
Oligomeric structure of the α1b-adrenoceptor: Comparisons with rhodopsin

Experimental Gerontology 45 (2010) 918–935
Contents lists available at ScienceDirect
Experimental Gerontology
j ourna l homepage: www.e lsev ie r.com/ locate /expgero
Age-associated plasticity of α1-adrenoceptor-mediated tuning of T-cell development
G. Leposavić a,⁎, V. Pešić a, Z. Stojić-Vukanić b, K. Radojević c, N. Arsenović-Ranin b, D. Kosec c,M. Perišić c, I. Pilipović c
a Department of Physiology, Faculty of Pharmacy, University of Belgrade, 450 Vojvode Stepe, 11221 Belgrade, Serbiab Department of Microbiology and Immunology, Faculty of Pharmacy, University of Belgrade, Belgrade, Serbiac Immunology Research Centre “Branislav Janković”, Institute of Virology, Vaccines and Sera “Torlak”, 458 Vojvode Stepe, 11221 Belgrade, Serbia
T-cell development
⁎ Corresponding author. Tel.: +381 11 3951 209.E-mail address: [email protected]
0531-5565/$ – see front matter © 2010 Elsevier Inc. Aldoi:10.1016/j.exger.2010.08.011
a b s t r a c t
a r t i c l e i n f oArticle history:Received 2 March 2010Received in revised form 10 August 2010Accepted 17 August 2010Available online 26 August 2010
Section Editor: R. Effros
Keywords:AgeingNoradrenalineα1-adrenoceptors
Rat thymus
Alpha1-adrenoceptors (α1-ARs) are involved in neuro-thymic and thymic intercellular communications, andconsequently modulation of T-cell development. Ageing is associated with a number of changes innoradrenergic neuro-effector transmission, and possibly intercellular noradrenaline (NA)-mediatedcommunication resulting in altered responses of target cells to NA. Thus, in old animals an altered NAmodulation of thymopoiesis via α1-ARs may be expected. To test this hypothesis, in old and young adultWistar rats we examined: 1) thymic NA levels, density of noradrenergic innervation and NA synthesizingcells, as well as α1-AR expression, and 2) then the effects of 14-day-long treatment with the α1-AR blocker,urapidil, on thymocyte development. Overall, the first part of study suggested augmented NA signalling tothymic cells via α1-ARs due to increased NA availability and α1-AR thymocyte surface density in old rats. Thesecond part of study supported this assumption. Namely, although in rats of both ages urapidil affected thesame thymocyte developmental steps ultimately leading to changes in the relative number of the mostmature single positive TCRαβhigh thymocytes, its effects were generally more prominent in old animals.Following urapidil treatment, the percentages of CD4+CD8− cells, including those showing a regulatoryCD4+CD25+RT6.1− phenotype, were increased, while CD4−CD8+ cells decreased. In old rats, anaugmented thymic escape of immature CD4+CD8+ cells was also registered. In rats of both ages the thymicchanges were accompanied by alterations in the proportions of major cell populations in the T-lymphocytecompartment of both peripheral blood and spleen, leading to an increase in the CD4+/CD8+ T-cell ratio.These alterations were also more pronounced in old rats. Moreover, in old rats following urapidil treatmentthe proportion of TCRαβ+cells in the periphery was slightly greater reflecting, most likely, partly enhancedthymic production of regulatory CD161+TCRαβ+cells. Thus, the study indirectly suggests an age-associated increase in the basal α1-AR-mediated inhibitory influence of NA on thymopoiesis.
.rs (G. Leposavić).
l rights reserved.
© 2010 Elsevier Inc. All rights reserved.
1. Introduction
Generally, ageing is associated with a number of changes innoradrenergic neuro-effector transmission, resulting in an alteredresponse to the neurotransmitter. Neurotransmission is likely to beaffected by changes in nerve activity, density of innervation andneurotransmitter disposition mechanisms, as well as by alterations intarget organ responsiveness, which may involve alterations at thereceptor and post-receptor level or even alterations in tissue structure(Docherty, 2002).
The thymus is richly innervated by postganglionic sympatheticnoradrenergic fibers. Noradrenergic nerve profiles have been ob-served not only within the thymic capsular and trabecular system, butalso within thymic parenchyma ending in close proximity to
thymocytes and non-lymphoid cells driving T-cell development(reviewed in Elenkov et al., 2000; Leposavić et al., 2008). In addition,noradrenaline (NA) was recently reported to be produced by thymiclymphoid and non-lymphoid cells (Pilipović et al., 2008). Moreover, itwas suggested that, apart fromNA of neural origin, thymic cellular NA,acting in both autocrine and paracrine ways, is involved inmodulationof T-cell development (Pilipović et al., 2008). This role of NA iscorroborated by the detection of β- and α1-adrenoceptors (ARs) onthymic lymphoid and non-lymphoid cells (Loveland et al., 1981;Marchetti et al., 1994; Kavelaars, 2002; Pešić et al., 2009). Further-more, pharmacological studies implicated both subtypes of ARs in NA-mediated tonic inhibition of T-cell development (Plećaš-Solarovićet al., 2004, 2005; Leposavić et al., 2006a; Pešić et al., 2009).
In rodent thymi, ageing has an impact on the density ofnoradrenergic nerve fibers and NA concentration (Bellinger et al.,1988; Madden et al., 1997; Cavallotti et al., 1999), but the influence ofageing on thymic cellular NA production has not been investigatedyet. Furthermore, age-associated alterations in the efficiency of β-AR-

919G. Leposavić et al. / Experimental Gerontology 45 (2010) 918–935
mediated modulation of T-cell development have been reported(Madden and Felten, 2001; Pešić et al., 2007). However, althoughmany types of cells have been demonstrated to exhibit age-relatedchanges in sensitivity to α1-AR signalling (Yokoyama et al., 1984;Basso and Piantanelli, 2002), there are no data related to the influenceof ageing on α1-AR-mediated intrathymic communication, andconsequently modulation of thymopoiesis.
Uncovering age-associated changes in NA-mediated intrathymiccommunications may be important for better understanding ofcomplex multifactorially driven alterations in thymopoiesis in oldage (Kelley et al., 1986; Li et al., 1992; Hirokawa et al., 1994; Fabriset al., 1997; Lau and Spain, 2000; Hsu et al., 2005; Leposavić andPerišić, 2008). Since T cells are central to adaptive immunity,knowledge about the mechanisms underlying detrimental changesin T-cell development with increasing age is crucial for understandingage-related decay in immune functions (Berzins et al., 2002;Globerson, 2002; Gruver et al., 2007). The changes in T-cellcompartment with age encompass narrowing diversity of the T-cellrepertoire and disruption of the major T-cell population balancewithin the peripheral lymphoid compartment (De Paoli et al., 1988;Gilman-Sachs et al., 1991; Pahlavani and Richardson, 1994; Aspinall andAndrew, 2000; Globerson, 2002; Clambey et al., 2007; Czesnikiewicz-Guzik et al., 2008), mainly due to the thymus-dependent decline inCD4+ lymphocyte abundance in combination with oligoclonalexpansion in the CD8 T-cell subset (De Paoli et al., 1988; Gilman-Sachs et al., 1991; Pahlavani and Richardson, 1994; Clambey et al.,2007; Czesnikiewicz-Guzik et al., 2008). Clinical consequences arepoor responses to new antigens or vaccinations, raised infection rateswith higher morbidity and mortality, and increasing incidence ofautoimmune diseases with advancing age (Aspinall and Andrew,2000; Pawelec et al., 2005; Aw et al., 2007).
Having all the previously stated in mind, we aimed to explore theputative influence of ageing on neuro-thymic and thymic intercellularα1-AR-mediated NA communications. To this end, in thymi fromyoung adult and old Wistar rats we examined: i) the density ofnoradrenergic nerve fibers and NA synthesizing cells, thymic andthymocyte NA levels and α1-AR expression and ii) the effects of 14-day-long treatment with urapidil on T-cell differentiation/maturation.In addition, to determine if the changes in thymopoiesis translatedinto the periphery, the proportions of major T-lymphocyte popula-tions in peripheral blood and spleen, as an example of a secondarylymphoid organ, were examined.
2. Materials and methods
2.1. Animals
In this study maleWistar rats purchased from the Medical MilitaryAcademy, Belgrade were used. This substrain of Wistar rats waschosen since thymi from these aged animals exhibit morphologicalcharacteristics similar to those found in aged human thymus (Plećaš-Solarović et al., 2004).
2.2. Experimental protocols
The first of two sets of experiments was undertaken to exploreage-associated changes in: i) thymic NA level; ii) thymocyte NAcontent and tyrosine hydroxylase (TH) expression, iii) distribution ofnoradrenergic nerve fibers and TH-immunoreactive cells and iv)distribution of α1-AR expressing cells and α1-AR thymocyte surfacedensity. For this purpose, old (18-month-old) and young (2.5-month-old) rats were killed and their thymi were dissected out. Thymic leftlobes were processed for quantification of NA by high performanceliquid chromatography (HPLC) or immunostaining of cells expressingTH andα1-ARs, while thymic right lobes were snap frozen for analysis
of fluorescent noradrenergic nerve distribution or processed for flowcytometric analysis (FCA) of thymocyte TH and α1-AR expression.
In the second set of experiments, old and young rats wererandomly assigned to an experimental condition: the α1-AR antag-onist, urapidil, or saline administration. Accordingly, over 14consecutive days the animals were injected subcutaneously withurapidil (Ebrantil, Byk Gulden, Germany) at 0.20 mg/kg BW/day orwith an equivalent volume of saline. This particular dose and durationof treatment were chosen on the basis of previous experimentsshowing that such treatment significantly affected thymocytedevelopment in young rats (Pešić et al., 2009). Following thetreatment all animals were euthanized by exposure to increasingdoses of CO2 followed by cardiac puncture exsanguination. The spleenand thymus were then carefully dissected out. The left thymic lobeswere processed for stereological and immunocytochemical analysesof apoptotic and proliferating cells, while the right lobes and spleenswere used for cell phenotyping by FCA. Each group consisted of atleast six animals.
Animal care and experimental procedures were carried out inaccordance with the principles described in the European Commu-nity's Council Directive of 24 November 1986 (86/609/EEC).
2.3. Chemicals, antibodies and immunoconjugates
All chemicals and standards (L-noradrenaline hydrochloride,L-adrenaline hydrochloride, 3,4-dihydroxybenzylamine) for HPLC,Concanavalin A (Con A) and RPMI 1640 powdered medium werepurchased from Sigma-Aldrich Chemie GmbH (Taufkirchen,Germany). RPMI 1640 powdered medium was dissolved in redistilledwater according to the manufacturer's instructions. To preparecomplete RPMI medium, 2 mM L-glutamine (Serva, Heidelberg,Germany), 1 mM sodium pyruvate (Serva), 100 units/ml penicillin(ICN, Costa Mesa, CA, USA), 100 μg/ml streptomycin (ICN) and 10%fetal calf serum (FCS) (Gibco, Grand Island, NY, USA) were added.Liquid DAB+substrate–chromogen system, LSAB+system, antibodydiluent, Faramount aqueous mounting medium and target retrievalsolution were obtained from DakoCytomation, Glostrup, Denmark.
For immunostaining the following first-step antibodies (Abs)were used: phycoerythrin (PE)-conjugated anti-CD4 (clone W3/25,Serotec, Oxford, UK), fluorescein isothiocyanate (FITC)-conjugatedanti-CD8 (clone MRC OX-8, Serotec), peridinin chlorophyll protein(PerCP)-conjugated anti-TCRαβ (clone R73, BD Biosciences Phar-mingen, Mountain View, CA, USA), biotin-conjugated anti-CD25(clone MRC OX-39, Serotec), biotin-conjugated anti-CD90 (Thy-1.1)(clone OX-7, BD Biosciences Pharmingen), anti-RT6.1 (clone P4/16,Serotec), FITC-conjugated anti-CD161 (clone 10/78, Serotec), anti-Ki-67 (clone B56, BD Biosciences Pharmingen), anti-rat tyrosinehydroxylase (TH, clone TOH A1, BD Biosciences Pharmingen), andgoat anti-rat α1B-AR polyclonal Ab (N-20, Santa Cruz Biotechnology,CA, USA). As second-step reagents were used: streptavidin-PerCP(BD Biosciences Pharmingen), streptavidin-PE (BD BiosciencesPharmingen), F(ab′)2-anti-rat IgG FITC (Serotec), FITC-conjugatedrabbit anti-goat IgG (Sigma-Aldrich Chemie) and FITC-conjugatedgoat anti-mouse IgG (BD Biosciences Pharmingen). Appropriate IgGisotype controls were obtained from BD Biosciences Pharmingen.
2.4. HPLC with electrochemical detection
Briefly, for measurement of NA level, thymic tissue and 1×108
thymocyte suspension homogenized in ice-cold 0.1 M perchloric acid(Sigma-Aldrich) containing 0.15% Na2S2O5 (Sigma-Aldrich) and 0.05%Na2EDTA (Sigma-Aldrich) were centrifuged at 30,000×g for 20 min aspreviously described (Pilipović et al., 2008). The supernatants werepassed through 0.2-μm filters and 25 μl aliquots were injected into aGBC high-performance liquid chromatograph (LC1120 pump, GBC,Victoria, Australia) with an electrochemical detector (LC1260, GBC)

920 G. Leposavić et al. / Experimental Gerontology 45 (2010) 918–935
and column Hibar® 125-4 LiChrospher® 100 RP-18 (5 μm) (MerckKGaA, Darmstadt, Germany).
2.5. Microscopy
2.5.1. Stereological analysisFive μm thick thymic sections were stainedwith haematoxylin and
eosin. Every 40th section (20 sections per organ) was subjected toanalysis of absolute volumes of lymphoid, so-called true thymic tissueand connective/adipose tissue. Stereological measurements wereperformed by the point counting method using an Olympus BX50microscope (Olympus, Tokyo, Japan) equipped with a digital cameraand image analysis software (Micro Image Version 4.0, Olympus,Hamburg, Germany) as previously described (Plećaš-Solarović et al.,2004). All the stereological data refer to fixed thymic tissue. Therelative shrinking of the thymic tissue was approximately 34%.
2.5.2. Immunohistochemical stainingBriefly, thymic sections were subjected to antigen retrieval by heat
pretreatment in Target Retrieval Solution. After quenching endogenousperoxidase activity (10 min, 3% H2O2 in water), the sections werewashed in phosphate-buffered saline (PBS, pH 7.4) and incubated withone of the following antibodies: anti-α1B-AR (1:40), anti-TH (1:1000)and anti-Ki-67 (1:150) overnight at 4 °C. After incubation, the sectionswere washed in PBS, and immunolabelling was visualized using acommercial LSAB+kit with horseradish peroxidase-based stainingsystem. The images of α1B-AR and TH staining were obtained with anOlympus BH-2 microscope (Olympus, Tokyo, Japan) equipped with adigital camera and AnalySIS 5.0 image analysis software (Olympus,Hamburg, Germany), while the images of Ki-67 staining were obtainedwith Olympus BX50 microscope (Olympus, Tokyo, Japan) equippedwith a digital camera and image analysis software (Micro Image Version4.0, Olympus, Hamburg, Germany). The immunostained cells werecounted using a 40x objective lens by an observer with no previousknowledge of which slide was being analyzed. The average of tenrandomly placedmicroscopic fields on lymphoid tissue was recorded infive sections of the thymus from each rat, and the number ofimmunoreactive cells was expressed per mm2.
2.5.3. Fluorescence microscopy
2.5.3.1. Analysis of catecholamine-containing nerve fibers. Thymicblocks were sectioned at 20 μm on a cryostat at −20 °C. The sectionswere thaw mounted onto slides and stained using a modification ofthe sucrose phosphate glyoxylic acid (SPG) method (Mićić et al.,1994). Briefly, the thymic sections were dipped in a solutioncontaining 1% glyoxylic acid (Sigma-Aldrich), 0.2 M sucrose (Sigma-Aldrich) and 0.236 M monobasic potassium phosphate (pH 7.4)(Sigma-Aldrich), drained, covered with non-autofluorescent immer-sion oil, heated at 95 °C for 2.5 min and coverslipped. Catecholamine(CA)-containing nerve terminals were visualized using an OlympusBH-2 microscope equipped with epi-illumination accessories, digitalcamera and AnalySIS 5.0 image analysis software.
2.5.3.2. Analysis of apoptotic cells. Apoptotic cells in situ in the thymuswere visualized using Annexin V staining. Five μm thick thymicsections were washed in PBS and then incubated with FITC-conjugated Annexin V (BD Biosciences Pharmingen, Germany) for1 h in a humidified chamber at room temperature. After incubationthe cells were washed in PBS and examined using an Olympus BX50fluorescence microscope equipped with epi-illumination accessories,digital camera and Micro Image Version 4.0 software (Olympus,Hamburg, Germany). Annexin V+cells were counted in the samewayas immunostained cells.
2.6. Flow cytometry
2.6.1. Single-cell suspension preparationFor single-cell suspensions, thymus and spleen tissues were
dissociated by grinding on a sterile 60-μm sieve screen submergedin ice-cold PBS, pH 7.3 supplemented with 2% FCS and 0.01% NaN3
(FACS buffer). Blood samples were taken by cardiac puncture. Bloodsamples and single-cell splenocyte suspensions were subjected toammonium chloride lysis to remove red blood cells. The resulting cellsuspensions were washed in ice-cold FACS buffer. The cells insuspension were enumerated using an improved Neubauer hemocy-tometer and trypan blue dye to exclude non-viable cells.
2.6.2. Analysis of surface antigen expressionThe immunolabeling was performed as previously described
(Pešić et al., 2009). Briefly, aliquots of 1×106 cells were incubatedfor 30 min on ice in the dark with fluorochrome-conjugated mAbs(direct staining) or with biotin-conjugated/unconjugated mAbs(indirect staining) before being washed twice with FACS buffer.When biotin-conjugated and unconjugated mAbs were used, the cellswere subsequently incubated with fluorochrome-conjugated strepta-vidin and FITC-conjugated anti-goat IgG/F(ab′)2-anti-rat IgG, respec-tively. After incubation, the cells were washed, and twenty thousandcells per sample were analyzed using a FACScan flow cytometer(Becton Dickinson, Mountain View, CA, USA). The percentage ofpositive cells for each labeling was determined using CellQuestsoftware (Becton Dickinson). In addition, age-associated changes inα1-AR expression and urapidil-induced changes in the density ofthymocyte surface CD90 were estimated by analyzing the relativechange in mean channel fluorescence intensity (MFI) according toKamath et al. (1998).
2.6.3. Analysis of tyrosine hydroxylase (TH) expressionFor TH staining, 1×107 cells were resuspended and fixed with
0.25% paraformaldehyde (Sigma-Aldrich) in PBS (30 min at 4 °C).Fixed cells were washed twice in ice-cold PBS containing 0.1% Tween-20 (Sigma-Aldrich) and 0.09% NaN3 (Sigma-Aldrich) (washingsolution) and incubated with permeabilizing solution (PBS containing0.2% Tween-20, 0.09% NaN3 and 2% FCS for 15 min at 4 °C.Permeabilized cells were washed twice in ice-cold washing solution,and then incubated subsequently with anti-rat TH mAbs and FITC-conjugated anti-mouse IgG. The cells were analyzed using a FACScanflow cytometer. The analysis included estimation of age-relatedchanges in MFI for TH (Kamath et al., 1998).
2.6.4. Analysis of thymocyte proliferation in vitroBromodeoxyuridine (BrdU)/7-Aminoactinomycin D (7-AAD) Flow
kit (BD Biosciences Pharmingen) was utilized to identify proliferatingcells in vitro. A total of 2×105 cells/well (100 μl) of thymocytes fromurapidil- or saline-injected rats were dispersed into plastic 96-wellplates (Nunc A/S, Roskilde, Denmark) and cultured for 48 h at 37 °C in5% CO2 humidified air atmospherewithout or with 2.5 μg/ml ConA in atotal volume of 200 μl of RPMI 1640 culturemedium. All cultures wererun in triplicate. During the last 18 h of culture the cells were pulsedby 1 μM BrdU/well. Staining was performed according to producer'smanual. Twenty thousand cells per sample were analyzed on aFACScan flow cytometer by CellQuest software (Becton Dickinson).
2.6.5. Analysis of thymocyte apoptosis in vitroThymocytes (2×105 cells/well) were cultivated in 96-well flat-
bottom plates (Nunc A/S) in RPMI 1640 for 18 h. Apoptotic cells wereidentified using FITC-conjugated Annexin V/propidium iodide (PI) kit(Apoptosis detection kit, BD Biosciences Pharmingen). Staining wasperformed according to the producer's manual. Twenty thousand cellsper sample were analyzed using a FACScan flow cytometer (BectonDickinson) and CellQuest software (Becton Dickinson).

921G. Leposavić et al. / Experimental Gerontology 45 (2010) 918–935
2.7. Statistical analysis
Data are reported as means±standard error of the mean (SEM).Statistical significance of age-related differences between young andold rats was evaluated using the Mann–Whitney U test, whilestatistical significance of the differences in urapidil-induced effectsbetween young and old rats was determined by two-way ANOVAfollowed by the Bonferroni test for post hoc comparisons. Allstatistical analyses were performed using GraphPad Prism 5.0software (GraphPad Software, Inc., USA). Differences were consideredto be significant when pb0.05.
3. Results
3.1. First set of experiments: age-associated changes in α1-adrenoceptor-mediated neuro-thymic and thymic intercellular communications
Different indices of neuro-thymic and thymic intercellular com-municationwere examined in young and old rats of aWistar substrainthat exhibits age-associated changes in thymic morphology similar tothat in humans (Plećaš-Solarović et al., 2004; Leposavić et al., 2006b).As earlier reported (Plećaš-Solarović et al., 2004; Leposavić et al.,2006b), despite changes characteristic for thymic involution, absolutethymic weight was not significantly reduced in old rats (433±50 mgin old vs. 590±71 mg in young rats). Namely, in old rats there was adramatic decrease in volume of true thymic lymphoid tissue (Fig. 1A)reflecting, at least partly, diminished thymocyte number (12×107±2.9×107 in old rats vs. 89.8×107±6.7×107 in young rats). The loss of
Fig. 1. Histology of the thymus from young and old Wistar rats. (Panel A) Graph bars show arats. Due to the fixation procedure, shrinking of thymic tissue was approximately 34%. Thephotomicrograph of thymus from a young rat. Note thin septa separating thymic lobi with shaan old rat with enlarged interlobulary space fulfilled with connective/adipose tissue. NoteHaematoxylin and eosin staining, original magnification ×100.
functional thymic tissue was acompanied by a substantial increase inthe amount of connective/adipose tissue (Fig. 1A). In addition,narrowing of the cortex with loss of a clearly discernible cortico-medullary junction was also evident (Fig. 1B).
3.1.1. Thymic noradrenaline levelSince there are intrastrain differences in basal neurochemistry of
Wistar rats (Portelli et al., 2009), to assess the influence of ageing onNA-mediated neuro-thymic and intrathymic cellular communica-tions, NA was measured in young and old rats exhibiting clear signs ofthymic involution. In such involuted thymi, NA levels were greater(pb0.05) than in thymi from young animals (Fig. 2).
3.1.2. Thymic noradrenergic innervationIn order to elucidate mechanisms underlying the increase in
thymic NA level, noradrenergic innervation was analyzed in youngand old rats. As in AO rats (Leposavić et al., 1992), fluorescent profileswere observed in capsule/trabeculae, subcapsulary/subtrabecularyand in cortico-medullary thymic parenchyma of adult Wistar rats,often adjacent to yellow autofluorescent cells. Very rare fluorescentnerve fibers were seen intracortically and in the thymic medulla. Wefailed to notice any significant difference in fluorescent nervedistribution between old and young rats. However, compared withyoung rats, the density of both fluorescent nerve profiles andautofluorescent cells was strikingly increased in old rats (Fig. 3A). Inaddition, the autofluorescent cells exhibited greater fluorescenceintensity in thymi from old rats (Fig. 3A).
bsolute volumes of lymphoid and connective/adipose tissue in thymi of old and youngdata are expressed as mean±SEM (n=7). ***pb0.001. (Panel B) (left) Representativerp cortico-medullary junction. (right) Representative photomicrograph of thymus fromnarrowing of the cortical area with a hardly discernible cortico-medullary junction.
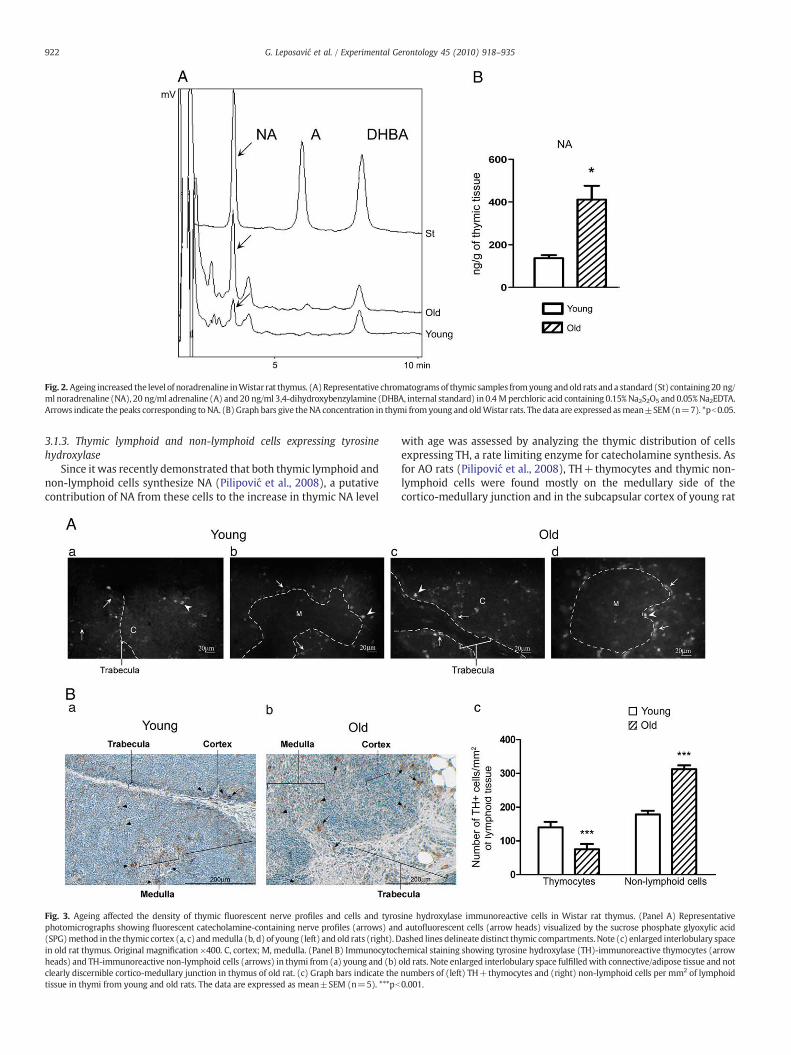
Fig. 2. Ageing increased the level of noradrenaline inWistar rat thymus. (A)Representative chromatogramsof thymic samples fromyoung andold rats anda standard (St) containing20 ng/ml noradrenaline (NA), 20 ng/ml adrenaline (A) and 20 ng/ml 3,4-dihydroxybenzylamine (DHBA, internal standard) in 0.4 Mperchloric acid containing 0.15%Na2S2O5 and0.05%Na2EDTA.Arrows indicate the peaks corresponding toNA. (B)Graph bars give theNA concentration in thymi from young and oldWistar rats. Thedata are expressed asmean±SEM(n=7). *pb0.05.
922 G. Leposavić et al. / Experimental Gerontology 45 (2010) 918–935
3.1.3. Thymic lymphoid and non-lymphoid cells expressing tyrosinehydroxylase
Since it was recently demonstrated that both thymic lymphoid andnon-lymphoid cells synthesize NA (Pilipović et al., 2008), a putativecontribution of NA from these cells to the increase in thymic NA level
Fig. 3. Ageing affected the density of thymic fluorescent nerve profiles and cells and tyrosphotomicrographs showing fluorescent catecholamine-containing nerve profiles (arrows) an(SPG)method in the thymic cortex (a, c) andmedulla (b, d) of young (left) and old rats (right). Din old rat thymus. Original magnification ×400. C, cortex; M, medulla. (Panel B) Immunocytochheads) and TH-immunoreactive non-lymphoid cells (arrows) in thymi from (a) young and (b)clearly discernible cortico-medullary junction in thymus of old rat. (c) Graph bars indicate thetissue in thymi from young and old rats. The data are expressed as mean±SEM (n=5). ***pb
with age was assessed by analyzing the thymic distribution of cellsexpressing TH, a rate limiting enzyme for catecholamine synthesis. Asfor AO rats (Pilipović et al., 2008), TH+thymocytes and thymic non-lymphoid cells were found mostly on the medullary side of thecortico-medullary junction and in the subcapsular cortex of young rat
ine hydroxylase immunoreactive cells in Wistar rat thymus. (Panel A) Representatived autofluorescent cells (arrow heads) visualized by the sucrose phosphate glyoxylic acidashed lines delineate distinct thymic compartments. Note (c) enlarged interlobulary spaceemical staining showing tyrosine hydroxylase (TH)-immunoreactive thymocytes (arrowold rats. Note enlarged interlobulary space fulfilledwith connective/adipose tissue and notnumbers of (left) TH+thymocytes and (right) non-lymphoid cells per mm2 of lymphoid0.001.

923G. Leposavić et al. / Experimental Gerontology 45 (2010) 918–935
thymi. Cortical and medullary thymic parenchyma was only sparselypopulated with these cells (Fig. 3B). Although TH+cells in old ratsshowed a similar distribution, their density quantified in thymicsections and expressed as number of TH+cells per mm2 of lymphoidtissue differed significantly from that in young rats. Namely, in old ratsa decrease (pb0.001) in density of TH+thymocytes and an increase(pb0.001) in that of non-lymphoid cells were found (Fig. 3B).
3.1.4. Tyrosine hydroxylase expression and noradrenaline content inthymocytes
Flow cytometric analysis confirmed that ageing is associated with adecrease in frequency of TH+thymocytes. Namely, in freshly isolatedthymocyte suspensions from old rats the frequency of TH+cells waslower (pb0.05) than that in thymocyte suspensions from young rats(7.03%±1.10%) (Fig. 4A). However, in old rats MFI for TH was increasedby 125%±17.70% (pb0.001) compared to the value for young rats,suggestingamore than two-fold increase inTHexpressionby thymocytes(Fig. 4A). This finding is supported by HPLC analysis of thymocyte NAcontent, which was greater (pb0.05) in old than in young rats (Fig. 4B).
3.1.5. α1-adrenoreceptor expression in thymusImmunocytochemical staining showed that the pattern of distri-
bution of α1-AR+cells did not differ between old and young rats. Asin young rats (Pešić et al., 2009), α1-AR+cells (mainly thymic non-lymphoid cells) were detected predominantly in the subcapsular/subtrabecular cortex and at the cortico-medullary junction in old
Fig. 4. Ageing increased the density of tyrosine hydroxylase expression and noradrenaline corepresents the expression of tyrosine hydroxylase (TH) on thymocytes isolated from (thin lin(broken line) was used to define background staining. Graph bars indicate (b) the percenta(c) mean fluorescence intensity (MFI) for TH as shown by flow cytometric analysis. The data achromatograms of thymocyte extracts from young and old rats superimposed with the chr10 ng/ml adrenaline (A) dissolved in 0.4 M perchloric acid containing 0.15% Na2S2O5 and 0.05NA content in thymocytes from young and old rats. The data are expressed as mean±SEM
Wistar rats, while only rareα1-AR+cells were observed in the thymicmedulla (Fig. 5A). The number of both α1-AR+thymocytes and non-lymphoid cells counted in thymic sections per mm2 of lymphoidtissue was greater (pb0.001) in old rats (Fig. 5A).
In agreement with the previous findings obtained in situ in thymi,an increased proportion of α1-AR+cells (pb0.001) was found infreshly isolated thymocyte suspensions from old rats compared withthe proportion in young rats (11.30%±1.02%) (Fig. 5B). Moreover, inold rats the MFI for α1-AR was greater (87.00%±9.00% increase;pb0.01) than for young rats (Fig. 5B) suggesting an almost two-foldincrease in α1-AR thymocyte surface density.
3.2. Second set of experiments: effects of long-lasting urapidil treatmenton thymocyte differentiation/maturation in young and old rats
To assess the functional relevance of age-associated changes inneuro-thymic and intrathymic cellular communication suggested bythe first set of experiments, the effects of urapidil treatment onthymocyte development were assessed in young and old rats withadvanced thymic involution (Fig. 1).
3.2.1. Thymic weight and thymocyte yieldThe effects of urapidil on absolute and relative thymusweight [age x
treatment interaction, F (1,24)=12.34, pb0.01 for absolute weight;F (1,24)=14.63, pb0.001 for relativeweight], aswell as on absolute andrelative thymocyte yield [age x treatment interaction, F (1,24)=8.08,
ntent in thymocytes of Wistar rats. (Panel A) (a) Overlaid flow cytometry histogram plote) young and (thick line) old rats. The control sample with omitted primary antibodiesge of change in frequency of TH+thymocytes in old relative to young rats (100%) andre expressed asmean±SEM (n=7). *pb0.05, ***pb0.001. (Panel B) (a) Representativeomatogram of the standard sample (St) containing 10 ng/ml noradrenaline (NA) and% Na2EDTA. Arrows indicate the peaks corresponding to NA. (b) Graph bars indicate the(n=7). *pb0.05.

Fig. 5. Influence of ageing on expression of α1-adrenoceptors in Wistar rat thymus. (Panel A) (a) Representative photomicrographs showing immunocytochemical staining ofthymocytes (arrow head) and non-lymphoid cells (arrows) expressing α1-adrenoceptors (AR) in thymi from (left) young and (right) old rats. Note enlarged interlobulary spacefulfilled with connective/adipose tissue and not clearly discernible cortico-medullary junction in thymus of old rat. (b) Graph bars indicate the number of α1-AR+thymocytes andnon-lymphoid cells per mm2 of lymphoid tissue in thymi from young and old rats. The data are expressed as mean±SEM (n=5). ***pb0.001. (Panel B) (a) The overlaid flowcytometry histogram plot represents the expression of α1-AR on thymocytes isolated from (thin line) young and (thick line) old rats. Control cells with omitted primary antibodies(broken lines) were used to define background staining. Graph bars indicate (b) the percentage change in frequency of α1-AR+thymocytes in old relative to young rats (100%) and(c) mean fluorescence intensity (MFI) for α1-AR obtained by flow cytometric analysis. The data are expressed as mean±SEM (n=7). **pb0.01, ***pb0.001.
924 G. Leposavić et al. / Experimental Gerontology 45 (2010) 918–935
pb0.01 for absolute yield; F (1,24)=12.87, pb0.01 for relative yield]were age-dependent. None of these thymic indices were significantlyaffected by blockade of α1-ARs in old rats, while the values of all theseparameters were significantly increased in young rats (Table 1).
3.2.2. Thymocyte proliferationThymocyte proliferation was assessed in vitro in cultures of
thymocytes isolated from urapidil-injected and control rats of bothages, as well as in thymi of these rats in situ.
In the absence of ConA, there was no significant difference in thefrequency of cells in S+G2/M phases of the cell cycle between culturesof thymocytes isolated fromurapidil-administered (8.21%±0.95% in oldand 10.51%±0.83% in young rats) and corresponding control rats
Table 1Thymic weight and thymocyte number in young and old rats subjected to 14-day long urap
Groups Absolute thymusweight (mg)
Relative thymus(mg/100 g BW)
Mean±SEM Mean±SEM
Young Saline 570±31 168±7Urapidil 706±39a* 232±13 a*
Old Saline 397±80 57±9 b***Urapidil 355±50 51±6
(a) Young controls vs urapidil treated rats, (b) young vs old rats; BW, body weight, * pb0.0
(6.06%±0.81% inold and11.54%±0.72% in young rats).However, in thepresence of ConA, the frequency of proliferating cells was greater(pb0.05) in thymocyte cultures from urapidil-injected than in thosefrom age-matched control cultures (Fig. 6A).
In thymic sections, irrespective of treatment, immunostaining ofnuclear cell proliferation-associated Ki-67 antigen, showed a similarpattern of Ki-67+ cell distribution in thymi from young and old rats.Namely, Ki-67+ thymocytes were mainly observed in the outercortex, with only rare Ki-67+ cells in the thymic medulla (Fig. 6B). Inrats of both ages increased the number of Ki-67+ thymocytes permm2 of lymphoid tissue from urapidil-injected rats was counted, butthe effect was more pronounced in old rats [age×treatmentinteraction, F (1,16)=5.402, pb0.05] (Fig. 6B).
idil treatment and in age-matched saline-injected controls.
weight Total number ofthymocytes (×107)
Relative number ofthymocytes (×107)/100 g BW
Mean±SEM Mean±SEM
84.1±5.2 20.6±1.4108.8±5.3 a* 26.2±0.5 a***
8±2.8 b*** 1.7±0.6 b***9.8±1.4 1.4±0.2
5, ***pb0.001.

Fig. 6. Effects of long-lasting urapidil treatment on thymocyte proliferation in young and old rats. (Panel A) Flow cytometric analysis of BrdU/7-AAD staining of thymocytes after 48 hculture in the presence of 2.5 μg/ml Concanavalin A (ConA). (a) Representative dot plots of BrdU/7-AAD staining of thymocytes of (left) control (Controls) and (right) urapidil-treated (Urapidil) young and old rats. (b) Graph bars show the percentages of cells in S and G2/M phases of the cell cycle in thymocyte suspensions from urapidil-treated (Urapidil)and control (Controls) old and young rats. The data are expressed as mean±SEM (n=7).*pb0.05, (Urapidil vs. Controls). (Panel B) (a) Representative photomicrograph of thymusfrom an old control rat showing localization of immunocytochemically stained Ki-67+ cells (arrows) (original magnification ×400). Note enlarged interlobulary space fulfilled withconnective/adipose tissue and not clearly discernible cortico-medullary junction. Dashed lines are used to delineate distinct thymic compartments. (b) Graph bars indicate thenumber of immunostained Ki-67+ cells per mm2 of lymphoid tissue in thymi from urapidil-treated (Urapidil) and control (Controls) old and young rats. The data are expressed asmean±SEM (n=7). ***pb0.001 (Urapidil vs. Controls). Cap, capsule; C, cortex; M, medulla.
925G. Leposavić et al. / Experimental Gerontology 45 (2010) 918–935
3.2.3. Thymocyte apoptosisIn rats of both ages thymocyte apoptosis was assessed in
thymocyte suspensions in vitro, as well as in situ in the thymus.Using Annexin V/PI staining we found that there was no significant
difference in the overall frequency of apoptotic cells between cultures ofthymocytes from urapidil-administered and age-matched control rats(Fig. 7A).Moreover, neither the frequency of cells in early (AnnexinV+/PI−) nor that of cells in advanced apoptosis (Annexin V+/PI+) differedsignificantly between thymocyte cultures from urapidil-treated andage-matched control rats (Fig. 7A).
In thymic sections, irrespective of urapidil treatment, thymocytesshowing membranous Annexin V staining and thymic non-lymphoidcells exhibiting cytoplasmic Annexin V staining, most likely, due tophagocytosis of Annexin V+thymocytes (Pešić et al., 2007)were foundin thymi from rats of both ages. The latter, probably macrophages andthymicnurse cells (Pešić et al., 2007), aswell asAnnexinV+thymocyteswere found throughout the thymic cortex, but mostly at the cortico-medullary junction and subcapsularly/subtrabecularly. Only very rareAnnexin V+cells were noticed in the thymic medulla (Fig. 7B). Theurapidil effect on the number of Annexin V+thymocytes per mm2 oflymphoid tissue was age-dependent [age× treatment interaction,F (1,16)=4.63, pb0.05]. Thus, urapidil diminished (pb0.05) thenumber of Annexin V+thymocytes per mm2 of lymphoid tissue inyoung rats, while it was ineffective in old rats (Fig. 7B). On the otherhand, neither in young nor in old rats urapidil affected the number ofAnnexin V+non-lymphoid cells per mm2 of lymphoid tissue (Fig. 7B).
3.2.4. Distribution of thymocyte subsets delineated by CD4/CD8/TCRαβexpression
As previously shown (Leposavić et al., 2006a), using FCA, twelvethymocyte subsets at distinct developmental stages were distin-guished by plotting CD4 vs. CD8 within each of three thymocytepopulations gated according to TCRαβ surface density, i.e. withinthymocytes with high (TCRαβhigh), low (TCRαβlow) and nondetect-able levels of TCRαβ (TCRαβ-) (Suppl. Fig. 1).
3.2.4.1. TCRαβ− thymocytes. The effect of urapidil on the relativeproportion of CD4+CD8+ double positive (DP) and CD4+CD8−single positive (SP) thymocytes was dependent on age [age× treatmentinteraction, F (1,24)=8.842, pb0.01 for DP; F (1,24)=96.82, pb0.001for CD4+CD8− SP]. Namely, urapidil increased the percentage of DPand CD4+CD8− SP cells in both old (pb0.001) and young rats (pb0.01and pb0.001, respectively), but more strongly in old ones (Fig. 8A). Inaddition, urapidil decreased (pb0.01) the percentage of CD4−CD8+ SPthymocytes but had no effect on the relative number of CD4−CD8−double negative (DN) thymocytes in rats of either age (Fig. 8A).
Urapidil affected the number of DP [age× treatment interaction,F (1,24)=703.3, pb0.001] and CD4+CD8− SP cells [age×treatmentinteraction, F (1,24)=72.02, pb0.001] differently in old comparedwith young rats. The number of DP cells was increased at both agesbut to a greater extent in old rats (pb0.01 and pb0.001, respectively),while the number of CD4+CD8− SP cells was elevated (pb0.01) only

Fig. 7. Effects of long-lastingurapidil treatment on thymocyte apoptosis inyoung andold rats. (PanelA)Flowcytometric analysisofAnnexinV/propidiumiodide (PI) stainingof thymocytesisolated fromurapidil-injected andcontrol young and old rats. (a)Representative dot-plots showingAnnexinV/PI stainingof thymocytes from(left) control (Controls) and (right) urapidil-treated (Urapidil) young and old rats. (b) Graph bars represent thepercentages of apoptotic thymocytes in (lower graph) all phases of apoptosis, (upper left graph) early (AnnexinV+/PI–)and (upper right graph) advanced (AnnexinV+/PI+) apoptosis from urapidil-treated (Urapidil) and control (Controls) old and young rats. The data are expressed asmean±SEM (n=7).(Panel B) (a) Representative photomicrograph of thymus from a control old rat showing localization of FITC-conjugated Annexin V binding cells (original magnification x100). Note non-lymphoid cells (arrows) and thymocytes (arrow heads) showing cytoplasmic and membrane Annexin V staining, respectively. Also, note enlarged interlobulary space fulfilled withconnective/adipose tissue. Insertion: (left) thymic nurse- and (right) macrophage-like cells showing cytoplasmic Annexin V staining of phagocytingmaterial at high powermagnification(original magnification ×1000). Graph bars indicate the number of Annexin V+(b) thymocytes and (c) non-lymphoid macrophage-like cells per mm2 of lymphoid tissue in thymi fromyoung and old rats injected with urapidil (Urapidil) or saline (Controls). The data are expressed as mean±SEM (n=7). *pb0.05 (Urapidil vs. Controls).
926 G. Leposavić et al. / Experimental Gerontology 45 (2010) 918–935
in young rats (Fig. 9A). Urapidil did not affect the number of DN orCD4−CD8+ SP thymocytes at either age (Fig. 9A).
3.2.4.2. TCRαβlow thymocytes. A significant age×treatment interactionfor the relative number of DN [F (1,24)=64.99, pb0.001] and DP[F (1,24)=79.93, pb0.001] thymocytes was found. Following urapidiltreatment, their relative number was reduced in both old (pb0.001)and young rats (pb0.05 and pb0.01, respectively), but to a greaterextent in old ones (Fig. 8B). Two-way ANOVA showed a significanteffect of urapidil on the percentage of CD4+CD8− SP cells in animalsof both ages, but with no significant age× treatment interaction.Namely, irrespective of age, urapidil increased (pb0.05) their relativenumber, but with no effect on the proportion of CD4−CD8+ SPthymocytes (Fig. 8B).
Urapidil influenced the cellularity of DP and CD4+CD8− SPthymocyte subsets differently in old and young rats [for DP, F (1,24)=201.1, pb0.001; for CD4+CD8−, F (1,24)=13.55, pb0.01]. In youngrats, it increased the number of DP and CD4+CD8− SP cells (pb0.01)compared to saline-injected controls, while in old rats the number of DPcells was diminished (pb0.05) (Fig. 9B). However, blockade of α1-ARs
didnot affect thenumberof cells belonging toeither theDNor theCD4−CD8+ SP thymocyte subset (Fig. 9B) at either age.
3.2.4.3. TCRαβhigh thymocytes. In rats of both ages urapidil increased(pb0.001) the percentage of CD4+CD8− SP cells, but to a greaterextent in old rats [for CD4+CD8−, F (1,24)=11.97, pb0.01](Fig. 8C). Following urapidil treatment the relative numbers of DNand CD4−CD8+ SP thymocytes were diminished to a similar extentin old and young rats, while the percentage of DP cells was notaffected (Fig. 8C).
The effects of urapidil on the number of DN cells and both subsetsof SP thymocytes were age-dependent [for DN, F (1,24)=17.26,pb0.001; for CD4+CD8−, F (1,24)=173.00, pb0.001 and for CD4−CD8+, F (1,24)=4.314, pb0.05]. In both young (pb0.01) and old rats(pb0.001) urapidil increased the number of CD4+CD8− SP cells, butmore strongly in old rats. In young animals the number of DN cellswas reduced (pb0.01) compared to the corresponding controls, whilein old rats their number remained unaltered (Fig. 9C). In contrast toyoung rats, where urapidil did not affect the number of CD4−CD8+SP cells, in old rats they were fewer (pb0.05) than in age-matched

927G. Leposavić et al. / Experimental Gerontology 45 (2010) 918–935
controls (Fig. 9C). In both groups of rats urapidil had no significanteffect on the number of DP cells (Fig. 9C).
3.2.5. Thy-1 (CD90) expression on thymocytesGiven that in young rats urapidil affected thymocyte surface
expression of Thy-1 (CD90) (Pešić et al., 2009), which is involved inregulation of TCRαβ signalling and selection thresholds (Hueber et al.,1997), we analyzed MFI for CD90 in the whole thymocyte populationand in TCRαβlow (mainly cortical DP cells undergoing selection) and
Fig. 8. Differential effect of long-lasting urapidil treatment on thymocyte subset distributthymocytes delineated by CD4/CD8 expression and surface density of TCRαβ in young andcytometric analysis. Data are expressed as means±SEM (n=7). *pb0.05; **pb0.01; ***pb0thymocytes with low surface density of TCRαβ; TCRαβhigh , thymocytes with high surface
TCRαβhigh thymocyte subsets (medullary post-selected DP andmature SP cells). Urapidil produced an age-dependent effect onCD90 MFI [age× treatment interaction for CD90 MFI on the wholethymocyte population, F (1,24)=64.22, pb0.001; for CD90 MFI onTCRαβlow thymocytes, F (1,24)=46.74, pb0.001 and for CD90MFI onTCRαβhigh thymocytes, F (1,24)=41.53, pb0.001]. Namely, in youngrats increases (pb0.001) in CD90 MFI were registered in all three cellpopulations after urapidil treatment (Fig. 10A,B), but not in old rats(Fig. 10B).
ion in young and old rats. Graph bars indicate the percentage of distinct subsets ofold rats injected with urapidil (Urapidil) or saline (Controls), as determined by flow.001 (Urapidil vs. Controls). TCRαβ–, thymocytes with undetectable TCRαβ; TCRαβlow,density of TCRαβ.

Fig. 8. (continued)
928 G. Leposavić et al. / Experimental Gerontology 45 (2010) 918–935
3.2.6. Relative and absolute numbers of CD4+CD25+RT6.1− andCD161+TCRαβ+thymocytes
3.2.6.1. CD4+CD25+RT6.1− thymocytes. Since the expression ofRT6.1 is restricted to the final stages of post-thymic T-cell develop-ment (Mojcik et al., 1991), its expression on CD4+CD25+ cells wasanalyzed in order to delineate mature activated CD4+CD25+ cellsre-entering the thymus (Agus et al., 1991) from regulatory cellsmaturing in the organ (Suppl. Fig. 2). Urapidil produced differentialeffects on the percentage [age×treatment interaction, F (1,24)=55.51, pb0.001] and number [age×treatment interaction, F (1,24)=158, pb0.001] of CD4+CD25+RT6.1− thymocytes in young and oldrats. Namely, although the relative and absolute numbers of CD4+CD25+RT6.1− thymocytes were increased in rats of both ages, theeffect was more pronounced in old rats (Fig. 11A).
3.2.6.2. CD161+TCRαβ+thymocytes. Urapidil increased the percent-age of CD161+TCRαβ+(NKT) cells in rats of both ages (pb0.05 andpb0.001, respectively), but more strongly in old rats [age×treatmentinteraction, F (1,24)=127.1, pb0.001] (Fig. 11B). Irrespective of age,the number of these cells was greater (pb0.01) in urapidil-treatedrats than in age-matched controls (Fig. 11B).
3.2.7. Phenotypic profile of T lymphocytes fromperipheral blood and spleenTo answer whether the changes observed in thymopoiesis
translated into the periphery, the proportion of major T-lymphocytepopulations in peripheral blood and spleen was estimated.
3.2.7.1. Peripheral blood. In old rats urapidil increased (pb0.05) thefrequency of TCRαβ+cells over that in corresponding controls, while itwas ineffective in this respect in young rats [age× treatment interaction,F (1,20)=4.29, pb0.05] (Fig. 12A). Following urapidil treatment theproportion of CD4+ cells within the TCRαβ+cell population waselevated in rats of both ages, but to a greater extent in old rats[age×treatment interaction, F (1,20)=4.261, pb0.05] (Fig. 12A). In oldurapidil-treated rats the proportion of CD4+CD8+ cells within the T-cell populationwasalso increased (pb0.001)over that in corresponding
controls, while it remained unaltered in young urapidil-treated animals[age×treatment interaction, F (1,20)=10.69, pb0.01] (Fig. 12A). Thesecells were shown to be partially immunocompetent thymus-derivedcells that undergo post-thymic maturation to become functionallymature CD4+ lymphocytes (Jimenez et al., 2002). The proportion ofCD8+ cells within the T-cell populationwas diminished in old urapidil-treated rats (pb0.001), while it was unaffected by urapidil treatment inyoung rats [age× treatment interaction, F (1,20)=4.29, pb0.05](Fig. 12A). In rats of both ages, urapidil-induced changes in theproportion of T-cell subsets led to increases in the CD4+/CD8+ T-cellratio, but more prominently in old rats [age× treatment interaction,F (1,20)=4.49, pb0.05] (Fig. 12A).
3.2.7.2. Spleen. Urapidil increased the frequency of TCRαβ+cells onlyin old rats [age× treatment interaction, F (1,20)=4.35, pb0.05](Fig. 12B). The proportion of CD4+ cells within TCRαβ+cells waselevated in rats of both ages, but to a greater extent in old ones [age×treatment interaction, F (1,20)=8.239, pb0.01] (Fig. 12B). Inaddition, in old animals urapidil increased (pb0.001) the proportionof CD4+CD8+ cells within the TCRαβ+subset [age×treatmentinteraction, F (1,20)=15.55, pb0.001] but not in young rats(Fig. 12B). Moreover, in urapidil-treated old rats the proportion ofCD8+ cells within the T-cell population was reduced (pb0.05), butthere was no affect of urapidil treatment in young rats [age×treatment interaction, F (1,20)=3.92, pb0.05] (Fig. 12B). In rats ofboth ages, urapidil increased the CD4+/CD8+ T-cell ratio, but to agreater extent in old rats [age× treatment interaction, F (1,20)=4.18,pb0.05] (Fig. 12B).
4. Discussion
The first part of our study demonstrating an age-associatedincrease in thymic NA availability and thymocyte surface density ofα1-ARs suggests altered NA neuro-thymic and thymic intercellularcommunications via α1-ARs at advanced ages. The second partconfirmed the age-associated plasticity of intrathymic communica-tions involving α1-ARs by showing differential effects of urapidil on

929G. Leposavić et al. / Experimental Gerontology 45 (2010) 918–935
thymopoiesis in old and young animals. Namely, although in rats ofboth ages urapidil increased the proportions of thymic CD4+CD8−cells, including CD4+CD25+RT6.1− cells, as well as NKT cells, thesechanges were more pronounced in old rats. In addition, an increasedproportion of phenotypically immature CD4+CD8+ T lymphocytesescaped from the thymus into periphery in old rats.
4.1. Age-associated changes in α1-adrenoceptor-mediated noradrenalineneuro-thymic and thymic intercellular communications
4.1.1. Neuro-thymic communicationsAn age-associated increase in NA level was found in thymi from
the substrain of Wistar rats used in our experiments, as in another
Fig. 9. Age-dependent effect of long-lasting urapidil treatment on number of cells within thymthe overall number of cells in distinct subsets of thymocytes delineated by CD4/CD8 expressor saline (Controls) as determined by flow cytometric analysis. Data are reported as mthymocytes with undetectable TCRαβ; TCRαβlow, thymocytes with low surface density of T
substrain (Cavallotti et al., 1999). The rise in thymic NA level with ageoccurred concomitantly with an increase in density of autofluorescentnerve fibers (Madden et al., 1997; Cavallotti et al., 1999). Sincenoradrenergic nerve fibers are the main source of NA in the thymus(Elenkov et al., 2000), the striking rise in NA level in thymi from oldrats mainly reflects increased density of these fibers.
However, besides the rise in noradrenergic nerve density and NAlevel in thymi from old rats, the density of lymphoid and non-lymphoidtarget cells expressing α1-ARs was also increased. Cells expressing α1-AR have been detected across all thymocyte subsets, but mainly withinlessmature CD3− andCD3low subsets (Pešić et al., 2009). In thymus,α1-AR immunoreactivity has also been observed in subsets of thymicepithelial and CD68+ cells (most likely macrophages) located mainly
ocyte subsets delineated according to CD4/CD8/TCRαβ expression. Graph bars indicateion and surface density of TCRαβ in young and old rats injected with urapidil (Urapidil)eans±SEM (n=7). *pb0.05; **pb0.01; ***pb0.001 (Urapidil vs. Controls). TCRαβ-,CRαβ; TCRαβhigh , thymocytes with high surface density of TCRαβ.

Fig. 9. (continued)
930 G. Leposavić et al. / Experimental Gerontology 45 (2010) 918–935
subcapsularly/subtrabecularly and at the cortico-medullary junction(Pešić et al., 2009). In old rats, apart from augmented frequency of α1-AR+thymocytes, a substantial increase in α1-AR surface density wasmeasured. Thus, an increased sensitivity of thymocytes from old rats toα1-AR-mediated signalling may be assumed. The previous finding iscorroborated by data indicating that α1-AR mRNA expression, andconsequently the receptor surface density, vary (increase, decrease orremain unchanged) with age in a tissue- and species-specific manner(Yokoyama et al., 1984; Rudner et al., 1999; Michelottia et al., 2000;Basso and Piantanelli, 2002). Collectively, the previous findingsindicated altered neuro-thymic communication in old compared withyoung rats and pose the question how far these changes are the cause oreffect of thymic involution, which will be subject of further studies.
4.1.2. Thymic intercellular communicationsIn addition to increaseddensity of noradrenergicfibers in thymi from
old rats, a rise indensity of TH-containingnon-lymphoid cellswas found.These cells belong to subsets ofmacrophages and thymic epithelial cells,including thymic nurse cells (Pilipović et al., 2008). The age-related risein density of non-lymphoid TH+cells was consistent with the increasein density of autofluorescent cells, whichmay bemast cells (Artico et al.,2002),macrophages (Maddenet al., 1997; Bellinger et al., 2008; Pilipovićet al., 2008) and thymic epithelial cells (Pilipović et al., 2008). In distinctsubsets of these cells α1-AR expression was observed, indicatingautocrine as well as paracrine action of NA in the thymus (Pešić et al.,2009). The increased density of non-lymphoid TH+cells in old thymimost likely reflected theunproportional loss of distinct subsets of thymicnon-lymphoid cells with age (reviewed in Leposavić and Perišić, 2008).The augmented intensity of SPG fluorescence evoked in non-lymphoidcells of old rats might be ascribed to an increase in their NA content.Thus, age-associated changes involvingTH+non-lymphoid thymic cellsmay have contributed to the increase in thymic NA level in old rats.Moreover, although frequency of TH+thymocytes was diminished,both increased expression of TH andmore NA in TH+thymocytes fromold rats were found. To envisage the possible functional implications ofall this, it should be mentioned that measurable levels of TH have beenfoundonly inmoremature subsets of thymocytes (Pilipović et al., 2008).
These findings together with previous data indicating an increasedsensitivity of thymocytes from old rats to α1-AR-mediated signalling
suggest that, besides neuro-thymic, NA-mediated intercellular thymiccommunications are altered in old rats.
4.2. Differential effects of long-lasting urapidil treatment on thymocytedevelopment in young and old rats
Further support for altered NA-mediated communications in oldthymi comes from our demonstration of a differential effect of urapidilon thymocyte number and differentiation/maturation pattern inyoung and old rats. In rats of both ages long-lasting urapidil treatmentincreased thymocyte number, but this increase reached statisticalsignificance only in young rats. Generally, an increase in thymocytenumber may be evoked by: i) increased entry of progenitor cells;ii) augmented proliferation of thymocytes; iii) decreased thymocyteapoptosis and iv) diminished egress ofmature cells into the periphery.Urapidil augmented thymocyte proliferation in response to ConA inthymocyte cultures, and increased the density of Ki-67+ cells inthymi from rats of both ages, but to a greater extent in old rats. On theother hand, urapidil augmented escape of immature CD4+CD8+thymocytes only in old rats, being ineffective in this respect in youngones. Collectively, all these findings may explain the differential effectof urapidil on thymocyte yield in rats of different age.
There are several explanations for the differential effect ofurapidil on thymocyte proliferation in young and old rats. Assimultaneous administration of NA and propranolol decreased thenumber of Ki-67+ cells per square mm in T-cell areas of rat spleen(Stevenson et al., 2001), a direct stimulatory influence of urapidil onthymocyte proliferation may be assumed. However, urapidil mayincrease thymocyte proliferation also indirectly, via central sup-pressive action on sympathetic flow (Sanders and Jurna, 1985;Anger et al., 1988; Castor and Schmidt, 1994; Pešić et al., 2009) andconsequently on β-AR-mediated inhibition of thymocyte prolifer-ation (Ray and Cramer, 1979; Leposavić et al., 2006a). Indeed, inyoung rats the increase in thymocyte proliferation followingurapidil treatment has been partly related to diminished β-AR-mediated thymic sympathetic tone (Pešić et al., 2009). However,the lack of increase in Thy-1 (CD90) thymocyte surface densityfollowing urapidil treatment in old rats, which in young rats wasreported and clearly ascribed to diminished β-AR-mediated thymicsympathetic tone (Pešić et al., 2009), speaks against significant
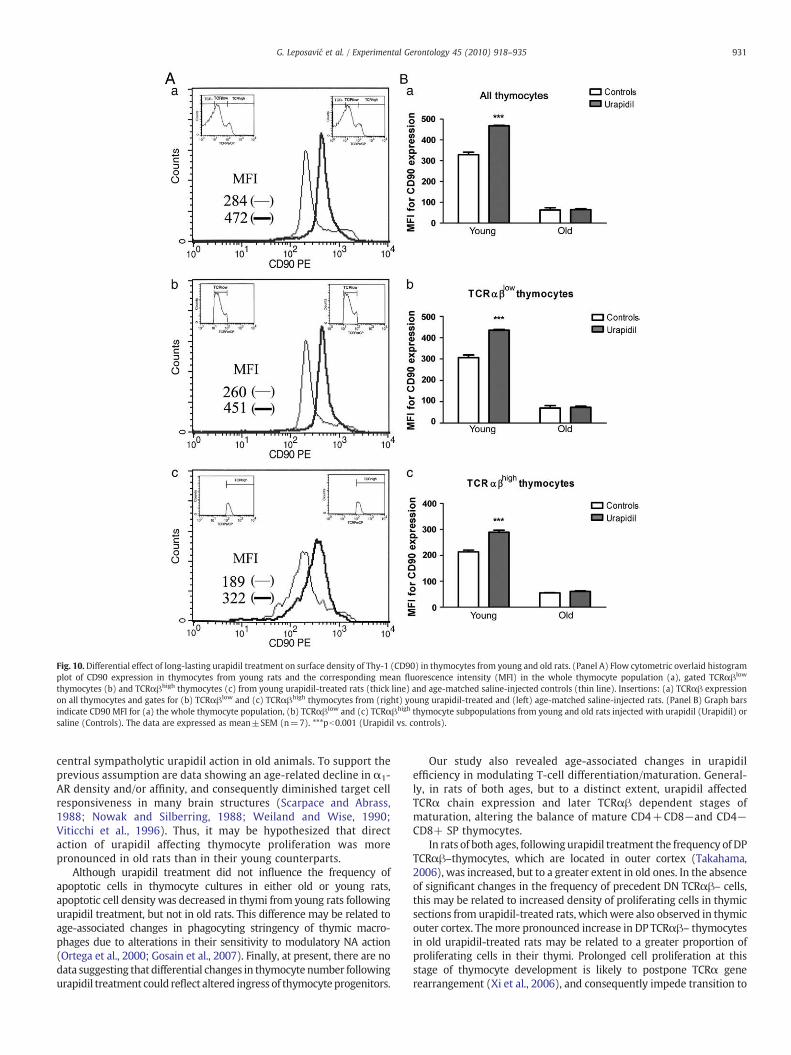
Fig. 10. Differential effect of long-lasting urapidil treatment on surface density of Thy-1 (CD90) in thymocytes from young and old rats. (Panel A) Flow cytometric overlaid histogramplot of CD90 expression in thymocytes from young rats and the corresponding mean fluorescence intensity (MFI) in the whole thymocyte population (a), gated TCRαβlow
thymocytes (b) and TCRαβhigh thymocytes (c) from young urapidil-treated rats (thick line) and age-matched saline-injected controls (thin line). Insertions: (a) TCRαβ expressionon all thymocytes and gates for (b) TCRαβlow and (c) TCRαβhigh thymocytes from (right) young urapidil-treated and (left) age-matched saline-injected rats. (Panel B) Graph barsindicate CD90 MFI for (a) the whole thymocyte population, (b) TCRαβlow and (c) TCRαβhigh thymocyte subpopulations from young and old rats injected with urapidil (Urapidil) orsaline (Controls). The data are expressed as mean±SEM (n=7). ***pb0.001 (Urapidil vs. controls).
931G. Leposavić et al. / Experimental Gerontology 45 (2010) 918–935
central sympatholytic urapidil action in old animals. To support theprevious assumption are data showing an age-related decline in α1-AR density and/or affinity, and consequently diminished target cellresponsiveness in many brain structures (Scarpace and Abrass,1988; Nowak and Silberring, 1988; Weiland and Wise, 1990;Viticchi et al., 1996). Thus, it may be hypothesized that directaction of urapidil affecting thymocyte proliferation was morepronounced in old rats than in their young counterparts.
Although urapidil treatment did not influence the frequency ofapoptotic cells in thymocyte cultures in either old or young rats,apoptotic cell density was decreased in thymi from young rats followingurapidil treatment, but not in old rats. This difference may be related toage-associated changes in phagocyting stringency of thymic macro-phages due to alterations in their sensitivity to modulatory NA action(Ortega et al., 2000; Gosain et al., 2007). Finally, at present, there are nodata suggesting that differential changes in thymocyte number followingurapidil treatment could reflect altered ingress of thymocyteprogenitors.
Our study also revealed age-associated changes in urapidilefficiency in modulating T-cell differentiation/maturation. General-ly, in rats of both ages, but to a distinct extent, urapidil affectedTCRα chain expression and later TCRαβ dependent stages ofmaturation, altering the balance of mature CD4+CD8−and CD4−CD8+ SP thymocytes.
In rats of both ages, following urapidil treatment the frequency of DPTCRαβ–thymocytes, which are located in outer cortex (Takahama,2006), was increased, but to a greater extent in old ones. In the absenceof significant changes in the frequency of precedent DN TCRαβ– cells,this may be related to increased density of proliferating cells in thymicsections from urapidil-treated rats, whichwere also observed in thymicouter cortex. Themore pronounced increase in DP TCRαβ– thymocytesin old urapidil-treated rats may be related to a greater proportion ofproliferating cells in their thymi. Prolonged cell proliferation at thisstage of thymocyte development is likely to postpone TCRα generearrangement (Xi et al., 2006), and consequently impede transition to

Fig. 11. Long-lasting urapidil treatment affected the percentage and number of CD4+CD25+RT6.1– and CD161+TCRαβ+NKT cells in thymi from young and old rats. (Panel A)Flow cytometric analysis of CD4+CD25+RT6.1– cells in thymi of young and old rats. Graph bars represent (left) the percentage and (right) the number of CD4+CD25+RT6.1–cells in thymi from young and old rats injected with urapidil (Urapidil) or saline (Controls), determined by flow cytometric analysis. The data are expressed as mean±SEM (n=7).**pb0.01; ***pb0.001 (Urapidil vs. Control). (Panel B) Flow cytometric analysis of CD161+TCRαβ+NKT cells in thymi of young and old rats. (a) Representative dot plots of CD161/TCRαβ expression in (left) saline-injected (Controls) and (right) urapidil-treated (Urapidil) young and old rats. (b) Graph bars represent (upper) the percentage and (lower) thenumber of CD161+TCRαβ+cells in thymi from young and old rats injected with urapidil (Urapidil) or saline (Controls), determined by flow cytometry analysis. The data areexpressed as mean±SEM (n=7). *pb0.05; **pb0.01; ***pb0.001 (Urapidil vs. Controls).
932 G. Leposavić et al. / Experimental Gerontology 45 (2010) 918–935
the next DP TCRαβlow developmental stage. Indeed, following urapidiltreatment the frequency of DP TCRαβlow cells entering positiveselection was lower in rats of both ages (Zamoyska and Lovatt, 2004;Miosge and Zamoyska, 2007), but this reduction wasmore pronouncedin old rats. In young rats, therewas anunaltered frequency ofDPTCRhigh
thymocytes that had just passed positive selection, being intermediarycells between the DP TCRαβlow and SP TCRαβhigh stages (Shortmanet al., 1991). In conjunctionwith increased thymocyte surface density ofThy-1, molecule that is implicated in negative regulation of TCRαβsignalling and thymocyte selection threshold (Hueber et al., 1997), thissuggests more efficient positive/reduced negative selection providingcompensation for the developmental block at the previous DP TCRαβ–developmental stage (Pešić et al., 2009). On the other hand, in old rats,the unaltered percentage of DP TCRαβhigh thymocytes in the absence ofsignificant alterations in Thy-1 thymocyte surface density suggests Thy-1 independent changes in thymocyte selection. In other words, the lackof significant central urapidil action in old rats mirrored unalteredthymocyte Thy-1 surface density, and pointed to urapidil capability, at
least in old rats, to affect the efficiency of thymocyte selection via directthymic action. This is fully consistent with the suggestion of Bellingeret al. (2008) that catecholamines acting viaα1-ARs on thymic epithelialcells essential to the thymocyte selection processes may affectthymocyte development. Considering that urapidil influences thymo-cyte selection in rats of both ages, the universal lack of substantialchanges in apoptosis following urapidil treatment may be because thefrequency of apoptotic cells was estimated within an unfractionatedthymocyte population. Namely, changes within selected thymocytesubpopulation/smay beminimized or even completely counteracted bychanges in the opposite direction within other thymocyte subpopula-tion/s, and so underestimated or even completely overlooked when anunfractionated population is analyzed. Moreover, we found that in ratsof both ages urapidil influenced lineage commitment causing anincrease in the frequency of CD4+CD8–TCRαβhigh, and decrease inthat of CD4−CD8+TCRαβhigh cells, which was much stronger in oldrats. Thisfinding provides further support for age-associated changes inα1-AR-mediated intrathymic NA communications.

Fig. 12. Long-lastingurapidil treatment affected thepercentage of cells delineated by expression of CD4/CD8molecules on TCRαβ+cells in peripheral blood and spleens fromyoung andold rats. (Panel A) Flow cytometric analysis of peripheral bloodlymphocytes delineated by expression of CD4/CD8/TCRαβ. Representative dot plots of CD4/CD8 expression on TCRαβ+lymphocytes from (a) young and (b) old rats gated as shown in the upper single parameter histograms of TCRαβ expression.Boundaries represent cut-offs of positive signal for themarker based on isotype control staining. Graphbars give thepercentage of (c) TCRαβ+cells, (d) CD4+cells, (e) CD8+cells, (f) CD4+CD8+cellswithin TCRαβ+lymphocytes and (g) the ratioof CD4+/CD8+ cells in urapidil-treated (Urapidil) and saline-injected (Controls) young and old animals. The data are expressed asmean±SEM (n=7). *pb0.05; ***pb0.001 (Urapidil vs. controls). (Panel B) Flow cytometric analysis of splenocytesdelineated by expression of CD4/CD8/TCRαβ. Representative dot plots of CD4/CD8 expression on TCRαβ+splenocytes from (a) young and (b) old rats gated as shown in the upper single parameter histograms of TCRαβ expression. Boundariesrepresent cut-offs of positive signal for themarker based on isotype control staining. Graph bars give the percentage of (c) TCRαβ+cells, (d) CD4+ cells, (e) CD8+ cells, (f) CD4+CD8+ cells within TCRαβ+splenocytes and (g) the ratio of CD4+/CD8+ cells in urapidil-treated (Urapidil) and saline-injected (Controls) young and old animals.The data are expressed as mean±SEM (n=7). *pb0.05; ***pb0.001 (Urapidil vs. Controls).
933G.Leposavić
etal./
Experimental
Gerontology
45(2010)
918–935

934 G. Leposavić et al. / Experimental Gerontology 45 (2010) 918–935
In addition, urapidil increased the frequencyof CD4+CD25+RT6.1–and NKT cells in thymi from rats of both ages, but to a greater extent inold rats. These data contradict our previous hypothesis based on thefinding that urapidil increases the frequency of these cells in young ratthymus via a central sympatholytic action (Pešić et al., 2009). In otherwords, the data reported herein clearly suggest that urapidil, acting atthe thymic level, may influence the maturation of regulatory T-cells,whose production decreases with advanced ages (Leposavić et al.,2006b). This warrants further investigations to understand the putativerelevance of urapidil treatment to susceptibility to autoimmune diseasedevelopment in old rats.
To assess how the alterations observed in thymopoiesis followingurapidil treatment translated to the periphery, the proportions of majorT-lymphocyte populations were estimated in peripheral blood andspleen. Differently from young rats, urapidil slightly, but significantly,increased the frequency of TCRαβ+cells in both peripheral blood andspleen from old rats. This was reflected by elevated levels of CD4+ andCD4+CD8+ cells, which are partially immunocompetent thymus-derived cells that undergo post-thymic maturation to become function-ally mature CD4+ lymphocytes in rats (Jimenez et al., 2002). Thisfinding seems to be particularly important considering the age-associated decline in CD4+ lymphocyte abundance in the periphery(De Paoli et al., 1988; Gilman-Sachs et al., 1991; Pahlavani andRichardson, 1994; Clambey et al., 2007; Czesnikiewicz-Guzik et al.,2008). Furthermore, in peripheral blood and spleen from old ratsurapidil diminished the proportion of CD8+ cells within the T-cellpopulation, raising theCD4+/CD8+T-cell ratio. In peripheral blood andspleen from young rats, urapidil evoked only a slight increase in theproportion of CD4+cellswithin the TCRαβ+population also leading toan elevated CD4+/CD8+ T-cell ratio. The functional implications ofthese changes should be also the subject of further investigations.
In conclusion, our study clearly demonstrated an age-associatedincrease in thymic NA level (due to alterations in both the thymicneural and non-neural noradrenergic network) and α1-AR thymocytesurface density. Furthermore, although this study did not answer howfar these changes are the cause or effect of thymic involution, it clearlyshowed that the α1-AR-mediated NA influence on T-cell differenti-ation/maturation is altered in the involuted thymus. Moreover, itpointed to the intrinsic potential of thymi in old rats, which might beemployed to modulate the quantity and quality of thymic output.
Supplementarymaterials related to this article can be found onlineat doi:10.1016/j.exger.2010.08.011.
Acknowledgement
This work was supported by grant 145049 from the Ministry ofScience and Technology of the Republic of Serbia.
References
Agus, D.B., Surh, C.D., Sprent, J., 1991. Re-entry of T cells to the adult thymus is restrictedto activated T cells. J. Exp. Med. 173, 1039–1046.
Anger, C., van Aken, H., Feldhaus, P., Wusten, R., Konig, H., Krahling, K.H., Lawin, P.,1988. Permeation of the blood-brain barrier by urapidil and its influence onintracranial pressure in man in the presence of compromised intracranialdynamics. J. Hypertens. Suppl. 6, S63–S64.
Artico, M., Cavallotti, C., Cavallotti, D., 2002. Adrenergic nerve fibres and mast cells:correlation in rat thymus. Immunol. Lett. 84, 69–76.
Aspinall, R., Andrew, D., 2000. Thymic involution in aging. J. Clin. Immunol. 20, 250–256.Aw, D., Silva, A.B., Palmer, D.B., 2007. Immunosenescence: emerging challenges for an
ageing population. Immunology 120, 435–446.Basso, A., Piantanelli, L., 2002. Influence of age on circadian rhythms of adrenoceptors in
brain cortex, heart and submandibular glands of BALB/c mice: when circadianstudies are not only useful but necessary. Exp. Gerontol. 37, 1441–1450.
Bellinger, D.L., Felten, S.Y., Felten, D.L., 1988. Maintenance of noradrenergic sympatheticinnervation in the involuted thymus of the aged Fischer 344 rat. Brain Behav.Immun. 2, 133–150.
Bellinger, D.L., Millar, B.A., Perez, S., Carter, J., Wood, C., ThyagaRajan, S., Molinaro, C.,Lubahn, C., Lorton, D., 2008. Sympathetic modulation of immunity: relevance todisease. Cell. Immunol. 252, 27–56.
Berzins, S.P., Uldrich, A.P., Sutherland, J.S., Gill, J., Miller, J.F.A.P., Godfrey, D.I., Boyd, R.L.,2002. Thymic regeneration: teaching an old immune system new tricks. TrendsMol. Med. 8, 469–476.
Castor, G., Schmidt, U., 1994. Urapidil permeates the intact blood–brain barrier.Intensive Care Med. 20, 278–281.
Cavallotti, C., Artico, M., Cavallotti, D., 1999. Occurrence of adrenergic nerve fibers andof noradrenaline in thymus gland of juvenile and aged rats. Immunol. Lett. 70,53–62.
Clambey, E.T., Kappler, J.W., Marrack, P., 2007. CD8 T cell clonal expansions and aging: aheterogeneous phenomenon with a common outcome. Exp. Gerontol. 42, 407–411.
Czesnikiewicz-Guzik, M., Lee, W.-W., Cui, D., Hiruma, Y., Lamar, D.L., Yang, Z.-Z.,Ouslander, J.G., Weyand, C.M., Goronzy, J.J., 2008. T cell subset-specific suscepti-bility to aging. Clin. Immunol. 127, 1107–1111.
De Paoli, P., Battistin, S., Santini, G.F., 1988. Age-related changes in human lymphocytesubsets: progressive reduction of CD4CD45R population. Clin. Immunol. Immuno-pathol. 48, 290–296.
Docherty, J.R., 2002. Age-related changes in adrenergic neuroeffector transmission.Autonomic Neurosci.: Basic Clin. 96, 8–12.
Elenkov, I.J., Wilder, R.L., Chrousos, G.P., Vizi, E.S., 2000. The sympathetic nerve — anintegrative interface between two supersystems: the brain and the immunesystem. Pharmacol. Rev. 52, 595–638.
Fabris, N., Mocchegiani, M., Provinciali, M., 1997. Plasticity of neuroendocrine-thymusinteractions during aging. Exp. Gerontol. 32, 415–429.
Gilman-Sachs, A., Kin,Y.B., Pollard,M., Snyder, D.L., 1991. Influence of aging, environmentalantigens and dietary restriction on expression of lymphocyte subsets in germ-free andconventional Lobund–Wistar rats. J. Gerontol. 46, B101–B106.
Globerson, A., 2002. Developmental aspects of the thymus in aging. Adv. Cell AgingGerontol. 13, 47–78.
Gosain, A., Muthu, K., Gamelli, R.L., DiPietro, L.A., 2007. Norepinephrine suppresseswound macrophage phagocytic efficiency through alpha- and beta-adrenoreceptordependent pathways. Surgery 142, 170–179.
Gruver, A.L., Hudson, L.L., Sempowski, G.D., 2007. Immunosenescence of ageing. J.Pathol. 211, 144–156.
Hirokawa, K., Utsuyama, M., Kasai, M., Kurashima, C., Ishijima, S., Zeng, Y.X., 1994.Understanding the mechanism of age-changes of thymic function to promote T celldifferentiation. Immunol. Lett. 40, 267–277.
Hsu, H.C., Li, L., Zhang, H.G., Mountz, J.D., 2005. Genetic regulation of thymic involution.Mech. Ageing Dev. 126, 87–97.
Hueber, A.O., Bernard, A.M., Battari, C.L., Marguet, D., Massol, P., Foa, C., Brun, N., Garcia,S., Stewart, C., Pierres, M., He, H.T., 1997. Thymocytes in Thy-1-/- mice showaugmented TCR signaling and impaired differentiation. Curr. Biol. 7, 705–708.
Jimenez, E., Sacedon, R., Vicente, A., Hernandez-Lopez, C., Zapata, A.G., Varas, A., 2002.Rat peripheral CD4(+)CD8(+) T lymphocytes are partially immunocompetentthymus-derived cells that undergo post-thymic maturation to become functionallymature CD4(+) T lymphocytes. J. Immunol. 168, 5005–5013.
Kamath, A.B., Nagarkatti, P.S., Nagarkatti, M., 1998. Characterization of phenotypicalterations induced by 2, 3, 7, 8-tetrachlorodibenzo-p-dioxin on thymocytes in vivoand its effect on apoptosis. Toxicol. Appl. Pharmacol. 150, 117–124.
Kavelaars, A., 2002. Regulated expression of alpha-1 adrenergic receptors in theimmune system. Brain Behav. Immun. 16, 799–807.
Kelley, K.W., Brief, S., Westly, H.J., Novakofski, J., Bechtel, P.J., Simon, J., Walker, E.B.,1986. GH3 pituitary adenoma cells can reverse thymic aging in rats. Proc. Natl Acad.Sci. USA 83, 5663–5667.
Lau, L., Spain, L.M., 2000. Altered ageing-related thymic involution in T cell receptortransgenic, MHC-deficient, and CD4-deficient mice. Mech. Ageing Dev. 114,101–121.
Leposavić, G., Perišić, M., 2008. Age-associated remodeling of thymopoiesis: role forgonadal hormones and catecholamines. Neuroimmunomodulation 15, 290–322.
Leposavić, G., Mićić, M., Ugrešić, N., Bogojević, M., Isaković, K., 1992. Components ofsympathetic innervation of the rat thymus during late fetal and postnataldevelopment: histofluorescence and biochemical study. Sympathetic innervationof the rat thymus. Thymus 19, 77–87.
Leposavić, G., Arsenović-Ranin, N., Radojević, K., Kosec, D., Pešić, V., Vidić-Danković, B.,Plećaš-Solarović, B., Pilipović, I., 2006a. Characterization of thymocyte phenotypicalterations induced by long-lasting beta-adrenoceptor blockade in vivo and itseffects on thymocyte proliferation and apoptosis. Mol. Cell. Biochem. 285, 87–99.
Leposavić, G., Pešić, V., Kosec, D., Radojević, K., Arsenović-Ranin, N., Pilipović, I., Perišić,M., Plećas-Solarović, B., 2006b. Age-associated changes in CD90 expression onthymocytes and in TCR-dependent stages of thymocyte maturation in male rats.Exp. Gerontol. 41, 574–589.
Leposavić, G., Pilipović, I., Radojević, K., Pešić, V., Perišić, M., Kosec, D., 2008.Catecholamines as immunomodulators: a role for adrenoceptor-mediatedmechan-isms in fine tuning of T-cell development. Auton. Neurosci. 144, 1–12.
Li, Y.M., Brunke, D.L., Dantzer, R., Kelley, K., 1992. Pituitary epithelial cell implantsreverse the accumulation of CD4–CD8-lymphocytes in thymus glands of aged rats.Endocrinology 130, 2703–2709.
Loveland, B.E., Jarrott, B., McKenzie, I.F.C., 1981. The detection of β-adrenoceptors onmurine lymphocytes. Int. J. Immunopharmacol. 3, 313–318.
Madden, K.S., Felten, D., 2001. β-adrenoceptor blockade alters thymocyte differenti-ation in aged mice. Cell. Mol. Biol. 47, 189–196.
Madden, K.S., Bellinger, D.L., Felten, S.Y., Snyder, E., Maida, M.E., Felten, D.L., 1997.Alterations in sympathetic innervation of thymus and spleen in aged mice. Mech.Ageing Dev. 94, 165–175.
Marchetti, B., Morale, M.C., Paradis, P., Bouvier, M., 1994. Characterization, expression,and hormonal control of a thymic beta 2-adrenergic receptor. Am. J. Physiol. 267,E718–E731.

935G. Leposavić et al. / Experimental Gerontology 45 (2010) 918–935
Michelottia, G.A., Priceb, D.T., Schwinna, D.A., 2000. Associate editor: D.R. Sibley. a1-Adrenergic receptor regulation: basic science and clinical implications. Pharmacol.Ther. 88, 281–309.
Mićić, M., Leposavić, G., Ugrešić, N., 1994. Relationships between monoaminergic andcholinergic innervation of the rat thymusduring aging. J. Neuroimmunol. 49, 205–212.
Miosge, L., Zamoyska, R., 2007. Signalling in T-cell development: is it all location,location, location? Curr. Opin. Immunol. 19, 194–199.
Mojcik, C.F., Greiner, D.L., Goldschneider, I., 1991. Characterization of RT6-bearing ratlymphocytes. II. Developmental relationships of RT6- and RT6+ T cells. Dev.Immunol. 1, 191–201.
Nowak, G., Silberring, J., 1988. Kinetics of [3H]-prazosin binding to the rat cortex duringaging. Pharmacol. Biochem. Behav. 31, 505–507.
Ortega, E., Garcia, J.J., Saez, M.C., De la Fuente, M., 2000. Changes with aging in themodulation of macrophages by norepinephrine. Mol. Cell. Biochem. 203, 113–117.
Pahlavani, M.A., Richardson, A., 1994. Age-related decrease in the naive (OX22+) Tcells in F344 rats. Mech. Ageing Dev. 74, 171–176.
Pawelec, G., Akbar, A., Caruso, C., 2005. Human immunosenescence: is it infectious?Immunol. Rev. 205, 257–268.
Pešić, V., Plećaš-Solarović, B., Radojević, K., Kosec, D., Pilipović, I., Perišić, M., Leposavić,G., 2007. Long-term beta-adrenergic receptor blockade increases levels of the mostmature thymocyte subsets in aged rats. Int. Immunopharmacol. 7, 674–686.
Pešić, V., Kosec, D., Radojević, K., Pilipović, I., Perišić, M., Vidić-Danković, B., Leposavić,G., 2009. Expression of alpha(1)-adrenoceptors on thymic cells and their role infine tuning of thymopoiesis. J. Neuroimmunol. 214, 55–66.
Pilipović, I., Vidić-Danković, B., Perišić, M., Radojevi, K., Čolić, M., Todorović, V.,Leposavić, G., 2008. Sexual dimorphism in the catecholamine-containing thymusmicroenvironment: a role for gonadal hormones. J. Neuroimmunol. 195, 7–20.
Plećaš-Solarović, B., Lalić, Lj., Leposavić, G., 2004. Age-dependent morphometricalchanges in the thymus of male propranolol-treated rats. Ann. Anat. 186, 141–147.
Plećaš-Soloravić, B., Hristić-Živković, I., Radojević, K., Kosec, D., Leposavić, G., 2005.Chronic alpha(1)-adrenoreceptor blockade produces age-dependent changes in ratthymus structure and thymocyte differentiation. Histol. Histopathol. 20, 833–841.
Portelli, J., Aourz, N., De Bundel, D., Meurs, A., Smolders, I., Michotte, Y., Clinckers, R.,2009. Intrastrain differences in seizure susceptibility, pharmacological responseand basal neurochemistry of Wistar rats. Epilepsy Res. 87, 234–246.
Ray, D.E., Cramer, J.E., 1979. The action of decamethrin (a synthetic pyrethroid) on therat. Pestic. Biochem. Physiol. 10, 330–340.
Rudner, X.L., Berkowitz, D.E., Booth, J.V., Funk, B.L., Cozart, K.L., D'Amico, E.B., El-Moalem, H., Page, S.O., Richardson, C.D., Winters, B., Marucci, L., Schwinn, D.A.,1999. Subtype specific regulation of human vascular alpha(1)-adrenergic receptorsby vessel bed and age. Circulation 100, 2336–2343.
Sanders, K.H., Jurna, I., 1985. Effects of urapidil, clonidine, prazosin and propranolol onautonomic nerve activity, blood pressure and heart rate in anaesthetized rats andcats. Eur. J. Pharmacol. 110, 181–190.
Scarpace, P.J., Abrass, I.B., 1988. Alpha- and beta-adrenergic receptor function in thebrain during senescence. Neurobiol. Aging 9, 53–58.
Shortman, K., Vremec, D., Egerton, M.J., 1991. The kinetics of T cell antigen receptorexpression by subgroups of CD4+8+ thymocytes: delineation of CD4+8+32+thymocytes as post-selection intermediates leading to mature T cells. J. Exp. Med.173, 323–332.
Stevenson, J.R., Westermann, J., Liebmann, P.M., Hörtner, M., Rinner, I., Felsner, P.,Wölfler, A., Schauenstein, K., 2001. Prolonged alpha-adrenergic stimulation causeschanges in leukocyte distribution and lymphocyte apoptosis in the rat. J.Neuroimmunol. 120, 50–57.
Takahama, Y., 2006. Journey through the thymus: stromal guides for T-celldevelopment and selection. Nat. Rev. Immunol. 6, 127–135.
Viticchi, C., Piantanelli, L., Rossanoa, F., Spector, N.H., 1996. Age-related regulation ofmouse brain cortex adrenoceptors following camphor vapor exposition. Arch.Gerontol. Geriatrics 23, 39–45.
Weiland, N.G., Wise, P.M., 1990. Aging progressively decreases the densities and altersthe diurnal rhythms of alpha 1-adrenergic receptors in selected hypothalamicregions. Endocrinology 126, 2392–2397.
Xi, H., Schwartz, R., Engel, I., Murre, C., Kersh, G.J., 2006. Interplay between RORgammat,Egr3, and E proteins controls proliferation in response to pre-TCR signals. Immunity24, 813–826.
Yokoyama, M., Kusui, A., Sakamoto, S., Fukuzaki, H., 1984. Age-associated increments inhuman platelet α-adrenoceptor capacity. Possible mechanism for platelethyperactivity to epinephrine in aging man. Thrombosis Res. 34, 287–295.
Zamoyska, R., Lovatt, M., 2004. Signalling in T-lymphocyte development: integration ofsignalling pathways is the key. Curr. Opin. Immunol. 16, 191–196.
Copyright © 2022 FDOKUMEN




















