
2011 Standard For Performance Rating Of Air Terminals - AHRI
Upload
independentCategory
view
0download
0
Protein synthesis in axons and terminals: signi®cance formaintenance, plasticity and regulation of phenotype
With a critique of slow transport theory
Jaime Alvareza,*, Antonio Giudittab, Edward Koenigc
aDepartamento de BiologõÂa Celular y Molecular, Facultad de Ciencias BioloÂgicas, Ponti®cia Universidad CatoÂlica de Chile, Santiago, ChilebDipartimento di Fisiologia Generale e Ambientale, Universita di Napoli, ``Federico II,'' 80134, Napoli, Italy
cDepartment of Physiology and Biophysics, University at Bu�alo, Bu�alo, NY, 14214, USA
Received 28 September 1999
Abstract
This article focuses on local protein synthesis as a basis for maintaining axoplasmic mass, and expression of plasticity inaxons and terminals. Recent evidence of discrete ribosomal domains, subjacent to the axolemma, which are distributed atintermittent intervals along axons, are described. Studies of locally synthesized proteins, and proteins encoded by RNA
transcripts in axons indicate that the latter comprise constituents of the so-called slow transport rate groups. A comprehensivereview and analysis of published data on synaptosomes and identi®ed presynaptic terminals warrants the conclusion that acytoribosomal machinery is present, and that protein synthesis could play a role in long-term changes of modi®able synapses.
The concept that all axonal proteins are supplied by slow transport after synthesis in the perikaryon is challenged because theunderlying assumptions of the model are discordant with known metabolic principles. The ¯awed slow transport model issupplanted by a metabolic model that is supported by evidence of local synthesis and turnover of proteins in axons. Acomparison of the relative strengths of the two models shows that, unlike the local synthesis model, the slow transport model
fails as a credible theoretical construct to account for axons and terminals as we know them. Evidence for a dynamic anatomyof axons is presented. It is proposed that a distributed ``sprouting program,'' which governs local plasticity of axons, is regulatedby environmental cues, and ultimately depends on local synthesis. In this respect, nerve regeneration is treated as a special case
of the sprouting program. The term merotrophism is proposed to denote a class of phenomena, in which regional phenotypechanges are regulated locally without speci®c involvement of the neuronal nucleus. 7 2000 Elsevier Science Ltd. All rightsreserved.
Contents
1. Overview . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31.1. Proposed model of axon . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
2. Maintenance of the axon: early views . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42.1. Historical synopsis. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
Progress in Neurobiology 62 (2000) 1±62
0301-0082/00/$ - see front matter 7 2000 Elsevier Science Ltd. All rights reserved.
PII: S0301-0082(99 )00062 -3
www.elsevier.com/locate/pneurobio
* Corresponding author. Tel.: +562-686-2609; fax: +562-686-2717.
E-mail address: [email protected] (J. Alvarez).
Abbreviations: AChE, acetylcholinesterase; ActD, actinomycin D; BAPTA-AM, a chelating agent that permeates the cell membrane;
BW284c51, speci®c inhibitor of AChE; CAP, chloramphenicol; CDCH, caudodorsal cell hormone; CTEM, conventional transmission electron
microscopy; CXM, cycloheximide; EM, electron microscope or microscopy; ER, endoplasmic reticulum; ESI, electron spectroscopic imaging;
ISH, in situ hybridization; LTP, long-term potentiation; mGluR, metabotropic glutamate receptor; NF-H, -M, -L, neuro®lament subunit, heavy,
medium, and light, respectively; PCR, polymerase chain reaction; RER, rough ER; RNP, ribonucleoprotein particle; RT-PCR, reverse transcrip-
tase PCR; S, svedberg; SER, smooth ER; SRP, signal recognition particle; TH, tyrosine hydroxylase.

2.2. Slow axoplasmic transport . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
2.2.1. Origin of slow transport model. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 52.2.2. Flaws of the slow transport model . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
3. Local protein synthesis in axons . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 73.1. Metabolic studies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7
3.1.1. The Mauthner (M-) cell axon as a model . . . . . . . . . . . . . . . . . . . . . . . . . . . . 73.1.2. Studies in mammalian nerves . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7
3.1.3. Protein synthesis in invertebrate axons . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 83.2. Restricted ribosomal domains in vertebrate axons . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
3.2.1. Periaxoplasmic plaques in gold®sh spinal axons . . . . . . . . . . . . . . . . . . . . . . . 12
3.2.2. Periaxoplasmic plaques in mammalian axons. . . . . . . . . . . . . . . . . . . . . . . . . 143.2.3. Implications of cortical actin cytoskeleton in plaque domains . . . . . . . . . . . . . 183.2.4. Ribosomes in terminal boutons of Mauthner axon collaterals . . . . . . . . . . . . . 18
3.3. RNA transcripts in axoplasm . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 183.3.1. Squid data . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 183.3.2. Mature vertebrate axons . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 193.3.3. Small cytoplasmic RNAs in axons . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
3.3.4. Neurons in culture . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 213.4.5. Growing versus mature axons . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 213.4.6. Glial source of axonal RNA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22
4. Local protein synthesis in presynaptic terminals . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 234.1. Historical introduction. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23
4.2. Early studies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 234.2.1. Factors a�ecting synaptosomal protein synthesis . . . . . . . . . . . . . . . . . . . . . . 254.2.2. Commentary. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26
4.3. Alternative interpretations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 264.3.1. CAP-resistant, CXM-sensitive protein synthesis in mitochondria and
synaptosomes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 264.3.2. A CAP-sensitive extramitochondrial system of protein synthesis . . . . . . . . . . . 26
4.3.3. Nonribosomal dependent incorporation of amino acids into synaptosomalproteins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27
4.3.4. Non-presynaptic ribosome-containing particles in synaptosomes account for most
of the CXM-sensitive translational activity . . . . . . . . . . . . . . . . . . . . . . . . . . 274.3.5. Commentary. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28
4.4. Recent ®ndings . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28
4.5. E�ects of chemical and electrical stimulation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 314.5.1. Commentary. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32
4.6. Miscellaneous observations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32
4.7. RNA in presynaptic endings. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 324.7.1. Synaptosomal mitochondrial RNA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 334.7.2. Synaptosomal extramitochondrial RNA . . . . . . . . . . . . . . . . . . . . . . . . . . . . 334.7.3. Commentary. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34
4.8. Concluding commentary . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34
5. Essay on the signi®cance of local protein synthesis in axons . . . . . . . . . . . . . . . . . . . . . . . . . 35
5.1. Local synthesis as a model for maintaining axoplasm . . . . . . . . . . . . . . . . . . . . . . . . . 355.1.1. The model . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 355.1.2. Slow wave of radioactivity and stationary proteins. . . . . . . . . . . . . . . . . . . . . 35
5.2. Explanatory power: slow transport versus local synthesis models. . . . . . . . . . . . . . . . . 375.3. Proposed experiment to test slow transport and local synthesis models . . . . . . . . . . . . 39
6. Dynamic anatomy of the axon and role of Schwann cells . . . . . . . . . . . . . . . . . . . . . . . . . . . 39
6.1. Axonal microtubules . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 396.1.1. Relation to axon caliber . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 396.1.2. Regional variations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40
6.1.3. Role in axons . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 406.1.4. Environmental regulation. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 406.1.5. Regulation by Schwann cell . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41
6.1.6. Regulation by electrical activity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±622

6.1.7. E�ects of inhibitors of protein synthesis or taxol . . . . . . . . . . . . . . . . . . . . . . 42
6.1.8. Slow transport model explanation of local changes . . . . . . . . . . . . . . . . . . . . 426.1.9. Local synthesis model explanation of local changes . . . . . . . . . . . . . . . . . . . . 43
6.2. Axon caliber . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44
6.2.1. Schwann cells and caliber . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 446.2.2. Trembler mouse as a case study . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 446.2.3. Axon caliber from perspectives of slow transport and local synthesis models . . 44
6.3. Sprouting of axons in vertebrates . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45
6.3.1. Altered Schwann cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 456.3.2. Protease inhibitors and acetylcholinesterase . . . . . . . . . . . . . . . . . . . . . . . . . . 456.3.3. Regeneration and the sprouting program . . . . . . . . . . . . . . . . . . . . . . . . . . . 46
6.3.4. Local synthesis and the sprouting program . . . . . . . . . . . . . . . . . . . . . . . . . . 466.3.5. The Wlds mutant mouse as a case study . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47
6.4. Sprouting of axons in invertebrates. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48
6.5. Plasticity of the terminal arborization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49
7. Autonomy of axons. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 51
8. Final commentary . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 52
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 52
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 52
1. Overview
What distinguishes the nerve cell from cells in gen-eral, and makes it unique is its cellular anatomy.While dimensions of most cells are in a range of tensof micrometers, nerve cells have axons which mayextend a meter or more in man and other large ver-tebrates. With growth and maturation during neuro-genesis, axons become extended cellular appendagesthat serve as the functional circuitry of the nervoussystem. These lines of communication must be stabil-ized and maintained throughout life, while still retain-ing a potential for structural and functional plasticity,since axons remodel as they encounter external modu-lating signals.
As the most highly polarized of cells, the anatomyof the neuron creates a number of logistical problemsfrom the standpoint of supplying proteins to distalreaches of axons. A supply of proteins is not only arequisite for maintaining axoplasmic mass and viabilityin a steady state, but serves also to satisfy growthrequirements associated with local dynamic structuralchanges that are de®ned ordinarily as plasticity. Axo-nal sprouting is a prominent example of such adynamic state. It entails a regional transformation of across-linked cytoskeleton, normally organized for stab-ility and high tensile strength that is characteristic ofthe mature axon, into a dynamically organized cytos-keleton characteristic of motile structures. This trans-formation is referred to in the present context as the`sprouting program'. We will present and illustrate
nerve regeneration as a special case of the sproutingprogram (see below).
For many years, a pervasive doctrine has held swayin the literature and textbooks, which states as itsbasic tenet that all axoplasmic proteins are supplied bythe cognate cell body at slow rates of transport. Theunderlying assumption that the axon lacks an en-dogenous protein synthesizing machinery was largelyarrived at by default because of a failure of conven-tional electron microscopy (EM) to identify ribosomesin axons (see below). Unfortunately, the dominance ofthis view not only grossly minimized the complexity ofthe biology of the axon, but also was instrumental ineclipsing a very substantial body of evidence docu-menting the presence of an endogenous protein synthe-sizing machinery in axons of principal neurons thatmake up the `hard wiring' of the nervous system.
1.1. Proposed model of axon
The evidence for protein synthesis in axons andterminals is reviewed, and our current understandingof the location and disposition of the translational ma-chinery is described. This body of evidence providesthe framework for a novel model of an axon, which isdepicted in Fig. 1. The model can account for thesupply of proteins requisite for supporting steady anddynamic states without invoking the concept of longrange slow transport as means of supply. For reasonsof parsimony and clarity, therefore, slow transport isexcluded from consideration in the context of the
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 3

model. The salient features of the model are that (1)each axonal segment maintains a steady state byundergoing local synthesis and degradation of intrinsicproteins, (2) axons can undergo transformation from asteady state to a dynamic state when a latent sproutingprogram is expressed, and (3) in the steady state, thedi�erentiated ensheathing Schwann cell maintains thesprouting program of axons in a latent state. Exper-imental support for the model will be presented in an
essay below, and explanatory and predictive powers ofthe latter will be contrasted to those o�ered by theprevailing slow transport model.
2. Maintenance of the axon: early views
2.1. Historical synopsis
Concepts of the neuron as a cell originated primarilywith the neurohistologists of the 19th century. The pre-vailing view of nervous system structure at the begin-ning of that century was that it was made up of twoprincipal components, globules, which later wererecognized as perikarya, and ®bers. Remak (1839)reported continuity between globules and ®bers, butthis notion did not gain acceptance until others repli-cated the results later, using improved histologicaltechniques. Waller (1850) showed in frog hypoglossaland glossopharyngeal nerves that the segment distal toa lesion degenerated, but that the proximal stump didnot, and proposed that the nutritional center of sen-sory ®bers was in the spinal ganglia, while that ofmotor ®bers was in the spinal cord. This idea met withresistance because ®bers were believed to be anasto-mosed with one another, and lacked a unique nutri-tional center. Marinesco (1909) described thechromatolytic response of perikarya following inter-ruption of ®bers, which he attributed to the loss of in-¯uence of distal ®bers. Cajal (1928) observed axonalballooning proximal to, and axonal thinning distal tonerve constrictions, and speculated that nutritionalassimilation occurred on a local basis, and that whileperikarya did not supply materials to ®bers, they hada dynamic in¯uence upon them. Parker (1932) sec-tioned the vagal nerve supply to the lateral line organof the cat®sh and found that the length of the ®berremaining attached to the receptor organ correlatedwith the delay of degeneration of neuroepithelial recep-tor cells. This led to the proposal of a trophic impulsein the nerve ®ber, which is consistent today with a roleplayed by rapid anterograde transport.
By the 1930s, it was established that the perikaryonwas necessary for survival of the axon, and that theintegrity of the e�ector depended on the nerve ®ber.The manner by which these interrelationships were sus-tained remained speculative until Paul Weiss initiateda line of research in the 1940s, designed to address thequestion of cellular mechanisms underlying the main-tenance of the axon. Nonetheless, even at the close ofthe 19th century, there already had been speculationby Goldscheider about the very likely `autochthonous'nature of local metabolism in the axon (Barker, 1899).In the 1950s, new concepts and experimental tech-niques expanded our understanding of cell biology,and provided new approaches of investigation.
Fig. 1. (A). Models for maintaining axoplasm. In the slow transport
model, proteins (P) are synthesized in the perikaryon and trans-
ported along the axon in so-called slow transport rate groups by
unknown mechanisms. To remove inherent inconsistencies, degra-
dation of proteins (d) was postulated to occur only in terminals, but
it was shown to occur in the axon (see text). In the local synthesis
model, amino acids (A) are taken up across the axolemma, di�use
within axoplasm, leak out, or are used for protein synthesis (s).
Local degradation (d) of axoplasmic proteins (P) re-supplies the pool
of amino acids. The slow transport model cannot account for the
steady state of the axoplasm as progressive protein degradation with-
out replenishment is inherent to the model; in contrast, the local syn-
thesis model explains maintenance of axoplasm by ordinary
mechanisms of metabolic turnover (see text). (B). Quantiative ex-
pression of the local synthesis model by di�erential equations
(Alvarez and Torres, 1985). The fate of free amino acids in the sol-
uble pool (Ai) and proteins (Pi) of the ith segment are modeled
where the kinetic rate constant (Ks), the di�usion rate constant
(Kdif), the metabolic decay constant (Kd), and leakage (KL), and
uptake (U ) parameters are taken into account. Equation (1)
describes the transfer of amino acids into, and release from proteins,
the di�usion of soluble amino acids into and from segments (i + 1)
and (i ÿ 1), and uptake and leakage across the axolemma. Eq. (2)
describes the transfer of amino acids into, and the release of the lat-
ter from proteins. (C). Sprouting program. The cartoon depicts a
regulatory pathway between Schwann cell and axon. Intact nerve: in
the normal condition, the axon has a distributed `sprouting program'
(tridents inside axon) and the di�erentiated Schwann cell (solid
blocks) represses this program (±q). Anticholinesterases or inhibitors
of exoproteases induce Schwann cells to proliferate and to dedi�er-
entiate (empty blocks), and the repression of the sprouting program
fades away; as a consequence, the axon extends sprouts (protruding
tridents). Regenerating nerve: distal to injury, the axon degenerates
(broken lines), and Schwann cells proliferate and dedi�erentiate; at
the injury, the blind end of the axon expresses the latent sprouting
program when the repression fades away in the distal stump.
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±624

In the vertebrate, current understanding of how axo-plasm is maintained must satisfy a minimum of twotheoretical boundary conditions: (1) nerve cells syn-thesize all of their proteins, and (2) axoplasmic mass inthe intact mature axon is in a steady state. Each ofthese boundary conditions can be satis®ed by eitherone of two mechanisms that have been proposed tosupply axoplasmic proteins; namely, by the prevailingdoctrinal view of slow axoplasmic transport, and bythe historically controversial view of local synthesis.While each mechanism will be considered independentof one another in the context of experimental ®ndingsbelow, in principle, there is no a priori reason why thetwo mechanisms cannot operate in a complementarymanner.
2.2. Slow axoplasmic transport
2.2.1. Origin of slow transport modelWeiss and Hiscoe (1948) reexamined the phenomena
of reversible ballooning of axons above, and thinningbelow nerve constrictions, and proposed that axoplasmwas ¯owing along the axon (Fig. 2). Because riboso-mal particles had been shown to mediate protein syn-thesis, but were not identi®ed in axons (Palay andPalade, 1955), the idea of a proximo-distal ¯ow sup-plying all axoplasmic proteins gained credence when
Droz and Leblond (1963) observed a slow wave ofradioactivity in axons after radiolabeled amino acidswere injected in the vicinity of perikarya. During the1970s, proteins in the slow wave were resolved intotwo transport rate groups, designated IV, or SCb (2±4mm/day), and V, or SCa (0.1±1 mm/day) (Willard etal., 1974; Ho�man and Lasek, 1975; Black and Lasek,1980). Neuro®lament triplet proteins and tubulin wereprincipal constituents of the slower moving com-ponent, while various elements of the actin ®lamentsystem and soluble enzymes were associated with thefaster moving component.
When RNA analysis of axoplasm extruded from thesquid and Myxicola giant ®bers yielded no evidence ofribosomal RNA (rRNA) in invertebrate axoplasm(Lasek et al., 1973) (however, see below), it seemed toprovide biochemical con®rmation that the axon lackeda protein synthesizing machinery. Thus, slow axoplas-mic transport, in which all axoplasmic proteins weresynthesized in cell bodies and supplied to their cognateaxons via the principal slow transport rate groups, wasa necessary theoretical construct to explain the main-tenance of the axoplasm, and became accepted dogma.As a corollary, it was also proposed that constituentsin the two slow transport rate subgroups were con-veyed as two independent moving matrices, mediatedby sliding polymer systems (Lasek, 1986).
Presently, direct evidence of slow transport is lack-ing, and the slow wave of radioactive proteins still pro-vides the best indirect evidence for it, although othertypes of indirect evidence have also been reported. Themajor line of investigation is largely concerned withmechanisms that underlie translocation of slowlytransported constituents. Recent years have seen theemergence of a debate as to whether assembled poly-mers in axoplasm are moving, or whether monomers/oligomers are moving and undergoing exchange withsubunits in assembled stationary polymers (see Nixon,1998). For example, using ¯uorescent probes underdirect visual monitoring, Okabe and Hirokawa (1990,1991, 1992) evaluated the displacement of microtubulesand actin ®laments in neurites, and concluded that theaxonal cytoskeleton was stationary. More recently,Hirokawa et al. (1997) proposed that proteins weretranslocated as monomers, as indicated in part by anapparent translocation of NF-M in transgenic miceinfected with a recombinant vector encoding an epi-tope-tagged protein (Terasa et al., 1996). On the otherhand, Baas and Brown (1997) have proposed that pro-teins were translocated as polymers on the basis ofindirect evidence.
2.2.2. Flaws of the slow transport modelFirst and foremost, the slow transport model must
be consistent with biological principles, and what isknown about real axons. A simpli®ed model of the
Fig. 2. Support and ¯aw of the slow transport model. Cartoon of
the early nerve constriction experiments by Weiss and Hiscoe (1948)
that gave rise to the notion of axoplasmic streaming, and later refor-
mulated as the slow transport model by Lasek and Ho�man (1976).
Chronic compression of a short segment of nerve (rectangular box)
causes ballooning of axoplasm on the proximal side, and thinning on
the distal side of the constricted region. These phenomena were
explained by the hindrance to a constant streaming of proteins com-
ing from the perikaryon (large arrow), which resulted in damming of
a fraction of proteins that distended axons (vertical arrows) proximal
to the constriction, while the unhindered fractional ¯ow (small
arrow) resulted in smaller caliber axons distal to the constriction
(Weiss and Hiscoe, 1948). This model assumes that (i) all axonal
proteins come from the perikaryon by slow transport, and (ii) pro-
teins are eliminated only at the terminals in order to maintain the
uniformity of the axon (Lasek and Ho�man, 1976). The model fails
to explain why ballooning does not continue ad in®nitum. This
inconsistency can be removed by postulating additional mechanisms,
such as degradation of proteins in the ballooned region, or an
unknown feedback mechanism to reduce the amount of protein
entering the axon from the perikaryon; the former explanation is
precluded by a basic premise of the model of metabolic stability of
proteins, while the latter would also result in smaller caliber axons in
the proximal nerve, which does not occur.
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 5

slow transport model is presented in Fig. 1, in order toinclude the possibility of degradation of axoplasmicproteins. Within the framework of the slow transportmodel, two important questions can be posed. (1) Canall axoplasmic proteins survive during a transit timethat may extend over a period of years? (2) Is the peri-karyon capable of supplying all of the proteins neededto maintain an axoplasmic mass that may be as muchas one-to-two thousand times greater than itself?
Real axons span distances from tens of micrometersin the case of local circuit neurons, to several meters,in the case of principal neurons of large vertebrates. Inall cases, the metabolic lifespan of axoplasmic proteinsbecomes the issue of paramount importance withwhich the adequacy of the slow transport model mustbe judged. If proteins move at rates of 1±4 mm/day,and have half-lives of 1±2 weeks, as reported for cyto-skeletal proteins in nerves and brain (Karlsson andSjoÈ strand, 1971a; Forgue and Dahl, 1978), axonscould not extend beyond a few centimeters. This para-dox was circumvented by Lasek and Ho�man (1976),and earlier by Weiss and Hiscoe (1948), by postulatingthat axonal proteins were stable during their transit,and were degraded ®nally in terminals. Nevertheless,another unexplained paradox arises in the context ofaxonal ballooning that results from constriction ofnerves. According to the slow transport model, bal-looning results from the progressive accumulation ofproteins due to hindrance to proximo-distal transport(Fig. 2). The model predicts that the ballooning ofaxons behind the constriction should continue to growad in®nitum. Inasmuch as they do not do so, anotherexplanation, such as protein degradation, or an arrestof slow transport would need to account for the stabil-ization of the ballooned axoplasm; however, the for-mer inference would contravene the assumption thataxoplasmic proteins are stable, and the latter would bean unspeci®able ad hoc mechanism.
The question of metabolic stability of proteins wasaddressed by Nixon (1980) who clearly showed thataxoplasmic proteins are not unique, because their esti-mated average half-life was not di�erent from that ofother cellular proteins. Moreover, amino acids fromradiolabeled neuro®laments were released into theextracellular space (Nixon and Logvinenko, 1986).Finally, an important ®nding was that amino acidsreleased during degradation were reutilized locally(Nixon, 1980). The latter had implications not only forthe issue of metabolic stability of axoplasmic proteins,but also for bringing into question the reliability ofspecifying the perikaryon as the exclusive source ofradiolabeled proteins in the axon (see below). In thiscontext, it should be noted that a previous estimate ofDroz et al. (1973) was that >90% of radiolabeling ofthe slow wave in preganglionic parasympathetic ®bersdisappeared en route before reaching terminals.
Miller and Samuels (1997) formulated a modelwhich acknowledges that axonal proteins are degraded,and that axons are maintained in a steady state. Themodel assumes that proteins enter the axon as a bolus,and that the transport velocity of proteins is highestproximal to the cell body, and decreases exponentiallyas proteins are degraded in a manner that maintainsgeometric uniformity of the axoplasm. Each bolus ofproteins leaving the perikaryon forms a moving cylin-der of axoplasm that decreases in length with time anddistance, and eventually vanishes when axon terminalsare reached. Thus, the model predicts that perikaryareceiving a pulse of radiolabeled amino acid shouldgenerate a square wave of radioactivity of constantamplitude that shortens progressively with distancealong the axons. This prediction, however, is not rea-lized by actual, experimentally produced waves ofradioactivity (cf. Fig. 21, Gar®sh).
In this model, the maximal axonal length is gov-erned by the amount of protein that the perikaryoncan deliver to the axon per unit of time. Axons 15meters in length in blue whales could be accounted forif the half-life of proteins were 2 years, and the initialtransport velocity were 7 mm/day or any combinationof numbers that would satisfy the equation: length =(2/ln(2)) � (initial velocity) � (t1/2) (K.E. Miller, per-sonal communication). Even if ad hoc values that arewell beyond reported ones were to be accepted withoutexperimental support, this slow transport model stillloses credibility as a sole mechanism for maintainingaxoplasm, because the perikaryon is required to deliverenough protein every day to replace that of the initial7 mm segment of the axon. The protein mass con-tained within this segment would be one-to-two-foldthat of the perikaryon. Such a metabolic load for acell would be unprecedented.
In the slow transport model, the perikaryon is thesole source of axoplasmic proteins whereby it main-tains the whole axoplasm in a steady state, no matterhow large the mass. As a consequence, the export ofrequisite proteins to the axon must impose a substan-tial metabolic load on perikarya. Karlsson (1982) esti-mated that in some neurons, over 99% of proteinsynthesized in the cell should be destined for the axon.However, most of the radioactivity remains in the peri-karya rather than being exported to axons, as shownin retina (Karlsson and SjoÈ strand, 1971a), and inspinal ganglia of the rat (Mori et al., 1979; Stromskaand Ochs, 1981). This issue was addressed speci®callyin spinal ganglia of toad and frog (von Bernhardi andAlvarez, 1989). Relative protein synthesizing activityof perikarya, which ranged from 1 to less than 40, wasnot commensurate with the relative axoplasmic massof cognate axons, which ranged from 1 to 5000. Con-trary to predictions arising from the tenets of the slowtransport model, these ®ndings indicate that there is
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±626

no direct, simple relationship between the maintenanceof axoplasm and the metabolic load that it imposes onthe perikaryon, which is contrary to what would bepredicted by slow transport theory. For additionalinconsistencies, the reader is referred to Alvarez andTorres (1985), and Alvarez (1992). Fast anterogradetransport cannot be invoked to supplement axoplasmicproteins because the estimated transport capacity is anorder of magnitude smaller than that of the slowwave, and it carries membrane and vesicular productsof the constitutive secretory pathways destined for thecell's periphery and terminals, and carries no cyto-skeletal proteins (McEwen et al., 1971; Droz et al.,1973).
In summary, available data indicate that the tenetsof the slow transport model alone cannot satisfy therequirements for steady state maintenance of axo-plasm. It is also clear that ad hoc assumptions, whichmay be added from time to time to address inadequa-cies of the model, or predictions that lack experimentalveri®cation raises fundamental questions about its val-idity. The view presented here is that the foundingtenets of the model are seriously ¯awed, and the chal-lenge is now to formulate a more biologically realisticparadigm.
3. Local protein synthesis in axons
3.1. Metabolic studies
In this section we will review experimental ®ndingsof a protein synthesizing machinery, and/or evidencefor de novo protein synthesis in mature axons, imma-ture growing axons, and presynaptic terminals. Thedata, moreover, demonstrate a compelling need tomodify and broaden current views of the axon inorder to recognize the much greater biological com-plexity than previously perceived.
3.1.1. The Mauthner (M-) cell axon as a modelThe M-cell axon has long served as a model for
studying RNA content and composition (see below),as well as endogenous protein synthesis in the axoncompartment. In teleost ®shes, the pair of M-neuronsmediate a so-called `C-bend' of the trunk, which is the®rst behavioral manifestation of an escape response.The M-perikarya are located in the rostral medulla,and after decussating, the large myelinated axon pro-jects the full length of the spinal cord. In its rostralextent in gold®sh, the M-cell axon may vary 40±80 mmin diameter (Funch et al., 1981), after which it tapersin the spinal cord. In its course, it gives o� very shortcollaterals that generally extend only through thethickness of the myelin sheath (Celio et al., 1979),where they form axo-axonic synapses with motoneur-
ons and interneurons. Under favorable conditions, M-cell axoplasm can be isolated as a wholemount withsome of its collaterals and presynaptic boutons stillattached (Koenig, unpublished; see below).
Studies of the M-cell axon by EdstroÈ m (1966) andEdstroÈ m and SjoÈ strand (1969) were among the earliestto provide evidence of an endogenous capacity to in-corporate amino acids into axoplasmic proteins in amanner that was sensitive to inhibitors of cytoriboso-mal, but not mitoribosomal dependent protein syn-thesis. The ®ndings in the M-axon were subsequentlycon®rmed and extended. Alvarez and Chen (1972a)showed incorporation of radiolabeled leucine into M-cell axoplasm by electrophoretic microinjection invivo. The low level of radioactivity in the surroundingmyelin sheath ruled out a glial origin of radiolabeledproteins in axoplasm. Alvarez and Benech (1983) latercon®rmed that the incorporation was sensitive to in-hibitors of protein synthesis. Although the estimatedrate of incorporation per unit of M-cell axoplasm was1.7±4.2% of that of the perikaryon, total incorpor-ation into axoplasm was 17 to 51-fold greater thanthat of the perikaryon when volumes of cell body andaxoplasm were taken into account; i.e., incorporationinto the axon per unit time was signi®cantly greaterthan that into the perikaryon. An analysis of radio-active axoplasmic proteins after CXM sensitive incor-poration in vitro by Koenig (1991) revealed a numberof radiolabeled polypeptides, including major cyto-skeletal polypeptides such as a- and b-tubulins, actin,and putative neuro®lament subunits.
Electrical stimulation in vivo was shown to increasethe rate of incorporation of labeled amino acids intoM-cell axoplasm (EugenõÂ n and Alvarez, 1995). After 4hours of continuous stimulation, the incorporationwas similar to that of control, but exceeded that ofcontrol by a factor of two after 18 hours of stimu-lation. This elevation, however, was transient, andreturned to the baseline within a day. This suggestedthat amino acid incorporation was likely to have beenregulated.
3.1.2. Studies in mammalian nervesAlthough the M-cell axon was a favorable model for
reasons of size and accessibility, direct analysis of pro-tein synthesis was also performed on axoplasm isolatedon a microscopic scale from myelinated ®bers ofselected mammalian species. Among early investi-gations was a study in the cat, showing leucine incor-poration into axoplasmic proteins of spinal accessorynerve root ®bers in vitro (Koenig, 1967). Later, a simi-lar approach on lesioned hypoglossal nerve in rabbitshowed that the rate of CXM sensitive incorporationinto axoplasmic proteins became very signi®cantly el-evated between 12 and 15 hours after nerve section,reaching a greater than 10-fold increase in speci®c
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 7

radioactivity compared to control by 18 hours (Tobiasand Koenig, 1975a). It is of interest that a comparabledramatic increase in protein and RNA synthesis wasalso documented in isolated squid nerves after about10 hours of incubation in vitro (Perrone Capano et al.,1999). Inasmuch as (i) the onset of the augmented rateof synthesis in the rabbit nerves was delayed, (ii) theresponse was independent of cell bodies, and (iii) themagnitude of the response diminished proximally withdistance from the injury site, it suggested that theinjury may have perturbed an intercellular Schwanncell regulatory pathway that normally down regulateslocal protein synthesis in intact axons (Tobias andKoenig, 1975a, 1975b; also, see below).
In vivo studies, using conventional historadiographictechniques, were able to con®rm that after radioactiveamino acid incorporation, grains overlay axons inlesioned (Benech et al., 1982), and unlesioned sciaticnerves (Contreras et al., 1983). Analysis of axoplasmicpolypeptides after incorporation of radiolabeled aminoacids in dorsal and ventral root ®bers in vitro (Koenig,1991) showed a number of labeled polypeptides thatincluded actin and both tubulin subunits; however,neuro®lament proteins were not among those thatcould be identi®ed on the basis of relative electrophor-etic mobilities. Nonetheless, the low molecular massneuro®lament protein (NF-L) was immunoadsorbedand identi®ed by 2D electrophoresis from radiolabedrat sciatic nerve (Sotelo et al., 1992).
Incorporation of amino acids was also evaluated inganglion cell axons regenerating in vitro from gold®shretinal explants. Growing axons radiate outward fromthe retinal explant in the absence of nonneural cells,and can be severed from the parent explant for pur-poses of microanalyzing incorporation into decentra-lized axons quantitatively. The incorporation wassensitive to inhibitors of cytoribosomal protein syn-thesis, and was not a�ected signi®cantly by inhibitionof mitoribosomal protein synthesis (Koenig andAdams, 1982). While prominently labeled polypeptidesincluded actin and b-tubulin (Koenig, 1989), a-tubulinwas generally not labeled, despite the apparent equival-ence in mass between the tubulin subunits in electro-phoretic patterns. This is in contrast to ®ndings inmature spinal root axons of the rat, in which bothtubulin subunits were similarly labeled (Koenig, 1991).The disparity in labeling of the two microtubule subu-nits in immature axons indicated that the source of a-tubulin is exogenous to the axon. This is consistentwith recent work that showed that a-tubulin mRNA isabsent in growing chick axons in vitro (Olink-Couxand Hollenbeck, 1996; see below). Because b-tubulincomprises a major fraction of labeled products ingrowing axons (Koenig, 1991), it would explain whynocodazole, a microtubule depolymerizing agent, pro-duced a signi®cant inhibition of protein synthesis
(Koenig and Adams, 1982), since an increase in tubu-lin monomer concentration diminishes tubulin mRNAstability (Cleveland et al., 1981).
Very recently, it was con®rmed that b-tubulin andb-actin are synthesized in intact, growing sympatheticganglion cell axons of the rat, using immunabsorptionto recover speci®c labeled products (Eng et al., 1999).Interestingly, while only distal axons were incubatedwith 35S-methionine in a compartmentalized chamber,some proteins labeled in axons were also labeled inperikarya, suggesting a retrograde transport of pro-teins after synthesis in axons (see below). Althoughthis in vitro study indicated that inhibitors of proteinsynthesis did not a�ect the rate of elongation over aperiod of 30 hours, an inhibition of growth in regener-ating axons of the rat was observed in vivo (Edbladhet al., 1994; Gaete et al., 1998). Indeed, the latter stu-dies emphasized the importance of a local source ofproteins in promoting axonal regeneration.
Despite di�erences in the conclusion drawn aboutthe importance of local protein synthesis for axongrowth based on di�erent preparations and experimen-tal paradigms, there are important qualifying distinc-tions between the two studies that should be noted.For example, in the culture system (Eng et al., 1999),immature neurons were taken from newborn pups,and axonal ®elds were in very close proximity to cellsof origin. In such preparations, there is a rapid bi-directional redistribution of axoplasmic aggregates thatoccurs in the form of motile varicosities (Koenig et al.,1985; Hollenbeck and Bray, 1987). Such `bulk' axo-plasmic tra�c in the immature state requires a `per-missive' cytoskeletal organization in order toaccommodate the movement of large axoplasmicaggregates. This type of activity in immature neuronsmay minimize the extent to which axonal growthduring development depends on local synthesis. Theexperiments in vivo, in contrast, were conducted onthe sciatic nerve in the adult rat after lesioning at con-siderable distance from cells of origin (Edbladh et al.,1994; Gaete et al., 1998). While newly formed sproutswould contain a cytoskeletal organization that is per-missive for rapid aggregate transport, proximal stumpaxons, intervening between sprouts and the perikar-yon, have a cytoskeleton that is `nonpermissive' forsuch transport. Thus, while rapid bulk transport maysatisfy requirements for axon growth in the immatureembryonic state, the mechanism is lacking in regener-ating axons of the adult, and a dependence on localsynthesis becomes manifest. Further discussion of therole of local protein synthesis in regeneration is pro-vided below (see Section 6.3.4).
3.1.3. Protein synthesis in invertebrate axonsFor more than a half century, the squid giant axon
has been exploited as a model for studying axonal
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±628
physiology; however, the ability to recover a fairly sub-
stantial amount of axoplasm for purposes of biochemi-cal, and molecular biological assays also made it a
very favorable preparation for studying the question of
local protein synthesis in an axonal compartment. Inthe squid, the giant axon used in these experiments
corresponds to the most medial (and largest) of ap-
proximately ten large axons which follow the course of
a corresponding number of stellate nerves, which pro-jects radially from the stellate ganglion. Each giant
axon originates by fusion of several axons that are de-
rived from perikarya segregated in a well de®ne region
of the stellate ganglion (i.e., the giant ®ber lobe).
When early work by Fisher and Litvak (1967), and
Giuditta et al. (1968), revealed that axoplasmic pro-
teins were radiolabed when the isolated giant ®ber wasincubated in vitro with radioactive amino acids, it set
the stage for a number of studies that yielded strong
evidence for local protein synthesis. Subsequent inves-tigations, however, gave rise to divergent views that
engendered controversy about the local source of axo-
plasmic proteins.
Initial biochemical analyses indicated that axoplasm
contained only 4S RNA, and apparently lacked rRNA
(Lasek et al., 1973). When the distribution of newly
synthesized radioactive proteins was evaluated by lightmicroscopic autoradiography after incubation, radioac-
tivity appeared to be largely restricted to the periaxo-
nal glial sheath (Lasek et al., 1974), and led to thehypothesis that proteins were synthesized in periaxonal
glial cells, and secondarily transferred to the subjacent
axon by intercellular transfer (Lasek et al., 1974). As
noted (Giuditta, 1980), however, the level of resolutionwas not su�cient to exclude the cortical zone of axo-
plasm near the plasma membrane as a potential contri-
buting site of incorporation. Subsequent work, whichincluded experiments in which labeled proteins were
recovered from internally perfused ®bers after extru-
sion of axoplasm, yielded additional evidence in sup-
port of the hypothesis of a glial-axonal transfer ofproteins (Gainer et al., 1977; Lasek et al., 1977; Tytell
and Lasek, 1984; Tytell et al., 1986), and indicated
that the process may be dependent on extracellular
Ca2+ (Tytell and Lasek, 1984). While the technique ofextruding axoplasm leaves a residual cortical rim of
axoplasm subjacent to the axonal membrane (Tsukita
et al., 1986), the inclusion of ribonuclease in some per-fusion experiments (Gainer et al., 1977) appeared to
indicate that the residual cortical layer was not a sig-
ni®cant source of newly synthesized proteins, although
satisfactory permeation of the cortical layer by ribonu-clease was not actually demonstrated. These studies on
glial-axonal protein transfer greatly contributed to the
widely held premise that proteins were not synthesizedendogenously in the giant axon, a view that was gener-
alized to all axons as well, and became commonlyaccepted doctrine in neurobiology.
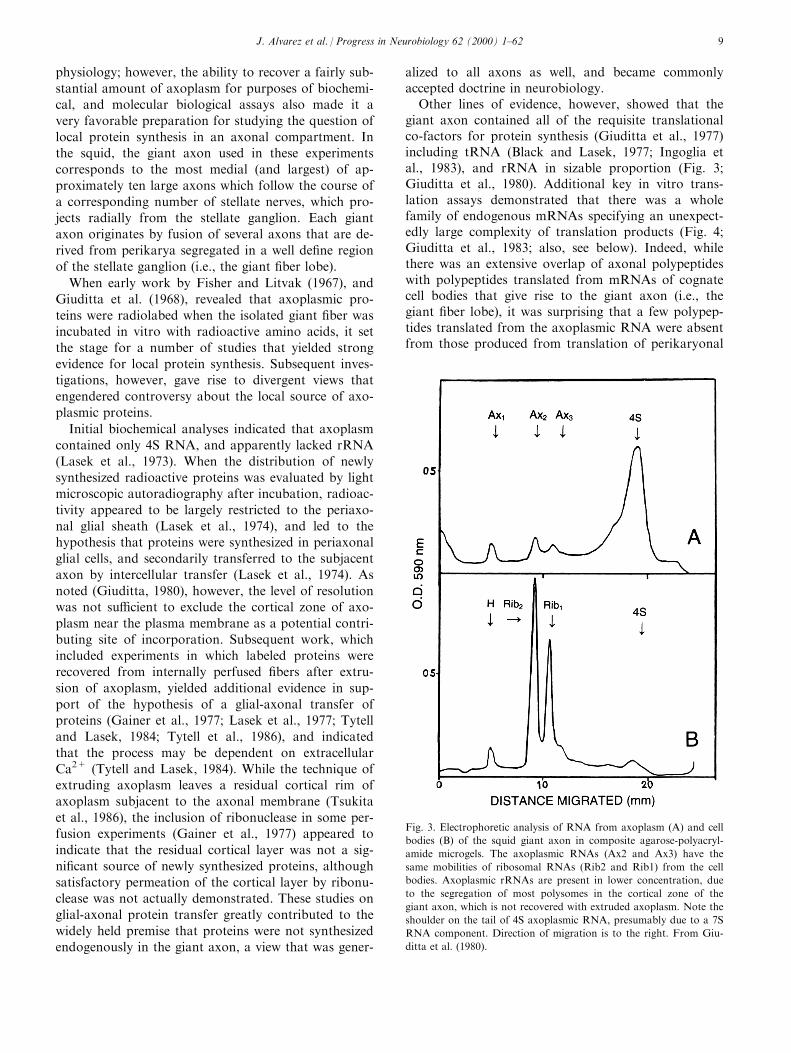
Other lines of evidence, however, showed that thegiant axon contained all of the requisite translationalco-factors for protein synthesis (Giuditta et al., 1977)including tRNA (Black and Lasek, 1977; Ingoglia etal., 1983), and rRNA in sizable proportion (Fig. 3;Giuditta et al., 1980). Additional key in vitro trans-lation assays demonstrated that there was a wholefamily of endogenous mRNAs specifying an unexpect-edly large complexity of translation products (Fig. 4;Giuditta et al., 1983; also, see below). Indeed, whilethere was an extensive overlap of axonal polypeptideswith polypeptides translated from mRNAs of cognatecell bodies that give rise to the giant axon (i.e., thegiant ®ber lobe), it was surprising that a few polypep-tides translated from the axoplasmic RNA were absentfrom those produced from translation of perikaryonal
Fig. 3. Electrophoretic analysis of RNA from axoplasm (A) and cell
bodies (B) of the squid giant axon in composite agarose-polyacryl-
amide microgels. The axoplasmic RNAs (Ax2 and Ax3) have the
same mobilities of ribosomal RNAs (Rib2 and Rib1) from the cell
bodies. Axoplasmic rRNAs are present in lower concentration, due
to the segregation of most polysomes in the cortical zone of the
giant axon, which is not recovered with extruded axoplasm. Note the
shoulder on the tail of 4S axoplasmic RNA, presumably due to a 7S
RNA component. Direction of migration is to the right. From Giu-
ditta et al. (1980).
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 9

mRNA. The latter ®ndings are interesting because theyindicated that there are mechanisms for selective vec-torial tra�cking of speci®c mRNAs to speci®c cellulardomains where translation of such mRNAs can be e�-ciently regulated according to local requirements.There is presently a growing body of evidence thatmRNAs contain so-called `zip codes' in the 3 ' untrans-lated region that confer speci®c subcellular localization(Oleynikov and Singer, 1998).
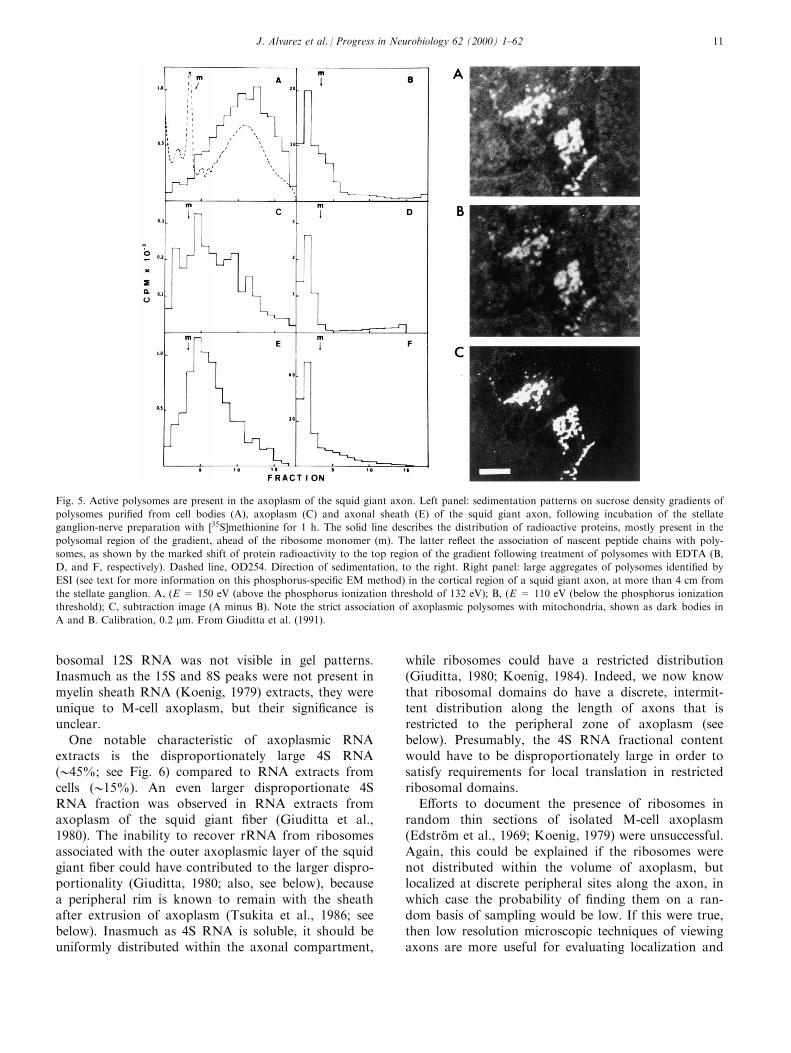
Subsequently, polyribosomes were isolated fromaxoplasm extruded from the giant ®ber, and shown tobe translationally active when assayed for labeling ofnascent polypeptide chains (Fig. 5; Giuditta et al.,
1991). In the same set of experiments, a riboprobe tomouse NF68 mRNA was shown to cross-hybridizedwith mRNA in the giant axon, as well as in stellateganglion cells, and ribosomal phosphorus signals wereshown in axoplasm in the vicinity of the axonal mem-brane using electron spectroscopic imaging (ESI)(Fig. 5).
The ESI method requires an EM equipped with anenergy spectrometer and ®lter systems. This type ofEM permits selection of a speci®c region of the energyloss spectrum to view an image produced by electronsinelastically scattered from elements of interest inultrathin sections (10±20 nm), free of heavy metal ®x-ation or staining, while ®ltering out unscattered andelastically scattered electrons. Nucleic acids are veryrich in phosphorus, and each ribosome, which contains>6800 phosphorus (P) atoms, produces a bright signalof 025 nm in a low contrast ®eld when the spectralenergy loss selected is above the P edge (i.e., E > 132eV; typically, E � 150 or 155 eV), and the signal disap-pears when the spectral energy loss selected is belowthe P edge (e.g., E � 110 eV). Comparison above andbelow the P edge, and size of the signal provide criteriafor identi®cation that are highly reliable (Korn et al.,1983; Ottensmeyer, 1986; Martin et al., 1989). Morerecently, the presence of axoplasmic polysomes in thesquid giant axon has been con®rmed by immunocyto-chemical analyses (Sotelo et al., 1999).
All the above data have provided the most directand compelling evidence for an endogenously activeprotein synthesizing machinery in the squid giantaxon, and were instrumental in abrogating a basic pre-mise of the glial-axon protein transfer hypothesis (seeabove).
3.2. Restricted ribosomal domains in vertebrate axons
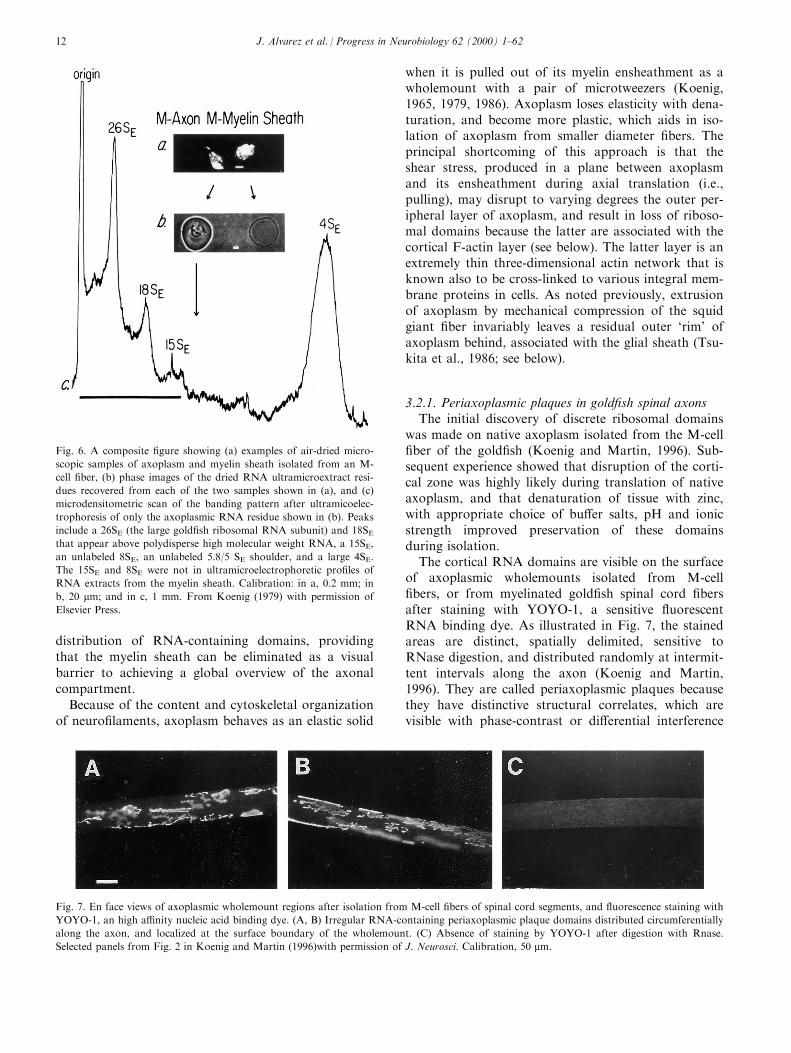
As previously noted, the M-cell axon in gold®sh wasthe ®rst vertebrate axon model in which analytical andexperimental metabolic studies were undertaken toanalyze RNA content, base composition, intercellularRNA transfer, and protein synthesizing activity(EdstroÈ m et al., 1962, 1969, 1964a, 1964b, 1966, 1967;EdstroÈ m and SjoÈ strand, 1969). Initially, RNA wasmicroextracted from formaldehyde-®xed dissected axo-plasmic samples (EdstroÈ m et al., 1962; EdstroÈ m,1964a). It was later shown that rRNA was indeed pre-sent in undegraded RNA extracts of un®xed isolatedaxoplasmic samples (Koenig, 1979). Typically, thepeaks that were resolved in the RNA pattern (seeFig. 6) included 26S and 18S cytoribosomal peaks, a15S peak, a small 8S peak (not labeled), an unresolvedshoulder (not labeled), presumably including 5.8S/5SRNA, and a very large 4S peak. While the 15S RNApeak in the RNA gel pattern could have representedthe larger 16S mitoribosomal RNA, the smaller mitori-
Fig. 4. Electrophoretic pattern of the translation products speci®ed
by puri®ed RNA from axoplasm (A) and cell bodies (L) of the squid
giant axon. Approximately ®fty translation products could be
detected in the axoplasmic lane with longer exposure times. Most
axoplasmic translation products have mobilities that correspond to
translation products from cell bodies, but some are unique, or domi-
nate in the axoplasmic pattern (e.g., the protein band somewhat lar-
ger than 116 kDa). Other translation products dominate in the cell
bodies lane. The mobility and size of MW markers (in kDa) is
shown on the left of lane A. From Giuditta et al. (1986).
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6210
bosomal 12S RNA was not visible in gel patterns.Inasmuch as the 15S and 8S peaks were not present inmyelin sheath RNA (Koenig, 1979) extracts, they wereunique to M-cell axoplasm, but their signi®cance isunclear.
One notable characteristic of axoplasmic RNAextracts is the disproportionately large 4S RNA(045%; see Fig. 6) compared to RNA extracts fromcells (015%). An even larger disproportionate 4SRNA fraction was observed in RNA extracts fromaxoplasm of the squid giant ®ber (Giuditta et al.,1980). The inability to recover rRNA from ribosomesassociated with the outer axoplasmic layer of the squidgiant ®ber could have contributed to the larger dispro-portionality (Giuditta, 1980; also, see below), becausea peripheral rim is known to remain with the sheathafter extrusion of axoplasm (Tsukita et al., 1986; seebelow). Inasmuch as 4S RNA is soluble, it should beuniformly distributed within the axonal compartment,
while ribosomes could have a restricted distribution(Giuditta, 1980; Koenig, 1984). Indeed, we now knowthat ribosomal domains do have a discrete, intermit-tent distribution along the length of axons that isrestricted to the peripheral zone of axoplasm (seebelow). Presumably, the 4S RNA fractional contentwould have to be disproportionately large in order tosatisfy requirements for local translation in restrictedribosomal domains.
E�orts to document the presence of ribosomes inrandom thin sections of isolated M-cell axoplasm(EdstroÈ m et al., 1969; Koenig, 1979) were unsuccessful.Again, this could be explained if the ribosomes werenot distributed within the volume of axoplasm, butlocalized at discrete peripheral sites along the axon, inwhich case the probability of ®nding them on a ran-dom basis of sampling would be low. If this were true,then low resolution microscopic techniques of viewingaxons are more useful for evaluating localization and
Fig. 5. Active polysomes are present in the axoplasm of the squid giant axon. Left panel: sedimentation patterns on sucrose density gradients of
polysomes puri®ed from cell bodies (A), axoplasm (C) and axonal sheath (E) of the squid giant axon, following incubation of the stellate
ganglion-nerve preparation with �35S]methionine for 1 h. The solid line describes the distribution of radioactive proteins, mostly present in the
polysomal region of the gradient, ahead of the ribosome monomer (m). The latter re¯ect the association of nascent peptide chains with poly-
somes, as shown by the marked shift of protein radioactivity to the top region of the gradient following treatment of polysomes with EDTA (B,
D, and F, respectively). Dashed line, OD254. Direction of sedimentation, to the right. Right panel: large aggregates of polysomes identi®ed by
ESI (see text for more information on this phosphorus-speci®c EM method) in the cortical region of a squid giant axon, at more than 4 cm from
the stellate ganglion. A, (E = 150 eV (above the phosphorus ionization threshold of 132 eV); B, (E = 110 eV (below the phosphorus ionization
threshold); C, subtraction image (A minus B). Note the strict association of axoplasmic polysomes with mitochondria, shown as dark bodies in
A and B. Calibration, 0.2 mm. From Giuditta et al. (1991).
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 11
distribution of RNA-containing domains, providingthat the myelin sheath can be eliminated as a visualbarrier to achieving a global overview of the axonalcompartment.
Because of the content and cytoskeletal organizationof neuro®laments, axoplasm behaves as an elastic solid
when it is pulled out of its myelin ensheathment as awholemount with a pair of microtweezers (Koenig,1965, 1979, 1986). Axoplasm loses elasticity with dena-turation, and become more plastic, which aids in iso-lation of axoplasm from smaller diameter ®bers. Theprincipal shortcoming of this approach is that theshear stress, produced in a plane between axoplasmand its ensheathment during axial translation (i.e.,pulling), may disrupt to varying degrees the outer per-ipheral layer of axoplasm, and result in loss of riboso-mal domains because the latter are associated with thecortical F-actin layer (see below). The latter layer is anextremely thin three-dimensional actin network that isknown also to be cross-linked to various integral mem-brane proteins in cells. As noted previously, extrusionof axoplasm by mechanical compression of the squidgiant ®ber invariably leaves a residual outer `rim' ofaxoplasm behind, associated with the glial sheath (Tsu-kita et al., 1986; see below).
3.2.1. Periaxoplasmic plaques in gold®sh spinal axonsThe initial discovery of discrete ribosomal domains
was made on native axoplasm isolated from the M-cell®ber of the gold®sh (Koenig and Martin, 1996). Sub-sequent experience showed that disruption of the corti-cal zone was highly likely during translation of nativeaxoplasm, and that denaturation of tissue with zinc,with appropriate choice of bu�er salts, pH and ionicstrength improved preservation of these domainsduring isolation.
The cortical RNA domains are visible on the surfaceof axoplasmic wholemounts isolated from M-cell®bers, or from myelinated gold®sh spinal cord ®bersafter staining with YOYO-1, a sensitive ¯uorescentRNA binding dye. As illustrated in Fig. 7, the stainedareas are distinct, spatially delimited, sensitive toRNase digestion, and distributed randomly at intermit-tent intervals along the axon (Koenig and Martin,1996). They are called periaxoplasmic plaques becausethey have distinctive structural correlates, which arevisible with phase-contrast or di�erential interference
Fig. 6. A composite ®gure showing (a) examples of air-dried micro-
scopic samples of axoplasm and myelin sheath isolated from an M-
cell ®ber, (b) phase images of the dried RNA ultramicroextract resi-
dues recovered from each of the two samples shown in (a), and (c)
microdensitometric scan of the banding pattern after ultramicoelec-
trophoresis of only the axoplasmic RNA residue shown in (b). Peaks
include a 26SE (the large gold®sh ribosomal RNA subunit) and 18SEthat appear above polydisperse high molecular weight RNA, a 15SE,
an unlabeled 8SE, an unlabeled 5.8/5 SE shoulder, and a large 4SE.
The 15SE and 8SE were not in ultramicroelectrophoretic pro®les of
RNA extracts from the myelin sheath. Calibration: in a, 0.2 mm; in
b, 20 mm; and in c, 1 mm. From Koenig (1979) with permission of
Elsevier Press.
Fig. 7. En face views of axoplasmic wholemount regions after isolation from M-cell ®bers of spinal cord segments, and ¯uorescence staining with
YOYO-1, an high a�nity nucleic acid binding dye. (A, B) Irregular RNA-containing periaxoplasmic plaque domains distributed circumferentially
along the axon, and localized at the surface boundary of the wholemount. (C) Absence of staining by YOYO-1 after digestion with Rnase.
Selected panels from Fig. 2 in Koenig and Martin (1996)with permission of J. Neurosci. Calibration, 50 mm.
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6212

contrast optics, and protrude above the surface bound-ary of the axoplasmic wholemount (Fig. 8). The struc-tural correlates overlie the ¯uorescent RNA domains,and are only preserved under optimum conditions ofisolation. Frequently, ¯uorescent `puncta' are visiblewithin the plaque formation, when partially disrupted,and in surrounding and underlying axoplasm (Fig. 9).As indicated by EM, the ¯uorescent puncta are mostlikely correlates of polyribosomal clusters (see below).When the axoplasmic wholemount is doubly stainedwith YOYO-1, and rhodamine-conjugated phalloidinto label RNA and F-actin, respectively, it is clear fromconfocal microscopic analysis that the plaque for-mation is intimately associated with the cortical F-actin layer, and that ¯uorescent puncta in subcorticalaxoplasm, representing putative polyribosomes (seebelow), are associated with F-actin (Fig. 10). The im-plications of this relationship are that the actin cytos-keleton probably plays an important role in governingthe puncta distribution, which is consistent with thegeneral association of so-called `free' ribosomes (anoperational de®nition that is an artifact of cell frac-tionation techniques) in cells with the cytoskeleton(Ornelles et al., 1986; Hesketh and Pryme, 1991; seebelow).
Electron density and size provide morphological cri-teria for identifying ribosomes at an EM level. How-ever, heavy metal staining, which is used to enhancecontrast in conventional transmission electron mi-croscopy (CTEM), can introduce structural ambigu-ities. ESI, on the other hand, provides a physicalmethod based on energy loss spectroscopy of rRNA Psignals (see above), in addition to size to identify ribo-somes, and circumvents potential problems arisingfrom heavy metal staining. Indeed, heavy metal stain-ing precludes application of ESI, and, for this reason,osmium ®xation, which is necessary to stabilize myelinlipids, can not be employed for examining intactnerves. Therefore, epon-embedded axoplasmic whole-mount specimens were used to study periaxoplasic pla-que domains.
A typical periaxoplasmic plaque domain consisted ofan unusual structural matrix with which speci®c rRNAP signals were associated, especially at the inneraspects of the matrix, in addition to polyribosomesscattered in subjacent axoplasm (Fig. 11). The matrixwas novel, and appeared to represent an ultrastruc-tural counterpart of plaque structural formations vis-ible with phase-contrast and di�erential interferencecontrast optics (Koenig and Martin, 1996; see Fig. 8).
Fig. 8. En face views of axoplasmic wholemount regions (A, B, C) containing single periaxoplasmic plaque formations stained with YOYO-1,
and exhibiting stuctrual correlates, as visualized with (A1) DIC optics, or (B1, C2) with phase-contrast optics. The structural correlates protrude
from the surface and lie subjacent to the axolemma (see also Fig. 8). From Fig. 4 in Koenig and Martin (1996)with permission of J. Neurosci.
Calibration, 10 mm.
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 13
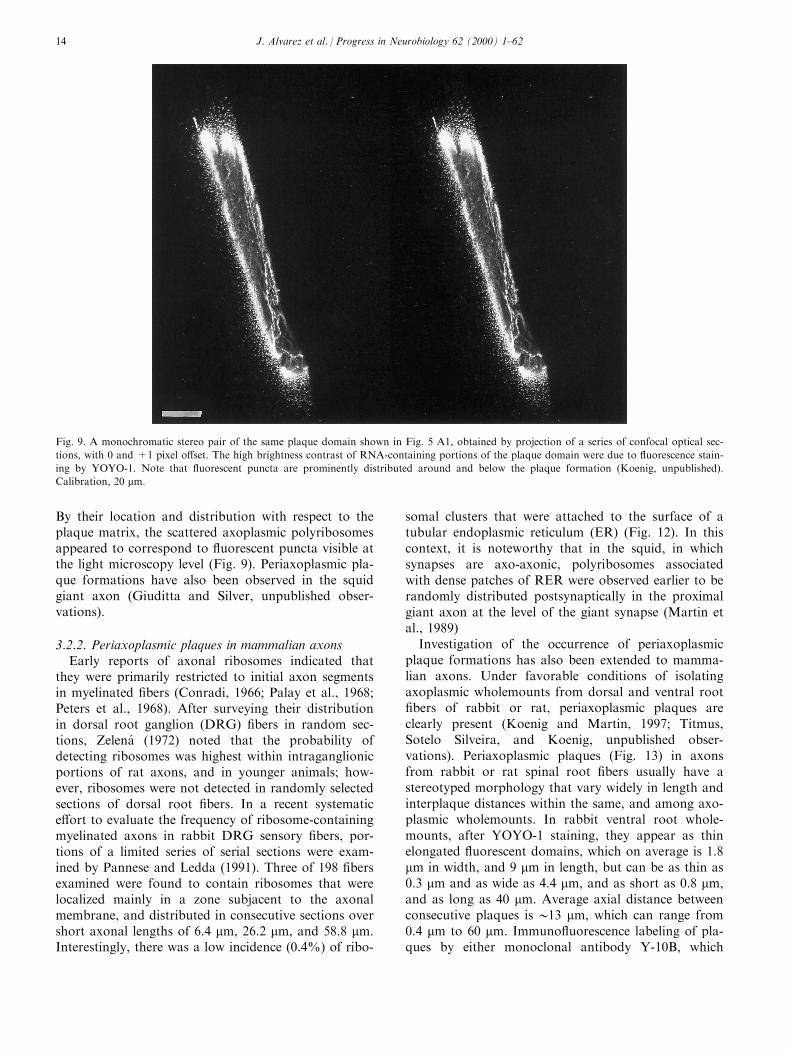
By their location and distribution with respect to theplaque matrix, the scattered axoplasmic polyribosomesappeared to correspond to ¯uorescent puncta visible atthe light microscopy level (Fig. 9). Periaxoplasmic pla-que formations have also been observed in the squidgiant axon (Giuditta and Silver, unpublished obser-vations).
3.2.2. Periaxoplasmic plaques in mammalian axonsEarly reports of axonal ribosomes indicated that
they were primarily restricted to initial axon segmentsin myelinated ®bers (Conradi, 1966; Palay et al., 1968;Peters et al., 1968). After surveying their distributionin dorsal root ganglion (DRG) ®bers in random sec-tions, Zelena (1972) noted that the probability ofdetecting ribosomes was highest within intraganglionicportions of rat axons, and in younger animals; how-ever, ribosomes were not detected in randomly selectedsections of dorsal root ®bers. In a recent systematice�ort to evaluate the frequency of ribosome-containingmyelinated axons in rabbit DRG sensory ®bers, por-tions of a limited series of serial sections were exam-ined by Pannese and Ledda (1991). Three of 198 ®bersexamined were found to contain ribosomes that werelocalized mainly in a zone subjacent to the axonalmembrane, and distributed in consecutive sections overshort axonal lengths of 6.4 mm, 26.2 mm, and 58.8 mm.Interestingly, there was a low incidence (0.4%) of ribo-
somal clusters that were attached to the surface of atubular endoplasmic reticulum (ER) (Fig. 12). In thiscontext, it is noteworthy that in the squid, in whichsynapses are axo-axonic, polyribosomes associatedwith dense patches of RER were observed earlier to berandomly distributed postsynaptically in the proximalgiant axon at the level of the giant synapse (Martin etal., 1989)
Investigation of the occurrence of periaxoplasmicplaque formations has also been extended to mamma-lian axons. Under favorable conditions of isolatingaxoplasmic wholemounts from dorsal and ventral root®bers of rabbit or rat, periaxoplasmic plaques areclearly present (Koenig and Martin, 1997; Titmus,Sotelo Silveira, and Koenig, unpublished obser-vations). Periaxoplasmic plaques (Fig. 13) in axonsfrom rabbit or rat spinal root ®bers usually have astereotyped morphology that vary widely in length andinterplaque distances within the same, and among axo-plasmic wholemounts. In rabbit ventral root whole-mounts, after YOYO-1 staining, they appear as thinelongated ¯uorescent domains, which on average is 1.8mm in width, and 9 mm in length, but can be as thin as0.3 mm and as wide as 4.4 mm, and as short as 0.8 mm,and as long as 40 mm. Average axial distance betweenconsecutive plaques is 013 mm, which can range from0.4 mm to 60 mm. Immuno¯uorescence labeling of pla-ques by either monoclonal antibody Y-10B, which
Fig. 9. A monochromatic stereo pair of the same plaque domain shown in Fig. 5 A1, obtained by projection of a series of confocal optical sec-
tions, with 0 and +1 pixel o�set. The high brightness contrast of RNA-containing portions of the plaque domain were due to ¯uorescence stain-
ing by YOYO-1. Note that ¯uorescent puncta are prominently distributed around and below the plaque formation (Koenig, unpublished).
Calibration, 20 mm.
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6214
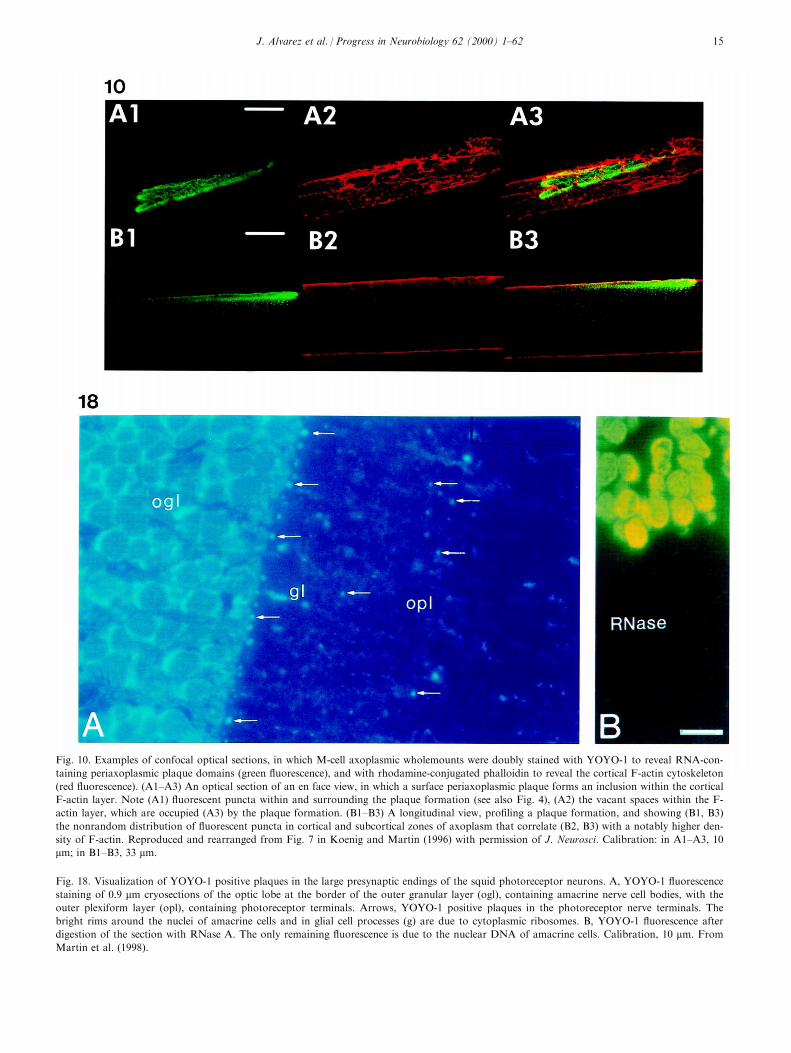
Fig. 10. Examples of confocal optical sections, in which M-cell axoplasmic wholemounts were doubly stained with YOYO-1 to reveal RNA-con-
taining periaxoplasmic plaque domains (green ¯uorescence), and with rhodamine-conjugated phalloidin to reveal the cortical F-actin cytoskeleton
(red ¯uorescence). (A1±A3) An optical section of an en face view, in which a surface periaxoplasmic plaque forms an inclusion within the cortical
F-actin layer. Note (A1) ¯uorescent puncta within and surrounding the plaque formation (see also Fig. 4), (A2) the vacant spaces within the F-
actin layer, which are occupied (A3) by the plaque formation. (B1±B3) A longitudinal view, pro®ling a plaque formation, and showing (B1, B3)
the nonrandom distribution of ¯uorescent puncta in cortical and subcortical zones of axoplasm that correlate (B2, B3) with a notably higher den-
sity of F-actin. Reproduced and rearranged from Fig. 7 in Koenig and Martin (1996) with permission of J. Neurosci. Calibration: in A1±A3, 10
mm; in B1±B3, 33 mm.
Fig. 18. Visualization of YOYO-1 positive plaques in the large presynaptic endings of the squid photoreceptor neurons. A, YOYO-1 ¯uorescence
staining of 0.9 mm cryosections of the optic lobe at the border of the outer granular layer (ogl), containing amacrine nerve cell bodies, with the
outer plexiform layer (opl), containing photoreceptor terminals. Arrows, YOYO-1 positive plaques in the photoreceptor nerve terminals. The
bright rims around the nuclei of amacrine cells and in glial cell processes (g) are due to cytoplasmic ribosomes. B, YOYO-1 ¯uorescence after
digestion of the section with RNase A. The only remaining ¯uorescence is due to the nuclear DNA of amacrine cells. Calibration, 10 mm. From
Martin et al. (1998).
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 15
forms an immune complex with rRNA (Lerner et al.,1981), or by human anti-ribosomal P protein, corre-
sponds to YOYO-1 ¯uorescence staining, and con®rmsthat ribosomes are distributed in plaque domains at alight microscopy level. While RNA ¯uorescence stain-
ing with YOYO-1 is di�use, immunostaining with Y-
10B, or anti-ribosomal P protein is punctate in charac-ter, suggesting a distribution of discrete polysomal
clusters (Fig. 13C) (Titmus, Sotelo Silveira, and Koe-nig, unpublished observations). As in the case of gold-®sh axons (Fig. 8), there are also structural correlates
associated with ¯uorescent plaque domains (Fig. 13B).
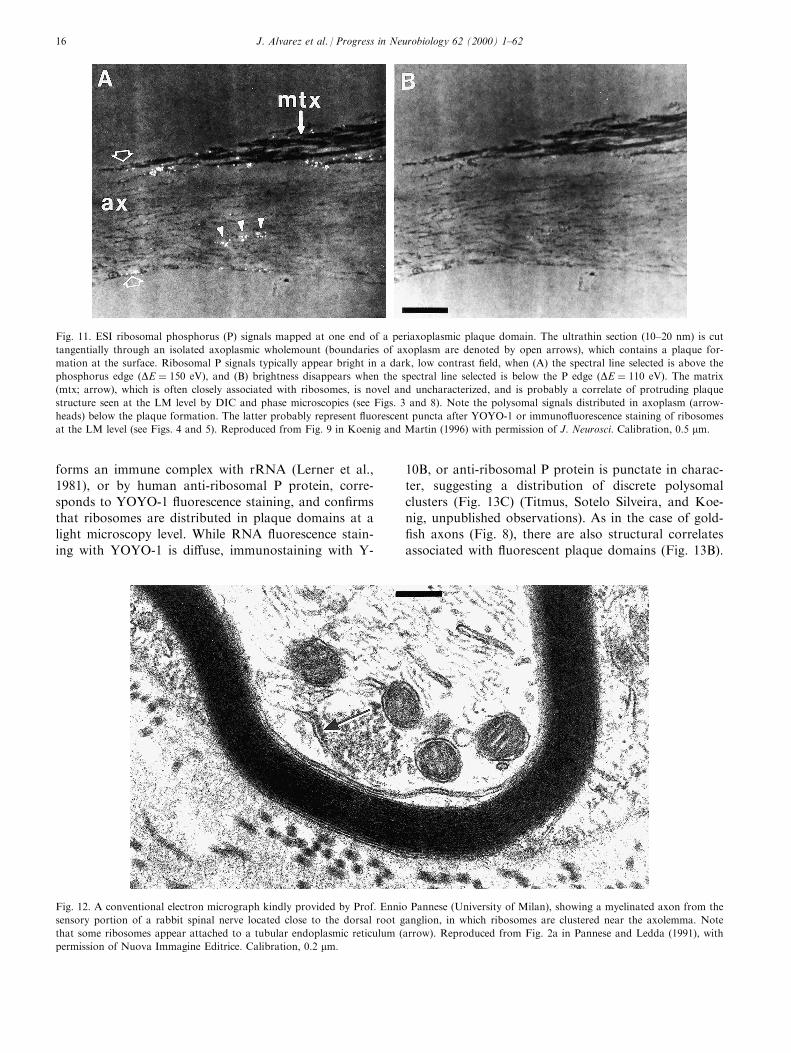
Fig. 11. ESI ribosomal phosphorus (P) signals mapped at one end of a periaxoplasmic plaque domain. The ultrathin section (10±20 nm) is cut
tangentially through an isolated axoplasmic wholemount (boundaries of axoplasm are denoted by open arrows), which contains a plaque for-
mation at the surface. Ribosomal P signals typically appear bright in a dark, low contrast ®eld, when (A) the spectral line selected is above the
phosphorus edge �DE � 150 eV), and (B) brightness disappears when the spectral line selected is below the P edge �DE � 110 eV). The matrix
(mtx; arrow), which is often closely associated with ribosomes, is novel and uncharacterized, and is probably a correlate of protruding plaque
structure seen at the LM level by DIC and phase microscopies (see Figs. 3 and 8). Note the polysomal signals distributed in axoplasm (arrow-
heads) below the plaque formation. The latter probably represent ¯uorescent puncta after YOYO-1 or immuno¯uorescence staining of ribosomes
at the LM level (see Figs. 4 and 5). Reproduced from Fig. 9 in Koenig and Martin (1996) with permission of J. Neurosci. Calibration, 0.5 mm.
Fig. 12. A conventional electron micrograph kindly provided by Prof. Ennio Pannese (University of Milan), showing a myelinated axon from the
sensory portion of a rabbit spinal nerve located close to the dorsal root ganglion, in which ribosomes are clustered near the axolemma. Note
that some ribosomes appear attached to a tubular endoplasmic reticulum (arrow). Reproduced from Fig. 2a in Pannese and Ledda (1991), with
permission of Nuova Immagine Editrice. Calibration, 0.2 mm.
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6216

They appear under optimal conditions of isolation asexcrescences, protruding above the cortical surfaceboundary of the axoplasmic wholemount.
There are two additional ®ndings of interest fromearlier studies that should be noted in this context,given what is presently know of the existence of corti-cal ribosomal plaque domains. Early quantitative EMautoradiographic studies showed that radiolabeledRNA was localized mainly at the periphery of opticnerve axons after intraocular injection of �3H]uridine inrabbits (Gambetti et al., 1973). Similarly, a later quan-titative EM autoradiographic analysis of radiolabeledproteins in the SCb component of the slow transportrate groups showed 2.5 times more radioactivity in thesubaxolemmal region than in the rest of the axoncross-section (Heriot et al., 1985). Thus, periaxoplas-mic plaques in the cortical zone provides a ready ex-planation for the peripheral localization of
radiolabeled RNA, and, as centers of translational ac-tivity, plaque domains very likely serve as a potentialsource of radioactively labeled proteins found in SCb(see Section 3.1).
Although plaques are typically found along inter-nodes, there are infrequent occasions when a plaquemay span a node of Ranvier and its two contiguousparanodes (Fig. 13D), or it may extend only partiallyacross a single paranode. The low incidence of plaquesin nodal/paranodal regions, however, would appear toindicate that their localization in this region is not ofspecial signi®cance. Periaxoplasmic plaque formationshave now been identi®ed by ESI in axoplasmic whole-mounts isolated from rabbit ventral root ®bers, and indelipidated ventral root ®bers (Martin and Koenig;unpublished ®ndings).
Preservation of the cortical zone during isolation isinconsistent. Nonetheless, it provides an unparalleled
Fig. 13. A gallery of videomicroscopic en face images, showing periaxoplasmic plaques in selected regions of axoplasmic wholemounts isolated
from myelinated motor ®bers of rabbit ventral root. (A) A typical intermittent distribution of periaxoplasmic plaques at the surface of an isolated
wholemount, as revealed by ¯uorescence staining with YOYO-1. (B) Under optimal conditions of isolating wholemounts, structural correlates, as
revealed by phase-contrast microscopy (see also Fig. 3), protrude above ¯uorescent plaque domains shown in (A). Some structural correlates
appear refractile (arrowheads). (C1) Immunostaining of ribosomes in a plaque formation with monoclonal Y-10B antibody. In contrast to di�use
¯uorescent staining of all RNA in plaque domains by YOYO-1 (see A), immuno¯uorescence staining by Y-10B, or anti-ribosomal P protein (not
shown; see text) yields discrete ribosomal clusters. (C2) The phase structural correlate of the plaque domain shown in (C1). (C3) Superimposition
of the ¯uorescence and phase images reveals that the longitudinal distribution of ribosome clusters in the plaque domain conforms to the contour
of the structural correlate. (D1) An example of a periaxoplasmic plaque that spans the region of a node of Ranvier and its two paranodes (Koe-
nig and Titmus, unpublished). Calibration, 10 mm.
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 17
appreciation of the systematic spatial distribution ofribosomal domains along axons. Inasmuch as periaxo-plasmic plaques have been documented in gold®sh(Koenig and Martin, 1996), and in mammalian axons(Koenig and Martin, 1997; Titmus, Sotelo Silveira,and Koenig, unpublished observations), it seems likelythat they are cortical specializations ubiquitous tomost, if not all mature axons of projection neurons.
3.2.3. Implications of cortical actin cytoskeleton inplaque domains
The thin cortical F-actin layer, where periaxoplasmicplaque formations are located, is ordinarily viewed asa dynamic structure in most cells (Stossel, 1993). Pla-ques may not be static in their spatial distribution andorganization. Indeed, their size and morphology arehighly variable even in the same axon. It is possiblethat plaques undergo dynamic changes, which mayinclude slow translocation or redistribution, an infer-ence that would be di�cult to demonstrate with pre-sent techniques. The co-distribution of F-actin with¯uorescent puncta in axoplasm (Fig. 10), which arepresumably polysomal clusters (Fig. 11), implicates theF-actin cytoskeleton in governing ribosome, and/ormRNA distributions, in agreement with a functionalrole in binding/anchorage and transport in other cells(Zambetti et al., 1990; Hesketh and Pryme, 1991;Singer, 1992; Bassell et al., 1994b). It is noteworthy inthis context that the highly abundant elongation factor1a (eEF-1a) (Slobin, 1980) is a F-actin binding proteinassociated with the actin cytoskeleton (see Condeelis,1995). Moreover, its activity is subject to regulation bythe actin cytoskeleton, and small shifts in intracellularpH (Liu et al., 1996). Finally, inasmuch as the actincytoskeleton has been shown to play a role in axoplas-mic transport (Kuznetsov et al., 1992), it may play animportant role as well in the local disposition of com-
ponents that o�-load from the microtubule transportsystem within the plaque domain, as well as in translo-cating translation products within the volume of axo-plasm locally.
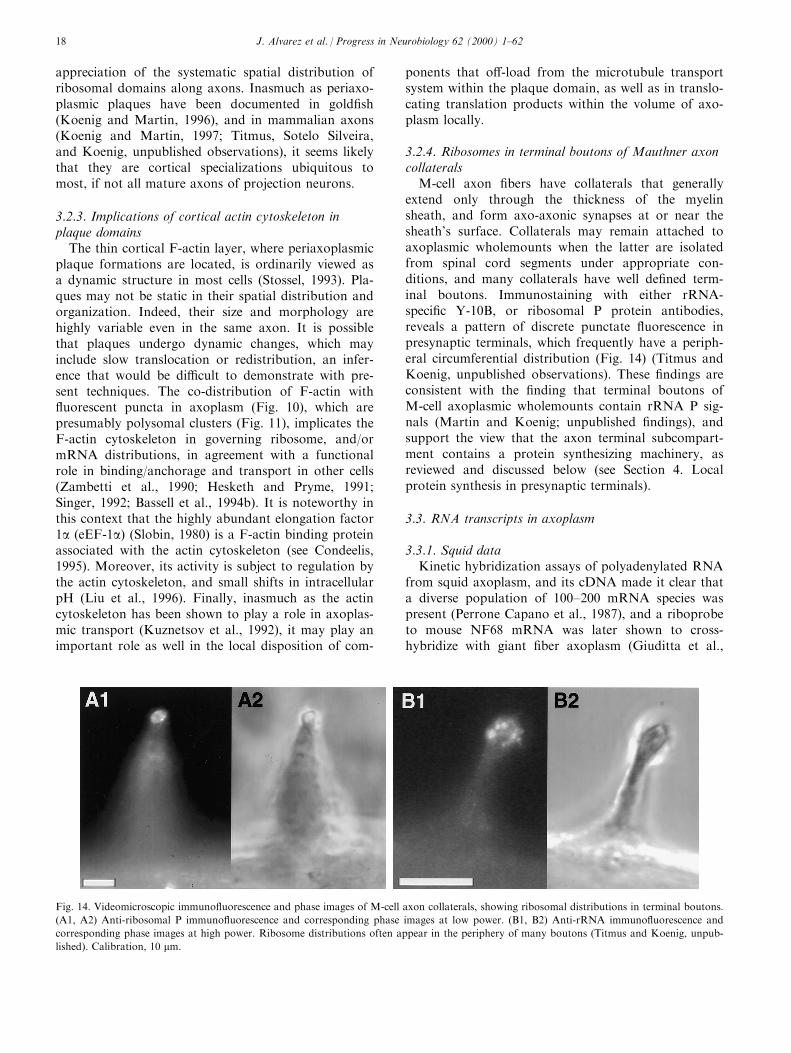
3.2.4. Ribosomes in terminal boutons of Mauthner axoncollaterals
M-cell axon ®bers have collaterals that generallyextend only through the thickness of the myelinsheath, and form axo-axonic synapses at or near thesheath's surface. Collaterals may remain attached toaxoplasmic wholemounts when the latter are isolatedfrom spinal cord segments under appropriate con-ditions, and many collaterals have well de®ned term-inal boutons. Immunostaining with either rRNA-speci®c Y-10B, or ribosomal P protein antibodies,reveals a pattern of discrete punctate ¯uorescence inpresynaptic terminals, which frequently have a periph-eral circumferential distribution (Fig. 14) (Titmus andKoenig, unpublished observations). These ®ndings areconsistent with the ®nding that terminal boutons ofM-cell axoplasmic wholemounts contain rRNA P sig-nals (Martin and Koenig; unpublished ®ndings), andsupport the view that the axon terminal subcompart-ment contains a protein synthesizing machinery, asreviewed and discussed below (see Section 4. Localprotein synthesis in presynaptic terminals).
3.3. RNA transcripts in axoplasm
3.3.1. Squid dataKinetic hybridization assays of polyadenylated RNA
from squid axoplasm, and its cDNA made it clear thata diverse population of 100±200 mRNA species waspresent (Perrone Capano et al., 1987), and a riboprobeto mouse NF68 mRNA was later shown to cross-hybridize with giant ®ber axoplasm (Giuditta et al.,
Fig. 14. Videomicroscopic immuno¯uorescence and phase images of M-cell axon collaterals, showing ribosomal distributions in terminal boutons.
(A1, A2) Anti-ribosomal P immuno¯uorescence and corresponding phase images at low power. (B1, B2) Anti-rRNA immuno¯uorescence and
corresponding phase images at high power. Ribosome distributions often appear in the periphery of many boutons (Titmus and Koenig, unpub-
lished). Calibration, 10 mm.
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6218

1991). Such ®ndings supported data from earlier exper-iments, in which at least 50 labeled polypeptides wereresolved by SDS polyacrylamide gel electrophoresisafter in vitro translation of axoplasmic RNA (Giudittaet al., 1983, 1986). In addition, in situ hybridization(ISH) with radiolabeled poly(U) probe con®rmed thepresence of poly(A)+ in the giant axon (PerroneCapano et al., 1987). Subsequently, when a cDNAlibrary was established from axoplasmic poly (A)+
RNA, several cDNA clones were identi®ed andsequenced, and corresponding mRNAs were shown tobe localized in axoplasm by ISH, including b-actin andb-tubulin (Kaplan et al., 1992), the heavy chain ofkinesin (Gioio et al., 1994), squid enolase (Chun et al.,1995), and a polypeptide having homology to a cal-cium channel protein of the sarcoplasmic reticulum(Chun et al., 1997). In all cases, riboprobes were pre-pared from axoplasmic clones. Some of these mRNAswere shown to be translated in the axon by PCR ana-lyses of puri®ed axoplasmic polyribosomes, such as b-actin and b-tubulin (Kaplan et al., 1992), and theheavy chain of kinesin (Gioio et al., 1994), or byimmunoadsorption analysis, such as NF protein (Cris-pino et al., unpublished work). These studies are themost comprehensive analyses of endogenous RNAtranscripts in axoplasm, and provide notable examplesof mRNAs coding for cytoskeletal and soluble proteinsthat are ordinarily reported to be constituents of theslow transport rate groups (Black and Lasek, 1980;Brady and Lasek, 1981). Of the two directionally dis-tinct microtubule based motor proteins, however, onlycytoplasmic dynein has been identi®ed with a slowtransport group (Dillman et al., 1996), while kinesinhas been shown to be associated with rapid antero-grade transported organelles (Hirokawa et al., 1991).
3.3.2. Mature vertebrate axonsEvidence of RNA transcripts in mature vertebrate
axons is limited for the most part to a few modelaxons in which ensheathment has not contributed toambiguity of interpretation, either from the standpointof subcellular localization, or neuronal speci®city ofthe transcript. Thus, in axons located in the posteriorneurohypophysis, several neuronal secretory proteinmRNAs have been reported that code for oxytocin(Jirikowski et al., 1990), provasopressin, prooxytocin,as well as prodynorphin (Mohr and Richter, 1992a,1992b), and vasopressin (Trembleau et al., 1994, 1996).A similar example of hormonal mRNA in axons wasdocumented in the case of the mollusc Lymnaea stag-nalis, in which hormone-secreting neurons in the CNSexpress caudodorsal cell hormone (CDCH), whichcomprises several neuropeptides that mediate egg lay-ing behavior. High resolution ISH showed that CDCHmRNA was localized in secretion granules distributedwithin axons and terminals (Dirks et al., 1993).
Finally, mRNAs coding for an olfactory marker pro-tein, for calcitonin gene-related peptide, and a numberof odorant receptors, which are distributed amongsubsets of neurons, were also found along olfactoryaxons (Vassar et al., 1994; Resseler et al., 1994; Denis-Donini et al., 1998).
The signi®cance of transcripts in olfactory axons isunknown. Similarly, there is no evidence that hormo-nal transcripts, which are presumably sequestered insecretory granules are translated within the axonalcompartment (van Minnen, 1994). Nevertheless, it isworth noting in this context that while ribosomes havenot been described in normal neurohypophyseal axons,they have been documented in hypertrophied vasopres-sin-containing axons after 7 days of osmotic challengeof drinking 2% NaCl (Martin, 1997). While the issuesof cause and e�ect cannot be addressed based on avail-able data, the appearance of ribosomes in the lattercondition must be considered as a potential contribut-ing factor in causing axonal hypertrophy, involvingincreased axoplasmic mass. This inference is consistentwith the increased rate of amino acid incorporation inaxoplasm promoted by hyperactivity (EugenõÂ n andAlvarez, 1995; see Section 6.1.9).
Galanin, a peptide that is synthesized also in magno-cellular neurons of the hypothalamus, is upregulated inresponse to salt loading. Unlike other neurohypophy-seal peptide mRNAs described above that are presum-ably sequestered within a vesicular subcompartmentwithin the axon (van Minnen, 1994), preprogalaninmRNA is present in neurohypophyseal axoplasm undersuch conditions, and appears to be transported by anonsecretory pathway (Landry and HoÈ kfelt, 1998).
Information about mRNAs coding for axoplasmicproteins in mature axons of vertebrates is very scanty.The neurohypophysis, which contains axons from mag-nocellular neurons, has been shown by RT-PCR ampli-®cation to contain NF-L mRNA (Mohr and Richter,1992a, 1992b). Similarly, NF-M mRNA was shown byRT-PCR ampli®cation in axoplasm isolated from thegold®sh M-cell ®ber (Weiner et al., 1996). ISH of actinmRNA in the M-cell axoplasmic wholemount revealeda nonrandom cortical distribution (Crispino, Giudittaand Koenig, unpublished data). Recent observations,in which ribosomal P proteins were immunolabeled incombination with ISH of b-actin mRNA, ISH signalsappeared to have a restricted distribution within theplaque domain (Sotelo Silveria and Koenig, unpub-lished observations).
Previous studies of NF transcripts in nerves havereported that NF-M and NF-L mRNAs are expressedin di�erentiating Schwann cells (Kelly et al., 1992),and during demyelination in response to ingestion oftellurium, or to transection of the sciatic nerve inweanling rats (Roberson et al., 1992). Unilateral nervetransection also induced a low level of expression of
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 19

NF-L mRNA in contralateral intact nerve (Robersonet al., 1992). These observations indicated that ex-pression of NF mRNAs is not necessarily restricted toneurons, but can be also present in a nonneural cell,such as the myelinating Schwann cells, albeit in anapparent transient manner. Recent extension of theseobservations in dedi�erentiating and redi�erentiatingSchwann cells of the distal nerve stump after sciaticnerve lesion in vivo, indicated that NF-M and NF-LmRNAs were transiently expressed during axonal de-generation, and, again, re-expressed at the time of my-elin deposition with the ingrowth of axons (Fabrizi etal., 1997). However, immunolabeling of NF-M or NF-L in Schwann cells was inconsistent and scatteredamong the degenerating ®bers in the distal nervestump, which indicated that translation was variableand not constant.
The elevated expression of NF-L mRNA in theproximal nerve stump in response to transection of thesciatic nerve in young rats (Roberson et al., 1992) wascon®rmed in adult rat very recently by northern blotanalysis; moreover, RT-PCR of RNA extracted fromproximal nerve stump, or from intact nerve yieldedevidence that NF-L, NF-M, and NF-H mRNAs wereall present (Sotelo Silveira; unpublished observations).ISH of NF-L mRNA indicated that signals were loca-lized in Schwann cells, myelin sheath, and in a regionbetween the inner border of the myelin sheath and theaxon (Sotelo Silveira, unpublished observations). Thelatter observation is of special interest in view of theapparent CXM-sensitive radiolabeling of putative neu-
ro®lament polypeptides in the M-cell axon (Koenig,1991). Future work should shed light on the questionsof origin, localization and signi®cance of NF RNAtranscripts in intact nerve. It remains to be demon-strated, however, that NF RNA transcripts are loca-lized in mammalian axons, and can be translated.
3.3.3. Small cytoplasmic RNAs in axonsPeriaxoplasmic plaque domains are presumably
restricted local centers of translational activity whichcan account for reports of protein synthesis in ver-tebrate axons (see above). It is reasonable to assumethat ribosomes and mRNAs (see above), as well asother potential RNA constituents involved in transla-tional activity or regulation of the latter are targetedto plaque domains. Recent experiments indicate thatBC1, a rat brain speci®c small cytoplasmic RNA,which is synthesized by RNA polymerase III, is loca-lized in dendrites of a subset of neurons (Tiedge et al.,1991), and in neurohypophyseal axons (Tiedge et al.,1993). BC1 is associated with a 10S ribonucleoproteinparticle (RNP) (Kobayashi et al., 1991, 1992), and isconsidered to be involved in protein synthesis in somemanner (Tiedge et al., 1991).
When full length radiolabeled BC1 was microin-jected into the M-cell perikaryon, it was transportedrapidly into the two dendrites and the axon (Muslimovet al., 1997). In some experiments, localization ofradiolabeled BC1 in M-cell axon appeared as focal ac-cumulations in the cortical zone, reminiscent of pla-que-like domains (Fig. 15). The targeting to the axon
Fig. 15. An autoradiogram (contrast is reversed), showing a focal distribution of silver grains overlying a region of a M-cell axoplasmic whole-
mount 2 hours after microinjection of 35S-labeled BC1 RNA in the M-cell body of the gold®sh. The spatially restricted clustering of grains is sug-
gestive of a speci®c accumulation of labeled BC1 RNA in a periaxoplasmic plaque domain, carried by rapid axonal transport (Muslimov,
Titmus, Koenig and Tiedge, unpublished). Calibration, 50 mm.
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6220

compartment was speci®c, and information appearedto be encoded in a 5 ' sequence because various trun-cated and irrelevant radiolabeled RNA constructs,including one with the 3 ' sequence were not trans-ported after microinjection (Muslimov et al., 1997). Itnow appears that an analogous small cytoplasmicRNA may also be expressed in gold®sh brain (Musli-mov and Tiedge, personal communication).
Recent studies of the electromotor system of Tor-pedo have shown that vault RNPs are highly enrichedin cholinergic terminals (Herrmann et al., 1996), andare actively transported bidirectionally, as indicated byprogressive accumulation of immunoreaction producton proximal and distal sides after the electric nervewas crushed or ligated (Li et al., 1999). Similar to BC1RNA, vault RNA is small, and is transcribed by poly-merase III. Although vaults have a close associationwith synaptic vesicles in the electric nerve terminals,their function is unknown (Herrmann et al., 1996).
Another small cytoplasmic RNA localized in thecortical zone of the M-cell axon, and in rabbit and ratventral root axons, is 7S RNA (Titmus et al., 1998), astructural RNA of the signal recognition particle(SRP). SRPs target ribosomes that are translatingmRNAs coding for secretory and integral membraneproteins to the ER. They function by recognizingemerging signal sequences of nascent polypeptides,arresting translation until the SRP-mRNA-ribosomecomplex binds to the SRP receptor of the ER, where-upon translation continues through the translocon(Walter and Johnson, 1994). One of the six componentpolypeptides associated with 7S RNA is SRP 54, towhich signal recognition activity maps (Walter andJohnson, 1994). Immunolabelling of SRP 54 localizesto plaque domains in axoplasmic wholemounts fromrabbit ventral root ®bers (Titmus et al., 1998). It is ofinterest in this context that one of the electron micro-graphs in the study by Pannese and Ledda (1991)documented ribosomes in contact with a tubular ER(Fig. 12). However, in cells, polypeptides synthesizedon the rough endoplasmic reticulum (RER) areshuttled to a Golgi apparatus for post-translationalmodi®cation and processing, including glycosylation.An equivalent system is not known to be in the axon.Nevertheless, if SRPs are present in the axonal com-partment, as preliminary ®ndings suggest, then theremay also be RNA transcripts coding for secretoryand/or membrane proteins. Although there is no evi-dence to support this inference, an investigation of thepossibility now seems warranted.
3.3.4. Neurons in cultureISH for poly(A)+ mRNA in cultured cortical neur-
ons from embryonic rat brain has indicated that thehybridization signal could only be detected in proximalaxons after several days in culture, apparently associ-
ated with microtubules (Bassell et al., 1994a). In chicksympathetic axons, Olink-Coux and Hollenbeck (1996)observed, however, that poly(A)+ mRNA was presentalong the length of the axon, although the signal inten-sity was greatest in the proximal segment. Further-more, the distribution was punctate, suggestinggranule-like clustering of mRNA, that was most fre-quently located at branch points, varicosities, and insome growth cones. Two additional RNA transcriptsthat have been identi®ed in growing axons are amRNA for the Tm-5 tropomyosin isoform (Hannan etal., 1995), and a 2 kb mRNA coding for SCG10, aprotein believed to be a member of an actin bindingfamily that may modulate membrane-cytoskeletal in-teractions (Hannan et al., 1996). Finally, RT-PCRrevealed that while b-actin mRNA was present, a-tubulin mRNA was not (Olink-Coux and Hollenbeck,1996). As noted previously, the latter observation wasconsistent with the apparent lack of a-tubulin labelingin growing gold®sh retinal ganglion cell axons (Koe-nig, 1991).
In a very recent in-depth study of cultured corticalrat brain neurons by Bassell et al. (1998), polyribo-somes were identi®ed in growth cones at an ultrastruc-tural level, while high resolution ISH showed that b-actin mRNA was localized in granule-like puncta dis-tributed within the shaft of the axon as well as ingrowth cones. Of interest also was that b-actinmRNA-containing `granules' exhibited a nonrandomproximity to microtubules, which indicated that micro-tubules probably play a major role in mediating distri-bution. The apparent absence of g-actin mRNA(Bassell et al., 1998), and a-tubulin mRNA (Olink-Coux and Hollenbeck, 1996) in growing axons sup-ports the concept of selective tra�cking of RNA tran-scripts to speci®c neuronal compartments in general,and to the axonal compartment that is developmen-tally regulated in particular.
3.4.5. Growing versus mature axonsGrowing axons of cultured neurons provide direct
accessibility to the axonal compartment without hin-drance of ensheathment, which provides a convenientmeans of evaluating the presence of endogenous RNAtranscripts. Uncritical extrapolation of ®ndings fromsuch studies to mature axons, however, is probablynot justi®ed. For example, both tubulin subunits showCXM-sensitive labeling in mature spinal root axons(Koenig, 1991). In contrast, however, although b-tubu-lin exhibits CXM-sensitive labeling in growing retinalganglion cell axons, a-tubulin is not labeled at all(Koenig, 1989; see above). The latter observation isalso consistent with ®ndings that a-tubulin mRNA islacking in growing chick sympathetic axons (Olink-Coux and Hollenbeck, 1996).
This particular example suggests that there may be
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 21

signi®cant qualitative and quantitative distinctions inthe mRNA population in growing axons compared tonongrowing, ensheathed axons. Another major distinc-tion is the systematic distribution of spatially restrictedribosomal domains comprising cortical periaxoplasmicplaque formations of ensheathed axons. Such special-ized formations have not been reported in growingaxons (Olink-Coux and Hollenbeck, 1996; Bassell etal., 1998). These di�erences may be related to distinc-tions in structural as well as functional propertiesexhibited by undi�erentiated and di�erentiated axons.For example, immature axons have a poorly organizedand dynamic cytoskeleton that is permissive for therapid redistribution of axoplasmic aggregate masses(Koenig et al., 1985; Hollenbeck and Bray, 1987),which contributes to net accretion of mass duringelongation, or loss of mass with rapid retraction.These are mechanisms that greatly enhance structuralplasticity associated with axon growth (see below). Incontrast, ensheathed, di�erentiated axons have ahighly organized, cross-linked stable cytoskeleton, inwhich an already established mass must be maintainedin a steady state, while lacking a discernible plasticityuntil it is transformed by extrinsic factors (see below).
3.4.6. Glial source of axonal RNAEarly evidence supported the view of a local extra-
axonal source of RNA along the axon. The ability toanalyze RNA content and base composition on amicroscopic scale prompted an early study of RNAchanges in the M-cell axon in response to spinal cordtransection (EdstroÈ m, 1964b). RNA was extracted byRNase digestion of dissected axon and myelin sheathsamples taken from the proximal spinal cord stump.Changes in RNA composition were evaluated in-directly in each sample type by determining the ade-nine/guanine (A/G) ratio after acid hydrolysis of RNAdigests and ultramicroelectrophoresis. The A/G ratioin the axon, which was 0.56 before transection, rapidlyincreased to 0.70 twelve hours after transection, andcontinued to increase to reach a peak of 0.80 after 2±3days. Subsequently, the A/G ratio diminished, butremained elevated for several days, and returned to apretransection value after 20±30 days. The myelinsheath also showed similar augmented changes in theA/G ratio in response to transection that were in-phase with those in the axon. RNA content, however,determined at 2 and 30 days after transection, was notchanged signi®cantly. A transient increase in A/G ratiowithout a signi®cant change in total RNA is consistentwith a relative increase in synthesis of an adenine-richRNA (i.e., mRNA). Moreover, the initial rapid rate ofchange, and the close parallel enrichment in axon andmyelin sheath appeared to suggest that metabolicchanges were locally induced and common to both
structures, which implicated a common cellular source,
such as myelin sheath oligodendrocytes.
A later study further tested this proposition by incu-
bating spinal cord segments with �3H]pyrimidines invitro for 7 hours, dissecting out the M-cell ®ber (i.e.,
axon with associated myelin sheath) from un®xed tis-
sue, and analyzing the distribution of radioactivity in asucrose gradient. Actinomycin D-sensitive, high mol-
ecular mass radioactive peaks, sedimenting in the 16
and 28±30 S regions, were present in the gradient. Inseparate experiments, it was determined that 50% or
more of the radiolabeled RNA was in the axon. Whenun®xed myelinated M-cell ®ber segments (i.e., axon
with myelin sheath, without glial cells) were dissected
and incubated, the preparation yielded only a labeledaxonal 4S RNA, presumably of mitochondrial origin
in the axon (EdstroÈ m et al., 1969). These results
appeared to support the earlier inference from spinalcord transection experiments (see above), that myelin
sheath associated oligodendrocyte nuclei were a prob-
able local source of axonal mRNA.
Results from in vitro pulse-chase experiments on the
11th cranial nerve root in the cat were also consistentwith the possibility of an extraaxonal source of RNA
(Koenig, 1970). Nerves were either `pulsed' for 1.5
hours with tritium-labeled RNA precursors, and`chased' for 4.5 hours in a nonradioactive incubation
medium containing unlabeled precursors in excess, or
pulsed only for 1.5 hours; a control nerve was alsoincubated for the full 6 hours in the radioactive med-
ium. Speci®c radioactivity (i.e., incorporated nucleotide
radioactivity/ng axoplasmic protein), assayed in thepulse-chase nerve was signi®cantly greater than in the
pulse-only nerve. Moreover, there was no signi®cant
di�erence between samples from the pulse-chase andcontrol nerves. The increase in speci®c radioactivity
during the chase period, which was similar in theunchased nerve, can only be explained by an enrich-
ment of radiolabeled RNA from an exogenous periax-
onal source.
In the squid, initial evidence for a glial-axonal trans-
fer of RNA was derived from an in vitro preparation
that lacked the cell bodies of the giant ®ber. �3H]uri-uridine uptake by the isolated giant ®ber resulted in a
modest level of radioactive RNA in axoplasm whenassayed at 4 hours, but which increased ®ve-fold in the
succeeding 4 hours, while RNA radioactivity in the
glial sheath remained essentially unchanged (Cutillo etal., 1983). A subsequent study characterized the radio-
labeled RNA species appearing in the axoplasm of the
isolated giant ®ber, and demonstrated that the threemajor classes, including tRNA, rRNA, and poly(A)+
RNA were present (Rapallino et al., 1988). Con®r-
mation that radiolabeled rRNA in axoplasm wasassembled into ribonucleoprotein particles was later
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6222

shown on the basis of sedimentation rate analysis, andCsCl2 density (Menichini et al., 1990).
It is clear that the most compelling evidence for alocal extraaxonal source of axoplasmic RNA comesfrom experiments in the squid giant ®ber system. Theclose apposition of glial and axonal membranes in thegiant ®ber (Villegas and Villegas, 1984) may well pro-vide a structural basis for permitting intercellulartransfer of various RNA species, including presumablyRNP particles as well (see above). As suggested by arecent report of apparent glial-axonal transfer of pro-teins, the mode of transfer does not appear to involveconventional secretory exo- and endocytic pathways(Sheller et al., 1995).
In vertebrate myelinated axons, there are specialstructural complexities at the interface between theaxon and its ensheathment. For example, at the para-node, which is where the myelin sheath attaches to theaxon, the cytoplasmic channel in the myelin sheath,comprising the so-called lateral belt, forms a spiral ofoverlapping cytoplasmic paranodal loops, in whicheach myelin lamella forms an axoglial junction. Theaxoglial junction is the only specialized zone of contactbetween the myelin sheath and the axon, a structurallynovel region where the two membranes are cross-bridged by periodic septa (Schnapp and Mugnaini,1978). The paranodal region, however, is at most afew micrometers in length, while the average internodeis 1±2 mm; therefore, the paranodal surface area isonly a very small fraction of the total membrane sur-face area of an internode. Although the almost crystal-line-like organization of the axoglial junction (Schnappand Mugnaini, 1978) would seem to argue against themembranes comprising the specialized adherent junc-tion engaging in dynamic exocytic/endocytic activity,the possibility that the junction could serve as a poten-tial pathway for intercellular macromolecular transfercannot be excluded. Neither, for that matter, can thepossibility be ruled out that there may be intercellularmacromolecular tra�c traversing the periaxonal cleftthat spans the internode between the inner cytoplasmicbelt and terminating Schmidt±Lanterman incisures ofthe myelin sheath and axon.
4. Local protein synthesis in presynaptic terminals
4.1. Historical introduction
Forty years ago, the mitochondrial fraction frombrain was found to contain a substantial proportion ofnonmitochondrial structures (Petrushka and Giuditta,1959), most of which were later identi®ed as shearedo� nerve terminals that had resealed into osmoticallysensitive particles that were referred to as synapto-somes (De Robertis et al., 1961; Gray and Whittaker,
1962). Criteria for their identi®cation were based oninclusion of synaptic vesicles and mitochondria, andoccasional attachment of postsynaptic membrane.When separated from free mitochondria, myelin andmembrane fragments by sedimentation throughsucrose (De Robertis et al., 1961; Gray and Whittaker,1962), or Ficoll gradients (Kurokawa et al., 1965;Abdel-Latif, 1966), nerve terminals represented themajor components of synaptosomal preparations, butremained contaminated to di�erent degrees by freemitochondria, membranes, and fragments of axons,dendrites, and glial processes (Autilio et al., 1968;Lemkey-Johnston and Larramendi, 1968; Lemkey-Johnston and Dekirmenjian, 1970; Morgan, 1970,1976; Cotman and Taylor, 1971; Gambetti et al., 1972;Joo and Karnushina, 1975; Henn et al., 1976; Verity etal., 1980; Rao and Steward, 1991; Whittaker, 1993).
These observations paved the way for a series of ex-periments to determine whether a system of proteinsynthesis was present in nerve terminals, as it had beenproposed for axons (Koenig and Koelle, 1961). An his-torical retrospective of developments in this ®eld indi-cates that it began as a brief initial phase, at whichtime the concept of presynaptic protein synthesisappeared to receive favorable support; a second, longerperiod of criticism ensued that ultimately led to a dis-crediting of the hypothesis, and a third, very recentphase, in which compelling evidence supporting theconcept once again became available, largely in invert-ebrate preparations. The very recent work in this areaindicates that a critical, comprehensive review of the®eld seems appropriate and warranted at this time.
4.2. Early studies
The investigation of protein synthesis in crude brainmitochondrial fractions served as the starting point ofthese studies. Except for the unusually high rates ofprotein synthesis, such brain fractions exhibited theusual features characteristic of mitochondrial prep-arations from other tissues, including resistance toRNase, lack of requirement for soluble factors, or ex-ogenous ATP, and dependence on oxidative phos-phorylation (Klee and Sokolo�, 1965; Campbell et al.,1966). A signi®cant di�erence emerged, however, whenprotein synthesis in brain mitochondrial fractions wasinhibited by CXM, a speci®c inhibitor of cytoplasmicribosomal protein synthesis (Yellin et al., 1967), in ad-dition to inhibition by chloramphenicol (CAP), aspeci®c inhibitor of mitochondrial protein synthesis.These observations implied that subcellular nonmito-chondrial particles, such as synaptosomes, were contri-buting to protein synthesizing activity.
The ®rst indication that synaptosomes were capableof protein synthesis came from experiments in whichrat brain slices were incubated with a radiolabeled
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 23

amino acid, and then subjected to subcellular fraction-ation to isolate synaptosomes (Austin and Morgan,1967). Under these conditions, radioactive proteinsincreased at a linear rate in synaptosomes, and in theirsoluble and particulate subfractions that were releasedby osmotic shock when synaptosomes were suspendedin a hypotonic bu�er. The absence of a lag periodbefore incorporation suggested that synaptosomal pro-teins were synthesized locally rather than having beenderived from neuronal cell bodies. The rapid appear-ance of newly synthesized synaptosomal proteins waslater con®rmed in brain slices (White et al., 1972), andin vivo (Droz and Barondes, 1969). On the otherhand, when radiolabeled proteins were recovered insynaptosomes following �14C]leucine injection in mousebrain, a long lag period of days was observed (Bar-ondes, 1964, 1966, 1967).
As a next logical step, brain synaptosomal fractionsfrom young rats were used in experiments of cell-freeprotein synthesis (Morgan and Austin, 1968). Inas-much as the rate of protein synthesis was known todecrease markedly with age (Verity et al., 1979), ratsless than one month old were used to obtain synapto-somal preparations, and this became common practicein most experiments on synaptosomal protein syn-thesis. It should be noted, however, that nerve term-inals undergo morphological modi®cations, and achange in their sedimentation properties during thisperiod (Gonatas et al., 1971). When puri®ed on a dis-continuous sucrose gradient and incubated in a lowionic strength medium, without soluble factorsrequired for protein synthesis, synaptosomes promotedthe incorporation of a radiolabeled amino acid intoproteins. The reaction remained linear for 30 min, wasresistant to RNase, was markedly inhibited by puro-mycin or CXM, and was partially inhibited by CAP,or other inhibitors of mitochondrial protein synthesis(Morgan and Austin, 1968). Fractionation of thelabeled synaptosomes indicated that only the synapticvesicle fraction lacked radioactive proteins. Thespeci®c radioactivity of soluble and membrane proteinswere comparable to those of intact synaptosomes,while it was approximately half as much in synaptoso-mal mitochondria. Interestingly, CXM blocked thesynthesis of soluble and membrane proteins, but didnot inhibit synthesis in synaptosomal mitochondria;however, CAP, or other mitochondrial inhibitors fullyblocked synthesis of mitochondrial proteins.
These data suggested that cytoplasmic and mito-chondrial systems of protein synthesis were present insynaptosomes, and that the soluble factors required bythe cytoplasmic system were intrinsic to synaptosomes.Inasmuch as RNA was in axons (EdstroÈ m, 1964a;Koenig, 1965; EdstroÈ m et al., 1969), and in synapto-somes (Austin and Morgan, 1967), and that membranefractions from other sources had been reported to lack
EM-detectable ribosomes while containing rRNA(Hallinan and Munro, 1965; Shapot and Pitot, 1966;Bergeron-Bouvet and MouleÁ , 1966), it was suggestedthat active cytoplasmic ribosomes were localized nearthe synaptosomal plasma membrane, and were therebyprotected from RNase hydrolysis, but remained unde-tected by conventional EM analyses (Morgan andAustin, 1968).
Synaptosomes prepared on a discontinuous Ficollgradient were better preserved, presumably becausethey were not exposed to an hypertonic sucrose med-ium (Abdel-Latif, 1966). They exhibited a markedlyhigher rate of protein synthesis when assayed in amedium containing 100 mM NaCl and 10 mM KCl(Autilio et al., 1968). In agreement with Morgan andAustin (1968), synaptosomal protein synthesis assayedunder these conditions remained linear for 20±30 min,was strongly inhibited by puromycin and CXM, mod-erately inhibited by CAP, and una�ected by RNase. Infurther agreement with Morgan and Austin (1968),bacterial or microsomal contamination was excluded,inasmuch as bacterial activity would have been prefer-entially inhibited by CAP, while microsomal basedsynthesis would have required the addition of ATPand soluble factors, and would also have been inhib-ited by RNase. EM analyses identi®ed 60% of synap-tosomal particulates as nerve terminals; i.e.,membrane-bound bodies 0.2±0.8 mm in diameter, sev-eral of which had attached postsynaptic structures.The remaining elements were considered postsynapticcomponents that only rarely contained ribosomes.
The subcellular localization of the system of proteinsynthesis in synaptosomes isolated by Ficoll was exam-ined shortly thereafter, using EM autoradiographicmethods (Cotman and Taylor, 1971; Gambetti et al.,1972). According to the former report, presynapticterminals accounted for 48% of the total proteinradioactivity, while membrane-bound vesicles contain-ing endoplasmic reticulum (ER) were much morelabeled, but accounted for only 9% of the total proteinradioactivity. The remaining radioactivity was associ-ated with unidenti®ed bodies, some of which was pre-sumed to derive from nerve endings. The authorsconcluded that presynaptic structures ``...de®nitely in-corporated label into polypeptides,'' but drew attentionto the heavily labeled vesicles containing ER and ribo-somes which resembled fragments of glial cells, and to``...some of the cytoplasmic vesicles [which] could arisefrom axonal or dendritic fragments.''
More detailed EM autoradiographic analyses werereported by Gambetti et al. (1972). When a mixture of�3H]amino acids was used to label proteins, 53% of thesilver grains was present in presynaptic endings, whichrepresented 70±75% of the identi®ed bodies, 34% waslocalized on non-presynaptic structures with ribosomes(representing 4±7% of the identi®ed bodies), 9% over
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6224

non-presynaptic structures without ribosomes (repre-senting 15±26% of the identi®ed bodies), and 5% overfragmented bodies. The grain density of presynapticendings was one-sixth that of non-presynaptic struc-tures with ribosomes, with the latter being suggestedto have been derived from glial cells or postsynaptic el-ements. Similar data were obtained using a single�3H]amino acid. The distribution of grains over presyn-aptic terminals was best explained by the assumptionthat 60% of the protein radioactivity was uniformlydistributed, and 40% was localized in the presynapticperiphery. The contribution of intraterminal mitochon-dria was quite limited, inasmuch as presynaptic end-ings lacking mitochondria were labeled, andsynaptosomes incubated with CAP exhibited a re-duction of only 20% of presynaptic silver grains.Under this condition, the distribution of radioactiveproteins was most consistent with the assumption ofan exclusive localization at the presynaptic periphery.The authors concluded that ``...presynaptic endings areindeed labeled in both experiments.''
The foregoing data were in agreement with thereport by Gilbert (1972) of a puri®ed synaptosomalmembrane fraction containing RNA (23 mg/mg pro-tein), which catalyzed the synthesis of protein whensupplemented with soluble factors, GTP, ATP, and anATP generating system. The activity was inhibited byRNase, CXM and puromycin, but not by CAP, oua-bain or KCN. Many of its translation products hadelectrophoretic mobilities di�erent from those ofmicrosomal translation products. This membrane frac-tion was likely to contain the polysomes responsiblefor the CXM-sensitive system of synaptosomal proteinsynthesis, although no evidence of its cellular or sub-cellular localization was provided.
Synaptosomal translation products resolved by gelelectrophoresis were attributed to the activity of thecytoplasmic or mitochondrial system of protein syn-thesis on the basis of their sensitivity to CXM or CAP,and in comparison with the translation products ofmodel systems, such as free mitochondria, culturedglial, or nerve cells. Newly-synthesized proteins withsizes below 50 kDa were considered mitochondrialtranslation products, while those with sizes above 50kDa were considered the products of cytoplasmic ribo-somes (Irwin, 1985; Loguercio Polosa and Attardi,1991).
4.2.1. Factors a�ecting synaptosomal protein synthesisSynaptosomal protein synthesis does not require ex-
ogenous ATP (Autilio et al., 1968; Morgan and Aus-tin, 1968), but is strongly dependent on oxidativephosphorylation, as shown by the marked inhibitionexerted by speci®c inhibitors of glycolysis, or of the tri-carboxylic acid cycle (Autilio et al., 1968; Morgan andAustin, 1969). These results suggested that synaptoso-
mal protein synthesis is driven by the ATP synthesizedby intrasynaptosomal mitochondria. The lack ofrequirement for exogenous ATP is due to the per-meability barrier of the synaptosomal membrane.When the membrane is damaged by osmotic shock,protein synthesis is almost completely lost, but can berecovered to a large extent upon addition of ATP andan ATP-generating system. Under these conditions,the activity becomes totally sensitive to RNase.
Synaptosomal protein synthesis is strongly depen-dent on the ionic composition of the incubation med-ium, and on its osmolarity (Autilio et al., 1968). Incomparison with the activity observed with 100 mMsucrose, a hypotonic condition, the addition of NaCl,or KCl induced a marked stimulation that was maxi-mal at 100 mM. A further increase was obtained whenboth salts were added together (i.e., 100 mM NaCl, 10mM KCl), while the reverse concentration (i.e., 100mM KCl, 10 mM NaCl) proved much less e�ective.The requirement for Na+ and K+ was in agreementwith the marked inhibition exerted by ouabain.
In a more detailed study (Appel et al., 1969), theincrement in synaptosomal protein synthesis broughtabout by Na+ and K+ was strongly correlated withthe activation of the membrane Na+, K+-ATPase,and with the resulting increase of synaptosomal K+
and O2 uptake. Increasing the concentration of exter-nal Na+ brought about a stimulation of mitochondrialstate 4, and of ouabain-sensitive Na+, K+-ATPase(Verity, 1972). The latter e�ect increased the pro-duction of ADP, and, in turn, of oxidative phosphoryl-ation, which eventually led to an enhanced proteinsynthesis.
In a later study, ouabain was found to inhibitboth CXM-sensitive and CAP-sensitive systems ofsynaptosomal protein synthesis, as a result of thedecrease in internal K+ (Verity et al., 1979). Indeed,a linear relationship was found between the rate ofprotein synthesis and the internal concentration ofK+. In addition, synaptosomal protein synthesis wasalso found to be developmentally regulated. In adultrats it was only 20% of that in 12 to 14 day-old rats,consistent with the marked reduction from 48 to 5%in response to ouabain inhibition. Ouabain also inhib-ited the increment in protein synthesis brought aboutby higher concentrations of external Ca2+.
Valinomycin, a K+ ionophore, inhibited synaptoso-mal protein synthesis (Verity et al., 1981), but thee�ect was independent of the external K+ concen-trations, and also independent of the increment in pro-tein synthesis that normally occurred at intermediateK+ concentrations (e.g., 2.6-fold at 25 mM). Thee�ect was not related to the concentration of intrasy-naptosomal K+, but to ATP production, inasmuch asmitochondrial uptake of K+ apparently interferedwith oxidative phosphorylation. However, as the
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 25

uptake of K+ by mitochondria would lead to a lowerconcentration of cytoplasmic K without changing theoverall synaptosomal K+ content, inhibition of proteinsynthesis might still be attributable to a decrease ofcytoplasmic K+ concentration. The relatively minordepletion of ATP (20%) in comparison with the stron-ger inhibition of protein synthesis (50%) appears tosupport this interpretation.
4.2.2. CommentaryOn the whole, this early evidence supported the pre-
sence of a cytoplasmic system of protein synthesis innerve terminals. However, doubts were soon raisedwith regard to the validity of this view, and severalalternative explanations were put forward to accountfor the data.
4.3. Alternative interpretations
4.3.1. CAP-resistant, CXM-sensitive protein synthesis inmitochondria and synaptosomes
One alternative explanation of the data was thatprotein synthesis in brain mitochondria and in synap-tosomes was resistant to CAP, and sensitive to CXM.It was based on experiments in which brain mitochon-dria and synaptosomes were incubated in a high Na+,low K+ medium (Gordon and Deanin, 1968; Gold-berg, 1971). It was suggested that synaptosomal pro-tein synthesis was only due to intraterminalmitochondria, since it was claimed that mitochondrialprotein synthesis was resistant to CAP and sensitive toacetoxyCXM. Previous data to the contrary wereattributed to bacterial contamination.
Nonetheless, when similar experiments were per-formed in the low Na+, high K+ medium required foroptimal mitochondrial activity, protein synthesis bycrude brain mitochondria was found to be sensitive toCAP, as in the case of liver mitochondria, althoughstill largely sensitive to CXM (Cunningham andBridgers, 1970). The latter feature was clearly re¯ectingthe presence of synaptosomes in the crude mitochon-drial fraction (Whittaker, 1968; Morgan, 1970). Com-parable data were reported for a brain mitochondrialfraction puri®ed on a sucrose gradient (Haldar, 1970),which was presumably contaminated by synaptosomes.On the other hand, brain mitochondria puri®ed on aFicoll gradient were entirely sensitive to CAP, andinsensitive to CXM (Haldar, 1971). Furthermore, sub-cellular fractions containing synaptosomes were sensi-tive to CXM in proportion to their synaptosomalcontent. Similar data were reported for brain mito-chondria and synaptosomes from mouse (Bridgers etal., 1971), and rat (Hernandez et al., 1971; Bosmanand Hemsworth, 1970).
The latter data were presumably not entirely convin-cing, because in a subsequent report (Deanin and Gor-
don, 1973) brain mitochondria and synaptosomes wereclaimed to exhibit evidence of a ``...hybrid particle con-taining both mitochondrial and cytoplasmic proteinsynthetic systems, structurally and functionally inte-grated.'' The authors added that ``...not all brain mito-chondria are associated with such a particle. Itsabsence in liver mitochondrial preparations is consist-ent with an assumption of its speci®c role in brain.''The main support for these claims came from theidenti®cation of 80S ribosomes but not mitochondrialribosomes in mitochondrial and synaptosomal frac-tions prepared on a sucrose gradient, while cyto-plasmic and mitochondrial rRNAs were both detected.Remarkably, however, while mitochondrial ribosomeswere not identi®ed in mitochondrial and synaptosomalfractions prepared on a sucrose gradient, cytoplasmicand mitochondrial rRNAs were both present. None-theless, although brain mitochondria, prepared on aFicoll gradient, exhibited only CAP-sensitive proteinsynthesis as reported by Haldar (1971) and Hernandezet al. (1971), and was even reported previously by thesame laboratory (Gordon and Deanin, 1968), thepossibility that the paradoxical discrepancy was due toexperimental artifact was not considered.
In the same paper (Deanin and Gordon, 1973), itwas also stated that ``...such particles exist in prep-arations of nerve endings and... contribute almost 50%of the radioactivity associated with these preparationswhile representing only 5±10% of the identi®ablestructures.'' This additional conclusion was based onunpublished autoradiographic results of Aghajanian,Gordon and Deanin, and claimed to be supported bydata in the literature (Morgan, 1970; Hernandez et al.,1971; Gambetti et al., 1972). However, it merits notingthat the ®rst two papers described the presence of only``rare'' ribosome-containing particles in brain mito-chondrial preparations, while in the latter publication,data/statements pertaining to the conclusion could notbe found.
4.3.2. A CAP-sensitive extramitochondrial system ofprotein synthesis
Another alternative proposal, sharply at variancewith the preceding speculation, was based on obser-vations of a synaptosomal membrane fraction puri®edon a CsCl gradient, which contained DNA (02 mg/mgprotein), and RNA (06 mg/mg protein) (Ramirez etal., 1972a, 1972b). The electrophoretic mobilities of theRNAs were comparable to those of mitochondrialrRNA, but 28S RNA, and some 9S and 4S RNAswere also present. Protein synthesis did not require theaddition of soluble factors, ATP or an ATP generatingsystem, and was strongly inhibited by CAP, and fullyresistant to CXM. The only three translation products(80, 62 and 17 kDA) were considered to be com-ponents of the synaptic junctional complex. The for-
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6226

mer two had mobilities di�erent from those of themore numerous mitochondrial translation products.Previous biochemical data supporting the presence of asynaptosomal CXM-sensitive system of protein syn-thesis were attributed to the variable purity of the frac-tions studied. Furthermore, EM autoradiographicdata, which provided evidence of a presynaptic CXM-sensitive system of protein synthesis (Cotman and Tay-lor, 1971; Gambetti et al., 1972), were mentioned onlyto point out that newly synthesized proteins in ident-i®ed synaptic endings accounted for merely 48% of thetotal synaptosomal protein radioactivity. The latterfraction, however, could hardly be considered insub-stantitial.
In a follow-up report (Ramirez, 1973), the conceptof an exclusive extramitochondrial CAP-sensitive sys-tem of synaptosomal protein synthesis was claimed tobe supported by the results of in vivo experiments inwhich cytoplasmic protein synthesis was suppressed byCXM. Under these conditions, CAP inhibited the syn-thesis of synaptosomal membrane proteins (notably ofthe junctional complex), and of free and synaptosomalmitochondrial proteins. Mitochondrial protein syn-thesis was only partially inhibited (050%), whichappeared to re¯ect a failure of either CAP or CXM toexert full inhibition; however, this result was not dis-cussed.
More recently, a system of protein synthesis, largelyinhibited by CAP, was described in cholinergic nerveterminals puri®ed from sheep synaptosomes by immu-noabsorption (Corbaton et al., 1991). In this exper-iment, synaptosomal protein synthesis was inhibitedapproximately 50% by either CXM or CAP, while thepuri®ed cholinergic terminals (less than 10% of thetotal synaptosomal population) were inhibited 75% byCAP and only 15% by CXM.
4.3.3. Nonribosomal dependent incorporation of aminoacids into synaptosomal proteins
A pH 5 precipitate prepared from the soluble frac-tion of rat brain synaptosomes was shown to incorpor-ate several radiolabeled amino acids into the threetroponin peptides (37, 26, and 20 kDa) (Carlin, 1977).The reaction proceeded linearly for only 10 min, wasnot inhibited by CXM, emetine, puromycin, RNase, orEDTA, and was moderately inhibited by CAP at veryhigh concentrations (25% at 0.8 mg/ml).
Comparable observations were reported with thesoluble fraction prepared from guinea pig brain synap-tosomes (Gower and Tytell, 1986), which catalyzed theincorporation of radiolabeled arginine into three pep-tides (50, 37 and 20 kDa), two of which with the sameMW of troponin peptides. This reaction, however, wasfully inhibited by RNase and puromycin, and appearedto require tRNA for the aminoacylation of preexistingproteins, in analogy with a similar reaction described
in mammalian brain (Barra et al., 1973; Laughrea,1982), and squid axoplasm (Ingoglia et al., 1983). Thisreaction was suggested to ``...explain amino acid ad-dition to macromolecules in the absence of ribosomes,thereby accounting for some of the prior observationsthat were interpreted as protein synthesis.'' Clearly, thelabeling of three peptides by a non-ribosomal mechan-ism cannot account for the synaptosomal CXM-sensi-tive synthesis of a much larger number ofsynaptosomal proteins (Gilbert, 1972; Irwin, 1985;Loguercio Polosa and Attardi, 1991; Rao and Steward,1991; Crispino et al., 1993a, 1993b, 1997).
4.3.4. Non-presynaptic ribosome-containing particles insynaptosomes account for most of the CXM-sensitivetranslational activity
Ribosome-containing particles of synaptosomalpreparations were suggested to be derived from glialcells, fragmented axons and/or dendrites (Morgan,1970; Cotman and Taylor, 1971; Gambetti et al.,1972). In the latter two reports, these particles wereshown by EM autoradiography to account for only afraction of the total proteins synthesized in synapto-some fractions. Nonetheless, on the basis of biochemi-cal observations, they were assumed to account formost of the CXM-sensitive synaptosomal activity(Hernandez, 1974; Jones et al., 1975; Verity et al.,1980).
In the former paper (Hernandez, 1974), this con-clusion was largely based on the claim that upon frac-tionation of a labeled synaptosomal fraction mostprotein radioactivity was associated with intraterminalmitochondria. Actually, only 30% of the newly syn-thesized proteins sedimented with intraterminal mito-chondria, while an additional 30% sedimented withdamaged synaptosomal particles, and most of theremaining radioactivity was associated with solubleand membrane synaptosomal fractions. In addition,the recovery of newly synthesized CXM-sensitive pro-teins was quite low, while that of the CAP-sensitiveproteins was more than expected. Furthermore, thesource of 21% of the total protein radioactivityremained unidenti®ed. In any case, the CXM sensi-tivity of synaptosomal protein synthesis was reportedin the same paper, in agreement with several otherreports, while relevant data from previous autoradio-graphic studies (Cotman and Taylor, 1971; Gambettiet al., 1972) were not discussed.
The presence of a cytoplasmic system of protein syn-thesis in synaptosomes was also claimed to be at oddswith an experiment in which less than 5% of the pro-teins synthesized by a crude mitochondrial fractioncould be recovered in the synaptosomal fraction puri-®ed after the incubation (Hernandez, 1974). Not con-sidered, however, was the possibility that subcellular
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 27

particles incubated at 378 might not retain the sedi-
mentation properties of native synaptosomes.
In the report by Jones et al. (1975), the origin of
presynaptic proteins was examined by EM autoradio-graphic analyses of the synaptosomal fraction prepared
from labeled brain slices. The data were interpreted to
indicate that ``...presynaptic nerve endings do not syn-thesize protein by any mechanism other than one due
to their mitochondria.'' However, ``...more than 50%
of the grains overlay particulate material, which couldnot be identi®ed on the basis of morphology... some of
which at least may well be contributed by nerve end-ings that have failed to retain their junction with post-
synaptic attachments.'' Thus, the conclusion cited
above was based on only 50% of the data!
In the paper by Verity et al. (1980), a labeled synap-
tosomal preparation fractionated on a linear sucrosegradient was found to exhibit a much higher speci®c
radioactivity in the bottom fractions than in the top
fractions. Cytochrome oxidase and the ribosome-con-taining non-presynaptic structures showed a similar
pattern. In addition, when native synaptosomes were
fractionated on the same sucrose gradient, the proteinsynthesizing capacity of the bottom fractions was
much higher than that of the top fractions. The
authors concluded that most of the synthesizing ac-tivity was due to subcellular structures other than pre-
synaptic endings. However, inasmuch as the data were
expressed in terms of protein speci®c radioactivityrather than as percent of the total translational ac-
tivity, they could o�er little, or no evidence that ribo-
some-containing elements accounted for the entiretranslational activity of synaptosomes.
In more general terms, it is not justi®ed to concludethat the high speci®c radioactivity of a single class of
synaptosomal particles containing ribosomes accounts
for most, or all of the translational activity of synapto-somal preparations. What is most relevant and import-
ant is the fraction of total synaptosomal radioactivity
that is accounted for by these particles. Moreover,ribosome-containing synaptosomal particles might not
be synthesizing proteins if soluble factors had leaked
out. Only EM autoradiographic analyses would pro-vide convincing evidence. In addition, the identi®cation
of presynaptic ribosomes cannot rest merely on the useof conventional EM methods, because unlike ESI
(Korn et al., 1983; Martin et al., 1989; see above), or
immunocytochemical methods (Sotelo et al., 1999),there are uncertainties regarding interpretation that
stem from potential ambiguities introduced by pre-
parative, or heavy metal staining procedures. Clearly,the widely accepted notion that presynaptic terminals
lack ribosomes may be traced to the general view
about the entire axonal compartment lacking ribo-somes; however, such a view is no longer tenable (for
recent review, see Koenig and Giuditta, 1999; also, seeabove).
Some of the ribosome-containing structures presentin synaptosomal fractions are derived from glial cells,as shown by experiments in which C-6 glial cells in cul-ture were used for the preparation of synaptosomes(Henn et al., 1976). Comparable results were obtainedusing a glia-enriched fraction from rat brain (Sieghartet al., 1978; Sieghart and Singer, 1979) and rat cerebralcortex, in which nerve endings and glial cells wereselectively labeled with radioactive GABA and b-ala-nine, respectively. On the other hand, fragments ofdendritic processes, called dendrosomes, were shownto be present in synaptosomal preparations from thesubstantia nigra (Hefti and Lichtensteiger, 1978). Den-drosomes are also present in brain cortex synapto-somes, in which they are actually considered to be themajor, if not the only structures capable of proteinsynthesis (Rao and Steward, 1991; Weiler and Green-ough, 1991, 1993). However, quantitative EM auto-radiographic data of the relative abundance of activedendrosomes, and of their contribution to the transla-tional capacity of synaptosomal preparations are notyet available.
4.3.5. CommentaryWhen examined critically, none of the data reviewed
above convincingly ruled out a presynaptic cyto-plasmic system of protein synthesis. The alternative hy-potheses on the nature of synaptosomal proteinsynthesis, attributed to a mitochondrial CXM-sensitivesystem, an hybrid mitochondrial/cytoplasmic system,or an extramitochondrial CAP-sensitive system, aremutually exclusive, lack adequate experimental sup-port, and disregard reproducible data in the literaturedemonstrating that a major cytoplasmic system of pro-tein synthesis coexists with a mitochondrial system insynaptosomes. As to the nonribosomal incorporationof amino acids into synaptosomal proteins, its charac-teristics are clearly distinctive from those of the synap-tosomal cytoplasmic translation system.
Finally, the conclusion that synaptosomal proteinsynthesis is not presynaptic, but is entirely due to glialfragments and/or post-synaptic elements was based ona biased selection of data that was then subject to overinterpretation and generalization. The data, moreover,were further conditioned by the invalid assumption ofa lack of protein synthesis in the axon compartment.Conclusive proof that cytoplasmic protein synthesisdoes exist in identi®ed presynaptic terminals is pre-sented in the following section.
4.4. Recent ®ndings
Convincing evidence that protein synthesis is presentin presynaptic endings has emerged recently from ana-
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6228
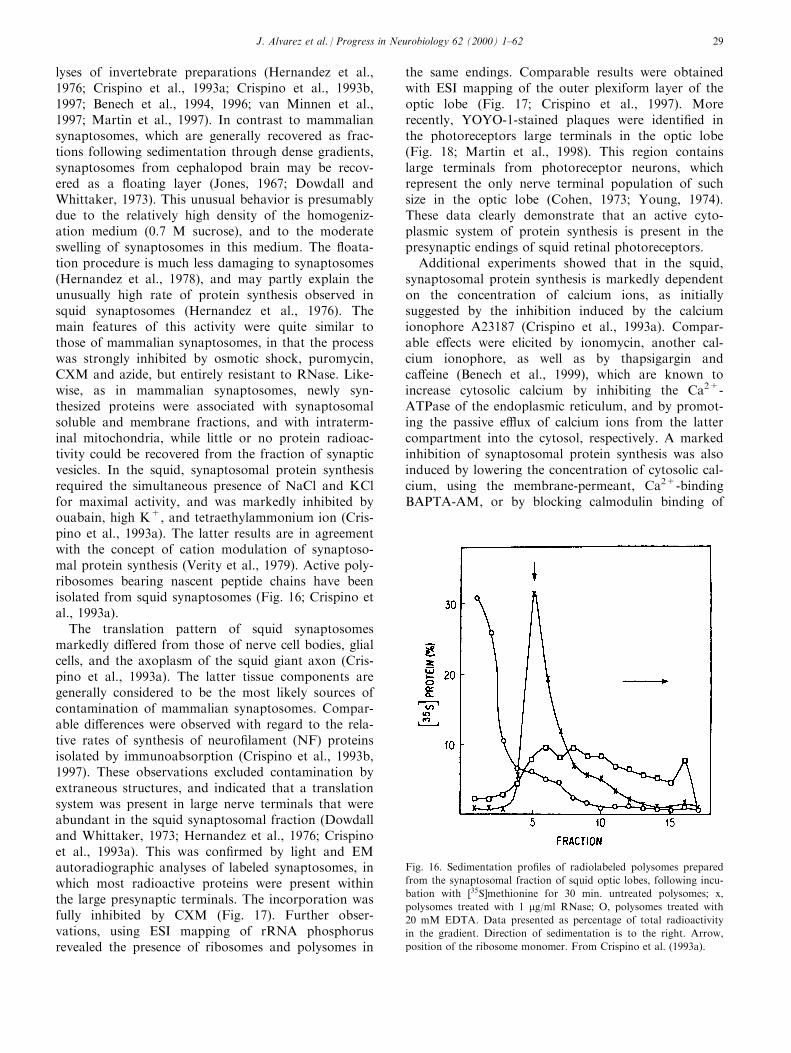
lyses of invertebrate preparations (Hernandez et al.,1976; Crispino et al., 1993a; Crispino et al., 1993b,1997; Benech et al., 1994, 1996; van Minnen et al.,1997; Martin et al., 1997). In contrast to mammaliansynaptosomes, which are generally recovered as frac-tions following sedimentation through dense gradients,synaptosomes from cephalopod brain may be recov-ered as a ¯oating layer (Jones, 1967; Dowdall andWhittaker, 1973). This unusual behavior is presumablydue to the relatively high density of the homogeniz-ation medium (0.7 M sucrose), and to the moderateswelling of synaptosomes in this medium. The ¯oata-tion procedure is much less damaging to synaptosomes(Hernandez et al., 1978), and may partly explain theunusually high rate of protein synthesis observed insquid synaptosomes (Hernandez et al., 1976). Themain features of this activity were quite similar tothose of mammalian synaptosomes, in that the processwas strongly inhibited by osmotic shock, puromycin,CXM and azide, but entirely resistant to RNase. Like-wise, as in mammalian synaptosomes, newly syn-thesized proteins were associated with synaptosomalsoluble and membrane fractions, and with intraterm-inal mitochondria, while little or no protein radioac-tivity could be recovered from the fraction of synapticvesicles. In the squid, synaptosomal protein synthesisrequired the simultaneous presence of NaCl and KClfor maximal activity, and was markedly inhibited byouabain, high K+, and tetraethylammonium ion (Cris-pino et al., 1993a). The latter results are in agreementwith the concept of cation modulation of synaptoso-mal protein synthesis (Verity et al., 1979). Active poly-ribosomes bearing nascent peptide chains have beenisolated from squid synaptosomes (Fig. 16; Crispino etal., 1993a).
The translation pattern of squid synaptosomesmarkedly di�ered from those of nerve cell bodies, glialcells, and the axoplasm of the squid giant axon (Cris-pino et al., 1993a). The latter tissue components aregenerally considered to be the most likely sources ofcontamination of mammalian synaptosomes. Compar-able di�erences were observed with regard to the rela-tive rates of synthesis of neuro®lament (NF) proteinsisolated by immunoabsorption (Crispino et al., 1993b,1997). These observations excluded contamination byextraneous structures, and indicated that a translationsystem was present in large nerve terminals that wereabundant in the squid synaptosomal fraction (Dowdalland Whittaker, 1973; Hernandez et al., 1976; Crispinoet al., 1993a). This was con®rmed by light and EMautoradiographic analyses of labeled synaptosomes, inwhich most radioactive proteins were present withinthe large presynaptic terminals. The incorporation wasfully inhibited by CXM (Fig. 17). Further obser-vations, using ESI mapping of rRNA phosphorusrevealed the presence of ribosomes and polysomes in
the same endings. Comparable results were obtainedwith ESI mapping of the outer plexiform layer of theoptic lobe (Fig. 17; Crispino et al., 1997). Morerecently, YOYO-1-stained plaques were identi®ed inthe photoreceptors large terminals in the optic lobe(Fig. 18; Martin et al., 1998). This region containslarge terminals from photoreceptor neurons, whichrepresent the only nerve terminal population of suchsize in the optic lobe (Cohen, 1973; Young, 1974).These data clearly demonstrate that an active cyto-plasmic system of protein synthesis is present in thepresynaptic endings of squid retinal photoreceptors.
Additional experiments showed that in the squid,synaptosomal protein synthesis is markedly dependenton the concentration of calcium ions, as initiallysuggested by the inhibition induced by the calciumionophore A23187 (Crispino et al., 1993a). Compar-able e�ects were elicited by ionomycin, another cal-cium ionophore, as well as by thapsigargin andca�eine (Benech et al., 1999), which are known toincrease cytosolic calcium by inhibiting the Ca2+-ATPase of the endoplasmic reticulum, and by promot-ing the passive e�ux of calcium ions from the lattercompartment into the cytosol, respectively. A markedinhibition of synaptosomal protein synthesis was alsoinduced by lowering the concentration of cytosolic cal-cium, using the membrane-permeant, Ca2+-bindingBAPTA-AM, or by blocking calmodulin binding of
Fig. 16. Sedimentation pro®les of radiolabeled polysomes prepared
from the synaptosomal fraction of squid optic lobes, following incu-
bation with �35S]methionine for 30 min. untreated polysomes; x,
polysomes treated with 1 mg/ml RNase; O, polysomes treated with
20 mM EDTA. Data presented as percentage of total radioactivity
in the gradient. Direction of sedimentation is to the right. Arrow,
position of the ribosome monomer. From Crispino et al. (1993a).
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 29
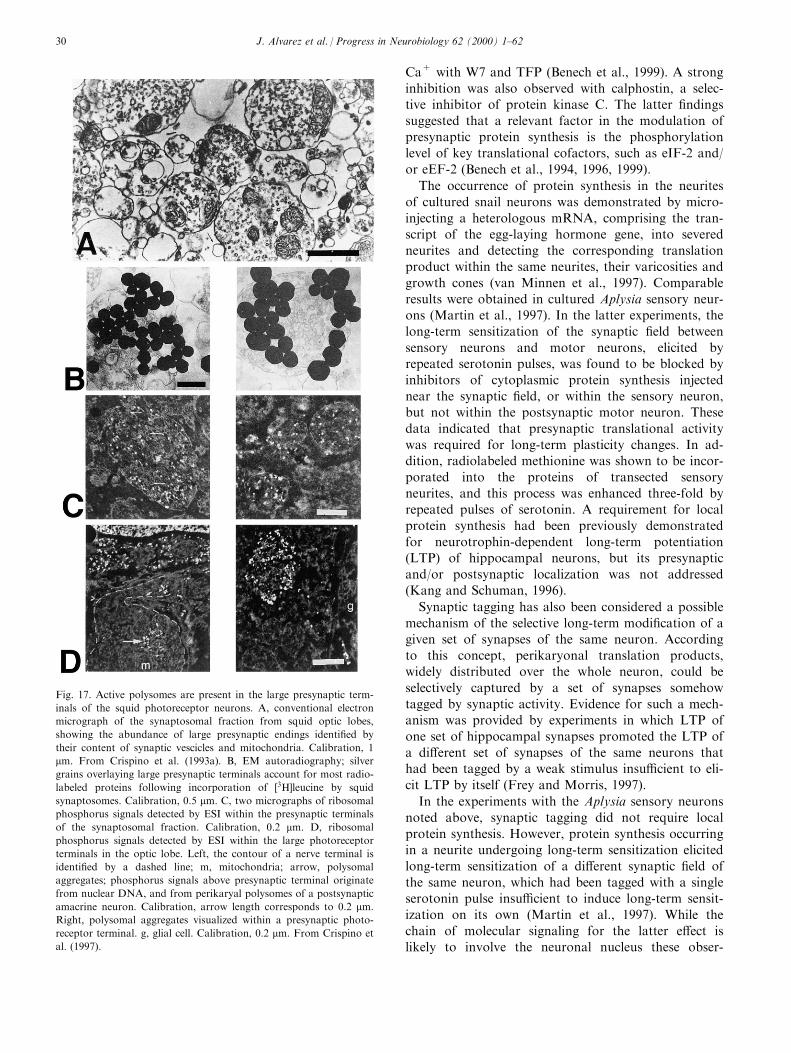
Ca+ with W7 and TFP (Benech et al., 1999). A strong
inhibition was also observed with calphostin, a selec-tive inhibitor of protein kinase C. The latter ®ndings
suggested that a relevant factor in the modulation ofpresynaptic protein synthesis is the phosphorylation
level of key translational cofactors, such as eIF-2 and/
or eEF-2 (Benech et al., 1994, 1996, 1999).
The occurrence of protein synthesis in the neurites
of cultured snail neurons was demonstrated by micro-
injecting a heterologous mRNA, comprising the tran-script of the egg-laying hormone gene, into severed
neurites and detecting the corresponding translationproduct within the same neurites, their varicosities and
growth cones (van Minnen et al., 1997). Comparable
results were obtained in cultured Aplysia sensory neur-ons (Martin et al., 1997). In the latter experiments, the
long-term sensitization of the synaptic ®eld between
sensory neurons and motor neurons, elicited byrepeated serotonin pulses, was found to be blocked by
inhibitors of cytoplasmic protein synthesis injected
near the synaptic ®eld, or within the sensory neuron,but not within the postsynaptic motor neuron. These
data indicated that presynaptic translational activity
was required for long-term plasticity changes. In ad-dition, radiolabeled methionine was shown to be incor-
porated into the proteins of transected sensory
neurites, and this process was enhanced three-fold byrepeated pulses of serotonin. A requirement for local
protein synthesis had been previously demonstrated
for neurotrophin-dependent long-term potentiation(LTP) of hippocampal neurons, but its presynaptic
and/or postsynaptic localization was not addressed(Kang and Schuman, 1996).
Synaptic tagging has also been considered a possible
mechanism of the selective long-term modi®cation of agiven set of synapses of the same neuron. According
to this concept, perikaryonal translation products,widely distributed over the whole neuron, could be
selectively captured by a set of synapses somehow
tagged by synaptic activity. Evidence for such a mech-anism was provided by experiments in which LTP of
one set of hippocampal synapses promoted the LTP of
a di�erent set of synapses of the same neurons thathad been tagged by a weak stimulus insu�cient to eli-
cit LTP by itself (Frey and Morris, 1997).
In the experiments with the Aplysia sensory neurons
noted above, synaptic tagging did not require local
protein synthesis. However, protein synthesis occurringin a neurite undergoing long-term sensitization elicited
long-term sensitization of a di�erent synaptic ®eld of
the same neuron, which had been tagged with a singleserotonin pulse insu�cient to induce long-term sensit-
ization on its own (Martin et al., 1997). While the
chain of molecular signaling for the latter e�ect islikely to involve the neuronal nucleus these obser-
Fig. 17. Active polysomes are present in the large presynaptic term-
inals of the squid photoreceptor neurons. A, conventional electron
micrograph of the synaptosomal fraction from squid optic lobes,
showing the abundance of large presynaptic endings identi®ed by
their content of synaptic vescicles and mitochondria. Calibration, 1
mm. From Crispino et al. (1993a). B, EM autoradiography; silver
grains overlaying large presynaptic terminals account for most radio-
labeled proteins following incorporation of �3H]leucine by squid
synaptosomes. Calibration, 0.5 mm. C, two micrographs of ribosomal
phosphorus signals detected by ESI within the presynaptic terminals
of the synaptosomal fraction. Calibration, 0.2 mm. D, ribosomal
phosphorus signals detected by ESI within the large photoreceptor
terminals in the optic lobe. Left, the contour of a nerve terminal is
identi®ed by a dashed line; m, mitochondria; arrow, polysomal
aggregates; phosphorus signals above presynaptic terminal originate
from nuclear DNA, and from perikaryal polysomes of a postsynaptic
amacrine neuron. Calibration, arrow length corresponds to 0.2 mm.
Right, polysomal aggregates visualized within a presynaptic photo-
receptor terminal. g, glial cell. Calibration, 0.2 mm. From Crispino et
al. (1997).
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6230

vations further extend the range of plastic eventsrequiring presynaptic protein synthesis.
4.5. E�ects of chemical and electrical stimulation
An early study of a crude brain mitochondrial frac-tion, most likely containing synaptosomes, may haveprovided the ®rst suggestive evidence of a neurotrans-mitter e�ect on synaptosomal protein synthesis (Camp-bell et al., 1966). More direct data were obtained witha synaptosomal fraction prepared on a Ficoll gradient(Goldberg, 1972), in which protein synthesis wasmarkedly inhibited by CXM. Under these conditions,dopamine, serotonin and DOPA produced 50% inhi-bition, while epinephrine and norepinephrine produced25% inhibition, and acetylcholine and GABA causedno e�ect.
Marked stimulatory e�ects were also elicited bydi�erent types of depolarizing treatments of a synapto-somal fraction, in which protein synthesis was likewisestrongly inhibited by CXM (Wedege et al., 1977). Thelargest increment of 400% was induced by electricalstimulation at a frequency of 100/s, applied during anincorporation period of 1 h. The e�ect was associatedwith an increased respiration. A smaller increase of150% was induced by veratrine. The enhanced syn-thesis elicited by electrical stimulation persisted in aCa2+-free medium, but was drastically reduced in aNa+-free medium. The largest increments of 488%and 361%, induced by electrical stimulation, occurredin the SDS-soluble and SDS-insoluble junctional com-plex proteins, respectively, while a less dramatic e�ectof 186% was obtained in Triton X-100-soluble pro-teins. The synthesis of the former proteins was directlyproportional to the amount of current passed throughsynaptosomes. Lower increments were obtained withveratrine and high K+. Tetrodotoxin did not a�ectprotein synthesis under conditions of high K+ concen-tration, but fully blocked the increment induced byveratrine, and markedly inhibited the e�ect of electri-cal stimulation. Interestingly, electrical stimulation, orveratrine did not modify protein synthesis in C-6glioma cells. Following electrical stimulation, severalsynaptosomal proteins in the size ranges of 10±18 kDaand 32±87 kDa became more prominently labeled.
The data described above led to the conclusion thatthe responsive structures were delimited by an electri-cally polarized plasma membrane enclosing activepolysomes and soluble factors, as expected of sealed-o� nerve terminals, or postsynaptic elements. Themarked response of junctional complex proteinssuggested that synthesis of the latter was likely tooccur at a postsynaptic site. Conversely, the lack ofstimulatory e�ects on glial cells suggested that glial-de-rived fragments did not contribute to the enhancedsynthesis, although intact glial processes could behave
in a di�erent way. It is of some interest that the inhi-
bition by CXM decreased to 58 from 70% followingelectrical stimulation, while CAP inhibition increased.
This suggested that mitochondria, or a mitochondrial-
like system, were more responsive than the cytoplasmicsystem of protein synthesis.
To further discriminate between nerve and glial cells
as sites of the stimulatory e�ects elicited by electricalstimulation, experiments were designed to take advan-
tage of the preferential uptake of glucose by neurons,
and its metabolic conversion to amino acids for incor-
poration into protein, and of acetate by glial cells(Abdul-Ghani et al., 1980). As expected, after intraper-
itoneal administration of �14C]glucose to adult rats,
electrical stimulation of the brachial plexus increased
radiolabeling of proteins in the sensory-motor cortexby 37%, an e�ect fully blocked by tetrodotoxin, while
it caused a 21% decrease when �14C]acetate was used.
A large share of the stimulatory e�ect observed withradiolabeled glucose was found in the synaptosomal
fraction of the cortex homogenate (Boyar et al., 1980).
Nonetheless, when electrical stimulation was directly
applied to the synaptosomal fraction, protein labelingwas enhanced to approximately the same degree when
�14C]glucose, or �14C]acetate was used (80% and 67%,
respectively). Thus, either precursor was capable ofreporting the stimulatory e�ect, presumably, as a result
of a dual stimulation of neuronal and glial elements,
although the capacity of nerve endings to incorporate
�14C]acetate into protein was not excluded.
A transient increment in content of synaptosomal
polysomes from rat cerebral cortex was induced by ex-
posure of synaptosomes to high K+ (Weiler andGreenough, 1991). Inasmuch as synaptosomes were
prepared by ®ltration of the homogenate through
pores of decreasing size, and elimination of the lowspeed sediment, the preparation contained mitochon-
dria, microsomes, and the cytosol. The largest increase
in polysomes of about 30% reached its maximum in
about 1 min, and decayed soon thereafter. Veratridineinduced a smaller increment, and tetraethylammonium
reduced, but did not block the increment in response
to high K+. The latter e�ect persisted in the presence
of CAP, thereby excluding a mitochondrial contri-bution. BAPTA-AM or EDTA inhibited the K+
e�ect, while the Ca2+ ionophore A23187 induced an
increased formation of polyribosomes, as measured bysedimentation analysis. Using UV irradiation to
``freeze'' the short-lasting K+-induced polysome aggre-
gation, the content of polysomes of small size was
found to increase 5 min after exposure of synapto-somes to high K+. In addition, 10 min afterward, the
incorporation of �35S]methionine increased 50%, but
soon reverted to the control value. These e�ects weretentatively attributed to the activation of initiation fac-
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 31

tors, but their subcellular localization was not investi-gated (Weiler and Greenough, 1991)
A de®nite postsynaptic localization was indicated,however, for the comparable e�ect induced by acti-vation of metabotropic glutamate receptors (mGluR)(Weiler and Greenough, 1993). In these experiments,an increase in synaptosomal polysomes was elicitedby glutamate or glutamate agonists, includingACPD, a selective mGluR agonist. This e�ect wassimilar to the K+ e�ect in extent and rate of onset,but displayed a more pronounced biphasic course, asthe polysome level decreased after 5 min, and wassoon followed by a second transient increment. Theglutamate stimulation was not blocked by APV, an an-tagonist of the NMDA receptor, and the quisqualatestimulation was not blocked by CNQX, a ionotropicreceptor antagonist. Further studies indicated that theactivation of protein kinase C induced a similar bipha-sic e�ect, while calphostin C, a speci®c inhibitor ofthis kinase, strongly inhibited the ACPD e�ect, and toa lesser extent the response to ibotenate or glutamate,but did not block the polysome aggregation inducedby high K+. The latter observations, and additionaldata, suggested that the K+ e�ect was a complexevent, not entirely correlated with the stimulation eli-cited by the activation of mGluR.
The above data stand in apparent contrast with theglutamate inhibition of synaptosomal protein synthesis(Autilio et al., 1968; Cheung, 1989), and of proteinsynthesis in rat brain slices (Orrego and Lipmann,1967). In the experiments by Cheung (1989), glutamateinhibition was limited to the fraction of synaptosomalprotein synthesis sensitive to ouabain, and was partlymediated by postsynaptic AMPA-speci®c quisqualatereceptors not involved in phospholipid breakdown.The apparent discrepancy between the latter data andthose by Weiler and Greenough (1993) may ®nd aplausible explanation in the complex time course ofthe glutamate e�ects, which induced more than oneround of polysome aggregation, each one lasting onlya few minutes. On the other hand, determination ofthe rate of protein synthesis required longer periods(Autilio et al., 1968; Cheung, 1989), which presumablyincluded inhibitory aftere�ects. Another di�erence wasconcerned with the age of the rats; namely, 18±20 dayold in the experiments by Autilio et al. (1968) andWeiler and Greenough (1993), but much older basedon body weights of 200±250 g in the experiments byCheung (1989).
4.5.1. CommentaryMost of the e�ects of chemical or electrical stimu-
lation of synaptosomal preparations have been attribu-ted to postsynaptic elements, based on the prevailingview that the axon compartment lacks the capacity tosynthesize proteins. Nonetheless, a potential contri-
bution by presynaptic structures has not beenexcluded, and should be further investigated by higherresolution methods of analysis. Indeed, the recent dem-onstration that presynaptic protein synthesis is essen-tial for the long-term sensitization of cultured Aplysianeurons (see above), which is associated with a majorincrease in presynaptic translational activity, is themost cogent demonstration to date that presynapticprotein synthesis may be up-regulated in response tofunctional activity.
4.6. Miscellaneous observations
In vivo administration of methyl mercury to adultrats induced a subacute intoxication, and a marked in-hibition of protein synthesis in cerebellar slices, and toa lesser extent, in cerebral slices (Verity et al., 1975,1977a). Following a latent period of 10 days, inhi-bition of protein synthesis was also observed in synap-tosomal preparations from either region. Methylmercury inhibited the rate of protein synthesis in cer-ebral slices or synaptosomes even when administeredin vitro. Replacement of Na+ and K+ by mannitolreduced synaptosomal protein synthesis by 40%, butthe residual activity was no longer inhibited by methylmercury.
An apparent inhibition of synaptosomal protein syn-thesis by thyroid hormones was traced to interferenceof the latter to the uptake of radiolabeled amino acids.In this respect, tri-iodothyronine compounds weremore e�ective than tetra- or di-iodothyronine (Verityet al., 1977b).
4.7. RNA in presynaptic endings
An interesting approach to studying presynapticRNAs entails identifying mRNAs expressed in axonal®elds of speci®c phenotype neurons in brain regionslacking the cognate cell bodies. A clear example is rep-resented by tyrosine hydroxylase (TH) mRNA, whichwas detected in brain regions of rat, such as cerebel-lum, striatum, and pituitary intermediate lobe, thatcontain catecholaminergic axonal projections but nocatecholaminergic cell bodies (Melia et al., 1994).These observations indicated that TH mRNA is pre-sent in the axons and presynaptic terminals of theseneurons. TH mRNA is locally regulated, because itsconcentration is doubled in the cerebellum after reser-pine administration, and is halved in the striatum afterselective lesioning of the cathecolaminergic input byipsilateral injection of 6-hydroxydopamine.
Data consistent with the presence and local trans-lation of TH mRNA in the terminal ®elds of periph-eral axons had been reported earlier in experiments inwhich TH activity markedly increased in cathecolami-nergic terminals in heart tissue following reserpine ad-
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6232

ministration (Thoenen et al., 1970). In contrast tocathecolaminergic cell bodies of stellate ganglia, inwhich TH mRNA also increased, the e�ect in heartoccurred with a delay of a few days, and persisted fora much longer time. Inasmuch as administration ofCXM during the period of TH increase signi®cantlyattenuated the increase of the heart enzyme, and therate of TH transport could only account for 12% ofsuch an increase, the data were interpreted to indicatea local synthesis of TH, rather than delivery of theenzyme by axonal transport.
The study of presynaptic RNA has more often beenconcerned with the analysis of synaptosomal prep-arations. Analyses of intraterminal mitochondria haveprovided the most straightforward results, but a siz-able body of data related to extramitochondrial RNAis also available.
4.7.1. Synaptosomal mitochondrial RNAIn brain synaptosomal preparations from young
rats, most newly synthesized RNA was identi®ed asmitochondrial rRNA (England and Attardi, 1976).RNA synthesis proceeded linearly for at least 2 h,yielding 16S, 12S and 4S RNAs as the prevalent radio-active species, but also trace amounts of RNAs sedi-menting ahead of 18S. Newly synthesized RNAssedimenting faster than 4S was fully suppressed byethidium bromide, and annealed to a satisfactoryextent with liver mitochondrial DNA. On the otherhand, newly synthesized 4S RNA was only partiallyinhibited by ethidium bromide and annealed less e�-ciently with mitochondrial DNA. The latter ®ndingsuggested that cytoplasmic 4S RNA was also syn-thesized. EM autoradiographic analyses indicated that50% of the newly synthesized ethidium bromide-sensi-tive RNA was associated with presynaptic endings.Interestingly, in comparison with 10 day-old rats, therate of RNA synthesis by synaptosomal mitochondriamarkedly decreased in rats of 30 days (i.e., a 5 to 6-fold decrease with regard to 12S and 16S rRNAs), andin rats of 60 days the labeling of mitochondrialrRNAs was barely detectable.
Likewise, a higher rate of mitochondrial protein syn-thesis was present in synaptosomal preparations fromyoung rats in comparison with older animals (Loguer-cio Polosa and Attardi, 1991). The rate markedlydecreased in the third week, but remained essentiallythe same in older animals up to 2 years of age. Insynaptosomal mitochondria, the peak of translationalactivity coincided with a marked increase in cyto-chrome oxidase, and with the period of intense synap-togenesis, but was not correlated with the level ofmitochondrial mRNAs. Notably, one of the peptidesof NADH dehydrogenase (ND5) could not be detectedin the mitochondrial translation products, and an ad-ditional peptide (ND3) was barely labeled, while the
concentration of the corresponding mRNAs was nor-mal. These observations indicated that gene expressionin synaptosomal mitochondria is mainly regulated atthe level of translation. It is unclear whether the sameconclusion applies to cell body mitochondria. Thetranslation products of synaptosomal mitochondria,resolved by gel electrophoresis, were strongly inhibitedby CAP, and, surprisingly, by emetine, although notby CXM. Most of the newly synthesized proteins sen-sitive to CXM migrated as high molecular weightbands, but a few of them displayed apparent molecularweights below 50 kDa, and were present among mito-chondrial translation products.
4.7.2. Synaptosomal extramitochondrial RNAA less de®ned set of data concerns synaptosomal
extramitochondrial RNA. In an early paper (Austinand Morgan, 1967), synaptosomal mitochondria andfree mitochondria were shown to contain similaramounts of RNA (12±13 mg/mg protein), and DNA(6.3 mg/mg protein). With the exception of synapticvesicles, most synaptosomal subfractions containedcomparable, or even higher concentrations of RNA(up to 20 mg/mg protein), but only trace amounts ofDNA. The nature of synaptosomal RNA, and its cel-lular site of synthesis, are only partially known.
In addition to mitochondrial rRNAs, synaptosomalpreparations were often reported to contain cyto-plasmic rRNAs and 4S RNA, which have generallybeen attributed to contamination by extraneous cellfragments (England and Attardi, 1976; LoguercioPolosa and Attardi, 1991; DeLarco et al., 1975;Cupello and Hyde n, 1975). In the latter report, how-ever, a 7S RNA (89 kDa) and an unusually largeRNA (>60S) were described as speci®cally localized insynaptosomal preparations from adult rats. The 7SRNA was occasionally detected in subsynaptosomalfractions, in association with cytoplasmic rRNAs and4S RNA (Di Battista and Cupello, 1988). As reportedin the same paper, intraterminal mitochondria con-tained cytoplasmic rRNAs and 4S RNA, as well asmitochondrial rRNAs; the fraction of synaptic vesiclescontained 5S RNA, in addition to 4S RNA and cyto-plasmic rRNAs, and the soluble synaptosomal fractioncontained only 4S RNA. The latter observation is inagreement with the presence of 4S RNA in the axonalcompartment (for review, see Koenig and Giuditta,1999). When considered with the reports of synaptoso-mal cytoplasmic rRNAs, it is also in agreement withthe presence in synaptosomes of a cytoplasmic systemof protein synthesis.
In young rats, brain synaptosomes and mitochon-dria were reported to contain a much higher pro-portion of poly(A)+ RNA than polysomes (18% ascompared to 7%) (DeLarco et al., 1975). Moreover,the poly(A)+ tail of synaptosomal RNA was half as
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 33

long as that in polysomal RNA (viz., 84 nt as opposedto 169 nt), in agreement with comparable observationson axonal poly(A)+ mRNAs (Mohr et al., 1991).Synaptosomal membranes and mitochondria had asimilar proportion of poly(A)+ RNA (15.5% and13.5%, respectively), but mitochondrial RNA was 14times more active with regard to template activity(DeLarco et al., 1975). Lower concentrations of 8.3%and 8.7% poly(A)+ RNA were later reported forsynaptosomes and mitochondria from adult rats, re-spectively (Cupello and Hyde n, 1977). In addition,upon electrophoretic fractionation, synaptosomalpoly(A)+ RNA appeared to be distributed in a widesize range (up to 30S) with a prevalence of high mol-ecular weight components, in analogy with the distri-bution of microsomal poly(A)+ RNA. A 7S RNAspecies was present in the synaptosomal poly(A)+
RNA fraction, and to a minor extent in the mitochon-drial fraction, but was absent in microsomes. In adultrats, the fraction of poly(A)+ RNA was somewhatmore abundant in brain mitochondria than in micro-somes (7.7% and 5.8%, respectively), but containedlower molecular weight components (Cupello andRosadini, 1976).
The synthesis of synaptosomal RNA was examinedin young rats, following the intracranial administrationof �3H]uridine and �3H]adenosine (DeLarco et al.,1975). At the earliest time period examined (1 h), thespeci®c radioactivity (SA; cpm/mg RNA) of synapto-somal RNA was lower than that of mitochondrialRNA, and polysomal RNA (one-half and one-fourth,respectively), but became higher than that of mito-chondrial RNA after 8 h (2.5-fold), and comparable tothat of polysomal RNA after 24 h (three-fourths).Throughout this period, the proportion of newly syn-thesized poly(A)+ RNA was higher in synaptosomes(26%, 38%, and 19%, respectively) than in mitochon-dria (17%, 27%, and 17%, respectively), polysomes(10%, 7%, and 6%, respectively), and nuclei (i.e.,17%, 18%, and 12%, respectively). As a result, the SAof poly(A)+ RNA calculated from the published litera-ture was markedly higher in synaptosomes than inpolysomes at 8 and 24 h (Fig. 19).
The subcellular distribution of newly synthesizedbrain RNA was also examined in adult rats 1 h afterthe intraventricular injection of �3H]uridine (Cupelloand Hyde n, 1982). Synaptosomes contained only aminor amount of newly synthesized RNA (0.3% ascompared to 8%, and 0.9% in microsomes and mito-chondria, respectively), but the proportion of newlysynthesized poly(A)+ RNA in synaptosomes (18%)was slightly higher than in mitochondria (15±16%),and markedly higher than in microsomes and nuclei(10% and 9%, respectively). While generally lowerthan those reported by DeLarco et al. (1975) foryoung rats, these values con®rmed the prevalence of
newly-synthesized poly(A)+ RNA in synaptosomes.The subcellular distribution of newly-synthesized RNAand poly(A)+ RNA was not modi®ed in rats trainedfor the reverse handedness task (Cupello and Hyde n,1982).
4.7.3. CommentaryObviously, the identi®cation of cytoplasmic RNAs
in synaptosomes (rRNA, tRNA, and mRNA) does noto�er any indication of their subcellular localization.Fragments of glial and postsynaptic processes are gen-erally assumed to contain these RNAs, while their ad-ditional localization in presynaptic elements has rarelybeen considered because of the prevailing prejudiceagainst protein synthesis in the axon compartment.However, ribosomes and polysomes have recently beenidenti®ed in presynaptic endings of squid (Figs. 16 and17), snail neurons (van Minnen et al., 1997), and term-inals of M-axon collaterals (Fig. 12). In addition, puta-tive presynaptic mRNAs, such as GAT-1 and synapsinmRNAs have been identi®ed in rat brain synapto-somes (Crispino, Perrone Capano, Aiello, Iannetti,Cupello and Giuditta, unpublished data). Therefore, itseems likely that the complement of cytoplasmicRNAs may also be localized in presynaptic terminals.Of the few studies on cytoplasmic RNA in synapto-somes, the most intriguing observation concerns itssurprisingly high content of newly synthesizedpoly(A)+ RNA (Fig. 19).This result may imply anunusually fast turnover of this RNA species. Whenconsidered together with other related data, it shouldstimulate further investigation.
4.8. Concluding commentary
After a long period of scepticism, recent data haverevived and con®rmed the original hypothesis of pre-synaptic protein synthesis. Furthermore, they have
Fig. 19. Speci®c activities of poly(A)+ RNA in synaptosomes (.)and polysomes (R) isolated from rat brain at di�erent times after
the intracranial injection of �3H]uridine. Recalculated from data pre-
sented in Table 2 of DeLarco et al. (1975).
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6234

demonstrated a functionally relevant role in long-termsynaptic events involving synaptic modi®cation.Although these data are derived from a limited num-ber of synaptic systems in invertebrates, it is reason-able to expect that local protein synthesis supports thesteady state of presynaptic terminals as well as theirmodi®cations. Nevertheless, whatever the ®nal out-come, it is fair to say that the importance of the datareviewed above rests on having revealed an entirelynew ®eld of inquiry dealing with molecular mechan-isms of synaptic plasticity elicited by appropriate phys-iological and pathological conditions. Previously, asearch for explanatory mechanisms have been soughton the premise that presynaptic proteins were exclu-sively derived from the neuronal perikaryon. It maynow be expected that the demonstration of a presyn-aptic translation system will generate a new impetusfor investigating the role that such a system couldplay, the cellular origin of its components, characteriz-ation of the translation products, and mechanisms forregulating activity and expression.
5. Essay on the signi®cance of local protein synthesis inaxons
In this section, an emphasis is placed on the conceptof a local source of proteins to explain the mainten-ance and dynamic responses of axons and terminals.Regional anatomical modi®cations of mature axonsinduced by experimental manipulation are reviewed,and the local protein synthesis model is contrastedwith that of the slow transport model in explaining ex-perimental data. Clearly, in order to withstand criticalscrutiny and have utility, a model must be capable ofexplaining experimental observations, and have predic-tive power. The comparative analysis below highlightsthe ¯aws of the slow transport model by showing thatit fails to take metabolic turnover of axoplasmic pro-teins into account, and to explain biological complex-ities of the axon and terminals without having toinvoke additional ad hoc assumptions. Thus, theanalysis shows that the slow transport model has out-lived its usefulness as an exclusive theoretical con-struct.
5.1. Local synthesis as a model for maintainingaxoplasm
5.1.1. The modelThe basic premise is that proteins are synthesized
and degraded within each axonal segment and in term-inals. The key features of the model, which are sup-ported by evidence reviewed in previous sections,include the following. First, amino acids taken up intoaxoplasm are free to di�use, and become incorporated
into proteins at sites of synthesis. Second, amino acidsliberated after protein degradation are either reutilized,metabolized, or leak out into the extracellular space.Third, the activity of the machinery is submaximalunder steady state conditions, and can be regulated byenvironmental signals (Fig. 1). Finally, translocationof proteins is limited to short range redistributionfrom site of synthesis to local sites of axoplasmic inte-gration. This model accounts for uniform axons of anylength, and for any potential regional structural vari-ations.
5.1.2. Slow wave of radioactivity and stationary proteinsThe local synthesis model explicitly excludes long
range slow rate translocation of proteins in the axon.Therefore, its ®rst challenge is to account for theobserved slow wave of radioactivity that invades theaxon when radiolabeled precursors are delivered inthe vicinity of perikarya. As described below, withonly basic assumptions of local synthesis and no longrange slow rate motility of proteins, the model gener-ates simulated waves of radioactivity that are com-parable to experimental waves (Alvarez and Torres,1985).
Under conditions of simulation, when a pulse oflabeled amino acid is delivered in vicinity of perikarya,the precursor is taken up by perikarya and proximalaxons.
Because proteins that are being synthesized re¯ectthe gradient of speci®c radioactivity of the precursorpool in axoplasm, the peak of radioactive axonal pro-teins is at a maximum in the initial segment, and thendecreases with distance (Fig. 20, Pulse).
At all times, the axial di�usion of the labeled aminoacid between any two adjacent axon segments resultsin a net transfer from the proximal region of higherspeci®c radioactivity to a contiguous distal region oflower speci®c radioactivity. In this manner, labeledproteins will appear distally by de novo synthesis.With time after the pulse, proteins undergoing degra-dation will release their labeled residues into the sol-uble amino acid pool. When the speci®c radioactivityof the amino acid pool falls in the perikaryon, therewill be a net retrograde transfer of radiolabeled aminoacid from the initial segment into the perikaryon, aswell as into the contiguous downstream axon segment.As a free radiolabeled amino acid, it may be eitherreutililized, metabolized, or it may leak out of theaxon. A greater loss of radiolabeled residues from pro-teins of segments close to the perikaryon will reducethe content of radioactivity, and the peak of radioac-tivity will be displaced to an adjacent downstream seg-ment (Fig. 20, late time point). As the speci®cradioactivity of the precursor pool diminishes due toconstant leakage out the axon, the total amount oflabeled amino acid in proteins decreases progressively.
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 35

The model can be expressed quantitatively by twosimultaneous di�erential equations (Fig. 1), in whichthe ®rst describes the bi-directional transfer of aminoacids between soluble pools of adjacent segments, andto the extracellular ¯uid, and the second describes thetransfer of amino acids (i.e., synthesis and degra-dation) between the amino acid pool and proteins(Alvarez and Torres, 1985).
To model a real axon using the equations, values foramount and half-life of proteins, size of the pool ofamino acids, and coe�cient of di�usion of aminoacids in axoplasm were taken from data published inthe literature. The system was assumed in a steadystate, whereby the rate of synthesis could be calcu-lated, and the leakage of amino acids was estimated.The numerical solution yielded the spatio-temporalevolution of labeled proteins in a 50 mm axon after apulse of presursors was supplied to the perikaryon.Fig. 21 depicts two simulated waves (Alvarez andTorres, 1985), which are to be compared to the elegantexperimental waves reported by Cancalon (1979) in astudy of the olfactory nerve in the gar®sh. The exper-imental waves were satisfactorily simulated by thelocal synthesis model, in which a distal displacement
of the peak occurred, notwithstanding the fact thataxoplasmic proteins were stationary.
A peak appears in the simulated waves because thenet loss of label from the segments in advance of thepeak was smaller than that in the peak, or in the seg-ments trailing it. This phenomenon was observed byHo�man and Lasek (1975); Fig. 1 of their paper) in anexperiment in which radiolabeled precursors were sup-plied to perikarya of the sciatic nerve of the rat. Atday 10, the peak was still close to the origin, and thesegment located 100 mm distally contained 5000 dpm.At day 135, when the peak of radioactivity wasmapped to the latter distal segment, it contained only500 dpm. It is clear, therefore, that the distal segmentdid not gain radioactivity, but rather that interveningsegments had lost relatively more radioactivity. Thesedata contradict the assumption of metabolic stabilityof axonal proteins required by the slow transportmodel (see above), and indicate that the observedwaves can be accounted for by ordinary turnover ofstationary proteins. The local synthesis model gener-ates slow moving waves that ¯atten with time, whiletotal radioactivity decreases continuously. In manyrespects, the model reveals details of experimentalresults that can easily escape attention.
Fig. 21. Experimental and simulated radioactive waves in axons, in
which normalized dpm (nDPM) is plotted against distance from the
perikaryon, in mm. Gar®sh: experimental results (curves redrawn
from Cancalon, 1979 with permission of Lippincott, Williams and
Wilkins), in which perikarya of the olfactory nerve were given a
pulse of radioactive amino acid at time 0. The curves show the distri-
bution of label along the nerve at the indicated days post-pulse.
Arrows have been added to highlight the two types of displacement
of the peak of radioactivity with time; i.e., the forward movement
and the progressive reduction of radioactivity. Note that all segments
on the proximal side of the peak lose label, while segments distal to
the peak gain label, and at the same time, the slope of the radio-
active front decreases. Note that segments more proximal to the peri-
karyon lose relatively more radioactivity than more distal segments
(compare peaks of radioactivity in corresponding segments at days
35, 45 and 56). Model: using the equations shown in Fig. 1, and par-
ameters described in Alvarez and Torres (1985 with permission of
Academic Press), the model simulates the salient features of the ex-
perimental curves shown in the left panel when a simulated pulse of
label is applied at time 0 (redrawn from Alvarez and Torres, 1985).
Note that although the peak of radioactivity moves at 0.85 mm/day,
axonal proteins are stationary.
Fig. 20. Diagrams that depict how a slow wave of radioactivity mov-
ing through stationary proteins arises in the local synthesis model
(redrawn from Alvarez and Torres, 1985). Upper diagram: the peri-
karyon and axon contain unlabeled proteins (P), and when a pulse
of radioactive amino acid initially enters the perikaryon (solid
arrow), it produces a proximo-distal gradient in the speci®c radioac-
tivity of the amino acid pool (A) along a longitudinal series of axon
segments. As proteins undergo synthesis (S) and degradation (D),
there is a corresponding gradient in radiolabeling of proteins (P�).Horizontal arrows depict bidirectional axial di�usion of A within the
axon, and uptake and leakage across the axolemma. The fractional
content of radioactivity is depicted by the solid proportion of each
arrow. Lower diagram: a later time point at which A� is lost from
the ®rst axonal segment to the perikaryon, to the extracellular space
and to the contiguous distal segment, and the rest is reutilized locally
for synthesis. The reduction in speci®c radioactivity of A results in a
net loss of label from P� in the segment. In comparison, the content
of radioactivity in proteins may be greater in the succeeding axonal
segments. Thus, the net loss of label from proteins in upstream seg-
ments, and the net gain in downstream segments produces a forward
displacement of the peak with time.
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6236

5.2. Explanatory power: slow transport versus localsynthesis models
The data presented in previous sections indicatebeyond reasonable doubt that axoplasm (i) incorpor-ates amino acids by a mechanism sensitive to inhibi-tors of cytoribosomal protein synthesis, (ii) containsrRNAs, a full complement of tRNAs, mRNAs cod-ing for axoplasmic proteins, and active polyribo-somes. Conventional interpretation of these ®ndingsleads to the local synthesis model. In contrast, adher-ents of an exclusive slow transport model assumethat there is no protein synthesis in axons, and thatthe incorporation of amino acids and the function ofaxoplasmic RNAs must be unrelated to protein syn-thesis. For example, incorporation of amino acids,which is sensitive to CXM, has been explained byamino acylation of axoplasmic tRNAs (Black andLasek, 1977), or of amino acylation of existing pro-teins (Black and Lasek, 1977; Chakraborty et al.,1986). The proposed role of tRNA has either beenthat it is of no functional signi®cance (Black andLasek, 1977), or that it may serve to amino acylateexisting proteins (Ingoglia et al., 1983; Zanakis et al.,1984). Axoplasmic mRNAs have been regarded asnontranslatable (Jirikowski et al., 1990), as an adven-titious over¯ow from the perikaryon into the axon(Mohr and Richter, 1992a, 1992b), or to serve evenas a reserve for the perikaryon (Wensley et al., 1995).To our knowledge, data documenting rRNA andribosomes in axons and terminals have not been for-mally assimilated into the slow transport model.
Important selected experimental ®ndings arebrie¯y reviewed below, which highlight signi®cantfailings of the slow transport model to providesatisfactory explanations, in contrast to the localsynthesis model.
1. In the slow transport model, the set of curves inFig. 21 from experiments in gar®sh by Cancalon(1979) is purported to represent moving (motile)proteins that have left the perikaryon. An equallyvalid interpretation consistent with the model is thatthe waves represent the spatial distributions of 35,45, and 56-day old proteins in olfactory axons.Thus, proteins ranging widely in age overlap andare interspersed in axons; therefore, the model pre-supposes that the amount of protein in a given axonsegment is the aggregate sum of proteins comprisingthe whole range of ages. It follows, then, that theprotein content in distal segments should be verysmall compared to that of proximal segments ofaxons, and the di�erence would be exaggerated ifwaves of unlabeled proteins of every age were to beincluded. Thus, while the slow transport model anddata predict tapering of axons, real axons are
cylindrical and nontapering, which gives rise to aconundrum that requires additional ad hoc assump-tions. The local synthesis model, in contrast, gener-ates waves similar to the experimental waves, andaccounts fully for the constant caliber of the axonbecause each segment is in a steady state, and allsegments are similar.
2. Radioactive amino acid derived from so-calledslowly transported proteins in optic nerves of themouse are released into the bathing medium, andthe rate of release increases three-fold after inhi-bition of protein synthesis (Nixon, 1980). These ob-servations indicate that these proteins undergodegradation, and that released residues are e�-ciently reutilized locally. The complementary pro-cesses of synthesis and degradation associated withprotein turnover of axoplasmic proteins have notbeen incorporated into the slow transport model. Incontrast, however, local turnover of proteins is abasic tenet of the local synthesis model (see above).
If axonal proteins undergo degradation and someof the labeled precursor leaks out of the axon, thelocal synthesis model predicts that a very smallamount of the radioactivity should reach the term-inals by the slow wave. This was, indeed, found byDroz et al. (1973), in parasympathetic preganglionic®bers, less than 15 mm long, in which over 90% ofthe radiolabeling disappeared en route to the term-inals.
3. Various components moving at di�erent rates in theslow wave have been resolved (Willard et al., 1974).According to the slow transport model, each com-ponent is an independent motile polymer matrix(Lasek, 1986). In the local synthesis model, how-ever, the displacement of the radioactive peak as-sociated with a protein is an inverse function of thelife span of that protein. In a computer simulation,proteins with di�erent half-lives generate indepen-dent simulated waves in the axon that move atdi�erent rates (Fig. 22).
4. In optic nerve ®bers of the gold®sh, Grafstein et al.(1981) reported that the peak of radioactivity wasalways in the ®rst segment, and that the slopediminished progressively with time (Fig. 23) tobecome horizontal. The decreasing slope was takenas an index of transport. In the local synthesismodel, this type of slope appears if the exchange ofradioactive amino acid between perikaryon and in-itial axon segment cancels out during the postpulseperiod (Fig. 23).
5. Grafstein and Alpert (1982) reported that nerve sec-tion did not a�ect transport in optic ®bers of thegold®sh, but that systemic inhibition of protein syn-thesis arrested the moving wave in intact axons. Toaccount for these results without compromisingbasic tenets of the slow transport model (Fig. 24),
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 37
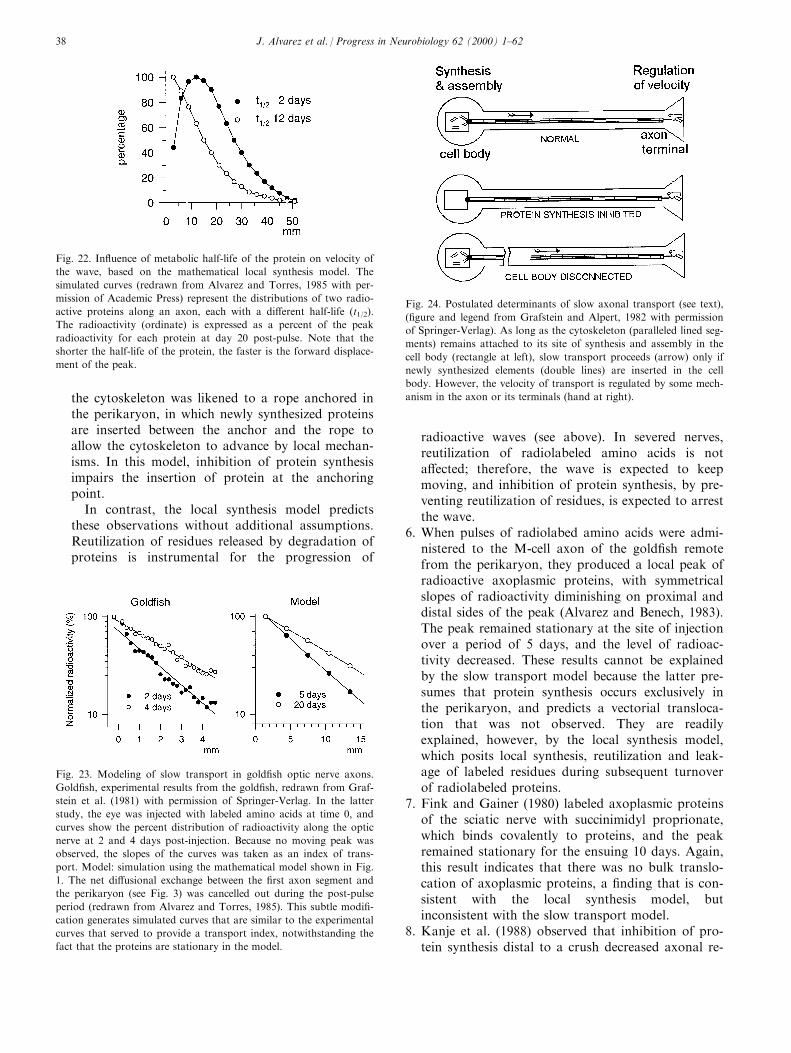
the cytoskeleton was likened to a rope anchored inthe perikaryon, in which newly synthesized proteinsare inserted between the anchor and the rope toallow the cytoskeleton to advance by local mechan-isms. In this model, inhibition of protein synthesisimpairs the insertion of protein at the anchoringpoint.
In contrast, the local synthesis model predictsthese observations without additional assumptions.Reutilization of residues released by degradation ofproteins is instrumental for the progression of
radioactive waves (see above). In severed nerves,reutilization of radiolabeled amino acids is nota�ected; therefore, the wave is expected to keepmoving, and inhibition of protein synthesis, by pre-venting reutilization of residues, is expected to arrestthe wave.
6. When pulses of radiolabed amino acids were admi-nistered to the M-cell axon of the gold®sh remotefrom the perikaryon, they produced a local peak ofradioactive axoplasmic proteins, with symmetricalslopes of radioactivity diminishing on proximal anddistal sides of the peak (Alvarez and Benech, 1983).The peak remained stationary at the site of injectionover a period of 5 days, and the level of radioac-tivity decreased. These results cannot be explainedby the slow transport model because the latter pre-sumes that protein synthesis occurs exclusively inthe perikaryon, and predicts a vectorial transloca-tion that was not observed. They are readilyexplained, however, by the local synthesis model,which posits local synthesis, reutilization and leak-age of labeled residues during subsequent turnoverof radiolabeled proteins.
7. Fink and Gainer (1980) labeled axoplasmic proteinsof the sciatic nerve with succinimidyl proprionate,which binds covalently to proteins, and the peakremained stationary for the ensuing 10 days. Again,this result indicates that there was no bulk translo-cation of axoplasmic proteins, a ®nding that is con-sistent with the local synthesis model, butinconsistent with the slow transport model.
8. Kanje et al. (1988) observed that inhibition of pro-tein synthesis distal to a crush decreased axonal re-
Fig. 22. In¯uence of metabolic half-life of the protein on velocity of
the wave, based on the mathematical local synthesis model. The
simulated curves (redrawn from Alvarez and Torres, 1985 with per-
mission of Academic Press) represent the distributions of two radio-
active proteins along an axon, each with a di�erent half-life (t1/2).
The radioactivity (ordinate) is expressed as a percent of the peak
radioactivity for each protein at day 20 post-pulse. Note that the
shorter the half-life of the protein, the faster is the forward displace-
ment of the peak.
Fig. 23. Modeling of slow transport in gold®sh optic nerve axons.
Gold®sh, experimental results from the gold®sh, redrawn from Graf-
stein et al. (1981) with permission of Springer-Verlag. In the latter
study, the eye was injected with labeled amino acids at time 0, and
curves show the percent distribution of radioactivity along the optic
nerve at 2 and 4 days post-injection. Because no moving peak was
observed, the slopes of the curves was taken as an index of trans-
port. Model: simulation using the mathematical model shown in Fig.
1. The net di�usional exchange between the ®rst axon segment and
the perikaryon (see Fig. 3) was cancelled out during the post-pulse
period (redrawn from Alvarez and Torres, 1985). This subtle modi®-
cation generates simulated curves that are similar to the experimental
curves that served to provide a transport index, notwithstanding the
fact that the proteins are stationary in the model.
Fig. 24. Postulated determinants of slow axonal transport (see text),
(®gure and legend from Grafstein and Alpert, 1982 with permission
of Springer-Verlag). As long as the cytoskeleton (paralleled lined seg-
ments) remains attached to its site of synthesis and assembly in the
cell body (rectangle at left), slow transport proceeds (arrow) only if
newly synthesized elements (double lines) are inserted in the cell
body. However, the velocity of transport is regulated by some mech-
anism in the axon or its terminals (hand at right).
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6238

generation by 70%, and proposed that the e�ectupon axons was mediated by cells resident in thedistal stump. Gaete et al. (1998) destroyed cells dis-tal to the crush by repeated freezing, which did nota�ect elongation of regenerating axons. Nonethe-less, when CXM was administered to the stumpregion, previously rendered acellular, the ingrowthof axons was reduced by 58%. In contrast, whenCXM was administered 1 mm central to the crush,axons regenerated normally. Interpretation of theseresults based on local synthesis is rather straightforward and predictable since the only possible tar-get for CXM in the tissue lacking cells is transla-tional machinery in growing sprouts. By contrast,the slow transport model requires ad hoc assump-tions to account for these observations.
5.3. Proposed experiment to test slow transport andlocal synthesis models
Both models deal with the maintenance of axoplasm,but their basic tenets predict mutually exclusive out-comes that can be tested experimentally. For example,if methionine and alanine are supplied to perikarya,the slow transport model predicts a constant ratiobetween amino acids in any given protein (e.g., b-tubu-lin) at the advancing margin of the peak of radioac-tivity because all molecules of the protein are assumedto have been synthesized in the perikaryon at the sametime. The local synthesis model, in contrast, predicts aprogressive increase in the methionine:alanine ratio,inasmuch as the alanine residue will be reutilized to alesser extent owing to metabolic conversion in the sol-uble pool. This type of experiment would distinguishwhich one of the two models is valid.
In summary, the concept of long range slow trans-port in axons as a mechanism for the maintenance ofaxoplasm was a logical outcome of assuming thataxons lacked an endogenous protein synthesizing ma-chinery, and rationalizing the proximo-distal pro-gression of waves of radioactive proteins originatingfrom the perikaryon. The dismissal or disregard of evi-dence of protein synthesis and turnover aside, themajor ¯aw of the slow transport model is its relianceon a single mechanism to explain axoplasmic renewal,which necessitates invoking additional, ad hoc assump-tions in order to account for experimental ®ndings thatare inconsistent with its basic premise. As a result, theslow transport model creates a biologically unrealisticprototype of the axon, and fails as a useful paradigm.
On the other hand, the local synthesis model (i) issupported by a substantial body of evidence, (ii)reveals that the waves of protein radioactivity may beobserved while proteins remain stationary, and (iii)provides an understanding of how the axon can be
maintained by mechanisms consonant with generalbiological principles of metabolic turnover. Thus, thelocal synthesis model has heuristic value, and canaccount for a wide range of phenomena. Explanatorycapabilities of the two models will be further comparedand contrasted below.
6. Dynamic anatomy of the axon and role of Schwanncells
Mature axons have a morphology that remainsessentially constant throughout life. In vertebrates,perturbations such as nerve section are followed bydestruction of a signi®cant portion of the axon, whichresults in a response that may, or may not restore theoriginal anatomy. This structural stability has engen-dered the idea that the axon is a passive appendage ofthe neuron, with either no, or minimal autonomy. Forexample, because the perikaryon is considered theexclusive source of axoplasmic proteins, the size of theaxon is assumed to be a consequence of a moving col-umn of axoplasm (Lasek et al., 1983), and that thissupply of proteins from the cell body is a preconditionfor regeneration of severed axons (cf. Rotshenker,1988; Bisby et al., 1995). It is possible to show, how-ever, that the axon has some degree of independencefrom the perikaryon. In the latter view, the constancyof the ®ber size is a result of a balance of conditionsthat regulate axonal structure and phenotype at a locallevel; i.e., local factors are assumed to e�ect changes inthe axon that are not mediated by the perikaryon.Data will be presented below in support of a localregulation with respect to the content of microtubules,axon caliber, and the ability of axons to form andextend sprouts anywhere along their length.
6.1. Axonal microtubules
6.1.1. Relation to axon caliberNeuro®laments and microtubules are the major
structural elements of the axonal cytoskeleton. Micro-tubules predominate in small caliber unmyelinatedaxons, while neuro®laments predominate in myelinatedaxons. There is a tightly linked relationship betweenmicrotubule content and axon size/caliber. In periph-eral nerves, while microtubules of myelinated andunmyelinated axons increase with size, their densitydecreases (Friede and Samorajski, 1970; Smith, 1973;Fadic et al., 1985; Pannese et al., 1984a); i.e., microtu-bule content increases at a rate that is slower than thatof axon size. Along the trajectory of myelinated andunmyelinated ®bers, microtubule density is constant(Alvarez and Zarour, 1983; Fadic et al., 1985). Forequal calibers, sensory, motor, and sympathetic ®bershave similar microtubule densities (Alvarez et al.,
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 39

1982; Fadic et al., 1985; Fadic and Alvarez, 1986). Themicrotubule density of 3 mm myelinated axons is 020microtubules/mm2 in laboratory animals (Fadic et al.,1985; Saitua and Alvarez, 1988), a value that is com-mon throughout vertebrates from ®sh to mammals,including man and cow (Vergara et al., 1991). Duringdevelopment and regeneration of nerves, the caliber ofthe axon increases, and microtubules do as well, result-ing in the usual correlation between caliber and density(Espejo and Alvarez, 1986; Fau ndez and Alvarez,1986). During severe undernutrition, caliber andmicrotubules of axons decrease but the correlationbetween them remains unaltered (Fau ndez et al.,1989). In peripheral nerves of frog, lizard, rat and®nch, the microtubule content is similar in a restrictedcaliber range, where unmyelinated and myelinated®bers overlap (Fau ndez and Alvarez, 1986; Pannese etal., 1988; Vergara et al., 1991).
In summary, axonal microtubule content correlateswith axon caliber, irrespective of whether the axon ismyelinated or unmyelinated, and remains remarkablyuna�ected by maturation, or adverse nutritional state.When axon caliber changes, microtubule content variesaccordingly. Nonetheless, this correlation may deviateunder particular circumstances. For example, the elec-trical activity of an axon increases the microtubulecontent (see below), suggesting that the correlationbetween caliber and microtubules is regulated dynami-cally.
6.1.2. Regional variationsThe microtubule content of stem axons is uniform;
however, there may be an abrupt change at particularpoints in the course of axons. For example, axonsemerging from retinal ganglion cells are unmyelinatedup to the lamina cribosa, and become myelinated there-after in the optic nerve. Upon myelination, the cross-sectional area of these axons increases by 59%, andthe microtubule content by 36%; however, the corre-lation between microtubule density and caliber is thesame in both domains (Herna ndez et al., 1989). Theterminal arborization is another example of abruptchange of the microtubule content. In motor axons ofthe rat, the sum total of microtubules in the terminalarborization is ten-fold greater than that of the stemaxon (Zenker and Hohberg, 1973). These regional vari-ations of the axonal cytoskeleton call for an expla-nation (see below).
6.1.3. Role in axonsThe inverse correlation between microtubules density
and axon caliber has been conserved throughout theevolution of vertebrates, and this relationship is likelyto signify a speci®c function that is conserved as well.While the cross-sectional area of myelinated axonsseems to depend on neuro®laments (Muma and Ho�-
man, 1993), axonal microtubules serve intracellulartransport functions, inasmuch as bidirectional microtu-bule based motors translocate organelles. In axons,microtubules support the rapid anterograde transportof vesicles targeted to presynaptic terminals and tointervening stem axon to a lesser extent (Droz et al.,1973), while also supporting rapid retrograde transporttargeted for the perikaryon. At face value, axonalmicrotubule content could be speci®ed by a transportload.
Within a given individual or between species, theamounts of axoplasm of stem axons of equal calibervary considerably owing to their di�erent lengths. Forexample, the axoplasm of a 3 mm axon of the sciaticnerve of cow may be 100-fold greater than that of thecorresponding axon of a lizard; nonetheless, the micro-tubule density is the same in both axons (Vergara etal., 1991). The terminal arborization varies also widely.Motor axons of extraocular muscles innervate a few®bers, while those innervating the diaphragm or anti-gravity muscles may innervate several hundred ®bers;however, the microtubule density is the same if thecaliber of the axon is the same (Alvarez and Zarour,1983). Thus, axonal microtubules correlate neitherwith the axoplasmic mass of the stem axon, nor withthe size of the synaptic ®eld. As a consequence, axonalmicrotubules do not seem to be governed by transportdemand related to the size of target structures, since,from a standpoint of satisfying organelle transportrequirements, axonal microtubules would seem to beoverly abundant. The inverse correlation between axonsize and microtubule density suggests that an architec-tural constraint is the primary determinant of axonalmicrotubules, albeit unknown at the present time.Nonetheless, microtubule density is a parameter thatcan be regulated by factors intrinsic, as well as extrin-sic to the axon (see below).
6.1.4. Environmental regulationThe constant correlation of axonal microtubules dis-
cussed above deviates signi®cantly in axons of ventraland dorsal roots. Motor axons are continuous fromspinal cord to neuromuscular junctions. For a givencaliber, the microtubule density of motor axons is thesame in the spinal cord and in peripheral nerve, but inthe ventral root, the density is one-half of that in otherregions (Saitua and Alvarez, 1989; Lo pez and Alvarez,1990). The anatomical organization of sensory axonsdi�ers from that of motor axons. Sensory perikaryagive rise to axons that bifurcate within the spinalganglion; one branch leaves the ganglion to enter thedorsal root, and thence into the spinal cord, while theother branch leaves the ganglion to enter the periph-eral nerve. In the dorsal root, central branches of sen-sory neurons contain one-half-to-one-third of themicrotubule density of peripheral axons of equal size
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6240

(Pannese et al., 1984b; Fadic et al., 1985; Saitua andAlvarez, 1989; Lo pez and Alvarez, 1990); however,upon entering the spinal cord, they acquire a microtu-bule density corresponding to that in peripheral nerveaxons (Saitua and Alvarez, 1989; Lo pez and Alvarez,1990).
The parallel manner in which the same variation inmicrotubule content occurs at the same particular ana-tomical locations in both sensory and motor axonssuggests that environmental cues regulate the axonalcytoskeleton. This inference was explored in exper-iments involving the vagal nerve of the cat (Serra andAlvarez, 1989), which are summarized in Fig. 25. Thenodose ganglion is composed of sensory neurons andlies in the vagus nerve of the neck. The centralbranches of these neurons constitute the bulk of theunmyelinated ®bers of the supranodosal nerve, whichis a peripheral nerve. These ®bers become radicular®bers after entering the skull and terminate in themedulla. In the supranodosal nerve and in the root,the central branches of vagal sensory axons exhibited amicrotubule density that was one-third that of the per-ipheral branches of equal size, and similar to the den-sity of dorsal root axons. Since the supranodosal nerveis a peripheral nerve from an anatomical standpoint,the low microtubule content of the ®bers seemed tocorrelate with their being central branches of sensoryneurons rather than with their peripheral location.However, when these central branches were allowed toregenerate along the hypoglossal nerve, they acquiredthe high microtubule density characteristic of periph-eral ®bers, but when regeneration occurred along theirnormal anatomical pathway, they retained their lowdensity. These regenerated axons neither made func-tional contact with the original target neurons in themedulla, nor with muscle ®bers after cross-suturing tothe hypoglossal nerve. This suggests that target cellsdo not play a role in specifying axonal microtubules.
On the whole, these observations indicate that themicrotubule content of axoplasm is modulated by cuesprovided by the local environment, probably involvingSchwann cells.
6.1.5. Regulation by Schwann cellIf Schwann cells regulate axonal microtubules, then
their selective destruction should a�ect the microtubulecontent. Local treatment of a nerve with actinomycinD (ActD) blocks transcriptional activity of residentcells, but spares axons since the only known axonalDNA is mitochondrial in location. One week afterlocal application of ActD to the nerve, tissue debrisand altered Schwann cells were present in the treatedregion, while axons exhibited an unusual abundance ofmicrotubules (Fig. 26). In unmyelinated ®bers, thetypical inverse relationship between axon caliber andmicrotubules was no longer valid, since the densityincreased throughout the spectrum of axon sizes tobecome the same for all axonal sizes. To reach thisapparent ceiling, axons with a cross sectional area of0.6±0.8 mm2 that had 30±40 microtubules mm2 at thebeginning of the experiment, increased their content to80±90 microtubules/mm2; i.e., there was a two-foldincrease (Fig. 26). The cytological appearance ofSchwann cells was normal 4 mm proximal and distalto the treated region, and so was the axonal microtu-bule content (Bustos et al., 1991). The local increase ofmicrotubule content was an indirect e�ect of ActDupon axons, presumably due to altered Schwann cellfunction. This coupled e�ect suggests that matureSchwann cells normally down-regulate the microtubulecontent of axons. If this is true, then the inverse re-lationship between microtubule density and axon sizewould suggest that the regulatory action of Schwanncells should be greater on larger axons than on thinnerones, and greater on axons of spinal roots than onthose of peripheral nerves.
6.1.6. Regulation by electrical activityAxonal microtubules can also be controlled by elec-
trical activity of the axon. For example, direct electri-cal stimulation of severed axons, or transsynapticstimulation of intact axons for two hours increasesaxonal microtubules by 24% (Alvarez, 1979; Alvarezand RamõÁ rez, 1979). Thus, dynamic instability ofmicrotubules and/or the tubulin-microtubule equili-brium at the steady state can be modulated on a localbasis by the frequency of action potentials. A re¯exhyperactivity of the sympathetic nerves for 2.5 daysraises microtubule content by 46% and axonal calibersby 19% (Alvarez et al., 1982). In motor nerves, hyper-activity produces changes in the anatomy of axons.When muscles are subjected to chronic overwork,axons involved grow larger (Edds, 1950), and axonschronically hyperactive as in spastic paralysis have a
Fig. 25. Diagrams, based on data of Serra and Alvarez (1989), depict
how microtubule density in axons of neurons in the nodose ganglion
can be a�ected by the local environment through which axons grow.
Left: nodose sensory perikaryon (circle) has peripheral and central
branches; the latter synapses in the medulla (M). Microtubule density
is high in peripheral branches and low in the central branches.
Middle: When central branches regenerate (dashed line) along their
normal pathway, they retains their low microtubule density. Right:
When central branches regenerate along the hypoglossal nerve (H)
that innervates the tongue (T), microtubule density in regenerated
axons is high (see text).
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 41

microtubule content twice that of a normal axon ofequal caliber (Vergara et al., 1992). In CNS neurons, asimilar pattern has been observed. In the hypotha-lamo-neurohypophyseal tract, hyperactivity caused byincreasing the sodium chloride intake resulted in largeraxons and an increased microtubular content (Grain-ger and Sloper, 1974; Dellmann et al., 1988). These®ndings suggest that the axonal cytoskeleton can bemodulated by the electrical activity of the axon invivo.
6.1.7. E�ects of inhibitors of protein synthesis or taxolAxonal microtubule content is also sensitive to in-
hibitors of cytoribosomal protein synthesis, and tostabilizers of microtubules. One week after adminis-tration of CXM or emetine to a 4-mm segment of ratsciatic nerve, microtubules were reversibly reduced byone-half, while adjacent segments remained una�ected(Bustos et al., 1991).
Taxol stabilizes microtubules and promotes tubulinpolymerization. Six hours after local administration oftaxol to a nerve, the number of microtubules wasslightly increased on visual inspection; however, themicrotubule content increased by 600% after one week(Fig. 26), while values were normal 4 mm central ordistal to the treated region (Bustos et al., 1991).
The observations discussed above indicate that cer-tain drugs can a�ect microtubule content in a delim-ited region of axons, such that it can be eitherdecreased or increased in a manner that re¯ects adegree of independence of axon from the cell body.Potential mechanisms o�ered by the local synthesis
and slow transport models for explaining the signi®-cant regional taxol-induced increase in microtubulecontent will be considered and contrasted below. If thedynamic cytoarchitecture of axoplasm is governed byan intrinsic program, and microtubule content is oneof its parameters, it follows that the program can bemodulated by environmental cues. It will be seen fromthe discussion below, however, that at least this par-ticular parameter of cytoarchitecture probably dependson local regulation of protein synthesis.
6.1.8. Slow transport model explanation of local changesTo explain the increase, or the decrease of the axo-
nal microtubule content, the slow transport model canonly posit changes in velocity of transport. Theincrease of microtubules by 600% one week after ad-ministration of taxol to rat sural nerves will beaddressed ®rst.
The slow transport model would predict that thesegment treated with taxol would receive a freshsupply of tubulin. In this model, tubulin is leaving theperikaryon constantly to maintain the steady state ofmicrotubules along the axon. The moving column ofmicrotubules/tubulin that had left the perikaryonweeks before the administration of taxol could havebeen arrested by the drug in the treated segment, andthe free subunits assembled into microtubules. The col-umn, progressing at 1±2 mm/day destined to replacedegraded subunits, could not convey enough tubulinto increase the microtubule content over a 4 mm spanof nerve by 600% in one week. Obviously, the arrestof transport would deprive downstream axonal seg-
Fig. 26. E�ects of actinomycin D and taxol on nerve ®bers. A, control unmyelinated ®ber bundle, in which axons contain neuro®laments and
microtubules, and are ensheathed by Schwann cell processes. B, an unmyelinated bundle after exposure to actinomycin D for 11 days. The axons
present an inordenately high density of microtubules, virtually no neuro®laments, and the Schwann cell has disappeared (remnants visible in
lower right corner). C, a myelinated ®ber after exposure to actinomycin D for 11 days. Axon sprouts are located between the myelin sheath and
basal lamina (arrowheads). Again, microtubules are very abundant, and there is a paucity of neuro®laments in the parent axon. Note that
although Schwann cell cytoplasm has been destroyed, the myelin sheath is still well organized and compact. A lipid droplet is located in extra-
cellular space (arrow). D, an unmyelinated ®ber bundle in a nerve segment that was exposed to taxol for 7 days. The axons contain a high den-
sity of microtubules and virtually no neuro®laments, and there is direct contact between axons. In contrast to the e�ects of actinomycin D,
however, the integrity of Schwann cell cytoplasm (arrowheads) is maintained with taxol treatment. Calibration, 0.5 mm for all panels.
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6242

ments of their normal supply of tubulin and would beunable to maintain their microtubule content in asteady state. This prediction was not realized sincemicrotubule content in segments proximal and distalto the treated segment was normal.
A similar reasoning applies to the increase in micro-tubules induced by ActD treatment, except thatSchwann cells, which would be the most likely targetof the drug, would have to regulate slow transport.The local reduction of microtubules in response toCXM or emetine would have to be attributed to anacceleration in slow transport velocity in the treatednerve segment. Alternatively, these inhibitors of pro-tein synthesis would have to stimulate a local degra-dation of tubulin; however, this would predict anadditional reduction in microtubule content distal tothe treated segment, which was not observed. All con-sidered, the slow transport model cannot o�er astraightforward explanation of the local e�ect of taxol,ActD, emetine, and CXM upon axonal microtubules.
Finally, with regard to the increase of axonal micro-tubules at speci®c regions along some nerve pathways,such as in the case of optic nerve axons (see above),the slow transport model must postulate a furtherslowing in the rate of slow transport velocity (Millerand Samuels, 1997) in the relevant segments.
6.1.9. Local synthesis model explanation of localchanges
In contrast to the explanation o�ered by the slowtransport model above, local synthesis can account forregional variations of axoplasm through an uncompli-cated mechanism in which axonal protein synthesiswould be regulated locally by external cues. Forexample, while two hours of steady ®ring of the axonresults in an increase of microtubule content by 25%that can be explained by a reduction in dynamicinstability, chronic hyperactivity doubles the microtu-bule content. The latter requires more protein thanthat existing as soluble dimers (see below). It also mer-its noting in this context that steady electrical stimu-lation enhances the rate of incorporation of aminoacids into axoplasmic proteins in the M-cell axon ofthe gold®sh (EugenõÂ n and Alvarez, 1995).
The local synthesis model does not invoke longrange vectorial displacement of proteins. Moreover, itassumes that inhibitors of protein synthesis operatelocally while they are present, and that synthesis willresume after their removal. Con®rming these expec-tations, inhibition of protein synthesis for a week,which reduced axonal microtubules by half in a revers-ible manner, had no e�ect on the microtubule contentin adjacent untreated segments. The reduction wasconsistent with an estimated tubulin half-life of 10days in brain, axons, and terminals (Karlsson and
SjoÈ strand, 1971b; Forgue and Dahl, 1978; Nixon,1980).
To explain the taxol induced increase in microtubulecontent, the mere promotion of assembly by this drugas a mechanism does not appear to be su�cient,owing to the latency of several days, and the inordi-nately large pool of soluble subunits required. Indeed,inasmuch as 10 microtubules/mm2 represents 3 mg ofassembled tubulin per ml of axoplasm (Fadic et al.,1985), an average unmyelinated ®ber in rat peripheralnerves has over 50 microtubules/mm2 (Fadic et al.,1985), which represents 15 mg/ml of assembled tubu-lin. Therefore, a 600% increase of microtubules wouldrequire 90 mg of soluble tubulin per ml of axoplasm,or 10.5 g% of total tubulin. Such a concentration isunprecedented when compared to only 0.8 mg per mlof cytoplasm, which is the total tubulin in mouse livercells based on the data of Pipeleers et al. (1977). In thesquid giant axon, assembled tubulin is 4.1 mg per mlof axoplasm, representing 14 microtubules/mm2, andthere is an additional 1.5 mg/ml of soluble tubulin(Morris and Lasek, 1984), which yields a maximalmicrotubule reserve of 26%. The accumulation ofmicrotubules in axons in response to taxol seems toolarge to be explained other than by a modi®ed metab-olism of tubulin, in which a combination of anincreased tubulin half-life, and/or an increased rate oftubulin synthesis could produce the observed elevatedmicrotubule content.
Thus, assuming that taxol shifts the monomer-poly-mer equilibrium in favor of the polymer, and that thepolymer does not undergo proteolysis, the microtubulecontent would rise by 75% in seven days. This wouldinclude 25%, corresponding to the original solublepool, and an additional 50% that would be syn-thesized de novo in order to satisfy the normal steadystate. This is far below the observed elevation of600%; however, an indirect e�ect of taxol would be toreduce tubulin monomer concentration, and therebyincrease mRNA stability (Cleveland et al., 1981; Yenet al., 1988; see Section 3.1.2). Thus, an increased over-all abundance of tubulin mRNAs would increase therate of tubulin synthesis.
Evidence has been reviewed above indicating thatenvironmental cues, probably originating fromSchwann cells, down regulate axonal microtubules.This view is supported by metabolic studies. Forexample, blunt dissection of a nerve segment was su�-cient to induce a local, delayed increase in the rate ofprotein synthesis in axons of the a�ected region, andthis type of metabolic stimulation has been interpretedas a `release' phenomenon, in which a negative regulat-ory pathway operating between Schwann/glial cellsand cognate axon is interrupted (Tobias and Koenig,1975b; Perrone Capano et al., 1999). Within the frame-work of the local synthesis model, and in accordance
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 43

with the experiments mentioned above, ActD treat-ment of a nerve segment would trigger a cascade ofe�ects in the a�ected region, including blockade oftranscription and loss of Schwann cells, which wouldcause a progressive disinhibition of translational ac-tivity in axons, an increase of tubulin synthesis and anaccummulation of microtubules.
6.2. Axon caliber
The caliber of the axon governs the velocity andsafety factor of the nerve impulse, which are vital forthe operation of the nervous system in processing in-formation. The mechanism by which axon caliber isspeci®ed remains poorly understood, notwithstandingthe several lines of evidence that indicate that the cali-ber is regulated locally, in addition to being controlledby the perikaryon.
6.2.1. Schwann cells and caliberThe microscopic anatomy of the nerve ®ber suggests
that the Schwann cell participates in the speci®cationof axon caliber, as exempli®ed by the fact that inter-nodes, which are myelinated are larger in diameterthan nodes of Ranvier. This is consistent with the hy-pothesis that Schwann cells regulate axon caliberlocally, as indicated by a number of documentedexamples, in which axon diameter increases duringmyelination (Pannese et al., 1988; Herna ndez et al.,1989; Windebank et al., 1985). Recently, myelin-associ-ated glycoprotein has been proposed to modulate theaxonal caliber (Yin et al., 1998). A complementary®nding is that axon caliber is reduced in intercalatedunmyelinated segments of myelinated peripheral ®bers(Yokota, 1984). While such observations indicate thatSchwann cells modulate axon diameter, it is the axonwhich induces Schwann cells to myelinate; however,the latter half of the regulatory loop is beyond thescope of this article.
6.2.2. Trembler mouse as a case studyThere are certain mutations that a�ect nerve ®bers
in mice, and such mutant strains have been used toisolate variables in the reciprocal intercellular regu-lation of axon and Schwann cell. A nerve segment of adonor strain grafted into a nerve of a recipient strainresults in a chimeric regenerated nerve, in which donorSchwann cells ensheath axons of the recipient strain.Data from studies on the Trembler (T) mouse will bediscussed.
The myelin of peripheral nerves in the T mouse isinsu�cient or nonexistent, and the caliber of axons issubnormal (Low, 1976). When a T nerve segment isgrafted into a nerve of a normal strain (N) to form achimeric regenerated nerve, consecutive longitudinalsegments of normal axon are ensheathed respectively
by N, T and N Schwann cells. Such a bioengineered®ber has three characteristics. First, central to the Tgraft, N axon and myelin are normal. Second, in thegraft, T Schwann cells form a thin myelin sheath cor-responding to its original phenotype; however, Naxons acquire the phenotype of the donnor strain bybeing reduced in diameter. Third, distal to the graft, Naxons increase in diameter again when they become as-sociated with N Schwann cells (Aguayo et al., 1977).
In the restricted segment in which the N-axon is sur-rounded by T Schwann cells, the neuro®lament densityis two-fold greater (De Waegh et al., 1992); however,due to the reduced cross-sectional area (Aguayo et al.,1977), total neuro®laments are half of those in the nor-mal axons. Therefore, the axonal caliber and its neuro-®lament content are regulated locally by Schwanncells.
6.2.3. Axon caliber from perspectives of slow transportand local synthesis models
Caliber should be determined by a rather complexset of factors that place constraints on various func-tional and structural properties of the axon, includingmechanical stability, a speci®c array of ion channels,transport systems, receptors, etc. The slow transportmodel regards axons as conduits, the cross sectionalarea of which is determined by cytoskeletal com-ponents moving within it (Lasek et al., 1983). In thechimeric ®ber segment, the reduction of neuro®lamentsin the T graft would be explained by an accelerationof slow transport (cf. Miller and Samuels, 1997). Con-trary to expectation, in normal axons surrounded by TSchwann cells, the slow wave follows the pattern ofintact T axons (De Waegh and Brady, 1991), whichfor neuro®lament polypeptides is 17% below normal(De Waegh and Brady, 1990). Thus, experimentalresults clearly reveal that axons do not satisfy predic-tions of the slow transport model. An alternative ex-planation would be that neuro®laments disassembleunder the in¯uence of T Schwann cells, move asmonomers, and reassemble at the end of the graft.
As already noted above, the local synthesis modelreadily accommodates regulation of axonal cytoarchi-tecture by Schwann cells. Thus, axon diameter andcontent of microtubules increase upon myelination,and mechanical perturbation can remove a steady statesource of translational inhibition, which would resultin an augmented rate of axonal protein synthesis (seeabove). Through the transcellular mechanism, chimericT Schwann cells would down regulate the cytoskeletalcontent of cognate axonal segments to a level lowerthan normal. As a result, the cross-sectional area ofthe axon would decrease, and the rate of the slowradioactive wave would be a�ected accordingly.
In this context, it is also worth noting the obser-vations described previously relating to the larger cali-
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6244

bers and increased microtubule content produced bychronic overwork of axons (Edds, 1950; Alvarez et al.,1982; Vergara et al., 1992), as well as those related tothe increased rate of incorporation of amino acids intothe axoplasm induced by hyperactivity (EugenõÂ n andAlvarez, 1995). Thus, in the local synthesis model,these ®ndings are complementary, and suggest that theaxon is modulated by the bioelectrical activity in ad-dition to environmental cues.
6.3. Sprouting of axons in vertebrates
In this section the ability of axons to extend sproutsanywhere along their course will be emphasized as anexample of local autonomy. This capability, in turn,can explain a number of observations that relate tonerve regeneration, inasmuch as the latter is regardedas a speci®c case of this ubiquitous property of axons.
6.3.1. Altered Schwann cellsAs early as 1904, Langley and Anderson found that
axons of a severed nerve anastomosed to an intactnerve did not invade the recipient nerve but did soafter axonal degeneration (Langley and Anderson,1904). On the other hand, a pre-degenerated nervegraft enhanced regeneration by shortening the initialdelay (Danielsen et al., 1994). The same pattern wasobserved under culture conditions in vitro, in whichgrowing axons associated with activated proliferatingSchwann cells, but failed to do so when Schwann cellswere quiescent, or di�erentiated (Baehr and Bunge,1989; Bedi et al., 1992; Agius and Cochard, 1998; cf.Shen et al., 1998).
Schwann cells may be selectively a�ected by variousagents that do not seem to act on axons directly. Onesuch example is the sprouting of myelinated andunmyelinated axons induced by local application ofActD to a segment of an intact nerve (Fig. 26) (Bustoset al., 1991). To explain this phenomenon, it was pro-posed that a sprouting potential or program distribu-ted throughout the length of axons was maintained ina latent state by mature Schwann cells. Because ActDinhibits transcriptional activity, Schwann cells will beselectively a�ected, resulting in a derepression of thesprouting program in the cognate axons.
Another example is when nerves are treated withlysophosphatidyl choline, a phospholipid with deter-gent-like properties, myelin is disrupted, Schwann cellsare altered, and axons sprout in the a�ected region(Gregson and Hall, 1973). These e�ects further high-light the apparent role of Schwann cells in repressingthe intrinsic sprouting program in axons.
Axons do not regenerate ordinarily in the CNS, butdo so when peripheral nerve is grafted in place(Aguayo et al., 1990). Oligodendrocytes are able tosuppress growth of CNS and PNS axons in vivo and
in culture (Schwab and Thoenen, 1985; Giftochristosand David, 1988; Schwab, 1996). Thus, both PNS andCNS ensheaththing cells regulate the ability of axonsto grow. In this regard, a major functional di�erencebetween them is that the Schwann cell becomes per-missive when it undergoes dedi�erentiation, while theoligodendrocyte does not.
6.3.2. Protease inhibitors and acetylcholinesteraseProtease inhibitors can elicit a sprouting response of
intact stem axons. Leupeptin, the amyloid precursorprotein (APP) with the Kunitz insert, or aprotininadministered to a short segment of an intact nerveinduces demyelination, Schwann cell proliferation, andsprouting of myelinated and unmyelinated axons.Axons remain intact, and associated with these alteredSchwann cells (Alvarez et al., 1992, 1995; Moreno etal., 1996). Nerve section induces similar e�ects, in thatmyelin is destroyed, Schwann cells proliferate (Bradleyand Asbury, 1970), and surviving axons sprout.Although axons induce Schwann cells to remain in anonproliferative state and myelinate, exogenous appli-cation of a protease inhibitor releases the Schwann cellfrom axonal control, and, reciprocally, releases theintrinsic sprouting program of axons from Schwanncell control. Because neither aprotinin nor APP perme-ate cells, the reciprocal regulation between axon andSchwann cell may involve an extracellular protease(see Fig. 1).
Several physiological proteases and their inhibitorshave also been shown to a�ect the microenvironmentof axons and Schwann cells. Thrombin, a protease inthe clotting cascade, binds to a cell surface receptorand inhibits neurite elongation, and glial derivednexin/protease nexin 1, which inhibits thrombin, is apotent promotor of neurite growth (Monard, 1988,1993). After peripheral nerve injury, prothrombin andprotease nexin-1 are altered (Meier et al., 1989; Smir-nova et al., 1996), as well as the expression of col-lagenase, stromelysin and the tissue inhibitor ofmetalloproteinases (La Fleur et al., 1996). Schwanncells secrete stromelysin, a member of the matrixmetalloproteinases family that cleaves ®bronectin, andthe released fragment inhibits the proliferation ofSchwann cells (Muir and Manthorpe, 1992). Lamininhas neurite promoting activity (Kuhn et al., 1998)which is neutralized by a chondroitin sulfate proteogly-can derived from Schwann cells (Zuo et al., 1998b).The activity is restored by metalloproteinase 2, orchondroitinase (Zuo et al., 1998a). PC12 cells thatoverexpress tissue plasminogen activator extend neur-ites to a greater extent than controls (Pittman andDiBenedetto, 1995). Thus, there is growing evidencethat an extracellular system of proteases, antiproteases,substrates and receptors are involved in the local regu-lation of axons and Schwann cells.
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 45

Acetylcholinesterase (AChE) also seems to play arole in the mutual regulation of axon and Schwanncell. The speci®c inhibitor of AChE, BW284c51,reduces neurite growth of dorsal root ganglion cells invitro (Dupree and Bigbee, 1994; Dupree and Bigbee,1996). In mid-brain dopaminergic neurons, AChEenhances neurite outgrowth, while BW284c51 reducescell body size, extent of neurite outgrowth, and num-ber of neurons (Jones et al., 1995). In chick neurons,AChE stimulates (Small et al., 1995), and BW284c51decreases neurite outgrowth (Layer et al., 1993). InAplysia neurons, AChE promotes, and BW284c51impairs neurite outgrowth (Srivatsan and Peretz,1997). AChE also has deleterious e�ects. The survivalof Neuro 2a and PC12 cell lines is shortened by AChE(Caldero n et al., 1998), and the toxic action of theamyloid b-peptide upon PC12 cells, and upon chickretinal neurons is potentiated (Alvarez et al., 1998).
Organophosphorus compounds that irreversiblyinhibit AChE and other esterases in vivo induce asprouting response of motor terminals (Kawabuchi etal., 1988). The G4 isoform of AChE is present in per-ipheral axons and interfaces with the extracellularspace (Iurato et al., 1974; Rotundo and Carbonetto,1987; Ferna ndez et al., 1996). Local inhibition ofAChE in the nerve trunk is followed within a few daysby demyelination, proliferation of Schwann cells andsprouting of axons (Keymer et al., 1999), as if mutualregulatory mechanisms between axon and Schwanncells were abrogated. Thus, AChE in the nerve trunkmay play a role in trophic regulation (see Fig. 1). Themechanism underlying this action of AChE does notseem to be related to hydrolysis of acetylcholine, andmay even be an activity that is nonenzymatic (Massou-lie et al., 1993). Other esterases may also participate introphic regulation in other regions of the nervous sys-tem since organophosphates induce a delayed neuropa-thy by inhibiting a putative ``neuropathy targetesterase'' in brain, the physiological substrate of whichis unknown (Lotti, 1991).
6.3.3. Regeneration and the sprouting programIn normal nerves, the regulatory loop between axon
and Schwann cell should be in e�ect generally, and thestability and constancy of the nerve must be the resultof a dynamically balanced regulation. The balance isperturbed by nerve section, resulting in the regenerat-ing response. Regeneration is believed to begin with adelay of 1±2 days after injury (Bisby and Keen, 1985;McQuarrie and Jacob, 1991), generally attributed tothe time required by the perikaryon to initiate theregenerative response (cf. Bisby et al., 1995). In themodel presented here, regeneration of severed axons isdelayed until Schwann cells distal to the injury acquirethe permissive phenotype. This was con®rmed by thefollowing experiment. Administration of aprotinin for
two days prior to a crush reduced the delay of regener-ation to two hours, while the velocity of elongationdid not change. In contrast, a crush remote from theconditioned segment, or administration of aprotinin atthe time of the crush resulted in a normal latency ofregeneration (Tapia et al., 1995). The brief latentperiod observed in the conditioned segment is prob-ably insu�cient for a retrograde signal to travel to theperikaryon, and another signal to return from the peri-karyon to initiate a sprouting response; therefore, thetissue distal, but adjacent to severed axons must be re-sponsible for the delay rather than the perikaryon. Theinhibition of proteases does not appear to a�ect axonalprocesses related to outgrowth, because the rate ofelongation is normal. It is noteworthy that condition-ing with aprotinin requires two days to eliminate thedelay, which is consistent with the time required bySchwann cells to acquire spontaneously a permissivephenotype after nerve section.
A similar pattern was observed when the role ofAChE was studied in regenerating nerves of the rat.Inhibition of AChE in the distal stump withBW284c51, edrophonium or propidium, for three daysstarting at the time of the crush, did not a�ect axonelongation. However, when the anticholinesterase wasapplied to the presumptive stump region two daysprior to the crush, elongation increased by 33%. Con-versely, administration of the bovine G4 form ofAChE to the distal stump after the crush decreasedelongation by 36%. Administration of vehicle, iso-OMPA (an inhibitor of pseudocholinesterases), orheated AChE, was without e�ect (Keymer et al.,1999). The fact that inhibition of AChE for two daysis required to increase axon elongation suggests thatthe action of AChE entails an unknown regulatoryprocess that has a slow rate of decay. These obser-vations highlight a role of AChE in this merotrophicregulation of peripheral nerves (Fig. 1; see below).
6.3.4. Local synthesis and the sprouting programSprouting is a necessary antecedent to growth, and
the spontaneous formation of sprouts in response tonerve lesioning in vivo appears to be accompanied byan increased rate of local protein synthesis. Thus, inresponse to peripheral nerve lesion, there is a morethan 10-fold increase in the rate of amino acid incor-poration into axonal proteins that appears suddenlyafter a latency of 12±15 hours (Tobias and Koenig,1975b). Comparable data have recently been reportedfor dissected squid nerves (Perrone Capano et al.,1999). The rate is maximal proximal to the lesion, anddiminishes with distance from the lesion site. Theonset after a latent period may re¯ect the timerequired for the decay of a negative regulation that theSchwann cell normally exerts on cognate axons (seeabove). Thus, the proximo-distal gradient of an
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6246

increasing rate of protein synthesis, as the lesion site isapproached, may re¯ect a corresponding increasingperturbation of the normal Schwann cell-axon re-lationship (see Section 6.1.9). That local protein syn-thesis is important for normal regeneration has beenshown by experiments in which local inhibition of pro-tein synthesis in a regenerating nerve reduces axongrowth by half or more (Kanje et al., 1988; Edbladh etal., 1994; Gaete et al., 1998; cf. Meyerson et al., 1994).
The formation of sprouts and the elongation pro-cess entails highly dynamic membrane activity. Indeveloping axons (Waxman, 1968), as in sproutsinduced by inhibitors of AChE, or by cerebrospinal¯uid from patients with tropical spastic paraparesis(Keymer et al., 1999; Nien et al., 1998), the axo-lemma exhibits an unusual abundance of coatedand pinocytic pits and vesicles. In preterminal axonsa dynamic interchange of components is likely tooccur between the axolemma and a subaxolemmalsmooth endoplasmic reticulum (SER), inasmuch asa branched tubular SER extends along the lengthof axons (Droz et al., 1975; Ellisman and Lindsey,1983) and periodically forms a spatially delimitedsubsurface network of tubules, called ``subaxolemmalplates'' (Droz et al., 1975). Moreover, the stacks oflumenless disks contained in growth cones of chickoptic axons have been proposed to be a reservoirfor expanding growth cone membrane (Cheng andReese, 1987; see Section 6.1.3).
The elongation of axons requires a whole arrayof essential molecular components, most of whichare axoplasmic, but some of which may alsoinclude membrane lipid constituents, such as phos-pho- and sphingolipids, in addition to selected inte-gral membrane proteins. The extent to which thecomplex balance of membrane and protein com-ponents requisite for growth and extension ofbranches depends on a local source is unknown atpresent. There is evidence, however, which suggeststhat axons may synthesize at least some of thecomponents necessary to sustain growth. Forexample, choline is incorporated into axoplasm afterionophoretic injection into the M-cell axon of thegold®sh, (Alvarez and Chen, 1972b), and the syn-thesis of choline phosphoglycerides has been studiedin the giant ®ber system of the squid (Brunetti etal., 1979). Moreover, sympathetic axons have alsobeen shown to synthesize phosphatidylcholine, phos-phatidylserine, phosphatidylethanolamine, sphingo-myelin (Vance et al., 1991, 1994), and localinhibition of phosphatidylcholine synthesis in distalgrowing axons in culture reduces growth by half(Posse de Chaves et al., 1995a, 1995b).
Finally, the apparent presence of 7SL RNA andSRP54 components of the signal recognition particle inaxons (Titmus et al., 1998; see Section 3.3.3), is con-
sistent with ultrastructural evidence of ER-attachedribosomes reminescent of a rough ER (Davis et al.,1992; Pannese and Ledda, 1991; see also Fig. 9), andfurther suggests the possibility that selected integralmembrane proteins may be synthesized in the axon.Although the question has an important bearing onthe issue of local autonomy of the axon, the availableevidence reviewed and commented upon in previoussections indicates that the axon does have considerablelocal independence in responding to regional environ-mental signals. The sprouting program intrinsic to theaxon may be considered a speci®c case in point. Re-gional modi®cation of axonal phenotype properties ishere referred to as merotrophism (see below), and regu-lation of local synthesis of proteins can provide amechanism for mediating some degree of autonomyfrom the perikaryon.
6.3.5. The Wlds mutant mouse as a case studyThe Wlds mutant mouse, known formerly as C57B1/
Ola mouse, is a strain in which severed, decentralizednerves neither degenerate nor regenerate for severalweeks (Lunn et al., 1989; Brown et al., 1991; Glassand Gri�n, 1991; Chen and Bisby, 1993) despite thevigorous production of sprouts at the crush site(Brown et al., 1994). The mutation, localized onchromosome 4 and named Wlds for Wallerian degener-ation-slow (Lyon et al., 1993), is a consequence of atandem triplication (Coleman et al., 1998). The strainhas been used extensively to study the relationshipbetween Schwann cell and axon.
Chimeric sciatic nerves were made by transplantinga nerve segment of normal or Wlds mouse into theother strain, and the nerve was allowed to regenerate,resulting in mutant axons surrounded by normalSchwann cells and vice versa. The pattern of axonaldegeneration of these engineered ®bers correlated withthat of the host strain (Glass et al., 1993), suggestingthat the long survival is a property of axons ratherthan an axonal response to external signals. This wascon®rmed by the observation that severed neurites ofWlds neurons in culture survive over 6 days comparedto 12±16 hours for neurites from wild type neurons(Buckmaster et al., 1995). When the Wlds nerve is fro-zen and thawed, axoplasm is degraded (Tsao et al.,1994). The same occurs when calcium ionophores areused to elevate intracellular Ca2+, although the cal-pains appear to require somewhat higher levels ofCa2+ (Glass et al., 1994). The mutation in Wlds micedoes not a�ect the onset of chromatolysis, a classicalresponse of the perikaryon to nerve transection (Bisbyet al., 1995).
When a short span of Wlds nerve is crushed, regen-eration is initiated, but is then halted when regrowingaxons reach the undamaged region of the nerve(Brown et al., 1994), which indicates that local factors
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 47

derived from undegenerated axons or Schwann cellsarrest the ingrowth of regenerating axons. As pre-viously noted (see Section 6.1.5), ActD destroysSchwann cells, but spares axons. Wlds nerves regener-ate at a normal rate if the crush is made in a segmentconditioned with ActD, but do not do so if the crushis placed distal to the conditioned segment (Benavidesand Alvarez, 1998; Court and Alvarez, unpublishedobservations). Therefore, destruction of resident cellsof the conditioned segment is su�cient to restore thenormal phenotype, at least with regards to rate ofregrowth in a strain in which axons do not regeneratespontaneously. Finally, when a nerve in a Wlds mouseis cut at the proximal end of an extended crushedregion, the undamaged axons of the distal stumpdevelop growth cones and extend in a central direction(InÄ iguez and Alvarez, 1999).
Fig. 27 summarizes the observations in the Wldsmouse. The latter con®rm that axons are endowedwith a sprouting program that can be expressed evenin the absence of perikarya, and that the program isordinarily repressed by normal resident cells. There-fore, the capacity to develop growth cones andelongate is a general intrinsic property of axons.
The directionality of growing axons is governed bythe permissive nature of the substratum lying in theirpath. In the Wlds mutant, the sequence of eventswould presumably be as follows: the mutation preventsentry of Ca2+ into severed axons, and degenerationdoes not occur. Axons, in the severed distal stump in
turn, continue to maintain Schwann cells in a di�eren-tiated state, and the latter repress ingrowth of regener-ating axons at the injury site, closing the regulatoryloop. In this view, elongation of the axon is subjectedto stringent local control, and the perikaryon's rolewould seem to be incidental, in that it would be re-sponsible for specifying the intrinsic basic programthat would be expressed locally in response to environ-mental conditions.
In summary, the sprouting of axons and its localregulation appears as a general property of the axon-ensheathing cell unit. In this scenario, nerve regener-ation is a special case in point, in which the sproutingprogram is expressed. A proposed sequence of eventswould be as follows. Axons degenerate distal to aninjury. As a result, Schwann cells dedi�erentiate, andthereby lose the ability to repress the sprouting pro-gram; as a consequence, sprouts elongate at an increas-ing rate until reaching a maximal value after a fewdays (cf. Wyrwicka, 1950; SjoÈ berg and Kanje, 1990).Thereafter they navigate along a permissive substratumuntil external stop signals arrest further growth. Re-generation, which requires accretion of axoplasmicproteins, can be controlled by local environmentalcues. From a logistical standpoint, a local source ofaxonal proteins is a key feature of ®ne-tuning regrowthin response to local modulatory factors, since it o�ersadvantages over that of the remotely situated perikar-yon.
6.4. Sprouting of axons in invertebrates
The paradigm presented above for mammalianaxons has also been observed in invertebrates. In theleech, an axon separated from the cell body and fromits terminal arborization extends sprouts at both ends(Mason and Muller, 1982), and when the glial cell isdestroyed by intracellular injection of a protease, thedisensheathed axon sprouts (Elliot and Muller, 1983).Laminin, secreted by glial cells, appears to be instru-mental in this sprouting (Masuda-Nakagawa et al.,1993). In Aplysia, transected axons in contact withother neurons survive for two weeks, and during thisperiod extend neurites, which may form electrotonicgap junctions (Benbassat and Spira, 1994).
The cut axon requires a rise in axoplasmic Ca2+ toreseal (Spira et al., 1993). An elevation in Ca2+ pre-cedes the formation of growth cones in transectedaxons, and a local exposure of intact axons to ionomy-cin elevates Ca2+ concentration in axoplasm, and alsoleads to the formation of growth cones (Ziv and Spira,1997). A focal axoplasmic injection of a protease is fol-lowed by formation of the growth cone (Ziv andSpira, 1998). Therefore, the cascade triggering the in-itial stages of regrowth seems to involve localized pro-teolytic activity, which would suggest that the rise of
Fig. 27. Diagrams representing the e�ects of Schwann cells in nerves
of Wlds mice on axon regeneration. A a nerve crush (vertical line) is
not followed by regeneration (x), and the distal stump does not
degenerate for 3±4 weeks. B an extended region of nerve crush
(dashed line), is followed by regrowth (arrow) that halts as axons
reach the undamaged segment. C a nerve crush (vertical line) made
through a segment previously in®ltrated with actinomycin D, an in-
hibitor of transcription (cross-hatched segment), to damage selec-
tively the resident cells is followed by regeneration (arrow). D, if the
crush (vertical line) is made distal to the region damaged with actino-
mycin D (dashed segment), no regeneration ensues (x). E, decentra-
lized axons grow centralward when they lie adjacent to a damaged
segment (dashed line). The diagrams are based on studies by Lunn et
al. (1989) (A); Brown et al. (1994) (B); Benavides and Alvarez (1998)
(C); Court and Alvarez (unpublished) (D); and Alvarez and InÄ iguez
(1999) (E).
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6248

Ca2+ during transection of the axon may be instru-mental in activating axoplasmic proteolytic activity.These ®ndings indicate that the intrinsic sprouting pro-gram, which is postulated to be repressed in mamma-lian axons by associated ensheathing cells, can also beextended to invertebrates. In this context, a reasonableconjecture is that the adaxonal cell may control thesprouting program by regulating the Ca2+ entry intothe intact axon.
6.5. Plasticity of the terminal arborization
The terminal arborization is generally recognized asa highly plastic region of axons. Even hard wired con-nections such as neuromuscular synapses (Barker andIp, 1966; Tu�ery, 1971; Pachter and Eberstein, 1984;Hill et al., 1991), or synapses of sympathetic ganglia(Purves et al., 1987) are being remodeled constantly.Several observations are noted here to emphasize thepossible connection between changes in the anatomicalcon®guration of terminal arbors and endogenous pro-tein synthesis described previously.
Partial denervation of peripheral ®elds induce sur-viving axons to sprout either from terminals or pre-terminal nodes of Ranvier that invade the denervatedzone, and is referred to as terminal or collateralsprouting, respectively. This is a general property ofsensory, motor and autonomic peripheral axons (Bisbyet al., 1996; Chung et al., 1996; Winkler et al., 1997).CNS axons present a similar pattern (Raisman, 1968).There are a few issues that merit highlighting here.First, there is the issue of the role that neighboring tis-sue plays. Partial denervation does not directly a�ectthe remaining intact axons because the stimulus thattriggers the sprouting response originates from dener-vated target cells, and Schwann cells in ®bers fromwhich axons have degenerated (i.e., ``widow'' Schwanncells). The latter Schwann cells proliferate, and expressa di�erent set of genes than those of di�erentiated,quiescent cells (Ide, 1996).
In the frog, a short nerve segment implanted in anormally innervated muscle induces collateral andterminal sprouting of neighboring axons. The e�ectrequires viable cells, but not actual contact betweentransplanted cells and responding axons (Ku�er, 1986;DõÂ az and Pecot-Dechavassine, 1990). In the rat, afterpartial denervation, or implant of a nerve segment intoa muscle, widow Schwann cells proliferate, migrate,and extend processes, which, upon close appositionwith an intact axon from an innervating nerve, inducethe latter to sprout (Son and Thompson, 1995a; Sonand Thompson, 1995b). Therefore, the minimum sys-tem that induces remodeling consists of intact terminalarbors and denervated resident cells of a nerve in thevicinity of the arbor. This suggests that axons respondto Schwann cells, and require local regulatory signals.
There is, in addition, the issue of whether speci®cparticipation of the neuronal perikaryon is requiredfor the terminal arbor to extend sprouts and grow. Itseems unlikely that the perikaryon can specify theexact locus within the terminal arborization fromwhich sprouts will develop. Although the perikaryonmay be involved in later stages of growth, it does notappear to be necessary for the initial formation ofsprouts in frog axons (Carlsen et al., 1982), and botuli-num toxin induces axonal sprouting of terminals, butthe cell body reaction is conspicuously absent (Pamph-lett, 1988; see also Section 7).
Local administration of colchicine to the phrenicnerve is followed by sprouting of motor terminals(Riley and Fahlman, 1985; Fahlman and Riley, 1986).Since colchicine arrests microtubule based transport,the perikaryon may not participate actively in thesprouting response of terminals, but may down regu-late their development through the fast transport,directly or by way of an intervening cell. Motor axonsof the frog survive for 10 days before degenerating,and extend sprouts before they undergo involution anddegeneration (Rotshenker, 1981, 1982), and in Aplysia,isolated axons grow and even form electrotonic junc-tions de novo with neurons (Benbassat and Spira,1994). Finally, as already noted, growth cones developwhen calcium, or a protease is injected locally into theaxon (see Section 6.4). Such observations substantiatethe conclusion that the perikaryon plays no speci®crole in the sprouting response, and that the local ma-chinery is su�cient to serve the local requirements forremodeling of terminals.
As discussed above for the preterminal axon, asimilar argument applies to nerve terminals; namely,that phenotype characteristics of terminals may bemodulated by local signals impinging upon endogen-ous machinery. The concept of a local responseindependent of the perikaryon is attractive since itconfers on the terminal a degree of freedom toadjust its states of growth/degrowth according toevolving conditions, which can account for somephenomena associated with plasticity. This issue hasrecently been addressed in an in vitro neuronalmodel from Aplysia. A single axonal branch canundergo long-term facilitation when serotonin isapplied to its synaptic ®eld, while other branchesremain una�ected; this response is abolished bylocal inhibition of protein synthesis (Martin et al.,1997).
The evidence presented indicates that terminals arenot static structures of the neuron, but that they are ina dynamic state that is far greater than that of thestem axon. Modi®cation of the phenotype of terminalsrequires a constellation of molecular and structuralcomponents. To account for this within the frameworkof the slow transport model, the only mechanisms
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 49

available are those that redistribute existing proteinsor reallocate transported proteins that were syn-thesized in the perikaryon weeks, months, or evenyears before the sprouting stimulus arose. In thismodel, proteins allocated to a newly formed regionmust be subtracted from another region, since the sumtotal cannot increase. Within the framework of thelocal synthesis model, local cues modulate the internalprogram of the terminals that may alter functionalproperties (cf. Martin et al., 1997).
Plasticity of axons and terminals is highlighted intwo conditions: (i) when an innervated target ®eld nextto normal axons loses its nerve supply, and (ii) in het-erologous regeneration. After partial denervation of asynaptic or a receptive ®eld, neighboring axons extendcollateral and terminal sprouts, which replace the orig-inal arbors in the vacated territory, and thereby restorefunction. In this type of reinnervation, axon and targetconserve their original functional class designation.Heterologous reinnervation provides a wider variety ofcombinations and will be examined in some detail. Inthe CNS, axons do not regenerate spontaneously, how-ever, they invade a sciatic nerve graft, which is per-missive for growth, and develop a terminal arbor whenthey reach a suitable cell (Aguayo et al., 1983). Centralbranches of the nodose ganglion of the vagal nervethat synapse normally in the brain stem can reinner-vate and establish cholinergic synapses with sympath-etic neurons (De Castro, 1951), or with skeletalmuscles (Falempin and Rousseau, 1983). In the cat,hypoglossal nerves cross-sutured to postganglionicsympathetic nerves form functional contacts betweenmotor axons and smooth muscle cells of the nicitatingmembrane, and induce the deposition of acetylcholin-esterase (Vera et al., 1957; Lennon et al., 1967; Ines-trosa et al., 1979); i.e., smooth muscle cells that wereinnervated originally by autonomic adrenergic ®bersbecome reinnervated by regenerated somatic motoraxons. In these heterologous reinnervations, axonsattain lengths longer than the original ones, anddevelop functional synaptic terminals on foreign post-synaptic cells, while conserving their original synaptictransmitter.
A�erents can also form connections with foreignreceptor cells. Axons of the superior laryngeal nerve,which are mainly mechanoreceptor a�erents, can re-innervate the carotid body and establish functionalcontacts with its chemoreceptors cells (Zapata et al.,1969). An even more interesting case is that of the car-otid body grafted onto the tenuissimus muscle of thecat. Three to six months later, the discharge of a givensensory axon of the tenuissimus nerve could be drivenboth by mechanical stimuli applied to the muscle, andby chemical stimuli applied to the transplanted carotidbody, but not when application of stimuli to the re-spective tissue targets was reversed. The receptor cells
in the carotid body were associated with nerve term-inals resembling normal ones (Monti-Bloch et al.,1983). Therefore, the carotid body induced propiocep-tive axons to extend branches that made functionalcontacts with chemoreceptor cells while maintainingthe original contact with muscle spindles, and eachreceptor arbor responded only to speci®c stimuli. Thisis a clear example of merotrophism in which the ulti-mate phenotype of a terminal arbor, its molecularanatomy, is speci®ed locally.
In the cases mentioned above, regenerated axonsestablish a functional connection upon an e�ector, ora receptor cell that conserves its original anatomy.However, both components of the functional contactmay be of a novel kind. Regenerating sensory axonsare sensitive to acetylcholine (Diamond, 1959) andregenerating motor axons can release acetylcholine(Evans and Saunders, 1974). When central stumps ofregenerating sensory and motor nerves are allowed toform a neuroma, containing a mass of tangled ®bers,motor, but not sensory axons, elicit a local electricalresponse that fatigues; moreover, a tetanizing stimulusproduces post tetanic potentiation, and the evoked ac-tivity is sensitive to curare and prostigmine (Alvarez,1968), i.e., regrowing axons establish an arti®cialcholinergic synapse in which the original anatomicalrelationship is lost.
The nature of the neuron imposes restrictions on thetype of terminal arbors possible. Pacinian corpusclestransplanted onto the dorsal column of the spinal cordare reinnervated and terminals appear morphologicallynormal (Zelena and Jirmanova, 1988), even thoughaxons of the spinal cord normally form presynapticcontacts with other neurons. On the other hand, sen-sory axons can regrow along motor nerves but areunable to form endplates with muscle ®bers, andmotor axons are unable to produce terminals of thereceptor type in Pacinian corpuscles (Zelena et al.,1990).
The evidence presented suggests that mechanisms ofelongation are common to all axons, but that thegrowth cones can di�erentiate into a few specializ-ations that are only functionally competent with arestricted set of target cells. In our model, the stableanatomy of the adult axon implies a stable balance inlocal regulation, notwithstanding that there may bemild perturbations, such as a change in the ongoingelectrical activity, alteration of cells in its vicinity, ordrastic perturbations, such as nerve section, whichupsets the balance, and leads to a release and ex-pression of latent programs. The terminal arbor, incontrast, is structurally more plastic, suggesting thatexternal regulatory signals are more variable.
In summary, the axon is neither constrained in itsgrowth potential to a designated length, nor predes-tined to having a speci®c trajectory, or target. Since
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6250

length, trajectory, and target are circumstantial, en-vironmental signals must govern the behavior of agrowing axon. In this scenario, a local source of pro-teins and phospholipids in axons and growth cones(see Section 6.3.4) would play a pivotal role in theplasticity of the synaptic arbor. Obviously, this is nota proof but a hypothesis amenable to investigation.
7. Autonomy of axons
The existence of a machinery for protein synthesis inaxoplasm endows the axon with a certain degree ofautonomy; i.e., RNA transcripts present in axon andterminals could be translated selectively, or at di�erentrates, depending on the regulators that are operatingat the moment. This autonomy is relative, however,since, as discussed in Section 3.3. RNA transcripts inaxoplasm, this machinery will ultimately depend onthe perikaryon, or possibly on the ensheathing cell.The autonomy can be measured by the repertoire ofresponses that the axon can produce without a directparticipation of the perikaryon, and by its survivalafter separation from the perikaryon.
The immediate e�ect of a nerve lesion is the loss ofmembrane integrity and a breach of its barrier func-tion, which promotes unhindered free exchangebetween intracellular and extracellular compartments.This condition is transient, in that continuity of theaxolemma is rapidly restored with the healing overprocess. Vesicles and electron dense materials accumu-late at open ends of proximal and distal stumps of ver-tebrate and invertebrate axons (Zelena et al., 1968;Godell et al., 1997). In Aplysia, axons reestablish mem-brane continuity within 0.5±2 min post-axotomy (Zivand Spira, 1993). Healing over requires a signi®cantlocal increase of calcium, as shown in axons of cray-®sh, squid, gold®sh, and Aplysia (Godell et al., 1997;Ziv and Spira, 1993), which results in activation ofCa2+ dependent enzymes. This includes phospholipaseA2, which generates micelle-forming lysophospholipidsto increase membrane ¯uidity, in addition to gelsolin,which severs actin ®laments and promotes depolymeri-zation, and calpain proteases that rapidly cleaveselected actin binding proteins such as ankyrin andspectrin, and neuro®laments. Ca2+ dependent calpainsin particular rapidly solate the axoplasmic gel by theiraction on neuro®laments (Gilbert et al., 1975).
As noted above (see Section 6.4), stem axons canform growth cones de novo, and this is preceded by alocal rise of calcium. For this response, the cell body isunnecessary as isolated axons extend terminal sprouts(Rotshenker, 1981; Benbassat and Spira, 1994). Iso-lated stem axons of Wld mice extend sprouts at theirdecentralized end (InÄ iguez and Alvarez, 1999) andgrowth cones of severed optic axons of Xenopus
embryos continue their navigation toward the tectumand even arborize upon reaching it (Harris et al.,1987). The growth cone, which is a structure special-ized for motility, is formed by the axon in response toappropriate external signals, and remains motile evenwhen disconnected from the perikaryon (Shaw andBray, 1977).
The survival of severed axons varies considerablybetween species. In rat, degeneration is extensive twodays after the lesion (Vial, 1958; Nathaniel and Pease,1963), while in man, severed axons still conduct actionpotentials 7±10 days after injury. In poikilotherms, thesurvival of severed axons is conspicuously longer thanin homeotherms. Optic nerve axons of the frog maysurvive for up to two months (Matsumoto and Scalia,1981), and motor axons, for about two weeks (Rot-shenker, 1981), while the M-cell axon of the gold®shcan survive for over two months in moderate ambienttemperatures (Blundon et al., 1990; Zottoli et al.,1987). In poikilotherms, the delay in degenerationshortens with higher temperatures (Blundon et al.,1990; Matsumoto and Scalia, 1981). In general, de-generation occurs in a spatially distributed fashion,with no apparent selected vectorial preference, as theaxon appears to collapse at the same time throughoutits extent (Vial, 1958; Donat and Wisniewski, 1973;Moehlenbruck et al., 1994), although ovoids form ®rstclose to the lesion, and progress at a rate of 46 to 250mm/day (Lubinska, 1977).
Axonal degeneration entails an active destruction ofaxoplasm, and is not due to a form of ``starvation''following decentralization from the cell body. Withprogressive loss of membrane polarization, calciumchannels become activated, and calcium concentrationrises in axoplasm. Under culture conditions, pharma-cological blockade of L-type Ca2+ channels, but notof the N-type channels, delays the onset of neurite de-generation (George et al., 1995). The elevated increasein Ca2+ concentration results in activation of calpains(Schlaepfer, 1971; Schlaepfer and Micko, 1979;Schlaepfer and Hasler, 1979; Zimmerman and Schlaep-fer, 1982; Schlaepfer et al., 1985; Raabe et al., 1995),which leads to global breakdown, and solation of axo-plasm alluded to above.
It is clear that calcium plays a key role in healingover, growth cone formation, and with the later acti-vation of calpains, is instrumental in causing degener-ation of the axon. In this respect the Wlds mutantprovides some insight because the ability to survive isan intrinsic property of Wlds axons; however, theydegenerate when an ionophore is used to elevate axo-plasmic Ca2+ concentration (see Section 6.3.5). Thus,the mutation delays the entry of calcium that wouldnormally precede solation of axoplasm and degener-ation. Since mutations usually produce a functionalde®cit rather than a novel function, the long survival
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 51

of decentralized Wld axons probably re¯ects a missingfunction that a�ects calcium permeation, which is pre-requisite to triggering the cascade leading to cytolysis.
Axotomy is followed by degeneration distal to theinjury. Colchicine, which selectively interrupts microtu-bule dependent intracellular transport, also causes axo-nal degeneration (Angevine, 1957; Singer andSteinberg, 1972). The linkage between axonal degener-ation and rapid anterograde transport was made byLubinska (1977) to account for the rapid progressionof degeneration ovoids in severed ®bers. It wouldappear, therefore, that rapid anterograde transportcould play a central role in the long-term regulation ofthe entry of calcium into the axoplasm. This depen-dence on the perkaryon would seem to impose a tem-poral limit to the autonomy of the axon.
8. Final commentary
Evidence has been reviewed which shows that theaxon and its terminals have a capacity to synthesizeproteins, and that a delimited region of the axon,including terminals, can undergo local changes withoutdirect involvement of the perikaryon. We propose theterm merotrophism (from the Greek, (mEroB) meros,meaning part) to denote phenomena entailing localregulation of phenotype characteristics of a restrictedregion of cells. This form of regulation does notinvolve the nucleus of the neuron, in contrast to the``classical'' meaning of trophism, which connotes selec-tive regulation of genes that a�ect expression in thecell as a whole. Merotrophism would operate region-ally on cells that have extended processes such as neur-ons and oligodendrocytes.
Experimental studies have clearly shown that thebiological complexity of maintaining the axon and itsterminal arborization is greater than that previouslyconsidered. Traditionally, the axon has been viewedsimply as a line of communication, bridging the peri-karyon and its terminal arborization, that conductselectrical signals, and conveys bidirectional trophic sig-nals between the neuron and its target. Notwithstand-ing these basic functions, the axon and its terminalarbors are intrinsically capable of altering their struc-ture, organization and spatial distribution in responseto local events, as conditioned by their phenotype andhistory. In contrast to the slow transport model, whichrequires supplemental stringent ad hoc assumptions inorder to explain maintenance and plasticity changesassociated with the axon, the local synthesis modelreadily accounts for them without straining credulity.Finally, local protein synthesis appears to play a pivo-tal role in explaining merotrophic phenomena associ-ated with the axon and terminals.
Acknowledgements
We are indebted to many colleages whose commentsenriched this article. This work was supported bygrants FONDECYT 1980973, Chile, to J.A., and IBN-9604841 from the National Science Foundation, toE.K; A.G. gratefully acknowledges the friendly co-operation of the sta� of the Marine Biological Labora-tory (Woods Hole, MA, USA) where most work onthe squid giant axon and nerve terminals was carriedout; this work was supported over the years by severalgrant from CNR, MURST, University of Naples``Federico II'', EC, NSF, and NIH.
References
Abdel-Latif, A.A., 1966. A simple method for isolation of nerve-end-
ing particles from rat brain. Biochim. Biophys. Acta 121, 403±
406.
Abdul-Ghani, A.S., Boyar, M.M., Coutinho-Netto, J., Bradford,
H.F., 1980. E�ect of stimulation on the incorporation of 14C
from glial and neuronal speci®c substrates into brain proteins in
vivo and in vitro. J. Neurochem 35, 170±175.
Agius, E., Cochard, P., 1998. Comparison of neurite outgrowth
induced by intact and injured sciatic nerves: a confocal and func-
tional analysis. J. Neurosci 18, 328±338.
Aguayo, A.J., Attiwel, M., Trecarten, J., Perkins, S., Bray, G.M.,
1977. Abnormal myelination in transplanted Trembler mouse
Schwann cells. Nature 265, 73±75.
Aguayo, A.J., Benfey, M., David, S., 1983. A potential for axonal re-
generation in neurons of the adult mammalian nervous system.
Birth defects: original article series 19, 327±340.
Aguayo, A.J., Bray, G.M., Rasminsky, M., Zwimpfer, T., Carter,
D., Vidal-Sanz, M., 1990. Synaptic connections made by axons
regenerating in the central nervous system of adult mammals. J.
Exp. Biol 153, 199±224.
Alvarez, A., Alarco n, R., Opazo, C., Campos, E.O., MunÄ oz, F.J.,
Caldero n, F.H., Dajas, F., Gentry, M.K., Doctor, B.P., De
Mello, F.G., Inestrosa, N.C., 1998. Stable complexes involving
acetylcholinesterase and amyloid-b peptide change the biochemi-
cal properties of the enzyme and increase the neurotoxicity of
Alzheimer's ®brils. J. Neurosci 18, 3213±3223.
Alvarez, J., 1968. A possible arti®cial axo-axonic chemical synapse.
Acta Physiol. Latamer 18, 366±367.
Alvarez, J., 1979. Microtubules in non-medullated ®bres: e�ect of
the action potential. Neurosci. Lett 15, 15±17.
Alvarez, J., 1992. Maintenance of the axoplasm: can neurones accord
with the accepted notions? Neurosci. Lett 144, 1±3.
Alvarez, J., Benech, C.R., 1983. Axoplasmic incorporation of amino
acids in a myelinated ®ber exceeds that of its soma: a radioauto-
graphic study. Exper. Neurol 79, 283±286.
Alvarez, J., Chen, W.Y., 1972a. Injection of leucine into a myelinic
axon: incorporation in the axoplasm and transfer to associated
cells. Acta Physiol. Latinoamer 22, 266±269.
Alvarez, J., Chen, W.Y., 1972b. Local incorporation of choline by a
myelinated ®bre. Acta Physiol. Latinoamer 22, 270±273.
Alvarez, J., RamõÁ rez, B.U., 1979. Axonal microtubules: their regu-
lation by the electrical activity of the nerve. Neurosci. Lett 15,
19±22.
Alvarez, J., Torres, J.C., 1985. Slow axoplasmic transport: a ®ction?
J. Theor. Biol 112, 627±651.
Alvarez, J., Zarour, J., 1983. Microtubules in short and long axons
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6252

of the same caliber: Implications for the maintenance of the neur-
on. Exper. Neurol 79, 283±286.
Alvarez, J., Arredondo, F., Espejo, F., Williams, V., 1982.
Regulation of axonal microtubules: e�ect of sympathetic hyperac-
tivity elicited by reserpine. Neuroscience 7, 2551±2559.
Alvarez, J., Moreno, R.D., Llanos, O., Inestrosa, N.C., Brandan, E.,
Colby, T., Esch, F.S., 1992. Axonal sprouting induced in the scia-
tic nerve by the amyloid precursor protein (APP) and other anti-
proteases. Neurosci. Lett 144, 130±134.
Alvarez, J., Moreno, R.D., Inestrosa, N.C., 1995. Mitosis of
Schwann cells and demyelination are induced by the amyloid pre-
cursor protein and other protease inhibitors in the rat sciatic
nerve. Eur. J. Neurosci 7, 152±159.
Angevine, J.B., 1957. Nerve destruction by colchicine in mice and
golden hamster. J. Exp. Zool 136, 607±634.
Appel, S.H., Autilio, L., Festo�, B.W., Escueta, A.V., 1969.
Biochemical studies of synapses in vitro. Part II: Ionic activation
of protein synthesis. J. Biol. Chem 244, 3166±3172.
Austin, L., Morgan, I.G., 1967. Incorporation of 14C-labelled leu-
cine into synaptosomes from rat cerebral cortex in vitro. J.
Neurochem 14, 377±387.
Autilio, L.A., Appel, S.H., Pettis, P., Gambetti, P.L., 1968.
Biochemical studies of synapses in vitro. Part I: Protein synthesis.
Biochem 7, 2615±2622.
Baas, P.W., Brown, A., 1997. Slow axonal transport: the polymer
transport model. Trends Cell Biol 7, 380±384.
Baehr, M., Bunge, R.P., 1989. Functional status in¯uences the ability
of Schwann cells to support adult rat retinal ganglion cell survival
and axonal regrowth. Exper. Neurol 106, 27±40.
Barker, D., Ip, M.C., 1966. Sprouting and degeneration of of mama-
lian motor axons in normal and de-a�erented skeletal muscles.
Proc. R. Soc. London, Series B 163, 538±554.
Barker, L., 1899. The Nervous System. Appleton, New York, p. 307.
Barondes, S.H., 1964. Delayed appearance of labeled protein in iso-
lated nerve endings and axoplasmic ¯ow. Science 146, 779±781.
Barondes, S.H., 1966. On the site of synthesis of the mitochondrial
protein of nerve endings. J. Neurochem 13, 721±727.
Barondes, S.H., 1967. Further studies of the transport of protein to
nerve endings. J. Neurochem 15, 343±350.
Barra, H.S., Rodriguez, J.A., Arce, C.A., Caputto, R., 1973. A sol-
uble preparation from rat brain that incorporates into its own
proteins �14C]arginine by a ribonuclease-sensitive system and
�14C]tyrosine by a ribonuclease-insensitive system. J. Neurochem
20, 97±108.
Bassell, G.J., Singer, R.H., Kosik, K.S., 1994a. Association of
poly(A). mRNA with microtubules in cultured neurons. Neuron
12, 571±582.
Bassell, G.J., Powers, C.M., Taneja, K.L., Kislauskis, E.H., Sundell
L., C., Powers, C.M., Ross, A., Singer, R.H., 1994b. Actin ®la-
ments and spatial positioning of mRNAs. Adv. Exp. Med. Biol
358, 183±189.
Bassell, G.J., Zhang, H., Byrd, A.L., Femino, A.M., Singer, R.H.,
Taneja, K.L., Lifshitz, L.M., Herman, I.M., Kosik, K.S., 1998.
Sorting of b-actin mRNA and protein to neurites and growth
cones in culture. J. Neurosci 18, 251±265.
Bedi, K.S., Winter, J., Berry, M., Cohen, J., 1992. Adult rat dorsal
root ganglion neurons extend neurites on predegenerated but not
on normal peripheral nerves in vitro. Eur. J. Neurosci 4, 193±200.
Benavides, E., Alvarez, J., 1998. Peripheral axons of Wlds mice,
which do not regenerate, do so when transcription is inhibited in
the distal stump. Neurosci. Lett 258, 77±80.
Benbassat, D., Spira, M.E., 1994. The survival of transected axonal
segments of cultured Aplysia neurons is prolonged by contact
with intact nerve cells. Eur. J. Neurosci 6, 1605±1614.
Benech, C., Sotelo Jr., J.R., Mene ndez, J., Correa-Luna, R., 1982.
Autoradiographic study of RNA and protein synthesis in sec-
tioned peripheral nerves. Exper. Neurol 76, 72±82.
Benech, J.C., Crispino, M., Chun, J.-T., Kaplan, B.B., Giuditta, A.,
1994. Protein synthesis in nerve endings from squid brain: modu-
lation by calcium ions. Biol. Bull 187, 269.
Benech, J.C., Crispino, M., Kaplan, B.B., Giuditta, A., 1999. Protein
synthesis in presynaptic endings from squid brain: modulation by
calcium ions. J. Neurosci. Res 55, 776±781.
Benech, J.C., Crispino, M., Martin, R., Alvarez, J., Kaplan, B.B.,
Giuditta, A., 1996. Protein synthesis in the presynaptic endings of
the squid photoreceptor neuron: in vitro and in vivo modulation.
Biol. Bull 191, 263.
Bergeron-Bouvet, C., MouleÁ , Y., 1966. Saccharose gradient sedimen-
tation of RNA from rat liver microsomal membranes. Biochim.
Biophys. Acta 123, 617±620.
Bisby, M.A., Keen, P., 1985. The e�ect of a conditioning lesion on
the regeneration rate of peripheral nerve axons containing sub-
stance P. Brain Res 336, 201±206.
Bisby, M.A., Tetzla�, W., Brown, M.C., 1995. Cell body response to
injury in motoneurons and primary sensory neurons of a mutant
mouse, Ola (Wld)., in which Wallerian degeneration is delayed. J.
Comp. Neurol 359, 653±662.
Bisby, M.A., Tetzla�, W., Brown, M.C., 1996. GAP-43 mRNA in
mouse motoneurons undergoing axonal sprouting in response to
muscle paralysis of partial denervation. Eur. J. Neurosci 8, 1240±
1248.
Black, M., Lasek, R.J., 1980. Slow components of axonal transport:
two cytoskeletal networks. J. Cell Biol 86, 616±623.
Black, M.M., Lasek, J., 1977. The presence of transfer RNA in the
axoplasm of the squid giant axon. J. Neurobiol 8, 229±237.
Blundon, J.A., Sheller, R.A., Moehlenbruck, J.W., Bittner, G.D.,
1990. E�ect of temperature on long-term survival of anucleate
giant axons in cray®sh and gold®sh. J. Comp. Neurol 297, 377±
391.
Bosman, H.B., Hemsworth, B.A., 1970. Incorporation of amino
acids and monosaccharides into macromolecules by isolated
synaptosomes and synaptosomal mitochondria. J. Biol. Chem
245, 363±371.
Boyar, M.M., Abdul-Ghani, A.-S., Bradford, H.F., 1980. Changes in
protein synthesis in cortical subcellular fractions after activation
of the brachial plexus. Biochem. Soc. Trans 8, 3±4.
Bradley, G.W., Asbury, A.K., 1970. Duration of synthesis phase in
neurilemma cells in mouse sciatic nerve during degeneration.
Exper. Neurol 26, 275±282.
Brady, S.T., Lasek, R.J., 1981. Nerve-speci®c enolase and creatine
phosphokinase in axonal transport: soluble proteins and the axo-
plasmic matrix. Cell 23, 515±523.
Bridgers, W.F., Cunningham, R.D., Gresset, G., 1971. Properties
distinguishing mitochondrial and synaptosomal protein synthesis.
Biochem. Biophys. Res. Commun 45, 351±357.
Brown, M.C., Lunn, E.R., Perry, V.H., 1991. Poor growth of mam-
malian motor and sensory axons into intact proximal nerve
stumps. Eur. J. Neurosci 3, 1366±1369.
Brown, M.C., Perry, V.H., Hunt, S.P., Laper, S.R., 1994. Further
studies on motor and sensory nerve regeneration in mice with
delayed Wallerian degeneration. Eur. J. Neurosci 6, 420±428.
Brunetti, M., Giuditta, A., Porcellati, G., 1979. The synthesis of cho-
line phosphoglycerides in the giant ®bre system of the squid. J.
Neurochem 32, 319±324.
Buckmaster, E.A., Perry, V.H., Brown, M.C., 1995. The rate of
Wallerian degeneration in cultured neurons from wild type and
C57BL/WldS mice depends on time in culture and may be
extended in the presence of elevated K+ levels. Eur. J. Neurosci
7, 1596±1602.
Bustos, J., Vial, J.D., Fau ndez, V., Alvarez, J., 1991. Axonal micro-
tubules decrease after local inhibition of protein synthesis and
increase after inhibition of trnascription: a morphometric study in
rat sural nerves. Eur. J. Neurosci 3, 1123±1133.
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 53

Cajal, S.R., 1959. Degeneration and Regeneration of the Nervous
System. Hafner, New York (R.M. May, Trans.).
Caldero n, F.H., von Bernhardi, R., DeFerrari, G., Luza, S.,
Aldunate, R., Inestrosa, N.C., 1998. Toxic e�ects of acetylcholin-
esterase on neuronal and glial cells in vitro. Molec. Psychiatry 3,
247±255.
Campbell, M.K., Mahler, H.R., Moore, W.J., Tewari, S., 1966.
Protein synthesis systems from rat brain. Biochem 5, 1174±1184.
Cancalon, P., 1979. In¯uence of temperature on the velocity and on
the isotope pro®le of slowly transported labeled proteins. J.
Neurochem 32, 997±1007.
Carlin, R.K., 1977. Nonribosomal incorporation of amino acids into
the troponin-like protein from synaptosomes. J. Neurochem 28,
355±360.
Carlsen, R.C., Ki�, J., Ryugo, K., 1982. Suppression of the cell
body response in axotomized frog spinal neurons does not pre-
vent initiation of nerve regeneration. Brain Res 234, 11±25.
Celio, M.R., Gray, E.G., Yasargil, G.M., 1979. Ultrastructure of the
Mauthner axon collateral and its synapses in the gold®sh spinal
cord. J. Neurocytol 8, 19±29.
Chakraborty, G., Leach, T., Zanakis, M.F., Ingoglia, N.A., 1986.
Posttranslational protein modi®cation by amino acid addition in
regenerating optic nerves of gold®sh. J. Neurochem 46, 726±732.
Chen, S., Bisby, M.A., 1993. Impaired motor axon regeneration in
the C57BL/Ola mouse. J. Comp. Neurol 333, 449±454.
Cheng, T.P., Reese, T.S., 1987. Recycling of plasmalemma in chick
tectal growth cones. J. Neurosci 7, 1752±1759.
Cheung, M.K., 1989. The speci®city of glutamate inhibition of pro-
tein synthesis in synaptosomal fractions from rat cerebral cortex.
Neurochem. Int 15, 293±300.
Chun, J.T., Gioio, A.E., Crispino, M., Eyman, M., Giuditta, A.,
Kaplan, B.B., 1997. Molecular cloning and characterization of a
novel mRNA present in the squid giant axon. J. Neurosci. Res
49, 144±153.
Chun, J.T., Gioio, A.E., Crispino, M., Giuditta, A., Kaplan, B.B.,
1995. Characterization of squid enolase mRNA: sequence analay-
sis, tissue distribution, and axonal localization. Neurochem. Res
20, 923±930.
Chung, K., Lee, B.H., Yoon, Y.W., Chung, J.M., 1996. Sympathetic
sprouting in the dorsal root ganglia of the injured peripheral
nerve in a rat neuropathic pain model. J. Comp. Neurol 376,
241±252.
Cleveland, D.W., Lopata, M.A., Sherline, P., Kirschner, M.W.,
1981. Unpolymerized tubulin modulates the level of tubulin
mRNAs. Cell 25, 537±546.
Cohen, A.I., 1973. An ultrastructural analysis of the photoreceptors
of the squid and their synaptic connections. III. Photoreceptor
terminations in the optic lobe. J. Comp. Neurol 147, 399±426.
Coleman, M.P., Conforti, L., Buckmaster, E.A., Tarlton, A., Ewing,
R.M., Brown, M.C., Lyon, M.F., Perry, V.H., 1998. An 85-kb
tandem triplication in the slow Wallerian degeneration (Wlds).
mouse. Proc. Natl. Acad. Sci. USA 95, 9985±9990.
Condeelis, J., 1995. Elongation factor 1a and the regulation of cyto-
skeletal dynamics, mRNS sorting and growth control. Trends
Biochem. Sci 233, 169±215.
Conradi, S., 1966. Ultrastructural specialization in the initial segment
of cat lumber motoneurons. Acta Soc. Med. Upsalien 71, 281±
284.
Contreras, G., Carrasco, O.C., Alvarez, J., 1983. Axoplasmic incor-
poration of amino acids in myelinated ®bers of the cat. Exper.
Neurol 82, 581±593.
Corbaton, V., MuinÄ o, M.T., Ferna ndez-Silva, P., Lo pez-Pe rez, M.J.,
Montoya, J., 1991. Estimation of the chloramphenicol and cyclo-
heximide inhibition of protein synthesis in brain cholinergic
synaptosomes. Brain Res 543, 351±353.
Cotman, C.W., Taylor, D.A., 1971. Autoradiographic analysis of
protein synthesis in synaptosomal fractions. Brain Res 29, 366±
372.
Crispino, M., Castigli, E., Perrone Capano, C., Martin, R.,
Menichini, E., Kaplan, B.B., Giuditta, A., 1993a. Protein syn-
thesis in a synaptosomal fraction from squid brain. Mol. Cell
Neurosci 4, 366±374.
Crispino, M., Kaplan, B.B., Martin, R., Alvarez, J., Chun, J.T.,
Benech, J.C., Giuditta, A., 1997. Active polysomes are present in
the large presynaptic endings of the synaptosomal fraction from
squid brain. J. Neurosci 17, 7694±7702.
Crispino, M., Perrone Capano, C., Kaplan, B.B., Giuditta, A.,
1993b. Neuro®lament proteins are synthesized in nerve endings
from squid brain. J. Neurochem 61, 1144±1146.
Cunningham, R.D., Bridgers, W.F., 1970. Brain and liver mitochon-
drial protein synthesis: potassium dependent chloramphenicol in-
hibition. Biochem. Biophys. Res. Commun 38, 99±105.
Cupello, A., Hyde n, H., 1975. Fractionation of RNA from brain
synaptosomes and cytoplasmic subcellular fractions. J.
Neurochem 25, 399±406.
Cupello, A., Hyde n, H., 1977. A comparison of poly(A).-associated
RNA from synaptosomes and cytoplasmic subcellular fractions of
rat brain. Biochem. Exper. Biol 13, 130±136.
Cupello, A., Hyde n, H., 1982. Labeling of poly(A).-associated RNA
in synaptosomes and the other subcellular fractions of rat cer-
ebral cortex in basal conditions and during training. J. Neurosci.
Res 8, 575±579.
Cupello, A., Rosadini, G., 1976. Poly(A).-associated RNA from
mitochondria and microsomes of rat brain. Experientia 32, 872±
874.
Cutillo, V., Montagnese, P., Gremo, F., Casola, L., Giuditta, A.,
1983. Origin of axoplasmic RNA in the squid giant ®ber.
Neurochem. Res 8, 1621±1634.
Danielsen, N., Kerns, J.M., Holmquist, B., Zhao, Q., Lundborg, G.,
Kanje, M., 1994. Pre-degenerated nerve grafts enhance regener-
ation by shortening the initial delay period. Brain Res 666, 250±
254.
Davis, L., Dou, P., DeWit, M., Kater, S.B., 1992. Protein synthesis
within neuronal growth cones. J. Neurosci 12, 4867±4877.
De Castro, F., 1951. Sur la structure de la synapse dans les chemore-
cepteurs: leur me canisme d'excitation et le roà le dans la circulation
sanguine locale. Acta Physiol. Scand 21, 14±43.
De Robertis, E., De Iraldi, A.P., Rodriguez, G., Gomez, J., 1961.
On the isolation of nerve endings and synaptic vesicles. J.
Biophys. Biochem. Cytol 9, 229±235.
De Waegh, S., Brady, S.T., 1990. Altered slow axonal transport and
regeneration in a myelin-de®cient mutant mouse: the trembler as
an in vivo model for Schwann cell-axon interactions. J. Neurosci
10, 1855±1865.
De Waegh, S.M., Brady, S.T., 1991. Local control of axonal proper-
ties by Schwann cells: neuro®laments and axonal transport in
homologous and heterologous nerve grafts. J. Neurosci. Res 30,
201±212.
De Waegh, S.M., Lee, V.M., Brady, S.T., 1992. Local modulation of
neuro®lament phosphorylation, axonal caliber, and slow axonal
transport by myelinating Schwann cells. Cell 68, 451±463.
Deanin, G.G., Gordon, M.W., 1973. Chloramphenicol- and cyclo-
heximide-sensitive protein synthetic systems in brain mitochon-
drial and nerve-ending preparations. J. Neurochem 20, 55±68.
DeLarco, J., Nakagawa, S., Abramowitz, A., Bromwell, K., Guro�,
G., 1975. Polyadenylic acid-containing RNA from rat brain
synaptosomes. J. Neurochem 25, 131±137.
Dellmann, H.-D., RodrõÂ guez, E.M., PenÄ a, P., Siegmund, I., 1988.
Immunohistochemical investigation of the magnocellular pepti-
dergic hypothalomo-neurohypophyseal system of the rat chroni-
cally stimulated by long-term administration of hypertonic saline.
Neuroendocrinol 47, 335±342.
DenisDonini, S., Branduardi, P., Campiglio, S., Carnevali, M.D.C.,
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6254

1998. Localization of calcitonin gene-related peptide mRNA in
developing olfactory axons. Cell. Tissue Res 294, 81±91.
Di Battista, A., Cupello, A., 1988. Microelectrophoretic fractionation
of RNA from rat brain subsynaptosomal fractions. Boll. Soc.
Ital. Biol. Sper 11, 1025±1032.
Diamond, J., 1959. The e�ects of injecting acetylcholine into normal
and regenerating nerves. J. Physiol., London 145, 611±629.
DõÂ az, J., Pecot-Dechavassine, M., 1990. Nerve sprouting induced by
a piece of peripheral nerve placed over a normally innervated
frog muscle. J. Physiol., London 421, 123±133.
Dillman, J.F., Dabney III, L.P., P®ster, K.K., 1996. Cytoplasmic
dynein is associated with slow axonal transport. Proc. Natl.
Acad. Sci. USA 93, 141±144.
Dirks, R.W., van Dorp, A.G.M., van Minnen, J., Fransen, J.A.M.,
van der Ploeg, M., Raap, A.K., 1993. Ultrastructural evidence
for the axonal localization of caudodorsal cell hormone mRNA
in the central nervous system of the mollusc Lymnaea stagnalis.
Microsc. Res. Tech 25, 12±18.
Donat, J.R., Wisniewski, H.M., 1973. The spatio-temporal pattern
of Wallerian degeneration in mammalian peripheral nerves. Brain
Res 53, 41±53.
Dowdall, M.J., Whittaker, V.P., 1973. Comparative studies in synap-
tosome formation: the preparation of synaptosomes from the
head ganglion of the squid Loligo pealii. J. Neurochem 20, 921±
935.
Droz, B., Leblond, C.B., 1963. Axonal migration of proteins in the
central nervous system and peripheral nerves as shown by radio-
autography. J. Comp. Neurol 121, 325±346.
Droz, B., Barondes, S.H., 1969. Nerve endings: rapid appearance of
labeled protein shown by electron microscope radioautography.
Science 165, 1131±1133.
Droz, B., Koenig, H.L., Di Giamberardino, L., 1973. Axonal mi-
gration of protein and glycoprotein to nerve endings. Part I:
Radioarutographic analysis of the renewal of protein in nerve
endings of chicken ciliary ganglion after intracerebaral injection
of 3H lysine. Brain Res 60, 93±127.
Droz, B., Rambourg, A.R., Koenig, H.L., 1975. The smooth endo-
plasmic reticulum: structure and role in the renewal of axonal
membrane and synaptic vesicles by fast axonal transport. Brain
Res 93, 1±13.
Dupree, J.L., Bigbee, J.W., 1994. Retardation of neuritic outgrowth
and cytoskeletal changes accompany acetylcholinesterase inhibitor
treatment in cultured rat dorsal root ganglion neurons. J.
Neurosci. Res 39, 567±575.
Dupree, J.L., Bigbee, J.W., 1996. Acetylcholinesterase inhibitor treat-
ment delays recovery from axotomy in cultures dorsal root
ganglion neurons. J. Neurocytol 25, 439±454.
Edbladh, M., Tonge, D., Golding, J., Ekstrom, A.R., Edstrom, A.,
1994. Early regeneration in vitro of adult mouse sciatic axons is
dependent on local protein synthesis but may not involve neuro-
trophins. Neurosci. Lett 168, 37±40.
Edds, M.V., 1950. Hypertrophy of nerve ®bers to functionally over-
loaded muscles. J. Comp. Neurol 93, 259±275.
EdstroÈ m, A., 1964a. The ribonucleic acid in the Mauthner neuron of
the gold®sh. J. Neurochem 11, 309±314.
EdstroÈ m, A., 1964b. E�ect of spinal cord transection on the base
composition and content of RNA in the Mauthner nerve ®bre of
the gold®sh. J. Neurochem 11, 557±559.
EdstroÈ m, A., 1967. Inhibition of protein synthesis in Mauthner nerve
®bre components by actinomycin-D. J. Neurochem 14, 239±243.
EdstroÈ m, A., 1966. Amino acid incorporation in isolated Mauthner
nerve ®bre of gold®sh. J. Neurochem 13, 315±321.
EdstroÈ m, A., SjoÈ strand, J., 1969. Protein synthesis in isolated
Mauthner nerve ®bre components. J. Neurochem 16, 67±81.
EdstroÈ m, A., EdstroÈ m, J.E., HoÈ kfelt, T., 1969. Sedimentation analy-
sis of ribonucleic acid extracted from isolated Mauthner nerve
®bre components. J. Neurochem 16, 53±66.
EdstroÈ m, J.-E., Eichner, D., EdstroÈ m, A., 1962. The ribonucleic acid
of axons and myelin sheaths from Mauthner neurons. Biochim.
Biophys. Acta 61, 178±184.
Elliot, E.J., Muller, K.J., 1983. Sprouting and regeneration of sen-
sory axons after destruction of ensheathing glial cells in the leech
central nervous system. J. Neurosci 3, 1994±2006.
Ellisman, M.H., Lindsey, J.D., 1983. The axoplasmic reticulum
within myelinated axons is not transported rapidly. J. Neurocytol
12, 393±411.
Eng, H., Lund, K., Campenot, R.B., 1999. Synthesis of b-tubulin,actin and other proteins in axons of sympathetic neurons in com-
partmentalized cultures. J. Neurosci 19, 1±19.
England, J.M., Attardi, G., 1976. Analysis of RNA synthesized by
an isolated rat brain synaptosomal fraction. J. Neurochem 27,
895±904.
Espejo, F., Alvarez, J., 1986. Microtubules and calibers in normal
and regenerating axons of the sural nerve of the rat. J. Comp.
Neurol 250, 65±72.
EugenõÂ n, J., Alvarez, J., 1995. Incorporation of amino acids into the
axoplasm is enhanced by electrical stimulation of the ®ber. Brain
Res 677, 319±325.
Evans, C.A.N., Saunders, N.R., 1974. An out¯ow of acetylcholine
from normal and regenerating ventral roots of the cat. J.
Physiol., London 240, 15±32.
Fabrizi, C., Kelly, B.M., Gillespie, C.S., Schlaepfer, W.W., Scherer,
S.S., Brophy, P.J., 1997. Transient expression of the neuro®la-
ment proteins NF-L and NF-M by Schwann cells is regulated by
axonal contact. J. Neurosci. Res 50, 291±299.
Fadic, R., Alvarez, J., 1986. Calibers and microtubules of sympath-
etic axons are not subject to trophic control by the preganglionic
nerve. Exper. Neurol 94, 237±240.
Fadic, R., Vergara, J., Alvarez, J., 1985. Microtubules and caliber of
central and peripheral processes of sensory axons. J. Comp.
Neurol 236, 258±264.
Fahlman, C.S., Riley, D.A., 1986. Colchicine-induced sprouting of
the neuromuscular junction in the pigeon extensor digitorum
longus muscle. Brain Res 363, 156±160.
Falempin, M., Rousseau, J.P., 1983. Reinnervation of skeletal
muscles by vagal sensory ®bres in the sheep, cat and rabbit. J.
Physiol. (Lond) 335, 467±479.
Fau ndez, V., Alvarez, J., 1986. Microtubules and calibers in develop-
ing axons. J. Comp. Neurol 250, 73±80.
Fau ndez, V., Cordero, M.E., Rosso, P., Alvarez, J., 1989. Calibers
and microtubules of nerve ®bers: di�erential e�ect of undernutri-
tion in developing and adults rats. Brain Res 509, 198±204.
Ferna ndez, H.L., Moreno, R.D., Inestrosa, N.C., 1996. Tetrameric
(G4). acetylcholinesterase: structure, localization, and physiologi-
cal regulation. J. Neurochem 66, 1335±1346.
Fink, H., Gainer, D.J., 1980. Axonal transport of proteins: a
new view using in vivo covalent labeling. J. Cell Biol 85, 175±186.
Fisher, S., Litvak, S., 1967. The incorporation of microinjected 14C-
aminoacid into TCA insoluble fraction of the giant axon in the
squid. J. Cell Physiol 70, 69±74.
Forgue, S.T., Dahl, J.L., 1978. The turnover rate of tubulin in rat
brain. Neurochem 31, 1289±1297.
Frey, U., Morris, R.G.M., 1997. Synaptic tagging and long-term po-
tentiation. Nature 385, 533±536.
Friede, R.L., Samorajski, T., 1970. Axon caliber related to neuro®la-
ments and microtubules in sciatic nerve ®bers of rat and mice.
Anat. Rec 167, 379±388.
Funch, G.P., Kinsman, S.L., Faber, D.S., Koenig, E., Zottoli, S.J.,
1981. Mauthner axon diameter and impulse conduction velocity
decrease with growth of gold®sh. Neurosci. Lett 27, 150±164.
Gaete, J., Kameid, G., Alvarez, J., 1998. Regenerating axons of the
rat require a local source of proteins. Neurosci. Lett 251, 197±
200.
Gainer, H., Tasaki, I., Lasek, R.J., 1977. Evidence for the glia-neur-
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 55

on protein transfer hypothesis from intracellular profusion studies
of squid giant axons. J. Cell Biol 74, 524±530.
Gambetti, P.L., Autilio-Gambetti, L.A., Gonatas, N.K., Shafer, B.,
1972. Protein synthesis in synaptosomal fractions, Ultrastructural
radioautographic study. J. Cell Biol 52, 526±535.
Gambetti, P., Autilio-Gambetti, L., Shafer, B., Pfa�, L., 1973.
Quantitative autoradiography of labeled RNA in rabbit optic
nerve after intraocular injection of [3H]uridine. J. Cell Biol 59,
677±684.
George, E.B., Glass, J.D., et al., 1995. Axotomy-induced axonal de-
generation is mediated by calcium in¯ux through ion-speci®c
channels. J. Neurosci 15, 6445±6452.
Giftochristos, N., David, S., 1988. Immature optic nerve glia of rat
do not promote axonal regeneration when transplanted into a
peripheral nerve. Brain Res 467, 149±153.
Gilbert, D.S., Newby, B.J., Anderton, B.H., 1975. Neuro®lament dis-
guise, destruction and discipline. Nature 256, 586±589.
Gilbert, J.M., 1972. Evidence for protein synthesis in synaptosomal
membranes. J. Biol. Chem 247, 6541±6550.
Gioio, A.E., Chun, J.T., Crispino, M., Capano, C.P., Giuditta, A.,
Kaplan, B.B., 1994. Kinesin mRNA is present in the squid giant
axon. J. Neurochem 63, 13±18.
Giuditta, A., 1980. Origin of axoplasmic protein in the squid giant
axon. Riv. Biol 73, 35±49.
Giuditta, A., Dettbarn, W.-D., Brzin, M., 1968. Protein synthesis in
the isolated giant axon of the squid. Proc. Natl. Acad. Sci. USA
59, 1284±1287.
Giuditta, A., Cupello, A., Lazzarini, G., 1980. Ribsomal RNA in the
axoplasm of the squidgiant axon. J. Neurochem 34, 1757±1760.
Giuditta, A., Hunt, T., Santella, L., 1983. Messenger RNA in squid
axoplasm. Biol. Bull 165, 526.
Giuditta, A., Hunt, T., Santella, L., 1986. Messenger RNA in squid
axoplasm. Neurochem. Intern 8, 435±442.
Giuditta, A., Menichini, E., Perrone Capano, C., Langella, M.,
Martin, R., Castigli, E., Kaplan, B.B., 1991. Active polysomes in
the axoplasm of the squid giant axon. J. Neurosc. Res 26, 18±28.
Giuditta, A., Metafora, S., Felsani, A., Del Rio, A., 1977. Factors
for protein synthesis in the axoplasm of the squid giant axon. J.
Neurochem 28, 1393±1395.
Glass, J.D., Gri�n, J.W., 1991. Neuro®lament redistribution in
transected nerves: evidence for bidirectional transport of neuro®-
laments. J. Neurosci 11, 3146±3154.
Glass, J.D., Brushart, T.M., George, E.B., Gri�n, J.W., 1993.
Prolonged survival of transected nerve ®bres in C57BL/Ola mice
is an intrinsic characteristic of the axon. J. Neurocytol 22, 311±
321.
Glass, J.D., Schryer, B.L., Gri�n, J.W., 1994. Calcium-mediated de-
generation of the axonal cytoskeleton in the Ola mouse. J.
Neurochem 62, 2472±2475.
Godell, C.M., Smyers, M.E., Eddleman, C.S., Ballinger, M.L.,
Fishman, H.M., Bittner, G.D., 1997. Calpain activity promotes
the sealing of severed giant axons. Proc. Natl. Acad. Sci. USA
94, 4751±4756.
Goldberg, M.A., 1971. Protein synthesis in isolated rat brain mito-
chondria and nerve endings. Brain Res 27, 319±328.
Goldberg, M.A., 1972. Inhibition of synaptosomal protein synthesis
by neurotransmitter substances. Brain Res 39, 171±179.
Gonatas, N.K., Autilio-Gambetti, L., Gambetti, P., Shafer, B., 1971.
Morphological and biochemical changes in rat synaptosome frac-
tions during neonatal development. J. Cell Biol 51, 484±498.
Gordon, M.W., Deanin, G.G., 1968. Protein synthesis by isolated
rat brain mitochondria and synaptosomes. J. Biol. Chem 243,
4222±4226.
Gower, D.J., Tytell, M., 1986. Protein modi®cation by RNA-depen-
dent posttranslational aminoacylation in synaptoplasm. J.
Neurochem 47, 389±395.
Grafstein, B., Alpert, R.M. 1982. Properties of slow axonal trans-
port: studies in gold®sh optic axons. In: Weiss, D.G. (Ed.),
Axoplasmic Transport. Springer-Verlag, Berlin, Heidelberg, pp.
226±231.
Grafstein, B., Edwards, D.L., Alpert, R.M. 1981. Axonal transport
and neuronal activity. In: Schweigger, H.G. (Ed.), International
Cell Biol. Springer-Verlag, Berlin, pp. 728±736.
Grainger, F., Sloper, J.C., 1974. Correlation between microtubular
number and transport activity of hypothalomo-neurohypophyseal
secretory neurons. Cell. Tiss. Res 153, 101±113.
Gray, E.G., Whittaker, V.P., 1962. The isolation of nerve endings
from brain: an electron-microscopic study of cell fragments de-
rived by homogenization and centrifugation. J. Anat 96, 79±88.
Gregson, N.A., Hall, S., 1973. A quantitative analysis of the e�ects
of the intraneural injection of lysophosphatidyl choline. J. Cell
Sci 13, 257±277.
Haldar, D., 1970. Protein synthesis in mammalian brain mitochon-
dria. Biochem. Biophys. Res. Commun 40, 129±134.
Haldar, D., 1971. Protein synthesis in isolated rat brain mitochon-
dria. Biochem. Biophys. Res. Commun 42, 899±904.
Hallinan, T., Munro, H.N., 1965. Protein synthesis and ribonucleic
acid turnover in rat liver microsome subfractions. Biochim.
Biophys. Acta 108, 285±296.
Hannan, A.J., Schevzov, G., Gunning, P., Je�rey, P.L., Weinberger,
R.P., 1995. Intracellular localization of tropomyosin mRNA and
protein is associated with development of neuronal polarity.
Molec. Cell Neurosci 6, 397±412.
Hannan, A.J., Henke, R.C., Weinberger, R.P., Sentry, J.W., Je�rey,
P.L., 1996. Di�erential induction and intracellular localization of
SCG10 messenger RNA is associated with neuronal di�eren-
tiation. Neurosci 72, 889±900.
Harris, W.A., Holt, C.E., Bonheo�er, F., 1987. Retinal axons with
and without their somata growing to and arborizing in the tectum
of Xenopus embryos: a time-laser video study of single ®bres in
vivo. Development 101, 123±133.
Hefti, F., Lichtensteiger, W., 1978. Subcellular distribution of dopa-
mine in substantia nigra of the rat brain: e�ects of .-butyrolac-tone and destruction of noradrenergic a�erents suggest formation
of particles from dendrites. J. Neurochem 30, 1217±1230.
Henn, F.A., Anderson, D.J., Rustad, D.G., 1976. Glial contami-
nation of synaptosomal fractions. Brain Res 101, 341±344.
Heriot, K., Gambetti, P., Lasek, R.J., 1985. Proteins transported in
slow components a and b of axonal transport are distributed dif-
ferently in the transverse plane of the axon. J. Cell Biol 100,
1167±1172.
Hernandez, A., Burdett, I., Work, T.S., 1971. Protein synthesis by
brain-cortex mitochondria. Biochem. J 124, 327±336.
Hernandez, A.G., 1974. Protein synthesis by synaptosomes from rat
brain, Contribution by intraterminal mitochondria. Biochem. J
142, 7±17.
Hernandez, A.G., Langford, G.M., Martinez Jr., J.L., Dowdall,
M.J., 1976. Protein synthesis by synaptosomes from the head
ganglion of the squid Loligo pealli. Acta Cient. Venez 27, 120±
123.
Hernandez, A.G., Sua rez, G., Roman, H., Dawidowicz, K., 1978.
Protein synthesis by synaptosomes from rat brain: the e�ect of
centrifugal forces. Exp. Brain Res 33, 325±335.
Herna ndez, C., Blackburn, E., Alvarez, J., 1989. Calibre and micro-
bules of the nonmedullated and myelinated domains of optic
nerve axons. Eur. J. Neurosci 1, 654±658.
Herrmann, C., Volknandt, W., Wittich, B., Kellner, R.,
Zimmermann, H., 1996. The major vault protein (MVP100). in
cholinergic nerve terminals of electric ray electric organ. J. Biol.
Chem 271, 13908±13915.
Hesketh, J.E., Pryme, I.F., 1991. Interaction between mRNA, ribo-
somes and the cytoskeleton. Biochem. J 277, 1±10.
Hill, R.R., Robbins, N., Fang, Z.P., 1991. Plasticity of presynaptic
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6256

and postsynaptic elements of neuromuscular junctions repeatdly
observed in living adult mice. J. Neurocytol 20, 165±182.
Hirokawa, N., Sato-Yoshitake, R., Kobayashi, N., P®ster, K.K.,
Bloom, G.S., Brady, S., 1991. Kinesin associates with anterogra-
dely transported membranous organelles in vivo. J. Cell Biol 114,
295±302.
Hirokawa, N., Terada, S., Funakoshi, T., Takeda, S., 1997. Slow
transport: the subunit transport model. Trends Cell Biol 7, 380±
384.
Ho�man, P.N., Lasek, R.J., 1975. The slow component of axonal
transport. J. Cell Biol 66, 351±366.
Hollenbeck, P.J., Bray, D., 1987. Rapidly transported organelles
containing membrane and cytoskeletal components: their relation
to axonal growth. J. Cell Biol 105, 2827±2835.
Ide, C., 1996. Peripheral nerve regeneration. Neurosci. Res 25, 101±
121.
Inestrosa, N.C., Me ndez, B., Luco, J.V., 1979. Acetylcholinesterase
like that of skeletal muscle in smooth muscle reinnervated by a
motor nerve. Nature (Lond) 280, 504±506.
Ingoglia, N.A., Giuditta, A., Zanakis, M.F., Babigian, A., Tasaki, I.,
Chakraborty, G., Sturman, J., 1983. Incorporation of �3H]amino
acids into proteins in a partially puri®ed fraction of axoplasm:
Evidence for transfer RNA mediated, post-translational protein
modi®cation in squid giant axon. J. Neurosci 3, 2463±2473.
InÄ iguez, A., Alvarez, J., 1999. Isolated axons of Wld(s). mice regrow
centralward. Neurosci. Lett 268, 108±110.
Irwin, C.C., 1985. Comparison of protein synthesis in mitochondria,
synaptosomes, and intact brain cells. J. Neurochem 44, 433±438.
Iurato, S., Luciano, L., Franke, K., Pannese, E., Reale, E., 1974.
Histochemical localization of acetylcholinesterase activity in the
cochlear and vestibular ganglion cells. Acta Otorlaryngol
(Stockholm) 78, 28±35.
Jirikowski, G.F., Sanna, P.P., Bloom, F.E., 1990. mRNA coding for
oxytocin is present in axons of the hypothalamo-neurohypophy-
sial tract. Proc. Natl. Acad. Sci. U.S.A 87, 7400±7404.
Jones, D.G., 1967. An electron microscopy study of subcellular frac-
tions of octopus brain. J. Cell Sci 2, 573±584.
Jones, L.R., Mahler, H.R., Moore, W.J., 1975. Synthesis of mem-
brane protein in slices of rat cerebral cortex. J. Biol. Chem 250,
973±983.
Jones, S.A., Holmes, C., Budd, T.C., Green®eld, S.A., 1995. The
e�ect of acetylcholinesterase on outgrowth of dopaminergic neur-
ons in organotypic slice culture of rat midbrain. Cell. Tiss. Res
279, 323±330.
Joo , F., Karnushina, I., 1975. Morphometric assessment of the com-
position of the synaptosomal fractions obtained by the use of
Ficoll gradients. J. Neurochem 24, 839±840.
Kang, H., Schuman, E.M., 1996. A requirement for local protein
synthesis in neurotrophin-induced hippocampal synaptic plas-
ticity. Science 273, 1402±1406.
Kanje, M., Lundborg, G., EdstroÈ m, A., 1988. A new method for the
studies of the e�ect of locally applied drugs on peripheral nerve
regeneration. Brain Res 439, 116±121.
Kaplan, B.B., Gioio, A.E., Perrone Capano, C., Crispino, M.,
Giuditta, A., 1992. b-actin and b-tubulin are components of a
heterogeneous mRNA population present in the squid giant
axon. Mol. Cell. Neurosci 3, 133±144.
Karlsson, J.-O. 1982. Axonal transport of macromolecules. In:
Brown, I.R. (Ed.), Molecular approaches to neurobiology.
Academic Press, New York, pp. 131±157.
Karlsson, J.-O., SjoÈ strand, J., 1971a. Synthesis, migration and turn-
over of protein in retinal ganglion cells. J. Neurochem 18, 749±
767.
Karlsson, J.-O., SjoÈ strand, J., 1971b. Transport of microtubular pro-
tein in axons of retinal ganglion cells. J. Neurochem 18, 975±982.
Kawabuchi, M., Boyne, A.F., Deshpande, S.S., Cintra, W.M.,
Brossi, A., Albuquerque, E.X., 1988. Enantiomer (+).physostig-
mine prevents organophosphate-induced subjunctional damage at
the neuromuscular synapse by a mechanism not related to cholin-
esterase carbamylation. Synapse 2, 139±147.
Kelly, B.M., Gillespie, C.S., Sherman, D.L., Brophy, P.J., 1992.
Schwann cells of the myelin-forming phenotype express neuro®la-
ment protein NF-M. J. Cell Biol 118, 397±410.
Keymer, J.E., Gaete, J., Kameid, G., Alvarez, J., 1999.
Acetylcholinesterase and inhibitors: e�ects upon normal and
regenerating nerves of the rat. Eur. J. Neurosci 11, 1049±1057.
Klee, C.B., Sokolo�, L., 1965. Amino acid incorporation into pro-
teolipid of myelin in vitro. Proc. Natl. Acad. Sci. USA 53, 1014±
1021.
Kobayashi, S., Goto, S., Anzai, K., 1991. Brain-speci®c small RNA
transcript of the identi®er sequences is present as a 10S ribonu-
cleoprotein particle. J. Bio. Chem 266, 4726±4730.
Kobayashi, S., Higashi, N., Susuki, K., Goto, S., Yumoto, K.,
Anzai, K., 1992. The 10S BC-1 ribonucleoprotein particle con-
tains identi®er sequence-binding proteins that interact with an
array of GCAAG/CTTGC motifs between split protomer
sequences for RNA polymerase III. J. Biol. Chem 267, 18291±
18297.
Koenig, E., 1965. Synthestic mechanisms in the axon. Part II: RNA
in myelin-free axons of the cat. J. Neurochem 12, 357±361.
Koenig, E., 1967. Synthetic mechanisms in the axon. Part IV: In
vitro incorporation into axonal protein and RNA. J. Neurochem
14, 437±446.
Koenig, E., 1970. Membrane protein synthesizing machinery of the
axon. In: Costa, E., Giacobini, E. (Eds.), Biochemistry of simple
neruonal models, Advances in Biochemical Psychopharmacology,
vol. 2. Raven, New York, pp. 305±315.
Koenig, E., 1979. Ribosomal RNA in Mauthner axon: implications
for a protein synthesizing machinery in the myelinated axon.
Brain Res 175, 95±107.
Koenig, E., 1984. Local synthesis of axonal protein. In: Lajtha, A.
(Ed.), Handbook of Neurochemistry, vol. 7. Plenum, New York,
pp. 315±340.
Koenig, E., 1986. Isolation of native Mauthner cell axoplasm and an
analysis of organelle movement in non-aqueous and aqueous
media. Brain Res 398, 288±297.
Koenig, E., 1989. Cycloheximide-sensitive [35S]methionine labeling
of proteins in gold®sh retinal ganglion cell axons in vitro. Brain
Res 481, 119±123.
Koenig, E., 1991. Evaluation of local synthesis of axonal proteins in
the gold®sh Mauthner cell axon and axons of dorsal and ventral
roots of the rat in vitro. Mol. Cell. Neurosci 2, 384±394.
Koenig, E., Adams, P., 1982. Local protein synthesizing activity in
axonal ®elds regenerating in vitro. J. Neurochem 39, 386±400.
Koenig, E., Giuditta, A., 1999. Protein-synthesizing machinery in the
axon compartment. Neuroscience 89, 5±15.
Koenig, E., Koelle, G.B., 1961. Acetylcholinesterase regeneration in
peripheral nerve after irreversible inactivation. Science 132, 1249±
1250.
Koenig, E., Martin, R., 1996. Cortical plaque-like structures identify
ribosome-containing domains in the Mauthner axon. J. Neurosci
16, 1400±1411.
Koenig, E., Martin, R., 1997. Ribosome-like ESI signals localized in
discrete cortical domains of rabbit ventral root axons. Mol. Biol.
Cell 8, 215a.
Koenig, E., Kinsman, S., Repasky, E., Sultz, L., 1985. Rapid moti-
lity of motile varicosities and inclusions containing a-spectrin,actin and calmodulin in regenerating axons in vitro. J. Neurosci
5, 715±729.
Korn, A.P., Spitnik-Elson, P., Elson, D., Ottensmeyer, F.P., 1983.
Speci®c visualization of ribosomal RNA in the intact ribosome
by electron spectroscopic imaging. Eur. J. Cell Biol 31, 334±340.
Ku�er, D.P., 1986. Isolated satellite cells of a peripheral nerve direct
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 57

the growth of regenerating frog axons. J. Comp. Neurol 249, 57±
64.
Kuhn, T.B., Williams, C.V., Dou, P., Kater, S.B., 1998. Laminin
directs growth cone navigation via two temporally and function-
ally distinct calcium signals. J. Neurosci 18, 184±194.
Kurokawa, M., Sakamoto, T., Kato, M., 1965. Distribution of
sodium-plus-potassium-stimulated adenosine-triphosphatase ac-
tivity in isolated nerve-ending particles. Biochem. J 97, 833±844.
Kuznetsov, S.A., Langford, G.M., Weiss, D.G., 1992. Actin-depen-
dent organelle movement in squid axoplasm. Nature 356, 722±
725.
La Fleur, M., Underwood, J.L., Rappolee, D.A., Werb, Z., 1996.
Basement membrane and repair of injury to peripheral nerve:
de®ning a potential role for macrophages, matrix metalloprotei-
nases, and tissue inhibitor of metalloproteinases-1. J. Exp. Med
184, 2311±2326.
Landry, M., HoÈ kfelt, T., 1998. Subcellular localization of preproga-
lanin messenger RNA in perikarya and axons of hypothalamo-
posthypophyseal magnocellular neurons: and in situ hybridization
study. Neurosci 84, 897±912.
Langley, J.N., Anderson, H.K., 1904. The union of di�erent kinds of
nerve ®bres. J. Physiol., London 31, 365±391.
Lasek, R.J., 1986. Polymer sliding in axons. J. Cell Sci. (Suppl.) 5,
161±179.
Lasek, R.J., Ho�man, P.N., 1976. The neuronal cytoskeleton, axonal
transport and axonal growth. In: Goldman, R., Pollard, T.D.,
Rosenbaum, J. (Eds.), Cell Motility, vol. 3. Cold Spring Harbor
Symposium, New York, pp. 1021±1049.
Lasek, R.J., Dabrowski, C., Nordlander, R., 1973. Analysis of axo-
plasmic RNA from invertebrate giant axons. Nature New Biol
244, 162±165.
Lasek, R.J., Gainer, H., Barker, J.L., 1977. Cell-to-cell transfer of
glial proteins to the squid giant axon. J. Cell Biol 74, 501±523.
Lasek, R.J., Gainer, H., Pryzblyski, R.J., 1974. Transfer of newly
synthesized proteins from Schwann cells to the squid giant axon.
Proc. Natl. Acad. Sci. U.S.A 71, 1188±1192.
Lasek, R.J., Oblinger, M.M., Drake, P.F., 1983. Molecular biology
of neuronal geometry: expression of neuro®lament genes in¯u-
ences axonal diameter. Cold Spring Harbor Symposia en
Quantitative Biology 48, 731±743.
Laughrea, M., 1982. Transfer ribonucleic acid-dependent but ribo-
some-independent leucine incorporation into rat brain protein.
Biochem 21, 5694±5700.
Layer, P.G., Weikert, T., Alber, R., 1993. Cholinesterases regulate
neurite growth in chick nerve cells in vitro by means of a non en-
zymatic mechanism. Cell. Tissue Res 273, 219±226.
Lemkey-Johnston, N., Dekirmenjian, H., 1970. The identi®cation of
fractions enriched in nonmyelinated axons from rat whole brain.
Exp. Brain Res 11, 392±410.
Lemkey-Johnston, N., Larramendi, L.M.H., 1968. The separation
and identi®cation of fractions of nonmyelinated axons from the
cerebellum of the cat. Exp. Brain Res 5, 326±340.
Lennon, A.M., Vera, C.L., Rex, A.L., Luco, J.V., 1967.
Cholinesterase activity of the nictitating membrane reinnervated
bycholinergic ®bers. J. Neurophysiol 30, 1523±1530.
Lerner, E.A., Lerner, M.R., Janeway, C.A., Steitz, J.A., 1981.
Monoclonal antibodies to nucleic acid-containing cellular con-
stituents: probes for molecular biology and autoimmune disease.
Proc. Natl. Acad. Sci. USA 78, 2737±2741.
Li, J.-Y., Volknandt, W., DahlstroÈ m, A., Herrmann, C., Blasi, J.,
Das, B., Zimmermann, H., 1999. Axonal transport of ribonucleo-
protein particles (Vaults). Neurosci 91, 1055±1065.
Liu, G., Edmonds, B.T., Condeelis, J., 1996. pH, EF1a and the
cytoskeleton. Trends Cell Biol 6, 168±171.
Loguercio Polosa, P., Attardi, G., 1991. Distinctive pattern and
translational control of mitochondrial protein synthesis in rat
brain synaptic endings. J. Biol. Chem 266, 10011±10017.
Lo pez, J.M.A., Alvarez, J., 1990. The microtubular pattern changes
at the spinal cord-root junction and reverts at the root-peripheral
nerve junction in sensory and motor ®bres of the rat. Eur. J.
Neurosci 2, 873±878.
Lotti, M., 1991. The pathogenesis of organophosphate polyneuropa-
thy. Crit. Rev. Toxicol 21, 465±487.
Low, P.A., 1976. Hereditary hypertrophic neuropathy in the trembler
mouse. Part 1, Histopathological studies: light microscopy. J.
Neurol. Sci 30, 327±341.
Lubinska, L., 1977. Early course of Wallerian degeneration in myeli-
nated ®bres of the rat phrenic nerve. Brain Res 130, 47±63.
Lunn, E.R., Perry, V.H., Brown, M.C., Rosen, H., Gordon, S.,
1989. Absense of Wallerian degeneration does not hinder regener-
ation in peripheral nerve ®bres. Eur. J. Neurosci 1, 27±33.
Lyon, M.F., Ogunkolade, B.W., Brown, M.C., Atherton, D.J.,
Perry, V.H., 1993. A gene a�ecting Wallerian nerve degeneration
maps distally on mouse chromosome 4. Proc. Natl. Acad. Sci.
USA 90, 17±20.
Marinesco, G. 1909. La Cellule nerveuse, O. Doin, Paris.
Martin, K.C., Casadio, A., Zhu, H.E.Y., Rose, J.C., Chen, M.,
Bailey, C.H., Kandel, E.R., 1997. Synapse-speci®c, long-term fa-
cilitation of aplysia sensory to motor synapses: a function for
local protein synthesis in memory storage. Cell 91, 927±938.
Martin, R. 1997. Ribosomes in peripheral and presynaptic domains
of axons. In: Teelken, A.W., Korf, J. (Eds.), Neurochemistry.
Plenum, New York, pp. 661±665.
Martin, R., Vaida, B., et al., 1998. Protein synthesizing units in pre-
synaptic and postsynaptic domains of squid neurons. J. Cell Sci
111, 3157±3166.
Martin, R., Fritz, W., Giuditta, A., 1989. Visualization of polyribo-
somes in the postsynaptic area of the squid giant synapse by elec-
tron spectroscopic imaging. J. Neurocytol 418, 11±18.
Mason, A., Muller, K.J., 1982. Axon segments sprout at both ends:
tracking growth with ¯uorescent D-peptides. Nature 296, 655±
657.
Massoulie , J., Pezzementi, L., Bon, S., Krejci, E., Vallete, F.M.,
1993. Molecular and cellular biology of cholinesterases. Prog.
Neurobiol 41, 31±91.
Masuda-Nakagawa, L.M., Muller, K.J., Nicholls, J.G., 1993. Axonal
sprouting and laminin appearance after destruction of glial
sheaths. Proc. Natl. Acad. Sci. USA 90, 4966±4970.
Matsumoto, D.E., Scalia, F., 1981. Long-term survival of centrally
projecting axons in the optic nerve of the frog following destruc-
tion of the retina. J Comp. Neurol 202, 135±155.
McEwen, B.S., Forman, D.S., Grafstein, B., 1971. Components of
fast and slow axonal transport in the gold®sh optic nerve. J.
Neurobiol 2, 361±377.
McQuarrie, I.G., Jacob, J.M., 1991. Conditioning nerve crush accel-
erates cytoskeletal protein transport in spronts that form after a
subsequent crush. J. Comp. Neurol 305, 139±147.
Meier, R., Spreyer, P., Ortmann, R., Harel, A., Monard, D., 1989.
Induction of glia-derived nexin after lesion of a peripheral nerve.
Nature 342, 548±550.
Melia, K.R., Thembleau, A., Oddi, R., Sanna, P.P., Bloom, E.,
1994. Detection and regulation of tyrosine hydroxylase mRNA in
catecholaminergic terminal ®elds: possible axonal compartmenta-
lization. Exper. Neurol 130, 394±406.
Menichini, E., Castigli, E., Kaplan, B.B., Giuditta, A., 1990.
Synthesis of axoplasmic RNA particles inthe isolated squid giant
axon. Neurosci. Res. Comm 7, 89±96.
Meyerson, G., Parrow, V., Gestblom, C., Johansson, I., Pahlman, S.,
1994. Protein synthesis and mRNA in isolated growth cones from
di�erentiating SH-SY5Y neuroblastoma cells. J. Neurosci. Res
37, 303±312.
Miller, K.E., Samuels, D.C., 1997. The axon as a metabolic compart-
ment: protein degradation, transport, and maximum length of an
axon. J. Theor. Biol 186, 373±379.
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6258

Moehlenbruck, J.W., Cummings, J.A., Bittner, G.D., 1994. Long-
term survival followed by degradation of neuro®lament proteins
in severed mauthner axons of gold®sh. J. Neurobiol 25, 1637±
1651.
Mohr, E., Richter, D., 1992a. Diversity of mRNAs in the axonal
compartment of peptidergic neurons in the rat. Eur. J. Neurosci
4, 870±876.
Mohr, E., Fehr, S., Richter, D., 1991. Axonal transport of neuropep-
tide encoding mRNAs within the hypothalamo-hypophyseal tract
of rats. EMBO J 10, 2419±2424.
Mohr, E., Richter, D., 1992b. Diversity of messenger RNAs in the
axonal compartment of peptidergic neurons in the rat. Eur. J.
Neurosci 4, 870±876.
Monard, D., 1988. Cell-derived proteases and protease inhibitors as
regulators of neurite outgrowth. Trends Neurosci 11, 541±544.
Monard, D., 1993. Tinkering with certain blood components can
engender distinct functions in the nervous system. Perspect Dev.
Neurobiol 1, 165±168.
Monti-Bloch, L., Stensaas, L.J., Eyzaguirre, C., 1983. Carotid body
grafts induce chemosensitivity in muscle nerve ®bers of the cat.
Brain Res 270, 77±92.
Moreno, R.D., Inestrosa, N.C., Culwell, A.R., Alvarez, J., 1996.
Sprouting and abnormal contacts of nonmedullated axons, and
deposition of extracellular material induced by the amyloid pre-
cursor protein (APP). and other protease inhibitors. Brain Res
718, 13±24.
Morgan, I.G., 1970. Protein synthesis in brain mitochondrial and
synaptosomal prepparations. FEBS Lett 10, 273±275.
Morgan, I.G., 1976. Synaptosomes and cell separation. Neurosci 1,
159±165.
Morgan, I.G., Austin, L., 1968. Synaptosomal protein synthesis in a
cell-free system. J. Neurochem 15, 41±51.
Morgan, I.G., Austin, L., 1969. Energy metabolism and synaptoso-
mal protein synthesis. Life Sci 8, 79±84.
Mori, H., Komiya, Y., Kurokawa, M., 1979. Slowly migrating axo-
nal polypeptides. J. Cell Biol 82, 174±184.
Morris, J.R., Lasek, R.J., 1984. Monomer-polymer equilibria in the
axon: direct measurement of tubulin and actin as polymer and
monomer in the axoplasm. J. Cell Biol 98, 2064±2076.
Muir, D., Manthorpe, M., 1992. Stromelysin generates a ®bronectin
fragment that inhibits Schwann cell proliferation. J. Cell Biol 116,
177±185.
Muma, N.A., Ho�man, P.N., 1993. Neuro®laments are intrinsic
determinants of axonal caliber. Micron 24, 677±683.
Muslimov, L.A., Titmus, M., Bremer, R., Koenig, E., Tiedge, H.,
1997. RNA transport in neurons: conservation in evolution. Soc.
Neurosci. Abstr 23 (Part 2), 2267.
Nathaniel, E.J.H., Pease, D.C., 1963. Degenerative changes in rat
dorsal roots during wallerian degeneration. J. Ultrastruct. Res 9,
511±532.
Nien, J.K., Schmidt, J., Cartier, L., Alvarez, J., 1998. Cerebrospinal
¯uid of HTLV-1 associated myelopathy patients induces axonal
sproutings and Schwann cell proliferation in the rat sciatic nerve.
J. Neurol. Sci 159, 17±24.
Nixon, R., 1998. The slow axonal transport of cytoskeletal proteins.
Curr. Opin. Cell Biol 10, 87±92.
Nixon, R.A., 1980. Protein degradation in the mouse visual system.
Part I: Degradation of axonally transported and retinal proteins.
Brain Research 200, 69±83.
Nixon, R.A., Logvinenko, K.B., 1986. Multiple fates of newly
synthezised neuro®lament proteins: evidence for a stationary neu-
ro®lament network distributed nonuniformly along axons of reti-
nal ganglion cell neurons. J. Cell Biol 102, 647±659.
Okabe, S., Hirokawa, N., 1990. Turnover of ¯uorescently labelled
tubulin and actin in the axon. Nature 343, 479±482.
Okabe, S., Hirokawa, N., 1992. Di�erential behavior of photoacti-
vated microtubules in growing axons of mouse and frog neurons.
J. Cell Biol 117, 105±120.
Okabe, S., Hirokawa, N., 1991. Actin dynamics in growth cones.
Neuroscience 11, 1918±1929.
Oleynikov, Y., Singer, R.H., 1998. RNA localization: di�erent zip-
codes, same postman. Trends Cell Biol 8, 381±383.
Olink-Coux, M., Hollenbeck, P.J., 1996. Localization and active
transport of mRNA in axons of sympathetic neurons in culture.
J. Neurosci 16, 1346±1358.
Ornelles, D.A., Fey, E.G., Penman, S., 1986. Cytochalasin releases
mRNA from the cytoskeletal framework and inhibits protein syn-
thesis. Mol. Cell. Biol 6, 1650±1662.
Orrego, F., Lipmann, F., 1967. Protein synthesis in brain slices.
E�ect of electrical stimulation and acidic amino acids. J. Biol.
Chem 242, 665±671.
Ottensmeyer, F.P., 1986. Elemental mapping by energy ®ltration: ad-
vantages, limitations and compromises. Ann. N.Y. Acad. Sci 483,
339±353.
Pachter, B.R., Eberstein, A., 1984. Neuromuscular plasticity follow-
ing limb immobilization. J. Neurocytol 13, 1013±1025.
Palay, S.L., Palade, G.E., 1955. The ®ne structure of neurons. J.
Biophys. Biochem. Cytol 1, 69±88.
Palay, S.L., Sotelo, C., Peters, A., Orkand, P.M., 1968. The axon hil-
lock and the initial segment. J. Cell Biol 38, 193±201.
Pamphlett, R., 1988. Axonal sprouting after botulinum toxin does
not elicit a histological axon reaction. J. Neurol Sci 87, 175±185.
Pannese, E., Ledda, M., 1991. Ribosomes inmyelinated axons of the
rabbit spinal ganglion neurons. J. Submicrosc. Cytol. Pathol 23,
33±38.
Pannese, E., Ledda, M., Matsuda, S., 1988. Nerve ®bres with myeli-
nated and unmyelinated portions in dorsal spinal roots. J.
Neurocytol 17, 693±700.
Pannese, E., Ledda, M., Arcidiacono, G., Rigamonti, L., Procacci,
P., 1984a. Density and distribution of microtubules in the axons
of the lizard dorsal root. J. Submicrosc. Cytol 13, 169±181.
Pannese, E., Ledda, M., Arcidiacono, G., Rigamonti, L., Procacci,
P., 1984b. A comparison of the density of microtubules in the
central and peripheral axonal branches of the pseudounipolar
neurons of lizard spinal ganglia. Anat. Rec 208, 595±605.
Parker, G.H., 1932. On the trophic impulse so-called, its rate and
nature. Am. Naturalist 66, 147±158.
Perrone Capano, C., Giuditta, A., Castigli, E., Kaplan, B.B., 1987.
Occurrence and sequence complexity of polyadenylated RNA in
squid axoplasm. J. Neurochem 49, 698±704.
Perrone Capano, C., Crispino, M., Menichini, E., Kaplan, B.B.,
Giuditta, A., 1999. Ribosomal RNAs synthesized by isolated
squid nerves and ganglia di�er from native ribosomal RNAs. J.
Neurochem 72, 910±918.
Peters, A., Proskauer, C.G., Kaiserman-Abramof, I.R., 1968. The
small pyramidal neuron of the cat cerebral cortex. The axon hil-
lock and the initial segment. J. Cell Biol 39, 604±619.
Petrushka, E., Giuditta, A., 1959. Electron microscopy of two sub-
cellular fractions isolated from cerebral cortex homogenate. J.
Biophys. Biochem. Cytol 6, 129±132.
Pipeleers, D.G., Pipeleers-Marichal, M.A., Kipnis, D.M., 1977.
Physiological regulation of total tubulin and polymerized tubulin
in tissues. J. Cell Biol 74, 351±357.
Pittman, R.N., DiBenedetto, A.J., 1995. PC12 cells overexpressing
tissue plasminogen activator regenerate neurites to a greater
extent and migrate faster than control cells in complex extracellu-
lar matrix. J. Neurochem 64, 566±575.
Posse de Chaves, E., Vance, D.E., Campenot, R.B., Vance, J.E.,
1995a. Axonal synthesis of phosphatidylcholine is required for
normal axonal growth in rat sympathetic neurons. J. Cell Biol
128, 913±918.
Posse de Chaves, E., Vance, D.E., Campenot, R.B., Vance, J.E.,
1995b. Alkylphosphocholines inhibit choline uptake and phospha-
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 59

tidylcholine biosynthesis in rat sympathetic neurons and impair
axonal extension. Biochem. J 312, 411±417.
Purves, D., Voyvodic, J.T., Magrassi, L., Yawo, H., 1987. Nerve
terminal remodeling visualized in the iving mice by repeated ob-
servations of the same neuron. Science 238, 1122±1126.
Raabe, T.D., Nguyen, T., Bittner, G.D., 1995. Calcium-activated
proteolysis of neuro®lament proteins in gold®sh Mauthner axons.
J. Neurobiol 26, 253±261.
Raisman, G., 1968. Neuronal plasticity in the septal nuclei of the
adult rat. Brain Res 14, 25±48.
Ramirez, G., 1973. Synaptic plasma membrane protein synthesis:
selective inhibition by chloramphenicol in vivo. Biochem.
Biophys. Res. Commun 50, 452±458.
Ramirez, G., Levitan, I.B., Mushynski, W.E., 1972a. Highly puri®ed
synaptosomal membranes from rat brain, Incorporation of amino
acids into membrane proteins in vitro. J. Biol. Chem 247, 5382±
5390.
Ramirez, G., Levitan, I.B., Mushynski, W.E., 1972b. Amino acid in-
corporation into nerve ending structures in vitro. FEBS Lett 21,
17±20.
Rao, A., Steward, O., 1991. Evidence that protein constituents of
postsynaptic membrane specializations are locally synthesized:
analysis of proteins synthesized within synaptosomes. J. Neurosci
11, 2881±2895.
Rapallino, M.V., Cupello, A., Giuditta, A., 1988. Axoplasmic RNA
species synthesized in the isolated giant axon. Neurochem. Res
13, 625±631.
Remak, R., 1839. Ueber die Verrichtungen des organischen
Nervensystems. Froriep's Notizen 7, 65±70.
Resseler, K.J., Sullivan, S.L., Buck, L.B., 1994. Information coding
in the olfactory system: evidence for a stereotyped and highly
organized epitope map in the olfactory bulb. Cell 79, 1245±1255.
Riley, D.A., Fahlman, C.S., 1985. Colchicine-induced di�erential
sprouting of the endplates on fast and slow muscle ®bers in rat
extensor digitorum longus, soleus and tibialis anterior muscles.
Brain Res 329, 83±95.
Roberson, M.D., Toews, A.D., Goodrum, J.F., Morell, P., 1992.
Neuro®lament and tubulin mRNA expression in Schwann cells.
J. Neurosci. Res 33, 156±162.
Rotshenker, S., 1981. Sprouting and synapse formation by motor
axons separated from their cell bodies. Brain Res 223, 141±145.
Rotshenker, S., 1982. Transneuronal and peripheral mechanisms for
the induction of motor neuron sprouting. J. Neurosci 2, 1359±
1368.
Rotshenker, S., 1988. Multiple modes and sites for the induction of
axonal growth. Trends Neurosci 11, 363±366.
Rotundo, R.L., Carbonetto, S.T., 1987. Neurons segregate clusters
of membrane-bound acetylcholinesterase along their neurites.
Proc. Nat. Acad.Sci. USA 84, 2063±2067.
Saitua, F., Alvarez, J., 1988. Do axons grow during adulthood? A
study of caliber and microtubules in sural nerve axons of young,
mature and aging rats. J. Comp. Neurol 269, 203±209.
Saitua, F., Alvarez, J., 1989. Microtubular packing varies along the
course of motor and sensory axons: possible regulation of micro-
tubules by environmental cues. Neurosci. Lett 104, 249±252.
Schlaepfer, W.W., 1971. Experimental alterations of neuro®laments
and neurotubules by calcium and other ions. Exp. Cell. Res 67,
73±80.
Schlaepfer, W.W., Hasler, M.B., 1979. Characterization of the cal-
cium-induced disruption of neuro®laments in rat peripheral
nerve. Brain Res 168, 299±309.
Schlaepfer, W.W., Micko, S., 1979. Calcium-dependent alterations of
neuro®lament proteins of rat peripheral nerve. J. Neurochem 32,
211±219.
Schlaepfer, W.W., Lee, C., Lee, V.M., Zimmerman, U.J., 1985. An
immunoblot study of neuro®lament degradation in situ and
during calcium-activated proteolysis. J. Neurochem 44, 502±509.
Schnapp, B., Mugnaini, E. 1978. Membrane architecture of myeli-
nated ®bers as seen by freeze-fracture. In: Waxman, S.G. (Ed.),
Physiology and Pathobiology of Axons. Raven Press, New York,
pp. 83±123.
Schwab, M.E., 1996. Structural plasticity of the adult CNS: negative
control by neurite growth inhibitory signals. Int. J. Dev. Neurosci
14, 379±385.
Schwab, M.E., Thoenen, H., 1985. Dissociated neurons regenerate
into sciatic but not optic nerve explants in culture irrespective of
neurotrophic factors. J. Neurosci 5, 2415±2423.
Serra, M., Alvarez, J., 1989. On the asymmetry of the primary
branching of vagal sensory axons: possible role of the supporting
tissue. J. Comp. Neurol 284, 108±118.
Shapot, V., Pitot, H.C., 1966. Isolation and fractionation of ribonu-
cleic acid from the smooth endoplasmic reticulum of rat liver.
Biochim. Biophys. Acta 119, 37±45.
Shaw, G., Bray, D., 1977. Movement and extension of isolated
growth cones. Exp. Cell Res 104, 55±62.
Sheller, R.A., Tytell, M., Smyers, M., Bittner, G.D., 1995. Glia-to-
axon communication: enrichment of glial proteins transferred to
the squid giant axon. J. Neurosci. Res 4, 324±334.
Shen, Y.J., DeBellard, M.E., Salzer, J.L., Roder, J., Filbin, M.T.,
1998. Myelin-associated glycoprotein in myelin and expressed by
Schwann cells inhibits axonal regeneration and branching. Molec.
Cell. Neurosci 12, 79±91.
Sieghart, W., Singer, E., 1979. Sedimentation and release properties
of glial particles present in P2-fractions isolated from rat cerebral
cortex. Brain Res 170, 203±208.
Sieghart, W., SellstroÈ m, A., Henn, F., 1978. Sedimentation character-
istics of subcellular vesicles derived from three glial systems. J.
Neurochem 30, 1587±1589.
Singer, M., Steinberg, M.C., 1972. Wallerian degeneration: a reeva-
luation based on transected and colchicine poisoned nerves in the
amphibian Triturus. Am. J. Anat 133, 51±85.
Singer, R.H., 1992. The cytoskeleton and mRNA localization. Curr.
Opin. Cell Biol 4, 15±19.
SjoÈ berg, J., Kanje, M., 1990. The initial period of peripheral nerve
regeneration and the importance of local environment for the
conditioning lesion e�ect. Brain Res 529, 79±84.
Slobin, L.I., 1980. The role of eucaryotic elongation factor 2 in pro-
tein synthesis. Eur. J. Biochem 110, 555±563.
Small, D.H., Reed, G., Whit®eld, B., Nurcombe, V., 1995.
Cholinergic regulation of neurite outgrowth for isolated chick
sympathetic neurons in culture. J. Neurosci 15, 144±151.
Smirnova, I.V., Ma, J.Y., Citron, B.A., Ratzla�, K.T., Gregory,
E.J., Akaaboune, M., Festo�, B.W., 1996. Neural thrombin and
protease nexin I kinetics after murine peripheral nerve injury. J.
Neurochem 67, 2188±2199.
Smith, R., 1973. Microtubule and neuro®lament densities in amphi-
bian spinal root nerve ®bers: relationship to axoplasmic trans-
port. Can. J. Physiol. Pharmacol 51, 798±806.
Son, Y.J., Thompson, W.J., 1995a. Schwann cell processes guide re-
generation of peripheral axons. Neuron 14, 125±132.
Son, Y.J., Thompson, W.J., 1995b. Nerve sprouting in muscle is
induced and guided by processes extended by Schwann cells.
Neuron 14, 133±141.
Sotelo, J.R., Benech, C.R., Kun, A., 1992. Local radiolabeling of the
68 kDa neuro®lament protein in rat sciatic nerves. Neurosci. Lett
144, 174±176.
Sotelo, J.R., Kun, A., Benech, J.C., Giuditta, A., Morillas, J.,
Benech, C.R., 1999. Ribosomes and polyribosomes are present in
the squid giant axon: an immunocytochemical study. Neurosci
90, 705±715.
Spira, M.E., Benbassat, D., Dormann, A., 1993. Resealing of the
proximal and distal cut ends of transected axons: electrophysio-
logical and ultrastructural analysis. J. Neurobiol 24, 300±316.
Srivatsan, M., Peretz, B., 1997. Acetylcholinesterase promotes regen-
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6260

eration of neurites in cultured adult neurons of Aplysia.
Neuroscience 77, 921±931.
Stossel, T.P., 1993. On the crawling of animal cells. Science 260,
1086±1094.
Stromska, D.P., Ochs, S., 1981. Patterns of slow transport in sensory
nerves. J. Neurobiol 12, 441±453.
Tapia, M., Inestrosa, N.C., Alvarez, J., 1995. Early regeneration:
repression by Schwann cells and a protease. Exper. Neurol 131,
124±132.
Terasa, S., Nakata, T., Peterson, A.C., Hirokawa, N., 1996.
Visualization of slow axonal transport in vivo. Science 273, 784±
788.
Thoenen, H., Mueller, R.A., Axelrod, J., 1970. Phase di�erence in
the induction of tyrosine hydroxylase in cell body and nerve
terminals of sympathetic neurons. Proc. Natl. Acad. Sci. USA 65,
58±62.
Tiedge, H., Fremeau Jr., R.T., Weinstock, P.H., Arancio, O.,
Brosius, J., 1991. Dendritic location of neural BC1 RNA. Proc.
Natl. Acad. Sci. USA 88, 2093±2097.
Tiedge, H., Zhou, A., Thorn, N.A., Brosius, J., 1993. Transport of
BC1 RNA in hypothalamo-neurohypophyseal axons. J. Neurosci
13, 4214±4219.
Titmus, M.J., Muslimov, I.A., Koenig, E., Tiedge, H., 1998. Signal
recognition particles (SRPs): in the axonal compartment. Mol.
Biol. Cell 9, 190a.
Tobias, G.S., Koenig, E., 1975a. Axonal protein synthesizing activity
during the early outgrowth period following neurotomy. Exper.
Neurol 49, 221±234.
Tobias, G.S., Koenig, E., 1975b. In¯uence of nerve cell body and
neurolemma cell on local axonal protein synthesis following neu-
rotomy. Exper. Neurol 49, 235±245.
Trembleau, A., Morales, M., Bloom, F.E., 1994. Aggregation of
vasopressin mRNA in a subset of axonal swellings of the median
eminence and posterior pituitary: light and electron microscopic
evidence. J. Neurosci 14, 39±53.
Trembleau, A., Morales, M., Bloom, F.E., 1996. Di�erential com-
partmentalization of vasopressin mRNA and neuropeptide within
the rat hypothalamo-neurohypophyseal axonal tracts: light and
electron microscopic evidence. Neurosci 70, 113±125.
Tsao, J.W., Brown, M.C., Carden, M.J., McLean, W.G., Perry,
V.H., 1994. Loss of the compound action potential: an electro-
physiological, biochemical and morphological study of early
events in axonal degeneration in the C57BL/Ola mouse. Eur. J.
Neurosci 6, 516±524.
Tsukita, S., Tsukita, S., Kobayashi, T., Matsumoto, G., 1986.
Subaxolemmal cytoskeleton in squid giant axon. Part II:
Morphological identi®cation of microtubule- and micro®lament-
associated domains of axolemma. J. Cell Biol 102, 1710±1725.
Tu�ery, A.R., 1971. Growth and degeneration of motor end plates
in normal cat hind limbs muscles. J. Anat 110, 221±247.
Tytell, M., Lasek, R.J., 1984. Glial polypeptides transferred into the
squid giant axon. Brain Res 324, 223±232.
Tytell, M., Greenberg, S.G., Lasek, R.J., 1986. Heat shock-like pro-
tein is transferred fromglia to axon. Brain Res 363, 161±164.
van Minnen, J., 1994. RNA in the axonal domain: a new dimension
in neuronal functioning. Histochem. J 26, 377±391.
van Minnen, J., Bergman, J.J., Van Kesteren, E.R., Smit, A.B.,
Geraerts, W.P., Lukowiak, K., Hasan, S.U., Syed, N.I., 1997. De
novo protein synthesis in isolated axons of identi®ed neurons.
Neuroscience 80, 1±7.
Vance, J.E., Pan, D., Vance, D.E., Campenot, R.B., 1991.
Biosynthesis of membrane lipids in rat axons. J. Cell Biol 115,
1061±1068.
Vance, J.E., Pan, D., Campenot, R.B., Bussiere, M., Vance, D.E.,
1994. Evidence that the major membrane lipids, except choles-
terol, are made in axons of cultured rat sympathetic neurons. J.
Neurochem 62, 329±337.
Vera, C.L., Vial, J.D., Luco, J.V., 1957. Reinnervation of nictitating
membrane of cat by cholinergic ®bers. J. Neurophysiol 30, 620±
627.
Vassar, R., Chao, S.K., Sitcheran, R., Nunez, J.M., Vosshall, L.B.,
Axel, R., 1994. Topographic organization of sensory projections
to the olfactory bulb. Cell 79, 981±991.
Vergara, J., Serra, J., Saitua, F., Iturriaga, R., Alvarez, J., 1991.
Axonal microtubules: comparative anatomy in vertebrates,
including man. J. Submicrosc. Cytol. Pathol 23, 357±363.
Vergara, J., Repetto, G., Alvarez, J., 1992. The axonal microtubular
density is higher than normal in ®bres innervating spastic
muscles. J. Submicrosc. Cytol. Pathol 24, 129±134.
Verity, M.A., 1972. Cation modulation of synaptosomal respiration.
J. Neurochem 19, 1305±1317.
Verity, M.A., Brown, W.J., Cheung, M., 1975. Organic mercurial
encephalopathy: in vivo and in vitro e�ects of methyl mercury on
synaptosomal respiration. J. Neurochem 25, 759±766.
Verity, M.A., Brown, W.J., Cheung, M., Czer, G., 1977a. Methyl
mercury inhibition of synaptosome and brain slice protein syn-
thesis: in vivo and in vitro studies. J. Neurochem 29, 673±679.
Verity, M.A., Brown, W.J., Cheung, M.K., Czer, G.T., 1977b.
Thyroid hormone inhibition of synaptosome amino acid uptake
and protein synthesis. J. Neurochem 29, 853±858.
Verity, M.A., Brown, W.J., Cheung, M.K., 1979. On the mechanism
of ouabain inhibition of synaptosome protein synthesis. J.
Neurochem 32, 1295±1301.
Verity, M.A., Brown, W.J., Cheung, M., 1980. Isolation of ribosome
containing synaptosome subpopulation with active in vitro pro-
tein synthesis. J. Neurosci. Res 5, 143±153.
Verity, M.A., Cheung, M.K., Brown, W.J., 1981. Studies on valino-
mycin inhibition of synaptosome-fraction protein synthesis.
Biochem. J 196, 25±32.
Vial, J.D., 1958. The early changes in the axoplasm during wallerian
degeneration. J. Biophys. Biochem. Cytol 4, 551±556.
Villegas, G.M., Villegas, R., 1984. Squid giant ultrastructure. In:
Baker, P.F. (Ed.), Current Topics in Membranes and Transport,
vol. 22. Academic Press, London, pp. 3±37.
von Bernhardi, R., Alvarez, J., 1989. Is the supply of axoplasmic
proteins a burden for the cell body? Morphometry of sensory ner-
urons and amino acid incorporation into their cell bodies. Brain
Res 478, 301±308.
Waller, A., 1850. Experiments on the section of the glossopharyngeal
and hypoglossal nerves of the frog, and observations of the
alterations produced thereby in the structure of their primitive
®bres. Phil. Trans. Roy. Soc. London Ser. B 140, 423±429.
Walter, P., Johnson, A.E., 1994. Signal sequence recognition and
protein targeting to the endoplasmic reticulum membrane. Annu.
Rev. Cell Biol 10, 87±119.
Waxman, S.G., 1968. Micropinocytic invaginations in the axolemma
of peripheral nerves. Z. Zellforsh 86, 571±573.
Wedege, E., Luqmani, Y., Bradford, H.F., 1977. Stimulated incor-
poration of amino acids into proteins of synaptosomal fractions
induced by depolarizing treatments. J. Neurochem 29, 527±537.
Weiler, I.J., Greenough, W.T., 1991. Potassium ion stimulation trig-
gers protein translation in synaptoneurosomal polyribosomes.
Mol. Cell Neurosci 2, 305±314.
Weiler, I.J., Greenough, W.T., 1993. Metabotropic glutamate recep-
tors trigger postsynaptic protein synthesis. Proc. Natl. Acad. Sci.
USA 90, 7168±7171.
Weiner, O.D., Zorn, A.M., Krieg, P.A., Bittner, G.D., 1996.
Medium-weight neuro®lament mRNA in gold®sh Mauthner axo-
plasm. Neurosci. Lett 213, 83±86.
Weiss, P., Hiscoe, H.B., 1948. Experiments on the mechanism of
nerve growth. J. Exp. Zool 107, 315±393.
Wensley, C.H., Stone, D.M., Baker, H., Kauer, J.S., Margolis, F.L.,
Chikaraishi, D.M., 1995. Olfactory marker protein mRNA is
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±62 61

found in axons of olfactory receptor neurons. J. Neurosci 15,
4827±4837.
White, F.P., McBride, W.J., Mahler, H.R., Moore, W.J., 1972.
Subcellular distribution of proteins synthesized in slices of rat cer-
ebral cortex. J. Biol. Chem 247, 1247±1256.
Whittaker, V.P., 1968. The morphology of fractions of rat forebrain
synaptosomes separated on continuous sucrose density gradients.
Biochem. J 106, 412±417.
Whittaker, V.P., 1993. Thirty years of synaptosome research. J.
Neurocytol 22, 735±742.
Willard, M., Cowan, W.M., Vagelos, P.R., 1974. The poypeptide
composition of intra-axonally transported proteins: evidence for
four transport velocities. Proc. Natl. Acad. Sci. USA 71, 2182±
2187.
Windebank, A.J., Wood, P., Bunge, R.P., Dyck, P.J., 1985.
Myelination determines the caliber of dorsal root ganglion neur-
ons in culture. J. Neurosci 5, 1563±1569.
Winkler, J., Ramirez, G.A., Kuhn, H.G., Peterson, D.A., Day-
Lollini, P.A., Stewart, G.R., Tuszynski, M.H., Gage, F.H., Thal,
L.J., 1997. Reversible Schwann cell hyperplasia and sprouting of
sensory and sympathetic neurites after intraventricular adminis-
tration of nerve growth factor. Ann. Neurol 41, 82±93.
Wyrwicka, W., 1950. On the rate of regeneration of the sciatic nerve
of the white mouse. Acta Biol. Exper 15, 147±153.
Yellin, T.O., Butler, B.J., Stein, H.H., 1967. Inhibition of protein
synthesis in mitochondria by cycloheximide. Fed. Proc 26 (3222),
833.
Yen, T.J., Machlin, P.S., Cleveland, D.W., 1988. Autoregulated
instability of beta-tubulin mRNAs by recognition of the nascent
amino terminus of beta-tubulin. Nature 334, 580±585.
Yin, X., Crawford, T.O., Gri�n, J.W., Tu, P., Lee, V.M.-Y., Li, C.,
Roder, J., Trapp, B.D., 1998. Myelin-associated glycoprotein is a
myelin signal that modeulates the caliber of myelinated axons. J.
Neurosci 18, 1953±1962.
Yokota, R., 1984. Occurrence of long non-myelinated axonal seg-
ments intercalated in myelinated, presumably sensory axons: elec-
tron microscopic observations in the dog atrial endocardium. J.
Neurocytol 13, 127±143.
Young, J.Z., 1974. The central nervous system of Loligo. Part I: The
optic lobe. Philos. Trans. R. Soc. (Biol) 267, 263±302.
Zambetti, G., Fey, E.G., Penman, S., Stein, J., Stein, G., 1990.
Di�erential association of membrane-bound and non-membrane-
bound polysomes with the cytoskeleton. Exp. Cell Res 191, 246±
255.
Zanakis, M.F., Chakraborty, G., Sturman, J.A., Ingoglia, N.A.,
1984. Posttranslational protein modi®cation by amino acid ad-
dition in intact and regenerating axons of the rat sciatic nerve. J.
Neurochem 43, 1286±1294.
Zapata, P., Hess, A., Eyzaguirre, C., 1969. Reinnervation of carotid
body and sinus with superior laryngeal nerve ®bers. J.
Neurophysiol 32, 215±228.
Zelena , J., 1972. Ribosomes in myelinated axons of dorsal root
ganglia. Z. Zellforsch 124, 217±229.
Zelena , J., Jirmanova, I., 1988. Grafts of pacinian corpuscles reinner-
vated by dorsal root axons. Brain Res 438, 165±174.
Zelena , J., Jirmanova, I., Lieberman, A.R., 1990. Reinnervation of
transplanted pacinian corpuscles by ventral root axons: ultra-
structure of the regenerated nerve terminals. J. Neurocytol 19,
962±969.
Zelena , J., Lubinska, L., Gutmann, E., 1968. Accumulation of orga-
nelles at the ends of interrupted axons. Z. Zellforsch. Mikrosk.
Anat 91, 200±219.
Zenker, W., Hohberg, E., 1973. A-a-nerve-®bre: number of neurotu-
bules in the stem ®bre and in the terminal branches. J.
Neurocytol 2, 143±148.
Zimmerman, U.J., Schlaepfer, W.W., 1982. Characterization of a
brain calcium-activated protease that degrades neuro®lament pro-
teins. Biochemistry 21, 3977±3982.
Ziv, N.E., Spira, M.E., 1993. Spatiotemporal distribution of Ca2+
following axotomy and throughout the recovery process of cul-
tured Aplysia neurons. Eur. J. Neurosci 5, 657±668.
Ziv, N.E., Spira, M.E., 1997. Localized and transient elevations o®n-
tracellular Ca2+ induce the dedi�erentiation of axonal segments
into growth cones. J. Neurosci 17, 3568±3579.
Ziv, N.E., Spira, M.E., 1998. Induction of growth cone formation by
transient and localized increases of intracellular proteolytic ac-
tivity. J. Cell Biol 140, 223±232.
Zottoli, S.J., Marek, L.E., Agostini, M.A., Strittmatter, S.L., 1987.
Morphological and physiological survival of gold®sh Mauthner
axons isolated from their somata by spinal cord crush. J. Comp.
Neurol 255, 272±282.
Zuo, J., Ferguson, T.A., Hernandez, Y.J., Stetler-Stevenson, W.G.,
Muir, D., 1998a. Neuronal matrix metalloproteinase-2 degrades
and inactivates a neurite-inhibiting chondroitin sulfate proteogly-
can. J. Neurosci 18, 5203±5211.
Zuo, J., Hernandez, Y.J., Muir, D., 1998b. Chondroitin sulfate pro-
teoglycan with neurite-inhibiting activity is up-regulated following
peripheral nerve injury. J. Neurobiol 34, 41±54.
J. Alvarez et al. / Progress in Neurobiology 62 (2000) 1±6262
Copyright © 2022 FDOKUMEN




















