
Estrogen receptor β2 is inversely correlated with Ki-67 in hyperplastic and noninvasive neoplastic...
10
1 3 J Cancer Res Clin Oncol (2014) 140:1057–1066 DOI 10.1007/s00432-014-1652-0 ORIGINAL ARTICLE – CLINICAL ONCOLOGY Estrogen receptor β2 is inversely correlated with Ki‑67 in hyperplastic and noninvasive neoplastic breast lesions Νiki Ι. Chantzi · Marina Palaiologou · Artemis Stylianidou · Nikos Goutas · Stamatis Vassilaros · Helen P. Kourea · Eugen Dhimolea · Dimitra J. Mitsiou · Dina G. Tiniakos · Μichael N. Alexis Received: 20 January 2014 / Accepted: 15 March 2014 / Published online: 27 March 2014 © Springer-Verlag Berlin Heidelberg 2014 in the whole set of cases and with high ERα expression and low ERβ2 expression in the postmenopausal cases of the subset that is generated by excluding AM and high-grade DCIS. Postmenopausal status correlated with low ERβ1 expression in the whole set and with higher than normal Ki-67 LI, high ERα expression and low ERβ1 expression in the subset. Conclusions These findings are in accordance with an ERα-opposing oncosuppressive role of ERβ2 in mammary carcinogenesis along the HUT-AH-DCIS1/2 pathway. Keywords Breast · Hyperplasia · Ductal carcinoma in situ · ERβ1 · ERβ2 · Κi-67 Abbreviations ERα Estrogen receptor α ERβ Estrogen receptor β ERβ1 Estrogen receptor β1 ERβ2 Estrogen receptor β2 HUT Hyperplasia of usual type AM Apocrine metaplasia Abstract Purpose The purpose of the study is to compare expres- sion levels of ΕRα, ERβ1, ERβ2 and cell proliferation marker Ki-67 in normal breast and hyperplastic and nonin- vasive neoplastic breast lesions. Materials and methods Routinely processed breast tis- sue from 55 patients provided 65 cases of noninvasive lesions, namely, epithelial hyperplasia of usual type (HUT), apocrine metaplasia (AM), atypical hyperplasia (AH) and ductal carcinoma in situ (DCIS) and 14 cases of adjacent normal breast tissue. Expression of ERα, ERβ1 and ERβ2 were evaluated using immunohistochemistry and correlated with Ki-67 labeling index (Ki-67 LI) and menopausal sta- tus of the patients. Results Compared with normal breast, ERα expression increased in low to intermediate-grade DCIS (DCIS1/2) and tended to decrease in high-grade DCIS, while ERβ1 expression decreased in DCIS irrespective of grade. Mean Ki-67 LI in HUT, low to intermediate-grade DCIS and high-grade DCIS was higher than in normal breast. Higher than normal Ki-67 LI correlated with low ERβ2 expression Ν. Ι. Chantzi · M. Palaiologou · D. G. Tiniakos (*) Laboratory of Histology and Embryology, Medical School, National and Kapodistrian University of Athens, 75 M. Asias str, Goudi, 11527 Athens, Greece e-mail: [email protected] Ν. Ι. Chantzi · E. Dhimolea · D. J. Mitsiou · Μ. N. Alexis (*) Institute of Biology, Medicinal Chemistry and Biotechnology, National Hellenic Research Foundation, 48, Vassileos Constantinou Av, 11635 Athens, Greece e-mail: [email protected] A. Stylianidou Histopathology Department, “Helena Venizelou” General and Maternity District Hospital, Athens, Greece N. Goutas Department of Pathology, Evgenidio Infirmary, Athens, Greece S. Vassilaros “Prolipsis” Diagnostic Center, Athens, Greece H. P. Kourea Department of Pathology, Medical School, University of Patras, Patras, Greece D. G. Tiniakos Institute of Cellular Medicine, Faculty of Medical Sciences, Newcastle University, Newcastle upon Tyne, UK
Transcript of Estrogen receptor β2 is inversely correlated with Ki-67 in hyperplastic and noninvasive neoplastic...

1 3
J Cancer Res Clin Oncol (2014) 140:1057–1066DOI 10.1007/s00432-014-1652-0
ORIgInal aRtICle – ClInICal OnCOlOgy
Estrogen receptor β2 is inversely correlated with Ki‑67 in hyperplastic and noninvasive neoplastic breast lesions
Νiki Ι. Chantzi · Marina Palaiologou · Artemis Stylianidou · Nikos Goutas · Stamatis Vassilaros · Helen P. Kourea · Eugen Dhimolea · Dimitra J. Mitsiou · Dina G. Tiniakos · Μichael N. Alexis
Received: 20 January 2014 / accepted: 15 March 2014 / Published online: 27 March 2014 © Springer-Verlag Berlin Heidelberg 2014
in the whole set of cases and with high eRα expression and low eRβ2 expression in the postmenopausal cases of the subset that is generated by excluding aM and high-grade DCIS. Postmenopausal status correlated with low eRβ1 expression in the whole set and with higher than normal Ki-67 lI, high eRα expression and low eRβ1 expression in the subset.Conclusions these findings are in accordance with an eRα-opposing oncosuppressive role of eRβ2 in mammary carcinogenesis along the HUt-aH-DCIS1/2 pathway.
Keywords Breast · Hyperplasia · Ductal carcinoma in situ · eRβ1 · eRβ2 · Κi-67
AbbreviationseRα estrogen receptor αeRβ estrogen receptor βeRβ1 estrogen receptor β1eRβ2 estrogen receptor β2HUt Hyperplasia of usual typeaM apocrine metaplasia
Abstract Purpose the purpose of the study is to compare expres-sion levels of ΕRα, eRβ1, eRβ2 and cell proliferation marker Ki-67 in normal breast and hyperplastic and nonin-vasive neoplastic breast lesions.Materials and methods Routinely processed breast tis-sue from 55 patients provided 65 cases of noninvasive lesions, namely, epithelial hyperplasia of usual type (HUt), apocrine metaplasia (aM), atypical hyperplasia (aH) and ductal carcinoma in situ (DCIS) and 14 cases of adjacent normal breast tissue. expression of eRα, eRβ1 and eRβ2 were evaluated using immunohistochemistry and correlated with Ki-67 labeling index (Ki-67 lI) and menopausal sta-tus of the patients.Results Compared with normal breast, eRα expression increased in low to intermediate-grade DCIS (DCIS1/2) and tended to decrease in high-grade DCIS, while eRβ1 expression decreased in DCIS irrespective of grade. Mean Ki-67 lI in HUt, low to intermediate-grade DCIS and high-grade DCIS was higher than in normal breast. Higher than normal Ki-67 lI correlated with low eRβ2 expression
Ν. Ι. Chantzi · M. Palaiologou · D. g. tiniakos (*) laboratory of Histology and embryology, Medical School, national and Kapodistrian University of athens, 75 M. asias str, goudi, 11527 athens, greecee-mail: [email protected]
Ν. Ι. Chantzi · e. Dhimolea · D. J. Mitsiou · Μ. n. alexis (*) Institute of Biology, Medicinal Chemistry and Biotechnology, national Hellenic Research Foundation, 48, Vassileos Constantinou av, 11635 athens, greecee-mail: [email protected]
a. Stylianidou Histopathology Department, “Helena Venizelou” general and Maternity District Hospital, athens, greece
n. goutas Department of Pathology, evgenidio Infirmary, athens, greece
S. Vassilaros “Prolipsis” Diagnostic Center, athens, greece
H. P. Kourea Department of Pathology, Medical School, University of Patras, Patras, greece
D. g. tiniakos Institute of Cellular Medicine, Faculty of Medical Sciences, newcastle University, newcastle upon tyne, UK

1058 J Cancer Res Clin Oncol (2014) 140:1057–1066
1 3
aH atypical hyperplasiaDCIS Ductal carcinoma in situDCIS1/2 low to intermediate-grade DCISDCIS3 High-grade DCISIHC ImmunohistochemistryΚi-67 lI Ki-67 labeling index
Introduction
Cancer development is the result of genetic alterations that confer proliferative and survival advantages to the trans-formed cell (Hanahan and Weinberg 2011). Based on the number and pattern of genetic alterations, breast cancer development is thought to reflect accumulation of changes leading from hyperplasia with or without atypia through carcinoma in situ to invasive carcinoma (lopez-garcia et al. 2010). HUt and cysts with aM are considered as risk factors of breast cancer development, while atypical ductal hyperplasia (aDH) and low-grade ductal carcinoma in situ (DCIS) are considered as non-obligate precursors of low-grade invasive ductal carcinoma (IDC). High-grade DCIS involves genetic alterations distinct from those of low-grade DCIS and is thought to arise de novo or from lesions other than atypical ductal hyperplasia (aDH) or low-grade DCIS and to be a non-obligate precursor of high-grade IDC (lopez-garcia et al. 2010).
the majority of human breast tumors arises from transformed ductal epithelial cells and displays estrogen-dependent growth predominantly regulated by the two subtypes of estrogen receptor (eR), eRα and eRβ, which are products of different genes (thomas and gustafsson 2011). although these receptors are structurally simi-lar, they are endowed with quite different functions and, while they are co-expressed in normal breast, expression of one in the absence of the other is commonly observed in benign breast lesions and breast tumors (Roger et al. 2001; Shaaban et al. 2003; Skliris et al. 2006). eRα is the principal regulator of estrogen-dependent proliferation of breast epithelial cells and has a key role in breast cancer prognosis and treatment (Patani et al. 2013; thomas and gustafsson 2011). Breast carcinomas are usually classified as eRα-positive or negative, largely encompassing low- and high-grade carcinomas of good and poor prognosis, respectively; similarly, eRα expression is most common in low-grade DCIS and least common in high-grade DCIS (Claus et al. 2001; lopez-garcia et al. 2010; Roger et al. 2001). In postmenopausal women, high eRα expression in normal breast epithelium is associated with increased breast cancer risk (Khan et al. 1998), probably as a result of increased accumulation of random genetic errors due to increased eRα-dependent proliferation of epithelial cells (Henderson and Feigelson 2000). accordingly, in
HUt, breast cancer risk is associated with high expres-sion of eRα and the cell proliferation marker Ki-67 (Shaaban et al. 2002). Ki-67 lI is considered an alterna-tive to mitotic index and in breast carcinoma is associated with eRα-negativity, high nuclear grade and poor progno-sis (Patani et al. 2013; van Diest et al. 2004). Response of invasive eRα-positive breast carcinoma to endocrine ther-apy has been associated with a decrease in Ki-67 expres-sion (Dowsett et al. 2009).
eRβ expression in human breast is located in stro-mal, myoepithelial and luminal epithelial cells (Roger et al. 2001); it comprises five different isoforms, namely eRβ1-5, products of alternative splicing of the last cod-ing exon of eRβ mRna (leung et al. 2006; Moore et al. 1998; Ogawa et al. 1998). all five isoforms share the same primary structure up to amino acid 468 but differ in their C-termini. the 530 amino acids-long eRβ1 is the longest of the five isoforms and the only one capable of binding 17β-estradiol (estradiol) with high affinity. eRβ1 and eRβ2 are the isoforms more frequently expressed in breast cancer and therefore more extensively studied so far (leung et al. 2012; Murphy and leygue 2012; thomas and gustafsson 2011). although eRβ2 is unable to bind estra-diol, it is known to heterodimerize with eRα and eRβ1, thereby inhibiting and enhancing, respectively, their estro-gen-dependent transcriptional activity (leung et al. 2006; Peng et al. 2003). In eRα-positive breast cancer cells, eRβ1 and eRβ2 display antiproliferative action (Ogawa et al. 1998; Paruthiyil et al. 2004; Ström et al. 2004; Zhao et al. 2007).
the clinical significance of eRβ expression in breast cancer has not been resolved so far (Patani et al. 2013). Using isoform-blind or inadequately validated antibodies or applying methodologies (e.g., PCR) that allow interfer-ence from eRβ expression in non-tumor cells may have seriously jeopardized assessment of the clinical signifi-cance of the differential expression of the isoforms (Skliris et al. 2008; Snyder et al. 2010). the clinical significance of eRβ1 and eRβ2 may differ depending on the type of mam-mary lesion (leygue and Murphy 2013). In eRα-positive breast cancer, eRβ1 and eRβ2 have been associated with response to endocrine therapy and longer disease-free and overall survival (Honma et al. 2008; nakopoulou et al. 2004; Palmieri et al. 2004; Shaaban et al. 2008). However, in eRα-negative breast cancer, eRβ1 expression was asso-ciated with response to tamoxifen (Honma et al. 2008) and eRβ2 with poor disease prognosis (Chantzi et al. 2013).
the role of eRβ in breast cancer development has been the focus of very few studies. Immunohistochemi-cal expression of eRα and eRβ (i.e., the sum of eRβ1-5 as assessed using antibodies unable to differentiate between the five isoforms) is known to increase and decrease, respectively, during mammary carcinogenesis (Roger et al.

1059J Cancer Res Clin Oncol (2014) 140:1057–1066
1 3
2001; Shoker et al. 1999). the decline in eRβ expression was found to inversely correlate with Ki-67 expression in high-grade DCIS (Roger et al. 2001), in accordance with the proposed tumor suppressor function of eRβ (Bardin et al. 2004; thomas and gustafsson 2011). Similarly, it has been reported that eRβ1 expression decreases during mam-mary carcinogenesis (Shaaban et al. 2003) and that cases of HUt with a low ratio of eRβ1 expression to eRα expres-sion (eRβ1/eRα expression) are more likely to progress to breast cancer (Shaaban et al. 2005). In contrast, expres-sion of eRβ decreased but expression of eRβ2 increased in DCIS, leading investigators to propose that eRβ2 may play an oncogenic role in mammary carcinogenesis (esslimani-Sahla et al. 2005).
In the present study, we determined expression levels of eRβ1, eRβ2, eRα and Ki-67 in normal breast and five types of hyperplastic or noninvasive neoplastic lesions, namely, HUt, aM, aH, low to intermediate-grade (i.e., grade 1 and 2) DCIS (DCIS1/2) and high-grade (i.e., grade 3) DCIS (DCIS3). nuclear expression levels were assessed by immunohistochemistry (IHC) using previously described isoform-specific antibodies to eRβ1 and eRβ2 (Chantzi et al. 2011, 2013). We found that higher than nor-mal Ki-67 lI is correlated with eRα expression and eRβ2 expression in a manner that is in accordance with an eRα-opposing oncosuppressive role of eRβ2 in mammary car-cinogenesis along the HUt, aH and DCIS1/2 pathway.
Methods
Case selection
We studied routinely processed surgical specimens of breast tissue from 55 female patients treated at “Helena Venizelou” general & Maternity District Hospital or at evgenidio Infirmary in athens between 2000 and 2006. the study was approved by the Institutes’ ethics commit-tees. Informed consent for anonymized use of the samples for research purposes was obtained from all patients. the median age of the patients was 49 years (range 32–84), with 27 (49.1 %) patients having entered menopause. In 10 of the specimens, 2 lesions were detected including, 7 spec-imens with HUt and aM, 2 with aDH and aM and 1 with HUt and grade 1 DCIS. as a whole, 79 cases were exam-ined including, 14 cases of normal breast adjacent to HUt lesions with or without adjacent aM (7 and 7 cases, respec-tively), 31 cases of epithelial hyperplasia with or without atypia (15 HUt, 13 aDH and 3 cases of atypical lobular hyperplasia), 13 cases of fibrocystic changes with aM, and 21 cases of DCIS (2 cases of grade 1, 8 of grade 2 and 11 of grade 3). all benign cases originated from specimens lacking invasive cancer foci and all normal cases from
specimens lacking aH or DCIS, since tissue adjacent to invasive breast carcinoma or DCIS is reportedly modified to support tumor aggressiveness (Román-Pérez et al. 2012).
antibodies
eRβ1 and eRβ2 were detected by IHC using previously reported (Chantzi et al. 2011) isoform-specific antibod-ies mB1C1 (mouse monoclonal) and pB2 (rabbit poly-clonal), respectively, as described already (Chantzi et al. 2013). the antibodies were raised against amino acids 517-530 of eRβ1 (mB1C1) and 469-495 of eRβ2 (pB2). the isoform specificity and immunohistochemical perfor-mance of mB1C1 and pB2 were validated as previously described (Chantzi et al. 2011, 2013). Commercially avail-able antibodies were used for immunohistochemical detec-tion of eRα (nCl-eR-6F11, novocastra, UK) and Ki-67 (Pa0118, novocastra, UK) as specified by the suppliers.
Immunohistochemistry
Serial 5-μm-thick tissue sections were heated at 60 °C, deparaffinized in xylene and rehydrated in graded alco-hols. endogenous peroxidase was deactivated by incuba-tion with a solution of 3 % H2O2 in methanol (30 min in the dark). Six microwave heating cycles (800 Watt, 4 min/cycle) with 0.01 Μ citrate buffer (pH 6.0) were used for antigen retrieval. Power Block reagent (Biogenex, USa) was used for 10 min to block non-specific protein bind-ing. antibodies were applied overnight (16–18 h) at 4 °C, at a dilution 2 μg/ml for mB1C1 and 10 μg/ml for pB2. the antibodies to eRα and Ki-67 were used at the dilution recommended by the supplier. Multilink (Biogenex, USa) was applied as secondary antibody for 25 min, followed by Strep-HRP label (Biogenex, USa; 25 min) and chro-mogen 3′,3′-diaminobenzidine tetrahydrochloride (Sigma, USa), as optimal detection system. Sections were coun-terstained with Mayer’s hematoxylin, dehydrated in graded alcohols and cover slipped. non-immune diluent (Re7133; novocastra) compatible with the immunohistochemical kit substituted for the primary antibody and was used as nega-tive control for each case. normal breast tissue was used as positive control.
evaluation of immunohistochemical staining
two independent investigators (n.I.C., D.g.t.) blinded to patient characteristics evaluated the immunohistochemi-cal results. Only nuclear immunostaining of eRα, eRβ1 and eRβ2 was assessed. Immunostaining was evaluated using quick score (QS; Rhodes et al. 2000). Briefly, the nuclear immunostaining intensity (range 0–3: 0 = negative, 1 = mild, 2 = moderate, 3 = strong) and the percentage of

1060 J Cancer Res Clin Oncol (2014) 140:1057–1066
1 3
positive epithelial cells (range 0–4: 0 = 0 %, 1 = 1–24 %, 2 = 25–49 %, 3 = 50–74 % and 4 = 75–100 %) were semi-quantitatively evaluated, and an additive score rang-ing from 0 to 7 was issued for each case. One representa-tive section from each case and an intensity score that cor-responded to that of the majority of tumor cells were used for scoring purposes. For Ki-67 lI, at least 1,000 epithelial cells in 5 different high-power optical fields (magnification 40×) were evaluated and used to calculate the percentage of cells with specifically immunostained nuclei.
Statistical analysis
Statistical analyses were performed using SPSS 13.0 (SPSS Inc., Chicago, Il, USa). the Kruskal–Wallis test was used to compare eRα, eRβ1 and eRβ2 QS across the six groups (i.e., normal breast, aM, HUt, aH, DCIS1/2 and DCIS3) and the Mann–Whitney U test to compare QS between two groups. Ki-67 lI values were compared using t test. Immu-nohistochemical expression of eRα, eRβ1 and eRβ2 was defined as positive using as cutoff the median value of the respective QS. Ki-67 lI was defined as positive using as cutoff the highest Ki-67 lI observed in normal breast. Cor-relations were examined using Spearman’s test. Statistical significance was claimed for p < 0.05.
Results
eRβ1 and eRβ2 immunostaining patterns in benign breast lesions and normal breast
the 65 cases of hyperplastic and noninvasive neoplastic breast lesions and the 14 cases of normal breast were evalu-ated using antibodies mB1C1 to eRβ1 and pB2 to eRβ2 (Chantzi et al. 2011). the isoform specificity and immuno-histochemical performance of the antibodies was assessed using cell lines transiently or stably expressing eRβ1, eRβ2 or neither of the two (Chantzi et al. 2011, 2013). as
already shown, mB1C1 and pB2 detected only the targeted isoform and failed to show positivity of immunostaining with cells expressing neither of eRβ1 and eRβ2 (Chantzi et al. 2011, 2013).
IHC using mB1C1 and pB2 showed predominantly nuclear immunostaining accompanied by mild cytoplasmic immunostaining of normal, hyperplastic and neoplastic epi-thelial cells (Figs. 1, 2, 3), in accordance with previous find-ings (esslimani-Sahla et al. 2005; Roger et al. 2001; Shaaban et al. 2003, 2005). Only nuclear immunostaining was evalu-ated, since cytoplasmic immunostaining is reportedly contro-versial (Roger et al. 2001) and was not taken into account in similar studies (Roger et al. 2001; Shaaban et al. 2003, 2005). Ki-67 was detected by IHC in the nuclei of epithelial cells.
eRβ1 was abundantly (QS = 6 or 7) expressed in nor-mal breast and the majority of breast lesions examined (Fig. 1). eRβ2 displayed by and large low (QS = 2 or 3) to moderate expression (QS = 4 or 5) in normal breast as well as in the lesions. However, we found that 3 out of 13 (3/13) cases (23 %) of aM, 2/16 cases (12.5 %) of aH, 5/10 cases (50 %) of DCIS, 1/2 and 1/11 cases (9.1 %) of high-grade DCIS, but not a single case of normal breast or HUt, expressed eRβ1 but not eRβ2. In fact, none of the 79 cases examined lacked expression of eRβ1 (Figs. 2, 3). all aM lesions lacked expression of eRα (Fig. 1a).
eRα, eRβ1, eRβ2 and Ki-67 expression in benign breast lesions compared with normal breast
table 1 displays the median and the range of Quick Score (Rhodes et al. 2000) of immunohistochemical expression of eRα, eRβ1 and eRβ2 and the mean of Ki-67 lI in normal breast, aM, HUt, aH, DCIS1/2 and DCIS3 and compares lesion scores to normal breast score. eRα expression was moderate (QS = 4) in normal breast and HUt, increased in DCIS1/2 (p = 0.005) and displayed a trend to decrease in DCIS3 (p = 0.064). eRβ1 expression was maximal (QS = 7) in normal breast, aM and HUt and decreased in DCIS1/2 and DCIS3 (p = 0.022 and 0.037, respectively).
Fig. 1 Immunohistochemical expression of eRα and eRβ1 in normal breast and apocrine metaplasia. Serial sections of apocrine metaplasia showing, a negative immunostaining for eRα and b positive nuclear immunostaining for eRβ1. Posi-tive nuclear immunostaining for both eRα and eRβ1 is noted in normal breast epithelium present in upper left corner. Magnification ×100
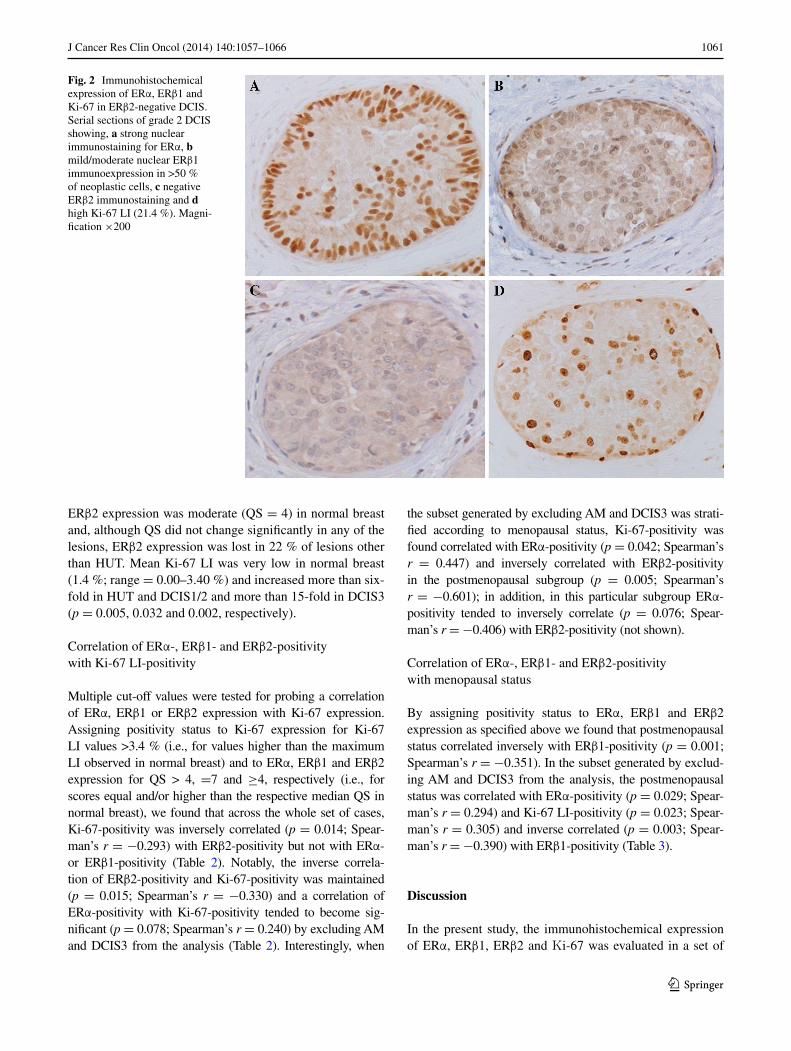
1061J Cancer Res Clin Oncol (2014) 140:1057–1066
1 3
eRβ2 expression was moderate (QS = 4) in normal breast and, although QS did not change significantly in any of the lesions, eRβ2 expression was lost in 22 % of lesions other than HUt. Mean Ki-67 lI was very low in normal breast (1.4 %; range = 0.00–3.40 %) and increased more than six-fold in HUt and DCIS1/2 and more than 15-fold in DCIS3 (p = 0.005, 0.032 and 0.002, respectively).
Correlation of eRα-, eRβ1- and eRβ2-positivity with Ki-67 lI-positivity
Multiple cut-off values were tested for probing a correlation of eRα, eRβ1 or eRβ2 expression with Ki-67 expression. assigning positivity status to Ki-67 expression for Ki-67 lI values >3.4 % (i.e., for values higher than the maximum lI observed in normal breast) and to eRα, eRβ1 and eRβ2 expression for QS > 4, =7 and ≥4, respectively (i.e., for scores equal and/or higher than the respective median QS in normal breast), we found that across the whole set of cases, Ki-67-positivity was inversely correlated (p = 0.014; Spear-man’s r = −0.293) with eRβ2-positivity but not with eRα- or eRβ1-positivity (table 2). notably, the inverse correla-tion of eRβ2-positivity and Ki-67-positivity was maintained (p = 0.015; Spearman’s r = −0.330) and a correlation of eRα-positivity with Ki-67-positivity tended to become sig-nificant (p = 0.078; Spearman’s r = 0.240) by excluding aM and DCIS3 from the analysis (table 2). Interestingly, when
the subset generated by excluding aM and DCIS3 was strati-fied according to menopausal status, Ki-67-positivity was found correlated with eRα-positivity (p = 0.042; Spearman’s r = 0.447) and inversely correlated with eRβ2-positivity in the postmenopausal subgroup (p = 0.005; Spearman’s r = −0.601); in addition, in this particular subgroup eRα-positivity tended to inversely correlate (p = 0.076; Spear-man’s r = −0.406) with eRβ2-positivity (not shown).
Correlation of eRα-, eRβ1- and eRβ2-positivity with menopausal status
By assigning positivity status to eRα, eRβ1 and eRβ2 expression as specified above we found that postmenopausal status correlated inversely with eRβ1-positivity (p = 0.001; Spearman’s r = −0.351). In the subset generated by exclud-ing aM and DCIS3 from the analysis, the postmenopausal status was correlated with eRα-positivity (p = 0.029; Spear-man’s r = 0.294) and Ki-67 lI-positivity (p = 0.023; Spear-man’s r = 0.305) and inverse correlated (p = 0.003; Spear-man’s r = −0.390) with eRβ1-positivity (table 3).
Discussion
In the present study, the immunohistochemical expression of eRα, eRβ1, eRβ2 and Κi-67 was evaluated in a set of
Fig. 2 Immunohistochemical expression of eRα, eRβ1 and Ki-67 in eRβ2-negative DCIS. Serial sections of grade 2 DCIS showing, a strong nuclear immunostaining for eRα, b mild/moderate nuclear eRβ1 immunoexpression in >50 % of neoplastic cells, c negative ΕRβ2 immunostaining and d high Ki-67 lI (21.4 %). Magni-fication ×200
1062 J Cancer Res Clin Oncol (2014) 140:1057–1066
1 3
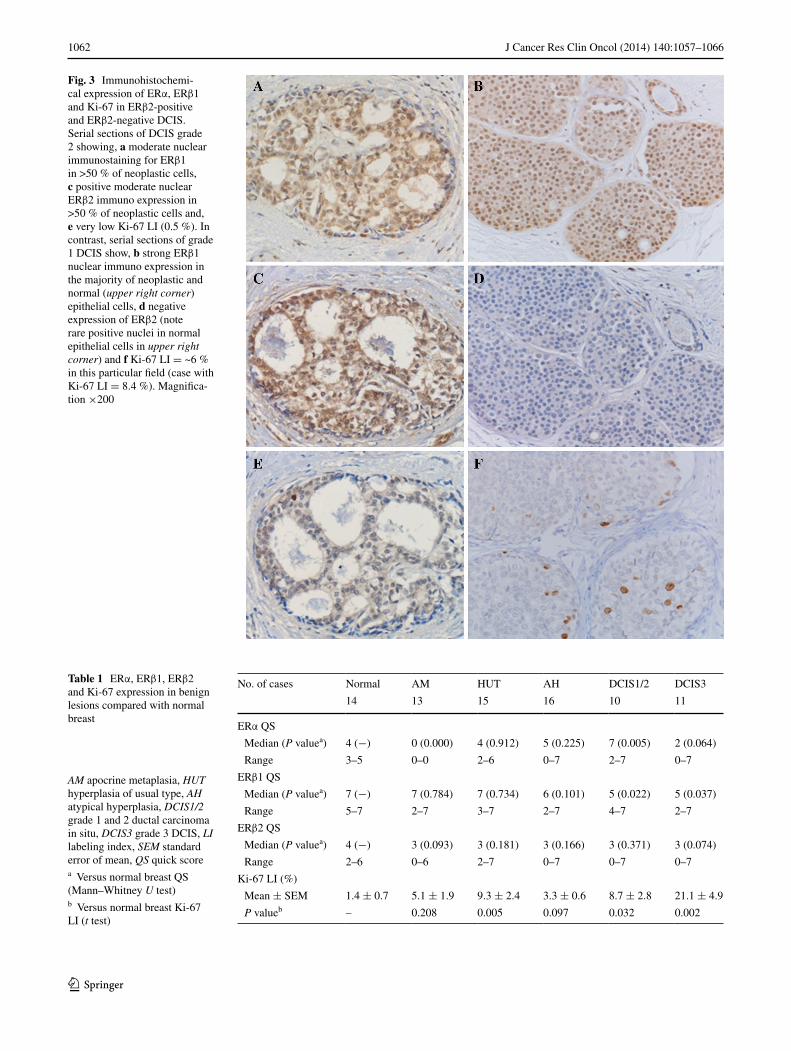
Fig. 3 Immunohistochemi-cal expression of eRα, eRβ1 and Ki-67 in eRβ2-positive and eRβ2-negative DCIS. Serial sections of DCIS grade 2 showing, a moderate nuclear immunostaining for eRβ1 in >50 % of neoplastic cells, c positive moderate nuclear eRβ2 immuno expression in >50 % of neoplastic cells and, e very low Ki-67 lI (0.5 %). In contrast, serial sections of grade 1 DCIS show, b strong eRβ1 nuclear immuno expression in the majority of neoplastic and normal (upper right corner) epithelial cells, d negative expression of eRβ2 (note rare positive nuclei in normal epithelial cells in upper right corner) and f Ki-67 lI = ~6 % in this particular field (case with Ki-67 lI = 8.4 %). Magnifica-tion ×200
Table 1 eRα, eRβ1, eRβ2 and Ki-67 expression in benign lesions compared with normal breast
AM apocrine metaplasia, HUT hyperplasia of usual type, AH atypical hyperplasia, DCIS1/2 grade 1 and 2 ductal carcinoma in situ, DCIS3 grade 3 DCIS, LI labeling index, SEM standard error of mean, QS quick scorea Versus normal breast QS (Mann–Whitney U test)b Versus normal breast Ki-67 lI (t test)
no. of cases normal aM HUt aH DCIS1/2 DCIS3
14 13 15 16 10 11
eRα QS
Median (P valuea) 4 (−) 0 (0.000) 4 (0.912) 5 (0.225) 7 (0.005) 2 (0.064)
Range 3–5 0–0 2–6 0–7 2–7 0–7
eRβ1 QS
Median (P valuea) 7 (−) 7 (0.784) 7 (0.734) 6 (0.101) 5 (0.022) 5 (0.037)
Range 5–7 2–7 3–7 2–7 4–7 2–7
eRβ2 QS
Median (P valuea) 4 (−) 3 (0.093) 3 (0.181) 3 (0.166) 3 (0.371) 3 (0.074)
Range 2–6 0–6 2–7 0–7 0–7 0–7
Ki-67 lI (%)
Mean ± SeM 1.4 ± 0.7 5.1 ± 1.9 9.3 ± 2.4 3.3 ± 0.6 8.7 ± 2.8 21.1 ± 4.9
P valueb – 0.208 0.005 0.097 0.032 0.002

1063J Cancer Res Clin Oncol (2014) 140:1057–1066
1 3
65 cases of hyperplastic and noninvasive neoplastic breast lesions and 14 cases of adjacent normal breast tissue. Com-pared with normal breast, eRα expression increased in DCIS1/2 and tended to decrease in DCIS3, while eRβ1 expression decreased in DCIS irrespective of grade and, although eRβ2 QS in the lesions was not significantly lower compared with normal breast, eRβ2 expression was lost with varying frequency in lesions other than HUt. In contrast, the mean Ki-67 lI in HUt, DCIS1/2 and DCIS3 was considerably higher than in normal breast. these find-ings suggest that expression of eRα and eRβ1 is dynami-cally regulated during mammary carcinogenesis and that expression of eRβ2 is occasionally lost in the more pro-gressed hyperplastic and neoplastic breast lesions.
It has been reported that development of eRα-positive breast carcinoma is associated with lower eRβ2 mRna expression compared with adjacent normal tissue (Park et al. 2006). thus, it appears that down-regulation of eRβ2 expression is relatively common in invasive breast carci-noma as well as in hyperplastic and noninvasive neoplas-tic breast lesions with the exception of HUt. In contrast, all lesions and adjacent normal breast that we exam-ined expressed eRβ1. as expected from previous studies (Selim and Wells, 1999), aM lacked expression of eRα. the absence of eRα in apocrine metaplastic cells report-edly reflects genetic differences from normal cells. at least a subset of lesions with apocrine morphology is known to display considerable genetic changes, whereas such
Table 2 Correlation of Ki-67-positivity with eRα-, eRβ1- and eRβ2-positivity
AM apocrine metaplasia, HUT hyperplasia of usual type, AH atypical hyperplasia, DCIS1/2 grade 1 and 2 DCIS, DCIS3 grade 3 DCISa Ki-67-positive cases are considered those with lI > 3.4 % (maximum lI in normal breast)b eRα, eRβ1 and eRβ2-positive cases are considered those with QS > 4, =7 and ≥4, respectively
Positivity no. of positive cases (% of all cases of the group) eR-positivity correlated with Ki-67-positivity
normal 14 (100) aM 13 (100) HUt 15 (100) aH 16 (100) DCIS1/2 10 (100) DCIS3 11 (100) Spearman’s R P value
Ki-67a 0 (0.00) 6 (46.2) 10 (66.7) 9 (56.3) 7 (70.0) 9 (81.8) – –
eRαb 5 (36.4) 0 (0.00) 5 (33.3) 10 (62.5) 9 (90.0) 2 (18.2) 0.190 0.120
eRβ1b 8 (57.1) 8 (61.5) 8 (53.3) 6 (37.5) 2 (20.0) 3 (27.3) −0.019 0.877
eRβ2b 11 (78.6) 5 (38.5) 7 (46.7) 6 (37.5) 5 (50.0) 3 (27.3) −0.293 0.014
eRαb 5 (36.4) – 5 (33.3) 10 (62.5) 9 (90.0) – 0.240 0.078
eRβ1b 8 (57.1) – 8 (53.3) 6 (37.5) 2 (20.0) – −0.099 0.473
eRβ2b 11 (78.6) – 7 (46.7) 6 (37.5) 5 (50.0) – −0.330 0.015
Table 3 Correlation of postmenopausal status with eRα-, eRβ1-, eRβ2- and Ki-67-positivity
AM apocrine metaplasia, HUT hyperplasia of usual type, AH atypical hyperplasia, DCIS1/2 grade 1 and 2 DCIS, DCIS3 grade 3 DCIS, PostMeS-tat* Postmenopausal Statusa Ki-67-positive cases are considered those with lI > 3.4 % (maximum lI in normal breast)b eRα, eRβ1 and eRβ2-positive cases are considered those with QS > 4, =7 and ≥4, respectively
Positivity no. of positive cases (% of all cases of the group) Postmenopausal status correlated with eR-positivity and Ki-67-positivity
normal 14 (100) aM 13 (100) HUt 15 (100) aH 16 (100) DCIS1/2 10 (100) DCIS3 11 (100) Spearman’s R P value
PostMeStat* 1 (7.10) 5 (38.5) 3 (20.0) 11 (68.8) 6 (60.0) 5 (45.5) – –
Ki-67a 0 (0.00) 6 (46.2) 10 (66.7) 9 (56.3) 7 (70.0) 9 (81.8) 0.385 0.099
eRαb 5 (36.4) 0 (0.00) 5 (33.3) 10 (62.5) 9 (90.0) 2 (18.2) 0.159 0.168
eRβ1b 8 (57.1) 8 (61.5) 8 (53.3) 6 (37.5) 2 (20.0) 3 (27.3) −0.351 0.001
eRβ2b 11 (78.6) 5 (38.5) 7 (46.7) 6 (37.5) 5 (50.0) 3 (27.3) −0.104 0.369
Ki-67a 1 (7.10) – 3 (20.0) 11 (68.8) 6 (60.0) – 0.305 0.023
eRαb 5 (36.4) – 5 (33.3) 10 (62.5) 9 (90.0) – 0.294 0.029
eRβ1b 8 (57.1) – 8 (53.3) 6 (37.5) 2 (20.0) – −0.390 0.003
eRβ2b 11 (78.6) – 7 (46.7) 6 (37.5) 5 (50.0) – −0.134 0.334

1064 J Cancer Res Clin Oncol (2014) 140:1057–1066
1 3
changes are either not observed or are very rare in HUt (lopez-garcia et al. 2010). notably, from all the lesions we examined only HUt failed to display loss of eRβ2 expres-sion. On the other hand, the significant decrease in eRβ1 expression in DCIS1/2 and DCIS3 may reflect methylation-induced silencing of eRβ gene promoter activity (Rody et al. 2005).
It has been reported that high eRα expression in normal breast and HUt is correlated with Ki-67 expression and increased breast cancer risk (Khan et al. 1998; Shaaban et al. 2002). We showed that higher than normal Ki-67 lI and eRα-positivity tended to be correlated in the subset generated by excluding aM and DCIS3 (i.e., eRα-negative lesions) and that they were correlated in the postmeno-pausal subgroup of this subset. the correlation of Ki-67 positivity with eRα-positivity in this subgroup is taken to reflect that higher levels of eRα can sensitize in situ pro-liferative lesions to postmenopausal levels of estrogen. We also showed that higher than normal Ki-67 lI and eRβ2-positivity were inversely correlated in the whole set, in the subset generated by excluding aM and DCIS3 and in the postmenopausal subgroup of this subset, in particular, where a tendency of inverse correlation of eRα-positivity and eRβ2-positivity was apparent as well. the inverse correlation of Ki-67 positivity with eRβ2-positivity could reflect that eRβ2 can challenge eRα integrity even in the presence of low levels of estrogen (Zhao et al. 2007) and thereby inhibit eRα-mediated stimulation of cell prolifera-tion. the tendency of inverse correlation of eRα-positivity and eRβ2-positivity we observed in this subgroup could reflect eRβ2-induced down-regulation of eRα by the pro-teasome (Zhao et al. 2007). these findings indicate that low eRβ2 expression is possibly correlated with high eRα and Ki-67 expression and consequently with increased rate of progression along the HUt-aH-DCIS1/2 pathway and are in accordance with an eRα-opposing oncosuppressive role of eRβ2 in mammary carcinogenesis. that eRβ2 expres-sion is associated with good clinical outcome in breast cancer and that both eRα- and eRβ2-positive tumors, in particular, display the best clinical outcome (Shaaban et al. 2008) are in accordance with an eRα-opposing oncosup-pressive role of eRβ2. However, others have reported that eRβ2 expression is up-regulated in spite of eRβ expres-sion being down-regulated in DCIS (esslimani-Sahla et al. 2005; Roger et al. 2001); and that the up-regulation may be taken to suggest that eRβ2 plays an oncogenic role in mammary carcinogenesis (esslimani-Sahla et al. 2005).
It has been reported that eRα expression in in situ prolif-erating atypical and/or hyperplastic breast lesions increases after menopause (Sharma et al. 2006); that eRβ1 and eRβ2 exert antiproliferative action on eRα-positive breast cancer cells (Paruthiyil et al. 2004; Ström et al. 2004; Zhao et al. 2007); that eRα/eRβ1 heterodimers are antiproliferative in
breast cancer cells that express both isotypes (Chang et al. 2006; Powell et al. 2012); and that eRβ2 heterodimerizes with eRα and inhibits its estrogen-dependent transcriptional activity in addition to challenging its integrity (Ogawa et al. 1998; Peng et al. 2003; Zhao et al. 2007). Since eRα stim-ulates and eRβ1 and eRβ2 inhibit cell proliferation, their relative levels are expected to affect the proliferative activity of atypical and/or hyperplastic breast lesions. In the present study we showed that postmenopausal status is correlated with low eRβ1 expression and high eRα and Ki-67 expres-sion in the subset generated by excluding aM and DCIS3; that high Ki-67 expression is correlated with high eRα expression and low eRβ2 expression in the postmenopausal subgroup of the this subset; and that high eRα expression and low eRβ2 expression tended to be correlated in this subgroup. It is therefore tempting to speculate that in this postmenopausal subgroup, in particular, eRβ2 could outper-form eRβ1 in forming heterodimers with eRα and in keep-ing antiproliferative control in spite of the presence of low levels of eRβ1; and that loss or down-regulation of eRβ2 expression would wave or weaken control over eRα expres-sion and eRα-dependent cell proliferation. these findings support an eRα-opposing oncosuppressive role of eRβ2 that is particularly important in preventing progression along the HUt-aH-DCIS1/2 pathway after menopause.
Recent reports suggested mechanisms likely affecting eRα and eRβ1 expression after menopause. It appears that in eRα-positive breast cancer cells estradiol up-regulates the expression of forkhead transcription factor FoxO3a (Morelli et al. 2010) which in turn interacts with eRα and eRβ1 and inhibits their estradiol-dependent tran-scriptional activities (Zou et al. 2008); in addition, eRβ1 down-regulates eRα promoter activity, eRα expression and eRα-dependent breast cancer cell proliferation (Bar-tella et al. 2012). It is therefore tempting to speculate that the postmenopausal decrease in circulating estrogen and the ensuing down-regulation of FoxO3a expression and of eRα/eRβ1-mediated repression of eRα expression on top of the decrease in eRβ1 expression could cooperatively lead to postmenopausal up-regulation of eRα expression. Considering the oncogenic potential of eRα (Khan et al. 1998), the oncosuppressive role of eRβ1 (Bardin et al. 2004; Bartella et al. 2012; thomas and gustafsson 2011) and the increased breast cancer risk of HUt with high eRα/eRβ1 expression ratio (Shaaban et al. 2005), the post-menopausal decrease in eRβ1 expression in hyperplastic and noninvasive neoplastic breast lesions could underlie the increased importance of an eRα-opposing oncosuppressive role of eRβ2 in preventing progression along the HUt-aH-DCIS1/2 pathway after menopause.
We recently reported that high nuclear eRβ2 expression is an independent marker of relapse of eRα-negative breast carcinoma (Chantzi et al. 2013). Others have reported that

1065J Cancer Res Clin Oncol (2014) 140:1057–1066
1 3
in eRα-positive breast cancer, eRβ2 expression is associ-ated with response to endocrine therapy, longer relapse-free and overall survival and lower rates of metastasis and vascu-lar invasion (Palmieri et al. 2004; Shaaban et al. 2008). On the other hand, Bozkurt and Kapucuoglu (2012) reported recently that, while eRβ2 expression is associated with breast cancer invasiveness, co-expression of eRα and eRβ2 in particular is associated with low histological grade. evi-dently, eRβ2 may take different roles in the absence and presence of eRα. the role of eRβ1 in breast cancer pro-gression may also differ depending on the type of mammary lesion (leygue and Murphy, 2013). For instance, it has been reported that high eRβ1 expression is associated with good prognosis in luminal a node-negative but not node-positive patients and with poor prognosis in luminal B node-positive but not node-negative patients (novelli et al. 2008).
the main limitation of our report is the relatively small number of lesions that we studied. In spite of this limita-tion, our findings are in full agreement with previous reports showing that expression of eRα and eRβ1 (as well as eRβ) increases and decreases, respectively, during mam-mary carcinogenesis (Roger et al. 2001; Shaaban et al. 2003; Shoker et al. 1999); that eRα expression is most common in low-grade and least common in high-grade DCIS (Claus et al. 2001; lopez-garcia et al. 2010; Roger et al. 2001); and that high Ki-67 lI in DCIS is associated with low eRα expression and high nuclear grade (Roger et al. 2001). In addition, we report for the first time that eRβ1 expression in normal breast and in hyperplastic and neoplastic breast lesions declines after menopause and that eRβ2 likely has an oncosuppressive role in mammary car-cinogenesis along the HUt-aH-low-grade DCIS pathway especially after menopause. Further studies with followed-up patients are expected to clarify the role of eRβ1 and eRβ2 in the pathogenesis of hyperplastic and neoplastic breast disease and enable assessment of their prognostic significance.
Acknowledgments this study was supported by eU Social Fund and greek national funds through the Operational Program ‘educa-tion and lifelong learning’ (Projects tHaleS 85355) and the nSRF Program (Project 03eD644).
Conflict of interest the authors declare that they have no conflict of interest.
References
Bardin a, Boulle n, lazennec g, Vignon F, Pujol P (2004) loss of eRbeta expression as a common step in estrogen-dependent tumor progression. endocr Relat Cancer 11:537–551. doi:10.1677/erc.1.00800
Bartella V, Rizza P, Barone I, Zito D, giordano F, giordano C, Cata-lano S, Mauro l, Sisci D, Panno Ml, Fuqua Sa, andò S (2012)
estrogen receptor beta binds Sp1 and recruits a corepressor com-plex to the estrogen receptor alpha gene promoter. Breast Cancer Res treat 134:569–581. doi:10.1007/s10549-012-2090-9
Bozkurt KK, Kapucuoglu n (2012) Investigation of immuno-histochemical eRα, eRβ and eRβcx expressions in normal and neoplastic breast tissues. Pathol Res Pract 208:133–139. doi:10.1016/j.prp.2011.12.015
Chang eC, Frasor J, Komm B, Katzenellenbogen BS (2006) Impact of estrogen receptor beta on gene networks regulated by estrogen receptor alpha in breast cancer cells. endocrinology 147:4831–4842. doi:10.1210/en.2006-0563
Chantzi nI, Meligova aK, Dhimolea e, Petrou CC, Mitsiou DJ, Magafa V, Pechtelidou a, Florentin I, Kitraki e, Cordopatis P, tini-akos Dg, alexis Mn (2011) Insights into ectopic estrogen recep-tor expression, nucleocytoplasmic distribution and interaction with chromatin obtained with new antibodies to estrogen receptors α and β. Steroids 76:974–985. doi:10.1016/j.steroids.2011.05.010
Chantzi nI, tiniakos Dg, Palaiologou M, goutas n, Filippidis t, Vassilaros SD, Dhimolea e, Mitsiou DJ, alexis Mn (2013) estrogen receptor beta 2 is associated with poor prognosis in estrogen receptor alpha-negative breast carcinoma. J Cancer Res Clin Oncol 139:1489–1498. doi:10.1007/s00432-013-1467-4
Claus eB, Chu P, Howe Cl, Davison tl, Stern DF, Carter D, Digiovanna MP (2001) Pathobiologic findings in DCIS of the breast: morphologic features, angiogenesis, HeR-2/neu and hormone receptors. exp Mol Pathol 70:303–316. doi:10.1006/exmp.2001.2366
Dowsett M, a’Hern R, Salter J, Zabaglo l, Smith Ie (2009) Who would have thought a single Ki67 measurement would pre-dict long-term outcome? Breast Cancer Res 11(Suppl 3):S15. doi:10.1186/bcr2434
esslimani-Sahla M, Kramar a, Simony-lafontaine J, Warner M, gustafsson Ja, Rochefort H (2005) Increased estrogen receptor betacx expression during mammary carcinogenesis. Clin Cancer Res 11:3170–3174. doi:10.1006/exmp.2001.2366
Hanahan D, Weinberg Ra (2011) Hallmarks of cancer: the next gen-eration. Cell 144:646–674. doi:10.1016/j.cell.2011.02.013
Henderson Be, Feigelson HS (2000) Hormonal carcinogenesis. Car-cinogenesis 21:427–433. doi:10.1093/carcin/21.3.427
Honma n, Horii R, Iwase t, Saji S, younes M, takubo K, Matsuura M, Ito y, akiyama F, Sakamoto g (2008) Clinical importance of estrogen receptor-beta evaluation in breast cancer patients treated with adjuvant tamoxifen therapy. J Clin Oncol 26:3727–3734. doi:10.1200/JCO.2007.14.2968
Khan Sa, Rogers Ma, Khurana KK, Meguid MM, numann PJ (1998) estrogen receptor expression in benign breast epithelium and breast cancer risk. J natl Cancer Inst 90:37–42. doi:10.1093/jnci/90.1.37
leung yK, Mak P, Hassan S, Ho SM (2006) estrogen receptor (eR)-beta isoforms: a key to understanding eR-beta signal-ing. Proc natl acad Sci USa 103:13162–13167. doi:10.1073/pnas.0605676103
leung yK, lee Mt, lam HM, tarapore P, Ho SM (2012) estro-gen receptor-beta and breast cancer: translating biology into clinical practice. Steroids 77:727–737. doi:10.1016/j.steroids.2012.03.008
leygue e, Murphy lC (2013) a bi-faceted role of estrogen receptor β in breast cancer. endocr Relat Cancer 20:R127–R139. doi:10.1530/eRC-12-0389
lopez-garcia Ma, geyer FC, lacroix-triki M, Marchió C, Reis-Filho JS (2010) Breast cancer precursors revisited: molecular features and progression pathways. Histopathology 57:171–192. doi:10.1111/j.1365-2559.2010.03568.x
Moore Jt, McKee DD, Slentz-Kesler K, Moore lB, Jones Sa, Horne el, Su Jl, Kliewer Sa, lehmann JM, Willson tM (1998) Clon-ing and characterization of human estrogen receptor beta iso-forms. Biochem Biophys Res Commun 247:75–78

1066 J Cancer Res Clin Oncol (2014) 140:1057–1066
1 3
Morelli C, lanzino M, garofalo C, Maris P, Brunelli e, Casaburi I, Cat-alano S, Bruno R, Sisci D, andò S (2010) akt2 inhibition enables the forkhead transcription factor FoxO3a to have a repressive role in estrogen receptor alpha transcriptional activity in breast cancer cells. Mol Cell Biol 30:857–8570. doi:10.1128/MCB.00824-09
Murphy lC, leygue e (2012) the role of estrogen receptor-β in breast cancer. Semin Reprod Med 30:5–13. doi:10.1055/s-0031-1299592
nakopoulou l, lazaris aC, Panayotopoulou eg, giannopoulou I, givalos n, Markaki S, Keramopoulos a (2004) the favourable prognostic value of oestrogen receptor beta immunohistochemi-cal expression in breast cancer. J Clin Pathol 57:523–528. doi:10.1136/jcp.2003.008599
novelli F, Milella M, Melucci e, Di Benedetto a, Sperduti I, Perrone-Donnorso R, Perracchio l, Venturo I, nisticò C, Fabi a, Buglioni S, natali Pg, Mottolese M (2008) a divergent role for estrogen receptor-beta in node-positive and node-negative breast cancer classified according to molecular subtypes: an observational pro-spective study. Breast Cancer Res 10:R74. doi:10.1186/bcr2139
Ogawa S, Inoue S, Watanabe t, Orimo a, Hosoi t, Ouchi y, Mura-matsu M (1998) Molecular cloning and characterization of human estrogen receptor betacx: a potential inhibitor of estrogen action in human. nucleic acids Res 26:3505–3512. doi:10.1093/nar/26.15.3505
Palmieri C, lam eW, Mansi J, MacDonald C, Shousha S, Madden P, Omoto y, Sunters a, Warner M, gustafsson Ja, Coombes RC (2004) the expression of eR beta cx in human breast cancer and the relationship to endocrine therapy and survival. Clin Cancer Res 10:2421–2428. doi:10.1158/1078-0432.CCR-03-0215
Park BW, Kim KS, Heo MK, yang WI, Kim SI, Kim JH, Kim ge, lee KS (2006) the changes of estrogen receptor-beta variants expression in breast carcinogenesis: decrease of estrogen recep-tor-beta2 expression is the key event in breast cancer develop-ment. J Surg Oncol 93:504–510. doi:10.1002/jso.20336
Paruthiyil S, Parmar H, Kerekatte V, Cunha gR, Firestone gl, leitman DC (2004) estrogen receptor beta inhibits human breast cancer cell proliferation and tumor formation by causing a g2 cell cycle arrest. Cancer Res 64:423–428. doi:10.1158/0008-5472.Can-03-2446
Patani n, Martin la, Dowsett M (2013) Biomarkers for the clini-cal management of breast cancer: international perspective. Int J Cancer 33:1–13. doi:10.1002/ijc.27997
Peng B, lu B, leygue e, Murphy lC (2003) Putative functional characteristics of human estrogen receptor-beta isoforms. J Mol endocrinol 30:13–29. doi:10.1677/jme.0.0300013
Powell e, Shanle e, Brinkman a, li J, Keles S, Wisinski KB, Huang W, Xu W (2012) Identification of estrogen receptor dimer selec-tive ligands reveals growth-inhibitory effects on cells that co-express eRα and eRβ. PloS One 7:e30993. doi:10.1371/journal.pone.0030993
Rhodes a, Jasani B, Balaton aJ, Miller KD (2000) Immunohisto-chemical demonstration of oestrogen and progesterone receptors: correlation of standards achieved on in house tumours with that achieved on external quality assessment material in over 150 labo-ratories from 26 countries. J Clin Pathol 53:292–301. doi:10.1136/jcp.53.4.292
Rody a, Holtrich U, Solbach C, Kourtis K, von Minckwitz g, engels K, Kissler S, gätje R, Karn t, Kaufmann M (2005) Methylation of estrogen receptor beta promoter correlates with loss of eR-beta expression in mammary carcinoma and is an early indication marker in premalignant lesions. endocr Relat Cancer 12:903–916. doi:10.1677/erc.1.01088
Roger P, Sahla Me, Mäkelä S, gustafsson Ja, Baldet P, Rochefort H (2001) Decreased expression of estrogen receptor beta pro-tein in proliferative preinvasive mammary tumors. Cancer Res 61:2537–2541
Román-Pérez e, Casbas-Hernández P, Pirone JR, Rein J, Carey la, lubet Ra, Mani Sa, amos KD, troester Ma (2012) gene
expression in extratumoral microenvironment predicts clini-cal outcome in breast cancer patients. Breast Cancer Res 14:R51. doi:10.1186/bcr3152
Selim ag, Wells Ca (1999) Immunohistochemical localisation of androgen receptor in apocrine metaplasia and apocrine adenosis of the breast: relation to oestrogen and progesterone receptors. J Clin Pathol 52:838–841. doi:10.1136/jcp.52.11.838
Shaaban aM, Sloane JP, West CR, Foster CS (2002) Breast cancer risk in usual ductal hyperplasia is defined by estrogen receptor-alpha and Ki-67 expression. am J Pathol 60:597–604
Shaaban aM, O’neill Pa, Davies MP, Sibson R, West CR, Smith PH, Foster CS (2003) Declining estrogen receptor-beta expression defines malignant progression of human breast neoplasia. am J Surg Pathol 27:1502–1512
Shaaban aM, Jarvis C, Moore F, West C, Dodson a, Foster CS (2005) Prognostic significance of estrogen receptor Beta in epithelial hyperplasia of usual type with known outcome. am J Surg Pathol 29:1593–1599
Shaaban aM, green aR, Karthik S, alizadeh y, Hughes ta, Harkins l, ellis IO, Robertson JF, Paish eC, Saunders Pt, groome nP, Speirs V (2008) nuclear and cytoplasmic expression of eRbeta1, eRbeta2, and eRbeta5 identifies distinct prognostic outcome for breast cancer patients. Clin Cancer Res 14:5228–5235. doi:10.1158/1078-0432.CCR-07-4528
Sharma P, Kimler BF, Warner C, Metheny t, Xue Q, Zalles CM, Fabian CJ (2006) estrogen receptor expression in benign breast ductal cells obtained from random periareolar fine needle aspi-ration correlates with menopausal status and cytomorphology index score. Breast Cancer Res treat 100:71–76. doi:10.1007/s10549-006-9234-8
Shoker BS, Jarvis C, Sibson DR, Walker C, Sloane JP (1999) Oestro-gen receptor expression in the normal and pre-cancerous breast. J Pathol 188:237–244. doi:10.1002/(SICI)10969896(199907)188:3<237:aIDPatH343>3.0.CO;2-8
Skliris gP, leygue e, Curtis-Snell l, Watson PH, Murphy lC (2006) expression of oestrogen receptor-beta in oestrogen receptor-alpha negative human breast tumours. Br J Cancer 95:616–626. doi:10.1038/sj.bjc.6603295
Skliris gP, leygue e, Watson PH, Murphy lC (2008) estrogen recep-tor alpha negative breast cancer patients: estrogen receptor beta as a therapeutic target. J Steroid Biochem Mol Biol 109:1–10. doi:10.1016/j.jsbmb.2007.12.010
Snyder Ma, Smejkalova t, Forlano PM, Woolley CS (2010) Multiple eRbeta antisera label in eRbeta knockout and null mouse tissues. J neurosci Methods 188:226–234. doi:10.1016/j.jneumeth.2010.02.012
Ström a, Hartman J, Foster JS, Kietz S, Wimalasena J, gustafsson Ja (2004) estrogen receptor beta inhibits 17beta-estradiol-stimu-lated proliferation of the breast cancer cell line t47D. Proc natl acad Sci USa 101:1566–1571. doi:10.1073/pnas.0308319100
thomas C, gustafsson JÅ (2011) the different roles of eR subtypes in cancer biology and therapy. nat Rev Cancer 11:597–608. doi:10.1038/nrc3093
van Diest PJ, van der Wall e, Baak JP (2004) Prognostic value of proliferation in invasive breast cancer: a review. J Clin Pathol 57:675–681. doi:10.1136/jcp.2003.010777
Zhao C, Matthews J, tujague M, Wan J, Ström a, toresson g, lam eW, Cheng g, gustafsson Ja, Dahlman-Wright K (2007) estro-gen receptor beta2 negatively regulates the transactivation of estrogen receptor alpha in human breast cancer cells. Cancer Res 67:3955–3962. doi:10.1158/0008-5472.Can-06-3505
Zou y, tsai WB, Cheng CJ, Hsu C, Chung yM, li PC, lin SH, Hu MC (2008) Forkhead box transcription factor FOXO3a sup-presses estrogen-dependent breast cancer cell proliferation and tumorigenesis. Breast Cancer Res 10:R21. doi:10.1186/bcr1872




















