) = 5.671, p = 0.024). Aswell, there was a significant interaction between CRF and TCAP-1 in the CA1 (F(1) = 5.834, p = 0.022), CA2 (F(1) = 7.276, p = 0.0114),and CA3 (F(1) = 13.77, p = 0.0008) subfields of the hippocampus, aswell as in the dentate gyrus (F(1) = 8.32, p = 0.0073). Bonferroni’spost hoc test revealed that CRF significantly increased c-Fos expres-sion relative to saline in the CA1, CA2, and CA3 subfields of thehippocampus and the dentate gyrus (p < 0.001 for all). Treatmentwith TCAP-1 in CRF-treated animals significantly reduced c-Fosexpression relative to CRF-only treated animals in the CA1 (p < 0.05),CA2 (p < 0.05), and CA3 (p < 0.001) subfields, and the dentate gyrus(p < 0.01).
In the amygdala, two-way ANOVA revealed that CRF had a
significant main effect of increasing c-Fos expression in the cen-tral nucleus (CeA, Fig. 4A; F(1) = 13.89, p = 0.0008) and basolateralnucleus (BLA, Fig. 4B; F(1) = 17.16, p = 0.0003). Bonferroni’s post hoctest revealed that treatment of CRF significantly increased c-Fosus showing c-Fos immunoreactivity in the subfields of the hippocampus: the CA1the lateral ventricle and treated with either saline only (SAL; A, E, I, and M), 3 �g of�g of TCAP-1 and 1 �g of CRF (TCAP + CRF; D, H, L, and P). There was no significanted rats (p > 0.05). CRF-treated rats had significantly increased c-Fos expression inbination of TCAP-1 and CRF had significantly reduced c-Fos expression relative to(p < 0.01). Scale bar = 100 �m.

L.A. Tan et al. / Behavioural Brain Research 201 (2009) 198–206 201
Fig. 2. Representative images of 25 �m coronal rat brain sections from the central nucleus of the amygdala (CeA; A–D), the basolateral nucleus of the amygdala (BLA; E–H), andthe medial prefrontal cortex (mPFC; I–L). Rats were cannulated into the lateral ventricle and treated with either saline only (SAL; A, E, and I), 3 �g of TCAP-1 and saline (TCAP;B, F, and J), saline and 1 �g of CRF (CRF; C, G, and K), or 3 �g of TCAP-1 and 1 �g of CRF (TCAP + CRF; D, H, and L). There was no significant difference in c-Fos immunoreactivitybetween TCAP-1-only treated rats and saline-treated rats (p > 0.05). CRF-treated rats displayed significantly greater c-Fos immunoreactivity relative to saline-treated rats inthe CeA (p < 0.01), BLA (p < 0.001) and mPFC (p < 0.01). Co-treatment of TCAP-1 and CRF attenuated c-Fos expression, such that c-Fos expression in rats co-treated with TCAP-1and CRF no longer differed from CRF-only treated rats. Scale bar = 100 �m.
Fig. 3. Cumulative c-Fos immunoreactivity counts in the subfields of the hippocampus: (A) the CA1, (B) CA2, (C) CA3, and (D) dentate gyrus (DG). Two-way ANOVA revealed asignificant main effect of CRF in the CA1 (p = 0.0003), CA2 (p = 0.0001), CA3 (p < 0.0001), and the DG (p < 0.0001), and a significant main effect of TCAP-1 in the CA1 (p = 0.0391),CA3 (p = 0.0037), and DG (p = 0.024). Furthermore, there was an antagonistic interaction between CRF and TCAP-1 in the CA1 (p = 0.022), CA2 (p = 0.0114), CA3 (p = 0.0008),and DG (p = 0.0073). Bonferonni’s post hoc test revealed that CRF significantly increased c-Fos expression relative to saline in all four areas, and co-treatment of TCAP-1 andCRF significantly reduced c-Fos expression relative to CRF-only treated animals in the hippocampus. Values represent means + SEM (*p < 0.05, **p < 0.01, ***p < 0.001).
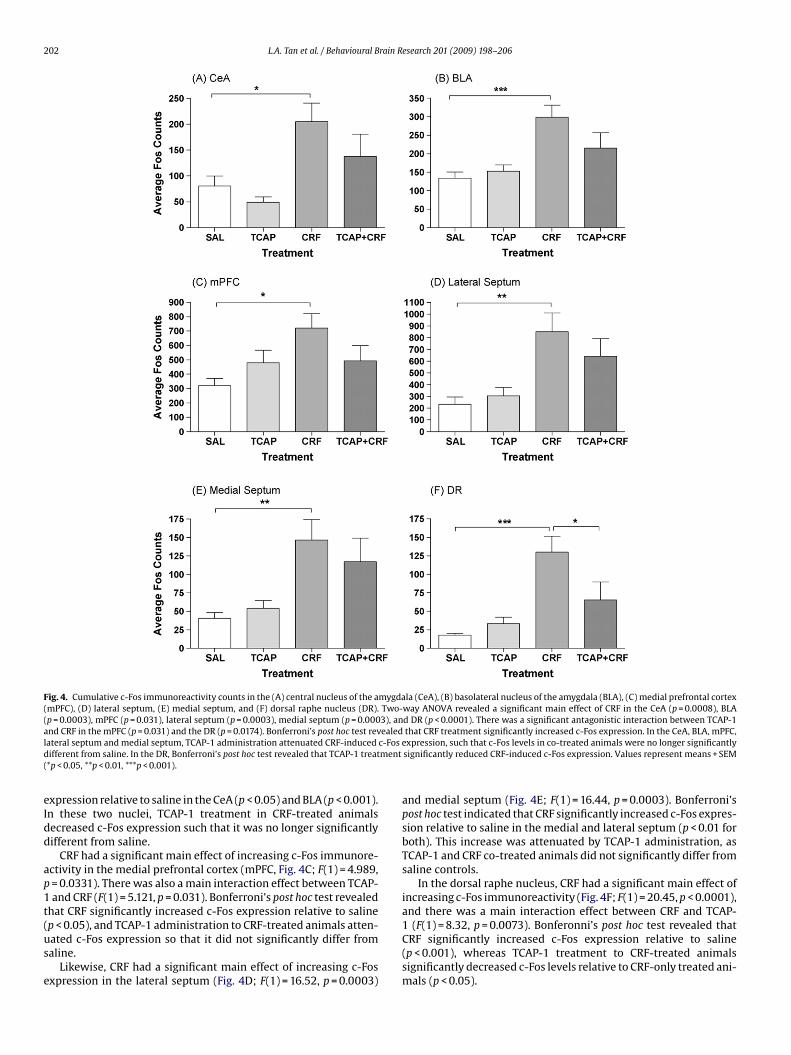
202 L.A. Tan et al. / Behavioural Brain Research 201 (2009) 198–206
Fig. 4. Cumulative c-Fos immunoreactivity counts in the (A) central nucleus of the amygdala (CeA), (B) basolateral nucleus of the amygdala (BLA), (C) medial prefrontal cortex(mPFC), (D) lateral septum, (E) medial septum, and (F) dorsal raphe nucleus (DR). Two-way ANOVA revealed a significant main effect of CRF in the CeA (p = 0.0008), BLA(p = 0.0003), mPFC (p = 0.031), lateral septum (p = 0.0003), medial septum (p = 0.0003), and DR (p < 0.0001). There was a significant antagonistic interaction between TCAP-1a ealedl c-Fosd ment(
eIdd
ap1t(us
e
nd CRF in the mPFC (p = 0.031) and the DR (p = 0.0174). Bonferroni’s post hoc test revateral septum and medial septum, TCAP-1 administration attenuated CRF-inducedifferent from saline. In the DR, Bonferroni’s post hoc test revealed that TCAP-1 treat*p < 0.05, **p < 0.01, ***p < 0.001).
xpression relative to saline in the CeA (p < 0.05) and BLA (p < 0.001).n these two nuclei, TCAP-1 treatment in CRF-treated animalsecreased c-Fos expression such that it was no longer significantlyifferent from saline.
CRF had a significant main effect of increasing c-Fos immunore-ctivity in the medial prefrontal cortex (mPFC, Fig. 4C; F(1) = 4.989,= 0.0331). There was also a main interaction effect between TCAP-and CRF (F(1) = 5.121, p = 0.031). Bonferroni’s post hoc test revealed
hat CRF significantly increased c-Fos expression relative to saline
p < 0.05), and TCAP-1 administration to CRF-treated animals atten-ated c-Fos expression so that it did not significantly differ fromaline.Likewise, CRF had a significant main effect of increasing c-Fosxpression in the lateral septum (Fig. 4D; F(1) = 16.52, p = 0.0003)
that CRF treatment significantly increased c-Fos expression. In the CeA, BLA, mPFC,expression, such that c-Fos levels in co-treated animals were no longer significantlysignificantly reduced CRF-induced c-Fos expression. Values represent means + SEM
and medial septum (Fig. 4E; F(1) = 16.44, p = 0.0003). Bonferroni’spost hoc test indicated that CRF significantly increased c-Fos expres-sion relative to saline in the medial and lateral septum (p < 0.01 forboth). This increase was attenuated by TCAP-1 administration, asTCAP-1 and CRF co-treated animals did not significantly differ fromsaline controls.
In the dorsal raphe nucleus, CRF had a significant main effect ofincreasing c-Fos immunoreactivity (Fig. 4F; F(1) = 20.45, p < 0.0001),and there was a main interaction effect between CRF and TCAP-
1 (F(1) = 8.32, p = 0.0073). Bonferonni’s post hoc test revealed thatCRF significantly increased c-Fos expression relative to saline(p < 0.001), whereas TCAP-1 treatment to CRF-treated animalssignificantly decreased c-Fos levels relative to CRF-only treated ani-mals (p < 0.05).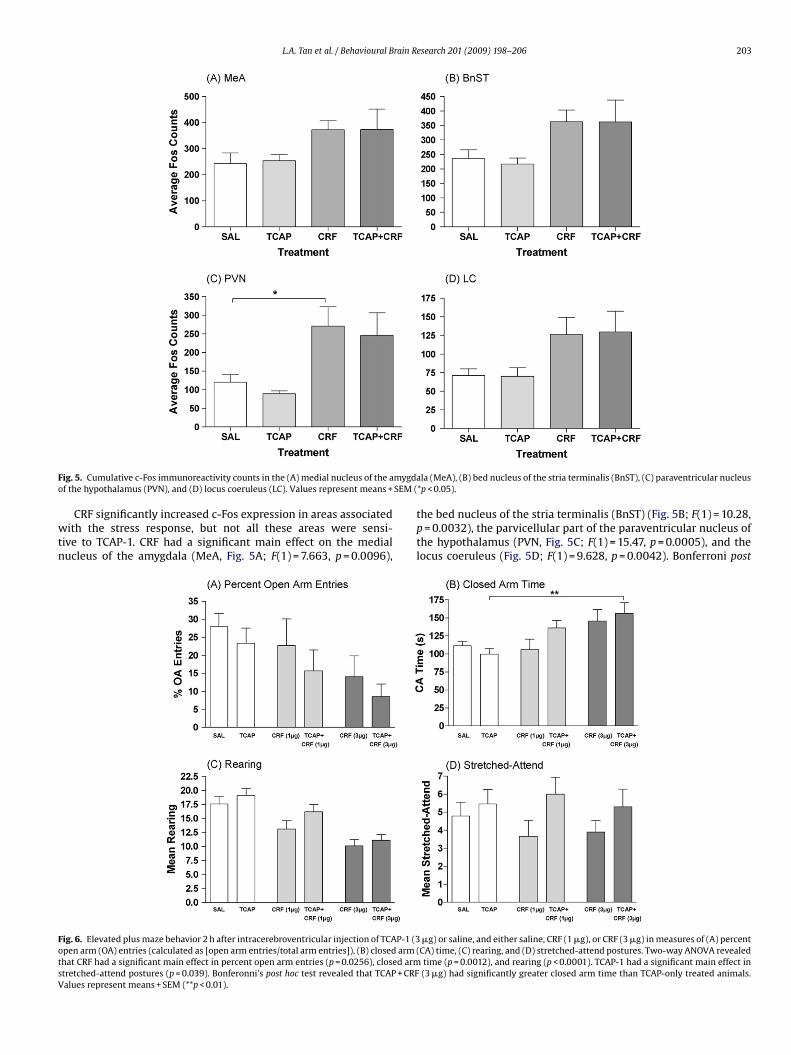
L.A. Tan et al. / Behavioural Brain Research 201 (2009) 198–206 203
F mygdao SEM (
wtn
FotsV
ig. 5. Cumulative c-Fos immunoreactivity counts in the (A) medial nucleus of the af the hypothalamus (PVN), and (D) locus coeruleus (LC). Values represent means +
CRF significantly increased c-Fos expression in areas associatedith the stress response, but not all these areas were sensi-
ive to TCAP-1. CRF had a significant main effect on the medialucleus of the amygdala (MeA, Fig. 5A; F(1) = 7.663, p = 0.0096),
ig. 6. Elevated plus maze behavior 2 h after intracerebroventricular injection of TCAP-1 (3pen arm (OA) entries (calculated as [open arm entries/total arm entries]), (B) closed armhat CRF had a significant main effect in percent open arm entries (p = 0.0256), closed armtretched-attend postures (p = 0.039). Bonferonni’s post hoc test revealed that TCAP + CRFalues represent means + SEM (**p < 0.01).
la (MeA), (B) bed nucleus of the stria terminalis (BnST), (C) paraventricular nucleus*p < 0.05).
the bed nucleus of the stria terminalis (BnST) (Fig. 5B; F(1) = 10.28,p = 0.0032), the parvicellular part of the paraventricular nucleus ofthe hypothalamus (PVN, Fig. 5C; F(1) = 15.47, p = 0.0005), and thelocus coeruleus (Fig. 5D; F(1) = 9.628, p = 0.0042). Bonferroni post
�g) or saline, and either saline, CRF (1 �g), or CRF (3 �g) in measures of (A) percent(CA) time, (C) rearing, and (D) stretched-attend postures. Two-way ANOVA revealed
time (p = 0.0012), and rearing (p < 0.0001). TCAP-1 had a significant main effect in(3 �g) had significantly greater closed arm time than TCAP-only treated animals.
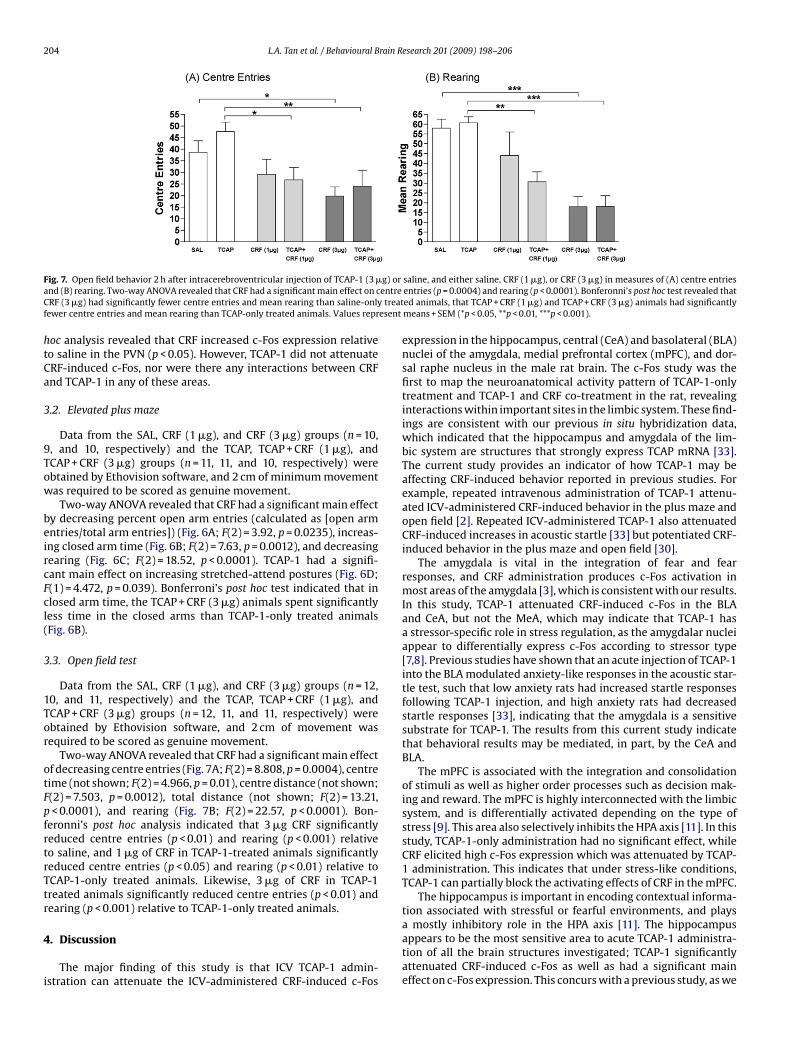
204 L.A. Tan et al. / Behavioural Brain Research 201 (2009) 198–206
F g) ora entreC treatf esent
htCa
3
9Tow
beircFcl(
3
1Tor
otFpfrtrTtr
4
i
ig. 7. Open field behavior 2 h after intracerebroventricular injection of TCAP-1 (3 �nd (B) rearing. Two-way ANOVA revealed that CRF had a significant main effect on cRF (3 �g) had significantly fewer centre entries and mean rearing than saline-only
ewer centre entries and mean rearing than TCAP-only treated animals. Values repr
oc analysis revealed that CRF increased c-Fos expression relativeo saline in the PVN (p < 0.05). However, TCAP-1 did not attenuateRF-induced c-Fos, nor were there any interactions between CRFnd TCAP-1 in any of these areas.
.2. Elevated plus maze
Data from the SAL, CRF (1 �g), and CRF (3 �g) groups (n = 10,, and 10, respectively) and the TCAP, TCAP + CRF (1 �g), andCAP + CRF (3 �g) groups (n = 11, 11, and 10, respectively) werebtained by Ethovision software, and 2 cm of minimum movementas required to be scored as genuine movement.
Two-way ANOVA revealed that CRF had a significant main effecty decreasing percent open arm entries (calculated as [open armntries/total arm entries]) (Fig. 6A; F(2) = 3.92, p = 0.0235), increas-ng closed arm time (Fig. 6B; F(2) = 7.63, p = 0.0012), and decreasingearing (Fig. 6C; F(2) = 18.52, p < 0.0001). TCAP-1 had a signifi-ant main effect on increasing stretched-attend postures (Fig. 6D;(1) = 4.472, p = 0.039). Bonferroni’s post hoc test indicated that inlosed arm time, the TCAP + CRF (3 �g) animals spent significantlyess time in the closed arms than TCAP-1-only treated animalsFig. 6B).
.3. Open field test
Data from the SAL, CRF (1 �g), and CRF (3 �g) groups (n = 12,0, and 11, respectively) and the TCAP, TCAP + CRF (1 �g), andCAP + CRF (3 �g) groups (n = 12, 11, and 11, respectively) werebtained by Ethovision software, and 2 cm of movement wasequired to be scored as genuine movement.
Two-way ANOVA revealed that CRF had a significant main effectf decreasing centre entries (Fig. 7A; F(2) = 8.808, p = 0.0004), centreime (not shown; F(2) = 4.966, p = 0.01), centre distance (not shown;(2) = 7.503, p = 0.0012), total distance (not shown; F(2) = 13.21,< 0.0001), and rearing (Fig. 7B; F(2) = 22.57, p < 0.0001). Bon-
eronni’s post hoc analysis indicated that 3 �g CRF significantlyeduced centre entries (p < 0.01) and rearing (p < 0.001) relativeo saline, and 1 �g of CRF in TCAP-1-treated animals significantlyeduced centre entries (p < 0.05) and rearing (p < 0.01) relative toCAP-1-only treated animals. Likewise, 3 �g of CRF in TCAP-1reated animals significantly reduced centre entries (p < 0.01) andearing (p < 0.001) relative to TCAP-1-only treated animals.
. Discussion
The major finding of this study is that ICV TCAP-1 admin-stration can attenuate the ICV-administered CRF-induced c-Fos
saline, and either saline, CRF (1 �g), or CRF (3 �g) in measures of (A) centre entriesentries (p = 0.0004) and rearing (p < 0.0001). Bonferonni’s post hoc test revealed thated animals, that TCAP + CRF (1 �g) and TCAP + CRF (3 �g) animals had significantlymeans + SEM (*p < 0.05, **p < 0.01, ***p < 0.001).
expression in the hippocampus, central (CeA) and basolateral (BLA)nuclei of the amygdala, medial prefrontal cortex (mPFC), and dor-sal raphe nucleus in the male rat brain. The c-Fos study was thefirst to map the neuroanatomical activity pattern of TCAP-1-onlytreatment and TCAP-1 and CRF co-treatment in the rat, revealinginteractions within important sites in the limbic system. These find-ings are consistent with our previous in situ hybridization data,which indicated that the hippocampus and amygdala of the lim-bic system are structures that strongly express TCAP mRNA [33].The current study provides an indicator of how TCAP-1 may beaffecting CRF-induced behavior reported in previous studies. Forexample, repeated intravenous administration of TCAP-1 attenu-ated ICV-administered CRF-induced behavior in the plus maze andopen field [2]. Repeated ICV-administered TCAP-1 also attenuatedCRF-induced increases in acoustic startle [33] but potentiated CRF-induced behavior in the plus maze and open field [30].
The amygdala is vital in the integration of fear and fearresponses, and CRF administration produces c-Fos activation inmost areas of the amygdala [3], which is consistent with our results.In this study, TCAP-1 attenuated CRF-induced c-Fos in the BLAand CeA, but not the MeA, which may indicate that TCAP-1 hasa stressor-specific role in stress regulation, as the amygdalar nucleiappear to differentially express c-Fos according to stressor type[7,8]. Previous studies have shown that an acute injection of TCAP-1into the BLA modulated anxiety-like responses in the acoustic star-tle test, such that low anxiety rats had increased startle responsesfollowing TCAP-1 injection, and high anxiety rats had decreasedstartle responses [33], indicating that the amygdala is a sensitivesubstrate for TCAP-1. The results from this current study indicatethat behavioral results may be mediated, in part, by the CeA andBLA.
The mPFC is associated with the integration and consolidationof stimuli as well as higher order processes such as decision mak-ing and reward. The mPFC is highly interconnected with the limbicsystem, and is differentially activated depending on the type ofstress [9]. This area also selectively inhibits the HPA axis [11]. In thisstudy, TCAP-1-only administration had no significant effect, whileCRF elicited high c-Fos expression which was attenuated by TCAP-1 administration. This indicates that under stress-like conditions,TCAP-1 can partially block the activating effects of CRF in the mPFC.
The hippocampus is important in encoding contextual informa-tion associated with stressful or fearful environments, and plays
a mostly inhibitory role in the HPA axis [11]. The hippocampusappears to be the most sensitive area to acute TCAP-1 administra-tion of all the brain structures investigated; TCAP-1 significantlyattenuated CRF-induced c-Fos as well as had a significant maineffect on c-Fos expression. This concurs with a previous study, as we
rain R
hTesia
tTapCTa
snpsTcaititalfnma
THpiccwtbtaib
sT1Wodtdciop(teptco
L.A. Tan et al. / Behavioural B
ave previously shown that 10 days of repeated ICV-administeredCAP-1 produces changes in CA3 neuron morphology (Al Chawaft al., manuscript in preparation). TCAP-1 appears to exert bothhort-term and long-term changes on this vital limbic structure,ndicating that the hippocampus is an important site for TCAP-1ction.
The sensitivity of the dorsal raphe nucleus to TCAP-1 suggestshat the serotonergic system may play a role in TCAP-1 signalling.his nucleus has serotonergic neurons that express high c-Fosctivity during stress [24] and has connections to the mPFC, hip-ocampus, and amygdala [20,32]. TCAP-1 significantly reducedRF-induced c-Fos in the dorsal raphe nucleus, indicating thatCAP-1 may be affecting serotonin pathways to other brain areasssociated with the modulation of the stress response.
Several regions of the brain that play a significant role in CRFignalling were not affected by TCAP-1 administration. TCAP-1 didot attenuate CRF-induced c-Fos in the parvicellular part of thearaventricular nucleus of the hypothalamus (PVN), which is theource of CRF to the hypothalamic–pituitary–adrenal (HPA) axis.his indicates that TCAP-1 may act independently of the classi-al HPA stress pathway. These results agree with corticosteronessays performed in a previous study [2], which showed that acutentravenous TCAP-1 did not attenuate intravenous CRF-induced cor-icosterone secretion. TCAP-1 did not attenuate CRF-induced c-Fosn the bed nucleus of the stria terminalis (BnST). This concurs withhe lack of effect of TCAP-1 on CRF-induced c-Fos seen in the PVN,s the BnST is a major relay station for communication between theimbic system and the PVN [12], and the PVN appears to be unaf-ected by acute TCAP-1. Likewise, the locus coeruleus, an area of highoradrenergic and behaviorally activating input into the hypothala-us and rest of the brain, showed no difference between CRF-only
nd animals co-treated with CRF and TCAP-1.rce of CRF to the hypothalamic–pituitary–adrenal (HPA) axis.
his indicates that TCAP-1 may act independently of the classicalPA stress pathway. These results agree with corticosterone assayserformed in a previous study [2], which showed that acute
ntravenous TCAP-1 did not attenuate intravenous CRF-inducedorticosterone secretion. TCAP-1 did not attenuate CRF-induced-Fos in the bed nucleus of the stria terminalis (BnST). This concursith the lack of effect of TCAP-1 on CRF-induced c-Fos seen in
he PVN, as the BnST is a major relay station for communicationetween the limbic system and the PVN [12], and the PVN appearso be unaffected by acute TCAP-1. Likewise, the locus coeruleus,n area of high noradrenergic and behaviorally activating inputnto the hypothalamus and rest of the brain, showed no differenceetween CRF-only and animals co-treated with CRF and TCAP-1.
When administered in the absence of CRF, TCAP-1 did not have aignificant effect on c-Fos expression relative to saline-treated rats.his is consistent with previous behavioral studies [2,30], as TCAP-has not elicited behavioral effects in the absence of a stressor.hile this result may suggest a direct or indirect role as an antag-
nist of CRF receptors, current in vitro data indicate that TCAP-1oes not bind or modify the activity of either CRF1 or CRF2 recep-ors (Barsyte, Nock, and Lovejoy, manuscript in preparation), noroes intravenously administered TCAP-1 affect CRF-induced corti-osterone secretion [2]. Therefore, it is possible that TCAP-1 may benteracting with elements of the CRF signalling pathway upstreamf c-Fos without affecting the CRF receptors themselves. For exam-le, previous in vitro studies have shown that high concentrations100 nM) of TCAP-1 can inhibit forskolin-induced cAMP accumula-ion [33] in immortalized hypothalamic neurons that do not possess
ither CRF receptor. This suggests that TCAP-1 on its own mayroduce a state that is only visible at times of neuronal activa-ion, and may explain why there were no significant changes of-Fos expression or behavior due to TCAP-1 administration on itswn.esearch 201 (2009) 198–206 205
The elevated plus maze and open field tests have been used inthe past to screen potential anxiolytic or anxiogenic drugs. In theelevated plus maze, more anxious rats will exhibit decreased openarm time and entries, preferring to stay within the confines of theclosed arms. More anxious rats will also display decreased rearing(standing up on hind legs) and increased stretched-attend postures(the rat stretching forward to full length with the back paws sta-tionary) [10,27]. In the open field test, more anxious rats will exhibitdecreased centre time and entries, and will instead remain in theperimeter of the maze [19]. The results from the elevated plus mazeindicated that, as expected, CRF had anxiogenic effects on tradi-tional measures of anxiety behavior, namely by decreasing percentof open arm entries, closed arm time, and rearing. However, CRFhad no effect on stretched-attend postures, which have been shownto increase in response to stressors [14,26]. TCAP-1 administrationproduced a significant main effect on stretched-attend postures,although post hoc tests did not reveal any significant differencesbetween TCAP-1 treated groups and their controls. In the open field,TCAP-1 had no significant effects when administered on its own,while CRF significantly increased anxiety-like behaviors as indi-cated by less time in and making fewer entries into the centre ofthe maze. TCAP-1 did not appear to have any significant effects inaltering CRF-induced anxiety behaviors.
TCAP-1 has been shown to have behavioral effects when admin-istered acutely into the BLA [33], such that high-emotionality ratsshowed decreased anxiety-like responses and low-emotionalityrats showed increased anxiety-like responses in the acoustic star-tle test, but the effects seen in our acute experiments are not aspronounced as we had expected. It is possible that the time courseused in this study was not appropriate to see behavioral modifica-tions to CRF-induced exploratory behavior. The anxiogenic effectsof CRF are still present 2 h after ICV administration [28], but otherpeptides may have shorter windows with which to see effects. Forexample, low doses of neuropeptide Y (NPY) in mice have anxio-genic effects on the elevated plus maze, but such effects are evident10 min after injection and are not detected 30 min after ICV injec-tion [21]. Therefore, although 2 h was sufficient to see the effectsof TCAP-1 on CRF-induced c-Fos, the behavioral effects of reducedc-Fos in specific areas of the brain may have already peaked andsubsided prior to testing.
In summary, acute intracerebral injections of TCAP-1 was able toattenuate the CRF-induced upregulation of c-Fos in specific areasof the brain associated with the extrahypothalamic stress circuit,particularly the limbic system and the dorsal raphe nucleus. TCAP-1 did not significantly alter CRF-induced c-Fos expression in allareas, such as the PVN, BnST, and locus coeruleus, which indicatesthat TCAP-1 is acting in an HPA axis-independent manner. Further-more, in the elevated plus maze, acute injections of TCAP-1 had asignificant main effect on stretched-attend postures, a type of risk-assessment behavior. However, no other measures of anxiety wereaffected, and there were no interactions with CRF. TCAP-1 did nothave any effects in the open field test. Together, these results indi-cate that TCAP-1 administration can attenuate the effects of CRFon c-Fos induction in specific areas in the brain associated with themodulation of behavioral responses to stress. These results indicatethat TCAP-1 may be acting as an endogenous modulator of the CRFsystem in key limbic regions to modulate behavior, which couldhave implications in the regulation of normal stress and anxiety, aswell as for mood and anxiety disorders.
Acknowledgements
This work was supported by Canadian Institutes of HealthResearch (CIHR) grant(s) FRN: 72027622 to Franco J. Vaccarinoand Susan Rotzinger, and by the National Sciences and Engineer-

2 rain R
iDtaa
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
endorphin. Science 1981;213:1394–7.[32] Vertes RP. A PHA-L analysis of ascending projections of the dorsal raphe nucleus
06 L.A. Tan et al. / Behavioural B
ng Research Council of Canada (NSERC) for a Discovery Grant toavid A. Lovejoy and a Ph.D. Scholarship to Laura A. Tan. We also
hank Paul C. Boutros for consultation regarding statistical analysis,nd thank Rasha Ahmed and Yvonne Kwok for excellent technicalssistance.
eferences
[1] Al Chawaf A, St Amant K, Belsham D, Lovejoy DA. Regulation of neurite growthin immortalized mouse hypothalamic neurons and rat hippocampal cultures byteneurin C-terminal-associated peptide-1. Neuroscience 2007;144:1241–54.
[2] Al Chawaf A, Xu K, Tan L, Vaccarino FJ, Lovejoy DA, Rotzinger S. Corticotropin-releasing factor (CRF)-induced behaviors are modulated by intravenousadministration of teneurin C-terminal associated peptide-1 (TCAP-1). Peptides2007;29:1406–15.
[3] Bittencourt JC, Sawchenko PE. Do Centrally administered neuropeptides accesscognate receptors?: an analysis in the central corticotropin-releasing factorsystem. J Neurosci 2000;20:1142–56.
[4] Britton DR, Koob GF, Rivier J, Vale W. Intraventricular corticotropin-releasingfactor enhances behavioral effects of novelty. Life Sci 1982;31:363–7.
[5] Crane JW, French KR, Buller KM. Patterns of neuronal activation in the rat brainand spinal cord in response to increasing durations of restraint stress. Stress2005;8:199–211.
[6] Cullinan WE, Herman JP, Battaglia DF, Akil H, Watson SJ. Pattern and time courseof immediate early gene expression in rat brain following acute stress. Neuro-science 1995;64:477–505.
[7] Dayas CV, Buller KM, Crane JW, Xu Y, Day TA. Stressor categorization: Acutephysical and psychological stressors elicit distinctive recruitment patternsin the amygdala and in medullary noradrenergic cell groups. Eur J Neurosci2001;14:1143–52.
[8] Dayas CV, Buller KM, Day TA. Neuroendocrine responses to an emotional stres-sor: evidence for involvement of the medial but not the central amygdala. EurJ Neurosci 1999;11:2312–22.
[9] Figueiredo HF, Bruestle A, Bodie B, Dolgas CM, Herman JP. The medial prefrontalcortex differentially regulates stress-induced c-fos expression in the forebraindepending on type of stressor. Eur J Neurosci 2003;18:2357–64.
10] Grant EC, Mackintosh JH. A comparison of the social postures of some commonlaboratory rodents. Behavior 1963;21:246–59.
11] Herman JP, Figueiredo H, Mueller NK, Ulrich-Lai Y, Ostrander MM, Choi DC, etal. Central mechanisms of stress integration: hierarchical circuitry controllinghypothalamo-pituitary-adrenocortical responsiveness. Front Neuroendocriol2003;24:151–80.
12] Herman JP, Ostrander MM, Mueller NK, Figueiredo H. Limbic system mech-anisms of stress regulation: hypothalamo-pituitary-adrenocortical axis. ProgNeuropsychopharmacol Biol Psych 2005;29:1201–13.
13] Imaki T, Shibasaki T, Hotta M, Demura H. Intracerebroventricular administra-tion of corticotrophin-releasing factor induces c-fos mRNA expression in brainregions related to stress responses: comparison with pattern of c-fos mRNAinduction after stress. Brain Res 1993;616:114–25.
14] Kaesermann HP. Stretched attend posture, a non-social form of ambiva-lence, is sensitive to a conflict-reducing drug action. Psychopharmacol (Berl)1986;89:31–7.
15] Koob GF, Heinrichs SC, Pich EM, Menzaghi F, Baldwin H, Miczek K, et al. The roleof corticotropin-releasing factor in behavioral responses to stress. Ciba FoundSymp 1993;172:277–89.
[
esearch 201 (2009) 198–206
16] Lee EH, Tang YP, Chai CY. Stress and corticotropin-releasing factor potentiatecenter region activity of mice in an open field. Psychopharmacology (Berl)1987;93:320–3.
[17] Liang KC, Lee EH. Intra-amygdala injections of corticotropin releasing factorfacilitate inhibitory avoidance learning and reduce exploratory behavior in rats.Psychopharmacology (Berl) 1988;96:232–6.
[18] Liang KC, Melia KR, Campeau S, Falls WA, Miserendino MJ, Davis M. Lesions ofthe central nucleus of the amygdala, but not the paraventricular nucleus of thehypothalamus, block the excitatory effects of corticotropin-releasing factor inthe acoustic startle reflex. J Neurosci 1992;12:2313–20.
19] Lister RG. Ethologically-based animal models of anxiety disorders. PharmacolTher 1990;46:321–40.
20] Lowry CA. Functional subsets of serotonergic neurones: implications forcontrol of the hypothalamic–pituitary–adrenal axis. J Neuroendocrinol2002;14:911–23.
21] Nakajima M, Inui A, Asakawa A, Momose K, Ueno N, Teranishi A, et al.Neuropeptide Y produces anxiety via Y2-type receptors. Peptides 1998:359–63.
22] Paxinos G, Watson C. The rat brain in stereotaxic coordinates. 3rd ed. Toronto:Academic Press; 1997.
23] Qian X, Barsyte-Lovejoy D, Wang L, Chewpoy B, Gautum N, Al Chawaf A, et al.Cloning and characterization of teneurin C-terminus associated peptide (TCAP)-3 from the hypothalamus of an adult rainbow trout (Oncorhynchus mykiss). GenComp Endocrinol 2004;137:205–16.
24] Rioja J, Santin LJ, Dona A, de Pablos L, Minano FJ, Gonzalez-Baron S, et al. 5-HT1Areceptor activation counteracts c-Fos immunoreactivity induced in serotoninneurons of the raphe nuclei after immobilization stress in the male rat. NeurosciLett 2006;397:190–5.
25] Rivest S, Laflamme N. Neuronal activity and neuropeptide gene transcrip-tion in the brains of immune-challenged rats. J Neuroendocrinol 1997;7:501–25.
26] Rodgers RJ, Cole JC. Anxiety enhancement in the murine elevated plus mazeby immediate prior exposure to social stressors. Physiol Behav 1993;53:383–8.
27] Rodgers RJ, Dalvi A. Anxiety, defence, and the elevated plus-maze. NeurosciBiobehav Rev 1997;21:801–10.
28] Spina MG, Merlo-Pich E, Akwa Y, Balducci C, Basso AM, Zorrilla EP, et al.Time-dependent induction of anxiogenic-like effects after central infusion ofurocortin or corticotropin-releasing factor in the rat. Psychopharamacology(Berl) 2002;160:113–21.
29] Sundquist SJ, Nisenbaum LK. Fast Fos: rapid protocols for single- anddouble-labeling c-Fos immunohistochemistry in fresh frozen brain sections.J Neuroscience Methods 2005;141:9–20.
30] Tan LA, Xu K, Vaccarino FJ, Lovejoy DA, Rotzinger S. Repeated intracerebralteneurin C-terminal associated peptide (TCAP)-1 injections produce enduringchanges in behavioral responses to corticotropin-releasing factor (CRF) in ratmodels of anxiety. Behav Brain Res 2008;188:195–200.
31] Vale W, Speiss J, Rivier C, Rivier J. Characterization of a 41-residue ovinehypothalamic peptide that stimulates secretion of corticotrophin and beta-
in the rat. J Comp Neurol 1991;313:643–68.33] Wang L, Rotzinger S, Al Chawaf A, Elias CF, Barsyte-Lovejoy D, Qian X, et al.
Teneurin proteins possess a carboxy terminal sequence with neuromodulatoryactivity. Brain Res Mol Brain Res 2005;133:253–65.
Copyright © 2022 FDOKUMEN



















