
ERβ1 and the ERβ2 Splice Variant (ERβcx/β2) Are Expressed in Distinct Cell Populations in the...
10
ER1 and the ER2 Splice Variant (ERcx/ 2) Are Expressed in Distinct Cell Populations in the Adult Human Testis PHILIPPA T. K. SAUNDERS, MICHAEL R. MILLAR, SHEILA MACPHERSON, D. STEWART IRVINE, NIGEL P. GROOME, LEE R. EVANS, RICHARD M. SHARPE, AND GRAEME A. SCOBIE Medical Research Council Human Reproductive Sciences Unit (P.T.K.S., M.R.M., S.M., D.S.I., R.M.S., G.A.S.), Centre for Reproductive Biology, Edinburgh EH3 9ET, United Kingdom; and School of Biological & Molecular Sciences (N.P.G., L.R.E.), Oxford Brookes University, Gypsy Lane Campus, Headington, Oxford OX3 0PB, United Kingdom Estrogens can regulate germ cell function. Estrogen action is mediated via high affinity ERs; two subtypes (ER and ER) have been identified. We have shown previously that ER is expressed in nuclei of multiple human testicular cells. A vari- ant isoform of human (h) ER (hERcx/2), formed by alterna- tive splicing, has been identified in testicular cDNA libraries by two laboratories. The present study examined the expres- sion of wild-type (ER1) and variant (ER2) receptors in human testes by 1) RT-PCR with isoform specific primers, and 2) single and double immunohistochemistry using monoclo- nal antibodies raised against peptides unique to the C termini of hER1 and hER2. PCR products specific for ER1 and ER2 were amplified from cDNA pools prepared from human testes and granulosa cells. On Western blots, the anti-ER1 monoclonal antibody bound to recombinant ER1 and the anti-ER2 monoclonal to recombinant hER2. Neither bound to the other ER isoform nor to recombinant ER. ER1 and ER2 proteins were both detected in human testis. Immuno- expression of ER1 was most intense in pachytene spermato- cytes and round spermatids, whereas low levels of expression were detected in Sertoli cells, spermatogonia, preleptotene, leptotene, zygotene, and diplotene spermatocytes. Highest levels of expression of ER2 protein were detected in Sertoli cells and spermatogonia with low/variable expression in preleptotene, pachytene, and diplotene spermatocytes. No im- munostaining was detected in elongating spermatids. Most interstitial cells expressed more ER2 than ER1. It is spec- ulated that the cells most susceptible to modulation by estro- genic ligands are round spermatids in which levels of expres- sion of ER1 are high. In contrast, expression of ER2, an isoform that may act as a dominant negative inhibitor of ER action, in Sertoli cells and spermatogonia, could protect these cells from adverse effects of estrogens. (J Clin Endocrinol Metab 87: 2706 –2715, 2002) I T HAS BEEN proposed that estrogens may play a role in regulating spermatogenesis (see review by Ref. 1). For example, Ebling et al. (2) found estrogen treatment of hpg mice stimulated spermatogenesis and Pentikainen et al. (3) reported that estrogen exposure reduced levels of germ cell apoptosis in isolated human seminiferous tubules consistent with estrogen acting as a germ cell survival factor. The bio- synthesis of estrogens from androgens is catalyzed by the aromatase complex. Levels of estrogen within the male re- productive tract are higher than in the general circulation consistent with production of estrogens (reviewed in Ref. 4). In the adult testis, the major site of aromatase expression appears to be the Leydig cells (5, 6); however, aromatase activity has also been detected within mature germ cells and spermatozoa (6 – 8). Male mice in which estrogen biosynthe- sis is impaired due to targeted disruption of the aromatase gene (ARKO) become infertile (9). Analyses of the testes of these animals has shown that round spermatids undergo apoptosis, show disturbances in acrosome formation, and fail to differentiate into mature elongate spermatids (9, 10). Consistent with estrogens acting as a germ cell survival fac- tor treatment of monkeys with an aromatase inhibitor also results in a reduction in the progression of round to elongate spermatids (11). Estrogen action is mediated via high affinity receptors that are shifted to a transcriptionally active state after ligand binding. Two forms of ER commonly known as (ER, NR3A1) and (ER, NR3A2) have been cloned from human (h) (12–14) and other species (15, 16). Both receptors are encoded by eight exons and are the products of two genes located on different chromosomes (17). Phylogenetic analysis suggests that they arose from a single duplication event at least 450 million years ago (18). Like other members of the steroid receptor superfamily, ER and ER share a common arrangement of five structure-function domains, denoted A to F (19). Key domains include the DNA binding domain (C), two transactivation domains [activation function (AF)-1 and AF-2, located in the N (A/B domain) and C-terminal (E/F domain) portions of the protein, and regions within the C and E domains involved in receptor dimerization (see reviews in Refs. 20 and 21). Studies in vitro have demonstrated that homodimers (ER-ER or ER-ER), or heterodimers (ER- ER), can be formed when both isoforms are expressed in the same cell (22, 23). In 1998, a novel human ER variant, named hERcx, was identified by screening a human testis cDNA library (Ref. 24; GenBank accession no. AB006589). The open reading frame of the protein was predicted to be identical to that of full-length hER (59 kDa, Ref. 14, GenBank accession Abbreviations: AF, Activation function; ARKO, aromatase gene; h, human; HRP, horseradish peroxidase; NRS, normal rabbit serum; TBS, Tris-buffered saline. 0013-7227/02/$15.00/0 The Journal of Clinical Endocrinology & Metabolism 87(6):2706 –2715 Printed in U.S.A. Copyright © 2002 by The Endocrine Society 2706 at Erskine Medical Library Univ. of Edinburgh on December 17, 2007 jcem.endojournals.org Downloaded from
Transcript of ERβ1 and the ERβ2 Splice Variant (ERβcx/β2) Are Expressed in Distinct Cell Populations in the...

ER�1 and the ER�2 Splice Variant (ER�cx/�2) AreExpressed in Distinct Cell Populations in the AdultHuman Testis
PHILIPPA T. K. SAUNDERS, MICHAEL R. MILLAR, SHEILA MACPHERSON, D. STEWART IRVINE,NIGEL P. GROOME, LEE R. EVANS, RICHARD M. SHARPE, AND GRAEME A. SCOBIE
Medical Research Council Human Reproductive Sciences Unit (P.T.K.S., M.R.M., S.M., D.S.I., R.M.S., G.A.S.), Centre forReproductive Biology, Edinburgh EH3 9ET, United Kingdom; and School of Biological & Molecular Sciences (N.P.G.,L.R.E.), Oxford Brookes University, Gypsy Lane Campus, Headington, Oxford OX3 0PB, United Kingdom
Estrogens can regulate germ cell function. Estrogen action ismediated via high affinity ERs; two subtypes (ER� and ER�)have been identified. We have shown previously that ER� isexpressed in nuclei of multiple human testicular cells. A vari-ant isoform of human (h) ER� (hER�cx/2), formed by alterna-tive splicing, has been identified in testicular cDNA librariesby two laboratories. The present study examined the expres-sion of wild-type (ER�1) and variant (ER�2) � receptors inhuman testes by 1) RT-PCR with isoform specific primers, and2) single and double immunohistochemistry using monoclo-nal antibodies raised against peptides unique to the C terminiof hER�1 and hER�2. PCR products specific for ER�1 andER�2 were amplified from cDNA pools prepared from humantestes and granulosa cells. On Western blots, the anti-ER�1monoclonal antibody bound to recombinant ER�1 and theanti-ER�2 monoclonal to recombinant hER�2. Neither boundto the other ER� isoform nor to recombinant ER�. ER�1 and
ER�2 proteins were both detected in human testis. Immuno-expression of ER�1 was most intense in pachytene spermato-cytes and round spermatids, whereas low levels of expressionwere detected in Sertoli cells, spermatogonia, preleptotene,leptotene, zygotene, and diplotene spermatocytes. Highestlevels of expression of ER�2 protein were detected in Sertolicells and spermatogonia with low/variable expression inpreleptotene, pachytene, and diplotene spermatocytes. No im-munostaining was detected in elongating spermatids. Mostinterstitial cells expressed more ER�2 than ER�1. It is spec-ulated that the cells most susceptible to modulation by estro-genic ligands are round spermatids in which levels of expres-sion of ER�1 are high. In contrast, expression of ER�2, anisoform that may act as a dominant negative inhibitor of ERaction, in Sertoli cells and spermatogonia, could protect thesecells from adverse effects of estrogens. (J Clin EndocrinolMetab 87: 2706–2715, 2002)
IT HAS BEEN proposed that estrogens may play a role inregulating spermatogenesis (see review by Ref. 1). For
example, Ebling et al. (2) found estrogen treatment of hpgmice stimulated spermatogenesis and Pentikainen et al. (3)reported that estrogen exposure reduced levels of germ cellapoptosis in isolated human seminiferous tubules consistentwith estrogen acting as a germ cell survival factor. The bio-synthesis of estrogens from androgens is catalyzed by thearomatase complex. Levels of estrogen within the male re-productive tract are higher than in the general circulationconsistent with production of estrogens (reviewed in Ref. 4).In the adult testis, the major site of aromatase expressionappears to be the Leydig cells (5, 6); however, aromataseactivity has also been detected within mature germ cells andspermatozoa (6–8). Male mice in which estrogen biosynthe-sis is impaired due to targeted disruption of the aromatasegene (ARKO) become infertile (9). Analyses of the testes ofthese animals has shown that round spermatids undergoapoptosis, show disturbances in acrosome formation, andfail to differentiate into mature elongate spermatids (9, 10).Consistent with estrogens acting as a germ cell survival fac-tor treatment of monkeys with an aromatase inhibitor also
results in a reduction in the progression of round to elongatespermatids (11).
Estrogen action is mediated via high affinity receptors thatare shifted to a transcriptionally active state after ligandbinding. Two forms of ER commonly known as � (ER�,NR3A1) and � (ER�, NR3A2) have been cloned from human(h) (12–14) and other species (15, 16). Both receptors areencoded by eight exons and are the products of two geneslocated on different chromosomes (17). Phylogenetic analysissuggests that they arose from a single duplication event atleast 450 million years ago (18). Like other members of thesteroid receptor superfamily, ER� and ER� share a commonarrangement of five structure-function domains, denoted Ato F (19). Key domains include the DNA binding domain (C),two transactivation domains [activation function (AF)-1 andAF-2, located in the N (A/B domain) and C-terminal (E/Fdomain) portions of the protein, and regions within the C andE domains involved in receptor dimerization (see reviews inRefs. 20 and 21). Studies in vitro have demonstrated thathomodimers (ER�-ER� or ER�-ER�), or heterodimers (ER�-ER�), can be formed when both isoforms are expressed in thesame cell (22, 23). In 1998, a novel human ER� variant, namedhER�cx, was identified by screening a human testis cDNAlibrary (Ref. 24; GenBank accession no. AB006589). The openreading frame of the protein was predicted to be identical tothat of full-length hER� (59 kDa, Ref. 14, GenBank accession
Abbreviations: AF, Activation function; ARKO, aromatase gene; h,human; HRP, horseradish peroxidase; NRS, normal rabbit serum; TBS,Tris-buffered saline.
0013-7227/02/$15.00/0 The Journal of Clinical Endocrinology & Metabolism 87(6):2706–2715Printed in U.S.A. Copyright © 2002 by The Endocrine Society
2706
at Erskine Medical Library Univ. of Edinburgh on December 17, 2007 jcem.endojournals.orgDownloaded from

AB006590) except for a truncation at the C-terminus leadingto the loss of 61 amino acids. The C-terminus of hER�cx wasfound to encode a unique 26 amino acid peptide, resultingin a protein containing 495 amino acids (�55 kDa). In aseparate series of experiments, Moore et al. (25) identifiedmRNAs encoding both this isoform of hER�, which theycalled hER�2 (GenBank accession no. AF051428), as well asthree further isoforms, which they named ER�3, ER�4, andER�5 in human tissues including testis. They showed that theER�2 variant could bind DNA both as a homodimer and asa heterodimer with hER�1 using gel shift assays. For the restof this paper, the hER� protein identified initially as thehomolog to rat ER� (13, 14, 17) will be referred to as ER�1and the ER�cx/ER�2 variant as ER�2 to distinguish themfrom each other. Note that the human ER�2 cloned by Lu etal. (GenBank accession no. AF124790) is not the same variantas the ER�cx/�2 described above, but a form of ER� thatlacks exon 5; this variant encodes a truncated ER� proteinlacking the entire ligand binding domain (26). A splice vari-ant isoform of ER� has also been identified in rodents and issometimes referred to as ER�2, or more recently as ER�ins(27–29). This variant is not identical to hER�2 but contains a54-bp insertion within the ligand binding domain (domain E)leading to the addition of 18 amino acids, this isoform has notbeen detected in human cDNAs prepared from normal hu-man ovary, endometrium, mammary gland (30, 31) or testis(our unpublished observations).
We have shown previously using both immunohistochem-istry and Western blotting that ER� protein is widely ex-pressed in the testis and reproductive system of the human,primate, and rodent (32–34). However, the antibody used forthose studies was raised against a peptide (P4) (35) in the Ddomain of hER� that is identical in both hER�1 and hER�2.We have recently confirmed that this antibody is capable ofrecognizing both these isoforms of ER� on Western blots(Scobie, G. S., and P. T. K. Saunders, unpublished observa-tions). As ER�2 was originally isolated from human testiscDNA libraries, and has been proposed to act as a negativeregulator of ER activity (24), we have extended our investi-gations using monoclonal antibodies specific for ER�1 andER�2. We show for the first time that both ER�1 and ER�2proteins are expressed in multiple cell types within the hu-man testis but that their patterns of expression are distinct.
Materials and MethodsTissue samples
Testicular tissues were obtained from men (n � 10) undergoingsurgical investigations for nonobstructive azoospermia or surgical cor-rection of vasectomy. The Lothian Reproductive Medicine Ethics com-mittee approved the protocols, and men gave informed consent. Addi-tional human tissues (n � 6) were obtained from the PeterboroughHospitals NHS Trust tissue bank. Samples for immunohistochemistrywere fixed in Bouins for 6–7 h and processed into paraffin wax usingstandard methods (36). A sample of RNA extracted from human gran-ulosa cells was a gift from Dr. Chris Harlow (Edinburgh University)human endometrial tissue was a gift from Prof. Hilary Critchley (Ed-inburgh University). Tissues for RNA extraction were snap frozen ondry ice or in liquid nitrogen and stored at �70 C before use.
Analysis of ER� mRNAs by RT-PCR
Frozen tissue samples (testicular biopsy and endometrium) wereground up under liquid nitrogen using a pestle and mortar, divided into
approximately 50 mg samples, and stored at �70 C. RNA was extractedusing Tri Reagent (Sigma, Poole, Dorset, UK) according to manufac-turer’s instructions, dissolved in ribonuclease-free water and 5 �g totalRNA subjected to reverse transcriptase using Expand Reverse Tran-scriptase (Roche Lewes, Sussex, UK) and an oligo deoxythymidineprimer according to the instructions supplied with the enzyme. First-strand cDNA was purified by heating to 100 C for 5 min, ribonucleasetreatment at 37 C for 15 min, and finally by passing through a “High PurePCR Purification” column (Roche). Purified cDNAs were quantified onAmersham Pharmacia Biotech Genequant (St. Albans, Hertfordshire,UK) and adjusted to a final concentration of 5 ng/�l in Tris/EDTA buffer(10 mm Tris/HCl, pH 7.5; 1 mm EDTA).
PCRs were performed using AGS Gold Taq (Hybaid, Ashford, Mid-dlesex, UK) with the following primers: ER� common 5� primer (exon 7)5�-GGCATCTCCTCCCAGCAGCA was used in combination with either aspecific ER�1 3� primer 5� CACTGAGACTGTGGGTTCTGGGA (amplifiedproduct 261 bp) or a specific ER�2 3� primer 5�-CACTGCTCCATCGTT-GCTTC (amplified product 156 bp). ER� cDNA was amplified with primersidentical to ER� specific set 1 (exon 1–3) published previously by Lau et al.(37) (amplified product 650 bp). Internal GAPDH primers were 5�-GAACGGGAAGCTCACTGGCAT and 5�-GTCCACCACCCTGTTGCTG-TAG (amplified product 301 bp). PCR conditions were as follows: 1 cycleof 94 C for 2 min followed by 30 cycles of 94 C for 30 sec, 58 C for 30 secand 72 C for 30 sec, with a final cycle of 72 C for 10 min, in a 0.2-ml HybaidSprint thermal cycler. PCR products were separated on 2% agarose gelsstained with ethidium bromide and then photographed.
Antibodies
Peptide P7 (CSPAEDSKSKEGSQNPQSQ) specific for hER�1 (wild-type,Refs. 14 and 17, GenBank accession no. AB006590) and peptide P8 (CMK-METLLPEATMEQ) specific for hER�2 (Refs. 24 and 25; GenBank accessionno. AB006589) were synthesized in the Center for Proteins and Peptides,Oxford Brookes University and conjugated to tuberculin. Conjugated pep-tides were used to immunize individual mice and monoclonal antibodiesprepared according to standard methods (38). Positive clones directedagainst ER�1 and ER�2 were identified by ELISA using recombinant hu-man ER�1 (P2466, PanVera, Madison, WI) or unconjugated peptide P8,respectively. Individual immunoglobulins (anti-ER�1 type IgG2a; antiER�2 type IgG1) were purified from culture media that had been dialyzedagainst 10 mm phosphate buffer (pH 7.2) using Hi-trap protein G Sepharosecolumns according to the manufacturer’s instructions (Amersham Phar-macia Biotech). The isolated IgGs were desalted on P10 columns (Amer-sham Pharmacia Biotech) that had been preequilibrated in PBS (pH 7.4)then mixed with 20% vol/vol glycerol and 0.02% wt/vol sodium azide(Sigma) and stored as 100 �l aliquots at �20 C. The ability of the antibodydirected against peptide P7 to detect expression of ER� in ovarian sectionsfrom human and marmoset monkey has been reported previously (35).
Preparation of recombinant ER�2
A full-length cDNA encoding hER�2 was amplified from humantestis cDNA prepared as described for RT-PCR analysis, using extensorHi-Fidelity PCR master Mix (ABgene, Epsom Surrey, UK) and 0.6 �m ofeach primer (5� ATG primer: 5�-GACATGGATATAAAAAACTCACCand 3� primer 5�-CACTGCTCCATCGTTGCTTC). PCR cycle conditionswere: 1 cycle of 94 C for 2 min followed by 30 cycles of 94 C for 30 sec,58 C for 30 sec and 72 C for 90 sec, with a final cycle of 72 C for 10 min.The resulting cDNA was cloned in frame into the pUni donor vector(pUni/V5-His-TOPO Echo Cloning System) according to the manufac-turer’s instructions (Invitrogen, Breda, The Netherlands). The full-lengthpUNI/�2 cDNA construct was recombined into the pRSET-E acceptorEcho bacterial expression vector (Invitrogen) by incubation of the do-nor/acceptor plasmids in the presence of CRE-recombinase according tothe protocol supplied by Invitrogen, transformed into TOP10 cells, andselected on kanamycin agar plates. Recombinant hER�2 protein wasmade by transforming the pRSETB2 plasmid into BL21(DE3)plysS (In-vitrogen) and growing the transformed bacteria in LB-medium contain-ing 1 mm IPTG for 2–3 h. Bacterial cells were separated by centrifugationand the resulting pellets were boiled in 1� lysis buffer (contains 50 mmTris/HCl, pH 6.8; 100 mm dithiothreitol; 2% SDS; 10% glycerol; and 0.1%bromophenol blue).
Saunders et al. • ER�1, and the ER�2 Splice Variant (ER�cx/�2) J Clin Endocrinol Metab, June 2002, 87(6):2706–2715 2707
at Erskine Medical Library Univ. of Edinburgh on December 17, 2007 jcem.endojournals.orgDownloaded from

Western analysis
Recombinant human ER�1 corresponding to short (�53 kDa) andlong (�59 kDa) forms of the wild-type form of the receptor (ER�1; 13,14) as well as full-length ER� were obtained from PanVera (MadisonWI). Recombinant ER�2 was prepared as above. Recombinant proteins(0.1 �g/lane ER�1L, ER�S, and ER� or 10 �l lystate ER�2) andprestained protein molecular weight markers (Bio-Rad Laboratories,Inc., Hercules, CA) were separated on denaturing minigels containingan acrylamide gradient from 4 to 20% (wt/vol) polyacrylamide (In-vitrogen) using Tris/glycine/SDS running buffer (25 mm Tris; 250 mmglycine, pH 8.3; 0.1% SDS). Proteins were transferred to immobilonmembrane using a Tris/glycine buffer (12 mm Tris; 96 mm glycine, pH8.3). Membranes were incubated overnight with the mouse monoclonalantibodies at 1 in 500 diluted in Tris-buffered saline (TBS)-Tween buffer(10 mm Tris; 150 mm NaCl; 0.05% Tween-20) containing 5% dried milk.Bound antibodies were detected using a horseradish peroxidase (HRP)-conjugated rabbit antimouse IgG and the electrochemiluminescencevisualization system (Amersham Pharmacia Biotech) according to themanufacturer’s instructions.
Immunohistochemistry
Immunolocalization of ER�1 and ER�2 used 5 �m paraffin tissuesections which were subjected to heat induced antigen retrieval (39) ina pressure cooker (Tefal, Nottingham, UK) containing 2 liters of nearboiling 0.01 m glycine/EDTA, pH 3.5. Thereafter, the sections werewashed and endogenous peroxidase blocked as described previously(33). Mouse anti-ER�1 (monoclonal 9) and anti-ER�2 (monoclonal 57/3)were each diluted 1 in 50 in normal rabbit serum (NRS, DiagnosticsScotland, Edinburgh, UK) diluted 1:4 in TBS (0.05 m Tris, pH 7.4; 0.85%saline) containing 5% BSA (NRS/TBS/BSA). Antibodies were incubatedon sections overnight at 4 C. Sections were washed twice for 5 min eachtime in TBS and incubated with biotinylated rabbit-antimouse IgG(DAKO Corp., Cambridge, UK) diluted 1:500 in NRS/TBS/BSA. Incu-bations lasted for 1 h and were followed by two washes in TBS (5 mineach). Thereafter, sections were incubated in avidin-biotin complex-HRP(DAKO Corp.) for 1 h, washed in TBS (2 � 5 min) and bound antibodiesvisualized by incubation with liquid 3,3�-diaminobenzidine tetra-hy-drochloride (catalog no. K3468, DAKO Corp.). Sections were counter-stained lightly with hematoxylin to enable identification of cell nucleicontaining low levels of immunopositive staining. Staining of sectionswas repeated at least twice; sections were then examined independentlyby two individuals and the identity of immunopositive cells and theintensity of immunostaining noted on a stage diagram of human sper-matogenesis (see Ref. 40). Images were captured using an OlympusCorp. Provis microscope (Olympus Corp. Optical Co., London, UK)equipped with a Kodak DCS330 camera (Eastman Kodak Co., Rochester,NY), stored on a Macintosh PowerPC computer and assembled usingPhotoshop 5.5 (Adobe, Mountain View, CA).
Double fluorescent immunohistochemistry
Sections were washed, subjected to antigen retrieval, and blocked asdescribed above. Washes detailed below were for 5 min each, andincubations were at room temperature unless otherwise specified.Mouse anti-ER�1 specific was diluted 1 in 20 in NRS/TBS/BSA (seeabove) and incubated on sections overnight at 4 C. Sections werewashed, incubated with biotinylated rabbit antimouse IgG (DAKOCorp.) diluted 1 in 500 in NRS/TBS/BSA for 30 min then washed in TBSfollowed by PBS. The fluorochrome streptavidin 546 Alexafluor (Mo-lecular Probes, Inc., Eugene, OR) diluted 1 in 200 in PBS was incubatedon slides for 2 h. Sections were washed in PBS then TBS, incubated withbiotin (4 drops/1 ml TBS) for 10 min, re-washed twice in TBS, andincubated with normal mouse IgG diluted 1 in 1000 in NRS/TBS/BSAfor 30 min. Sections were reblocked with NRS/TBS/BSA for 30 min thenincubated with mouse anti-ER�2 diluted 1 in 50 in NRS/TBS/BSAovernight at 4 C. Sections were washed twice in TBS, incubated withHRP conjugated rabbit-antimouse IgG1 subtype specific antiserum(Zymed Laboratories, Inc., South San Francisco, CA) diluted 1 in 100 inNRS/TBS/BSA for 1 h. Sections were washed in TBS containing 0.1%Tween followed by TBS and incubation with Tyr-Cy5 (from kit NEL745,NEN Life Science Products) for 10 min, which produced and enhanced
the second fluorescent signal. Sections were washed and mounted inPermafluor (Immunotech-Coulter, High Wycomb, Buckinghamshire,UK). Fluorescent images were captured on a Carl Zeiss (Jena, Germany)laser scanning microscope LSM 510. The alexafluor 546 was detectedusing the helium/neon 1 laser (excitation beam, 543 nm) and an emissionband pass filter 560–615 nm. The Cy 5 was detected with the helium/neon 2 laser (excitation beam 633 nm) and emission long band pass filter650.
ResultsDetection of both ER�1 and ER�2 in testicular samples
cDNAs specific for both ER�1 and ER�2 isoforms wereamplified from cDNA prepared by RT of total RNA fromhuman testis obtained from BD Biosciences Clontech (PaloAlto, CA) (Fig. 1, lanes 1 and 2) or prepared from a humantesticular biopsy (Fig. 1, lane 9), human granulosa cells(Fig. 1, lane 5) or human endometrium (Fig. 1, lane 8). Bothisoforms were identified in a cDNA library obtained fromStratagene (La Jolla, CA) (lane 4) and a pool of marathon-ready cDNAs prepared from human testes by BD BiosciencesClontech (lane 3). In all samples, the amount of the ER�1cDNA amplified appeared to be slightly higher than that ofER�2; this difference was most marked in the StratagenecDNA library and in granulosa cells but was not quantified.When the same panel of samples was tested using ER�-specific primers (Fig. 1C), the highest levels of ER� cDNAwas amplified from human endometrial tissue (lane 8, pos-itive control; Ref. 41) and low levels of expression detectedin pooled human granulosa cells (lane 6; Ref. 35). Amountsof ER� cDNA amplified from testicular samples after 30cycles of PCR were highly variable with no detectable signalin a testicular biopsy (lane 9), low levels in cDNAs preparedfrom testis RNA obtained from BD Biosciences Clontech(lanes 1 and 2), moderate amounts in a cDNA library fromStratagene (lane 4) and significant signal in a sample ofmarathon-ready cDNA (BD Biosciences Clontech) (lane 3).
Specificity of ER� monoclonal antibodies
On Western blots, the monoclonal antibodies directedagainst peptides at the C termini of ER�1 or ER�2 recognizedthe appropriate full-length recombinant proteins (Fig. 2, Aand B) but did not cross-react either with the alternativeisoform nor with recombinant ER� (lane �). Absorption withthe immunizing peptide abolished the positive reaction (notshown).
Immunoexpression of ER�1 and ER�2 in adulthuman testis
Although proteins recognized by the monoclonal antibod-ies specific for ER�1 and ER�2 were expressed in multiplecell nuclei within the human seminiferous epithelium, theirpattern of expression was strikingly different (Fig. 3, a andb). The most intense immunostaining for ER�1 was detectedin round spermatids (R, Fig. 3, c and e, g), but this was lostas they started to undergo nuclear condensation (Fig. 3e, s4).Round spermatids were immunonegative or very slightlyimmunopositive for ER�2 (Fig. 3d, R). ER�1 and ER�2 pro-teins were not detected in elongate spermatids (E). Pachytenespermatocytes (P) contained intense immunopositive stain-ing for ER�1 (Fig. 3e) but stained only slightly when incu-
2708 J Clin Endocrinol Metab, June 2002, 87(6):2706–2715 Saunders et al. • ER�1, and the ER�2 Splice Variant (ER�cx/�2)
at Erskine Medical Library Univ. of Edinburgh on December 17, 2007 jcem.endojournals.orgDownloaded from

bated with ER�2 monoclonal (see Fig. 3, b and f, labeled P),whereas diplotene pachytene spermatocytes contained lowlevels of both proteins (Fig. 3, labeled D). All A type sper-matogonia contained ER�1 and ER�2; immunoexpression ofER�2 was generally more intense and was particularly strik-ing in B type spermatogonia (Fig. 3f, arrows). A clear differ-
ential was seen between the levels of expression of the iso-forms within Sertoli cell nuclei (arrowheads); ER�1 was low/barely detectable (Fig. 3, c and e, g) but staining for ER�2 wasintense (Fig. 3, d and f, h). Low and variable levels of ex-pression of both isoforms were detected in peritubular my-oid and Leydig cells.
Fluorescent immunohistochemistry
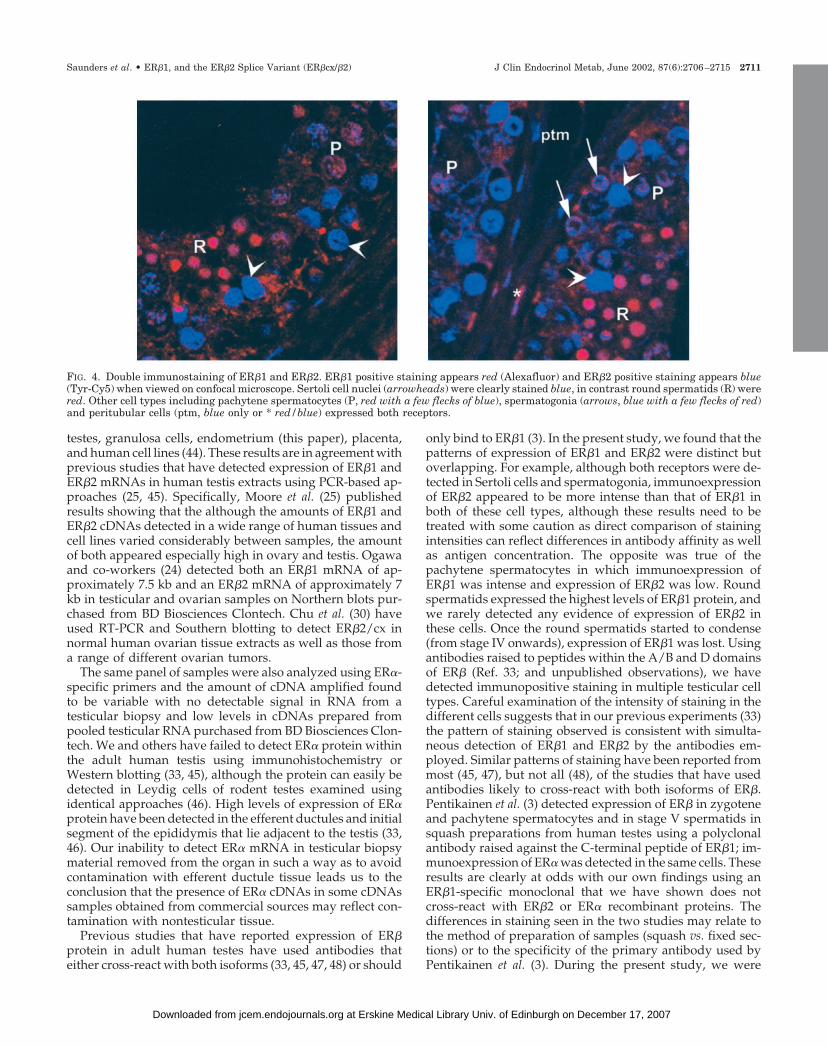
Colocalization of ER�1 and ER�2 on the same sections ofhuman testes proved technically challenging because bothantibodies were raised in mouse and the fluorescent detec-tion systems were less sensitive than those using color end-points. Results obtained (Fig. 4) confirmed and extendedresults of single immunohistochemistry. For example, Sertolicell nuclei (arrowheads) were clearly stained blue (ER�2 pos-itive), in contrast round spermatids (R) were red (ER�1 pos-itive). Other cell types including pachytene spermatocytes (P,red with a few flecks of blue), spermatogonia (arrows, blue witha few flecks of red) and peritubular myoid cells (ptm, blue or* red/blue) expressed both receptors in variable proportions.
Summary of patterns of expression of ER�1 and ER�2
To provide a summary of the patterns of expression of theER subtypes, the intensity of immunostaining observed (each
FIG. 1. ER�1 and ER�2 mRNAs are expressedin adult human testis. PCR amplification withprimers specific for the ER�1 (panel A, 261bp)and �2 (panel B, 156 bp) revealed that both weredetected in cDNA pools prepared from humantestis RNA (lanes 1 and 2), in marathon-readycDNA from human testis (BD Biosciences Clon-tech, lane 3), a cDNA library (Stratagene, lane4) and a testicular biopsy obtained in house(lane 9). Control tissues (human granulosa celland endometrial cDNAs) contained both ER�1and ER�2 (lanes 5 and 8). Negative controlsprepared without reverse transcriptase (lane 6)or lacking input cDNA (lane 7) were included.ER� cDNA (panel C) was amplified from humanendometrium (lane 8, positive control). Theamounts of ER� detected in testicular sampleswas highly variable and ranged from negative(biopsy, lane 9), or low (BD Biosciences ClontechRNA, lanes 1 and 2) to moderate (cDNA library,lane 4; marathon-ready cDNA, lane 3). GAPDHwas amplified from all samples containingcDNA (panel D, but not from the negative con-trols, lanes 6 and 7). One hundred-base pairmarkers were run on all gels.
FIG. 2. Specificity of monoclonal antibdodies. Monoclonal antibodiesdirected against peptides at the C-terminus of ER�1 and ER�2 wereincubated with recombinant ER�1 [long(L) and short (s)]; ER�2 orER�. No cross reactivity with ER� (�) was observed but anti-ER�1 (A)and anti-ER�2 (B) reacted specifically with the appropriate recom-binant protein(s) to which they were directed.
Saunders et al. • ER�1, and the ER�2 Splice Variant (ER�cx/�2) J Clin Endocrinol Metab, June 2002, 87(6):2706–2715 2709
at Erskine Medical Library Univ. of Edinburgh on December 17, 2007 jcem.endojournals.orgDownloaded from
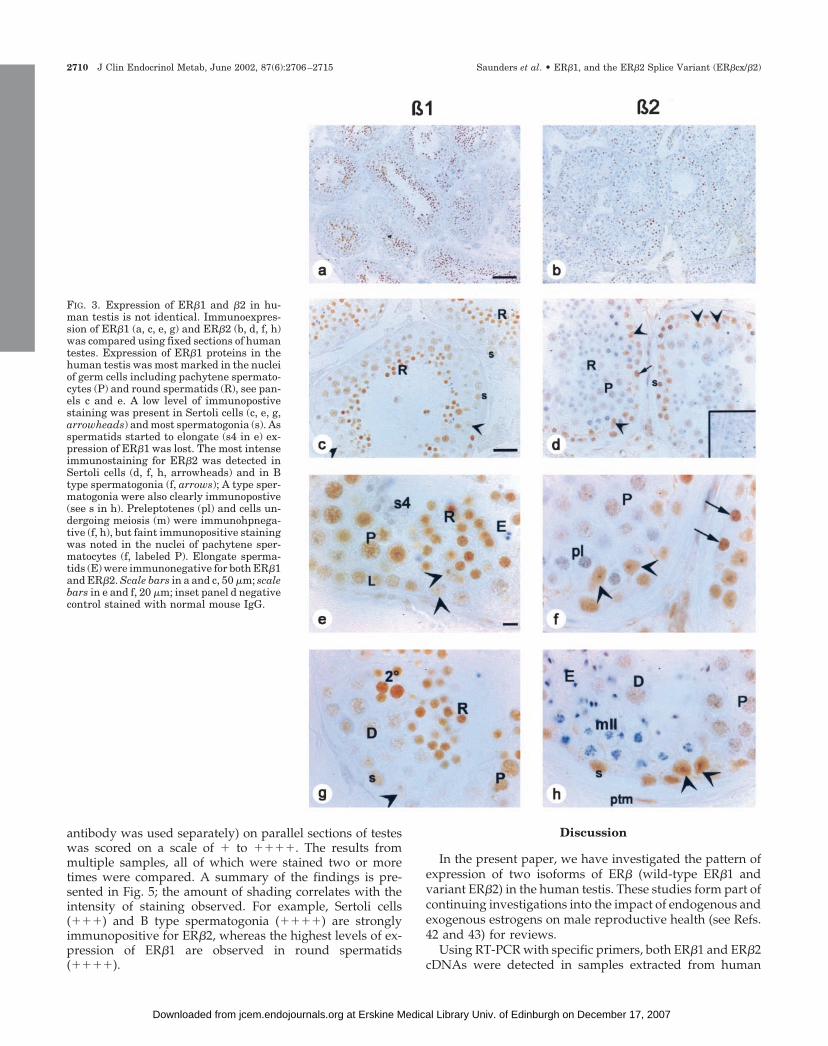
antibody was used separately) on parallel sections of testeswas scored on a scale of � to ����. The results frommultiple samples, all of which were stained two or moretimes were compared. A summary of the findings is pre-sented in Fig. 5; the amount of shading correlates with theintensity of staining observed. For example, Sertoli cells(���) and B type spermatogonia (����) are stronglyimmunopositive for ER�2, whereas the highest levels of ex-pression of ER�1 are observed in round spermatids(����).
Discussion
In the present paper, we have investigated the pattern ofexpression of two isoforms of ER� (wild-type ER�1 andvariant ER�2) in the human testis. These studies form part ofcontinuing investigations into the impact of endogenous andexogenous estrogens on male reproductive health (see Refs.42 and 43) for reviews.
Using RT-PCR with specific primers, both ER�1 and ER�2cDNAs were detected in samples extracted from human
FIG. 3. Expression of ER�1 and �2 in hu-man testis is not identical. Immunoexpres-sion of ER�1 (a, c, e, g) and ER�2 (b, d, f, h)was compared using fixed sections of humantestes. Expression of ER�1 proteins in thehuman testis was most marked in the nucleiof germ cells including pachytene spermato-cytes (P) and round spermatids (R), see pan-els c and e. A low level of immunopostivestaining was present in Sertoli cells (c, e, g,arrowheads) and most spermatogonia (s). Asspermatids started to elongate (s4 in e) ex-pression of ER�1 was lost. The most intenseimmunostaining for ER�2 was detected inSertoli cells (d, f, h, arrowheads) and in Btype spermatogonia (f, arrows); A type sper-matogonia were also clearly immunopostive(see s in h). Preleptotenes (pl) and cells un-dergoing meiosis (m) were immunohpnega-tive (f, h), but faint immunopositive stainingwas noted in the nuclei of pachytene sper-matocytes (f, labeled P). Elongate sperma-tids (E) were immunonegative for both ER�1and ER�2. Scale bars in a and c, 50 �m; scalebars in e and f, 20 �m; inset panel d negativecontrol stained with normal mouse IgG.
2710 J Clin Endocrinol Metab, June 2002, 87(6):2706–2715 Saunders et al. • ER�1, and the ER�2 Splice Variant (ER�cx/�2)
at Erskine Medical Library Univ. of Edinburgh on December 17, 2007 jcem.endojournals.orgDownloaded from
testes, granulosa cells, endometrium (this paper), placenta,and human cell lines (44). These results are in agreement withprevious studies that have detected expression of ER�1 andER�2 mRNAs in human testis extracts using PCR-based ap-proaches (25, 45). Specifically, Moore et al. (25) publishedresults showing that the although the amounts of ER�1 andER�2 cDNAs detected in a wide range of human tissues andcell lines varied considerably between samples, the amountof both appeared especially high in ovary and testis. Ogawaand co-workers (24) detected both an ER�1 mRNA of ap-proximately 7.5 kb and an ER�2 mRNA of approximately 7kb in testicular and ovarian samples on Northern blots pur-chased from BD Biosciences Clontech. Chu et al. (30) haveused RT-PCR and Southern blotting to detect ER�2/cx innormal human ovarian tissue extracts as well as those froma range of different ovarian tumors.
The same panel of samples were also analyzed using ER�-specific primers and the amount of cDNA amplified foundto be variable with no detectable signal in RNA from atesticular biopsy and low levels in cDNAs prepared frompooled testicular RNA purchased from BD Biosciences Clon-tech. We and others have failed to detect ER� protein withinthe adult human testis using immunohistochemistry orWestern blotting (33, 45), although the protein can easily bedetected in Leydig cells of rodent testes examined usingidentical approaches (46). High levels of expression of ER�protein have been detected in the efferent ductules and initialsegment of the epididymis that lie adjacent to the testis (33,46). Our inability to detect ER� mRNA in testicular biopsymaterial removed from the organ in such a way as to avoidcontamination with efferent ductule tissue leads us to theconclusion that the presence of ER� cDNAs in some cDNAssamples obtained from commercial sources may reflect con-tamination with nontesticular tissue.
Previous studies that have reported expression of ER�protein in adult human testes have used antibodies thateither cross-react with both isoforms (33, 45, 47, 48) or should
only bind to ER�1 (3). In the present study, we found that thepatterns of expression of ER�1 and ER�2 were distinct butoverlapping. For example, although both receptors were de-tected in Sertoli cells and spermatogonia, immunoexpressionof ER�2 appeared to be more intense than that of ER�1 inboth of these cell types, although these results need to betreated with some caution as direct comparison of stainingintensities can reflect differences in antibody affinity as wellas antigen concentration. The opposite was true of thepachytene spermatocytes in which immunoexpression ofER�1 was intense and expression of ER�2 was low. Roundspermatids expressed the highest levels of ER�1 protein, andwe rarely detected any evidence of expression of ER�2 inthese cells. Once the round spermatids started to condense(from stage IV onwards), expression of ER�1 was lost. Usingantibodies raised to peptides within the A/B and D domainsof ER� (Ref. 33; and unpublished observations), we havedetected immunopositive staining in multiple testicular celltypes. Careful examination of the intensity of staining in thedifferent cells suggests that in our previous experiments (33)the pattern of staining observed is consistent with simulta-neous detection of ER�1 and ER�2 by the antibodies em-ployed. Similar patterns of staining have been reported frommost (45, 47), but not all (48), of the studies that have usedantibodies likely to cross-react with both isoforms of ER�.Pentikainen et al. (3) detected expression of ER� in zygoteneand pachytene spermatocytes and in stage V spermatids insquash preparations from human testes using a polyclonalantibody raised against the C-terminal peptide of ER�1; im-munoexpression of ER� was detected in the same cells. Theseresults are clearly at odds with our own findings using anER�1-specific monoclonal that we have shown does notcross-react with ER�2 or ER� recombinant proteins. Thedifferences in staining seen in the two studies may relate tothe method of preparation of samples (squash vs. fixed sec-tions) or to the specificity of the primary antibody used byPentikainen et al. (3). During the present study, we were
FIG. 4. Double immunostaining of ER�1 and ER�2. ER�1 positive staining appears red (Alexafluor) and ER�2 positive staining appears blue(Tyr-Cy5) when viewed on confocal microscope. Sertoli cell nuclei (arrowheads) were clearly stained blue, in contrast round spermatids (R) werered. Other cell types including pachytene spermatocytes (P, red with a few flecks of blue), spermatogonia (arrows, blue with a few flecks of red)and peritubular cells (ptm, blue only or * red/blue) expressed both receptors.
Saunders et al. • ER�1, and the ER�2 Splice Variant (ER�cx/�2) J Clin Endocrinol Metab, June 2002, 87(6):2706–2715 2711
at Erskine Medical Library Univ. of Edinburgh on December 17, 2007 jcem.endojournals.orgDownloaded from

unable to obtain sufficient fresh testicular biopsies to allowus to undertake Western blotting experiments. We have pre-viously reported that proteins with molecular masses in therange 53–57 kDa were detectable in extracts from two humantesticular biopsies using a polyclonal antibody that willcross-react with both ER�1 and ER�2 (33). It is thereforepossible that the smaller protein could either be the short
form of ER�1 (53 kDa) or ER�2 (55 kDa), which we havesuccessfully identified in nuclear extracts from human en-dometrial tissue on Western blots using the specific ER�2monoclonal antibody (Scobie, G., unpublished observations).
In considering how expression of ER�2 might influenceestrogen activation of gene expression in the human testis, itis useful to review the information available on the functional
FIG. 5. Summary diagrams of the pattern of expression ofER�1 and ER�2 in human testes determined by specificimmunohistochemistry. The diagram was redrawn fromthat published by Clermont (40). The intensity of shadingreflects the intensity of the immunpositive reaction withinthe nuclei of the different cell types (negative to ����).
2712 J Clin Endocrinol Metab, June 2002, 87(6):2706–2715 Saunders et al. • ER�1, and the ER�2 Splice Variant (ER�cx/�2)
at Erskine Medical Library Univ. of Edinburgh on December 17, 2007 jcem.endojournals.orgDownloaded from

role of the region of ER�1 that is not present in hER�2 dueto the presence of an ER�2 specific exon 8 (24). Briefly, inhER�1 the wild-type exon 8 encodes part of domain E andall of domain F (amino acids 468 to 530, Ref. 14). Criticalamino acids within this region include several involved inligand binding that are conserved with ER�, for example,ER� G472 (� G521), ER� H475 (� 524) and ER� L476 (� 525)(49–51). The AF-2 domain that includes the twelfth helix ofthe protein, is also encoded by exon 8 (hER�1 amino acids476–500). Furthermore, the conformation adopted by helix12 following ligand binding is critical in determining recruit-ment of cofactors to the ligand-receptor complex (see reviewby Ref. 52). Consistent with the loss of these amino acids,Ogawa et al. (24) reported that when they transfected anER�2 construct into COS cells they were unable to detectbinding of E2 to whole cell extracts and that ER�2 was unableto induce expression of an ERE-CAT reporter construct in thepresence or absence of E2. All but one of the amino acidsidentified as key to the dimerization of ERs (50, 53) are notencoded by exon 8 and are therefore identical in both ER�1and ER�2. Consistent with the presence of both the DNAbinding and dimerization domains, Moore et al. (25) used gelshift assays to demonstrate that ER�2 bound to DNA con-taining a consensus ERE both as a homo- and as a het-erodimer with either ER� or ER�1. Ogawa et al. (24) foundthat ER�2 was incapable of interacting with TIF1a, a coregu-lator that has been shown to interact with ligand-activatedhER� via the AF-2 domain. Studies on murine ER� haveshown that the absence of an intact AF-2 domain in onepartner within the steroid receptor dimer is sufficient toimpair transcriptional activity in the full-length het-erodimeric partner (54). When ER�2 was cotransfected withER� (ratios 1:100 to 1:1), it acted as a dominant negativeinhibitor of ER�-induced transactivation (24), although, sur-prisingly, in similar assays cotransfection of ER�2 with ER�1did not appear to have any affect on reporter gene activationin the presence of E2. Alternative splicing of exons at the 3�ends of other steroid hormone receptor superfamily mem-bers such as the glucocorticoid receptor (55) has been re-ported previously. Loss of the 50 amino acids from the Cterminus of hGR� and their replacement with a novel 15amino acid results in formation of an hGR� isoform that, likeER�2, does not bind steroid (56). hGR� has been shown toheterodimerize with hGR� and to act as a dominant negativeregulator of hGR� activity (57). Taken together with thelimited data on ER�2 from transfection studies (24), thesefindings suggest that expression of ER�2 in cells containingER�1 might impair gene activation by the wild-type receptorfollowing ligand binding.
In the course of the present study, when we used primersspecific for the 5� and 3� ends of ER�1 and ER�2 to amplifyfull-length cDNAs, as well as products of the expected size,we usually amplified shorter cDNA products. Sequencing ofseveral of these confirmed that they lacked sequences cor-responding to exon 5 but included those within exon 6 (ourunpublished observations). ER� and ER� isoforms that con-tain exon 5 deletions have been identified previously inbreast cancer cell lines and tumors (58–60). Inoue et al. (26)have identified ER�1 transcripts lacking exon 5 (ER�1�5) inhuman testis cDNA, and we have shown that ER�2�5 tran-
scripts are also formed. The loss of exon 5 results in intro-duction of a premature stop codon in exon 6; the proteinformed from the ER�1�5 and ER�2�5 cDNAs should beidentical. The ER�1�5 and ER�2�5 proteins would not bedetected by either of the antibodies used in the current study,as their sequence does not include the peptides used forimmunization. Hall and McDonnell (61) have demonstratedthat, unlike ER�, the A/B region of ER� does not contain astrong AF-1 domain, but instead they suggest this regionmight act as a repressor. The presence of intact AF-1 andDNA binding domains in both ER�2, and in the exon 5deleted variants, would be consistent with the in vitro datashowing they can act as dominant negative receptors whencotransfected with wild-type ER� and/or ER� (24, 26). Weplan further studies to investigate whether ER�2 can alsoinhibit transactivation by ER�1 at AP-1 sites.
Based on the results from several studies including thosein �ERKO mice (62), it has been proposed that one of thephysiological roles of ER� is to act as a negative regulatorypartner for ER� (29, 61, 62). It has been reported that therodent ER� variant (ER�ins), which has lower affinity for E2than wtER�, also acts as a dominant negative receptor whenheterodimerized with wild-type ER� (28, 63). Expression ofER�ins in the rat mammary gland is up-regulated duringlactation, and double immunohistochemistry has found thevariant colocalized with ER� in up to 80% of the epithelialcells. Saji and colleagues (29) have proposed that expressionof ER�ins quenches ER� in these cells making them insen-sitive to estrogens. Expression of ER�ins in the rodent testishas not been investigated to date, but we speculate thatexpression of ER�2 in the human testis may play a similarrole to that of ER�ins in the rodent mammary gland andserve to protect cells from estrogen action. If this is the case,then in adult human testes both Sertoli cells and spermato-gonia may be insensitive to estrogens as they express anexcess of ER�2 compared with ER�1.
In conclusion, we have demonstrated that ER�1, the func-tional wild-type receptor, and ER�2, a splice variant receptorthat lacks the ability to bind E2 or to recruit cofactors via theAF-2 domain, are both expressed in the human testis. Wespeculate that the expression of ER�2 in some cells may affecttheir ability to respond to endogenous or exogenous estro-gens. Expression of ER�1 in pachytene spermatocytes andround spermatids would be consistent with evidence fromrodent studies that points to estrogens having a direct impacton the survival and maturation of germ cells.
Acknowledgments
We thank Prof. H. Critchley and Dr. C. Harlow for making samplesavailable.
Received October 11, 2001. Accepted March 7, 2002.Address all correspondence and requests for reprints to: Dr. Philippa
T. K. Saunders, Medical Research Council Human Reproductive Sci-ences Unit, Center for Reproductive Biology, 37 Chalmers Street, Ed-inburgh EH3 9ET, United Kingdom. E-mail: [email protected].
References
1. O’Donnell L, Robertson KM, Jones ME, Simpson ER 2001 Estrogen andspermatogenesis. Endocr Rev 22:289–318
Saunders et al. • ER�1, and the ER�2 Splice Variant (ER�cx/�2) J Clin Endocrinol Metab, June 2002, 87(6):2706–2715 2713
at Erskine Medical Library Univ. of Edinburgh on December 17, 2007 jcem.endojournals.orgDownloaded from

2. Ebling FJP, Brooks AN, Cronin AS, Ford H, Kerr JB 2000 Estrogenic inductionof spermatogenesis in the hypogonadal mouse. Endocrinology 141:2861–2869
3. Pentikainen V, Erkkila K, Suomalainen L, Parvinen M, Dunkel L 2000Estradiol acts as a germ cell survival factor in the human testis in vitro. J ClinEndocrinol Metab 85:2057–2067
4. Hess RA 2000 Oestrogen in fluid transport in efferent ducts of the malereproductive tract. Rev Reprod 5:84–92
5. Payne A, Kelch R, Musich S, Halpern M 1976 Intratesticular site of aroma-tization in the human. J Clin Endocrinol Metab 42:1081–1087
6. Turner KJ, Macpherson S, Millar MR, McNeilly AS, Williams K, Cranfield M,Groome NP, Sharpe RM, Fraser HM, Saunders PT 2002 Development andvalidation of a new monoclonal antibody to mammalian aromatase. J Endocrinol172:21–30
7. Nitta H, Bunick D, Hess RA, Janulis L, Newton SC, Millette CF, Osawa Y,Shizuta Y, Toda K, Bahr JM 1993 Germ cells of the mouse testis express P450aromatase. Endocrinology 132:1396–1401
8. Hess RA, Bunick D, Bahr JM 1995 Sperm, a source of estrogen. Environ HealthPerspect 103:59–62
9. Robertson K, O’Donnell L, Jones ME, Meachem SJ, Boon WC, Fisher CR,Graves KH, McLachlan RI, Simpson ER 1999 Impairment of spermatogenesisin mice lacking a functional aromatase (CYP19) gene. Proc Natl Acad Sci USA96:7986–7991
10. Robertson KM, Simpson ER, Lacham-Kaplan O, Jones MEE 2001 Charac-terization of the fertility of male aromatase knockout mice. J Androl 22:825–830
11. Shetty G, Krishamurthy H, Krishnamurthy HN, Bhatnagar AS, Moudgal NR1998 Effect of long-term treatment with aromatase inhibitor on testicular func-tion of adult bonnet monkeys (M. radiata). Steroids 63:414–420
12. Green S, Walter P, Kumar V, Krust A, Bornert J-M, Argos P, Chambon P 1986Human oestrogen receptor cDNA: sequence, expression and homology tov-erb-A. Nature 320:134–139
13. Mosselman S, Polman J, Dijkema R 1996 ER�: identification and character-ization of a novel human estrogen receptor. FEBS Lett 392:49–53
14. Ogawa S, Inoue S, Watanabe T, Hiroi H, Orimo A, Hosoi T, Ouchi Y,Muramatsu M 1998 The complete primary structure of human estrogen re-ceptor beta (hER�) and its heterodimerization with ER � in vivo and in vitro.Biochem Biophys Res Commun 243:122–126
15. Kuiper GGJM, Enmark E, Pelto-Hukko M, Nilsson S, Gustafsson J-A 1996Cloning of a novel estrogen receptor expressed in rat prostate. Proc Natl AcadSci USA 93:5925–5930
16. White R, Lees J, Needham M, Ham J, Parker M 1987 Structural organizationand expression of the mouse estrogen receptor. Mol Endocrinol 1:735–744
17. Enmark E, Pelto-Huikko M, Grandien K, Lagercrantz S, Lagercrantz J, FriedG, Nordenskjold M, Gustafsson JA 1997 Human estrogen receptor �-genestructure, chromosomal localization, and expression pattern. J Clin EndocrinolMetab 82:4258–4265
18. Kelley ST, Thackray VG 1999 Phylogenetic analyses reveal ancient duplica-tion of estrogen receptor isoforms. J Mol Evolution 49:609–614
19. Beato M, Klug J 2000 Steroid hormone receptors: an update. Hum ReprodUpdate 6:225–236
20. Saunders PTK 1998 Oestrogen receptor � (ER�). Rev Reprod 3:164–17121. Pettersson K, Gustafsson J-A 2001 Role of estrogen receptor beta in estrogen
action. Annu Rev Physiol 63:165–19222. Cowley SM, Hoare S, Mosselman S, Parker SG 1997 Estrogen receptors � and
� form heterodimers on DNA. J Biol Chem 272:19858–1986223. Pettersson K, Grandien K, Kuiper GGJM, Gustafsson J-A 1997 Mouse es-
trogen receptor � forms estrogen receptor response element-binding het-erodimers with estrogen receptor �. Mol Endocrinol 11:1486–1496
24. Ogawa S, Inoue S, Watanabe T, Orimo A, Hosoi T, Ouchi Y, Muramatsu M 1998Molecular cloning and characterization of human estrogen receptor bcx: a po-tential inhibitor of estrogen action in human. Nucleic Acids Res 26:3505–3512
25. Moore JT, McKee DD, Slentz-Kesler K, Moore LB, Jones SA, Horne EL, SuJL, Kliewer SA, Lehmann JM, Willson TM 1998 Cloning and characterisationof human estrogen receptor � isoforms. Biochem Biophys Res Commun 247:75–78
26. Inoue S, Ogawa S, Horie K, Hoshino S, Goto W, Hosoi T, Tsutsumi O,Muramatsu M, Ouchi Y 2000 An estrogen receptor � isoform that lacks exon5 has dominant negative activity on both ER� and ER�. Biochem Biophys ResCommun 279:814–819
27. Chu S, Fuller PJ 1997 Identification of a splice variant of the rat estrogenreceptor � gene. Mol Cell Endocrinol 132:195–199
28. Petersen DN, Tkalcevic GT, Koza-Taylor PH, Turi TG, Brown TA 1998Identification of estrogen receptor �2, a functional variant of estrogen receptor� expressed in normal rat tissues. Endocrinology 139:1082–1092
29. Saji S, Sakaguchi H, Andersson S, Warner M, Gustafsson J-A 2001 Quan-titative analysis of estrogen receptor proteins in rat mammary gland. Endo-crinology 142:3177–3186
30. Chu S, Mamers M, Burger HG, Fuller PJ 2000 Estrogen receptor isoform geneexpression in ovarian stromal and epithelial tumors. J Clin Endocrinol Metab85:1200–1205
31. Lu B, Leygue E, Dotzlaw H, Murphy LC, Watson PH 1998 Estrogen receptor-�mRNA variants in human and murine tissues. Mol Cell Endocrinol 138:199–203
32. Saunders PT, Fisher JS, Sharpe RM, Millar MR 1998 Expression of oestrogenreceptor � (ER �) occurs in multiple cell types, including some germ cells, inthe rat testis. J Endocrinol 156:R13–R17
33. Saunders PTK, Sharpe RM, Williams K, Macpherson S, Urquart H, IrvineDS, Millar MR 2001 Differential expression of oestrogen receptor alpha andbeta proteins in the testes and male reproductive system of human and non-human primates. Mol Hum Reprod 7:227–236
34. Atanassova N, McKinnell C, Williams K, Turner KJ, Fisher JS, SaundersPTK, Millar MR, Sharpe RM 2000 Age-, cell- and region-specific immuno-expression of ER� (but not ER�) during postnatal development of the epi-didymis and vas deferens of the rat and disruption of this pattern by neonataltreatment with diethylstilbestrol. Endocrinology 142:874–876
35. Saunders PTK, Millar MR, Williams K, Macpherson S, Harkiss D, AndersonRA, Orr B, Groome NP, Scobie G, Fraser HM 2000 Differential expression ofestrogen receptor-� and -� and androgen receptor in the ovaries of marmosetand human. Biol Reprod 63:1098–1105
36. Millar MR, Sharpe RM, Maguire SM, Saunders PTK 1993 Cellular localisa-tion of messenger RNAs in rat testis: application of digoxigenin labelled probesto embedded tissue. Cell Tissue Res 273:269–277
37. Lau K-M, Mok SC, Ho S-M 1999 Expression of human estrogen receptor a and-�, progesterone receptor, and androgen receptor mRNA in normal and ma-lignant ovarian epithelial cells. Proc Natl Acad Sci USA 96:5722–5727
38. Groome NP, Lawrence M 1991 Preparation of monoclonal antibodies to the�A subunit of ovarian inhibin using a synthetic peptide immunogen. Hybrid-oma 10:309–312
39. Norton AJ, Jordan S, Yeomans P 1994 Brief, high-temperature heat denatur-ation (pressure cooking): a simple and effective method of antigen retrieval forroutinely processed tissues. J Pathol 173:371–379
40. Clermont Y 1963 The cycle of the seminiferous epithelium in man. AmJ Anat112:35–51
41. Critchley HOD, Brenner RM, Drudy TA, Williams KA, Nayak NR, MillarMR, Saunders PTK 2001 Estrogen receptor �, but not estrogen receptor �, ispresent in the vascular endothelium of the human and nonhuman primateendometrium. J Clin Endocrinol Metab 86:1370–1378
42. Miller WR, Sharpe RM 1998 Environmental oestrogens and human repro-ductive cancers. Endocr-Rel Cancer 5:69–96
43. Williams K, McKinnell C, Saunders PTK, Walker M, Fisher JS, Turner KJ,Atanassova N, Sharpe M 2001 Neonatal exposure to potent and environmentaloestrogens and abnormalities of the male reproductive system in the rat:evidence for the importance of the androgen-oestrogen balance and assess-ment of relevance to man. Hum Reprod Update 7:236–247
44. Scobie GS, Macpherson S, Millar MR, Groome NP, Romana PG, SaundersPTK, Human estrogen receptors: differential expression of ER� and � and theidentification of ER� variants. Steroids, in press
45. Makinen S, Makela S, Weihua Z, Warner M, Rosenlund B, Salmi S, HovattaO, Gustafsson JA 2001 Localization of oestrogen receptors � and � in humantestis. Mol Hum Reprod 7:497–503
46. Fisher J, Millar MR, Majdic G, Saunders PTK, Fraser HM, Sharpe RM 1997Immunolocalisation of oestrogen receptor-� (ER�) within the testis and ex-current ducts of the rat and marmoset monkey from perinatal life to adulthood.J Endocrinol 153:485–495
47. Taylor AH, Al-Azzawi F 2000 Immunolocalisation of oestrogen receptor � inhuman tissues J Mol Endocrinol 24:145–155
48. Pelletier G, El-Alfy M2 2000 Immunocytochemical localization of estrogenreceptor � and � in human reproductive organs. J Clin Endocrinol Metab85:4835–4840
49. Ekena K, Weis KE, Katzenellenbogen JA, Katzenellenbogen BS 1997 Dif-ferent residues of the human estrogen receptor are involved in the recognitionof structurally diverse estrogens and anti-estrogens. J Biol Chem 272:5069–5075
50. Tanenbaum DM, Wang Y, Williams SP, Sigler PB 1998 Crystallographiccomparison of the estrogen and progesterone receptor’s ligand binding do-mains. Proc Natl Acad Sci USA 95:5998–6003
51. Pike ACW, Brzozowski AM, Hubbard RE, Bonn T, Thorsell AG, EngstromO, Ljunggren J, Gustafsson JA, Carlquist M 1999 Structure of the ligand-binding domain of oestrogen receptor � in the presence of a partial agonist anda full antagonist. EMBO J 18:4608–4618
52. Klinge CM 2000 Estrogen receptor interaction with co-activators and co-repressors. Steroids 65:227–251
53. Lees JA, Fawell SE, White R, Parker MG 1990 A 22-aminoacid peptide restoresDNA-binding activity to dimerization-defective mutants of the estrogen re-ceptor. Mol Cell Biol 10:5529–5531
54. Tremblay GB, Tremblay A, Labrie F, Giguere V 1999 Dominant activity ofactivation function (AF-1) and differential stoichiometric requirements forAF-1 and -2 in the estrogen receptor �-� heterodimeric complex. Mol Cell Biol19:1919–1927
55. Hollenberg SM, Wienberger C, Ong ES, Cerelli G, Oro A, Lebo R, ThompsonEB, Rosenfeld MG, Evans RM 1985 Primary stucture and expression of afunctional human glucocorticoid receptor cDNA. Nature 318:635–641
56. Oakley RH, Sar M, Cidlowski JA 1996 The human glucocorticoid receptor �isoform. Expression, biochemical properties and putative function. J Biol Chem271:9550–9559
2714 J Clin Endocrinol Metab, June 2002, 87(6):2706–2715 Saunders et al. • ER�1, and the ER�2 Splice Variant (ER�cx/�2)
at Erskine Medical Library Univ. of Edinburgh on December 17, 2007 jcem.endojournals.orgDownloaded from

57. Oakley RH, Jewell CM, Yudt MR, Bofetiado DM, Cidlowski JA 1999 Thedominant negative activity of the human glucocorticoid receptor � isoform.J Biol Chem 274:27857–27866
58. Fuqua SA, Schiff R, Parra I, Friedrichs WE, Su JL, McKee DD, Slentz-Kesler K,Moore LB, Willson TM, Moore JT 1999 Expression of wild-type estrogen receptor� and variant isoforms in human breast cancer. Cancer Res 59:5425–5428
59. Fuqua SA, Fitzgerald SD, Chamness GC, Tandon AK, McDonnell DP,Nawaz Z, O’Malley BW, McGuire WL 1991 Variant human breast tumorestrogen receptor with constitutive transcriptional activity. Cancer Res 51:105–109
60. Vladusic EA, Hornby AE, Guerra-Vladusic FK, Lupu R 1998 Expression ofestrogen receptor � messenger RNA variant in breast cancer. Cancer Res58:210–214
61. Hall JM, McDonnell DP 1999 The estrogen receptor �-isoform (ER�) of thehuman estrogen receptor modulates ER� transcriptional activity and is a keyregulator of the cellular response to estrogens and antiestrogens. Endocrinol-ogy 140:5566–5578
62. Weihua Z, Saji S, Makinen S, Cheng G, Jensen EV, Warner M, GustafssonJA 2000 Estrogen receptor (ER) �, a modulator of ER� in the uterus. Proc NatlAcad Sci USA 97:5936–5941
63. Maruyama K, Endoh H, Sasaki-Iwaoka H, Kanou H, Shimaya E, HashimotoS, Kato S, Kawashima H 1998 A novel isoform of rat estrogen receptor � with18 amino acid insertion in the ligand binding domain as a putative dominantnegative regulator of estrogen action. Biochem Biophys Res Commun 246:142–147
Saunders et al. • ER�1, and the ER�2 Splice Variant (ER�cx/�2) J Clin Endocrinol Metab, June 2002, 87(6):2706–2715 2715
at Erskine Medical Library Univ. of Edinburgh on December 17, 2007 jcem.endojournals.orgDownloaded from




















