
Quantitative Two-Process Analysis of Avoidance Conditioning in Goldfish
12
Journal of Experimental Psychology: Animal Behavior Processes 1994, Vol. 20. No. 1,32^(3 Copyright 1994 by the American Psychological Association, Inc. 0097-7403/94/53.00 Quantitative Two-Process Analysis of Avoidance Conditioning in Goldfish A. Y. Zhuikov, P. A. Couvillon, and M. E. Bitterman The shuttlebox performance of goldfish was studied under standardized conditions in a variety of problems—with or without an avoidance contingency, a conditioned stimulus (CS)-termination contingency, and an escape contingency. The effects of CS-only, unconditioned stimulus (US)-only, and explicitly unpaired training were also examined. All the data could be simulated quantitatively with a version of O. H. Mowrer's (1947) 2-process theory expressed in 2 learning equations (1 classical, the other instrumental) and a performance equation. The good fit suggests that the theory is worth developing further with new experiments designed to challenge it. Familiar objections to Mowrer's (1947) early two-process theory of avoidance conditioning were recently reviewed by Levis (1989), in whose opinion they reflect misunderstand- ings of the theory that are attributable at least in part to the loose language in which the theory has been expressed. Our purpose in this work is to explore the possibilities of a math- ematical formulation of the theory designed to yield entirely unambiguous predictions of experimental outcomes. Efforts at quantitative simulation of real avoidance data on the basis of theories resembling Mowrer's have been made in the past (Atkinson, Bower, & Crothers, 1965; Bush & Mosteller, 1955; Hoffman, 1965; Theios, 1963) but were not carried very far. What is required first in such a project is a large and readily expandable database, and for that reason we turned to shuttlebox experiments with fishes. Fishes are inexpen- sive to acquire and to maintain properly, and the shut- tlebox technique developed for them (Horner, Longo, & Bitterman, 1961) is efficient, easily standardized, and well- suited to a variety of species (Klinman & Bitterman, 1963; Leschova & Zhuikov, 1989; Wodinsky, Behrend, & Bitter- man, 1962). Another important advantage of the technique is that shuttling, at least in goldfish, is produced by purely classical as well as instrumental conditioning (Woodard & Bitterman, 1971, 1973). Although there is much to be gained from the study of avoidance responses different from, and even largely incompatible with, responses that are classically conditioned with the same aversive stimulus (Brogden, Lipman, & Culler, 1938; Hunter, 1935; Kamin, A. Y. Zhuikov, Severtsov Institute of Evolutionary Animal Mor- phology and Ecology, Moscow, Russia; P. A. Couvillon and M. E. Bitterman, Be'ke'sy Laboratory of Neurobiology, University of Hawaii. This research was supported by Research Centers for Minority Institutions Grant RR03061 from the National Institutes of Health. The collaboration of A. Y. Zhuikov was made possible by Fulbright Grant 15405. We thank F. Robert Brush, Howard Hoffman, J. Bruce Overmier, and J. E. Sigurdson for helpful suggestions. Correspondence concerning this article should be addressed to M. E. Bitterman, Bekesy Laboratory of Neurobiology, 1993 East- West Road, Honolulu, Hawaii 96822. Electronic mail may be sent to [email protected]. 1956), it may be more instructive at the outset, and cer- tainly much simpler, to use a situation in which classical and instrumental influences are indexed by the same re- sponse (Gormezano, 1965; Logan, 1951). A Quantitative Restatement of Two-Process Avoidance Theory The first main premise of the theory is that pairing a neutral stimulus with shock (Pavlovian reinforcement) results in the formation of an association between them. In consequence of that association and to an extent proportional to its strength (c), scaled from 0 to 1, the conditioned stimulus (CS) has two properties: It is aversive (Mowrer's conditioned fear), and it tends to evoke shuttling. Change in c with reinforcement and nonreinforcement (presentation of the CS without shock) is described by the equation, Ac = /3 C (A C - c), (1) with Ac representing the change in c, /3 C the rate of change, and A c the asymptotic value of c, which is determined by the functional intensity of shock (s). When (A c - c) > 0, c in- creases, and c decreases when (A c - c) < 0, with the pos- sibility considered that the rate of increase (the incremental /3 or t/p c ) and the rate of decrease (the decremental /3 or D/3 C ) are different. No inhibitory process is assumed. The second main premise of the theory is that an asso- ciation between the CS and the shuttling response develops when shuttling in the presence of the CS is followed by re- duction in the discomfort generated by the CS (Hullian re- inforcement). In consequence of that association, too, and again in proportion to its strength (i), scaled from 0 to 1, the CS tends to evoke shuttling. Change in i with reinforcement and nonreinforcement is described by the equation, Ai =ft(A,- i). (2) with At representing the change in i, and 0 ; the rate of change, the incremental and decremental values of which (t/j3, and Dft) may differ from each other as well as from Ufi c and D/3 C . When (A, - i) > 0, i increases, and it decreases when (A, - 0 < 0. Scaled from 0 to 1, A, is the asymptotic value of i, which is determined by the magnitude of reinforcement. 32
Transcript of Quantitative Two-Process Analysis of Avoidance Conditioning in Goldfish

Journal of Experimental Psychology:Animal Behavior Processes1994, Vol. 20. No. 1,32^(3
Copyright 1994 by the American Psychological Association, Inc.0097-7403/94/53.00
Quantitative Two-Process Analysis of Avoidance Conditioning in Goldfish
A. Y. Zhuikov, P. A. Couvillon, and M. E. Bitterman
The shuttlebox performance of goldfish was studied under standardized conditions in a variety ofproblems—with or without an avoidance contingency, a conditioned stimulus (CS)-terminationcontingency, and an escape contingency. The effects of CS-only, unconditioned stimulus (US)-only,and explicitly unpaired training were also examined. All the data could be simulated quantitativelywith a version of O. H. Mowrer's (1947) 2-process theory expressed in 2 learning equations (1classical, the other instrumental) and a performance equation. The good fit suggests that the theoryis worth developing further with new experiments designed to challenge it.
Familiar objections to Mowrer's (1947) early two-processtheory of avoidance conditioning were recently reviewed byLevis (1989), in whose opinion they reflect misunderstand-ings of the theory that are attributable at least in part to theloose language in which the theory has been expressed. Ourpurpose in this work is to explore the possibilities of a math-ematical formulation of the theory designed to yield entirelyunambiguous predictions of experimental outcomes. Effortsat quantitative simulation of real avoidance data on the basisof theories resembling Mowrer's have been made in the past(Atkinson, Bower, & Crothers, 1965; Bush & Mosteller,1955; Hoffman, 1965; Theios, 1963) but were not carriedvery far.
What is required first in such a project is a large andreadily expandable database, and for that reason we turnedto shuttlebox experiments with fishes. Fishes are inexpen-sive to acquire and to maintain properly, and the shut-tlebox technique developed for them (Horner, Longo, &Bitterman, 1961) is efficient, easily standardized, and well-suited to a variety of species (Klinman & Bitterman, 1963;Leschova & Zhuikov, 1989; Wodinsky, Behrend, & Bitter-man, 1962). Another important advantage of the techniqueis that shuttling, at least in goldfish, is produced by purelyclassical as well as instrumental conditioning (Woodard &Bitterman, 1971, 1973). Although there is much to begained from the study of avoidance responses differentfrom, and even largely incompatible with, responses thatare classically conditioned with the same aversive stimulus(Brogden, Lipman, & Culler, 1938; Hunter, 1935; Kamin,
A. Y. Zhuikov, Severtsov Institute of Evolutionary Animal Mor-phology and Ecology, Moscow, Russia; P. A. Couvillon and M. E.Bitterman, Be'ke'sy Laboratory of Neurobiology, University ofHawaii.
This research was supported by Research Centers for MinorityInstitutions Grant RR03061 from the National Institutes of Health.The collaboration of A. Y. Zhuikov was made possible by FulbrightGrant 15405. We thank F. Robert Brush, Howard Hoffman, J.Bruce Overmier, and J. E. Sigurdson for helpful suggestions.
Correspondence concerning this article should be addressed toM. E. Bitterman, Bekesy Laboratory of Neurobiology, 1993 East-West Road, Honolulu, Hawaii 96822. Electronic mail may be sentto [email protected].
1956), it may be more instructive at the outset, and cer-tainly much simpler, to use a situation in which classicaland instrumental influences are indexed by the same re-sponse (Gormezano, 1965; Logan, 1951).
A Quantitative Restatement of Two-ProcessAvoidance Theory
The first main premise of the theory is that pairing a neutralstimulus with shock (Pavlovian reinforcement) results in theformation of an association between them. In consequenceof that association and to an extent proportional to its strength(c), scaled from 0 to 1, the conditioned stimulus (CS) has twoproperties: It is aversive (Mowrer's conditioned fear), and ittends to evoke shuttling. Change in c with reinforcement andnonreinforcement (presentation of the CS without shock) isdescribed by the equation,
Ac = /3C(AC - c), (1)
with Ac representing the change in c, /3C the rate of change,and Ac the asymptotic value of c, which is determined by thefunctional intensity of shock (s). When (Ac - c) > 0, c in-creases, and c decreases when (Ac - c) < 0, with the pos-sibility considered that the rate of increase (the incremental/3 or t/pc) and the rate of decrease (the decremental /3 or D/3C)are different. No inhibitory process is assumed.
The second main premise of the theory is that an asso-ciation between the CS and the shuttling response developswhen shuttling in the presence of the CS is followed by re-duction in the discomfort generated by the CS (Hullian re-inforcement). In consequence of that association, too, andagain in proportion to its strength (i), scaled from 0 to 1, theCS tends to evoke shuttling. Change in i with reinforcementand nonreinforcement is described by the equation,
Ai = ft(A, - i). (2)
with At representing the change in i, and 0; the rate of change,the incremental and decremental values of which (t/j3, andDft) may differ from each other as well as from Ufic and D/3C.When (A, - i) > 0, i increases, and it decreases when (A, -0 < 0. Scaled from 0 to 1, A, is the asymptotic value of i,which is determined by the magnitude of reinforcement.
32

QUANTITATIVE ANALYSIS OF AVOIDANCE CONDITIONING 33
Magnitude of reinforcement is defined as the extent to whichthe aversiveness of the antedating situation is reduced. Inavoidance experiments, in some of which the CS is turnedoff by the response and in others of which it is changed bythe addition of a "safety signal" or by feedback from theresponse alone, A, = (c - g X c); g (scaled from 0 to 1) isa measure of the similarity of the sensory conditions fol-lowing the response (CS') to those preceding it (CS) andhence the extent to which c generalizes from CS to CS',which itself is never paired with shock (Bower, Starr, & Laz-arovitz, 1965). Where g is large, the magnitude of reinforce-ment is small; where the response terminates the CS, g istaken as 0. It may be well to emphasize that no inhibition offear is assumed (cf. Mackintosh, 1983), only reduced exci-tation. In classical conditioning (where shock occurs inde-pendently of shuttling), A., = 0 whatever the value of g, be-cause CS and CS' are not differentially paired with shock.
In some experiments (e.g., Hunter, 1935), the programmedshock is brief and terminates independently of response,leaving /' unchanged. In others (e.g., Warner, 1932), there isan escape contingency; that is, the shock, once turned on,remains on until the animal responds. Where shock is ter-minated by shuttling, Ai is computed with A, taken as thefunctional intensity of the shock (s). In the first of the presentexperiments, one group of animals had classical training inthe sense that avoidance was impossible, but there was anescape contingency that was expected to permit the devel-opment of i.
The third main premise of the theory is that the probabilityof a shuttling response (R) in the presence of the CS is a jointfunction of three probabilities: one (C) associated with c, asecond (/) with i, and a basal shuttling tendency ([/). Noeffort is made at the outset to decompose U, which may bein part unconditioned and in part a product of contextualconditioning. It is simply defined as the probability of re-sponding in a period of time equal to the CS-US interval, thetime between the onset of the CS and the onset of the un-conditioned stimulus (US), which is sampled repeatedly inthe intertrial intervals of each experimental session.
The probabilities, C, I, and U, are summed in the con-ventional manner (Feller, 1950) with the equation
R = C+I+U - CXI - CXU - IXU + CXIXU, (3)
which, of course, restricts values of R to the range of 0 to 1.Mowrer (1947) assumed that fear is required both for in-strumental conditioning and for the activation of the instru-mental response, or, in our terms, that i is potentiated by c(cf. Hoffman, 1965). There is some reason, however, to be-lieve that although the development of i depends on c, i, oncedeveloped, may contribute to R independently of c (Mack-intosh, 1983), and the simplicity of the autonomy assumptionrecommends its consideration. For the reason of simplicityalso, no parameters are introduced to shape the contributionsof c and i to performance; as a first approximation to reality,C is taken as c, and / as i.
To compute R, it is necessary to evaluate the six parameters(t/ft, Dfic, Ufa D&i, S, and g) that affect the growth anddecline of c and /', and it was to that end that we directed the
present experiments. Our strategy was to collect some dataon shuttlebox performance in a wide range of problems in thehope that a fitting process would permit reasonable estimatesof the parameter values. Those estimates would permit rig-orous quantitative predictions from the theory that we couldthen test in further experiments. We also had to consider thepossibility that no reasonably good fit to the data would befound with any set of parameter values, which would showthe theory to be deficient in one or more respects and raisethe question of how it might be improved.
We should note that new experiments were required be-cause the data in the literature are fragmentary and some-times inconsistent, probably because they were obtained un-der different circumstances. To appreciate the importance ofa standardized training environment in which quantitativerather than merely ordinal prediction is to be attempted, it isnecessary to consider only how performance may be ex-pected to vary, for example, with the size of the shuttleboxcompartments, with the difficulty of response, and with theway in which shock is delivered. The four experiments re-ported here provide a large block of data obtained under thesame, precisely described conditions.
Experiment 1
The purpose of this experiment was to sample the behaviorof subjects in a wide variety of problems in our standardizedtraining environment. We used both classical and avoidanceprocedures, with and without an escape contingency, and inthe avoidance case, with and without a CS-termination con-tingency. Probability of reinforcement was varied in the clas-sical case, and repeated conditioning and extinction werestudied in both cases.
Method
Subjects. The subjects were 24 10-cm goldfish (Carassius au-ratus), all experimentally naive. They were maintained in indi-vidual enclosures made by partitioning large tanks, the water ofwhich was continuously aerated, filtered, and recirculated with par-tial replacement. A 24-hr feeding schedule was used. Note that it isespecially important in such experiments to minimize variation inthe size of the subjects. Given the fixed level of water over thehurdle, both the effortfulness of response and the salience of thefeedback from response may increase with size. The effectivenessof shock, which depends on the voltage drop across the animal, alsoincreases with size.
Apparatus. The animals were trained in a set of four identicalshuttleboxes of the same general design as those described by Hor-ner et al. (1961), although substantially larger in size (Figure 1).Each box was constructed entirely of black Plexiglas except for thewalls of milk-Plexiglas between the animal's compartment and thelight-boxes at either end. The light-boxes contained colored 7.5-WChristmas tree lamps that served as CSs. The electrodes used forshock were stainless steel rods, 3 mm in diameter, set against thelong walls. As in the earlier model of the apparatus, there werephotodetectors on both sides of the hurdle, which made it possibleto define response as a complete crossing from one end of the

34 A. ZHUIKOV, P. COUVILLON, AND M. BITTERMAN
PHOTOEMITTERSELECTRODE
n n 1
CS LAMPS
O/
\0
1 I1
111
o\
/o
CS LAMPS
PHOTODETECTORS
ELECTRODE
-
CS LAMPS
MWATER LEVEL y
• • | CS LAMPS
^/HURDLE^^.
u T
Figure 1. Diagram of the apparatus, an overhead view in the toppanel and a side view below. The arrows show the inflows andoutflows of the water circulating through the animal's compart-ment. The opaque cover is not shown.
shuttlebox to the other; a response was registered only when thesubject had passed through both photobeams and not by mere ap-proach to the hurdle. Continuously circulating through each cham-ber was water coming from and returning to the same system thatsupplied the living tanks. A single desktop computer programmedthe stimuli and monitored behavior in all four shuttleboxes. (Allexperiments were fully automated.)
Procedure. Two groups of 12 animals, each group divided into3 subsets of 4 animals, were given three daily 40-min periods ofadjustment to the apparatus. This pretraining was followed by 114daily training sessions; in each session there were 20 trials with amean intertrial interval of 90 s. Of the 20 trials, 10 were widely
separated blank trials (each providing a 10-s sample of activity) and10 were CS trials (scheduled in balanced quasirandom orders). TheCS was general illumination provided by red lamps turned on si-multaneously at both ends of the shuttlebox. The mean intervalbetween CS presentations was 180 s, with a range of 90 to 290 s.
The training of one group was classical in that the US was turnedon 10 s after the onset of the CS regardless of whether a shuttlingresponse occurred in that interval, but to begin with there was anescape contingency as well (classical-escape or C/E training). TheUS was a train of .25-s pulses of ac shock (.33 v/cm) delivered atintervals of 2.5 s across the long walls of the shuttlebox; the CSremained on until it was terminated along with the train of shockpulses by the first shuttling response following the 10-s CS-USinterval, or after 30 s. After 10 sessions (see Figure 2 for details),the animals were shifted to purely classical (C) training for 15 ses-sions, that is, the escape contingency was removed; each 10-s CSterminated with a single .25-s pulse of shock. In the subsequent 21sessions, the percentage of reinforcement was reduced progres-sively to 60% for 2 sessions, to 40% for 2 sessions, and to 30% forthe rest (classical-partial or C/P training); with 30% reinforcement,the frequency of shock was almost as low as in an avoidance groupthat was trained concurrently. A series of 18 extinction sessionsfollowed, after which there were 5 series of 5 reacquisition sessions(with 100% reinforcement) alternating with 5 series of 5 extinctionsessions.
The second group had avoidance training. In the first 10 sessions,the CR both terminated the CS and avoided the US, and on trialson which the CR did not occur, the escape contingency was in force(avoidance-escape or A/E training). In the next 5 sessions (see Fig-ure 2), the escape contingency was eliminated (pure avoidance orA training) and then, for 31 sessions, the CS-termination contin-gency was eliminated as well, that is, the CR avoided shock but didnot terminate the CS, which stayed on during the scheduled 10-s CSinterval (A/N training). As with the classical group, a series of 18extinction sessions followed in which there still was no CS-termination contingency (making the extinction conditions exactlythe same for both groups), and then there were 5 series of 5 reac-quisition (A/N) sessions alternating with 5 series of 5 extinctionsessions.
(REPEATED ACQ AND EXT)
•-• CLASSICAL
O-O AVOIDANCE
10 20 30 40 50 60 70 80 90 100 110
SESSIONS
Figure 2. The session-by-session performance of the two groups under the various conditions oftraining in Experiment 1. A/E = avoidance training with CS-termination and escape; C/E = classicaltraining with escape; A = avoidance training with CS-termination; C = classical training; A/N =avoidance training without a CS-termination contingency; C/P = classical training with partialreinforcement; EXT = extinction; ACQ = acquisition.
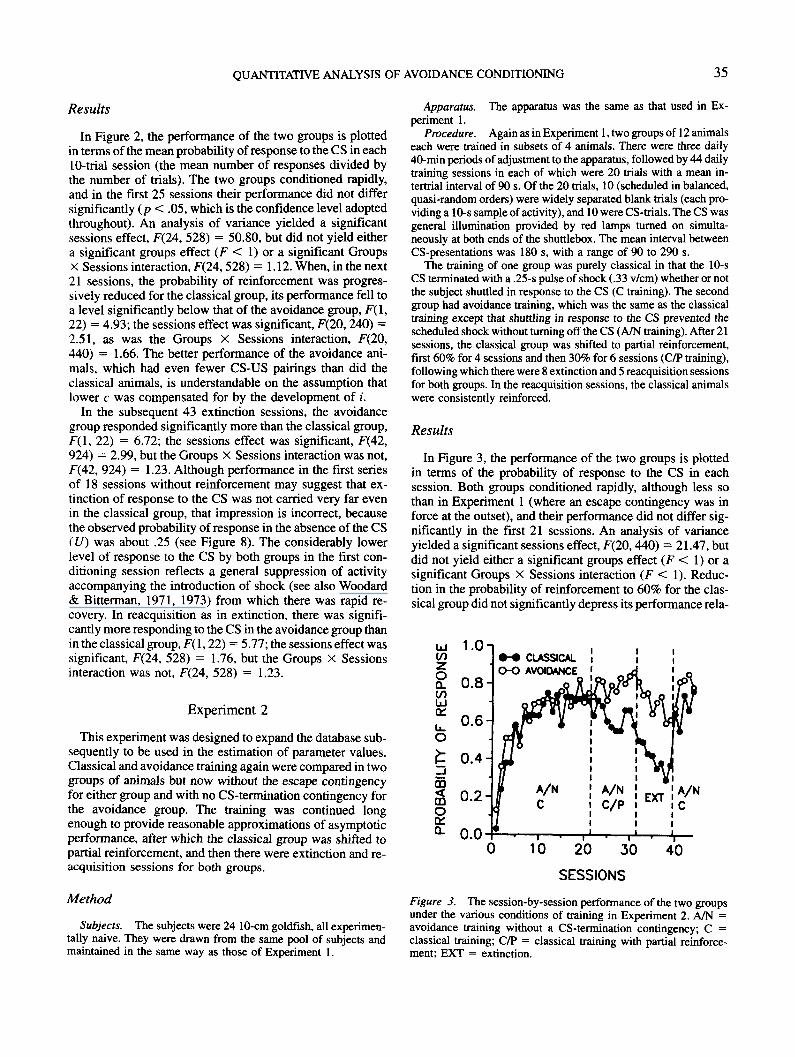
QUANTITATIVE ANALYSIS OF AVOIDANCE CONDITIONING 35
Results
In Figure 2, the performance of the two groups is plottedin terms of the mean probability of response to the CS in eachLO-trial session (the mean number of responses divided bythe number of trials). The two groups conditioned rapidly,and in the first 25 sessions their performance did not differsignificantly (p < .05, which is the confidence level adoptedthroughout). An analysis of variance yielded a significantsessions effect, F(24, 528) = 50.80, but did not yield eithera significant groups effect (F < 1) or a significant GroupsX Sessions interaction, F(24,528) = 1.12. When, in the next21 sessions, the probability of reinforcement was progres-sively reduced for the classical group, its performance fell toa level significantly below that of the avoidance group, F(l,22) = 4.93; the sessions effect was significant, F(20,240) =2.51, as was the Groups X Sessions interaction, F(20,440) = 1.66. The better performance of the avoidance ani-mals, which had even fewer CS-US pairings than did theclassical animals, is understandable on the assumption thatlower c was compensated for by the development of i.
In the subsequent 43 extinction sessions, the avoidancegroup responded significantly more than the classical group,F(l, 22) = 6.72; the sessions effect was significant, F(42,924) = 2.99, but the Groups X Sessions interaction was not,F(42, 924) = 1.23. Although performance in the first seriesof 18 sessions without reinforcement may suggest that ex-tinction of response to the CS was not carried very far evenin the classical group, that impression is incorrect, becausethe observed probability of response in the absence of the CS(U) was about .25 (see Figure 8). The considerably lowerlevel of response to the CS by both groups in the first con-ditioning session reflects a general suppression of activityaccompanying the introduction of shock (see also Woodard& Bitterman, 1971, 1973) from which there was rapid re-covery. In reacquisition as in extinction, there was signifi-cantly more responding to the CS in the avoidance group thanin the classical group, F(l, 22) = 5.77; the sessions effect wassignificant, F(24, 528) = 1.76, but the Groups X Sessionsinteraction was not, F(24, 528) = 1.23.
Experiment 2
This experiment was designed to expand the database sub-sequently to be used in the estimation of parameter values.Classical and avoidance training again were compared in twogroups of animals but now without the escape contingencyfor either group and with no CS-termination contingency forthe avoidance group. The training was continued longenough to provide reasonable approximations of asymptoticperformance, after which the classical group was shifted topartial reinforcement, and then there were extinction and re-acquisition sessions for both groups.
Method
Subjects. The subjects were 24 10-cm goldfish, all experimen-tally naive. They were drawn from the same pool of subjects andmaintained in the same way as those of Experiment 1.
Apparatus. The apparatus was the same as that used in Ex-periment 1.
Procedure. Again as in Experiment 1, two groups of 12 animalseach were trained in subsets of 4 animals. There were three daily40-min periods of adjustment to the apparatus, followed by 44 dailytraining sessions in each of which were 20 trials with a mean in-tertrial interval of 90 s. Of the 20 trials, 10 (scheduled in balanced,quasi-random orders) were widely separated blank trials (each pro-viding a 10-s sample of activity), and 10 were CS-trials. The CS wasgeneral illumination provided by red lamps turned on simulta-neously at both ends of the shuttlebox. The mean interval betweenCS-presentations was 180 s, with a range of 90 to 290 s.
The training of one group was purely classical in that the 10-sCS terminated with a .25-s pulse of shock (.33 v/cm) whether or notthe subject shuttled in response to the CS (C training). The secondgroup had avoidance training, which was the same as the classicaltraining except that shuttling in response to the CS prevented thescheduled shock without turning off the CS (A/N training). After 21sessions, the classical group was shifted to partial reinforcement,first 60% for 4 sessions and then 30% for 6 sessions (C/P training),following which there were 8 extinction and 5 reacquisition sessionsfor both groups. In the reacquisition sessions, the classical animalswere consistently reinforced.
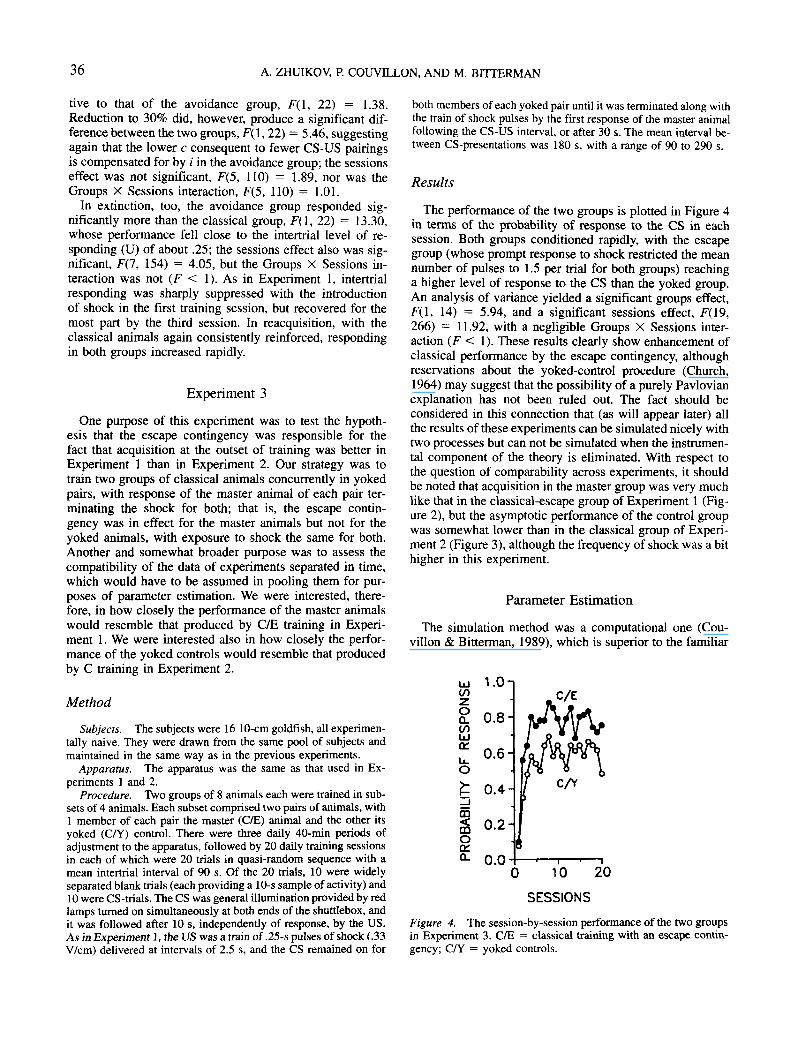
Results
In Figure 3, the performance of the two groups is plottedin terms of the probability of response to the CS in eachsession. Both groups conditioned rapidly, although less sothan in Experiment 1 (where an escape contingency was inforce at the outset), and their performance did not differ sig-nificantly in the first 21 sessions. An analysis of varianceyielded a significant sessions effect, F(20,440) = 21.47, butdid not yield either a significant groups effect (F < 1) or asignificant Groups X Sessions interaction (F < 1). Reduc-tion in the probability of reinforcement to 60% for the clas-sical group did not significantly depress its performance rela-
UJ 1.0-V)
I 0.8-CO
o:U.o
0.6-
0.4-
m< 0.2-oo:°- 0.0
CLASSICAL iO-O AVOIDANCE
10 20 30 40
SESSIONS
Figure 3. The session-by-session performance of the two groupsunder the various conditions of training in Experiment 2. A/N =avoidance training without a CS-termination contingency; C =classical training; C/P = classical training with partial reinforce-ment; EXT = extinction.
36 A. ZHUIKOV, P. COUVILLON, AND M. BITTERMAN
live to that of the avoidance group, F(l, 22) = 1.38.Reduction to 30% did, however, produce a significant dif-ference between the two groups, F(l, 22) = 5.46, suggestingagain that the lower c consequent to fewer CS-US pairingsis compensated for by i in the avoidance group; the sessionseffect was not significant, F(5, 110) = 1.89, nor was theGroups X Sessions interaction, F(5, 110) = 1.01.
In extinction, too, the avoidance group responded sig-nificantly more than the classical group, F(l, 22) = 13.30,whose performance fell close to the intertrial level of re-sponding (U) of about .25; the sessions effect also was sig-nificant, F(7, 154) = 4.05, but the Groups X Sessions in-teraction was not (F < 1). As in Experiment 1, intertrialresponding was sharply suppressed with the introductionof shock in the first training session, but recovered for themost part by the third session. In reacquisition, with theclassical animals again consistently reinforced, respondingin both groups increased rapidly.
Experiment 3
One purpose of this experiment was to test the hypoth-esis that the escape contingency was responsible for thefact that acquisition at the outset of training was better inExperiment 1 than in Experiment 2. Our strategy was totrain two groups of classical animals concurrently in yokedpairs, with response of the master animal of each pair ter-minating the shock for both; that is, the escape contin-gency was in effect for the master animals but not for theyoked animals, with exposure to shock the same for both.Another and somewhat broader purpose was to assess thecompatibility of the data of experiments separated in time,which would have to be assumed in pooling them for pur-poses of parameter estimation. We were interested, there-fore, in how closely the performance of the master animalswould resemble that produced by C/E training in Experi-ment 1. We were interested also in how closely the perfor-mance of the yoked controls would resemble that producedby C training in Experiment 2.
Method
Subjects. The subjects were 16 10-cm goldfish, all experimen-tally naive. They were drawn from the same pool of subjects andmaintained in the same way as in the previous experiments.
Apparatus. The apparatus was the same as that used in Ex-periments 1 and 2.
Procedure. Two groups of 8 animals each were trained in sub-sets of 4 animals. Each subset comprised two pairs of animals, with1 member of each pair the master (C/E) animal and the other itsyoked (C/Y) control. There were three daily 40-min periods ofadjustment to the apparatus, followed by 20 daily training sessionsin each of which were 20 trials in quasi-random sequence with amean intertrial interval of 90 s. Of the 20 trials, 10 were widelyseparated blank trials (each providing a 10-s sample of activity) and10 were CS-trials. The CS was general illumination provided by redlamps turned on simultaneously at both ends of the shuttlebox, andit was followed after 10 s, independently of response, by the US.As in Experiment 1, the US was a train of .25-s pulses of shock (.33V/cm) delivered at intervals of 2.5 s, and the CS remained on for
both members of each yoked pair until it was terminated along withthe train of shock pulses by the first response of the master animalfollowing the CS-US interval, or after 30 s. The mean interval be-tween CS-presentations was 180 s, with a range of 90 to 290 s.
Results
The performance of the two groups is plotted in Figure 4in terms of the probability of response to the CS in eachsession. Both groups conditioned rapidly, with the escapegroup (whose prompt response to shock restricted the meannumber of pulses to 1.5 per trial for both groups) reachinga higher level of response to the CS than the yoked group.An analysis of variance yielded a significant groups effect,F(l, 14) = 5.94, and a significant sessions effect, F(19,266) = 11.92, with a negligible Groups X Sessions inter-action (F < 1). These results clearly show enhancement ofclassical performance by the escape contingency, althoughreservations about the yoked-control procedure (Church,1964) may suggest that the possibility of a purely Pavlovianexplanation has not been ruled out. The fact should beconsidered in this connection that (as will appear later) allthe results of these experiments can be simulated nicely withtwo processes but can not be simulated when the instrumen-tal component of the theory is eliminated. With respect tothe question of comparability across experiments, it shouldbe noted that acquisition in the master group was very muchlike that in the classical-escape group of Experiment 1 (Fig-ure 2), but the asymptotic performance of the control groupwas somewhat lower than in the classical group of Experi-ment 2 (Figure 3), although the frequency of shock was a bithigher in this experiment.
Parameter Estimation
The simulation method was a computational one (Cou-villon & Bitterman, 1989), which is superior to the familiar
I .OnC/E
0 10 20
SESSIONS
Figure 4. The session-by-session performance of the two groupsin Experiment 3. C/E = classical training with an escape contin-gency; C/Y = yoked controls.

QUANTITATIVE ANALYSIS OF AVOIDANCE CONDITIONING 37
Monte Carlo method both because it is much faster and be-cause it is free of the random error of simulation inherent inthe Monte Carlo method. The Monte Carlo method is to dealwith each of a set of stat-animals in turn, using a probability-generator set at R to determine whether or not the animalresponds on a given trial. If, in avoidance training, the animaldoes not respond on a given trial, c is incremented. If theanimal does respond c is decremented, and i either is incre-mented when i < c (that is, i < A,) or decremented when i> c, as might be the case, for example, where c has beenseparately extinguished following avoidance training (e.g.,Wilson, 1973). Then R is calculated for the next trial, theprobability-generator is consulted again, and so forth. Thesimulated performance is the average performance of a largenumber of stat-animals; the larger the number, the smaller therandom error of simulation. Our own method is to calculatemean associative strengths on the assumption that the pro-portion of animals responding on each trial is R. The changein c on an avoidance trial is the sum of a decrement equal toR X Dfa. X (0 - c) and an increment equal to (1 - R) X UfaX (Ac - c). When i < c, there is an increment in i equal toR X Ufa X (A, - i); when i > c, there is decrement equal toR X Dfa X (A, - 0- It should be obvious that in the classicalcase, where the CS and US are paired (or not) independentlyof response, the Monte Carlo procedure would be pointless.
Efforts were made to simulate the results of ExperimentsI and 2 together. (Experiment 3 was redundant for thepurpose, because the training of the master group was likethe initial training of the classical group in Experiment 1 andthe training of the yoked group was much like the initialtraining of the classical group in Experiment 2.) In the firstsimulations, the six parameters of the theory (the four learn-ing rates, the effective intensity of the shock, and the gen-eralization constant) were varied factorially over a widerange. The variation in parameters was coarse at first andthen was narrowed as the computer homed in on the bestfits, defined in terms of minimal root-mean-square (RMS)deviation of the simulated session-by-session performancefrom the obtained performance of the two groups in each ofthe two experiments.
Shown in Table 1 are the parameter values that yielded thesmall cluster of best fits, 26 of which (.04% of 57,875 simu-lations) have RMS-deviations < .087. Ufa, which provednot to be very closely determined by the data, is much largerthan the other learning rates. The small values of g suggestthat feedback from the shuttling response was itself highlydiscriminable and that CS-termination contributed little tothe growth of i, although there was not a great deal in thetraining procedure to constrain g. The values of s suggest that
Table 1Estimates of Parameter Values
Parameters
i//3c Z?pc t//3, Z>/3, g s
.2-.6 .02-.03 .02-.04 .01 O-.l .50-.55Note. I//3C = incremental 0 for c; D$c — decremental 0 for c;t/j3, = incremental j3 for i; D/3, = decremental /3 for i; g =generalization constant; 5 = effective intensity of shock.
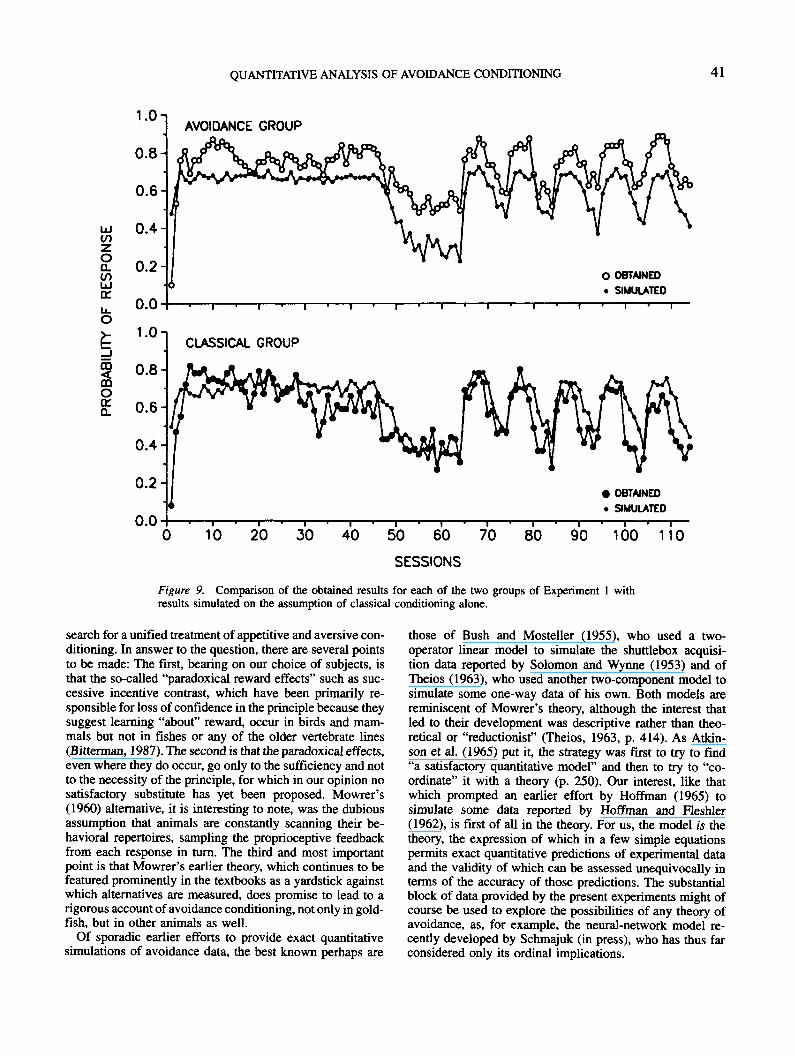
the shock-intensity chosen supported the development onlyof intermediate levels of c. The small variations in parametervalues shown in Table 1 actually made little difference. Theset of values chosen for the purpose of illustration (Ufa =.6, Dfa = .02, Ufa = .04, DP, = .01, g = 0, and s = .5, withRMS = .086) is the one that yielded the marginally very bestfit to the avoidance data of Experiment 1, which proved tobe the most constraining. The fits with these values to theclassical and avoidance data of both experiments (involving3,160 training trials) are shown in Figures 5 (Experiment 1)and 6 (Experiment 2).
It is evident that the main features of this large set of data,not only ordinal, but exact, are captured fairly well in thesimulations, with only three discrepancies worth noting. Oneis that the performance at the very outset of training is con-sistently overestimated; the second is that the performance ofthe avoidance group of Experiment 1 is underestimated atseveral points, and the third is that the asymptotic perfor-mance of the classical group of Experiment 2 is underesti-mated. Such discrepancies may be due to either error in thetheory, error in the data (a certain amount of which is, ofcourse, to be expected in work with small groups), or both.The first discrepancy points to theoretical insufficiency,seeming to reflect a general suppression of activity concomi-tant with the introduction of shock on which the theory issilent; the suppression was studied in Experiment 4 (de-scribed next). The interpretation of the second discrepancyis in doubt; we are inclined now to attribute it to random errorin the data, although it may be resolved with changes in thetheory required by the results of new experiments designedto introduce further constraints. That the third discrepancymay be due to error in the data is suggested by the lower levelof performance in the comparably trained control (CAT) ani-mals of Experiment 3.
A simulation of the results of Experiment 3 with thesame parameters is plotted in Figure 7. The satisfactory fit(RMS = . 106) might be taken as a successful quantitativeprediction in the sense that the results of Experiment 3were not used in the choice of parameters, although theachievement is perhaps not very dramatic because thetraining procedures of Experiment 3 essentially replicatedsome of those in the previous experiments. An interestingoutcome is that the fit to the asymptotic performance ofthe control (C/Y) group of Experiment 3 is better than tothat of the purely classical group of Experiment 2, becausethe level of performance was lower in the third experimentthan in the second. If the data of Experiment 3 are taken asa better estimate of classical performance under these con-ditions than the data of Experiment 2, the third of the threediscrepancies in the fit to the data of Experiments 1 and 2disappears.
The way in which the model works is illustrated by Fig-ure 8, which shows the hypothetical changes in c, i, and Rduring the initial extinction of the avoidance group overSessions 47-64 of Experiment 1. (The plotted values of Uare based, of course, on observed intertrial responding.)There is rapid decline in c, but R declines less rapidly thanit does in the classical group (see Figure 2) because of the

38 A. ZHUIKOV, P. COUVILLON, AND M. BITTERMAN
0.8-
0.6-
LJ 0.4 Hto
1 0.2 HinLUIf 0.0
AVOIDANCE GROUP
O OBTAINED
• SIMULATED
m
oa:a.
1.0-
0.8-
0.6-
0.4-
0.2-
0.0
CLASSICAL GROUP
• OBTAINED
• SIMULATED
0 10 20 30 40 50 60 70 80 90 100 110
SESSIONS
Figure 5. Comparison of the simulated and obtained results for each of the two groups ofExperiment 1.
(autonomous) contribution of i. It is evident here that, de-spite the relatively small values of £7/3, and D/3, required tofit the data of Experiments 1 and 2, the instrumental com-ponent of the theory cannot be dispensed with entirely.Simulations of the data of the two experiments with onlythe classical process yield a minimal RMS of .135 forUfic = .5, I>/3C = .02, and s = .6. The obtained and simu-lated results of Experiment 1 are compared in Figure 9,which shows, as might be expected, large and consistentunderestimation of the performance of the avoidance groupwith the instrumental process excluded.
Experiment 4
This experiment was prompted by the principal discrep-ancy between the simulated and obtained results of thethree experiments, which was the overprediction of re-sponse to the CS at the outset of conditioning. The dis-crepancy seemed to reflect a general suppression of activ-ity produced by shock, and we thought further study of thecircumstances under which suppression occurs was war-ranted. Our plan was to compare intertrial activity in four
groups of animals, a group that was classically conditionedwith a brief shock, a CS-only group, a group for which theCS and US were explicitly unpaired, and an unstimulatedcontrol group.
Method
Subjects. The subjects were 48 10-cm goldfish, all experimen-tally naive. They were drawn from the same pool of subjects andmaintained in the same way as those of the previous experiments.
Apparatus. The apparatus was the same as that used in theprevious experiments.
Procedure. The design called for four groups of 12 animalseach to be trained in subsets of 4 animals, but 1 animal in each groupexcept the CS-only group was eliminated because of bacterial in-fection developed in the course of the training. There were 40 dailytraining sessions of about 40 min in length, during each of whichactivity was sampled in 10 widely separated 10-s intervals (blanktrials). In the first 8 sessions, all animals were unstimulated. In eachof the next 17 sessions, the paired group had, in addition to the blanktrials, 10 classical training trials as in Experiment 2 (10 trials onwhich a 10-s illumination of the red lamps was followed by a singleunavoidable .25-s pulse of .33-V/cm shock). The CS-only grouphad, in addition to the blank trials, 10 unreinforced exposures of the

QUANTITATIVE ANALYSIS OF AVOIDANCE CONDITIONING 39
1.0-
0.8-
0.6-
uj 04-to
1 0.2 HCOLJ
If 0.0
AVOIDANCE GROUP
O OBTAINED
. SIMULATED
1.0-
0.8-
0.6-
0.4-
0.2-
0.0
CLASSICAL GROUP
• OBTAINED
• SIMULATED
10 20 30
SESSIONS
40
Figure 6. Comparison of the simulated and obtained results foreach of the two groups of Experiment 2.
CS alone, the unpaired group had 10 exposures to the CS and 10brief shocks in quasi-random sequence, and the activity controlgroup (with 20 blank trials) was unstimulated as before. In the final15 sessions, the training of all groups was classical. The meaninterval between CS-presentations was 180 s, with a range of90 to 240 s.
Results
In Figure 10, the intertrial activity of the four groups isplotted in terms of the probability of response on the 10 blanktrials programmed in each of the 40 sessions. The probabilityof response both in the activity group and in the CS-onlygroup increased at first and then declined gradually oversessions to a fairly stable level. With the introduction ofshock in Session 26, the intertrial responding in both groupsfell off sharply and then recovered in the next few sessions.An analysis of variance showed a significant sessions effect,F(39, 819) = 5.12, but did not show either a significantgroups effect (F < 1) or a significant Groups X Sessionsinteraction (F < 1). The probability of response both in thepaired group and in the unpaired group also fell sharply withthe introduction of shock in the 9th session and then recov-
ered in the next few sessions, soon reaching the level of theunshocked groups. Here again, an analysis of varianceshowed a significant sessions effect, F(39, 780) = 4.02, butnot a significant groups effect (F < 1) or a significant GroupsX Sessions interaction, F(39, 780) = 2.97.
Performance in the classical training, which closely re-sembled the classical performance shown in Figure 3, wasmuch the same in the four groups; there was no evidenceeither of latent inhibition (CS-only vs. activity, F < 1) or ofretarded acquisition produced by the explicitly unpairedtraining (unpaired vs. activity, F < 1). These results give noindication of contextual conditioning, perhaps because thebrief shocks were relatively weak and the shock-free periodsrelatively long. The observed suppression seems then to bea nonassociative effect of shock that soon habituates andwhose only significance for the theory is that it leads to over-prediction of response to the CS for a few sessions. Beforeformalizing that assumption, however, we think it might bewise to study the effect under a wider range of conditions.
General Discussion
Apart from the usefulness of the results of these experi-ments in the development of a quantitative theory, they addsubstantially to the store of qualitative information on theshuttlebox performance of goldfish.
1. It is now much clearer than before (cf. Scobie & Fallen,1974; Woodard & Bitterman, 1973) that the rates of acqui-sition in classical and avoidance training may be under cer-tain circumstances very much the same, and the stage is setfor an inquiry into the conditions under which the rates maybe different.
2. Performance in classical conditioning is impaired bypartial reinforcement even with frequencies of reinforcementno less than those experienced in avoidance conditioning,and resistance to extinction is reduced as well.
C/E GROUP C/Y GROUP
O OBTAINED
• SIMULATED
20 0 10 20
SESSIONS
Figure 7. Comparison of the simulated and obtained results foreach of the two groups of Experiment 3. C/E = classical trainingwith escape; C/Y = yoked controls.

40 A. ZHUIKOV, P. COUVILLON, AND M. BITTERMAN
~'j 0.8-ui ̂to _z jE
t/> 2 0-6"
u_ w
O UJs. ^ 0.4-
ii8Q.
0.2-
0.0 J
1 3 5 7 9 11 13 15 17
EXTINCTION SESSIONS
Figure 8. Theoretical changes in c (the classical component), i(the instrumental component), and R (response to the CS), andobserved changes in U (intertrial responding) during the initialextinction of the avoidance group in Experiment 1.
3. When blocks of conditioning and extinction sessions arealternated, performance both in classical and avoidancetraining changes appropriately, and the overall level of re-sponding is higher in avoidance training even with consistentreinforcement in the classical training.
4. Shuttlebox performance is facilitated by an escape con-tingency; an indication of such an effect in avoidance trainingappeared some years ago (Behrend & Bitterman, 1964), andhere it appears also in classical training.
5. The suppression of activity with the onset of condi-tioning, which itself has been observed before (Woodard &Bitterman, 1973), appears now to be a nonassociative effectof shock.
6. Classical conditioning is not retarded by frequent priorexposure to the CS alone, to the US alone, or to the CS andUS explicitly unpaired, with no indication of the unusualsuppressing effect of CS-preexposure on asymptotic perfor-mance found by Shishimi (1985); null results also have beenreported for avoidance training (Scobie & Fallen, 1974), andboth sets of results are intriguing in the light of the markedimpairments commonly found in work with other vertebrates(Mackintosh, 1983).
There has been little other study of avoidance under cir-cumstances in which the avoidance behavior tends to be pro-duced also by CS-US contiguity alone, that is, under cir-cumstances in which classical and instrumental influencesare indexed by the same response. General activity in gold-fish was conditioned by Greenway and Bitterman (1972),who found better acquisition in avoidance groups than inyoked controls and some indication of better performance
with a CS-termination contingency. Leg-flexion in dogs wasconditioned by Wahlsten and Cole (1965), who found betterperformance in avoidance training than in classical trainingbut no reliable effect of a CS-termination contingency. Ex-periments on human eyelid conditioning (Gormezano, 1965;Logan, 1951) have shown poorer acquisition but greater re-sistance to extinction in avoidance as compared with clas-sical training, and experiments both on human eyelid con-ditioning and on nictitating-membrane conditioning inrabbits (Gormezano, 1965) have shown better performancein avoidance animals than in yoked controls. In recent workon cheliped-extension in crayfish, Hoshino (1988) found nodifferences in the performance of classical and avoidancegroups trained with or without a CS-termination contin-gency; classical acquisition was as good after CS-only ex-perience, US-only experience, and explicitly unpaired train-ing as after mere exposure to the experimental situation. Ingeneral, these results seem to present no serious challenge totwo-process theory as formulated here in the sense that mostof them can readily be simulated with parameter values tai-lored to the different species and training conditions used.
Nor is it difficult to imagine that it might be possible with-out any fundamental change in the theory to deal with resultsobtained under circumstances in which the response pro-duced by CS-US pairing alone is different from the avoid-ance response, or even to some extent incompatible with it.Although our shuttlebox results for goldfish can be simulatedfairly accurately on the assumption of a substantial contri-bution of c to R, that may not be true of other animals or othersituations. Consider, for example, the fact that the perfor-mance of rats in a conventional shuttlebox as distinct froma "one-way" apparatus is poorer with a higher intensity ofshock than with a lower intensity (Anisman & Waller, 1972;Moyer & Korn, 1964; Theios, Lynch, & Lowe, 1966) or thefact that the two-way shuttlebox performance of dogs bothin avoidance training and in extinction is nonmonotonicallyrelated to shock intensity (Brush, 1957): c may increase asshock intensity increases, and with it the aversiveness of theCS and the reinforcing effect of its termination (A.,), but at thesame time the contribution to R may decline, or even becomenegative, if the overt response classically conditioned to theCS at the higher intensity is less compatible with the avoid-ance response. Fukui and Shishimi (1984) reported a mono-tonic relation between Pavlovian two-way shuttlebox per-formance and shock intensity in goldfish that probably wasnot independent of the structure of their apparatus. Largerend-compartments in relation to the size of the subjectswould be expected to provide other behavioral outlets for cand thus to reduce its contribution to R, perhaps even to apoint at which C training would produce no better shuttleboxperformance in goldfish than in rats (Bolles, Stokes, &Younger, 1966; Kamin, 1956). Such an experiment might bewell worth doing.
With confidence in the S-R reinforcement principle prob-ably lower now than ever before, there may be reason to askwhy serious consideration should be given to a theory inwhich the principle plays a central role. Mowrer (1956,1960)himself soon was tempted to abandon the principle in his
QUANTITATIVE ANALYSIS OF AVOIDANCE CONDITIONING 41
10 30 40 50 60 70
SESSIONS
80 90 100 110
Figure 9. Comparison of the obtained results for each of the two groups of Experiment 1 withresults simulated on the assumption of classical conditioning alone.
search for a unified treatment of appetitive and aversive con-ditioning. In answer to the question, there are several pointsto be made: The first, bearing on our choice of subjects, isthat the so-called "paradoxical reward effects" such as suc-cessive incentive contrast, which have been primarily re-sponsible for loss of confidence in the principle because theysuggest learning "about" reward, occur in birds and mam-mals but not in fishes or any of the older vertebrate lines(Bitterman, 1987). The second is that the paradoxical effects,even where they do occur, go only to the sufficiency and notto the necessity of the principle, for which in our opinion nosatisfactory substitute has yet been proposed. Mowrer's(1960) alternative, it is interesting to note, was the dubiousassumption that animals are constantly scanning their be-havioral repertoires, sampling the proprioceptive feedbackfrom each response in turn. The third and most importantpoint is that Mowrer's earlier theory, which continues to befeatured prominently in the textbooks as a yardstick againstwhich alternatives are measured, does promise to lead to arigorous account of avoidance conditioning, not only in gold-fish, but in other animals as well.
Of sporadic earlier efforts to provide exact quantitativesimulations of avoidance data, the best known perhaps are
those of Bush and Mosteller (1955), who used a two-operator linear model to simulate the shuttlebox acquisi-tion data reported by Solomon and Wynne (1953) and ofTheios (1963), who used another two-component model tosimulate some one-way data of his own. Both models arereminiscent of Mowrer's theory, although the interest thatled to thek development was descriptive rather than theo-retical or "reductionist" (Theios, 1963, p. 414). As Atkin-son et al. (1965) put it, the strategy was first to try to find"a satisfactory quantitative model" and then to try to "co-ordinate" it with a theory (p. 250). Our interest, like thatwhich prompted an earlier effort by Hoffman (1965) tosimulate some data reported by Hoffman and Fleshier(1962), is first of all in the theory. For us, the model is thetheory, the expression of which in a few simple equationspermits exact quantitative predictions of experimental dataand the validity of which can be assessed unequivocally interms of the accuracy of those predictions. The substantialblock of data provided by the present experiments might ofcourse be used to explore the possibilities of any theory ofavoidance, as, for example, the neural-network model re-cently developed by Schmajuk (in press), who has thus farconsidered only its ordinal implications.

42 A. ZHUIKOV, P. COUVILLON, AND M. BITTERMAN
PAIREDO-O UNPAIRED
* 0.61
0.4-
0.2-
0.00 10 20 f30 40
SESSIONS
Figure 10. Intertrial levels of responding in the four groups ofExperiment 4. The arrows show the point at which shock wasintroduced for the paired and unpaired groups (above) and for theCS-only and activity groups (below).
Shown to be tenable by reasonably good quantitative fitsto our data, the model described here seems worth tryingto develop in new experiments designed to shape it further,to refine the estimates of its parameter values, and to testits limits. A good place to begin might be with independentmanipulations of c and i, as in experiments in which shut-tlebox performance has already been studied as a functionof "off-baseline" classical training (Anisman & Waller,1972; Behrend & Bitterman, 1964; Gallon, 1972; Over-mier & Starkman, 1974). Another good place to begin,which would make contact with the substantial rat litera-ture, might be with a structural modification of the appara-tus intended to reduce the probability of shuttling in purelyclassical training. Experiments of both kinds would be ex-pected to bear on the validity of the performance rule,which the results of shuttlebox experiments withtelencephalon-ablated goldfish (Overmier & Hollis, 1990)may seem already to challenge. The determinants of U,which may be in part associative, also should be studiedsoon, perhaps in work on shock intensity and the distribu-tion of training trials (Brush, 1962). Whatever the outcome
of further experiments, the continuing strategy will be toask whether the model predicts each new set of data and, ifnot, how the model can be changed to fit the new data to-gether with all the data already at hand (Couvillon & Bit-terman, 1991).
References
Anisman, H., & Waller, T. G. (1972). Facilitative and disruptiveeffects of prior exposure to shock on subsequent avoidance per-formance. Journal of Comparative and Physiological Psychol-ogy, 78, 113-122.
Atkinson, R. C., Bower, G. H., & Crothers, E. J. (1965). An in-troduction to mathematical learning theory. New York: Wiley.
Behrend, E. R., & Bitterman, M. E. (1964). Avoidance-conditioningin the fish: Further studies of the CS-US interval. American Jour-nal of Psychology, 77, 15-28.
Bitterman, M. E. (1987). Evidence of divergence in vertebratelearning. Behavioral and Brain Sciences, 10, 659-660.
Bolles, R. C., Stokes, L. W, & Younger, M. S. (1966). Does CStermination reinforce avoidance behavior? Journal of Compara-tive and Physiological Psychology, 62, 201-207.
Bower, G., Starr, R., & Lazarovitz, L. (1965). Amount of response-produced change in the CS and avoidance learning. Journal ofComparative and Physiological Psychology, 59, 13-17.
Brogden, W. J., Lipman, E. A., & Culler, E. (1938). The role ofincentive in conditioning and extinction. American Journal ofPsychology, 51, 109-117.
Brush, F. R. (1957). The effects of shock intensity on the acquisitionand extinction of an avoidance response in dogs. Journal of Com-parative and Physiological Psychology, 50, 547-552.
Brush, F. R. (1962). The effects of intertrial interval on avoidancelearning in the rat. Journal of Comparative and PhysiologicalPsychology, 55, 888-892.
Bush, R. R., & Mosteller, F. (1955). Stochastic models of learning.New York: Wiley.
Church, R. M. (1964). Systematic effect of random error in theyoked control design. Psychological Bulletin, 62, 122-131.
Couvillon, P. A., & Bitterman, M. E. (1989). Reciprocal overshad-owing in the discrimination of color-odor compounds by hon-eybees: Further tests of a continuity model. Animal Learning &Behavior, 17, 213-222.
Couvillon, P. A., & Bitterman, M. E. (1991). How honeybees makechoices. In J. L. Goodman & R. C. Fischer (Eds.), The behaviourandphysiology of bees (pp. 116-130). Wallingford, United King-dom: CAB International.
Feller, W. (1950). An introduction to probability theory and its ap-plications. New York: Wiley.
Fukui, H., & Shishimi, A. (1984). Effects of US and CS intensitieson classical conditioning in goldfish (Carassius auratus). Annualof Animal Psychology, 34, 75-86. (In Japanese)
Gallon, R. L. (1972). Effects of pretraining with fear and escapeconditioning on shuttlebox avoidance acquisition by goldfish.Psychological Reports, 31, 919-924.
Gormezano, I. (1965). Yoked comparisons of classical and instru-mental conditioning of the eyelid response; and an addendum on"voluntary responders." In W. Prokasy (Ed.), Classical condi-tioning: A symposium (pp. 48-70). New York: Appleton-Century-Crofts.
Greenway, P., & Bitterman, M. E. (1972). Avoidance conditioningof general activity in the goldfish. Behavior Research Methodsand Instrumentation, 4, 201-202.
Hoffman, H. S. (1965). Theory construction through computersimulation. In W. Prokasy (Ed.), Classical conditioning: A sym-

QUANTITATIVE ANALYSIS OF AVOIDANCE CONDITIONING 43
posium (pp. 107-117). New York: Appleton-Century-Crofts.Hoffman, H. S., & Fleshier, M. (1962). The course of emotionality
in the development of avoidance. Journal of Experimental Psy-chology, 64, 288-294.
Horner, J. L., Longo, N., & Bitterman, M. E. (1961). A shuttleboxfor the fish and a control circuit of general applicability. AmericanJournal of Psychology, 74, 114-120.
Hoshino, Y. (1988). Reflexivity of avoidance behavior in the cray-fish. Annual of Animal Psychology, 38, 107-120. (In Japanese)
Hunter, W. S. (1935). Conditioning and extinction in the rat. BritishJournal of Psychology, 26, 135-148.
Karnin, L. J. (1956). The effects of termination of the CS and avoid-ance of the US on avoidance learning. Journal of Comparativeand Physiological Psychology, 49, 420-424.
Klinman, C. S., & Bitterman, M. E. (1963). Classical conditioningin the fish: The CS-US interval. Journal of Comparative andPhysiological Psychology, 56, 578-583.
Leschova, T. S., & Zhuikov, A. Y. (1989). Fish learning: Ecologicaland applied aspects. Moscow: Nauka. (In Russian)
Levis, D. J. (1989). The case for a return to a two-factor theory ofavoidance: The failure of non-fear interpretations. In S. B. Klein& R. B. Mowrer (Eds.), Contemporary theories of learning: Pav-lovian conditioning and the status of traditional teaming theory(pp. 227-275). Hillsdale, NJ: Erlbaum.
Logan, F. A. (1951). A comparison of avoidance and nonavoidanceeyelid conditioning. Journal of Experimental Psychology, 42,390-393.
Mackintosh, N. J. (1983). Conditioning and associative learning.New York: Oxford University Press.
Mowrer, O. H. (1947). On the dual nature of learning—a reinter-pretation of "conditioning" and "problem-solving." HarvardEducational Review, 17, 102-148.
Mowrer, O. H. (1956). Two-factor learning theory reconsidered,with special reference to secondary reinforcement and the con-cept of habit. Psychological Review, 63, 114—128.
Mowrer, O. H. (1960). Learning theory and behavior. New York:Wiley.
Moyer, K. E., & Korn, J. H. (1964). Effect of UCS intensity on theacquisition and extinction of an avoidance response. Journal ofExperimental Psychology, 67, 352-359.
Overmier, J. B., & Hollis, K. L. (1990). Fish in the think tank:Learning, memory, and integrated behavior. In R. P. Kesner & D.S. Olton (Eds.), Neurobiology of cognition (pp. 205-236).Hillsdale, NJ: Erlbaum.
Overmier, J. B., & Starkman, N. (1974). Transfer of control ofavoidance behavior in normal and telencephalon ablated goldfish(Carassius auratus). Physiology and Behavior, 12, 605-608.
Schmajuk, N. A. (in press). The frightening complexity of avoid-ance: An adaptive network approach. In C. D. L. Wynne & J. E.R. Staddon (Eds.), Models of action. Hillsdale, NJ: Erlbaum.
Scobie, S. R., & Fallen, D. (1974). Operant and Pavlovian controlof a defensive shuttle response in goldfish (Carassius auratus).Journal of Comparative and Physiological Psychology, 86, 858-866.
Shishimi, A. (1985). Latent inhibition experiments with goldfish(Carassius auratus). Journal of Comparative Psychology, 99,316-327.
Solomon, R. L., & Wynne, L. C. (1953). Traumatic avoidance learn-ing: Acquisition in normal dogs. Psychological Monographs, 67,No. 4.
Theios, J. (1963). Simple conditioning as two-stage all-or-nonelearning. Psychological Review, 70, 403—417.
Theios, J., Lynch, A. D., & Lowe, W. F., Jr. (1966). Differentialeffects of shock intensity on one-way and shuttle avoidance con-ditioning. Journal of Experimental Psychology, 72, 294-299.
Wahlsten, D. L., & Cole, M. (1965). Classical and avoidance train-ing of leg flexion in the dog. In W. Prokasy (Ed.), Classicalconditioning: A symposium (pp. 379-408). New York: Appleton-Century-Crofts.
Warner, L. H. (1932). The association span of the white rat. Journalof Genetic Psychology, 41, 57-90.
Wilson, G. T. (1973). Counterconditioning versus forced exposurein extinction of avoidance responding in rats. Journal of Com-parative and Physiological Psychology, 82, 105-114.
Wodinsky, J., Behrend, E. R., & Bitterman, M. E. (1962). Avoidanceconditioning in two species of fish. Animal Behaviour, 10, 76-78.
Woodard, W. T, & Bitterman, M. E. (1971). Classical conditioningof goldfish in the shuttlebox. Behavior Research Methods andInstrumentation, 3, 193-194.
Woodard, W. T, & Bitterman, M. E. (1973). Pavlovian analysis ofavoidance conditioning in the goldfish (Carassius auratus). Jour-nal of Comparative and Physiological Psychology, 82, 123-129.
Received January 1, 1993Revision received July 15, 1993
Accepted July 15, 1993 •
Correction to Author Index for Volume 19
The article "Temporal Encoding as a Determinant of Blocking," by Robert C. Barnet,Nicholas J. Grahame, and Ralph R. Miller (Journal of Experimental Psychology: AnimalBehavior Processes, 1993, Vol. 19, No. 4, pp. 327-341) was inadvertently omitted from theauthor index to the volume.




















