
Removal of the N-terminal hexapeptide from human β2-microglobulin facilitates protein aggregation...
15
Removal of the N-terminal hexapeptide from human b2-microglobulin facilitates protein aggregation and fibril formation G. ESPOSITO, 1 R. MICHELUTTI, 1 G. VERDONE, 1 P. VIGLINO, 1 H. HERNÁNDEZ, 2 C.V. ROBINSON, 2 A. AMORESANO, 3 F. DAL PIAZ, 3 M. MONTI, 3 P. PUCCI, 3 P. MANGIONE, 4,5 M. STOPPINI, 4 G. MERLINI, 4,5 G. FERRI, 4 and V. BELLOTTI 4,5 1 Dipartimento di Scienze e Tecnologie Biomediche, Università di Udine, P. le Kolbe 4, 33100 Udine, Italy 2 Oxford Centre for Molecular Sciences, University of Oxford, South Parks Road, Oxford OX1 3QT, United Kingdom 3 Dipartimento di Chimica Organica e Biologica and Centro Internazionale Servizi Spettrometria di Massa, Università di Napoli Federico II, Via Mezzocannone 16, 80124 Napoli, Italy 4 Dipartimento di Biochimica, Università di Pavia, via Taramelli 3b, 27100 Pavia, Italy 5 Laboratorio di Biotecnologie IRCCS, Policlinico S. Matteo, 27100 Pavia, Italy ~Received November 16, 1999; Final Revision February 23, 2000; Accepted March 3, 2000! Abstract The solution structure and stability of N-terminally truncated b2-microglobulin ~DN6b2-m!, the major modification in ex vivo fibrils, have been investigated by a variety of biophysical techniques. The results show that DN6b2-m has a free energy of stabilization that is reduced by 2.5 kcal 0mol compared to the intact protein. Hydrogen exchange of a mixture of the truncated and full-length proteins at mM concentrations at pH 6.5 monitored by electrospray mass spectrometry reveals that DN6b2-m is significantly less protected than its wild-type counterpart. Analysis of DN6b2-m by NMR shows that this loss of protection occurs in b strands I, III, and part of II. At mM concentration gel filtration analysis shows that DN6b2-m forms a series of oligomers, including trimers and tetramers, and NMR analysis indicates that strand V is involved in intermolecular interactions that stabilize this association. The truncated species of b2- microglobulin was found to have a higher tendency to self-associate than the intact molecule, and unlike wild-type protein, is able to form amyloid fibrils at physiological pH. Limited proteolysis experiments and analysis by mass spectrometry support the conformational modifications identified by NMR and suggest that DN6b2-m could be a key intermediate of a proteolytic pathway of b2-microglobulin. Overall, the data suggest that removal of the six residues from the N-terminus of b2-microglobulin has a major effect on the stability of the overall fold. Part of the tertiary structure is preserved substantially by the disulfide bridge between Cys25 and Cys80, but the pairing between b-strands far removed from this constrain is greatly perturbed. Keywords: amyloidosis; b2-microglobulin; hydrogen exchange mass spectrometry; limited proteolysis; NMR; protein folding Amyloidoses are diseases caused by tissue deposition of protein aggregate organized in an ordered b-sheet structure. The conver- sion of globular proteins to insoluble fibrillar aggregates requires significant conformational changes, such as the loss of tertiary and quaternary interactions or conversion of a to b secondary structure ~Sunde & Blake, 1998!. Of the 17 or so proteins implicated in amyloidoses the fibril morphology is indistinguishable and there does not appear to be any common features that link the soluble precursor proteins. For many of these proteins, the amyloid fibril formation is facilitated by amino acid mutations that destabilize the native state and confer a structural flexibility to the molecule, but other proteins like IAPP, wild-type TTR, and b2-microglobulin Reprint requests to: Dr. Vittorio Bellotti, Dipartimento di Biochimica, Università di Pavia, via Taramelli 3b, 27100 Pavia, Italy; e-mail: [email protected]. Abbreviations: b2-m, b2-microglobulin; DN6b2-m, truncated species of b2-m comprising residues 7–99; EDTA, ethylenediaminetetraacetic acid; FID, free induction decay; IPTG, isopropyl b-d-thiogalactopyranoside; PMSF, phenylmethylsulfonyl fluoride; GdnHCl, guanidine hydrochlo- ride; HPLC, high-performance liquid chromatography; TFA, trifluoroace- tic acid; CD, circular dichroism; MD, molecular dynamics; MS, mass spectrometry; NOE, nuclear Overhauser effect; NOESY, NOE spectros- copy; PCR, polymerase chain reaction; PDB, Protein Data Bank; TOCSY, total correlation spectroscopy; C m , midpoint of GdnHCl un- folding transition; 1D, one-dimensional; 2D, two-dimensional; 3D, three-dimensional. Protein Science ~2000!, 9:831–845. Cambridge University Press. Printed in the USA. Copyright © 2000 The Protein Society 831
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Removal of the N-terminal hexapeptide from human β2-microglobulin facilitates protein aggregation...

Removal of the N-terminal hexapeptide from humanb2-microglobulin facilitates protein aggregationand fibril formation
G. ESPOSITO,1 R. MICHELUTTI,1 G. VERDONE,1 P. VIGLINO,1 H. HERNÁNDEZ,2
C.V. ROBINSON,2 A. AMORESANO,3 F. DAL PIAZ,3 M. MONTI,3 P. PUCCI,3 P. MANGIONE,4,5
M. STOPPINI,4 G. MERLINI,4,5 G. FERRI,4 and V. BELLOTTI 4,5
1Dipartimento di Scienze e Tecnologie Biomediche, Università di Udine, P. le Kolbe 4, 33100 Udine, Italy2Oxford Centre for Molecular Sciences, University of Oxford, South Parks Road, Oxford OX1 3QT, United Kingdom3Dipartimento di Chimica Organica e Biologica and Centro Internazionale Servizi Spettrometria di Massa,Università di Napoli Federico II, Via Mezzocannone 16, 80124 Napoli, Italy
4Dipartimento di Biochimica, Università di Pavia, via Taramelli 3b, 27100 Pavia, Italy5Laboratorio di Biotecnologie IRCCS, Policlinico S. Matteo, 27100 Pavia, Italy
~Received November 16, 1999;Final Revision February 23, 2000;Accepted March 3, 2000!
Abstract
The solution structure and stability of N-terminally truncatedb2-microglobulin~DN6b2-m!, the major modification inex vivo fibrils, have been investigated by a variety of biophysical techniques. The results show thatDN6b2-m has a freeenergy of stabilization that is reduced by 2.5 kcal0mol compared to the intact protein. Hydrogen exchange of a mixtureof the truncated and full-length proteins atmM concentrations at pH 6.5 monitored by electrospray mass spectrometryreveals thatDN6b2-m is significantly less protected than its wild-type counterpart. Analysis ofDN6b2-m by NMRshows that this loss of protection occurs inb strands I, III, and part of II. At mM concentration gel filtration analysisshows thatDN6b2-m forms a series of oligomers, including trimers and tetramers, and NMR analysis indicates thatstrand V is involved in intermolecular interactions that stabilize this association. The truncated species ofb2-microglobulin was found to have a higher tendency to self-associate than the intact molecule, and unlike wild-typeprotein, is able to form amyloid fibrils at physiological pH. Limited proteolysis experiments and analysis by massspectrometry support the conformational modifications identified by NMR and suggest thatDN6b2-m could be a keyintermediate of a proteolytic pathway ofb2-microglobulin. Overall, the data suggest that removal of the six residuesfrom the N-terminus ofb2-microglobulin has a major effect on the stability of the overall fold. Part of the tertiarystructure is preserved substantially by the disulfide bridge between Cys25 and Cys80, but the pairing betweenb-strandsfar removed from this constrain is greatly perturbed.
Keywords: amyloidosis;b2-microglobulin; hydrogen exchange mass spectrometry; limited proteolysis; NMR; proteinfolding
Amyloidoses are diseases caused by tissue deposition of proteinaggregate organized in an orderedb-sheet structure. The conver-sion of globular proteins to insoluble fibrillar aggregates requiressignificant conformational changes, such as the loss of tertiary andquaternary interactions or conversion ofa to b secondary structure~Sunde & Blake, 1998!. Of the 17 or so proteins implicated inamyloidoses the fibril morphology is indistinguishable and theredoes not appear to be any common features that link the solubleprecursor proteins. For many of these proteins, the amyloid fibrilformation is facilitated by amino acid mutations that destabilizethe native state and confer a structural flexibility to the molecule,but other proteins like IAPP, wild-type TTR, andb2-microglobulin
Reprint requests to: Dr. Vittorio Bellotti, Dipartimento di Biochimica,Università di Pavia, via Taramelli 3b, 27100 Pavia, Italy; e-mail:[email protected].
Abbreviations:b2-m, b2-microglobulin; DN6b2-m, truncated speciesof b2-m comprising residues 7–99; EDTA, ethylenediaminetetraaceticacid; FID, free induction decay; IPTG, isopropylb-d-thiogalactopyranoside;PMSF, phenylmethylsulfonyl fluoride; GdnHCl, guanidine hydrochlo-ride; HPLC, high-performance liquid chromatography; TFA, trifluoroace-tic acid; CD, circular dichroism; MD, molecular dynamics; MS, massspectrometry; NOE, nuclear Overhauser effect; NOESY, NOE spectros-copy; PCR, polymerase chain reaction; PDB, Protein Data Bank;TOCSY, total correlation spectroscopy;Cm, midpoint of GdnHCl un-folding transition; 1D, one-dimensional; 2D, two-dimensional; 3D,three-dimensional.
Protein Science~2000!, 9:831–845. Cambridge University Press. Printed in the USA.Copyright © 2000 The Protein Society
831
are amyloidogenic in the wild-type form~Sunde & Blake, 1998!.Despite extensive study of the fibril formation process, a generalmechanism has not yet emerged. There is, however, a growingawareness that the process may constitute an alternative proteinfolding pathway through which partially folded intermediates aredriven to an aggregate state with favorable overall free energy~Radford & Dobson, 1999!.
Human b2-microglobulin ~b2-m! is one of the group of 17proteins associated with disease and this type of amyloidosis rep-resents an inevitable and severe complication of long-term hae-modialysis~Gejyo et al., 1985!. Under native conditions,b2-mforms a predominantlyb-sheet structure organized in a jellyroll-like topology stabilized by a single disulfide bridge between Cys25and Cys80~Bjorkman et al., 1987!. No amyloidogenic mutationshave been described forb2-m, although chemical modificationssuch as glycation~Miyata et al., 1993!, oxidation ~Capeiller-Blandin et al., 1991!, and limited proteolysis have been reported~Linke et al., 1986, 1987, 1989!. Previously, we have shown thatproteolysis of the N-terminal region ofb2-m results in a truncationof the six N-terminal residues~DN6b2-m! in ;30% of theb2-mmolecules extracted from ex vivo fibrils~Bellotti et al., 1998!. Thistruncated species was shown to have a higher tendency to self-aggregate than the full-length protein and did not form a fullyfolded native state at the end of the refolding procedure~Bellottiet al., 1998!. This observation prompted us to examine further thestructural modifications induced by the removal of six N-terminalresidues to investigate the possible implications of thisb2-m frag-ment in the dynamics of fibril formation and stability.
Results
Reduced secondary structure inDN6b2-m
The effect of the removal of the N-terminal hexapeptide on thefolding stability was studied at increasing concentrations of Gdn-HCl. We have measured aCm ~unfolding midpoint! of 2.23 M for
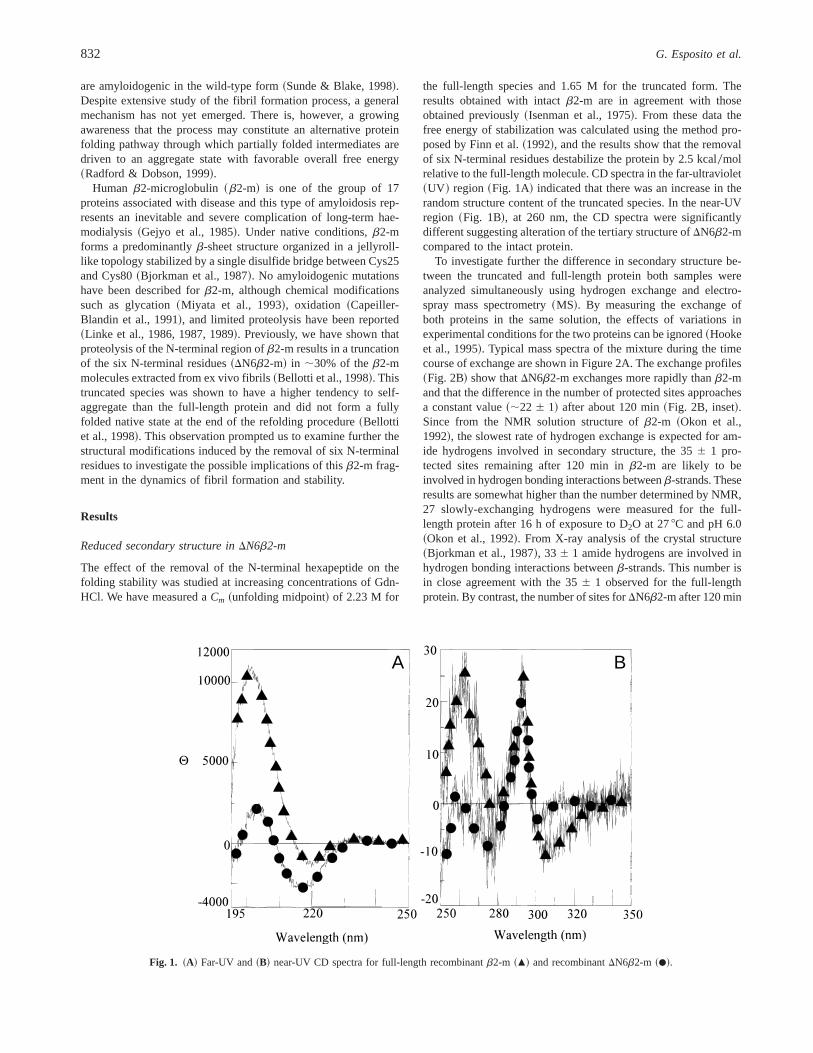
the full-length species and 1.65 M for the truncated form. Theresults obtained with intactb2-m are in agreement with thoseobtained previously~Isenman et al., 1975!. From these data thefree energy of stabilization was calculated using the method pro-posed by Finn et al.~1992!, and the results show that the removalof six N-terminal residues destabilize the protein by 2.5 kcal0molrelative to the full-length molecule. CD spectra in the far-ultraviolet~UV ! region~Fig. 1A! indicated that there was an increase in therandom structure content of the truncated species. In the near-UVregion ~Fig. 1B!, at 260 nm, the CD spectra were significantlydifferent suggesting alteration of the tertiary structure ofDN6b2-mcompared to the intact protein.
To investigate further the difference in secondary structure be-tween the truncated and full-length protein both samples wereanalyzed simultaneously using hydrogen exchange and electro-spray mass spectrometry~MS!. By measuring the exchange ofboth proteins in the same solution, the effects of variations inexperimental conditions for the two proteins can be ignored~Hookeet al., 1995!. Typical mass spectra of the mixture during the timecourse of exchange are shown in Figure 2A. The exchange profiles~Fig. 2B! show thatDN6b2-m exchanges more rapidly thanb2-mand that the difference in the number of protected sites approachesa constant value~;22 6 1! after about 120 min~Fig. 2B, inset!.Since from the NMR solution structure ofb2-m ~Okon et al.,1992!, the slowest rate of hydrogen exchange is expected for am-ide hydrogens involved in secondary structure, the 356 1 pro-tected sites remaining after 120 min inb2-m are likely to beinvolved in hydrogen bonding interactions betweenb-strands. Theseresults are somewhat higher than the number determined by NMR,27 slowly-exchanging hydrogens were measured for the full-length protein after 16 h of exposure to D2O at 278C and pH 6.0~Okon et al., 1992!. From X-ray analysis of the crystal structure~Bjorkman et al., 1987!, 336 1 amide hydrogens are involved inhydrogen bonding interactions betweenb-strands. This number isin close agreement with the 356 1 observed for the full-lengthprotein. By contrast, the number of sites forDN6b2-m after 120 min
A B
Fig. 1. ~A! Far-UV and~B! near-UV CD spectra for full-length recombinantb2-m ~m! and recombinantDN6b2-m ~d!.
832 G. Esposito et al.

exchange is considerably reduced, indicating that the extent ofhydrogen bonded structure inDN6b2-m is significantly lower thanthat measured for the full-length protein.
The solution structure ofDN6b2-m
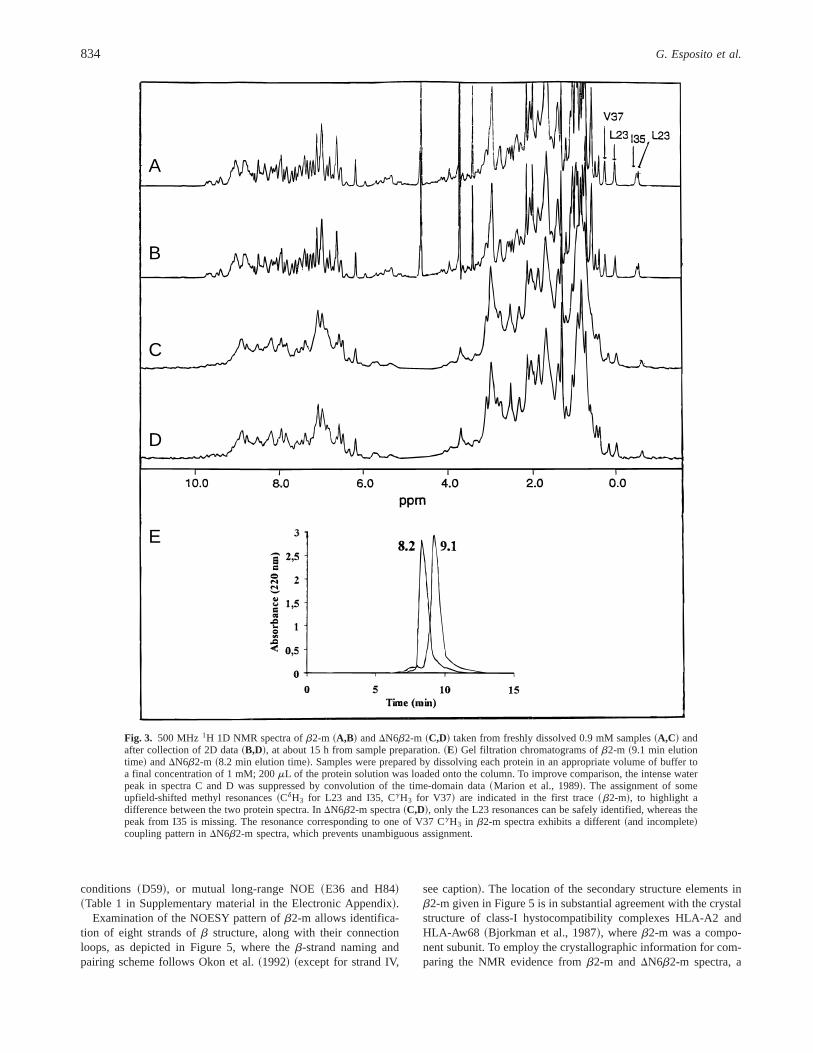
To examine in more detail the origin of the reduced hydrogenexchange protection in the truncated protein, the solution structureof DN6b2-m was investigated by NMR. The1H NMR assignmentof the full-length protein and its secondary structure determinationhave been reported~Okon et al., 1992!. Although some difficultieswere experienced with precipitation 50–60 h after sample prepa-ration, a 0.9 mM sample ofb2-m, at pH 6.6 and 378C, did notexhibit significant broadening during two-dimensional~2D! spec-tra collection~Fig. 3A,B!. For DN6b2-m precipitation was more
extensive and rapid~50% loss of signal at 15 h! although thespectrum did not change~Fig. 3C,D!. It can be concluded that slowdenaturation or oligomerization processes affecting linewidthsand0or resonance chemical shifts did not occur during spectraacquisition.
The 2D NMR spectra ofb2-m were compared with reportedassignments~Okon et al., 1992!. Spin systems were first identifiedin the TOCSY spectrum~Fig. 4A! and then confirmed by checkingthe sequential connectivity pattern in the NOESY maps~Fig. 4C!~Wüthrich, 1986!. For b-sheet regions, the presence of the inter-strand contacts was ascertained. This led to the identification of theresonance pattern for 93 residues~plus Met0! out of 99 and alsoconfirmed the integrity of the disulfide bridge. No consistent res-onance pattern was observed for the remaining six residues~I1,N17, D53, S61, P72, K91! due to exchange broadening and0orinherent line width problems as reported also previously~Okonet al., 1992!.
The 1D1H spectra ofDN6b2-m differ in two respects comparedwith those ofb2-m ~Fig. 3!. First, the resonance linewidths inDN6b2-m appear broader than inb2-m, a feature that may arisefrom either aggregation0association or conformational exchange,or a contribution from both these effects. The oligomeric nature ofDN6b2-m was investigated by gel filtration chromatography at thesame protein concentration used for NMR analysis and the resultsare shown in Figure 3E.DN6b2-m is eluted with a retention timeconsistent with the presence of trimers and tetramers, whereas thefull length b2-m is eluted with a retention time consistent with anequilibrium of monomeric and dimeric forms. The second differ-ence between the two protein spectra concerns the overall reso-nance envelope. This suggests that, besides aggregation,b2-mundergoes a conformational change upon deletion of the N-terminalhexapeptide. For instance, the typical features observed in theupfield region of the NMR spectrum of the full-length protein~dueto shifts induced by buried aromatic rings! arise from hydrogennuclei of residues far removed from the deleted fragment and,consequently, might be expected to be similar for the truncatedspecies. However, in the spectra ofDN6b2-m, these resonances areonly partially retained.
To investigate further the issue 2D TOCSY and NOESY spectraof DN6b2-m were analyzed in detail~Fig. 4B,D! to identify changesin scalar and dipolar patterns with respect to the full-length protein.The results~see Table 1 in Supplementary material in the Elec-tronic Appendix! can be summarized by splitting the overall evi-dence into two broad resonance categories: those that retain thechemical shift and dipolar connectivity patterns of the intact pro-tein and those that indicate disordered fragments inDN6b2-mthrough resonance overlap within the typical spectral regions ofaminoacyl residues of structureless oligopeptides~Wishart & Sykes,1994!. Besides these two resonance categories, it is also possible toobserve a third less populated class ofDN6b2-m resonances thatexhibit sizable deviations from the limiting random coil chemicalshift values~Wishart et al., 1995!. However, no firm identificationcould be reached for this latter group, from either pattern similarityrecognition or de novo assignment procedures due to either theambiguity or incompleteness of the scalar connectivity pattern orthe failure to follow sequential NOE contacts leading to the crowdedrandom coil region, etc.~Fig. 4!. However, 38 of the 93 residueswere assigned with complete or partial reconstruction of the spin-system pattern. In addition, four other residue spin-systems couldbe totally or partially assigned, based either on the uniqueness oftheir chemical shift pattern~L23!, favorable resonance resolution
A
B
Fig. 2. Hydrogen-deuterium exchange at pH 7 and 238C monitored bynanoflow MS.A: ESI mass spectrum of both proteins in H2O ~upper trace!and D2O after 120 min exchange~lower trace!. Charge states are labeledfor b2-m ~series A! andDN6b2-m ~series B!. B: Exchange profiles~threereplicates! as the number of protected sites~H! with respect to time forb2-m ~m! andDN6b2-m ~d!. The predicted random coil exchange profile~—!, calculated using HXPep1.1, is also shown. Inset: The difference in thenumber of protected sites betweenb2-m andDN6b2-m for each time point.
Role of truncated species ofb2-m in fibrillogenesis 833
conditions ~D59!, or mutual long-range NOE~E36 and H84!~Table 1 in Supplementary material in the Electronic Appendix!.
Examination of the NOESY pattern ofb2-m allows identifica-tion of eight strands ofb structure, along with their connectionloops, as depicted in Figure 5, where theb-strand naming andpairing scheme follows Okon et al.~1992! ~except for strand IV,
see caption!. The location of the secondary structure elements inb2-m given in Figure 5 is in substantial agreement with the crystalstructure of class-I hystocompatibility complexes HLA-A2 andHLA-Aw68 ~Bjorkman et al., 1987!, whereb2-m was a compo-nent subunit. To employ the crystallographic information for com-paring the NMR evidence fromb2-m andDN6b2-m spectra, a
A
B
C
D
E
Fig. 3. 500 MHz 1H 1D NMR spectra ofb2-m ~A,B! andDN6b2-m ~C,D! taken from freshly dissolved 0.9 mM samples~A,C! andafter collection of 2D data~B,D!, at about 15 h from sample preparation.~E! Gel filtration chromatograms ofb2-m ~9.1 min elutiontime! andDN6b2-m ~8.2 min elution time!. Samples were prepared by dissolving each protein in an appropriate volume of buffer toa final concentration of 1 mM; 200mL of the protein solution was loaded onto the column. To improve comparison, the intense waterpeak in spectra C and D was suppressed by convolution of the time-domain data~Marion et al., 1989!. The assignment of someupfield-shifted methyl resonances~CdH3 for L23 and I35, CgH3 for V37! are indicated in the first trace~b2-m!, to highlight adifference between the two protein spectra. InDN6b2-m spectra~C,D!, only the L23 resonances can be safely identified, whereas thepeak from I35 is missing. The resonance corresponding to one of V37 CgH3 in b2-m spectra exhibits a different~and incomplete!coupling pattern inDN6b2-m spectra, which prevents unambiguous assignment.
834 G. Esposito et al.

Fig. 4. 500 MHz 1H fingerprint regions from 2D NMR TOCSY and NOESY spectra ofb2-m ~A,C! andDN6b2-m ~B,D!. The Ha-HN
TOCSY ~A,B! connectivity assignments are indicated by amino acid single letter code. Only the interresidue sequential Hia-Hi11
N
connectivities are labeled in NOESY contour plots~C,D!, using just the residue number to limit graphic crowding. Prior to 2D-FT, all2D experimental data were processed with right shift int1 dimension by 1 point to remove baseline offsets~Marion & Bax, 1989!, zerofilling and apodization with square sinebell function shifted by 608 in both dimensions, to end up with final matrix dimensions of2,0483 2,048 real points. The quality ofDN6b2-m spectra was affected by transient field inhomogeneity due to precipitation occurringthroughout data collection. For example, the isolated NOESY cross peak occurring at approximately 9.7104.36 ppm for both molecules~C,D!, i.e., at quite peculiar frequency coordinates, could be attributed to the sequentialai -NHi11 contact between K41 and N42 onlyfor b2-m, in spite of being observed also in the NOESY fingerprint ofDN6b2-m. The scalar patterns of the corresponding spin systems,in fact, do not overcome the noise threshold in the TOCSY contour plot ofDN6b2-m ~B! and, therefore, could not be listed amongthe attributions of Table 1~see Supplementary material in the Electronic Appendix!. ~Figure continues on next page.!
Role of truncated species ofb2-m in fibrillogenesis 835
consistency check was first undertaken, in the absence of an NMR-restrained solution structure of the parent molecule. In addition toverifying theb2-m assignments with respect to the NOE restraintset, the crystallographic secondary structure elements were checkedagainst the experimental chemical shift set; by plotting the Ha
chemical shift deviations with respect to the limiting values~Dd!,it is possible to readily visualize a gross distribution of secondarystructure elements along the protein sequence~Wishart & Sykes,1994; Wishart et al., 1995!. For b2-m, such a plot~Fig. 6! showsa good agreement with the X-ray data and represents an additional
Fig. 4. Continued.
836 G. Esposito et al.

piece of evidence that is consistent with the results of the quali-tative NOE analysis~Okon et al., 1992!. Small discrepancies amongNOE restraints, Ha chemical shifts, and crystal structure are de-tailed in Supplementary material in the Electronic Appendix. It canbe concluded, however, that the conformation of the isolated pro-tein in solution is closely related to its crystallographic structure inthe complex, which can be employed as a model at this stage.
The subsequent step in the analysis of theDN6b2-m NMR datawas to determine structural inferences for the identified spin-system that were invariant with respect to the parent molecule. Forinstance, the isolated resonance of L23 CdH3 showed a uniquechemical shift, with an upfield deviation of;1.5 ppm with respectto the limiting value, which can only arise from retention of thespatial proximity relationships responsible for that deviation, i.e.,the ring current shift due to the proximity and orientation of Y67aromatic ring~strand VI! with respect to L23~strand II! ~Fig. 7A!.This, besides accounting for the deviations of L23 and E69 Ha Ddvalues from secondary-structure diagnostic values~Fig. 6!, alsoimplies that the spatial geometry ofb2-m around the single disul-fide bridge of the molecule, joining C25 and C80, must be con-
served inDN6b2-m and must involve the intervening segments ofstrand II and strand VI.
The absence or the partial pattern variation, in theDN6b2-mspectra, of two other characteristically shifted resonances ofb2-mspectra, i.e., I35 CdH3 and V37 CgH3 ~Fig. 3!, must reflect astructural change undergone by strand III ofb2-m on deletion ofthe N-terminal hexapeptide. Coupled with the failure to observeany invariant spin-system from that fragment~except E36!, theresults indicate that strand III is either lost or paired differently inDN6b2-m. In general, the majority of the 42 assigned residues ofDN6b2-m exhibited high chemical shift invariance compared tob2-m. Most of these residues are clustered along strands VI, VII,and VIII of the secondary structure ofb2-m ~Fig. 5, Table 1 inSupplementary material in the Electronic Appendix!, with a few atthe initial segment of strand II and the final segment of strand I,i.e., a region of the 3D structure that may be constrained by theC25–C80 disulfide bridge~Fig. 7A!. For the four isolated spin-systems of the pool recovered fromDN6b2-m spectra, i.e., L23~strand II!, E36 ~strand III!, H84 ~strand VII!, and D59 ~loopbetween strands V and VI!, failure to identify at least one adjacent
Fig. 5. Synopsis of the NMR assignments obtained fromb2-m andDN6b2-m spectra. Dotted-contour shaded symbols indicateresidues that were not assigned in either protein spectra. White-filled symbols indicate residues that could be assigned only inb2-mspectra. Grey-filled0white characters symbols indicate the residues that were identified inb2-m andDN6b2-m spectra. The secondarystructure designation of the individual residues and theb-strand naming and pairing scheme follow from Okon et al.~1992!, exceptfor the orientation of strand IV, which was reported as paired to strand III in Okon’s et al. scheme. In the above picture, the orientationof strand IV is intended to represent its nearly perpendicular arrangement with respect to strand III in the crystal structure.
Role of truncated species ofb2-m in fibrillogenesis 837

or closely-spaced residue for E36 and D59, and also the large Ha
shift deviation for H84~Table 1 in Supplementary material in theElectronic Appendix!, suggested that they should be excluded fromthe constraint list for subsequent calculations. However, L23 wasincluded due to the invariance of the upfield shift experienced byall resonances of this residue inDN6b2-m spectra~see previousdiscussion!.
Molecular modeling
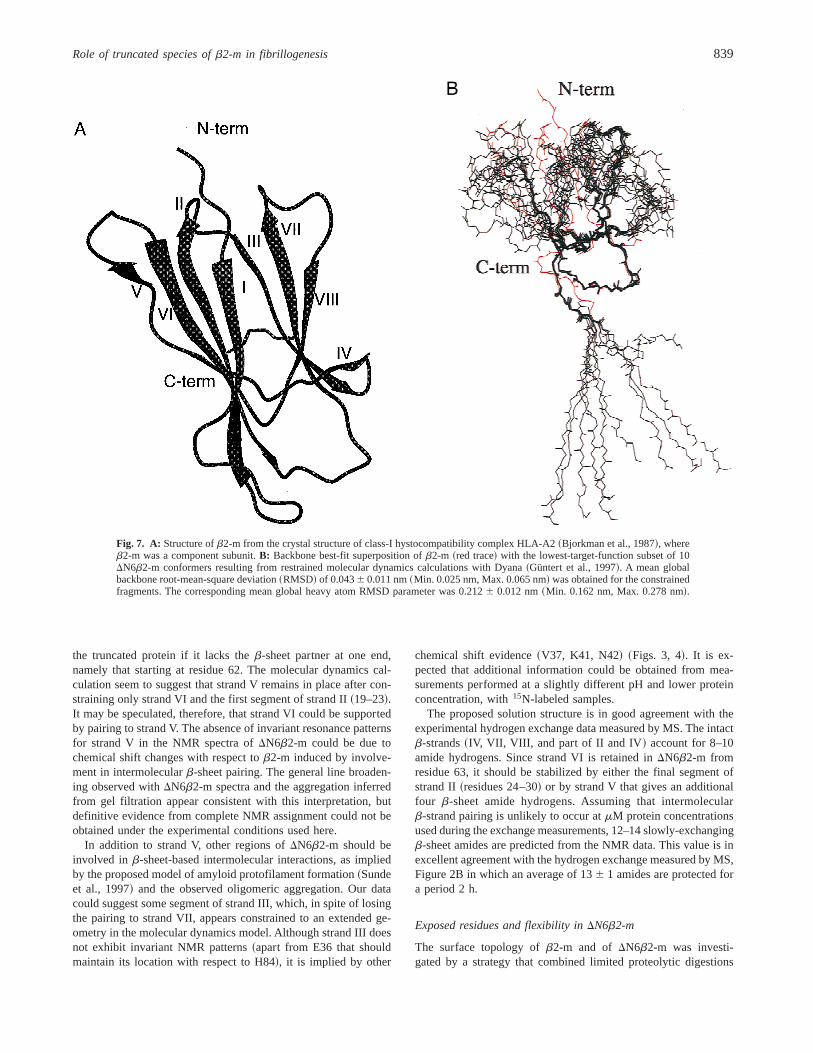
Assuming that the invariance of chemical shifts inDN6b2-m re-flects the persistence of local spatial arrangements, molecular dy-namics simulations were performed using the crystallographic intraand interresidue Ca–Cb separations as restraints for fragments19–23, 44–49, 63–81, 87–89, and 93–99, which, according toOkon’s et al. scheme~Fig. 5!, are located in strands II, IV, VI, VII,and VIII, the loops between strands I and II~residue 19!, strandsIV and V ~fragment 47–49!, and strands VI and VII~fragment72–76!, the bulge of strand VIII~fragment 87–89!, and at theC-terminal tail ~fragment 96–99!. In addition to E36, D59, andH84, also fragment 11–13 was not constrained due to the largedeviation of R12 Ha chemical shift with respect tob2-m and thefailure to detect the R12 HN resonance. In total, 3,003 restraintswere introduced in the calculation input and 200 initial randomstructures were generated. The results, depicted in Figure 7B, arethe best-fit superposition ofb2-m crystal structure and the 10 con-formers ofDN6b2-m with lowest target function~Güntert et al.,
1997!. As expected, the relative arrangement of the restrainedstrands and loops is maintained within the selected conformerfamily. The organization and relative orientation of strands I, II,and III are totally lost or substantially altered, except for the initialsegment of strand II, which, besides the imposed restraints, isconstrained also by the disulfide bridge to retain its position withrespect to strands VI and VII. A relatively limited conformationalspread is observed for the unrestrained strand V, which retains itsinitial orientation with respect to strand VI, in spite of the absenceof properb-sheet pairing. It is noteworthy that in the crystal struc-ture ofb2-m, strand V is formed by two dipeptide fragments inbstructure~50–51 and 55–56!, paired to strand VI and separated bya b-bulge centered at residues 53–54. Evidence in favor of thisb-bulge in b2-m is also observed in solution from NOEs~Okonet al., 1992! and from our data, based on the further upfield de-viation of L64 Ha ~Fig. 5; Table 1 in Supplementary material in theElectronic Appendix! with respect to the reported chemical shift~Okon et al., 1992!. It is very likely, therefore, that also in thesolution state ofb2-m, the b-sheet organization of strand V islimited and discontinuous. The NMR data forDN6b2-m show thata similarb-structure conformation is retained for strand VI and forthe first segment of strand II compared to the parent molecule~Table 1 in Supplementary material in the Electronic Appendix;Fig. 7B!. No resonance group is observed that matches the patternof the initial segment of strand V~50–51! or the final segment ofstrand II ~26–30!, both paired to strand VI in the full-length pro-tein. The question arises as to how strand VI can be conserved in
Fig. 6. Histogram ofb2-m Ha Dd values vs. the residue number.Dd is a Ha chemical shift difference defined asDd 5 dexpti 2 dlimit
i ,where the superscripti refers to the amino acid residue and the subscriptsexpt and limit stand for experimental and limiting,respectively~Wishart et al., 1995!. The reported secondary structure classifications follow from the PDB record~3hla! of HLA-A2crystal structure~Bjorkman et al., 1987!. Missing Ha chemical shift~residues I1, P5, N17, D53, S61, P72, P90, K91, and W95! werereplaced by Okon et al.’s values~white bars!. Gaps correspond to glycine residues, namely 18, 29, and 43, for which no stereospecificassignment was obtained.
838 G. Esposito et al.
the truncated protein if it lacks theb-sheet partner at one end,namely that starting at residue 62. The molecular dynamics cal-culation seem to suggest that strand V remains in place after con-straining only strand VI and the first segment of strand II~19–23!.It may be speculated, therefore, that strand VI could be supportedby pairing to strand V. The absence of invariant resonance patternsfor strand V in the NMR spectra ofDN6b2-m could be due tochemical shift changes with respect tob2-m induced by involve-ment in intermolecularb-sheet pairing. The general line broaden-ing observed withDN6b2-m spectra and the aggregation inferredfrom gel filtration appear consistent with this interpretation, butdefinitive evidence from complete NMR assignment could not beobtained under the experimental conditions used here.
In addition to strand V, other regions ofDN6b2-m should beinvolved in b-sheet-based intermolecular interactions, as impliedby the proposed model of amyloid protofilament formation~Sundeet al., 1997! and the observed oligomeric aggregation. Our datacould suggest some segment of strand III, which, in spite of losingthe pairing to strand VII, appears constrained to an extended ge-ometry in the molecular dynamics model. Although strand III doesnot exhibit invariant NMR patterns~apart from E36 that shouldmaintain its location with respect to H84!, it is implied by other
chemical shift evidence~V37, K41, N42! ~Figs. 3, 4!. It is ex-pected that additional information could be obtained from mea-surements performed at a slightly different pH and lower proteinconcentration, with15N-labeled samples.
The proposed solution structure is in good agreement with theexperimental hydrogen exchange data measured by MS. The intactb-strands~IV, VII, VIII, and part of II and IV! account for 8–10amide hydrogens. Since strand VI is retained inDN6b2-m fromresidue 63, it should be stabilized by either the final segment ofstrand II~residues 24–30! or by strand V that gives an additionalfour b-sheet amide hydrogens. Assuming that intermolecularb-strand pairing is unlikely to occur atmM protein concentrationsused during the exchange measurements, 12–14 slowly-exchangingb-sheet amides are predicted from the NMR data. This value is inexcellent agreement with the hydrogen exchange measured by MS,Figure 2B in which an average of 136 1 amides are protected fora period 2 h.
Exposed residues and flexibility inDN6b2-m
The surface topology ofb2-m and of DN6b2-m was investi-gated by a strategy that combined limited proteolytic digestions
Fig. 7. A: Structure ofb2-m from the crystal structure of class-I hystocompatibility complex HLA-A2~Bjorkman et al., 1987!, whereb2-m was a component subunit.B: Backbone best-fit superposition ofb2-m ~red trace! with the lowest-target-function subset of 10DN6b2-m conformers resulting from restrained molecular dynamics calculations with Dyana~Güntert et al., 1997!. A mean globalbackbone root-mean-square deviation~RMSD! of 0.0436 0.011 nm~Min. 0.025 nm, Max. 0.065 nm! was obtained for the constrainedfragments. The corresponding mean global heavy atom RMSD parameter was 0.2126 0.012 nm~Min. 0.162 nm, Max. 0.278 nm!.
Role of truncated species ofb2-m in fibrillogenesis 839

with mass spectrometric identification of the fragments. The ra-tionale behind the method is that the amino acid residues thatare located within exposed and flexible regions of the proteinare more susceptible to proteolysis~Zappacosta et al., 1996;Scaloni et al., 1998!. Moreover, conformational changes affect-ing protein topology can be monitored by the appearance0disappearance of preferential cleavage sites~Orrù et al., 1999;Scaloni et al., 1999!. Limited proteolysis experiments were per-formed on bothb2-m andDN6b2-m at a protein concentrationof 10 mM by using a series of enzymes as conformational probes.At this protein concentration, both proteins showed the sameretention time by gel filtration, consistent with the presence ofmonomers and dimers. As an example, when using chymotryp-sin a rapid hydrolysis of intactb2-m at Y10 and L87 was in-ferred by the identification of the peptide pairs 1–10, 11–99,1–87, and 88–99. A further component displaying a molecularmass of 11,877.76 1.0 Da was detected in the early stages ofdigestion and assigned to a nicked form of the protein. Reduc-tion of this fragment with dithiothreitol followed by MALDI0MSanalysis revealed the occurrence of a further preferential cleav-age site at W60.
Chymotryptic proteolysis ofDN6b2-m required a much lowerenzyme-to-substrate ratio compared tob2-m, suggesting a higheraccessibility of the truncated protein. In common withb2-m,DN6b2-m was readily cleaved at Y10 and L87. However, no fur-ther cleavage sites were detected even after prolonged digestionindicating that in the truncated protein W60 is no longer accessibleto proteases. Similar results were obtained with the other proteo-lytic enzymes, and the cleavage sites are reported in Scheme 1. Inthe N-terminal region, both proteins were cleaved at Y10 but onlyDN6b2-m underwent hydrolysis at Q8, K19, and E36. This stronglysuggests that the N-terminal region of the truncated protein is moredisordered than the corresponding region of the parent molecule, inagreement with the NMR results. The cleavage at K19, only ob-served inDN6b2-m is particularly interesting because a fragmentstarting at position 20 is frequently identified in natural fibrils asreported by Linke et al.~1989! and recently by our group~Stoppiniet al., 2000!. The proteolytic patterns in the regions of strands Vand VI fromb2-m indicate that cleavage occurs at the loop~S57,K58, D59, and W60! and theb-bulge of strand V~D53!, i.e., atrather exposed and flexible locations of the parent structure. InDN6b2-m, only the initial part of strand V undergoes hydrolysis~E50, D53!, while theb-bulge, loop~except S57!, and strand VIappear to be protected, suggesting that this region is endowed withless conformational flexibility. This interpretation agrees with theinference from NMR of an additional, novel pairing-arrangementproposed for strand V inDN6b2-m.
Proteolysis at the C-terminus occurred at L87, K91, V93, andW95 for both proteins, while onlyDN6b2-m underwent cleavageat D96 and D98. According to the NMR results,DN6b2-m retains
the paring between strands VII~fragment 78–81! and VIII ~frag-ment 93–95!, which is consistent with the proteolysis at the end ofthe intervening turn and within strand VIII. The accessibility ofboth locations is a feature that may also be appreciated by inspec-tion of the crystal structure ofb2-m ~Fig. 7A!. In the latter, theC-terminal fragment 95–99 adopts a well-defined conformation, inspite of not being involved in anyb-sheet, as reflected by theinterresidue H-bonds D96 Od2–D98 HN and D96 O–M99 HN. TheNOESY spectra ofb2-m showed intra- and interresidue correla-tions that are compatible with the crystallographic value ofx96
1
~2171.48!, while no clear conclusion could be obtained for theside-chain conformation of D98~x98
1 5 264.88!. Hence, it is notsurprising that the Asp-specific protease that was employed for theassay failed to hydrolyseb2-m at the aspartyl residues in posi-tion 96 and 98, whereas location W95 underwent processing. Onthe other hand, from the NMR data obtained withDN6b2-m, onlythe HN and Ha resonance of D96 could be assigned, while for D98the whole spin system pattern could be observed without anychanges. This suggests that the conformation of D96 side chainmight be changed inDN6b2-m and thus explain the different re-sponse to proteolysis.
DN6b2-m has a greater propensity to form amyloid fibrils
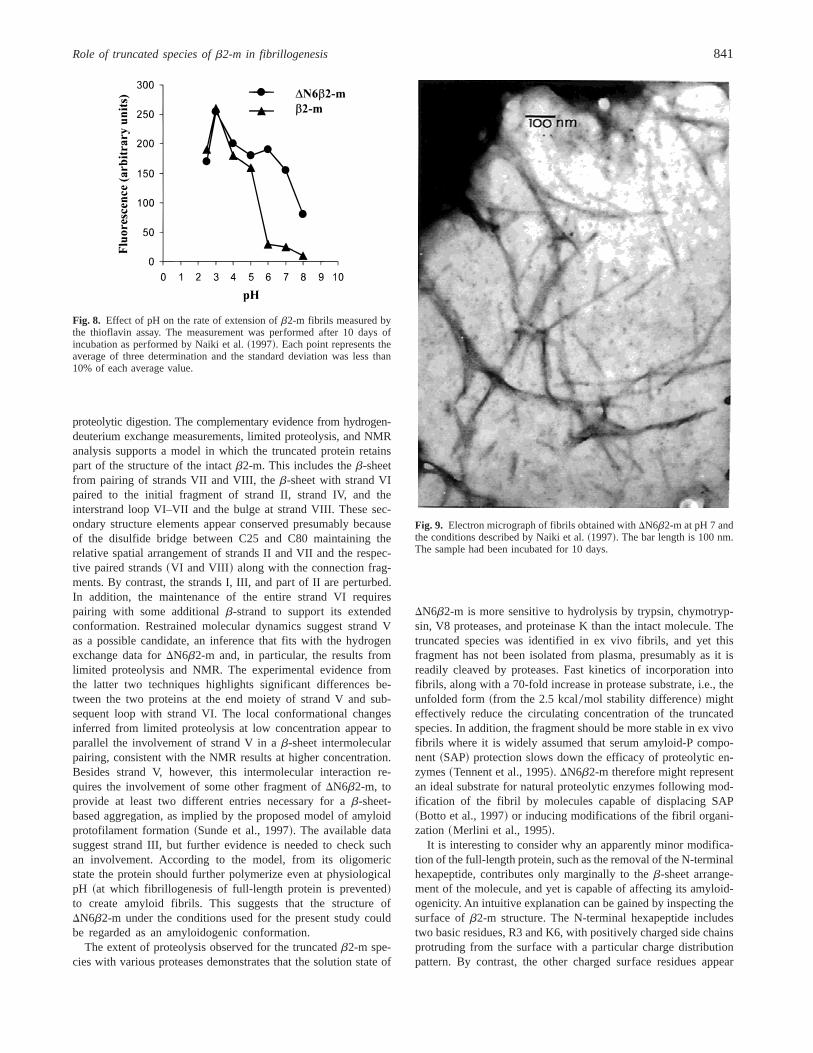
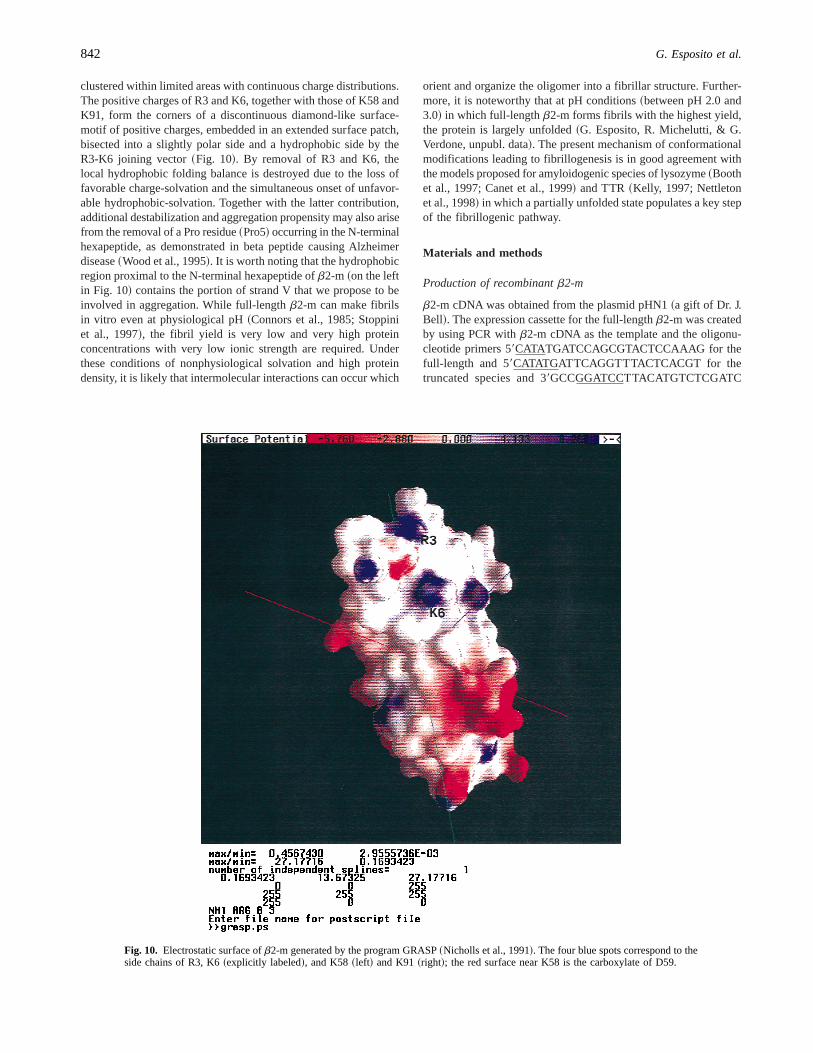
The tendency of full-length andDN6b2-m to self-aggregate wasmonitored by light scattering at various pH from 8 to 4. The resultsshowed that, at a pH lower than 7,DN6b2-m aggregates andprecipitates. Electron microscopy of the protein aggregate revealedthe coexistence of an amorphous precipitate and short fibrillarstructure having a diameter of;10 nm. This material formed acomplex with Congo red and showed the bathochromic shift of theCongo red spectrum typical for amyloid fibrils. However, the ma-terial analyzed by polarized light microscopy did not show theclassical red-green birefringency. Using the fibrillogenesis methodproposed by Naiki et al.~1997!, we found that both theb2-mspecies can extend the natural fibrils. In our hands the fibrils werevisible after 24 h of incubation and a continuous fibril growthcould be monitored for 2 weeks. Quantification of fibril formationis in good agreement with inspection of the samples by electronmicroscopy and demonstrates that, at neutral pH, onlyDN6b2-mmaintains the capacity to extend the natural fibrils. Figure 8 reportsthe quantification of fibril extension at different pHs, and Figure 9shows the fibrils obtained withDN6b2-m at pH 7.
Discussion
The combination of results of this study demonstrates that lossof the first six residues ofb2-m not only modifies its 3D struc-ture but also reduces its folding stability, facilitates fibril forma-tion at physiological pH, and enhances its susceptibility to
10 20 30 40 50b2m: MIQRTPKIQVYSRHPAENGKSNFLNCYVSGFHPSDIEVDLLKNGERIEKVEb2mDN6: MIQVYSRHPAENGKSNFLNCYVSGFHPSDIEVDLLKNGERIEKVE
60 70 80 90b2m: HSDLSFSKDWSFYLLYYTEFTPTEKDEYACRVNHVTLSQPKI VKWDRDMb2mDN6: HSDLSFSKDWSFYLLYYTEFTPTEKDEYACRVNHVTLSQPKI VKWDRDM
Scheme 1.Amino acid sequences ofb2-m andDN6b2-m. Preferential cleavage sites detected in each protein during limited prote-olysis experiments are in bold.
840 G. Esposito et al.
proteolytic digestion. The complementary evidence from hydrogen-deuterium exchange measurements, limited proteolysis, and NMRanalysis supports a model in which the truncated protein retainspart of the structure of the intactb2-m. This includes theb-sheetfrom pairing of strands VII and VIII, theb-sheet with strand VIpaired to the initial fragment of strand II, strand IV, and theinterstrand loop VI–VII and the bulge at strand VIII. These sec-ondary structure elements appear conserved presumably becauseof the disulfide bridge between C25 and C80 maintaining therelative spatial arrangement of strands II and VII and the respec-tive paired strands~VI and VIII ! along with the connection frag-ments. By contrast, the strands I, III, and part of II are perturbed.In addition, the maintenance of the entire strand VI requirespairing with some additionalb-strand to support its extendedconformation. Restrained molecular dynamics suggest strand Vas a possible candidate, an inference that fits with the hydrogenexchange data forDN6b2-m and, in particular, the results fromlimited proteolysis and NMR. The experimental evidence fromthe latter two techniques highlights significant differences be-tween the two proteins at the end moiety of strand V and sub-sequent loop with strand VI. The local conformational changesinferred from limited proteolysis at low concentration appear toparallel the involvement of strand V in ab-sheet intermolecularpairing, consistent with the NMR results at higher concentration.Besides strand V, however, this intermolecular interaction re-quires the involvement of some other fragment ofDN6b2-m, toprovide at least two different entries necessary for ab-sheet-based aggregation, as implied by the proposed model of amyloidprotofilament formation~Sunde et al., 1997!. The available datasuggest strand III, but further evidence is needed to check suchan involvement. According to the model, from its oligomericstate the protein should further polymerize even at physiologicalpH ~at which fibrillogenesis of full-length protein is prevented!to create amyloid fibrils. This suggests that the structure ofDN6b2-m under the conditions used for the present study couldbe regarded as an amyloidogenic conformation.
The extent of proteolysis observed for the truncatedb2-m spe-cies with various proteases demonstrates that the solution state of
DN6b2-m is more sensitive to hydrolysis by trypsin, chymotryp-sin, V8 proteases, and proteinase K than the intact molecule. Thetruncated species was identified in ex vivo fibrils, and yet thisfragment has not been isolated from plasma, presumably as it isreadily cleaved by proteases. Fast kinetics of incorporation intofibrils, along with a 70-fold increase in protease substrate, i.e., theunfolded form~from the 2.5 kcal0mol stability difference! mighteffectively reduce the circulating concentration of the truncatedspecies. In addition, the fragment should be more stable in ex vivofibrils where it is widely assumed that serum amyloid-P compo-nent ~SAP! protection slows down the efficacy of proteolytic en-zymes~Tennent et al., 1995!. DN6b2-m therefore might representan ideal substrate for natural proteolytic enzymes following mod-ification of the fibril by molecules capable of displacing SAP~Botto et al., 1997! or inducing modifications of the fibril organi-zation ~Merlini et al., 1995!.
It is interesting to consider why an apparently minor modifica-tion of the full-length protein, such as the removal of the N-terminalhexapeptide, contributes only marginally to theb-sheet arrange-ment of the molecule, and yet is capable of affecting its amyloid-ogenicity. An intuitive explanation can be gained by inspecting thesurface ofb2-m structure. The N-terminal hexapeptide includestwo basic residues, R3 and K6, with positively charged side chainsprotruding from the surface with a particular charge distributionpattern. By contrast, the other charged surface residues appear
Fig. 8. Effect of pH on the rate of extension ofb2-m fibrils measured bythe thioflavin assay. The measurement was performed after 10 days ofincubation as performed by Naiki et al.~1997!. Each point represents theaverage of three determination and the standard deviation was less than10% of each average value.
Fig. 9. Electron micrograph of fibrils obtained withDN6b2-m at pH 7 andthe conditions described by Naiki et al.~1997!. The bar length is 100 nm.The sample had been incubated for 10 days.
Role of truncated species ofb2-m in fibrillogenesis 841
clustered within limited areas with continuous charge distributions.The positive charges of R3 and K6, together with those of K58 andK91, form the corners of a discontinuous diamond-like surface-motif of positive charges, embedded in an extended surface patch,bisected into a slightly polar side and a hydrophobic side by theR3-K6 joining vector~Fig. 10!. By removal of R3 and K6, thelocal hydrophobic folding balance is destroyed due to the loss offavorable charge-solvation and the simultaneous onset of unfavor-able hydrophobic-solvation. Together with the latter contribution,additional destabilization and aggregation propensity may also arisefrom the removal of a Pro residue~Pro5! occurring in the N-terminalhexapeptide, as demonstrated in beta peptide causing Alzheimerdisease~Wood et al., 1995!. It is worth noting that the hydrophobicregion proximal to the N-terminal hexapeptide ofb2-m ~on the leftin Fig. 10! contains the portion of strand V that we propose to beinvolved in aggregation. While full-lengthb2-m can make fibrilsin vitro even at physiological pH~Connors et al., 1985; Stoppiniet al., 1997!, the fibril yield is very low and very high proteinconcentrations with very low ionic strength are required. Underthese conditions of nonphysiological solvation and high proteindensity, it is likely that intermolecular interactions can occur which
orient and organize the oligomer into a fibrillar structure. Further-more, it is noteworthy that at pH conditions~between pH 2.0 and3.0! in which full-lengthb2-m forms fibrils with the highest yield,the protein is largely unfolded~G. Esposito, R. Michelutti, & G.Verdone, unpubl. data!. The present mechanism of conformationalmodifications leading to fibrillogenesis is in good agreement withthe models proposed for amyloidogenic species of lysozyme~Boothet al., 1997; Canet et al., 1999! and TTR~Kelly, 1997; Nettletonet al., 1998! in which a partially unfolded state populates a key stepof the fibrillogenic pathway.
Materials and methods
Production of recombinantb2-m
b2-m cDNA was obtained from the plasmid pHN1~a gift of Dr. J.Bell!. The expression cassette for the full-lengthb2-m was createdby using PCR withb2-m cDNA as the template and the oligonu-cleotide primers 59CATATGATCCAGCGTACTCCAAAG for thefull-length and 59CATATGATTCAGGTTTACTCACGT for thetruncated species and 39GCCGGATCCTTACATGTCTCGATC
Fig. 10. Electrostatic surface ofb2-m generated by the program GRASP~Nicholls et al., 1991!. The four blue spots correspond to theside chains of R3, K6~explicitly labeled!, and K58~left! and K91~right!; the red surface near K58 is the carboxylate of D59.
842 G. Esposito et al.

CCAC for both species and containing the underlined NdeI restric-tion sites at the 59 and the BamHI restriction site at the 39. Theamplification mixture~40mL! contained dGTP, dATP, dTTP, dCTP~each 200mM !, oligonucleotide primers~1 mM!, template DNA~10 ng!, Thermus aquaticus~Taq! DNA polymerase~Pharmacia,Uppsala, Sweden! in 40 mM NaCl, 10 mM MgCl2 50 mM Tris-HCl, pH 9. The reaction mixture were subjected to 2 cycles of948C for 1 min, 458C for 1 min, 728C for 1 min and 30 cycles inwhich the annealing was obtained at 508C. A final incubation at728C for 10 min ended the cycles. The amplified DNA was gelpurified, digested with BamHI and NdeI, and ligated into pet 11avector that has been digested with the same enzymes. TheEsch-erichia coli strain BL21DE3 was transformed with the plasmidsand clones containing inserts and producing protein upon induc-tion with isopropylb-d-thiogalactopyranoside~IPTG! were iden-tified. The correct nucleotide sequence of the insert was confirmedby DNA sequencing. A methionine residue was always present atthe N-terminal position of the recombinant products that will bereferred to as M0.
Protein reconstitution and purification
One liter of cells transformed with either the full-lengthb2-m andDN6b2-m was incubated at 378C and induced to express the pro-tein by addition of 1 mM IPTG. The cells were harvested bycentrifugation and the cell pellets containing the recombinantb2-mas inclusion bodies were resuspended in 10 mM Tris HCl, pH 8~20 mL! containing lysozyme at 100mg0mL and phenylmethyl-sulfonyl fluoride ~PMSF! at 50mg0mL. The cells were lysed bysonication and then centrifuged~10,0003 g! for 20 min. The pelletcontaining recombinant protein was washed with 10 mM Tris HCl,pH 8 ~20 mL!, dissolved in 100 mM Tris HCl, pH 806 M GdnHCl~10 mL!, and centrifuged~100,0003 g ! for 1 h at 48C. Thesupernatant was applied to a Sephacryl S300 column equilibratedwith the 6 M GdnHCl buffer. The identification of chromato-graphic fractions containingb2-m was accomplished by ELISA.b2-m isolated from gel filtration was then dialyzed against 10 mMTris HCl, pH 8 and further purified by ion-exchange chromatog-raphy with an UNO column~Bio-Rad, Hercules, California!. Elec-trospray mass spectrometry and N-terminal amino acid sequencingwere used to confirm the identity of the recombinant proteins andcompared with wild-typeb2-m. The experimentally determinedaverage molecular masses were 11,136.2 and 11,860.3 Da andconsistent with the calculated masses of 11,137.5 Da forDN6b2-mand 11,861.4 Da forb2-M with an N-terminal methionine residue.
CD spectra
Circular dichroism~CD! measurements were performed with aJasco 710 spectropolarimeter. Measurements were performed at208C in cells with 0.1 cm pathlength in the far-UV~190–250 nm!at a protein concentration of 4.2mM and in cells with 1 cm path-length in the near-UV~250–310 nm! at a protein concentration of42 mM. The ellipticity was recorded every 1 s with a response timeof 1 s and a bandwidth of 2 nm. Ellipticities in deg cm2 dmol21
were expressed as~Q!.
Gel filtration assay
Gel filtration experiments were performed with an HPLC system~Hewlett Packard HP1100! using a Phenomenex Biosep-Sec-S 2000
column~3003 7.50 mm! equilibrated and eluted in 15 mM phos-phate buffer, pH 7.5, containing 12 mM NaCl. Samples ofb2-mandDN6b2-m ~500 mg! were dissolved in an appropriate volumeof buffer to a final concentration of either 1 mM or 10mM. Allexperiments were carried out at 258C with a flow rate of 1 mL0min. The column was calibrated under the same conditions withproteins of known molecular mass: bovine serum albumin~66kDa!, lysozyme~14.3 kDa!, and insulin~5.7 kDa!.
Determination of stability
Thermodynamic stability ofb2-m was determined by monitoringthe red shift oflmax and the dependence of intrinsic tryptophanfluorescence on GdnHCl concentration. The protein was dissolvedat a concentration of 5mM in 10 mM potassium phosphate frompH 4 to 8. Excitation was at 295 nm and emission monitored in therange 300–500 nm~Tsunenaga et al., 1987!. The transition curvesobtained by increasing the concentration of GdnHCl were analysedas suggested by Finn et al.~1992!.
Light scattering
Light scattering measurements were performed in a Perkin ElmerLS50 spectrofluorimeter equipped with a temperature-regulatedcell holder and a temperature-controlled system. Aggregation wasmonitored at different pH values. Samples, at a concentration of40 mM in 0.1 M potassium phosphate buffer at pH 4–7, wereexcited at 510 nm and the scattered light was detected at 908 to theincident beam; the spectral band was 2.5 nm.
Electron microscopy
Samples for transmission electron microscopy were prepared byfloating the protein suspension on formvar0carbon coated grids for2–3 min, then air dried and stained with 2% uranyl acetate. Sam-ples were examined in a Jeol JEM 1200 EX electron microscopeoperating at 80 kV.
Fibrillogenesis: Effect of pH
The experiments were performed according to the method of Naikiet al.~1997!. The buffers were: 50 mM HCl citrate buffer~pH 1.5,2, 3!, 50 mM phosphate buffer~pH 4, 5, 6, 7!, and 50 mM Tris HClfor pH 8. All the samples contained 100 mM NaCl. Ex vivob2-mfibrils were added to the reaction mixture in order to obtain a finalconcentration of 25mg0mL. Full-lengthb2-m andDN6b2-m wereat a concentration of 1 mg0mL.
Thioflavine assay
Quantitation of amyloid fibril formation was performed with themethod described by LeVine~1993!. Thioflavine concentration was5 mM and the buffer used was 50 mM glycine NaOH, pH 8.5.Measurements were made using a LS50 Perkin Elmer spectroflu-orometer with excitation at 455 nm and emission was collected at485 nm. The slits were set at 5 nm.
Hydrogen exchange MS
Both recombinant protein samples were washed three times withwater at pH 7.0 using SR3 centricon concentrators~Amicon Inc.,
Role of truncated species ofb2-m in fibrillogenesis 843

Beverly, Massachusetts! to give final concentrations of;160and 250mM for b2-m andDN6b2-m, respectively. To initiatethe hydrogen exchange reaction, 1mL aliquots of each proteinsolution were mixed and diluted into 48mL D2O ~Apollo Sci-entific Ltd., Whaley Bridge, UK! at pH 7 ~uncorrected pH meterreading! to give concentrations of 3mM ~b2-m! and 5 mM~DN6b2-m!. The total number of exchangeable sites for recom-binant b2-m andDN6b2-m are 189 and 175, respectively: fromthe level of H2O present in the exchange solution~4%!, fully-exchangedb2-m andDN6b2-m would correspond to 181.4 and168 protected sites, respectively. All data were acquired in pos-itive ion mode using a Platform II mass spectrometer~Micro-mass, Manchester, United Kingdom! configured for nanoflowelectrospray. Sample aliquots for each time-point were loadedinto gold-coated borosilicate glass capillaries~0.5 mm ID, ClarkeElectromedical Instruments, Reading, United Kingdom!, whichhad been drawn out and cut to give a tip ID of,10 mm.Sample flow into the source was initiated by applying a back-pressure of nitrogen to the sample solution. Source conditionswere optimized forb2-m: the same capillary and cone voltageswere used for all time points~capillary 1 kV, cone 80 V!, anddesolvation was achieved using nitrogen gas at room temperature.
Time points were taken every 3 min for the first 20 min, fol-lowed by sampling at 6, 12, and 30 min as the exchange proceeded.Spectra were acquired in continuum mode for 1 min over the massrange 960–1,560m0z at a scan rate of 3 s0scan. For each timepoint, the masses of the two protein were calculated as an averageof the masses determined from charge states~18 to 112 for b2-mand 18 to 111 for DN6b2-m!. The instrument was calibratedusing sodium iodide at unit resolution. Data processing parameterswere the same for all spectra~background subtraction, smoothing,and peak centroid!.
Limited proteolysis MS
Both b2-m andDN6b2-m were digested with trypsin, endoprote-ase AspN, endoprotease V8, chymotrypsin, proteinase K, and elas-tase. Enzymatic hydrolysis was performed at 378C in 100 mMsodium phosphate, pH 7.5, using enzyme-to-substrate ratios rang-ing between 1:5 and 1:700. The extent of proteolysis was moni-tored by sampling the incubation mixture at different time intervals.Proteolytic fragments were separated by reversed-phase HPLC ona Phenomenex Jupiter C18 column~250 3 2.1 mm, 300 Å poresize! with a linear gradient from 5 to 50% acetonitrile in 0.1%trifluoroacetic acid~TFA! over 40 min, at a flow rate of 200mL 0min. Peaks were monitored at 220 and 280 nm, and the individualfractions were collected manually. Proteolytic fragments were an-alyzed by electrospray MS using either a BIO-Q triple quadrupolemass spectrometer~Micromass! or an API-100 single quadrupoleinstrument~Perkin Elmer, Foster City, California; PE Biosystems,Foster City, California!. Samples were injected directly into the ionsource via a loop injection at a flow rate of 5mL 0min. Data wereacquired and processed using either Mass-Lynx~Micromass! orBiomultiviewer~Perkin Elmer; PE Biosystems! software. The cal-ibrant was horse heart myoglobin~average molecular mass 16,951.5Da!; all masses are reported as average mass.
When necessary, identification of disulfide-bridged intermedi-ates was carried out by reduction of samples with 10 mM dithio-threitol in 50 mM Tris HCl, 1 mM EDTA, pH 7.5, containing6 M GdnHCl, for 2 h at 378C under nitrogen, followed by analy-sis by MALDI mass spectrometry~MALDI 0MS! using a Voyager
DE instrument~Perkin Elmer; PE Biosystems!. Typically, 1 mLof analyte solution was mixed with 1mL of a-cyano-4-hydroxycinammic acid 10 mg0mL in acetonitrile00.2% TFA, 70:30v0v, containing 250 fmol of bovine insulin. The mixture was ap-plied onto the metallic sample plate and air dried. Mass calibrationwas performed using insulin~m0z 5,734.5! and a matrix peak atm0z 379.1. All mass values are reported as average masses.
NMR spectroscopy
1H-NMR spectra were obtained with a Bruker Avance 500 NMRspectrometer at a frequency of 500.13 MHz. Aqueous solutionswere prepared by adding;5 mg of the lyophilyzed samples to450 mL 50 mM perdeuterated ammonium acetate in H2O0D2O~90010!; the solution pH values were 6.6 and 6.3 forb2-m andDN6b2-m, respectively. Freshly prepared protein solutions~0.9 mM!were submitted to 2D NMR experiments after equilibrating theprobe temperature at 378C. For each sample, the measurementtime was;15 h and control 1D spectra were recorded before andafter 2D data acquisition to monitor possible spectral changes oc-curring during data collection. Two-dimensional TOCSY~Braun-schweiler & Ernst, 1983! and 2D NOESY~Jeener et al., 1979!spectra were acquired for both protein samples. The adopted ex-perimental schemes included solvent suppression by pulsed gra-dient field spin-echo~WATERGATE, Piotto et al., 1992!, 1 s steadystate recovery time, 50 and 100 ms mixing time~tm!, respectively,for TOCSY and NOESY, TPPIt1 quadrature detection~Marion &Wüthrich, 1983!. The spin-lock mixing of the TOCSY experimentwas obtained with an MLEV17 pulse train~Bax & Davis, 1985! atgB202p 5 9.4 kHz. The acquisitions were performed over a spec-tral width of 7,002.801 Hz in both dimensions with matrix arraysize of 2,0483 512 in t2 andt1, respectively, and 32–64 scans0t1FID. Data processing and analysis were performed using Felix~MSI, San Diego, California! software. All spectra were refer-enced by comparison with the data reported by Okon et al.~1992!for aqueous solution ofb2-m, obtained under similar conditions asthose adopted in the present study. In particular, the L23 CdH3
resonance at20.58 ppm was chosen as a reference peak for thespectra.
Molecular dynamics simulations
MD simulations were performed using DYANA 1.4~Güntert et al.,1997! software following the default protocol for simulated an-nealing~4,000 steps MD, 1,000 conjugate gradients minimizationsteps!. Molecular structures were generated with InsightII~MSI,San Diego, California! and MOLMOL ~Koradi et al., 1996!.
Supplementary material in the Electronic Appendix
1H NMR chemical shifts ofDN6b2-m andb2-m ~Table 1! anddifferences betweenb2-m crystal structure and NMR data.
Acknowledgments
We would like to thank Dr. Lia Asti of Centro Grandi Strumenti Universitàdi Pavia and Dr. Patricia Arcidiaco. This study was supported by Ministerodell Università e della Ricerca Scientifica~PRIN project: Protein Foldingand misfolding!, Università di Pavia~Progetto d’Ateneo!, CNR, Telethon~Grant E. 793!, the Royal Society and Oxford Centre for Molecular Sciences.
844 G. Esposito et al.

References
Bax A, Davis DG. 1985. MLEV-17 based two dimensional homonuclear mag-netization transfer spectroscopy.J Mag Reson 65:355–360.
Bellotti V, Stoppini M, Mangione P, Sunde M, Robinson CV, Asti L, BrancaccioD, Ferri G. 1998.b2-microglobulin can be refolded into a native state fromex vivo amyloid fibrils.Eur J Biochem 258:61–67.
Bjorkman PJ, Saper MA, Samaroui B, Bennet WS, Strominger JL, Wiley DC.1987. Structure of the human class I histocompatibility antigen, HLA-A2.Nature 329:506–512.
Booth D, Sunde M, Bellotti V, Robinson CV, Hutchinson WL, Fraser P, Hawk-ins PN, Dobson CM, Radford SE, Blake CCF, Pepys M. 1997. Instability,unfolding and aggregation of human lysozyme variants underlying amyloidfibrillogenesis.Nature 385:787–793.
Botto M, Hawkins PN, Bickerstaff MC, Herbert J, Bygrave AE, McBride A,Hutchinson WL, Tennent GA, Walport MJ, Pepys MB. 1997. Amyloiddeposition is delayed in mice with targeted deletion of the serum amyloid Pcomponent gene. Nat Med 8:855–859.
Braunschweiler L, Ernst RR. 1983. Coherence transfer by isotropic mixing:Application to proton correlation spectroscopy.J Magn Reson 53:521–528.
Canet D, Sunde M, Last AM, Miranker A, Spencer A, Robinson C, Dobson CM.1999. Mechanistic studies of the folding of human lysozyme and the originof amyloidogenic behavior in its disease related variants.Biochemistry38:6419–6427.
Capeiller-Blandin C, Delaveau T, Descamps-Latscha B. 1991. Structural mod-ifications of humanb2-microglobulin treated with oxygen-derived radicals.Biochem J 277:175–182.
Connors LH, Shirahama T, Skinner M, Fenves A, Cohen AS. 1985. In vitroformation of amyloid fibrils from intact beta2-microglobulin.Biochem Bio-phys Res Commun 131:1063–1068.
Finn BE, Chen X, Jennings PA, Saalau-Bethell SM. 1992. Principles of proteinstability. Part 1: Reversible unfolding of protein: Kinetic and thermo-dynamic analysis. In: Rees AR, Stenderberg MJE, Wetzel R, eds.Proteinengineering. Oxford: IRL Oxford University Press. pp 167–189.
Gejyo F, Yamada T, Odani S, Nakagawa Y, Arakawa M, Kunitomo T, KataokaH, Suzuki M, Hirasawa Y, Shirahama T, et al. 1985. A new form of amyloidprotein associated with hemodialysis was identified asb2-microglobulin.Biochem Biophys Res Commun 129:701–706.
Güntert P, Mumenthaler C, Wüthrich K. 1997. Torsion angle dynamics for NMRstructure calculation with the new program DYANA.J Mol Biol 273:283–298.
Hooke SD, Eyles SJ, Miranker A, Radford SE, Robinson CV, Dobson CM.1995. Co-operative elements in protein folding monitored by electrosprayionization mass spectrometry. J Am Chem Soc 117:7548–7549.
Isenman DE, Painter RH, Dorrington KJ. 1975. The structure and function ofimmunoglobulin domains: Studies withb2-microglobulin on the role of theintrachain disulfide bond.Proc Natl Acad Sci USA 72:548–552.
Jeener J, Meier BH, Bachmann P, Ernst RR. 1979. Investigation of exchangeprocesses by two-dimensional NMR spectroscopy.J Chem Phys 71:286–292.
Kelly JW. 1997. Amyloid fibril formation and protein misassembly: A structuralquest for insights into amyloid and prion disease.Structure 5:595–600.
Koradi R, Billeter M, Wüthrich K. 1996. MOLMOL, a program for display andanalysis of macromolecular structure.J Mol Graphics 14:51–55.
LeVine H. 1993. Thioflavine T interaction with synthetic Alzheimer’s diseaseb-amyloid peptide: Detection of amyloid aggregation in solution.ProteinSci 2:404–410.
Linke RP, Bommer J, Ritz E, Waldherr R, Eulitz M. 1986. Amyloid kidneystones of uremic patients consist of beta 2-microglobulin fragments.Bio-chem Biophys Res Commun 136:665–671.
Linke RP, Hampl H, Bartel-Schwarze S, Eulitz M. 1987. Beta 2-microglobulin,different fragments and polymers thereof in synovial amyloid in long-termhemodialysis.Biol Chem Hoppe Seyler 368:137–144.
Linke RP, Hampl H, Lobeck H, Ritz E, Bommer J, Waldherr R, Eulitz M. 1989.Lysine-specific cleavage ofb2-microglobulin in amyloid deposits associ-ated with haemodialysis.Kidney Int 36:675–681.
Marion D, Bax A. 1989. Baseline correction of 2D FT NMR spectra using asimple linear prediction extrapolation of the time-domain data.J MagnReson 83:205–211.
Marion D, Ikura M, Bax A. 1989. Improved solvent suppression in one- andtwo-dimensional NMR spectra by convolution of time-domain data.J MagnReson 84:425–430.
Marion D, Wüthrich K. 1983. Application of phase sensitive two-dimensionalcorrelated spectroscopy~COSY! for measurements of proton-proton spin-
spin coupling constants in proteins.Biochem Biophys Res Commun 113:967–974.
Markley JL, Bax A, Arata Y, Hilbers CW, Kaptein R, Sykes BD, Wright PE,Wüthrich K. 1998. Recommendations for the presentation of NMR struc-tures of proteins and nucleic acids. IUPAC-IUBMB-IUPAB Inter-UnionTask Group on the standardization of data bases of protein and nucleic acidstructures determined by NMR spectroscopy.J Biomol NMR 12:1–23.
Merlini G, Ascari E, Amboldi N, Bellotti V, Arbustini E, Perfetti V, Ferrari M,Zorzoli I, Marinone MG, Garini P, et al. 1995. Interaction of the anthra-cycline 49-iodo-49-deoxydoxorubicin with amyloid fibrils: Inhibition of am-yloidogenesis.Proc Natl Acad Sci USA 92:2959–2963.
Miyata T, Oda O, Inagi R, Iida Y, Araki N, Yamada N, Horiuchi S, Taniguchi N,Maeda K, Kinoshita T. 1993.b2-microglobulin modified with advancedglycation end products is a major component of haemodialysis-associatedamyloidosis.J Clin Invest 92:1243–1252.
Naiki H, Hashimoto N, Suzuki S, Kimura H, Nakakuki K, Gejyo F. 1997.Establishment of a kinetic model of dialysis-related amyloid fibril extensionin vitro. Amyloid Int J Exp Clin Invest 4:223–232.
Nettleton EJ, Sunde M, Lai Z, Kelly JW, Dobson CM, Robinson CV. 1998.Tetramer to monomer dissociation and structural integrity of amyloidogenictransthyretins—Evidence from electrospray mass spectrometry.J Mol Biol281:553–564.
Nicholls A, Sharp K, Honig B. 1991. Protein folding and association: Insightsfrom the interfacial and thermodynamic properties of hydrocarbons.Pro-teins 11:281–296.
Okon M, Bray P, Vucelic D. 1992. 1H NMR assignments and secondary struc-ture of humanb2-microglobulin in solution.Biochemistry 31:8906–8915.
Orrù S, Dal Piaz F, Casbarra A, Biasol G, De Francesco R, Steinkuhler C, PucciP. 1999. Conformational changes in the NS3 protease from hepatitis C virusstrain Bk monitored by limited proteolysis and mass spectrometry.ProteinSci 8:1445–1454.
Piotto M, Saudek V, Sklenar V. 1992. Gradient-tailored excitation for single-quantum NMR spectroscopy of aqueous solutions.J Biomol NMR 2:661–665.
Radford SE, Dobson CM. 1999. From computer simulations to human disease:Emerging themes in protein folding.Cell 97:291–298.
Scaloni A, Miraglia N, Orrù S, Amodeo P, Motta A, Marino G, Pucci P. 1998.Topology of calmodulin-melittin complex.J Mol Biol 277:945–958.
Scaloni A, Monti M, Acquaviva R, Tell G, Damante G, Formisano, Pucci P.1999. Topology of the thyroid transcription factor 1 homeodomain-DNAcomplex.Biochemistry 38:64–72.
Stoppini M, Arcidiaco P, Mangione P, Giorgetti S, Brancaccio D, Bellotti V.2000. Detection of fragments ofb2-microglobulin in amyloid fibrils.Kid-ney Int 57:349–350.
Stoppini M, Bellotti V, Mangione P, Merlini G, Ferri G. 1997. Use of anti-beta2 microglobulin mAb to study formation of amyloid fibrils.Eur J Biochem249:21–26.
Sunde M, Blake CCF. 1998. From the globular to the fibrous state: Proteinstructure and structural conversion in amyloid formation.Quart Rev Bio-phys 31:1–39.
Sunde M, Serpell LC, Bartlam M, Fraser PE, Pepys MB, Blake CCF. 1997.Common core structure of amyloid fibrils by synchrotron X-ray diffraction.J Mol Biol 273:729–739.
Tennent GA, Lovat LB, Pepys MB. 1995. Serum amyloid P component preventsproteolysis of the amyloid fibrils of Alzheimer disease and systemic amy-loidosis.Proc Natl Acad Sci USA 92:4299–4303.
Tsunenaga M, Goto Y, Kawata Y, Hamaguchi K. 1987. Unfolding and refoldingof a type kappa immunoglobulin light chain and its variable and constantfragments.Biochemistry 26:6044–6051.
Wishart DS, Bigam CG, Holm A, Hodges RS, Sykes BD. 1995.1H, 13C and15Nrandom coil chemical shifts of common amino acids. I. Investigations ofnearest-neighbor effects.J Biomol NMR 5:67–81.
Wishart DS, Sykes BD. 1994. Chemical shifts as a tool for structure determi-nation.Methods Enzymol 239:363–392.
Wood SJ, Wetzel R, Martin JD, Hurle MR. 1995. Prolines and amyloidogenicityin fragments of the Alzheimer’s peptideb0A4. Biochemistry 34:724–730.
Wüthrich K. 1986.NMR spectroscopy of proteins and nucleic acids. New York:Wiley & Sons.
Zappacosta F, Pessi A, Bianchi E, Ingallinella P, Scaloni A, Venturini S, SollazzoM, Tramontano A, Marino G, Pucci P. 1996. Probing the tertiary stucture ofproteins by limited proteolysis and mass spectrometry: The case of Mini-body.Protein Sci 5:802–813.
Role of truncated species ofb2-m in fibrillogenesis 845
![Selective and high affinity labeling of neuronal and recombinant nociceptin receptors with the hexapeptide radioprobe [3H]Ac-RYYRIK-ol](https://static.fdokumen.com/doc/165x107/633741c44554fe9f0c05ba75/selective-and-high-affinity-labeling-of-neuronal-and-recombinant-nociceptin-receptors.jpg)



















