
Regulation of endothelial permeability by β-adrenoceptor agonists: Contribution of β1- and...
13
286 Biochimica et Biophysica Acta, 1178 (1993) 286-298 © 1993 Elsevier Science Publishers B.V. All rights reserved 0167-4889/93/$06.00 BBAMCR 13433 Regulation of endothelial permeability by/3-adrenoceptor agonists: contribution of ill- and fl2-adrenoceptors Sigrid Zink a, Peter R6sen a, Beate Sackmann b and Horst Lemoine u a Diabetes Research Institute, Diisseldorf (Germany) and t, Molecular Drug Research Group, Biomedical Research Center, Heinrich-Heine- Universitiit, Diisseldorf (Germany) (Received 23 February 1993) Key words: Endothelial cell; Permeability regulation;/3-Adrenoceptor subtype; Catecholamine; Sympathomimetics The barrier function of cultured, macrovascular cndothelial cells derived from bovine aorta was analyzed using confluent monolayers of cells and measuring the exchange of fluorescein dextrans of different molecular masses. The effects of fl-adrenoceptor agonists with different selectivity for ill- and /32-adrenoceptors (AR)were investigated. Formoterol, a novel high-affinity agonist for fl2AR recently introduced in the treatment of bronchial asthma, showed a significant reduction of cell permeability with subnanomolar concentrations, whereas the catecholamines (-)-isoproterenol and (-)-norepinephrine only showed significant effects with micromolar concentrations. In order to elucidate if this difference in potential to regulate cell permeability is related to appropriate changes in the selectivity and affinity of the agonists for fl2AR, we investigated thc /3AR-coupled adenylate cyclase (AC) in membranes from endothelial cells and compared AC stimulation with the binding of agonists to the receptors using [1251](-)-iodopindolol as radioligand, fl-Adrenoceptors revealed to be closely coupled to AC as assessed by a similar magnitude of effects by receptor agonists in comparison to GTP analogues and direct stimulants of AC activity. AC activity was increased by formoterol in parallel to its receptor occupancy of fl2AR with nanomolar concentrations which were 50-fold higher than those used for the regulation of cell permeability indicating the existence of spare receptors. In contrast to formoterol, the catecholamines (-)-isoproterenol and (-)-norepinephrine stimulated AC activity through both [31AR and fl2AR. From the overproportionai high contribution of fl1AR to AC stimulation (42%) in comparison to its low fraction (13%) in receptor binding we calculated that fllAR is 3-4-fold more effectively coupled to AC than fl2AR. Introduction The vascular endothelium builds up a selective bar- rier between the vascular lumen and the extravascular space and plays a central role in regulating the ex- change of fluid, solutes and cells. In the last decade, a variety of hormones, neurotransmitters and regulators were found to be involved in the modulation of en- dothelial permeability. Under physiological conditions, concentration gradients across the endothelial barrier are build-up for nutrients, hormones, proteins and other macromolecules, and blood cells are excluded from the passage across the vessel wall [1-3]. Under Correspondence to: P. R/Ssen, Diabetes Research Institute, Auf'm Hennekamp 65, 4000 Diisseldorf l, Germany. Abbreviations: AC, adenylate cyclase; AR, adrenoceptor; ASD, asymptotic standard deviation; BAEC, bovine aortic endothelial cells; cAMP, cyclic AMP; IBMX, 3-isobutyl-l-methylxanthine; IA, intrinsic activity; ISO, (-)-isoproterenol; [1251]IPIN, [12sI](-)-iodo- pindolol; FOR, (+)-formoterol; E, (-)-epinephrine; NE, (-)-nor- epinephrine; FITC, fluorescein-isothiocyanate. pathophysiological conditions, such as ischemia [4], ar- teriosclerosis [5], bronchial asthma [6] and in tumors [7], the endothelium can loose its barrier function and allow the extravasation of macromolecules. Such a loss of tightness of endothelial cells seems to result specifi- cally from an opening of intracellular clefts and, thus, mostly reflects an increase in the paracellular exchange of molecules [2,3]. A variety of substances like histamine, thrombin and bradykinin [8-10] has been shown to be involved in the increase of endothelial permeability, but only few have been suggested to induce the opposite effect. Among these, flAR agonists belong to the most potent sub- stances to reduce endothelial cell leakage. In animal models, e.g., the canine forelimb [11], hamster cheek- pouch preparation [10] and guinea-pig lung [6], it was found that the efflux of protein induced by histamine and bradykinine could be reduced by non-selective (isoprenaline) and fl2-selective (terbutaline) flAR ago- nists. New developments in cell culture techniques make it possible to study the permeability properties of an endothelial monolayer in vitro [3], thereby avoiding
-
Upload
uni-duesseldorf -
Category
Documents
-
view
1 -
download
0
Transcript of Regulation of endothelial permeability by β-adrenoceptor agonists: Contribution of β1- and...

286 Biochimica et Biophysica Acta, 1178 (1993) 286-298 © 1993 Elsevier Science Publishers B.V. All rights reserved 0167-4889/93/$06.00
BBAMCR 13433
Regulation of endothelial permeability by/3-adrenoceptor agonists: contribution of ill- and fl2-adrenoceptors
Sigrid Zink a, Peter R6sen a, Beate Sackmann b and Horst Lemoine u a Diabetes Research Institute, Diisseldorf (Germany) and t, Molecular Drug Research Group, Biomedical Research Center,
Heinrich-Heine- Universitiit, Diisseldorf (Germany)
(Received 23 February 1993)
Key words: Endothelial cell; Permeability regulation;/3-Adrenoceptor subtype; Catecholamine; Sympathomimetics
The barrier function of cultured, macrovascular cndothelial cells derived from bovine aorta was analyzed using confluent monolayers of cells and measuring the exchange of fluorescein dextrans of different molecular masses. The effects of fl-adrenoceptor agonists with different selectivity for ill- and /32-adrenoceptors (AR)were investigated. Formoterol, a novel high-affinity agonist for fl2AR recently introduced in the treatment of bronchial asthma, showed a significant reduction of cell permeability with subnanomolar concentrations, whereas the catecholamines (-)-isoproterenol and (-)-norepinephrine only showed significant effects with micromolar concentrations. In order to elucidate if this difference in potential to regulate cell permeability is related to appropriate changes in the selectivity and affinity of the agonists for fl2AR, we investigated thc /3AR-coupled adenylate cyclase (AC) in membranes from endothelial cells and compared AC stimulation with the binding of agonists to the receptors using [1251](-)-iodopindolol as radioligand, fl-Adrenoceptors revealed to be closely coupled to AC as assessed by a similar magnitude of effects by receptor agonists in comparison to GTP analogues and direct stimulants of AC activity. AC activity was increased by formoterol in parallel to its receptor occupancy of fl2AR with nanomolar concentrations which were 50-fold higher than those used for the regulation of cell permeability indicating the existence of spare receptors. In contrast to formoterol, the catecholamines (-)-isoproterenol and (-)-norepinephrine stimulated AC activity through both [31AR and fl2AR. From the overproportionai high contribution of fl1AR to AC stimulation (42%) in comparison to its low fraction (13%) in receptor binding we calculated that fllAR is 3-4-fold more effectively coupled to AC than fl2AR.
Introduction
The vascular endothelium builds up a selective bar- rier between the vascular lumen and the extravascular space and plays a central role in regulating the ex- change of fluid, solutes and cells. In the last decade, a variety of hormones, neurotransmitters and regulators were found to be involved in the modulation of en- dothelial permeability. Under physiological conditions, concentration gradients across the endothelial barrier are build-up for nutrients, hormones, proteins and other macromolecules, and blood cells are excluded from the passage across the vessel wall [1-3]. Under
Correspondence to: P. R/Ssen, Diabetes Research Institute, Auf'm Hennekamp 65, 4000 Diisseldorf l, Germany. Abbreviations: AC, adenylate cyclase; AR, adrenoceptor; ASD, asymptotic standard deviation; BAEC, bovine aortic endothelial cells; cAMP, cyclic AMP; IBMX, 3-isobutyl-l-methylxanthine; IA, intrinsic activity; ISO, (-)-isoproterenol; [1251]IPIN, [12sI](-)-iodo- pindolol; FOR, (+)-formoterol; E, (-)-epinephrine; NE, (-)-nor- epinephrine; FITC, fluorescein-isothiocyanate.
pathophysiological conditions, such as ischemia [4], ar- teriosclerosis [5], bronchial asthma [6] and in tumors [7], the endothelium can loose its barrier function and allow the extravasation of macromolecules. Such a loss of tightness of endothelial cells seems to result specifi- cally from an opening of intracellular clefts and, thus, mostly reflects an increase in the paracellular exchange of molecules [2,3].
A variety of substances like histamine, thrombin and bradykinin [8-10] has been shown to be involved in the increase of endothelial permeability, but only few have been suggested to induce the opposite effect. Among these, f lAR agonists belong to the most potent sub- stances to reduce endothelial cell leakage. In animal models, e.g., the canine forelimb [11], hamster cheek- pouch preparat ion [10] and guinea-pig lung [6], it was found that the efflux of protein induced by histamine and bradykinine could be reduced by non-selective (isoprenaline) and fl2-selective (terbutaline) f lAR ago- nists. New developments in cell culture techniques make it possible to study the permeability properties of an endothelial monolayer in vitro [3], thereby avoiding

the complications of changes in hydrostatic pressure produced in whole animal models, and to correlate endothelial cell function with biochemical events, such as receptor binding and receptor coupled AC stimula- tion, occurring in the plasma membrane. If endothelial OAR were of the 132 subtype as supposed by some authors [12-14] quantitative differences in functional responses should be expected corresponding to differ- ent affinities and selectivities of the used/3 agonists for B1AR and O2AR. Thus, non-selective (e.g., (-)-iso- proterenol), O~-selective (e.g., (-)-norepinephrine) or O2-selective OAR-agonists (e.g., formoterol) should clearly differ in the properties to regulate endothelial cell permeability or to stimulate OAR-coupled AC [15-17]. If, however, the OAR-population of endothe- lial cells is heterogeneous [14], as found in various organs of many species, it is of great interest to investi- gate the distribution of OAR, its coupling to AC and its role in regulating endothelial cell permeability. The study of regulation of endothelial permeability and its relation to the activation of AC was enabled by the use of formoterol, a novel highly 02-selective and high-af- finity agonist [18,19] which is currently introduced in the therapy of bronchial asthma [20], and antagonists highly selective for OIAR (CGP 20712A [17,21]) and for 02AR (ICI 118551 [15]).
Materials and Methods
Experiments in endothelial cell monolayers Cell isolation and culture conditions. Bovine aortic
endothelial cells (BAEC) were isolated using the method of Booyse and co-workers [22] with slight modifications. In brief, aortas of freshly slaughtered calves were collected and washed under sterile condi- tions with phosphate-buffered saline (PBS (pH 7.4)) consisting of (raM) 140 NaCl, 4 KCI, 1 Na2HPO 4. 2H20, 1 KH2PO 4 and 12 glucose. The adventitia was removed and the remaining part of media and intima was clamped into an aluminium tub. The endothelial cells built the bottom of the tub, which was filled with 5 ml dispase solution (0.5 mg/ml) and incubated at 37°C. After 15 min the enzyme solution was removed and the cells were collected by washing the intima with DMEM (Dulbecco's Modified Eagle's Medium) con- taining 20% fetal calf serum. The cells were spun down at 100 × g for 10 min. The pellet was disperged and the cells were seeded in DMEM enriched with 10% fetal calf serum on 25-cm 2 dishes coated with 0.2% gelatine for 30 min at room temperature. The plating density was approx. 10 000 cells/cm 2.
BAEC were cultured at 37°C in 5% CO 2 and 95% air. The cells were passaged every four or five days and diluted 1:7, i.e., the cells need three divisions to reach confluency. Maximum lifespan for BAEC is 55 pas-
287
sages. In this study, cells were used up to the 25th passage.
Characterization of endothelial cells. The cells were identified as endothelial cells by their morphology, immunostaining of factor-VIII-related antigen and up- take of acetylated low-density lipoproteins. To study the synthesis of a specific pattern of prostglandins by macrovascular endothelial cells [23], cell monolayers cultivated in 25-cm 2 dishes were stimulated with 10 /zM [14C]arachidonic acid for 15 min. The prosta- glandins were separated by HPLC using a ClS reverse- phase column and an acetonitrile gradient as mobile phase (30%-100% acetonitrile in water with 0.1% acetic acid) [24]. Radioactivity was detected by a con- tinuous flow monitor after addition of liquid scintilla- tion fluid in a ratio of 1 : 3.
Transendothelial exchange. Cells were seeded on polycarbonate filters (0.4-/zm pores, 4.71 cm 2 area). At confluency, filters were set in a frame, which was placed in a 600-ml beaker with 40 ml DMEM (ablumi- nai compartment). The temperature was regulated at 37°C with a waterbath. The abluminal solution was stirred with a magnetic stirrer (Variomag, H + P, Miinchen, Germany). The luminal compartment con- tained 1.5 ml DMEM with several marker substances (tracers) such as [14C]sucrose (1 #M) and fluorescein- isothiocyanate (FITC) labelled dextrans (10 ~M) of different molecular masses as described [25]. Aliquots of 1 ml were taken from the abluminal compartment at 5, 10, 20, 30, 40, 60 and 90 min and were analyzed in a O-scintillation counter or in a fluorimeter (excitation wavelength, 495 nm; emission wavelength, 530 nm). Data were calculated in two different ways: (1) The amount of substance diffused into the abluminal (lower) compartment is calculated as percentage of the total amount in the luminal (upper) chamber at t = 0 min. (2) The permeability (PEc (cm/s)) of the endothelial monolayer was calculated according to Del Vecchio and co-workers [26]. The experimentally-determined permeabili ty (PEc + F) was corrected for the perme- ability of the filters without cells (PF) as
1/PEc = I /PEc+F-- I / P F (1)
where the permeability P is defined as P = dV/d t "A. dV/d t stands for the exchanged volume per time and A stands for the area of exchange (4~71 cm2).
Modulation of transendothelial exchange. Nore- pinephrine (NE) and synthetic OAR-agonists (ISO and FOR) were used in concentrations from 0.01 nM up to 100 /~M and added to the luminal and abluminal compartment at t = 0 min. Aliquots of the abluminal compartment were taken and analyzed as described. To show the reversibility of effects induced by OAR agonists, cells were treated with the OAR-antagonist alprenolol after preincubation with NE for 60 min.

288
Measurement of intracellular cAMP. Cyclic AMP was quantified using a radioimmunoassay kit. Cells were incubated concomittantly with 0.2 mM 3-isobutyl-l- methylxanthine (IBMX) and with different concentra- tions of the used catecholamines for 10 min. There- after, the incubation was stopped by removing the incubation medium and addition of 1 ml ice-cold ethanol (100%) per dish (9.6 cm2), the cells were removed from the dish with a rubber policeman and sonicated three times for 3 s and centrifuged at 1000 x g for 10 min. The ethanolic extract was dried under a stream of N 2 at 60°C and then dissolved in acetate buffer (pH 5.8). Aliquots of this solution were used to quantify the amount of endogenous cAMP in the ra- dioimmunoassay kit.
Experiments in membranes from endothelial cells Preparation of cell membranes. Endothelial cells were
cultured in 58.1-cm 2 cell-culture dishes under the con- ditions detailed above. Cells were harvested at conflu- ency 5 days after the culture was set up. Membranes from cultured cells were prepared after removing the culture medium and rinsing the cell monolayer with serum-free medium. Thereafter, the cells were gently removed from the dish surface with a rubber police- man, transferred on ice and washed by centrifugation at 40000 ×g for 30 min at 4°C. After decanting the supernatant the cell pellet was resuspended in a buffer containing 3 mM EGTA and 5 mM Tris-HCi at pH 7.4 and gently homogenized (10 strokes) in a glass-glass homogenizer. The suspension was centrifuged again at 40000 x g for 30 min at 0°C. The resulting pellet was resuspended in Tris-EGTA buffer at a protein concen- tration of approx. 1 mg/ml and stored at -80°C until use .
Radioligand binding. Experiments were carried out at 37°C in 50 mM Tris-HCl (pH 7.4), 2 mM MgCI, 1 mM EGTA, 0.1 mM ascorbic acid and in the presence of 0.2 mM GTP. In order to estimate the/3AR density and the affinity of/3AR agonists for/31AR and /32AR, /3AR were radiolabelled by [125I]IPIN. The radioligand was incubated for 30 min, a time found sufficient for complete equilibration with the receptors [27]. Binding observed in the presence of 0.2 mM ISO was regarded to be non-specific. Bound radioligand was separated from free radioligand by rapid vacuum filtration through Whatman U F / A glass-fiber filters. The filters were counted in a y-counter with an efficiency of 70%.
Saturation binding experiments were analyzed ac- cording to the equation
Bt([L* ] ) = B s + B m = s lope ' [L* ] + Brn ~ [ L * ] / ( [ L * ] + KL. )
(2)
where Bt, Bns , B s and Bma x represent total, non- specific, specific and maximum specific binding of
[125I]IPIN (L*), respectively, KL. is the equilibrium dissociation constant of L* and slope stands for the steepness of Bns.
Competition binding experiments were performed (i) to estimate the relative proportion of/3AR-subtypes using a /31-selective (CGP 20712 A) and a /3E-selective (ICI 118551)/3AR-antagonist and (ii) to determine the affinity of/3AR-agonists for/31AR and flEAR. Specific binding of [125I]IPIN was measured in the presence of increasing concentrations of unlabelled ligands (L). Competition binding experiments for n unlabelled lig- ands Lj ( j = 1 . . . . . n) were analyzed by a common multiple non-linear regression as reported by Ehle and co-workers [28,29] according to the set of n equations
2 f , ' [ L , ] Bs ( [LJ ] ) = B°j-B°", •=1 [LA+ KLj,'( I + [L* ]/KI* )
(3)
where i = 1, 2 indicate/31AR and fl2AR, respectively, /3o) and Bs([Lj]) represent the specific binding of L* to /3AR in the absence and presence of L j, respectively, KL, represents the dissociation constants of [~zsI]IPIN estimated independently in saturation binding experi- ments, KLj i represents equilibrium dissociation con- stants of Lj for fllAR and /32AR, and fi (fl = 1 - f2) represents fractions of receptor subtypes. The experi- ment depicted in Fig. 4 was analyzed with a set of two equations setting L~ and L 2 as CGP 20712 A and ICI 118551, respectively. Competition binding experiments for each agonist were analyzed with a set of three equations setting L 1 and L 2 as the highly selective antagonists CGP 20712 A and ICI 118551, respectively, and L 3 as the agonist of interest (FOR, ISO or NE). Thus, intercorrelations between the estimates of fl and PKL, and between f : and PKL~ of the agonist could be minimized, resulting in reliable estimates of pgL~ and PKL2. Experimental data were analyzed after transformation of data to obtain homoscedasticity [28], resulting in reliable estimates of parameters and asymptotic standard deviations (ASD).
Adenylate cyclase. Assays were carried out in a final volume of 60 /xl containing (mM): 0.1 [a-32p]ATP (100-140 cpm/pmoi), 100 Tris-HCl (pH 7.4), 2 MgCl 2, 1 EGTA, 0.1 ascorbic acid, 0.01 GTP, 1 [3H]cAMP (5000 cpm per tube), 20 creatine phosphate as sub- strate of the ATP-regenerating enzyme system and the enzymes (IU/ml): 15 creatine phosphokinase, 9.8 myokinase. The reaction was started by the addition of the membrane suspension and continued for 20 min at 37°C. Accumulation of cAMP was linear during this period, both in the absence and in the presence of 0.2 mM ISO. The reaction was terminated by the addition of 100 ~1 of a solution containing 10 mM cAMP, 40 mM ATP and 1% SDS. [32p]cAMP was purified by double chromatography as described [30] and mea- sured by liquid scintillation counting.

Concentration-effect curves for AC stimulation by flAR-agonists were fitted by non-linear regression ac- cording to the following equation [17,31]:
2 /s," [AI s = s ~ + (s~.~ - s ~ ) . ~ ,
[A] + ECs0.,(1 + [B]/K,.,) (4)
where Sba s, Sma x and S represent the basaI, maximum and resultant AC activity, respectively, subindices i represent fllAR (i = 1) and fl2AR (i = 2), fsi is the respective fraction of AC stimulation via /3~AR and fl2AR by an agonist with ECs0,i as half-maximum con- centrations and Ka.i is the respective dissociation con- stants of an antagonist B.
Drugs and materials Adenosine 5'-triphosphate Tris salt (ATP), adeno-
sine 3'-5'-monophosphoric acid (cAMP), guanosine 5'- triphosphate Tris salt (GTP), guanyl-5'-yl imidodiphos- phate sodium salt (GPP-NH-P), creatine phosphate, myokinase, forskolin, trypsin and 3-isobutyl-l-methyl- xanthine (IBMX) were from Sigma (St. Louis, MO, USA). Creatine phosphokinase was from Calbiochem (La Jolla, CA, USA). [a-32p]ATP (22.2 TBq/mmol), 3'-8'-[3H]cAMP (1.33 TBq/mmol), the [tzSI]cAMP as- say system for cAMP and [14C]arachidonic acid were from the Radiochemical Center (Amersham, UK). [t25I](-)-Iodopindolol ([t2sI]IPIN, 81.4 TBq/mmol) was from Du Pont de Nemours (Dreieich, Germany). ICI 118551 (erythro-(+)-l-(7-methylindan-4-yloxy)-3- isopropylamin obutan-2-ol hydrochloride) was from ICI (Macclesfieid, UK). CGP 20712 A (1-[2(3-carbamoyi- 4-hydroxyphenoxy)-et hylamino]-3-[4-( 1 -methyl-4-triflu- oromethyl-2-imidazolyl) phenoxy]-2-propanolmethane- sulfonate) and (±)-formoterol were from Ciba-Geigy (Basle, Switzerland). (-)-Norepinephrine bitartrate monohydrate and (-)-isoproterenol bitartrate were from Sterling-Winthrop (Rensselaer, NY, USA). Phe- noxybenzamine hydrochloride was from Smith, Kline and French (Philadelphia, PA, USA). Dispase (grade II) and fetal calf serum were from Boehringer-Man- nheim (Mannheim, Germany). Dulbecco's Modified Eagle's Medium (DMEM) was from Flow Laboratories (Meckenheim, Germany). Anti-human-factor VIII was from the Red Cross (Amsterdam, The Netherlands) and FITC-anti-rabbit-IgG was from (Dakopatts, Den- mark). Acetylated low-density lipoproteins (LDL) were from Paesel (Frankfurt am Main, Germany). Other chemicals were obtained from local commercial sources.
C18 reverse-phase columns (LiChrosorb) were from Merck (Darmstadt, Germany). Polycarbonate filters (0.4-/.~m pores; 4.71 cm 2 area) were from Costar Eu- rope (The Netherlands). The fluorimeter 650-40 was from Perkin-Elmer (0berlingen, Germany). The HP-
289
TABLE 1
Transendothelial exchange of various tracer molecules across endothe- lial cell monolayers within 20 rain
The exchange of tracer molecules was calculated as the amount of tracer in the abluminal compartment as percentage of the initial amount of tracer in the luminal compartment set to 100%. Data are mean + S.E. of n observations. The radii of tracer molecules given in nm were calculated using the Stokes-Einstein equation and data from the literature [34,35].
Tracer Radius exchange n (nm) (%)
[3H]NE 0.41 36.9:1:1.5 5 [ 14C]Sucrose 0.52 19.6:1:0.2 31 [ 125 l]lnsulin 1.32 2.2 ± 0.1 3 FITC-dextran (4 kDa) 1.56 1.4 ± 0.4 3 FITC-dextran (40 kDa) 4.30 1.3 ± 0.02 29 FITC-dextran (70 kDa) 5.92 0.4 ± 0.02 10
LC-radioactivity monitor LB 506c was from Berthold (Wildbad, Germany).
Results
Functional experiments Characterisation of macrovascular endothelial cells
(BAEC). The cells grew in a typical cobblestone pat- tern, forming a confluent monolayer. The growth of the endothelial cells was contact inhibited. At the state of confluency, the ceils reached a density of approx. 10 5 cells/cm 2. Transmission electron microscopy (data not shown) showed a regular distribution of junctions between the cells. The cells rapidly took up acetylated low-density lipoproteins [32] and showed a spot-like pattern after immunostaining of factor-VIII-related antigen, indicating the presence of Weibel-Palade bod- ies [33].
Additionally, the synthesis of eicosanoids was stud- ied. It is known that the predominant synthesis of prostacyclin (PGI 2) is a typical property of BAEC [23]. Stimulation of cells with 10 ~M arachidonic acid led to the synthesis of 13.6 + 0.9 pmol PGI2/105 ceils within 15 min. This amount represents 64.5% of the whole production of prostaglandins.
Transendothelial exchange. The permeability of BAEC monolayers depended on the molecular mass, i.e., on the size of molecules being exchanged (Table I); 19.6 + 0.2% and 0.40 + 0.02% of the total amounts of sucrose and of 70 kDa FITC-dextran passed the monolayers in 20 min, respectively.
The relation between radius and permeability of molecules of different molecular mass is in line with the assumption of two classes of intercellular clefts in endothelial cells (Fig. 1). The relation can be approxi- mated by two straight lines with different steepness (Fig. 1, insets). The crossing points of the straight lines

290
1000
800 U3
E
u 600 ~D
40O 8 U . I
o_ 200 .~ .
oo ol o~ o* os 1o
0 @O I I i I I I
0 1 2 3 4 5 6 radius of t racer (nm)
Fig. 1. Relation between the radius of tracer molecules and the permeability of endothelial cell monolayers (BAEC). As tracer molecules [3H}(-) -norepinephr ine , [~4C]sucrose, [125Ilinsulin and FITC-dextrans with a molecular mass of 4, 40 and 70 kDa were used. The permeability PEC was calculated as described in Materials and Methods. The insets represent linearizations of the original data and reveal two components of the relation. The steep line (top inset) and the fiat line (bottom i n , t ) are characterized by abzissa intercepts of 0.54 and 8.77 nm (radii), respectively, hinting at the coexistence of
small and large pores in cultured endothelial cell monolayers.
with the abscissa (that means PEC of this class of pores becomes zero) indicate two limiting pore-sizes, the diameters (2r) of which can be estimated to be 1.1 and 17.5 nm.
The total area of intercellular clefts which con- tribute to the exchange of molecules through endothe- lial cell monolayers (pathlength dx = 2 . 1 0 -4 cm) can be roughly approximated by comparing the permeabil- ity ( P ) of sucrose in water (Pu,o = DH,o/dx = 5.82. 10 -6 /2 • 10 -4 = 2.91 • 10 -2 c m / s at 37°C3 with its per- meability experimentally determined in endothelial cell monolayers ( P E c = 117" 10 -6 cm/s) . From the ratio of PEC and Plt2o (PEc/PHzo =4"10-3) it can be as- sumed that 0.4% of the total area of the monolayers contributes to the exchange of molecules across the endothelial cell monolayers.
The exchange of molecules such as sucrose and FITC-labelled dextrans increased proportionally to the tracer concentration in the luminal compartment (sucrose, 10-300 /~M; 70 kDa FITC-dextran, 1-20 /zM) and exhibited linear kinetics up to 180 min (data not shown). These observations indicate that the ex- change of molecules across endothelial cell monolayers mainly reflects diffusion processes.
Modulation of transendothelial exchange by fl AR ag- onists. The tightness of cell monolayers increased with increasing concentrations of OAR agonists, as shown for the OAR agonists FOR, ISO and NE in Fig. 2. Half-maximal reduction of permeability is already achieved by 0.42 nM FOR, whereas about 10000-fold higher concentrations are necessary to get the same effect with ISO and NE. In contrast to the large
E
o
~q cO
b (D
lO0
-- 80
6O %
40
20
0 - 4 / i i i i i i i i
11 10 9 8 7 6 5 4 -log [agonist] M
Fig. 2. Dose-dependent reduction of permeability of endothelial cell monolayers (BAEC) by the f lAR-agonists norepinephrine (o), iso- proterenol (* ) and formoterol (*). Endothelial cells were incubated for 1 h with 10 p M of 41) kDa FITC-dextran and the indicated concentrations of agonist. The amount of tracer in the abluminal compar tment was determined. The permeability is given as percent- age of basal tracer-exchange set to 100%. Symbols indicate mean + S.E. of 4 -10 observations. A non-linear regression analysis yielded the concentrations of the half-maximal effects (ECs0) of 0.42 + 0.01 nM, 2.49+0.02 /~M and 3.70+0.13 /zM for FOR, ISO and NE,
respectively.
difference in potency between FOR and I S O / N E , the potencies of ISO and NE are only slightly different. The decrease in permeability was more striking for 40 kDa FITC-dextran than for small molecules such as sucrose (Table II).
The decrease of permeability induced by 10/zM NE could be abolished by incubating the cells with the /3AR blocker alprenolol (Fig. 3), but not with the a A R blocker phentolamine, indicating that the changes in permeability observed with NE were mediated by acti- vation of /3AR.
Radioligand binding /3AR from endothelial cells was identified in cell
membranes by radioligand binding using [125I]IPIN as radioligand (Fig. 4). Non-specific binding in the pres- ence of a flAR-saturating concentration of ISO was
TABLE II
Comparison of the effects of ( - )-norepinephrine on the permeability of endothelial cell monolayers for tracer molecules of different sizes
Monolayers of BAEC were incubated with 10 /~M ( - ) -no rep in ep h - rine and tracer. The amount of tracer in the abluminal compar tment was determined at 60 min and calculated as percentage of the exchange of tracer in the absence of ( - ) - no rep ineph r in e set to 100%. Data represent means_+ S.E. of n different cell preparations.
Tracer exchange n (%)
[ 14C]Sucrose 71.7 + 1.2 7 FITC-dextran (20 kDa) 63.6:1:2.3 3 FITC-dextran(40 kDa) 45.1 5- 3.9 3

291
_,~ 8
e o o T o C o 4 / l ,e , •
6 ~ ° . .~/~
2 / ~ 0 ~ "addition of c a / 0 ~ I ~ ' - ' ~ O ~ olprenolol
eJ 0 i i i i i i i i
0 I0 20 30 40 50 60 70 80 90
time (rain)
Fig. 3. Kinetic of the reduction of endothelial cell (BAEC) perme- ability by (-)-norepinephrine. Blockade by a BAR antagonist. Monolayers were incubated with 10/zM 40 kDa FITC-dextran up to 90 min and the exchange of the tracer between the luminal (1.5 ml) and the abluminal (40 ml) compartment was measured. The ex- change was calculated as the amount of tracer in the abluminal compartment at the indicated time points as percentage of the initial (total) amount of tracer in the luminal compartment set to 100%. The curves represent the FD-exchange in the absence (o) and presence (o) of l0 /zM (-)-norepinephrine. At 60 rain, 100 #,M alprenolol was added resulting in an increase of the FD exchange up to control levels. Each point represents the mean of six observa-
tions + S.E.
low and increased linearly with increasing concentra- tions of the radioligand. Specific binding was saturable and characterized by a hyperbolic concentration-de- pendence. The non-linear regression analysis yielded a high affinity of the radioligand (K d = 0.2 nM) and a Bma x value of 48.7 + 1.9 fmol/mg protein. Scatchard transformation of the data (not shown) revealed that []25I]IPIN bound to a homogeneous class of binding sites as inferred from the linearity of the Scatchard plot.
To characterize the subtypes of /3AR on the en- dothelial cell the binding of [t25I]IPIN was dose-depen- dently inhibited by antagonists highly selective for /31AR (CGP 20712 A) or /3zAR (ICI 118551). The experiment is depicted in the bottom part of Fig. 4. Whereas ICI 118551 competed with nanomolar con- centrations for 87% of the receptors, CGP 20712 A only exerted high affinity for 13% of the receptors. By these experiments the predominant receptor subtype was characterized as /32AR. Simultaneous analysis of both binding inhibition curves by non-linear regression according to Eqn. 3 confirmed that ICI 118551 and CGP 20712 A competed with [t25I]IPIN for 87% (i.e., /32AR) of total/3AR with high and low affinity, respec- tively, as expected from their selectivity for flAR sub- types. The remainder part of 13% of total /3AR were characterized as /3tAR. The estimates of parameters are listed in Table III.
The binding of/3AR agonists was investigated in the presence of a saturating concentration of GTP, a con-
60.
B
i 0.0 0.3 0.6
[ 1251-H-iodopindolol] nM 019
B% I c ~ 2 x7 (Sr~ek~'tI~J /
5 0
loo
-log [antagonistl M Fig. 4. Estimation of the BAR density and chararacterization of BAR subtypes of endothelial cells. Upper panel: Saturation binding of [125I]IPIN. Binding of [125I]IPIN was assayed in the absence and presence of 0.2 mM (-)-isoproterenol yielding total (o) and non- specific binding (D). Specific binding (o) is plotted as broken line. Curves were analyzed according to Eqn. 2 by non-linear regression yielding a Bmax of 48.74- 1.9 fmol/mg protein and a dissociation constant ( - l o g (M)) of 9.71:I:0.05 for [125I]IPIN. Lower panel: Competition of subtype-selective antagonists with [~251]IPIN for binding to BAR. Binding of 0.375 nM []2-Sl]IPIN was inhibited with increasing concentrations of a BtAR-selective (CGP 20712 A, El) and a BzAR-selective (ICI 118551, o) antagonist. Both curves were analyzed with a common non-linear regression procedure according to Eqn. 3, assuming that fractions of receptor-subtypes were the same for both antagonists. The analysis revealed that 134-2% of total BAR were of the Bl type. Estimates of dissociation constants for the antagonists are given in Table III. Dotted curves represent computer simulations according to Eqn. 3, assuming that only/32AR contributes to the binding of [125I]IPIN (i.e., f l = 0, ]'2 = 1). Symbols
represent means ± S.E. of four determinations.
TABLE III
Dissociation constants of agonists and antagonists for f l lAR and fl 2 AR estimated from radioh'gand binding
pK a values are dissociation constants ( - log (M)) of antagonists and agonists for BIAR and BzAR. ]'1 values (]'1 + I"2 = 1) are fractions of B1AR. Parameters were estimated by a common non-linear regres- sion according to Eqns. 2 and 3 and assuming a common receptor fraction (.fl) for all curves with selective ligands. Experiments are shown in Figs. 4 and 5.
Ligand Dissociation constant f] 4- ASD (pK a ±ASD)
fllAR B2AR
CGP 20712 A 8.79:t:0.65 5.484-0.04 "~ ICI 118551 6.884-0.28 8.53-1-0.05 ( - )-Norepinephrine 6.54 4- 0.62 4.71:1:0.03 0.13 :t: 0.02 ( 4- )-Formoterol 6.12 4- 0.26 7.89 4- 0.05 ( - )-Isoproterenol 6.97 :t: 0.03 6.97 ± 0.03

292
dition known to induce a complete interconversion of receptors into the low-affinity binding state which en- ables activation of adenylate cyclase. The experiments are demonstrated in Fig. 5. The binding-inhibition curve with ISO which is non-selective for /3~AR and /32AR revealed a normal steepness (Hill coefficient = 1) characteristic for the interaction with a homogeneous class of binding sites. Formoterol and NE exerted shallow binding inhibition curves, indicating that FOR and NE bound to /3~AR and /32AR with different affinities. Binding inhibition curves of FOR and NE were analyzed by non-linear regression simultaneously with the curves of the selective antagonists (Fig. 4), thus constraining the estimation of f~ (fraction of /31AR) to the value ( f l = 0.13) estimated for the selec- tive antagonists alone. The non-linear regression analy- sis based upon the constrained model converged and allowed the estimation of dissociation constants for the agonists listed in Table III. For FOR, 87% and 13% of total /gAR were of high ( K a = 15 nM) and low ( K a =
800 nM) affinity, respectively, characterizing FOR as highly selective for /32AR. These values can be con- trasted to those of NE, an agonist selective for/3~AR, which showed high affinity ( K a = 0.3 /zM) only for 13% of total /gAR (i.e.,/31AR) and low affinity ( K a = 20/zM) for the remaining 87%.
Stimulat ion o f intracellular c A M P To prove whether the mechanism for the /3AR-
agonist response on permeability was related to an elevation of intracellular cAMP, intracellular levels of cAMP were measured at 10 min after incubation with 0.1 and 10/zM ISO, NE and E. Only the high concen- trations of catecholamines induced a 10-15-fold in-
B%
50,
0 ( - r - n . ~ L '100. a t-~,,oml~m~,~,-~ i
- l o g [agonist] M
Fig. 5. Competit ion of /3AR agonists with [1251]IPIN for binding to /3AR subtyes in membranes of endothelial cells. Increasing concen- trations of formoterol (El), ( - ) - isoproterenol ( o ) and ( - ) - no r - epinephrine ( = ) were used to inhibit binding of 0.423 nM [ I~ ] ] IP IN to /3AR. Protein contents per assay tube w e r e 12.8 (El), 10.7 (©) and 12.8 (zx) /zg. Inhibition curves were analyzed according to Eqn. 3 assuming that the fractions o f / 3 A R subtypes were identical to those est imated with highly selective antagonists (see Fig. 4). Estimates of dissociation constants are listed in Table III. Dotted curves represent computer simulations according to Eqn. 2 assuming that only /3AR contribute to the binding of selective antagonists (i.e., f, = 0, ]'2 = 1).
Symbols represent means + S.E. of four determinations.
10~M
4O
"i 3oL
0 ~ C AL PA
E ISO NE NE
Fig. 6. Stimulation of intracellular cAMP in endothelial cells (BAEC) by catecholamines. Confluent cells were incubated with the indicated concentrations of ( - )-epinephrine (E), ( - )-isoproterenol (ISO) and (-)-norepinephrine (NE) in the absence and presence of the aAR- antagonist phentolamine (PA, 100 p.M) or the ~8AR-antagonist al- prenolol (AL, 100 p.M) for 10 min. Cells were preincubated with the antagonists 30 min before challenge with/3AR agonists. Controls (C) were in the absence of any addition of drug. Each incubation was carried out in the presence of 0.2 mM IBMX. The reaction was stopped by removing the medium and addition of 1 ml ice-cold ethanol. Thereafter, the cells were scraped off the dishs and broken by sonification. Cell extracts were used for determination of cAMP by a radioimmunoassay. Columns represent means+S.E, of 4-6
determinations.
crease of intracellular cAMP, independent of the kind of catecholamine. As seen in the permeability experi- ments this increase in intracellular cAMP was abol- ished by preincubation of cells with alprenolol (100 /.tM), whereas the aAR-antagonist phentolamine did not inhibit the stimulation of intracellular cAMP (Fig. 6).
The used catecholamines (10/zM) stimulated intra- cellular cAMP to a similar extent, though the degree of receptor occupancy strongly differed. With the dissoci- ation constants of ISO and NE for /32AR estimated in radioligand binding (Table III) it can be calculated that 10 /zM ISO and 10 /zM NE occupy 99% and 34% of the available /32AR , respectively. Furthermore, a con- centration of ISO (0.1 /zM)which occupies 50% of the /32AR did not elevate the intracellular cAMP content at all. Reasons which may cause these inconsistencies are discussed.
Adenylate cyclase experiments
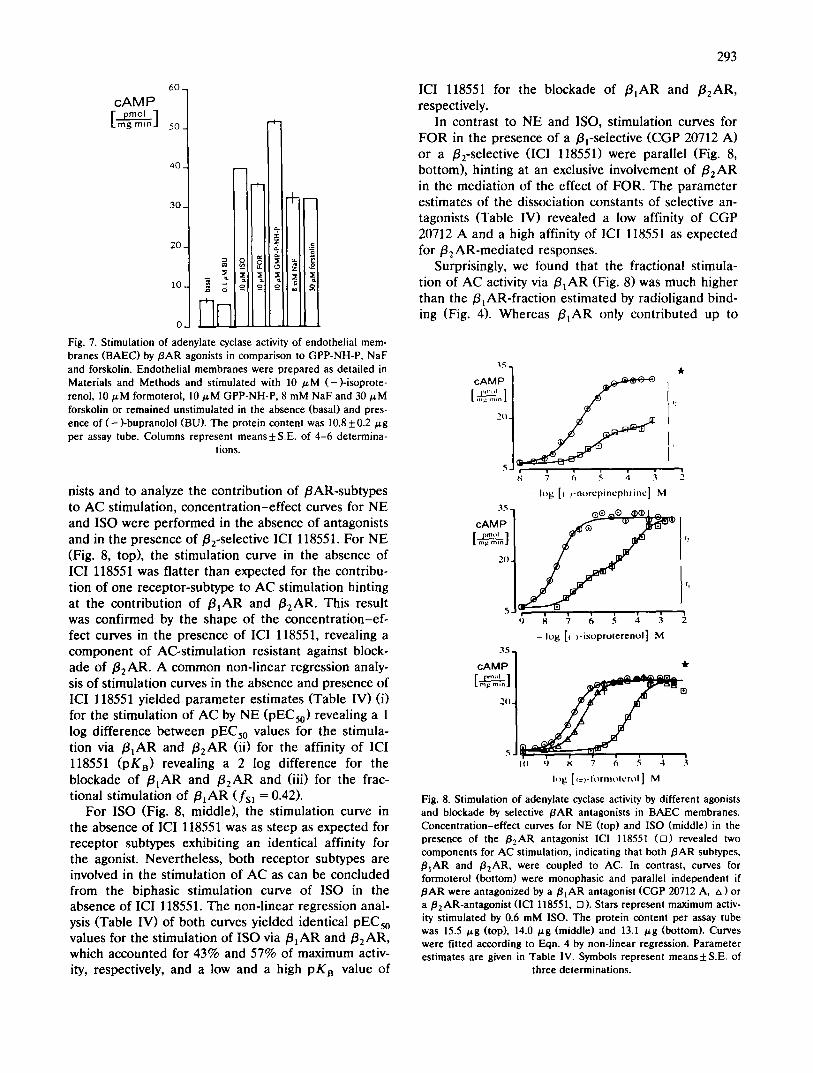
The coupling of /3AR to AC could directly be demonstrated in membranes from BAEC in the pres- ence of 10/zM GTP (Fig. 7). 10/zM ISO increased the rate of cAMP production more than 6-fold over basal activity. Cyclase activity was stimulated to a similar rate by GPP-NH-P, a non-hydrolysable GTP-analogue, NaF and forskolin, a direct stimulant of the catalytic subunit of AC, hinting at a close coupling of /BAR to AC. A similar pattern of AC stimulation was found in differ- ent cell lines from different aortae (data not shown).
In order to measure the concentration dependence of the stimulation of adenylate cyclase by /3AR ago-
c A M P [ mg mmJ
6 0 _
50_
40_
30-
20-
10 , o ~ 1 ~ _
0
-r z ¢
z
Fig. 7. Stimulation of adenylate cyclase activity of endothelial mem- branes (BAEC) by/3AR agonists in comparison to GPP-NH-P, NaF and forskolin. Endothelial membranes were prepared as detailed in Materials and Methods and stimulated with 10 /aM (-)-isoprote- renol, 10/aM formoterol, 10/,M GPP-NH-P, 8 mM NaF and 30/ ,M forskolin or remained unstimulated in the absence (basal) and pres- ence of (-)-bupranolol (BU). The protein content was 10.8 _+ 0.2/zg per assay tube. Columns represent means ± S.E. of 4-6 determina-
tions.
nists and to analyze the contribution of BAR-subtypes to AC stimulation, concentration-effect curves for NE and ISO were performed in the absence of antagonists and in the presence of ~2-selective ICI 118551. For NE (Fig. 8, top), the stimulation curve in the absence of ICI 118551 was flatter than expected for the contribu- tion of one receptor-subtype to AC stimulation hinting at the contribution of /3tAR and /32AR. This result was confirmed by the shape of the concentration-ef- fect curves in the presence of ICI 118551, revealing a component of AC-stimulation resistant against block- ade of fl2AR. A common non-linear regression analy- sis of stimulation curves in the absence and presence of ICI 118551 yielded parameter estimates (Table IV) (i) for the stimulation of AC by NE (pECs0) revealing a 1 log difference between pECs0 values for the stimula- tion via /31AR and /32AR (ii) for the affinity of ICI 118551 (PKB) revealing a 2 log difference for the blockade of /31AR and fl2AR and (iii) for the frac- tional stimulation of fllAR (fsl --- 0.42).
For ISO (Fig. 8, middle), the stimulation curve in the absence of ICI 118551 was as steep as expected for receptor subtypes exhibiting an identical affinity for the agonist. Nevertheless, both receptor subtypes are involved in the stimulation of AC as can be concluded from the biphasic stimulation curve of ISO in the absence of ICI 118551. The non-linear regression anal- ysis (Table IV) of both curves yielded identical pEC50 values for the stimulation of ISO via ,0~AR and/32AR, which accounted for 43% and 57% of maximum activ- ity, respectively, and a low and a high pK B value of
293
ICI 118551 for the blockade of /31AR and /32AR, respectively.
In contrast to NE and ISO, stimulation curves for FOR in the presence of a /31-selective (CGP 20712 A) or a /32-selective (ICI 118551) were parallel (Fig. 8, bottom), hinting at an exclusive involvement of/32AR in the mediation of the effect of FOR. The parameter estimates of the dissociation constants of selective an- tagonists (Table IV) revealed a low affinity of CGP 20712 A and a high affinity of ICI 118551 as expected for fl2AR-mediated responses.
Surprisingly, we found that the fractional stimula- tion of AC activity via flIAR (Fig. 8) was much higher than the/3~AR-fraction estimated by radioligand bind- ing (Fig. 4). Whereas /31AR only contributed up to
3 5 ; .
c A M P
m g m m j
211.
35.
c A M P [ ~_v.v.~_]
mg m l n j
2 0 .
35.
c A M P [ ~._e.~_]
mg r n m j
2 ( I .
- log [i )-norepincphrinc] M
- log [( )-isoproterenol] M
10 ~) X 7 6 5; 4 3
-- h'g [(=>fonnolcrol] M
Fig. 8. Stimulation of adenylate cyclase activity by different agonists and blockade by selective /3AR antagonists in BAEC membranes. Concentration-effect curves for NE (top) and ISO (middle) in the presence of the ,82AR antagonist ICI 118551 (El) revealed two components for AC stimulation, indicating that both ,BAR subtypes, BtAR and ~82AR, were coupled to AC. In contrast, curves for formoterol (bottom) were monophasic and parallel independent if ,BAR were antagonized by a ,81AR antagonist (CGP 20712 A, A ) or a )gzAR-antagonist (ICI 118551, []). Stars represent maximum activ- ity stimulated by 0.6 mM ISO. The protein content per assay tube was 15.5 ~,g (top). 14.0 /,g (middle) and 13.1 ,~g (bottom). Curves were fitted according to Eqn. 4 by non-linear regression. Parameter estimates are given in Table IV. Symbols represent means ± S.E. of
three determinations.

294
T A B L E IV
Dissociation constants of antagonists and ECso ~:alues of agonists for stimulation of fl AR-coupled adenylate cyclase
Values are means and s tandard deviations (ASD) est imated by non-linear regression from experiments depicted in Fig. 8 according to Eqn. 4. pECs0 values ( - l o g (M)) represent ECs0 values of the agonist-induced stimulation of AC. pK n values are dissociation constants ( - log (M)) of antagonists (B) est imated from the blockade of agonist-induced effects by ICI 118551 and CGP 20712 A. fs values ( f s t + fs2 = 1) represent the fractional stimulation of AC through f l lAR and fl2AR, respectively. * FOR did not st imulate AC
via f l IAR.
f l IAR f l2AR
( - )-Norepinephrine ICI 118551 stimulation fraction
pEC50 6.30 _+ 0.08 5.43 +_ 0.06 pKi~ 7.19_+0.10 9.27_+0.16 f s 0.41 + 0.02 0.59 _+ 0.02
( - )-Isoproterenol ICI 118551 stimulation fraction
p E C ~ 7.41 _+ 0.03 7.41 _+ 0.03 pK n 6.98 + 0.12 9.49 + (I.08 f s 0.43 _+ 0.02 0.57 + 0.02
( +_ )-Formoterol (1) ICI 118551 (2) CGP 20712 A
pEC50 - * 7.87 + I).03 pK m - * 8.65+0.03 PKB2 - * 5.32+0.04
13% to maximum binding to f lAR ( B m a x = 4 8 i - 2 fmo l /mg protein, Table III), up to 42% of Vm~ (27.2 _+ 0.3 pmol c A M P / m i n per mg protein) was stimulated through activation of fl~AR. With these data, it can be calculated that maximally activated flzAR stimulates the formation of up to 500 cAMP molecules per rain, whereas maximally activated fl1AR stimulates up to 1800 cAMP molecules per min. This finding is incom- patible with the assumption of a pure additivity of /3~AR-coupled and /32AR-coupled activation of AC.
Discussion
Transendothelial exchange across monolayers of macrovascular endothelial cells (BAEC)
To study the influence of/3-agonists on the endothe- lium we used monolayers built up by macrovascular bovine aortic endothelial cells. The cells showed the typical cobblestone pattern, were stained by antibodies directed against factor VIII antigen and took up acety- lated LDL as has been described to be characteristic for endothelial cells [32,33]. Since it has been suggested that the production of PGI 2 is a characteristic property of endothelial cells [23], the synthesis of prostaglandins by BAEC was determined. As to be expected, BAEC
mainly synthesized PGI 2 if incubated with arachidonic acid. Only trace amounts of TXA 2 and other prosta- glandins could be detected.
The transendothelial exchange of various markers across monolayers of BAEC was quite similar as previ- ously described for this and other types of macrovascu- lar endothelial cells. The exchange rate depends on concentration and molecular mass of the marker molecules, as well as on time and tcmperature [3,25,37]. Molecules with a molecular mass higher than 70 kDa could nearly not pass the endothelial monolayer indi- cating the tightness of the continuous endothelial layer, in agreement with electron microscopic observations (data not shown). From these observations and from the experimental data demonstrating that transendo- thelial exchange of tracer molecules is linear with time (not saturable), we conclude that the transendothelial exchange across endothelial monolayers reflects pure diffusion processes through intercellular clefts [2,3,38]. Since we mainly used FITC-dextrans for the determi- nation of transendothelial exchange, additional forces such as charge selectivity or uptake of the tracer molecules by endothelial cells can be neglected in our model.
The dependences of transendothelial exchange on molecular mass of tracer molecules are compatible with the assumption of two classes of pores [39] with diameters of 1.1 and 17.5 nm for small and large pores, respectively. These diameters are in the same range as those already published from studies using isolated capillaries and monolayers of macrovascular endothe- lial cells [3,37,40,41]. However, a direct comparison of these results is difficult because different experimental models and tissues were used to determine pore sizes. From our data on endothelial permeability it follows that about 0.4% of the endothelial surface is taken for the formation of pores. Similar data have been re- ported for endothelial cells from pulmonar arteries [42]. Thus, our data indicate that the used model of monolayers built up by bovine aortic endothelial cells is suitable to study the regulation of permeability by /3-agonist-mediated processes.
We here present direct evidence that the endothe- lial permeability is directly modulated by activation of fl2AR. This conclusion is based firstly on the use of FOR, a novel highly specific fl2-agonist which has been shown to selectively activate the fl2AR in other tissues with a K d in the range of 10 nM [19]. In good agree- ment with these data, we observed in bovine aortic endothelial cells a K d for receptor binding and activa- tion of adenylate cyclase by the /32AR in the range of 12-14 nM. The presence of /32AR on the surface of macrovascular endothelial cells is further supported by findings previously published by others [12-14]. In con- trast to the K d determined for occupancy of fl2AR and activation of adenalyte cyclase (15 nM), the half-

maximal increase in endothelial tightness was, how- ever, already obtained by 0.4 nM FOR, indicating that a partial occupancy of/32AR is sufficient for maximal reduction of endothelial permeability. Secondly, a re- duction of endothelial permeability can also be achiev- ed by other, less selective catecholamines and dibu- tyryl-cAMP. However, high concentrations (> 1 ~M) of natural occurring (NE) or synthetic (ISO) catechol- amines are necessary to modify endothelial permeabil- ity. Similar results were reported in endothelial cells derived from pig aortae [43], bovine pulmonary artery [9] and human umbilical artery [38], indicating that supra-micromolar concentrations of catecholamines had to be used to modulate endothelial permeability in contrast to the highly specific fl2AR-agonist FOR which reduced endothelial permeability dose-depen- dently with 6000-fold lower concentration (0.42 nM). This latter finding suggests that the endothelial perme- ability is specifically modified by fl2AR-specific action. Lastly, the increase in endothelial tightness was selec- tively prevented and reversed by incubation of the endothelial cells with a /3-antagonist.
To understand the differences in the biological effi- cacy of the specific /32-agonist FOR and the naturally occurring catecholamines in reducing endothelial per- meability, we decided to study the mode of action in more detail. Therefore, we determined the flAR popu- lation on the used endothelial ceils, the coupling be- tween the BAR subtypes and AC and compared the potencies of agonists in receptor-binding and stimula- tion of AC with their functional role in the modulation of endothelial permeability (Fig. 9).
Endothelial cell fl AR and their coupling to AC The observations reported herein provide direct evi-
dence for the existence of two /3AR-subtypes on en- dothelial cells, for the coupling of both subtypes, fl]AR and fl2AR, to AC, and for the the first time for a preferential coupling of the subdominant subtype, /3~AR, to AC. Previous attempts to demonstrate en- dothelial cell f lAR were performed using the racemate of [125I]iodocyanopindolol as radioligand [13,14] which exerts extremely slow dissociation kinetics [44]. In con- trast, we used [25I](-)-iodopindoloi as pure enan- tiomer which exerts rapid binding reactions reaching equilibrium of association and dissociation after 15 and 60 min, respectively [27], and therefore appears to be more appropriate to detect /3AR-subtypes. [~zsI]IPIN binding is specific, saturable, reversible and of high affinity. We decided to assay the binding of [I25I]IPIN in membranes of endothelial cells because of the marked improvement of the ratio of specific to non- specific binding as compared to studies in intact cells [13,14]. This fact improves the determination of recep- tor-subtypes with low binding capacity.
295
/ /
| w , i , • • w , , ,
| | i , , w J ~ ' ~ " j " i , w ,
, , , ~ - ~ : - ~ ' , . . . .
/ /
11 lO 9 a z 6 5 4 3 2
-log [selective agonist] rno~
Fig. 9. Comparison of agonist-induced decrease of endothelial per- meability and stimulation of AC with receptor binding to fl2AR. Computer simulations were performed using ECs0 data from the regulation of endothelial permeability (Fig. 2), ECs0 data from AC stimulation (Table IV) and K d data from radioligand binding (Table III) depicted as full, broken and dotted lines, respectively. Note that for FOR functional effects occur with much lower concentrations than receptor binding and AC stimulation, indicating the existence of
spare receptors.
The contribution of receptor subtypes was analyzed using high affinity, highly-selective antagonists for fl~AR and /32AR. Nanomolar concentrations of CGP 20712 A [21] specifically bind to fllAR, whereas 3000- fold higher concentrations have to be used to displace [12sI]IPIN binding from/32AR [17,45]. For an indepen- dent prove of coexisting /3~AR and /32AR a ligand with an inverse selectivity, ICI 118551, was used; nanomolar and 100-fold higher concentrations of ICI 118551 specifically bind to /32AR and fllAR, respec- tively [15,19]. Competition-binding experiments with both antagonists analyzed by a common non-linear regression procedure clearly indicated the coexistence of/31AR and fl2AR in a proportion of/3~AR to fl2AR of 13%:87%. The 13% component of/31AR in BAEC was lower than the 25% /3~AR-fraction estimated by Ahmad and co-workers in bovine pulmonary endothe- lial cells [14]; however, this difference may possibly not reflect a difference in aortic and pulmonary cells, but may depend on methodological differences: Ahmad and co-workers [14] (i) used intact cells instead of membranes resulting in higher degree of non-specific binding, (ii) they used /3tAR ligands with a 100-fold lower selectivity than observed with CGP 20712 A and (iii) they did not submit their data to an objective non-linear regression analysis. The estimated dissocia-

296
tion constants of both antagonists for the OAR sub- types on BAEC (Table III) match the respective con- stants estimated for pulmonary and myocardial 01AR and 02AR [15,17,19,45]. These data ensure the identity of the detected EAR subtypes in endothelial cell with the classical subtype of the 0-adrenergic receptor, /31AR and 02AR, as characterized in many organs of mammals, including humans. No indication was found hinting at a contribution of 03AR postulated by others [14].
The identification of EAR subtypes as 0~AR and 32AR is further supported by their affinity for EAR- agonists. Formoterol, known as high-affinity agonist for 02AR with a 300-fold lower affinity for 31AR as as- sessed in guinea-pig pulmonary and myocardial mem- branes [19], exerted high and low affinity for the main (87%) and the minor (13%) component of [125I]IPIN binding in endothelial cells. In contrast, ISO was non- selective for 3~AR and 02AR as found in many stud- ies in heart and lung [15,46-48]. The above conclusion was further supported by the findings that NE exerted a 50-fold higher affinity for endothelial 0 ,AR than for O2AR; the respective dissociation constants match data reported for lung and heart flAR [15,46-48].
Similarly to radioligand binding, the presence of both 0-adrenergic receptor subtypes could be demon- strated in assaying receptor-stimulated AC activity. Comparing the EC50 values of concentration-effect curves for EAR agonists and the dissociation constants for blockade by highly selective antagonists with data from heart [17,31,45,49] and lung [16,19,49] supported our conclusion that endothelial cell EAR are identical with 01AR and 0zAR. Surprisingly, however, we found for both catecholamines that /3~AR are coupled to endothelial AC much more effectively (Vm,x= 1800 cAMP molecules per stimulated B1AR) than 02AR (V,,ax = 500 cAMP molecules per stimulated 02AR). This observation is in contrast to the assumption of additivity of stimuli, usually expected for coexisting 31AR and 02AR which are coupled to the same sig- nal-transduction pathway.
FOR, a high-affinity 02AR agonist [19], stimulated AC activity significantly less than NE and ISO. Further insight into that apparent loss of intrinsic activity of FOR compared to NE and ISO was found by inhibi- tion of FOR effects on AC by highly-selective antago- nists. These experiments elucidated that only 02AR were involved in the stimulation of AC by FOR even when high FOR-concentrations (> 10 tzM) were used which also significantly bind to 0 ,AR (Ka = 5 ~M). The maximum stimulation of AC by FOR matched the stimulation maximum by ISO and NE via 02AR, i.e., in the presence of a highly effective 0~AR-blockade (80% of Vma x, see Table IV), indicating that FOR exhibited full agonistic potency for AC activation via /32AR. Similar results were found in membranes from
guinea-pig lung and from desaggregated tracheal smooth muscle cells [19] characterizing FOR as full agonist for 02AR.
The relation between the regulation of endothelial perme- ability, receptor binding and stimulation of AC by EAR agonists
Evidence is presented, for the first time, that spare receptors [50] are involved in the 0-adrenergic regula- tion of endothelial cell permeability by 0zAR-agonists. Less than 10% receptor occupancy of 02AR by FOR was sufficient to induce an effective reduction of per- meability (Fig. 2). The spare receptor capacity amount- ing to a 50-fold amplification of receptor signals was of similar height as estimated from tracheal relaxation by FOR compared to its binding to pulmonary mem- branes [19]. Based upon these findings it may be as- sumed that inhaled FOR used in pharmacotherapy of bronchial asthma [20] not only causes a relaxation of bronchial smooth muscle tone but also prevents the developement of lung edema by reducing transvascular flux. The latter mechanism is thought to contribute to the beneficial role of 0-adrenergic bronchodilator therapy [6]. Our finding of a very sensitive modulation of endothelial permeability by a OzAR agonist are in support to the conclusions made by Minnear and co- workers [9], who postulate that 0AR-agonists reduce the transvascular flux not only by reducing microvascu- lar pressure but also by a direct action on endothelial cells.
Extrapolating the very sensitive modulation of en- dothelial permeability by FOR (a 50-fold amplification between binding (K d) and function (ECs0)) to the experiments with catecholamines it should be expected from the dissociation constants (K d) for the occupancy of O2-AR by ISO (100 nM) and NE (30 ~M) that half maximum effects (ECs0) for the reduction of perme- ability should occur in the range of 3 nM (ISO) and 1 /zM (NE). In contrast to the above hypothesis, how- ever, supramicromolar concentrations had to be used to modulate permeability by ISO and NE, indicating a significant loss of efficacy (ECs0 >> K d) which becomes most evident with ISO. What are the reasons for that loss of efficacy occurring with catecholamines but not with a non-catecholamine 02AR-agonist? A first hint at the cause for that discrepancy was found in studying catecholamine-induced elevation of intracellular cAMP. A concentration of ISO (0.1 /xM) which is expected from its dissociation constant in binding (Kj = 0.1 ~.M) to induce a half-maximal increase in cAMP, was rather ineffective in stimulating intracellular cAMP though phosphodiesterases were blocked with IBMX. Only concentrations 100-fold higher than Kj signifi- cantly stimulated intracellular cAMP. Thus, we assume that the free concentrations of catecholamines were markedly reduced by degradation by catecholamine-dc-

297
grading enzymes such as catechol-O-methyl transferase 11 (COMT) [51] and/or by monoamine oxidase (MAP) [52]. Such a catalytic activity of catecholamine-degrad- • 12 ing enzymes was not only found in endothelial cells 13 derived from cerebral arteries [53] but also in rat aorta [54] and bovine aortic cells [51]. The above assumption 14 is further supported by the finding, that the effects of FOR, which is a non-catecholamine/32AR agonist, was 15 not attenuated in functional experiments on endothe- 16 lial cell monolayers.
Surprisingly, changes in endothelial permeability ex- 17 erted by NE and the increase in cAMP content oc- curred in the same dose-range as with ISO though the 18
19 affinity of NE for f l E A R was 100-fold lower that of ISO (Table III). This finding can be taken as an indirect 20 hint at the contribution of/31AR to the noradrenergic modulation of permability in accordance to the highly- 21
effective coupling of flIAR to AC and to the 50-fold 22 higher affinity of NE for/31AR. A matching concentra- tion-dependence of NE and ISO for the elevation of 23
cAMP, which is not characteristic for/32AR [46], was also found by others [55]. Further experiments, which are designed to elucidate the contribution of degrada- tive enzymic activity in endothelial cell monolayers and to demonstrate /3tAR-mediated reduction of perme- ability by synthetic, non-catecholamine /31AR agonists are currently under investigation.
Acknowledgements
The writers thank Thilo Salmon for computer work, Sylvia Berger for photography (Institute for Laser- medicine, Diisseldorf, Germany) and Ciba-Geigy (Basel, Switzerland) for a gift of (+)-formoterol. This work was supported by the Wissenschaftsministerium of Nordrhein-Westfalen and the Bundesministerium fiir Jugend, Familie und Gesundheit.
References
1 Furie, M.B., Cramer, E.B., Naprstek, B.L. and Silverstein, S.C. (1984) J. Cell Biol. 98, 1033-1041.
2 Simionescu, M. and Simionescu N. (1986) Annu. Rev. Physiol. 48, 279-293.
3 Abelda, S.M., Sampson, P.M., Haselton, T.R., McNiff, J.M., Mueller, S.N., Williams, S.K., Fishman, A.P. and Levine, E.M. (1988) J. Appl. Physiol. 64, 308-322.
4 Suval, W.D., Duran, W.N., Boric, M.P., Horson, R.W., Berend- sen, P.B. and Ritter, A.B. (1987) Am. J. Surg. 154, 211-218.
5 Stender, S. and Hjelms, E. (1987) Scand. J. Clin. Lab. Invest. 47 (Suppl. 186), 21-29.
6 Persson, C.G.A., Ekman M. and Erjefalt, I. (1979) Acta Pharma- col. Toxicol. 44, 216-220.
7 Dvorak, H.F. (1986) N. Engl. J. Med. 315, 1650-1659. 8 Majno, G. and Palade, G.E. (1961) J. Biophys. Biochem. Cytol.
11,571-605. 9 Minnear, F.L., Demichele, M.A.A., Moon, D.G., Rieder, D.L.
and Fenton II J.W. (1989) Am. J. Physiol. 257, H1613-H1623. 10 Svensjo, E., Persson, D.G.A. and Rutili, G. (1976) Acta Physiol.
Scand. 101,504-506.
Grega, G.J. Maciejko, J.J., Raymond, R.M. and Sak, D.P. (1980) Circ. Res. 46, 264-275. Schafer, A.I., Gimbrone, M.A. and Handin, R.I. (1980) Biochem. Biophys. Res. Commun. 4, 1640-1647. Steinberg, S.F., Jaffe, E.A. and Bilezikian, J.P. (1984) Naunyn- Schmiedeberg's Arch. Pharmacol. 325, 310-313. Ahmad, S., Chretien, P., Daniel, E.E. and Shen, S.H. (1990) Life Sci. 47, 2365-2370. Lemoinc, H., Ehle, B. and Kaumann, A.J. (1985) Naunyn- Schmiedeberg's Arch. Pharmacol. 331, 40-51. Lemoine, H., Novotny, G.E.K. and Kaumann, A.J. (1989) Naunyn-Schmiedeberg's Arch. Pharmacol. 339, 85-98. Lemoine, H. and Kaumann, A.J. (1991) Naunyn-Schmiedeberg's Arch. Pharmacol. 344, 56-69. Ida, H. (1976) Drug Res. 26, 839-842. Lemoine, H. and Overlack, C. (1992) J. Pharmacol. Exp. Ther. 261,258-270. Becker, A.B. and Simons, F.E.R. (1989) J. Allergy Clin. Immunol. 84, 891-895. Dooley, D.J., Bittiger, H. and Reymann, N.C. (1986) Eur. J. Pharmacol. 130, 137-139. Booyse, F.M.," Sedlak, B.J. and Rafelson, M.E. (1975) Thrombos. Diathes. Haemorrh. 31,825-839. Oestreich, R., Zink, S. and R6sen, P. (1989) in Progress in Clinical and Biological Research, Prostaglandins in Clinical Re- search, Cardiovascular System, Vol. 301 (Schr6r, K. and Sinzinger H., eds.), pp. 371-375, Alan R. l_,iss, New York.
24 Hohl, C.M. and R6sen, P. (1987) Biochim. Biophys. Acta 921, 356-363.
25 Siflinger-Birnboim, A., Del Vecchio, P.J., Cooper, J.A., Blumen- stock, F.A., Shepard, J.M. and Malik, A.B. (1987) J. Cell. Physiol. 132, 111-117.
26 Del Vecchio, P.J., Siflinger-Birnboim, A., Shepard, J.M., Bizios R., Cooper, J.A. and Malik, A.B. (1987) Fed. Proc. 46, 2511-2515.
27 Hedberg, A., Kempf, F., Josephson, M.E. and Molinoff, P.B. (1985) J. Pharmacol. Exp. Ther. 234, 561-568.
28 Ehle, B., Lemoine, H. and Kaumann, A.J. (1985) Naunyn- Schmiedeberg's Arch. Pharmacol. 331, 52-59.
29 Lemoine, H. (1992) Ouant. Struct. Act. Relat. 11,211-218. 30 Salomon, Y., Londos, C. and Rodbell, M. (1974) Anal. Biochem.
58, 541-548. 31 Gille, E., Lemoine, H., Ehle, B. and Kaumann, A.J. (1985)
Naunyn-Schmiedeberg's Arch. Pharmacol. 331, 60-70. 32 Voyta, J.C., Via, P., Buttertield, C.E. and Zetter, B.R. (1984) J.
Cell Biol. 99, 2034-2040. 33 Jaffe, E.A., Hoyer, L.W. and Nachman, R.L. (1973) J. Clin.
Invest. 52, 2757-2764. 34 Van Brec, J.B.M.M., de Boer, A.G., Danhof, M., Ginsel, L.A.
and Breimer, D.D. (1988) J. Pharmacol. Exp. Ther. 247, 1288- 1239.
35 Peters, W. and Wiese, B. (1986) J. Insect. Physiol. 32, 43-49. 36 Gerritsen, M.E. (1987) Biochem. Pharmacol. 36, 2701-2711. 37 Shasby, D.M. and Roberts, R.L. (1987) Fed Proc. 46, 2506-2510. 38 Langeler, E.G. and Van Hinsbergh, V.W.M. (1991) Am. J. Phys-
iol. 260, C1052-C1059. 39 Pappenheimer, J.R. (1953) Physiol. Rev. 33, 387-444. 40 Curry, F.E., Huxley, V.H. and Adamson, R.H. (1983) Am. J.
Physiol. 245, H495-H505. 41 Renkin, E.M. (1985) J. Gen. Physiol. 52, 96-107. 42 Powers, M.R., Blumenstock, F.A., Cooper, J.A. and Malik, A.B.
(1989) J. Cell Physiol. 141,558-564. 43 Gudgeon, J.R. and Martin, W. (1989) Br. J. Pharmacol. 98,
1267-1274. 44 Engel, G., Hoyer, D., Berthold, R. and Wagner, H. (1981)
Naunyn-Schmiedeberg's Arch. Pharmacol. 317, 277-285. 45 Kaumann, A.J. and Lemoine, H. (1987) Naunyn-Schmiedeberg's
Arch. Pharmacol. 335, 403-411.

298
46 Lands, A.M., Arnold, A., McAuliff, J.P., Luduena, F.P. and Brown, T.G. (1967) Nature 214, 597-598.
47 Minneman, K.P., Hegstrand, L.R. and Molinoff, P.B. (1979) Mol. Pharmacol. 16, 21-33.
48 Frielle, T., Caron, M.G. and Lefkowitz, R.J. (1989) Clin. Chem. 35, 721-725.
49 Kaumann, A.J. and Lemoine, H. (1985) Naunyn-Schmiedeberg's Arch. Pharmacol. 331, 27-39.
50 Stephenson, R.P. (1956) Br. J. Pharmacol. 11,379-393.
51 Shepro, D. and Dunham, B. (1986) Annu. Rev. Physiol. 48, 335 -345.
52 Roth, J.A. and Venter, J.C. (1978) Biochem. Pharmacol. 27, 2371-2773.
53 Scriba, G.K.E. and Borchardt, R.T. (1989) J. Neurochem. 53, 610-615.
54 Head, R.J., Panek, R., Reid, J., Stitzel, R.E. and Barone, S. (1986) Blood Vessels 23, 279-287.
55 Makarski, J.S. (1981) In Vitro 17, 450-458.




















