
Previous experience of ethanol withdrawal increases withdrawal-induced c-fos expression in limbic...
13
Psychopharmacology (2006) 185: 188–200 DOI 10.1007/s00213-005-0301-3 ORIGINAL INVESTIGATION Gilyana G. Borlikova . Julie Le Merrer . David N. Stephens Previous experience of ethanol withdrawal increases withdrawal-induced c-fos expression in limbic areas, but not withdrawal-induced anxiety and prevents withdrawal-induced elevations in plasma corticosterone Received: 2 September 2005 / Accepted: 21 December 2005 / Published online: 10 February 2006 # Springer-Verlag 2006 Abstract Rationale: Increased anxiety is a characteristic of the acute ethanol withdrawal syndrome. Repeated exposure of rats to withdrawal from chronic ethanol increases sensitivity to seizures. Objectives: We investi- gated whether repeated withdrawal experience increases withdrawal-induced anxiety and stress, and if it changes withdrawal-induced activation of related brain areas. Methods: Rats were chronically treated with an ethanol- containing liquid diet either for 24 days continuously (single withdrawal, SWD) or interspersed with 2×3-day withdrawal periods (repeated withdrawal, RWD), or with a control diet. Eight hours after ethanol withdrawal, anxiety- like behaviour was tested in the elevated plus-maze, blood corticosterone levels were measured, and expression level of markers of neuronal activity and plasticity, c-fos and zif268, was assessed. Results: Eight hours after ethanol withdrawal, SWD rats showed increased anxiety on the elevated plus-maze relative to control rats. Rats given previous withdrawal experiences did not show further increases in measures of anxiety. Corticosterone levels were elevated during withdrawal in SWD rats but not in RWD rats. RWD resulted in marked increases in c-fos expression in amygdala, hippocampus, nucleus accumbens and dorsolateral periaqueductal grey. In contrast, zif268 expression was not increased after RWD, and in central amygdala the marked increase in zif268 seen after SWD was absent after RWD. Conclusions: The data suggest increased ability of withdrawal to activate neuronal circuits but reduced plasticity after RWD. We suggest parallels between the consequences of repeated ethanol withdrawal and repeated exposure to stress, and discuss implications of withdrawal for brain plasticity. Keywords Amygdala . Detoxification . Dependence . Repeated withdrawal . Anxiety . Stress . c-fos . zif268 Introduction Repeated experience of detoxification from ethanol de- pendence leads to increased severity of withdrawal symp- toms (Baker and Cannon 1979; Brown et al. 1988; Lechtenberg and Worner 1991). Although most studies have concentrated on the effects of repeated ethanol withdrawal on seizure activity during withdrawal (Brown et al. 1988; Lechtenberg and Worner 1991), a phenomenon readily modelled in rodents (Becker and Hale 1993; Ghozland et al. 2005; Mhatre and Gonzalez 1999; Stephens et al. 2001; Veatch and Becker 2002), attention has been given more recently to the effects of repeated withdrawal on other behavioural and physiological symptoms of withdrawal (Holter et al. 1998; Overstreet et al. 2002) and to long-term consequences of repeated withdrawal experience on emotional and cognitive function (Duka et al. 2004; Ripley et al. 2003a; Stephens et al. 2001; Townshend and Duka 2003). A link has also been drawn between repeated withdrawal and binge drinking (Duka et al. 2004; Stephens et al. 2005; Townshend and Duka 2005). We have previously reported that a long-term consequence of repeated withdrawal from ethanol is the impairment of fear conditioning in both rodents (Ripley et al. 2003b; Stephens et al. 2001) and humans (Stephens et al. 2005), though aspects of appetitive conditioning may also be influenced (Ripley et al. 2004). Increased anxiety is a characteristic of the acute ethanol withdrawal syndrome (DSM-IV 1994). Nevertheless, there is mixed evidence in the literature that the number of previous detoxification experiences influences anxiety levels in alcoholics undergoing acute detoxification. In one study, several days into detoxification of alcoholic inpatients, although we found increased levels of anxiety in patients undergoing detoxification, anxiety levels were not exacerbated by prior withdrawal experience (Duka et al. G. G. Borlikova . J. Le Merrer . D. N. Stephens (*) Department of Psychology, School of Life Sciences, University of Sussex, Falmer, Brighton, BN1 9QG, UK e-mail: [email protected] Tel.: +44-1273-678638 Fax: +44-1273-876619
Transcript of Previous experience of ethanol withdrawal increases withdrawal-induced c-fos expression in limbic...

Psychopharmacology (2006) 185: 188–200DOI 10.1007/s00213-005-0301-3
ORIGINAL INVESTIGATION
Gilyana G. Borlikova . Julie Le Merrer .David N. Stephens
Previous experience of ethanol withdrawal increaseswithdrawal-induced c-fos expression in limbic areas, but notwithdrawal-induced anxiety and prevents withdrawal-inducedelevations in plasma corticosteroneReceived: 2 September 2005 / Accepted: 21 December 2005 / Published online: 10 February 2006# Springer-Verlag 2006
Abstract Rationale: Increased anxiety is a characteristicof the acute ethanol withdrawal syndrome. Repeatedexposure of rats to withdrawal from chronic ethanolincreases sensitivity to seizures. Objectives: We investi-gated whether repeated withdrawal experience increaseswithdrawal-induced anxiety and stress, and if it changeswithdrawal-induced activation of related brain areas.Methods: Rats were chronically treated with an ethanol-containing liquid diet either for 24 days continuously(single withdrawal, SWD) or interspersed with 2×3-daywithdrawal periods (repeated withdrawal, RWD), or with acontrol diet. Eight hours after ethanol withdrawal, anxiety-like behaviour was tested in the elevated plus-maze, bloodcorticosterone levels were measured, and expression levelof markers of neuronal activity and plasticity, c-fos andzif268, was assessed. Results: Eight hours after ethanolwithdrawal, SWD rats showed increased anxiety on theelevated plus-maze relative to control rats. Rats givenprevious withdrawal experiences did not show furtherincreases in measures of anxiety. Corticosterone levelswere elevated during withdrawal in SWD rats but not inRWD rats. RWD resulted in marked increases in c-fosexpression in amygdala, hippocampus, nucleus accumbensand dorsolateral periaqueductal grey. In contrast, zif268expression was not increased after RWD, and in centralamygdala the marked increase in zif268 seen after SWDwas absent after RWD. Conclusions: The data suggestincreased ability of withdrawal to activate neuronalcircuits but reduced plasticity after RWD. We suggestparallels between the consequences of repeated ethanolwithdrawal and repeated exposure to stress, and discussimplications of withdrawal for brain plasticity.
Keywords Amygdala . Detoxification . Dependence .Repeated withdrawal . Anxiety . Stress . c-fos . zif268
Introduction
Repeated experience of detoxification from ethanol de-pendence leads to increased severity of withdrawal symp-toms (Baker and Cannon 1979; Brown et al. 1988;Lechtenberg and Worner 1991). Although most studieshave concentrated on the effects of repeated ethanolwithdrawal on seizure activity during withdrawal (Brownet al. 1988; Lechtenberg and Worner 1991), a phenomenonreadily modelled in rodents (Becker and Hale 1993;Ghozland et al. 2005; Mhatre and Gonzalez 1999; Stephenset al. 2001; Veatch and Becker 2002), attention has beengiven more recently to the effects of repeated withdrawalon other behavioural and physiological symptoms ofwithdrawal (Holter et al. 1998; Overstreet et al. 2002)and to long-term consequences of repeated withdrawalexperience on emotional and cognitive function (Duka etal. 2004; Ripley et al. 2003a; Stephens et al. 2001;Townshend and Duka 2003). A link has also been drawnbetween repeated withdrawal and binge drinking (Duka etal. 2004; Stephens et al. 2005; Townshend and Duka 2005).We have previously reported that a long-term consequenceof repeated withdrawal from ethanol is the impairment offear conditioning in both rodents (Ripley et al. 2003b;Stephens et al. 2001) and humans (Stephens et al. 2005),though aspects of appetitive conditioning may also beinfluenced (Ripley et al. 2004).
Increased anxiety is a characteristic of the acute ethanolwithdrawal syndrome (DSM-IV 1994). Nevertheless, thereis mixed evidence in the literature that the number ofprevious detoxification experiences influences anxietylevels in alcoholics undergoing acute detoxification. Inone study, several days into detoxification of alcoholicinpatients, although we found increased levels of anxiety inpatients undergoing detoxification, anxiety levels were notexacerbated by prior withdrawal experience (Duka et al.
G. G. Borlikova . J. Le Merrer . D. N. Stephens (*)Department of Psychology, School of Life Sciences,University of Sussex,Falmer, Brighton, BN1 9QG, UKe-mail: [email protected].: +44-1273-678638Fax: +44-1273-876619

2002). In contrast, (Malcolm et al. 2002) found someevidence for a mild increase in anxiety in this populationvery early in withdrawal. Nevertheless, in a rodent model,Breese and colleagues (Breese et al. 2005; Overstreet et al.2002, 2003, 2004a,b) consistently found a decrease insocial interaction, which has been characterised as ananimal model of anxiety (File 1980; File and Hyde 1978),in rats subjected to repeated ethanol withdrawal, andconcluded that multiple withdrawals from chronic ethanolled to a sensitisation of withdrawal-induced anxiety. Usinga quite different model of lengthy (11-month) chronicethanol intake, Holter et al. 1998 also reported thatwithdrawal anxiety was higher after many (perhaps nine)withdrawal episodes than on the first.
We found in previous experiments that repeated with-drawal from another class of sedative/hypnotic drugs, thebenzodiazepines, whilst like ethanol withdrawal, sensi-tising mice to convulsants, reduced their anxiety-likebehaviour in the elevated plus-maze and four-plate tests(Ward and Stephens 1998). If ethanol withdrawal resem-bles benzodiazepine withdrawal, repeated withdrawalexperience from ethanol may even result in habituation tothe withdrawal-induced stress and perhaps lower respon-siveness to stress-related anxiety. In this context, it isinteresting that repeated exposure to restraint stress alsoprotects against future stressors (Belda et al. 2004; Carter etal. 2004; Chen and Herbert 1995; Stamp and Herbert1999).
In the present study, we therefore tested whetherrepeated ethanol withdrawal led to the sensitisation of theanxiety state, as suggested by Overstreet, Breese andcolleagues (Overstreet et al. 2002; Breese et al. 2005), or,as we have reported for repeated benzodiazepine with-drawal, if it lowered anxiety level in rodents. For thispurpose, we tested animals in the elevated plus-maze, atask frequently used to assess anxiety (Pellow et al. 1985;Rodgers et al. 1999), including anxiety induced by ethanolwithdrawal (Baldwin et al. 1991). Additionally, we mea-sured blood levels of the stress-related hormone, cortico-sterone (Herman and Cullinan 1997). To understand moreprecisely the brain processes influenced by repeatedwithdrawal, the ability of the final withdrawal to affectexpression levels of the immediate early genes that aremarkers of neuronal activity, c-fos and zif268, wasscreened in different brain areas related to stress, fear andanxiety. The amygdala is strongly implicated in fear- andanxiety-related processes, as well as showing activationduring stress processing (Dayas et al. 2001; LeDoux 2000;Singewald et al. 2003), and the hippocampus also plays arole in these processes, especially in case of conditionedcues and cognitive stressors (Herman et al. 1998; Linden etal. 2004). The hypothalamus is important for regulation ofthe hormonal response to stress and to fear-related stimuli(Dayas et al. 2001; Herman and Cullinan 1997; Linden etal. 2004; Singewald et al. 2003), and the periaqueductalgrey is implicated in the response to stressors and anxiety-related behaviour (Dayas et al. 2001; Linden et al. 2004;Salome et al. 2004). Changes in the expression of c-fos andzif268 are widely used as indicators of brain activation as a
result of different manipulations (Herdegen and Leah 1998;Knapska and Kaczmarek 2004; Mutschler et al. 2000).Stress-related changes in the level of c-fos and zif268expression are well documented (Day et al. 2005;Herdegen and Leah 1998; Keay et al. 2001; Knapska andKaczmarek 2004; Melia et al. 1994; Senba and Ueyama1997).
Materials and methods
Animals
The subjects of experiment 1 were 36 male Lister hoodedrats (130–160 g at the beginning of the experiment, Harlan,Bicester, UK). For experiments 2 and 3, 54 male Listerhooded rats (350–400 g at the beginning of the experiment,University of Sussex, Brighton, UK) were used. Allanimals were pair-housed and maintained on a 12-h light/dark cycle (lights off at 19:00; temperature 21±2°C;humidity 50±10%) with ad libitum access to water. Allexperiments were carried out under the authority of the UKAnimal (Experimental Procedures) Act, 1986.
Chronic ethanol treatment
For each of the experiments described below, separategroups of rats were randomly assigned to one of threetreatment conditions: single withdrawal (SWD), repeatedwithdrawal (RWD) and control (CON) (according toRipley et al. 2004). Briefly, the animals in the first twogroups received a nutritionally complete liquid diet (Dyets,Bethlehem, PA, USA), containing 7% ethanol as their onlyfood source. Control animals were pair-fed with acalorifically matched control diet. During 30 days oftreatment, the RWD group had two intermediate with-drawal episodes (days 11–13 and 21–23), when animalsreceived the control diet instead of the ethanol diet. TheSWD group remained on ethanol-containing diet for 24consecutive days.
On the final day, animals were withdrawn from theliquid diet at 06:00–08:00 h (counterbalanced betweengroups) and remained in their home cages with ad libitumaccess to water but no food for 8 h until they were tested inthe elevated plus-maze or killed for blood collection orimmunohistochemistry. In our experience, food restrictionduring this period heightens the ethanol withdrawalsyndrome, so that signs of withdrawal could be observedthroughout the 8-h period (unpublished observations fromRipley et al. 2004).
Experiment 1
Behaviour in the elevated plus-maze
The animals (n=12 per treatment group) were testedbetween 14:00 and 17:00 h in the plus-maze, 8 h after
189
the final withdrawal. The plus-maze consisted of two open(44×12 cm) and two enclosed (44×12×26 cm) arms thatwere connected by a common central platform (12×12 cm).The arms and central platform had white Plexiglascovering; the walls of the enclosed arms were from greyPlexiglas. The maze was elevated 82 cm above the floorlevel and illuminated using diffuse overhead fluorescentlighting. Testing was conducted under low light illumina-tion (2×32 W, 2 m above the maze). The animals wereplaced on the central platform facing an open arm and theirbehaviour in the maze was recorded for 5 min by a videocamera. The maze was wiped with cleaning liquid aftereach animal was tested. Behavioural parameters, scored byobservation of videotapes, (according to Cruz et al. 1994,Rodgers et al. 1999), were the number of open and closedarm entries, time spent in different arms, the number ofcrossings of the hypothetical line dividing an arm intoequal proximal and distal parts, time spent in distal parts ofopen and closed arms, the number of head dips from openarms and the number of “protected” dips, the number ofrears, the number of risk assessments [investigations of theopen arms from the central platform or a closed arm withthe forepaws and head only (Cruz et al. 1994; Rodgers et al.1999)] and the number of grooming episodes. Time spentin distal parts of open arms was calculated as percentage ofthe total time spent in distal parts of both types of arms. Thenumber of rears and head dips was calculated as a ratio ofthe number of rears (dips) in the open/closed arms to thetotal time spent in that kind of arm/area.
Experiment 2
Collection of blood samples and determinationof plasma corticosterone
The animals (n=10 per group) were killed by decapitation8 h after ethanol withdrawal. Trunk blood was collectedinto iced, heparinised tubes and centrifuged at 3,500×g for15 min at room temperature to remove cells, and the plasmawas stored at −18°C. Plasma corticosterone levels weredetermined using a commercially available enzyme immu-noassay kit (Immunodiagnostic Systems, UK). Intra- andinter-assay coefficients of variation were less than 5%.
Experiment 3
c-fos and zif268 immunohistochemistry
c-fos and zif268 expression was assessed in the sameanimals (n=8 per group). Eight hours after final ethanolwithdrawal, animals were deeply anaesthetised with Avert-in (40 mg/kg, i.p.) and immediately transcardially perfusedwith paraformaldehyde in 0.1 M phosphate-buffered saline(PBS). The brains were excised and placed in 4%paraformaldehyde in PBS for 24 h, then cryoprotectedin sucrose before freezing in isopentane at −50°C andstored at −80°C for up to 2 months before being
coronally sectioned at 30 μm using a freezing micro-tome. Sections were collected in PBS (alternativesections through the region of interest stained for c-fosor for zif268), incubated with 0.3% H2O2 for 10 min,and washed in PBS. After 1 h incubation with 1.5%normal goat serum (NGS, Vector Laboratories), sectionswere washed again and incubated overnight in PBScontaining 0.5% NGS, 0.02% sodium azide (Sigma),0.3% Triton X-100 (Sigma) and 1:10,000 c-fos primaryantibody (rabbit polyclonal serum, Oncogene, Ab-5) or1:1,600 zif268 primary antibody (Egr-1 (588), rabbitpolyclonal serum, Santa Cruz Biotechnology, sc-110).On the next day, sections were washed and incubated for1 h in PBS containing 1.5% NGS and 1:300 biotinylatedanti-rabbit secondary antibody (Vector laboratories,BA-1000). After washing, sections were incubated for1 h in PBS containing 1:1,000 avidin-biotinylated–horseradish peroxidase complex (Vectastain ABC kitElite, Vector Laboratories). The reaction was visualisedusing a standart glucose oxidase-3,3-diaminobenzidinemethod. The reaction was terminated by extensivewashing.
Fos- and zif-positive nuclei were quantified from imagesof sections captured using AxioCam HRc digital cameramounted on a Zeiss Akioskop 2 plus microscope (CarlZeiss, UK) using AxioVision 3.1 software (ImagingAssociates, Bicester, UK) with ×100 magnification. Theregions studied were basolateral (BLA, 0.15 mm2 region),central (CeA, 0.13 mm2) and lateral (LA, 0.18 mm2)nuclei of the amygdala, core (AcbC, 0.13 mm2) and dorsalshell of nucleus accumbens (AcbSh, 0.13 mm2), CA1(0.12 mm2) and CA3 (0.08 mm2) fields of hippocampus,medial parvacellular nucleus of paraventricular nucleus of
Ethanol consumption
0 2 4 6 8 10 12 14 16 18 20 22 240
2
4
6
8
10
12
14
16
18
20
22
24
SWD
RWD
**
*
**
day
g/k
g
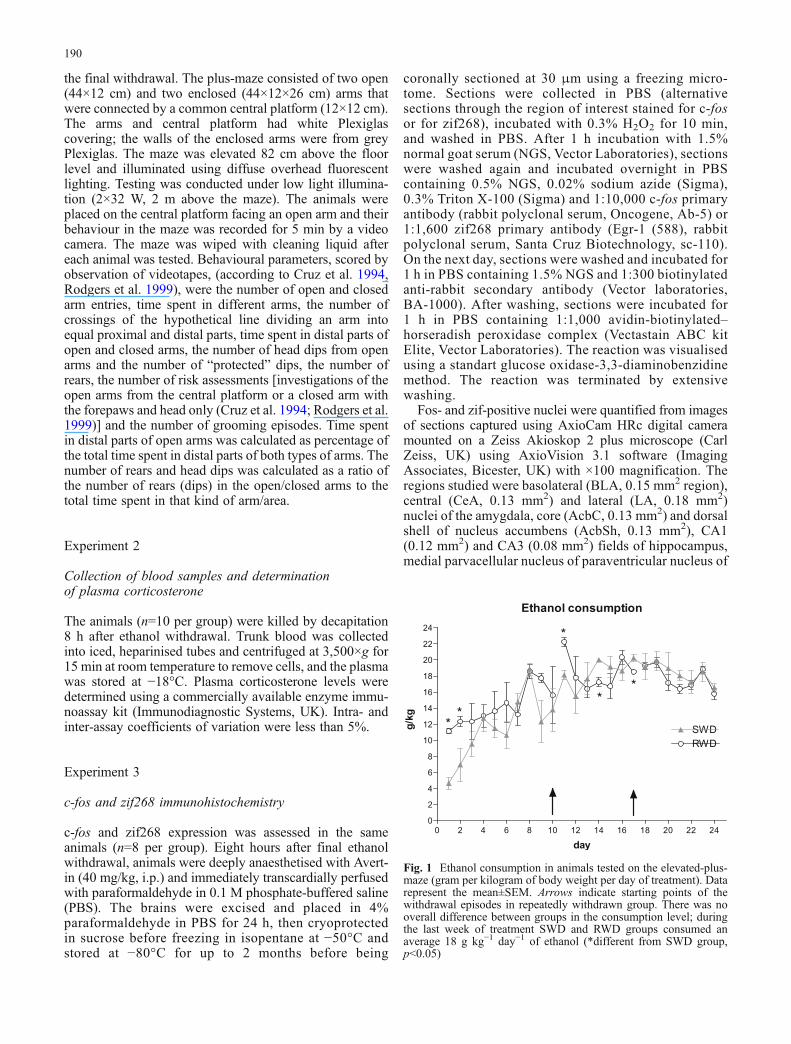
Fig. 1 Ethanol consumption in animals tested on the elevated-plus-maze (gram per kilogram of body weight per day of treatment). Datarepresent the mean±SEM. Arrows indicate starting points of thewithdrawal episodes in repeatedly withdrawn group. There was nooverall difference between groups in the consumption level; duringthe last week of treatment SWD and RWD groups consumed anaverage 18 g kg−1 day−1 of ethanol (*different from SWD group,p<0.05)
190

hypothalamus (PVN, 0.05 mm2), dorsal caudate putamen(dCPu, 0.18 mm2) and dorsolateral periaqueductal grey(dlPAG, 0.23 mm2). Counting was performed using ScionImage for Windows (Scion Corporation, MD, USA). Thelocations of the areas used for quantifying Fos- and zif-positive nuclei were taken from Paxinos and Watson(1998) and are shown in Fig. 3. Counts were takenbilaterally in two sections for each brain region and themean of these four values was taken as the number ofnuclei for that brain region.
Statistics
Data collected were analysed using SPSS 11.5 for Windows.For each set of data, means were calculated and data thatwere greater than or less than mean ±2 SD were excludedfrom the analysis. All data analysed were initially checkedfor normality of distribution by Kolmogorov–Smirnov’s testof normality. Then data were analysed using one-way ortwo-way ANOVA and repeated measures ANOVA. Signif-icant interactions were followed by post hoc tests (Student–
Time spent in different arms
0
100
200CON
SWD
RWD
closed open
** **
arm
sec
Time spent in distal parts ofopen arms
CON SWD RWD0
25
50
75
**
group%
Entries into different arms
0.0
2.5
5.0
7.5CON
SWD
RWD
closed openarm
nu
mb
er
of
en
trie
s
Entries into distal parts of arms
0
25
50
75CON
SWD
RWD
closed open
* *
arm
%
Rears
0.00
0.05
0.10
0.15CON
SWD
RWD
closed open
*
arm
rears
/min
Dips
0
5
10
15
20CON
SWD
RWD
protected open
****
*
area
dip
s/m
in
A B
C D
E F
Fig. 2 Measures of anxiety-related behaviour in the plus-maze.Data show the mean±SEM, n=11–12 per group. a Time spent indifferent arms (**p<0.01 compared to closed arms). b Time spent indistal parts of open arms as percentage from the total time in distalparts of arms (*p<0.05 compared to CON). c The number of entriesinto different arms (SWD and RWD groups less active than CON,p<0.05). d The number of entries into distal parts of different armsas percentage of the total number of arm entries (all animals entered
distal parts of open arms on fewer occasions, p<0.001; *p<0.05compared to CON). e Rates of rearing in open and closed arms (allanimals performed more rearings in closed arms, p<0.001; *p<0.05compared to SWD in open arms). f Rates of dipping from the“protected” (central platform and proximal parts of closed arms) andopen areas (*p<0.05 compared to CON in “protected” area;**p<0.01 compared to CON in open arms)
191

Newman–Keuls test, t test). All data analysed using arepeated measures ANOVA were initially checked byMauchly’s test of sphericity. A significant p valueindicated that there was sufficient evidence to reject thesphericity assumption and the p value for repeatedmeasures was calculated using the Greenhouse–GeisserEpsilon correction. All tests of significance wereperformed at α values of 0.05.
Results
Chronic ethanol treatment
In the subgroup of rats later tested on the elevated plus-maze,themean ethanol consumption over the final week of chronictreatment in both SWD and RWDgroups was approximately18 g ethanol per kilogram body weight per day. An analysisof the overall ethanol consumption with repeated measuresANOVA showed that, whilst consumption changed withtreatment (Fig. 1; main effect of day F23, 230=13.16, p<0.05),there was no significant difference in ethanol intake betweenthe two groups (F1,10=4.39, ns), but there was a significantgroup by day interaction (F23,230=2.00, p<0.05). Post hocanalysis revealed that SWD and RWD groups consumeddifferent amount of ethanol on days 1, 2, 11, 15 and 17(F1,10=69.63, F1,10=7.54, F1,10=33.12, F1,10=24.72, p<0.05)of the 24-day treatment. In experiments 2 and 3, over thefinal week of chronic treatment, SWD and RWD animalsconsumed approximately 12.5 g kg−1 day−1 of ethanol.Analysis of the 24-day consumption showed changes in theconsumption level with treatment (F23,138=6.97, p<0.05),but no difference between SWD and RWD groups and nogroup by day interaction (F1,6=1.09, F23,138=0.83, ns).
Experiment 1
Behaviour in the elevated plus-maze
The effect of single and repeated withdrawal from chronicethanol on behaviour in the elevated plus-maze is shown inFig. 2. A significant effect of arm (open vs closed)(F1,64=27.36, p<0.001) indicated less exploration of theopen arms (Fig. 2a). Whilst there was no main effect oftreatment (F2,64=0.14, ns), a significant group by arminteraction (F2,64=3.39, p<0.05) suggested that ethanolwithdrawal decreased exploration of the open arms. Whilst
under the experimental conditions used, controls showed nodifferences in time spent exploring open and closed arms(T20=1.27, ns), both SWD and RWD rats spent significantlyless time exploring the open arms (SWD: T22=4.82,p<0.001; RWD: T22=2.88, p<0.01). A clear group differenceemerged in the percentage of time that animals fromdifferent groups spent in the distal part of open arms
CON SWD RWD0
50
100
150
200
250CON
SWD
RWD
*
group
Co
rtic
oste
ron
en
g/m
l
Fig. 3 Blood corticosterone levels, 8-h after withdrawal. SWDanimals had higher level of corticosterone than both CON and RWDanimals. n=9–10 per group (*p<0.05)
Fig. 4 Schematic diagrams, adapted from the atlas of Paxinos andWatson (1998), showing the areas in which c-fos and zif268expression was quantified. Bregma 1.60: AcbC Nucleus accumbenscore, AcbSh nucleus accumbens dorsal shell, dCPu dorsal caudateputamen. Bregma −1.80: PVN Medial parvacellular part ofparaventricular nucleus of hypothalamus. Bregma −3.14: LA lateralamygdala nucleus, BLA basolateral amygdala nucleus, CeA centralamygdala nucleus, CA1 CA1 field of hippocampus, CA3 CA3 fieldof hippocampus. Bregma −6.8: dlPAG dorsolateral periaqueductalgrey
Table 1 Additional exploratory measures from the plus-maze
Measures Treatment group
CON SWD RWD
Risk assessments 13.18±1.02 12.08±0.99 13.67±1.12Groomings 1.00±0.45 0.75±0.48 0.58±0.20
Data presented as means±SEM. Risk assessments representedinvestigation of the open arms from the central platform or a closedarm (Cruz et al. 1994; Rodgers et al. 1999)
192

(Fig. 2b, main effect of treatment; F2,32=4.52, p<0.05), posthoc tests revealing that animals from SWD and RWDgroups spent significantly less time in the distal parts ofopen arms than control animals (p<0.05). The analysis of thenumber of closed and open arm entries (Fig. 2c) showedsignificant effect of treatment group (F2,64=6.21, p<0.01),but no effect of arm (F1,64=1.46, ns), and no group by arminteraction (F2,64=1.03, ns). Post hoc tests revealed that bothethanol-treated groups were significantly different from theCON group, indicating reduced activity, but did not differfrom each other. The analysis of the entries into the distalparts of arms as a percentage of the total number of armentries (Fig. 2d) further supported findings from the analysis
of the time spent in different arms. It showed that all animalsvisited distal parts of the open arms on fewer occasionsthan the distal parts of the closed arms (arm: F1,64=57.33,p<0.001; group: F2,64=2.21, ns; group by arm interaction:F2,64=4.58, p<0.05). Consequent separate analysis for eachtype of arms revealed that both RWD and SWD animalsavoided the most anxiogenic distal parts of the open arms(F2,32=4.15, p<0.05; post hoc p<0.05), whilst in the closedarms, animals from all treatment groups showed the sameactivity (F2,32=2.39, ns).
All animals performed more rears (Fig. 2e) in the closedthan in the open arms (F1,64=203.18, p<0.001). Although noeffect of treatment group was found, (F2,32=0.90, ns), there
Amygdala
0
50
100
150
200
250CON
SWD
RWD*
*
CeA BLA LA
subdivision
c-fo
sce
lls
per
mm
2
Hippocampus
0
25
50
75CON
SWD
RWD
CA1 CA3
**
field
c-fo
sce
lls
per
mm
2
Nucleus Accumbens
0
100
200
300CON
SWD
RWD
AcbC AcbSh
**
subdivision
c-fo
sce
lls
per
mm
2
Paraventricular nucleus ofHypothalamus
CON SWD RWD0
50
100
150
200
250
300
350CON
SWD
RWD
group
c-fo
sce
lls
per
mm
2
Periaqueductal Grey
CON SWD RWD0
25
50
75
100 **
group
c-fo
sce
lls
per
mm
2
Caudate Putamen
CON SWD RWD0
10
20
30
40
50CON
SWD
RWD
group
c-fo
sce
lls
per
mm
2
A B
C D
E F
Fig. 5 Effect of single and repeated withdrawal from ethanol on thenumber of c-fos positive cells in amygdala (a), hippocampus (b),nucleus accumbens (c), paraventricular nucleus of hypothalamus(d), periaqueductal grey (e) and caudate putamen (f). Data represent
the mean±SEM of the number of c-fos positive cells in an indicatedbrain region, n=7–8 per group (*different from CON and SWDgroups, **different from CON group, p<0.05)
193
was a significant group by arm interaction (F2, 64= 3.59,p<0.05). Subsequent analyses of each arm separately showedno group differences in the number of rears in the closedarms, but a significant difference in the rate of rearing betweenSWD and RWD groups in the open arms (F2,32=1.60, ns, closedarms and F2,32=3.89, p<0.05; post hoc p<0.05, open arms). Thepicture for head dips was rather different (Fig. 2f). Unsurpris-ingly, head dipping was mostly confined to the open arms. One-way ANOVA of dips from the open arms showed an effect oftreatment, with both alcohol-treated groups exhibiting fewerhead dips (F2,32=6.32, p<0.01; post hoc p<0.05, RWD andSWD vs CON). Additionally, SWD animals performedfewer dips than control animals from the central area ofthe maze (F2,32=5.36, p<0.05). There was no differencebetween experimental groups in the number of riskassessments and groomings (Table 1, risk assessments:F2,32=0.67, ns; groomings: F2,32 =0.30, ns).
Experiment 2
Blood corticosterone level
Differences in the plasma corticosterone level between thethree chronic treatment groups (Fig. 3) were significant(F2,26=4.08, p<0.05), post hoc Student–Newman–Keulsindicating that SWD animals differed from both CON andRWD animals in having higher level of corticosterone at8 hr after withdrawal, whilst there was no differencebetween RWD and CON groups.
Experiment 3
c-fos and zif268 expression
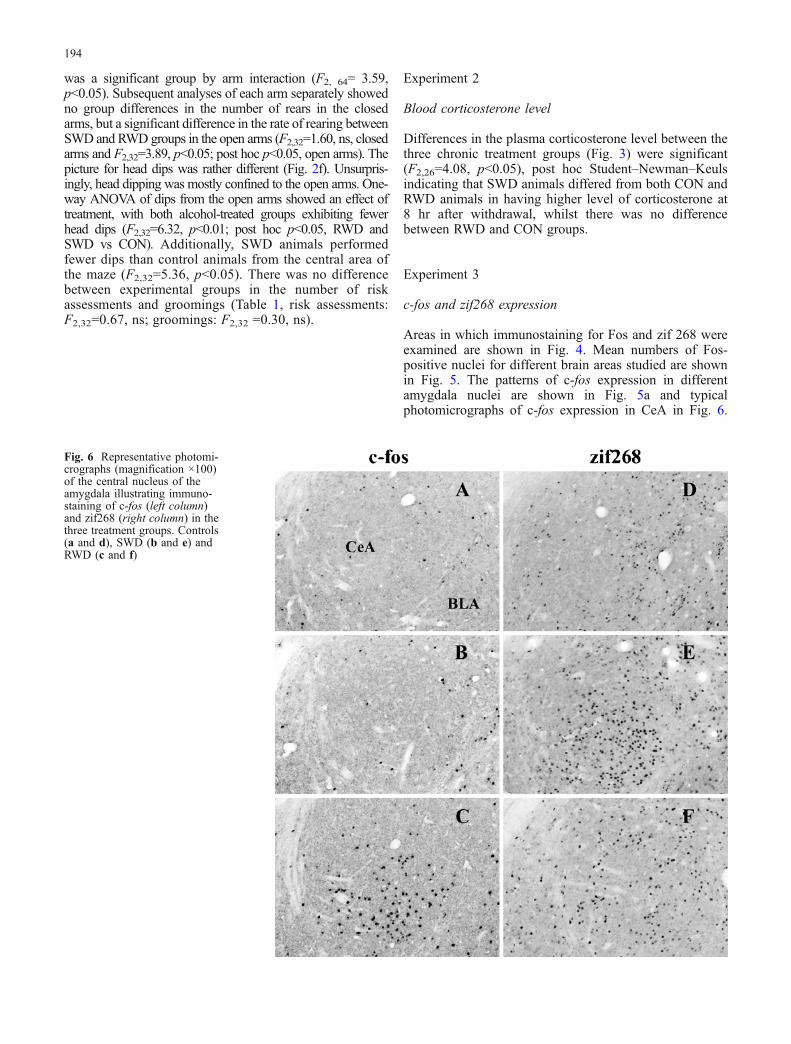
Areas in which immunostaining for Fos and zif 268 wereexamined are shown in Fig. 4. Mean numbers of Fos-positive nuclei for different brain areas studied are shownin Fig. 5. The patterns of c-fos expression in differentamygdala nuclei are shown in Fig. 5a and typicalphotomicrographs of c-fos expression in CeA in Fig. 6.
Fig. 6 Representative photomi-crographs (magnification ×100)of the central nucleus of theamygdala illustrating immuno-staining of c-fos (left column)and zif268 (right column) in thethree treatment groups. Controls(a and d), SWD (b and e) andRWD (c and f)
194

Two-way ANOVA revealed significant main effect ofamygdala region (F2,34=15.88, p<0.01), region by groupinteraction (F4,34=5.47, p<0.01) and a significant differ-ence between treatment groups (F2,17=16.88, p<0.01). Posthoc test showed that the RWD group differed from bothCON and SWD animals. Subsequent separate analysis of c-fos data from each nucleus showed that, whilst there was nodifference among groups in the level of this gene expressionin LA, in CeA and BLA nuclei of RWD animals, level of c-fos expression was much higher than in CON and SWDanimals (one-way ANOVA, LA: F2,20=0.64, ns; BLA:F2,18=8.36, p<0.05, post hoc RWD differed from CON and
SWD; CeA: F2,18=16.82, p<0.01, post hoc RWD differedfrom CON and SWD; see Fig. 5 for typical photomicro-graphs of Fos immunostaining in CeA).
In the hippocampus (Fig. 5b), analysis showed adifference between c-fos expression in CA1 and CA3field (F1,17=12.8, p<0.01), but no field by group interac-tion (F2,17=1.62, ns), and no effect of treatment group(F2,17=2.17, ns). Subsequent one-way ANOVA found nosignificant difference between groups in the CA1 field(F2,19=1.41, ns), whereas in the CA3 field pattern wasrather different—the RWD group had a significantly higherlevel of c-fos expression than the CON group, with SWD
Amygdala
0
500
1000
1500CON
SWD
RWD
*
CeA BLA LA
subdivision
zif
268
cell
sp
erm
m2
Hippocampus
0
500
1000
1500
2000
2500CON
SWD
RWD
CA1 CA3
field
zif
268
cell
spe
rm
m2
Nucleus Accumbens
0
500
1000
1500
2000
2500CON
SWD
RWD
AcbC AcbSh
subdivision
zif
268
cell
spe
rm
m2
Paraventricular nucleus ofHypothalamus
CON SWD RWD0
20
40
60
group
zif
268
cell
sp
erm
m2
Periaqueductal Grey
CON SWD RWD0
100
200
300
group
zif
268
cell
sp
erm
m2
Caudate Putamen
CON SWD RWD0
500
1000
1500
2000
2500
group
zif
268
cell
sp
erm
m2
A B
C D
E F
Fig. 7 Effect of single and repeated withdrawal from ethanol on thenumber of zif268 positive cells in amygdala (a), hippocampus (b),nucleus accumbens (c), paraventricular nucleus of hypothalamus(d), periaqueductal grey (e) and caudate putamen (f). Data represent
the mean±SEM of the number of zif268 positive cells in anindicated brain region, n=7–8 per group (*different from CONgroup, p<0.05)
195

group intermediate (F2,18=4.63, p<0.05, post hoc RWDdiffered from CON).
In nucleus accumbens (Fig. 5c), there was a significantdifference in c-fos expression between core and shellregions (F1,19=62.14, p<0.01), no region by group interac-tion (F2,19=2.86, ns) and a significant main effect oftreatment group (F2,19=5.44, p<0.05). Post hoc testsshowed that RWD group differed from CON. One-wayANOVA clarified that in AcbC, RWD animals had higher c-fos expression than CON, whereas SWD animals wereagain intermediate (F2,20=3.63, p<0.05, post hoc RWDdiffered from CON); there was no significant differencebetween treatment groups in AcbSh (F2,19= 2.32, ns).
One-way ANOVA did not reveal any significantdifference between c-fos expression in PVN (Fig. 5d) ofethanol- and control-diet treated animals (F2,18=0.35, ns).Nevertheless, RWD animals had higher c-fos level indorsolateral PAG (Fig. 5e, F2,18=4.61, p<0.05, post hocRWD differed from CON). In the dorsal part of caudateputamen (Fig. 5f), despite some tendency, ANOVA analysisdid not show any significant difference between groups(F2,20=1.71, ns).
The profile of zif268 expression 8 h after the finalwithdrawal from chronic ethanol in different brainstructures is shown in Fig. 7. The patterns of zif268expression in different amygdala nuclei are shown inFig. 7a, and typical photomicrographs of zif268 expressionin CeA in Fig. 6. Analysis of the data with two-wayANOVA showed that level of zif268 expression variedbetween different amygdala nuclei (F2,38=106.43, p<0.01)and there was significant nucleus by group interaction(F4,38=3.43, p<0.05), but no main effect of group(F4,19=2.15, ns). Subsequent analysis showed that in theCeA, single withdrawal from ethanol caused significantincrease in zif268 expression; however, experience ofrepeated withdrawal did not affect expression of this gene(F2,20=5.47, p<0.05, post hoc SWD differed from CON andRWD). There was no difference in the level of zif268expression between treatment groups in the other amygdalanuclei (one-way ANOVA, BLA: F2,20=0.48, ns; LA:F2,21=0.13, ns).
In hippocampus (Fig. 7b), expression of zif268 wassignificantly higher in the CA1 field than in the CA3 field(F1, 21=594.58, p<0.01, nearly all nuclei were stained), butthere were no differences in expression between groups(F2,21=1.06, ns) and no region by group interaction(F2,21=0.75, ns). One-way ANOVA analysis confirmed,that, in contrast to the c-fos pattern, zif268 expression in thebrains of RWD and SWD rats was not changed in eitherfield of hippocampus examined (CA1: F2,21=1.05, ns;CA3: F2,21=0.79, ns).
In nucleus accumbens (Fig. 7c), the pattern was similar:There was main effect of region (F1,21=54.3, p<0.01) but nodifferences between groups (F2, 21=0.11, ns) and no regionby group interaction (F2,21=0.08, ns). Further analysis onceagain confirmed absence of differences between treatmentgroups (AcbC: F2,21=0.09, ns; AcbSh: F2,21=0.12, ns).
In PVN of hypothalamus (Fig. 7d), there was anumerically higher level of zif268 expression in SWDrats, but it was not significant (F2,18=0.79, ns). Level ofzif268 expression in dlPAG (Fig. 7e) and dCPu (Fig. 7f)was similar in all treatment groups (dlPAG: F2,19=1.24, ns;dCPu: F2,21=0.2, ns).
Discussion
In keeping with previous studies demonstrating increasedanxiety-like behaviour during alcohol withdrawal, rats, ontheir first experience of ethanol withdrawal, showedincreased anxiety-like behaviour (spent less time in theopen arms of the elevated plus-maze) than controls. Thisbecame especially apparent when time spent in and entriesinto the distal parts of the open arms were considered.Although entries to the distal parts of the arms did not formpart of the classical analyses of plus-maze behaviour(Handley and Mithani 1984; Pellow et al. 1985), a numberof researchers have subsequently introduced this measure(Dere et al. 2002; File et al. 1999), which factor analysisreveals to be heavily loaded under the factor anxiety (Cruzet al. 1994). Rats that had previously experiencedwithdrawal (RWD group) showed similar anxious-likebehaviour in these measures to SWD rats. Data on thenumber of entries into different arms showed that bothethanol-treated groups were generally less active thancontrols. Similar reductions in activity in the plus-mazeafter withdrawal from chronic ethanol treatment have beenreported previously (File et al. 1993; Gatch et al. 1999,2000; Watson et al. 1997; Wilson et al. 1998). The fact that,in our experiment, ethanol-treated groups tended to showreduced locomotor activity in some measures puts someconstraints on the interpretation of the elevated plus-mazedata. But as a greater locomotor suppression in RWD groupshould bias the entry towards a higher anxiety-like profile(Dawson and Tricklebank 1995; Kliethermes 2005;Kliethermes et al. 2005), our data are at least clear inshowing no evidence for sensitisation of the anxiety-likebehaviour with repeated withdrawal.
Considering the so-called ethological measures ofbehaviour in the plus-maze, animals from all threetreatment groups emitted similar numbers of rears, indi-cating little effect of ethanol withdrawal in this type ofexploratory activity (Rodgers et al. 1999); however, a smallincrease in numbers of rears was seen in RWD, comparedto SWD animals, in open arms. The number of head dipsfrom open arms was lower in the both RWD and SWDgroups than controls. Surprisingly, there was no significantdifference between groups in the number of risk assess-ments; number of grooming episodes was also similar in allgroups.
In summary then, whilst withdrawal from chronictreatment with ethanol gave rise to anxious-like behaviourin the plus-maze, there was no evidence that rats that hadpreviously experienced withdrawal show heightened levels
196

of anxiety, compared to SWD rats, even though thisregimen of repeated withdrawal from alcohol sensitises ratsto seizures (Stephens et al. 2001). This conclusion also fitswith our clinical observations that withdrawal-associatedanxiety is not increased following multiple withdrawals(Duka et al. 2002).
Nevertheless, the results obtained here using a 7%ethanol diet differ from those of Overstreet et al. (2002,2004a,b) when lower ethanol concentrations were given. Inthis latter case, an increase in anxiety-like behaviour afterrepeated withdrawal was found. There are several possibleaccounts of these discrepancies. Firstly, differences may bea result of different treatment protocols, as levels of intakeinduced by Overstreet and colleagues (Overstreet et al.2002) were lower than those that we achieve. Indeed, whena liquid diet regimen using 7% ethanol concentrations wasused (Overstreet et al. 2002), whilst withdrawal increasedanxiety relative to controls, no differences were seenbetween SWD and RWD groups. Secondly, there is apossibility that the elevated plus-maze and the socialinteraction test model different aspects of anxiety (File1992), and repeated withdrawal from ethanol affects one ofthese aspects more than the other. Thirdly, in theexperiments of Overstreet, Knapp and Breese, RWDanimals not only exhibited suppressed social interactioncompared with controls and SWD animals, which wasconsidered as an indication of increased anxiety, butsimultaneously ethanol-withdrawn animals showed adecrease in locomotion (Overstreet et al. 2002, 2003,2004a,b), a common observation during ethanol withdraw-al (File et al. 1993; Gatch et al. 1999; 2000; Watson et al.1997; Wilson et al. 1998). Reductions in activity may giverise to a false positive result in the social interaction test(File and Seth 2003; Kliethermes 2005; Koss et al. 2004),though by including the reduced locomotion as a factor inan analysis of covariance, Breese, Overstreet and col-leagues attempted to control for this aspect. Although othergroups have reported increased anxiety-like behaviour afterrepeated withdrawal from ethanol (Ghozland et al. 2005;Holter et al. 1998). Holter et al. (1998) used a much longerperiod of ethanol exposure, while Ghozland et al. (2005)did not control for length of exposure to ethanol, so that itcannot be interpreted whether the increased anxiety-likeprofile is the result of multiple withdrawals or of a moreprolonged ethanol exposure.
In addition to behavioural indices of anxiety, we alsoinvestigated the effect of withdrawal on blood levels ofthe stress-related hormone corticosterone (Herman andCullinan 1997). In keeping with the aversive and stressfulnature of withdrawal and as has been reported by others(Janis et al. 1998; Tabakoff et al. 1978), corticosteronelevels were increased in SWD animals. However, therewas no evidence for increased circulating levels of corti-costerone in RWD rats. The failure of withdrawal to in-crease corticosterone levels in RWD rats suggests that thewithdrawal experience was less stressful in this group.These results fit with more general observations of habit-uation to repeated stress (Belda et al. 2004; Carter et al.2004; Chen and Herbert 1995; Martinez et al. 1998; Melia
et al. 1994; Senba and Ueyama 1997; Stamp and Herbert1999; Umemoto et al. 1997).
Withdrawal from ethanol in RWD rats (but not in SWDanimals) led to a significant increase in the number ofneurons expressing Fos in the central and the basolateralnuclei of amygdala, in the CA3 field of hippocampus, inthe core of nucleus accumbens and in the dorsolateral PAG.Surprisingly, no evidence was seen of withdrawal-inducedactivation of the PVN. Fos expression in dCPu was also notchanged. Zif268 expression was increased only in thecentral nucleus of amygdala of the SWD group. In all otherstructures studied, level of zif268 expression after bothRWD and SWD was similar to that in the control group.
Fos expression increases as a result of neuronal activa-tion, and Fos is widely used for monitoring the areas of thebrain activated as a result of different manipulations (Chenand Herbert 1995; Dunworth et al. 2000; Herdegen andLeah 1998). Surprisingly, a single withdrawal did not giverise to reliable increases in Fos expression in any area,though trends were observed. We used an immunohisto-chemical method to visualise Fos protein, and the timecourse of expression therefore reflects the time required forthe translation and transcription processes. We have chosen8 h after withdrawal of ethanol as a time point for all ourmeasures on the basis of data showing that blood alcohollevels are similar immediately after withdrawal from SWDand RWD treatment (Stephens et al. 2001), the rate ofreduction of blood alcohol levels after withdrawal of 7%liquid diet treatment does not differ between SWD andRWD rats, and elimination is largely complete at 4 h(Overstreet et al. 2002), and correspondingly, this is a timeat which withdrawal of rats from liquid diet gives rise toseveral withdrawal signs that are both intense and stableover a few hours (Macey et al. 1996; Rassnick et al. 1992).Perhaps a later time point may have allowed Fosaccumulation to have occurred even in the SWD group.Nevertheless, the numbers of neurons expressing Fos wasreliably increased in several areas of the RWD group,reflecting the expected increases in excitability followingsensitisation, or “kindling” of withdrawal that results inincreased seizure sensitivity (Ballenger and Post 1978;Becker and Hale 1993; Brown et al. 1988; Duka et al.2004; Lechtenberg andWorner 1991; Mhatre and Gonzalez1999; Pinel et al. 1975; Veatch and Becker 2002). Areasshowing clear effects of repeated withdrawal included CeAand BLA nuclei of amygdala, n. accumbens core, CA3field of hippocampus, and the periaqueductal grey of themidbrain, though other areas showed similar, if non-significant trends. Thus, although we found no evidencethat anxiety-inducing effects of withdrawal increased withRWD, there was clear evidence for increased withdrawal-induced neuronal activation in several brain areas. Suchincreases may thus reflect the kindling of seizure activityinduced by repeated withdrawal from ethanol.
In contrast, expression of another immediate early gene,zif268, was generally insensitive to withdrawal. Never-theless, zif268 showed a reliable increase in the centralamygdala after single withdrawal but failed to increase afterrepeated withdrawal. This pattern of zif268 expression in
197

CeA parallels the corticosterone effects. Zif268 has beensuggested to be a marker of synaptic plasticity rather thanneuronal activity per se (Cole et al. 1989; Jones et al. 2001;Knapska and Kaczmarek 2004; Lee et al. 2004; Thomas etal. 2003). We have recently reported that withdrawal fromethanol results in reduced capacity of rat hippocampus andlateral amygdala to display long-term potentiation, and that,at least in hippocampus, the extent of this deficit increaseswith increased numbers of withdrawals (Stephens et al.2005). The pattern of zif268 expression in CeA, showing anincrease following a single, but not repeated withdrawal,may thus reflect withdrawal-induced plasticity after a singlewithdrawal (withdrawal non-specifically strengthens weaksynapses) but reduced capacity for plasticity followingmultiple withdrawals. That is, following several with-drawals as a result of previous strengthening, the pool ofsynapses available for strengthening with future learningmay decrease, and the room for further plasticity narrows,thus reducing the capacity for further LTP, as seen in theprevious study. These neuronal processes in the amygdalamay constitute the basis for impaired fear conditioningassociated with multiple withdrawals observed in rats(Ripley et al. 2003b; Stephens et al. 2001) and human bingedrinkers (Duka et al. 2004; Stephens et al. 2005).
This suggestion that the absence of an increase in zif268expression after RWD reflects impaired plasticity isweakened by our surprising failure to find reliable effectsof withdrawal on zif268 expression in other brain areas.However, in certain areas such as hippocampus CA1 andlateral amygdala, zif268 expression was already high in thecontrol animals, so that further increases induced bywithdrawal may have been obscured. As with c-fos, wealso do not know whether immunohistochemistry forzif268 was performed at the optimal withdrawal time.
In conclusion, the present report demonstrates that whilstwithdrawal from chronic ethanol treatment gives rise toanxiety-like behaviour in the elevated plus-maze, increasescorticosterone blood level, and zif268 expression in centralamygdala, previous experience of withdrawal from chronicethanol treatment does not exacerbate withdrawal-inducedanxiety, protects against increases in corticosterone level,possibly as a result of habituation to ethanol withdrawal-induced stress, and simultaneously gives rise to increasedneuronal activity as indicated by c-fos expression inamygdala, hippocampus, nucleus accumbens and dorsalPAG. This complex pattern indicates there is no simplerelationship between behavioural measures thought toreflect anxiety (such as the plus-maze), hormonal stressresponses and activation of brain areas involved inmediating responses to fearful stimuli.
Acknowledgement Supported by MRC Programme GrantG9806260.
References
Baker TB, Cannon DS (1979) Potentiation of ethanol withdrawal byprior dependence. Psychopharmacology (Berl) 60:105–110
Baldwin HA, Rassnick S, Rivier J, Koob GF, Britton KT (1991)CRF antagonist reverses the ‘‘anxiogenic’’ response to ethanolwithdrawal in the rat. Psychopharmacology (Berl) 103:227–232
Ballenger JC, Post RM (1978) Kindling as a model for alcoholwithdrawal syndromes. Br J Psychiatry 133:1–14
Becker HC, Hale RL (1993) Repeated episodes of ethanolwithdrawal potentiate the severity of subsequent withdrawalseizures:an animal model of alcohol withdrawal ‘‘kindling’’.Alcohol Clin Exp Res 17:94–98
Belda X, Marquez C, Armario A (2004) Long-term effects of asingle exposure to stress in adult rats on behavior andhypothalamic–pituitary–adrenal responsiveness:comparison oftwo outbred rat strains. Behav Brain Res 154:399–408
Breese GR, Overstreet DH, Knapp DJ (2005) Conceptual frame-work for the etiology of alcoholism:a ‘‘kindling’’/stresshypothesis. Psychopharmacology (Berl) 178:367–380
Brown ME, Anton RF, Malcolm R, Ballenger JC (1988) Alcoholdetoxification and withdrawal seizures:clinical support for akindling hypothesis. Biol Psychiatry 23:507–514
Carter RN, Pinnock SB, Herbert J (2004) Does the amygdalamodulate adaptation to repeated stress? Neuroscience 126:9–19
Chen X, Herbert J (1995) Regional changes in c-fos expression inthe basal forebrain and brainstem during adaptation to repeatedstress:correlations with cardiovascular, hypothermic and endo-crine responses. Neuroscience 64:675–685
Cole AJ, Saffen DW, Baraban JM, Worley PF (1989) Rapid increaseof an immediate early gene messenger RNA in hippocampalneurons by synaptic NMDA receptor activation. Nature340:474–476
Cruz AP, Frei F, Graeff FG (1994) Ethopharmacological analysis ofrat behavior on the elevated plus-maze. Pharmacol BiochemBehav 49:171–176
Dawson GR, Tricklebank MD (1995) Use of the elevated plus mazein the search for novel anxiolytic agents. Trends Pharmacol Sci16:33–36
Day HE, Nebel S, Sasse S, Campeau S (2005) Inhibition of thecentral extended amygdala by loud noise and restraint stress.Eur J Neurosci 21:441–454
Dayas CV, Buller KM, Crane JW, Xu Y, Day TA (2001) Stressorcategorization:acute physical and psychological stressors elicitdistinctive recruitment patterns in the amygdala and in med-ullary noradrenergic cell groups. Eur J Neurosci 14:1143–1152
Dere E, Topic B, De Souza Silva MA, Srejic M, Frisch C,Buddenberg T, Huston JP (2002) The graded anxiety test:anovel test of murine unconditioned anxiety based on theprinciples of the elevated plus-maze and light–dark test.J Neurosci Methods 122:65–73
DSM-IV (1994) Diagnostic and statistical manual of mental disorders,4th edn. American Psychiatric Association, Washington
Duka T, Townshend JM, Collier K, Stephens DN (2002) Kindling ofwithdrawal:a study of craving and anxiety after multipledetoxifications in alcoholic inpatients. Alcohol Clin Exp Res26:785–795
Duka T, Gentry J, Malcolm R, Ripley TL, Borlikova G, StephensDN, Veatch LM, Becker HC, Crews FT (2004) Consequencesof multiple withdrawals from alcohol. Alcohol Clin Exp Res28:233–246
Dunworth SJ, Mead AN, Stephens DN (2000) Previous experienceof withdrawal from chronic diazepam ameliorates the aver-siveness of precipitated withdrawal and reduces withdrawal-induced c-fos expression in nucleus accumbens. Eur J Neurosci12:1501–1508
File SE (1980) The use of social interaction as a method fordetecting anxiolytic activity of chlordiazepoxide-like drugs.J Neurosci Methods 2:219–238
File SE (1992) Behavioural detection of anxiolitic action. In:Marsden CA (ed) Experimental approaches to anxiety anddepression. Willey, Chichester, pp 25–44
File SE, Hyde JR (1978) Can social interaction be used to measureanxiety? Br J Pharmacol 62:19–24
File SE, Seth P (2003) A review of 25 years of the social interactiontest. Eur J Pharmacol 463:35–53
198

File SE, Andrews N, al-Farhan M (1993) Anxiogenic responses ofrats on withdrawal from chronic ethanol treatment:effects oftianeptine. Alcohol Alcohol 28:281–286
File SE, Gonzalez LE, Gallant R (1999) Role of the dorsomedialhypothalamus in mediating the response to benzodiazepines ontrial 2 in the elevated plus-maze test of anxiety. Neuropsycho-pharmacology 21:312–320
Gatch MB, Wallis CJ, Lal H (1999) Effects of NMDA antagonistson ethanol-withdrawal induced ‘‘anxiety’’ in the elevated plusmaze. Alcohol 19:207–211
Gatch MB, Wallis CJ, Lal H (2000) Effects of ritanserin on ethanolwithdrawal-induced anxiety in rats. Alcohol 21:11–17
Ghozland S, Chu K, Kieffer BL, Roberts AJ (2005) Lack ofstimulant and anxiolytic-like effects of ethanol and accelerateddevelopment of ethanol dependence in mu-opioid receptorknockout mice. Neuropharmacology 49:493–501
Handley SL, Mithani S (1984) Effects of alpha-adrenoceptoragonists and antagonists in a maze-exploration model of‘fear’-motivated behaviour. Naunyn Schmiedebergs ArchPharmacol 327:1–5
Herdegen T, Leah JD (1998) Inducible and constitutive transcriptionfactors in the mammalian nervous system:control of geneexpression by Jun, Fos and Krox, and CREB/ATF proteins.Brain Res Brain Res Rev 28:370–490
Herman JP, Cullinan WE (1997) Neurocircuitry of stress:central control of the hypothalamo–pituitary–adrenocorticalaxis. Trends Neurosci 20:78–84
Herman JP, Dolgas CM, Carlson SL (1998) Ventral subiculumregulates hypothalamo–pituitary–adrenocortical and behaviour-al responses to cognitive stressors. Neuroscience 86:449–459
Holter SM, Engelmann M, Kirschke C, Liebsch G, Landgraf R,Spanagel R (1998) Long-term ethanol self-administration withrepeated ethanol deprivation episodes changes ethanol drinkingpattern and increases anxiety-related behaviour during ethanoldeprivation in rats. Behav Pharmacol 9:41–48
Janis GC, Devaud LL, Mitsuyama H, Morrow AL (1998) Effects ofchronic ethanol consumption and withdrawal on the neuroac-tive steroid 3alpha-hydroxy-5alpha-pregnan-20-one in maleand female rats. Alcohol Clin Exp Res 22:2055–2061
Jones MW, Errington ML, French PJ, Fine A, Bliss TV, Garel S,Charnay P, Bozon B, Laroche S, Davis S (2001) A requirementfor the immediate early gene Zif268 in the expression of lateLTP and long-term memories. Nat Neurosci 4:289–296
Keay KA, Clement CI, Depaulis A, Bandler R (2001) Differentrepresentations of inescapable noxious stimuli in the periaque-ductal gray and upper cervical spinal cord of freely moving rats.Neurosci Lett 313:17–20
Kliethermes CL (2005) Anxiety-like behaviors following chronicethanol exposure. Neurosci Biobehav Rev 28:837–850
Kliethermes CL, Cronise K, Crabbe JC (2005) Home cage activityand ingestive behaviors in mice following chronic ethanolvapor inhalation. Physiol Behav 85:479–488
Knapska E, Kaczmarek L (2004) A gene for neuronal plasticity inthe mammalian brain:Zif268/Egr-1/NGFI-A/Krox-24/TIS8/ZENK? Prog Neurobiol 74:183–211
Koss WA, Gehlert DR, Shekhar A (2004) Different effects ofsubchronic doses of 17-beta estradiol in two ethologicallybased models of anxiety utilizing female rats. Horm Behav46:158–164
Lechtenberg R, Worner TM (1991) Relative kindling effect ofdetoxification and non-detoxification admissions in alcoholics.Alcohol Alcohol 26:221–225
LeDoux JE (2000) Emotion circuits in the brain. Annu RevNeurosci 23:155–184
Lee JL, Everitt BJ, Thomas KL (2004) Independent cellularprocesses for hippocampal memory consolidation and reconsol-idation. Science 304:839–843
Linden AM, Greene SJ, Bergeron M, Schoepp DD (2004)Anxiolytic activity of the MGLU2/3 receptor agonistLY354740 on the elevated plus maze is associated with thesuppression of stress-induced c-Fos in the hippocampus andincreases in c-Fos induction in several other stress-sensitivebrain regions. Neuropsychopharmacology 29:502–513
Macey DJ, Schulteis G, Heinrichs SC, Koob GF (1996) Time-dependent quantifiable withdrawal from ethanol in the rat:effectof method of dependence induction. Alcohol 13:163–170
Malcolm R, Myrick H, Roberts J, Wang W, Anton RF (2002) Thedifferential effects of medication on mood, sleep disturbance,and work ability in outpatient alcohol detoxification. Am JAddict 11:141–150
Martinez M, Phillips PJ, Herbert J (1998) Adaptation in patterns ofc-fos expression in the brain associated with exposure to eithersingle or repeated social stress in male rats. Eur J Neurosci10:20–33
Melia KR, Ryabinin AE, Schroeder R, Bloom FE, Wilson MC(1994) Induction and habituation of immediate early geneexpression in rat brain by acute and repeated restraint stress.J Neurosci 14:5929–5938
Mhatre MC, Gonzalez LP (1999) Increased Ro15-4513 inducedseizures following multiple ethanol withdrawals. PharmacolBiochem Behav 63:93–99
Mutschler NH, Miczek KA, Hammer RP Jr (2000) Reduction ofzif268 messenger RNA expression during prolonged with-drawal following ‘‘binge’’ cocaine self-administration in rats.Neuroscience 100:531–538
Overstreet DH, Knapp DJ, Breese GR (2002) Accentuated decreasein social interaction in rats subjected to repeated ethanolwithdrawals. Alcohol Clin Exp Res 26:1259–1268
Overstreet DH, Knapp DJ, Moy SS, Breese GR (2003) A 5-HT1Aagonist and a 5-HT2c antagonist reduce social interactiondeficit induced by multiple ethanol withdrawals in rats.Psychopharmacology (Berl) 167:344–352
Overstreet DH, Knapp DJ, Breese GR (2004a) Modulation ofmultiple ethanol withdrawal-induced anxiety-like behaviorby CRF and CRF1 receptors. Pharmacol Biochem Behav77:405–413
Overstreet DH, Knapp DJ, Breese GR (2004b) Similar anxiety-likeresponses in male and female rats exposed to repeatedwithdrawals from ethanol. Pharmacol Biochem Behav78:459–464
Paxinos W, Watson C (1998) The rat brain in stereotaxiccoordinates, 4th edn. Academic, New York
Pellow S, Chopin P, File SE, Briley M (1985) Validation of open:closed arm entries in an elevated plus-maze as a measure ofanxiety in the rat. J Neurosci Methods 14:149–167
Pinel JP, Van Oot PH, Mucha RF (1975) Intensification of thealcohol withdrawal syndrome by repeated brain stimulation.Nature 254:510–511
Rassnick S, Koob GF, Geyer MA (1992) Responding to acousticstartle during chronic ethanol intoxication and withdrawal.Psychopharmacology (Berl) 106:351–358
Ripley TL, Brown G, Dunworth SJ, Stephens DN (2003a) Aversiveconditioning following repeated withdrawal from ethanol andepileptic kindling. Eur J Neurosci 17:1664–1670
Ripley TL, O’Shea M, Stephens DN (2003b) Repeated withdrawalfrom ethanol impairs acquisition but not expression of condi-tioned fear. Eur J Neurosci 18:441–448
Ripley TL, Borlikova G, Lyons S, Stephens DN (2004) Selectivedeficits in appetitive conditioning as a consequence of ethanolwithdrawal. Eur J Neurosci 19:415–425
Rodgers RJ, Haller J, Holmes A, Halasz J, Walton TJ, Brain PF(1999) Corticosterone response to the plus-maze:high correla-tion with risk assessment in rats and mice. Physiol Behav68:47–53
Salome N, Salchner P, Viltart O, Sequeira H, Wigger A, Landgraf R,Singewald N (2004) Neurobiological correlates of high (HAB)versus low anxiety-related behavior (LAB):differential Fosexpression in HAB and LAB rats. Biol Psychiatry 55:715–723
199

Senba E, Ueyama T (1997) Stress-induced expression of immediateearly genes in the brain and peripheral organs of the rat.Neurosci Res 29:183–207
Singewald N, Salchner P, Sharp T (2003) Induction of c-Fosexpression in specific areas of the fear circuitry in rat forebrainby anxiogenic drugs. Biol Psychiatry 53:275–283
Stamp JA, Herbert J (1999) Multiple immediate-early gene expres-sion during physiological and endocrine adaptation to repeatedstress. Neuroscience 94:1313–1322
Stephens DN, Brown G, Duka T, Ripley TL (2001) Impaired fearconditioning but enhanced seizure sensitivity in rats givenrepeated experience of withdrawal from alcohol. Eur J Neurosci14:2023–2031
Stephens DN, Ripley TL, Borlikova G, Schubert M, Albrecht D,Hogarth L, Duka T (2005) Repeated ethanol exposure andwithdrawal impairs human fear conditioning and depresseslong-term potentiation in rat amygdala and hippocampus. BiolPsychiatry 58:392–400
Tabakoff B, Jafee RC, Ritzmann RF (1978) Corticosteroneconcentrations in mice during ethanol drinking and withdrawal.J Pharm Pharmacol 30:371–374
Thomas KL, Arroyo M, Everitt BJ (2003) Induction of the learningand plasticity-associated gene Zif268 following exposure to adiscrete cocaine-associated stimulus. Eur J Neurosci 17:1964–1972
Townshend JM, Duka T (2003) Mixed emotions:alcoholics’impairments in the recognition of specific emotional facialexpressions. Neuropsychologia 41:773–782
Townshend JM, Duka T (2005) Binge drinking, cognitiveperformance and mood in a population of young socialdrinkers. Alcohol Clin Exp Res 29:317–325
Umemoto S, Kawai Y, Ueyama T, Senba E (1997) Chronicglucocorticoid administration as well as repeated stress affectsthe subsequent acute immobilization stress-induced expressionof immediate early genes but not that of NGFI-A. Neuroscience80:763–773
Veatch LM, Becker HC (2002) Electrographic and behavioral indicesof ethanol withdrawal sensitization. Brain Res 946:272–282
Ward BO, Stephens DN (1998) Sensitisation of withdrawal signsfollowing repeated withdrawal from a benzodiazepine:differ-ences between measures of anxiety and seizure sensitivity.Psychopharmacology (Berl) 135:342–352
Watson WP, Robinson E, Little HJ (1997) The novel anticonvulsant,gabapentin, protects against both convulsant and anxiogenicaspects of the ethanol withdrawal syndrome. Neuropharmacol-ogy 36:1369–1375
Wilson J, Watson WP, Little HJ (1998) CCK(B) antagonists protectagainst anxiety-related behaviour produced by ethanol with-drawal, measured using the elevated plus maze. Psychophar-macology (Berl) 137:120–131
200




















