
Plasticity of Somatosensory Cortex in Primates
10
Plasticity of Somatosensory Cortex in Primates Sherre L. Florence, 1 Neeraj Jain, and Jon H. Kaas Department of Psychology, Vanderbilt University, Nashville Tennessee 37240 Over recent years, we have come to the surprising realization that sensory cortex is highly plastic in functional organization, even in adult brains. Much of the evidence for this conclusion comes from studies of the effects of peripheral deafferentation or sensory experience on the somatotopy of primary somatosensory cortex (area 3b) of monkeys. Local modifications in cortical representations occur rapidly after sensory loss or more gradually during altered sensory experience. These changes depend on reductions in lateral inhibition and other dynamic adjustments in sensory networks, as well as Hebbian-like modifications of synaptic strengths. Activity-dependent alterations in the expression of neurotransmitters and modulators may also play a role. After major deactivations, such as those produced by amputation or section of dorsal column afferents, cortex regains responsiveness over a much longer time period as a result, at least in part, from the growth of new connections. r 1997 Academic Press KEY WORDS: Cortical topography; cuneate nucleus; peripheral nerve transection; spinal cord injury; ventroposterior nucleus. The developing brain is extremely plastic. Most notably, the organization of sensory systems can be greatly altered early in development by injury to part of the pathway or by sensory deprivation. In contrast, some of the manipulations that have great impact during development have little effect in the adult brain (1). Therefore, the mature brain has been considered to be rather stable. Yet, recovery from brain damage, alterations of sensory inputs, and the acquisition of new skills and abilities must accompany changes in the adult brain. Thus, the recent evidence of the mutability of sensory representations in mature cortex has had considerable impact. Much of the research has involved primary somatosensory cortex, where the normal representation of the contralateral body sur- face can be mapped in great detail with microelec- trodes, and even minor changes in topography can be appreciated. Manipulations of a small portion of the sensory inputs alter the normal maps in minor ways, so that little or no change in the structural framework of the brain would seem to be involved. Instead, local modifications in synaptic efficacy and the dynamic balance of circuits could adequately account for the new topographies. In contrast, some manipulations that deactivate large parts of somatosensory cortex are followed by massive reactivations, changes that seem too large to be mediated by previously existing cir- cuitry, and thus would require the growth of new connections. New growth is unexpected for the mature brain, but there is now clear evidence that new growth does occur under some conditions. In this review, we discuss the reorganization of somatosensory cortex that follows nerve and spinal cord injury, both because the reorganizations are more massive and more obvious after such deprivations and also because these larger modifications of cortex may yield insights as to how and when new connections form in a damaged nervous system, and if they can be functionally useful. While our emphasis is on cortex, where sensory maps are large and often conveniently exposed on the brain surface so that they are easy to explore, some cortical reorganization may result from modifications that occur earlier in the processing sequence. Thus, we also consider subcortical modifica- tions of sensory maps. We have concentrated on studies in primates because they are especially rel- evant to the human brain. 1 To whom correspondence and reprint requests should be ad- dressed. Fax: 615-343-8449. Email: fl[email protected]. Seminars in NEUROSCIENCE 9, 3–12 (1997) Article No. SN970101 1044-5765/97 $25.00 Copyright r 1997 by Academic Press All rights of reproduction in any form reserved. 3
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Plasticity of Somatosensory Cortex in Primates

Plasticity of Somatosensory Cortex in Primates
Sherre L. Florence,1 Neeraj Jain, and Jon H. KaasDepartment of Psychology, Vanderbilt University, Nashville Tennessee 37240
Over recent years, we have come to the surprising realization that sensory cortex is highly plastic infunctional organization, even in adult brains. Much of the evidence for this conclusion comes fromstudies of the effects of peripheral deafferentation or sensory experience on the somatotopy of primarysomatosensory cortex (area 3b) of monkeys. Local modifications in cortical representations occurrapidly after sensory loss or more gradually during altered sensory experience. These changes dependon reductions in lateral inhibition and other dynamic adjustments in sensory networks, as well asHebbian-like modifications of synaptic strengths. Activity-dependent alterations in the expression ofneurotransmitters and modulators may also play a role. After major deactivations, such as thoseproduced by amputation or section of dorsal column afferents, cortex regains responsiveness over amuch longer time period as a result, at least in part, from the growth of new connections. r 1997 Academic
Press
KEY WORDS: Cortical topography; cuneate nucleus; peripheral nerve transection; spinal cord injury;ventroposterior nucleus.
The developing brain is extremely plastic. Mostnotably, the organization of sensory systems can begreatly altered early in development by injury to partof the pathway or by sensory deprivation. In contrast,some of the manipulations that have great impactduring development have little effect in the adult brain(1). Therefore, the mature brain has been considered tobe rather stable. Yet, recovery from brain damage,alterations of sensory inputs, and the acquisition ofnew skills and abilities must accompany changes inthe adult brain. Thus, the recent evidence of themutability of sensory representations in mature cortexhas had considerable impact. Much of the research hasinvolved primary somatosensory cortex, where thenormal representation of the contralateral body sur-face can be mapped in great detail with microelec-trodes, and even minor changes in topography can beappreciated. Manipulations of a small portion of thesensory inputs alter the normal maps in minor ways,so that little or no change in the structural frameworkof the brain would seem to be involved. Instead, localmodifications in synaptic efficacy and the dynamic
balance of circuits could adequately account for thenew topographies. In contrast, some manipulationsthat deactivate large parts of somatosensory cortex arefollowed by massive reactivations, changes that seemtoo large to be mediated by previously existing cir-cuitry, and thus would require the growth of newconnections. New growth is unexpected for the maturebrain, but there is now clear evidence that new growthdoes occur under some conditions.In this review, we discuss the reorganization of
somatosensory cortex that follows nerve and spinalcord injury, both because the reorganizations are moremassive and more obvious after such deprivations andalso because these larger modifications of cortex mayyield insights as to how and when new connectionsform in a damaged nervous system, and if they can befunctionally useful. While our emphasis is on cortex,where sensory maps are large and often convenientlyexposed on the brain surface so that they are easy toexplore, some cortical reorganization may result frommodifications that occur earlier in the processingsequence. Thus, we also consider subcortical modifica-tions of sensory maps. We have concentrated onstudies in primates because they are especially rel-evant to the human brain.
1To whom correspondence and reprint requests should be ad-dressed. Fax: 615-343-8449. Email: [email protected].
Seminars in NEUROSCIENCE 9, 3–12 (1997)
Article No. SN970101
1044-5765/97 $25.00Copyright r 1997 by Academic PressAll rights of reproduction in any form reserved. 3
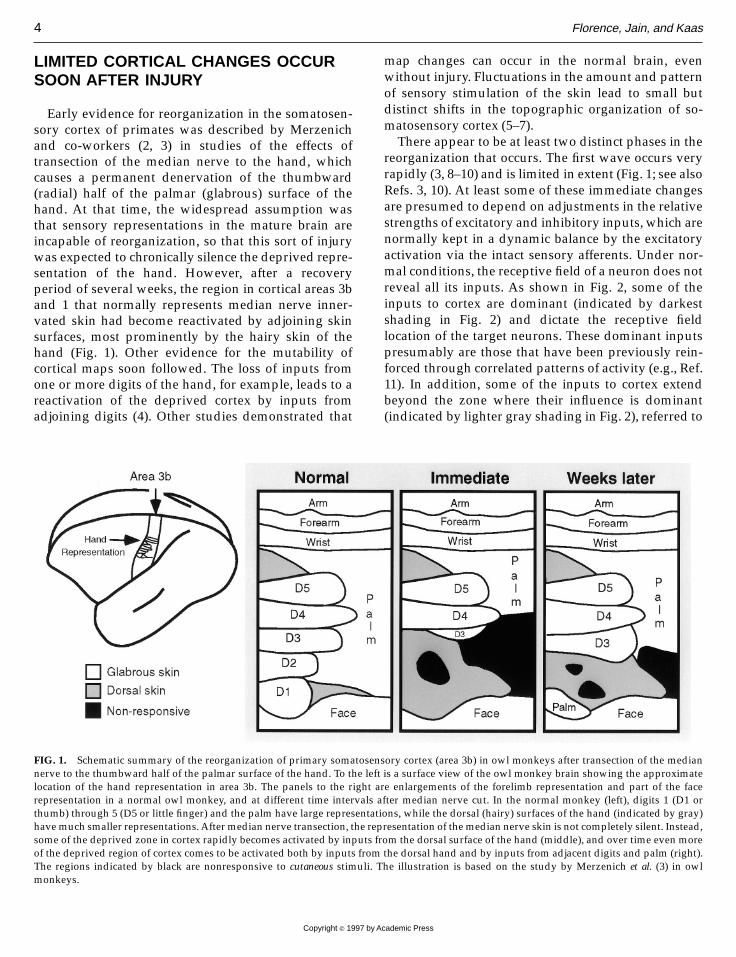
LIMITED CORTICAL CHANGES OCCURSOON AFTER INJURY
Early evidence for reorganization in the somatosen-sory cortex of primates was described by Merzenichand co-workers (2, 3) in studies of the effects oftransection of the median nerve to the hand, whichcauses a permanent denervation of the thumbward(radial) half of the palmar (glabrous) surface of thehand. At that time, the widespread assumption wasthat sensory representations in the mature brain areincapable of reorganization, so that this sort of injurywas expected to chronically silence the deprived repre-sentation of the hand. However, after a recoveryperiod of several weeks, the region in cortical areas 3band 1 that normally represents median nerve inner-vated skin had become reactivated by adjoining skinsurfaces, most prominently by the hairy skin of thehand (Fig. 1). Other evidence for the mutability ofcortical maps soon followed. The loss of inputs fromone or more digits of the hand, for example, leads to areactivation of the deprived cortex by inputs fromadjoining digits (4). Other studies demonstrated that
map changes can occur in the normal brain, evenwithout injury. Fluctuations in the amount and patternof sensory stimulation of the skin lead to small butdistinct shifts in the topographic organization of so-matosensory cortex (5–7).There appear to be at least two distinct phases in the
reorganization that occurs. The first wave occurs veryrapidly (3, 8–10) and is limited in extent (Fig. 1; see alsoRefs. 3, 10). At least some of these immediate changesare presumed to depend on adjustments in the relativestrengths of excitatory and inhibitory inputs, which arenormally kept in a dynamic balance by the excitatoryactivation via the intact sensory afferents. Under nor-mal conditions, the receptive field of a neuron does notreveal all its inputs. As shown in Fig. 2, some of theinputs to cortex are dominant (indicated by darkestshading in Fig. 2) and dictate the receptive fieldlocation of the target neurons. These dominant inputspresumably are those that have been previously rein-forced through correlated patterns of activity (e.g., Ref.11). In addition, some of the inputs to cortex extendbeyond the zone where their influence is dominant(indicated by lighter gray shading in Fig. 2), referred to
FIG. 1. Schematic summary of the reorganization of primary somatosensory cortex (area 3b) in owl monkeys after transection of the mediannerve to the thumbward half of the palmar surface of the hand. To the left is a surface view of the owl monkey brain showing the approximatelocation of the hand representation in area 3b. The panels to the right are enlargements of the forelimb representation and part of the facerepresentation in a normal owl monkey, and at different time intervals after median nerve cut. In the normal monkey (left), digits 1 (D1 orthumb) through 5 (D5 or little finger) and the palm have large representations, while the dorsal (hairy) surfaces of the hand (indicated by gray)havemuch smaller representations.After median nerve transection, the representation of themedian nerve skin is not completely silent. Instead,some of the deprived zone in cortex rapidly becomes activated by inputs from the dorsal surface of the hand (middle), and over time even moreof the deprived region of cortex comes to be activated both by inputs from the dorsal hand and by inputs from adjacent digits and palm (right).The regions indicated by black are nonresponsive to cutaneous stimuli. The illustration is based on the study by Merzenich et al. (3) in owlmonkeys.
4 Florence, Jain, and Kaas
Copyright r 1997 by Academic Press

as a subthreshold ‘‘fringe.’’ At least some of the inputsin this fringe region rapidly come to be expressed aftermanipulations, such as injury, because of a reductionof the excitatory drive of inhibitory cortical neurons,called afferent driven inhibition (12). Other inputs inthe subthreshold fringe may not have sufficient synap-tic strength to activate cortical neurons immediatelyafter inhibition is reduced. Instead, these inputs maycome to be expressed over time if the synaptic strengthof the inputs is reinforced (see below).An additional, but less well understood, influence
on rapidly occurring synaptic plasticity is a modifica-tion of the effectiveness of excitatory inputs by thebehavioral state of the animal (13). The level of atten-tion devoted to a task during its execution appears tobe strongly correlated with the degree of change thatcan be produced in cortex by the sensory stimulation(6, 14). Perhaps, attention and other motivationalfactors, including noxious stimuli, alter the release ofneuromodulators that affect local circuits.Over the course of days and weeks subsequent to
sensory deprivation, a second type of reorganizationoccurs (3, 10). This second phase in cortical plasticityleads to somewhat more extensive map reorganization(Fig. 1) as well as to what appears to be a progressive‘‘fine-tuning’’ of the topographic order within reorga-nized cortex (3). The substrate for this more slowlydeveloping plasticity probably involves Hebbian-typemechanisms for synaptic strengthening of previouslyexisting, latent inputs (11). Presumably, the inputs arepart of the subthreshold fringe schematized in Fig. 2,and they may comprise a wider fringe area than thatoccupied by the inputs that can be expressed followinga reduction of intracortical inhibition. The synapticstrengthening is thought to be based on correlations inthe pattern of afferent-driven activity and appears toinvolve the NMDA receptor complex, since only lim-ited reorganization occurs in somatosensory cortex ifNMDA receptors are blocked during the recoveryperiod (15, 16). Additional slowly evolving changescould result from alterations in the expression ofmolecules involved in the synaptic circuitry. For ex-ample, after injury there appears to be a down-regulation of inhibitory neurotransmitters (17) so thatthe impact of even weak excitatory inputs will bepotentiated. Also, the local release of acetylcholine isthought to be an important factor in experience-basedplasticity (18–20).Although it has become apparent that sensory repre-
sentations in the adult brain are dynamically main-tained, rather than being stable as it had previouslybeen assumed, it was not clear from the earlier demon-strations of adult cortical plasticity how changes insensory maps impact on sensory function. Evidencethat disordered sensory representations correlate withpoor sensory function indicates that there is an impor-tant relationship between the cortical maps and sen-sory perception. In individuals born with webbedfingers, called syndactyly, the cortical representation ofthe webbed fingers is compressed and topographicallyabnormal, and these individuals apparently do nothave a perception of the individual, albeit fused,fingers (21). However, within a week after surgical
FIG. 2. Schematic depiction of the proposed topographic distribu-tion of thalamic afferents to layer IV of primary somatosensorycortex (area 3b). The evidence for this proposal is, first, thatindividual thalamic axons have a wider projection zone than isreflected in the topographic map in cortex under normal conditions(44, 70). Second, after manipulations that reduce the effectiveness ofinhibitory neurotransmitters, a larger region of excitatory activationis expressed in cortex (12). Therefore, thalamic afferents are thoughtto have a subthreshold fringe of influence. In this example, thedominant influence (indicated by dark gray shading) of two thalamo-cortical afferents representing digit 3 (D3) is within the D3 represen-tation, and their subthreshold fringes (indicated by lighter grayshading) each extend into one of the adjacent digit representations,so that if the levels of inhibition are reduced, by injury for example,the representation of D3 could expand immediately into the adjacentdigit representation. Similar topographic changes likely result fromsynaptic strengthening that results from use-dependent changes, butmay require days to weeks to develop.
Plasticity of Somatosensory Cortex 5
Copyright r 1997 by Academic Press

separation of the fingers, the map of the hand in cortexachieved a more normal organization, and in at leastone of the two individuals studied, separate fingerscould be perceived for the first time (21).The significance of cortical topography for sensory
function also is apparent from studies of the effects ofsensory nerve transection followed by surgical repairand regeneration. Such an injury in adult humansproduces persisting functional abnormalities, includ-ing sensory mislocalizations and poor two-point touchdiscrimination (22). In adult monkeys, the topographicorder of the reinnervated skin of the hand in somato-sensory cortex is markedly abnormal (23, 24), and thecortical disorder is the likely source of the sensorydisturbances. In contrast, nerve transection, repair, andregeneration in children leaves only mild sensorydisturbances (22), and in monkeys that had nerve cutand repair either in fetal or early postnatal life, topo-graphic organization of somatosensory cortex is muchmore normal than after similar injuries in adults (25).Although indirect, these correspondences indicate thatnormal sensory function is critically dependent on theorderliness of cortical representations.The most detailed study of the relationship between
sensory performance and cortical map changes wasdone in monkeys trained to detect small differences inthe flutter-vibration rate of tactile stimuli by Recan-zone and colleagues (6, 14). In individual monkeys,improvements in discrimination performance pro-duced by the training were correlated with changes invarious aspects of the response of cortical neurons,including receptive field size and overlap, temporalresponse, and the overall size of the representation ofthe portion of the hand used in the detection task.Although there were indications of weak correlationsbetween the improvement in behavioral performanceand some of response properties, none of the physi-ological changes could completely account for thesensory improvement. Therefore, Recanzone et al. (14)concluded that the improvements in sensory functionresult from changes in multiple response propertiesover a distributed neural network.
CORTICAL CHANGES MAY REFLECTSUBCORTICAL MODIFICATIONS
Originally, only the higher centers of the brain werethought to have the potential for plasticity. The spinalcord, brain stem, and thalamus were thought to havelittle adaptive flexibility (26) and were viewed assimple relays of afferent activity from the periphery to
cortex. Consistent with these well-entrenched notions,the early descriptions of change in somatosensorycortex were interpreted as cortical in origin (2). Subcor-tical reorganization produced by sensory deprivationhad been reported in rats and cats (for review see Refs.27, 28); however, the reported changes were typicallysmall and there were disagreements about the interpre-tation of the findings (29). However, it has now becomeabundantly clear that topographic reorganization canoccur at all levels of the somatosensory pathway.In the spinal cord, most of the recent evidence for
plasticity is from studies in cats following sensorymanipulations which deprive cells in the dorsal hornof most or all dominant excitatory inputs (e.g., Refs. 30,31). Over time, neurons in the deprived portion of thedorsal horn respond to previously unexpressed inputs,presumably through mechanisms much like thosedescribed above for cortex.Also, in the gracile and cuneate nuclei of the brain
stem, which contain representations of the lower andupper extremities, respectively, dorsal root transec-tions lead to topographic changes (32–34). However,the effects of less severe denervations appear to bespecies variable. For example, in cats, subcutaneousinjections of a local anesthetic are rapidly followed bythe appearance of a new receptive field for cuneateneurons that previously responded to the anesthetizedskin (35). In raccoons, no receptive field changes wereproduced using the same approach (36). Neurons inthe dorsal column nuclei of rats show receptive fieldchanges only after dorsal root transection, but not aftersmaller denervations such as those produced by nervecut (34).There appears to be much more conclusive evidence
for plastic changes in the somatosensory thalamus,although extensive comparisons of the extent and timecourse of the changes across species have not beenmade. The general conclusion from all studies to dateis that there is remarkable potential for change in thethalamus as a result of peripheral denervations (inmonkeys: Refs. 37, 38; in raccoons: Refs. 39–41; and inrats: Refs. 9, 42).Few studies had tested directly whether the subcor-
tical changes could substantially affect cortical organi-zation. Recently, however, it has become apparent thatsome of the topographic reorganization in cortex isprobably caused by changes at one or more of thesubcortical somatosensory nuclei. For example, insquirrel monkeys, if one or more nerves to the glabroussurface of the hand are transected, leaving intact onlythe innervation to the hairy surface of the hand, thedeprived zone in cortex comes to be completely reacti-
6 Florence, Jain, and Kaas
Copyright r 1997 by Academic Press

vated by inputs from hairy hand (43). The width of thereorganized zone in cortex is over 3 mm, larger thanwould be expected if the reorganization were based onthe extent of overlap of thalamocortical arbors in thesemonkeys (44). However, in the cuneate nucleus wherethe hand representation is compressed, inputs fromthe glabrous and hairy surfaces of the hand are directlyadjacent to one another (45, 46). Consequently, small(on the order 50 µm) adjustments in the strength ofexisting inputs in the brain stem could allow for theobserved topographic shifts in cortex (see also Ref. 47).A second example of the relationship between subcor-tical changes and those found in cortex is that thepattern of reorganization in cortex after nerve sectionis very similar to that found in the ventroposteriornucleus of the thalamus (38, 43), and we have postu-lated that such reorganization reflects synaptic modifi-cations that occur even earlier in the pathway, at thelevel of the cuneate nucleus of the brain stem (48).Thus, topographic reorganizations based on local ad-justments in synaptic strengths that occur amongnear-neighbors subcortically can appear as relativelylarge reorganizations in cortex.
NEW AXON GROWTH MAY FOLLOWMASSIVE INJURY
Since most of the early evidence for map reorganiza-tion could be explained by local modifications within adynamic synaptic framework, there was no indicationthat the connectional organization of the somatosen-sory pathway changed. The assumption that newconnections do not form in the adult brain wasreinforced by results from studies of the effects of moreextensive denervations. For example, after the loss ofseveral digits, the denervated zone in cortex typicallyis not completely reactivated by other inputs, and asilent zone in cortex may persevere for months (4).These results were taken to indicate that reactivation islimited by the overlap of existing connections so thatcortical reorganization occurs only near the borders ofthe deprived zone.The reactivation of larger zones of denervated cortex
was considered unlikely in adults because this wouldinvolve substantial growth of new connections, andextensive new growth was thought to occur only in thedeveloping brain. The assumption that large areas ofcortex could not be reactivated became untenable afew years ago when Pons and co-workers (49) usedmicroelectrode mapping to study the topographicorganization of somatosensory cortex in monkeys that
had a long-standing loss of all sensory inputs from theentire forelimb by a procedure known as ‘‘dorsalrhizotomy.’’ They discovered that the large zone ofcortex where the entire forelimb would normally berepresented had been completely reactivated by inputsfrom the face, an expansion of such massive propor-tions that it could not be explained by mechanismsinvolving previously existing connections and localadjustments in synaptic efficacy. Thus, Pons et al. (49)postulated that new connections formed somewhere inthe somatosensory system, but there was no directevidence to support this assumption.We now have found that such massive reorganiza-
tions involve the growth of new connections in thebrain stem where the body representation is quitecompact (50). In monkeys that had amputations ofeither a hand or forearm many years previously asveterinary treatment for traumatic injury to the arm,we first looked for evidence of reorganization inprimary somatosensory cortex. We found that most ofthe deprived hand representation had regained respon-siveness to the remaining portion of the amputatedarm (Fig. 3). In addition, there appeared to be someexpansion of the face representation into the lateralportion of the deprived hand representation in cortex.In these same animals, we also studied the anatomicaldistribution of afferent inputs from the skin of the arminto the cuneate nucleus of the brain stem, since thetopography of these inputs appears to have a signifi-cant impact on topographic organization in cortex (24).The results showed that the axon terminals of thenerves fibers from the remaining portion of the ampu-tated arm were no longer confined to small zones atthe border of the cuneate nucleus where they arelocated in normal animals (50). Instead, the axonterminals of the nerves from the arm had sprouted adistance of 1–2 mm into the region where inputs fromthe hand normally terminate; as a result neurons thatoriginally responded to stimulation of the hand couldbe activated by the arm. While this anatomical changewas relatively limited at the level of the brain stem, thenew patterns of activation produced by the changewould be magnified as the information was relayed tocortex. The impact on cortical topography is presumedto be the remarkable expansion of the representation ofthe arm in cortex.Given the evidence that massive cortical reorganiza-
tion can occur, and that it can be based on the growthof new connections, a reevaluation of the effects ofdorsal spinal cord injury was needed. Transection ofthe dorsal columns, the predominant source of tactileinformation to somatosensory cortex, should have a
Plasticity of Somatosensory Cortex 7
Copyright r 1997 by Academic Press

major impact on the responsiveness and function ofsomatosensory cortex. Yet, behavioral defects afterdorsal column section have sometimes been reportedas mild or transitory (for review, see Ref. 51). The fewpersisting deficits include a loss of temporal resolution
(52) and loss of the ability to discriminate direction ofmovement of the stimulus on the skin (53). The effectsof dorsal column section on the response properties ofneurons in somatosensory cortex have been variouslydescribed as a small to marked decrease in the number
FIG. 3. Demonstration of the effect of long-standing amputation of the hand in macaque monkeys. The normal distribution of sensory inputsfrom the forelimb to the cuneate nucleus is very orderly; inputs from the hand terminate in the ventral portion of the nucleus, as shown in theenlargement of the cuneate nucleus on the left. Inputs from the arm and forearm are restricted to the dorsal extreme of the cuneate nucleus(outlined by the thin black lines in the cuneate nucleus on the left). However, after hand amputation, the inputs from the arm and forearm sproutinto the region where inputs from the hand are normally located (enlargement on the right), and as a result, the target neurons in what wasformerly the hand representation of the cuneate nucleus come to respond to stimulation of the forearm. This abnormal activation patternpresumably is relayed through the thalamus to area 3b (shown at the top right) so that inputs to cortex that formerly were activated only whenthe hand was touched come to be activated when the arm is touched. The perceptual effect of these anatomical and physiological changes is thattactile stimulation of the amputated stump will produce neuronal activity that is perceived as on the hand as well as the arm (shown on the‘‘monkunculus’’ at the top right) and will cause phantom sensations. Revised from Kaas and Florence (71).
8 Florence, Jain, and Kaas
Copyright r 1997 by Academic Press

of neurons that respond to light cutaneous stimuli(54–56). The issue of cortical plasticity has been givenlittle attention in these studies.To test whether there is large-scale reorganization
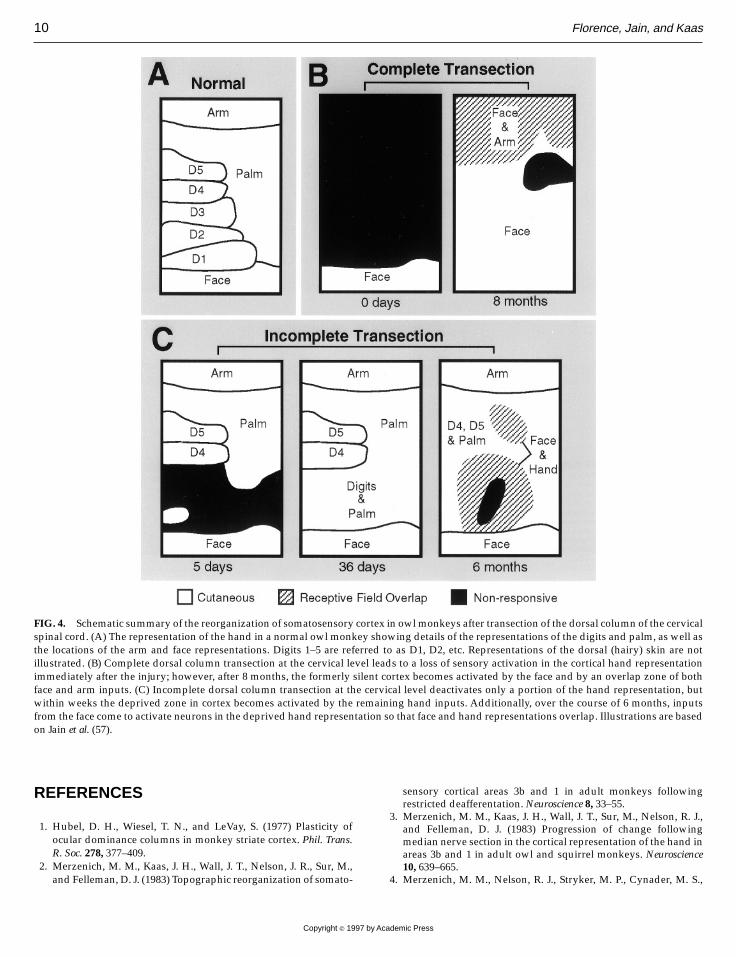
after dorsal column transection, the topographic orga-nization of cortical area 3b was studied in monkeysimmediately after transection of the dorsal columns atthe C3/C4 level of the cervical spinal cord and atvarious times up to 8months later (57).Additionally, toevaluate whether the spinal cord lesions transected allthe ascending fibers in the dorsal columns, the skin ofthe hand was injected with tracers that are carried bythe sensory afferents in the hand to their targets in thespinal cord and brain stem nuclei. Since the cervicallesions were high enough to disrupt the afferents fromthe hand as well as the lower body, the tracer from thehand injections would not be transported to the brainstem nuclei if the lesions were complete. Such a controlwas essential since incomplete transection of the dor-sal columns would be expected to have a muchdifferent impact on the organization of area 3b than thecomplete removal of all dorsal column inputs. Indeed,this was found to be a very important distinction. Inmonkeys with incomplete dorsal spinal cord transec-tion, the remaining inputs activated not only theneurons in the expected cortical locations, but alsosoon came to activate neurons that were driven byother inputs prior to the lesion (Fig. 4C). By about 5weeks after an incomplete cervical spinal cord lesion,the deprived zone in cortex had been reactivated;however, the representation of the hand was com-pletely abnormal. There was a markedly expandedand abnormal representation of the ulnar edge of thehand—the portion of the hand that had not beendeactivated by the lesion (Fig. 4C). A second, lateroccurring pattern of cortical reorganization was evenmore remarkable in the monkeys with incompletedorsal column transection. After a period of about 6months, some neurons in the cortical region thatoriginally contained the hand representation re-sponded not only to stimulation of the few remaininginputs from the hand (the reactivation that occurredwithin weeks of the injury), but also to stimulation ofthe face (Fig. 4C).In cases with total elimination of all dorsal column
inputs from the hand, Jain et al. (57) found that neuronsthroughout the entire region of deprived cortex wereinitially unresponsive to tactile stimuli, but over aperiod of months, these neurons developed responsesto light touch on the face (Fig. 4B). This type ofreactivation would seem to depend on the growth ofnew connections so that axons related to the face
innervated structures formerly related to the hand. Alikely place for this to happen is in the cuneate nucleus,with axons from the trigeminal system sprouting intothe cuneate nucleus. However, there is yet no directevidence for this type of new growth.An important future goal is to understand the
mechanisms that allow for new growth, so that desir-able growth can be facilitated. Previously, new growthin the adult central nervous system has been demon-strated for the central terminations of injured periph-eral nerves (e.g., Refs. 58, 59). Direct injury may triggerthe up-regulation of genes that are normally associatedwith developmental growth (i.e., GAP-43), thus result-ing in new growth (60). However, our finding of newgrowth in the cuneate nucleus after amputation wasnot produced by axons that had been directly injured(50). Indeed, subsequent research has shown thatneurons can sprout new processes without being‘‘induced’’ to grow by injury (61). What could be thesource of triggers other than direct injury to the neuronthat can lead to the extension of long axonal processes?It is unlikely that growth factors produced by theinjured neurons are available to noninjured neighbors,since naive (uninjured) neurons grown in culture withpreviously injured cells do not produce long axonextensions (61). Perhaps, vacant synaptic sites on thepostsynaptic neurons are the source of a trigger for thenew growth.Also, the production of growth inhibitors,which normally suppress neurite growth (62), may bedown-regulated or blocked after injury, so that thegrowth of new connections is no longer prevented.Another important goal is to understand the behav-
ioral consequences of new axon growth. After injuriessuch as amputation, the changes in primary somatosen-sory cortex likely contribute to abnormal ‘‘phantomlimb’’ sensations (63, 64). Such phantoms also occurafter spinal cord injuries (65). Noninvasive imagingtechniques, used to observe the pattern of corticalactivity in patients who have had hand or arm amputa-tions, have shown that there is a cortical expansion ofthe face and arm representations into the deprivedhand representation (66–68). Presumably, activation ofthe cortical neurons in the location where the handwasrepresented prior to the injury is being interpreted as asensation on the hand, even though the source of theexcitatory activitymight be a touch to the upper arm orface (see, for example, Fig. 3). These abnormal sensa-tions can be very disturbing and quite painful (69).Thus, an important direction for future research is tolearn not only how to promote brain reorganization, ifsuch an outcome is desirable, but also how to control itif such an outcome is detrimental.
Plasticity of Somatosensory Cortex 9
Copyright r 1997 by Academic Press
REFERENCES
1. Hubel, D. H., Wiesel, T. N., and LeVay, S. (1977) Plasticity ofocular dominance columns in monkey striate cortex. Phil. Trans.R. Soc. 278, 377–409.
2. Merzenich, M. M., Kaas, J. H., Wall, J. T., Nelson, J. R., Sur, M.,and Felleman, D. J. (1983) Topographic reorganization of somato-
sensory cortical areas 3b and 1 in adult monkeys followingrestricted deafferentation.Neuroscience 8, 33–55.
3. Merzenich, M. M., Kaas, J. H., Wall, J. T., Sur, M., Nelson, R. J.,and Felleman, D. J. (1983) Progression of change followingmedian nerve section in the cortical representation of the hand inareas 3b and 1 in adult owl and squirrel monkeys. Neuroscience10, 639–665.
4. Merzenich, M. M., Nelson, R. J., Stryker, M. P., Cynader, M. S.,
FIG. 4. Schematic summary of the reorganization of somatosensory cortex in owlmonkeys after transection of the dorsal column of the cervicalspinal cord. (A) The representation of the hand in a normal owl monkey showing details of the representations of the digits and palm, as well asthe locations of the arm and face representations. Digits 1–5 are referred to as D1, D2, etc. Representations of the dorsal (hairy) skin are notillustrated. (B) Complete dorsal column transection at the cervical level leads to a loss of sensory activation in the cortical hand representationimmediately after the injury; however, after 8 months, the formerly silent cortex becomes activated by the face and by an overlap zone of bothface and arm inputs. (C) Incomplete dorsal column transection at the cervical level deactivates only a portion of the hand representation, butwithin weeks the deprived zone in cortex becomes activated by the remaining hand inputs. Additionally, over the course of 6 months, inputsfrom the face come to activate neurons in the deprived hand representation so that face and hand representations overlap. Illustrations are basedon Jain et al. (57).
10 Florence, Jain, and Kaas
Copyright r 1997 by Academic Press

Schoppmann, A., and Zook, J. M. (1984) Somatosensory corticalmap changes following digit amputation in adult monkeys. J.Comp. Neurol. 224, 591–605.
5. Jenkins, W. M., Merzenich, M. M., Ochs, M. T., Allard, T., andGuic-Robles, E. (1990) Functional reorganization of primarysomatosensory cortex in adult owl monkeys after behaviorallycontrolled tactile stimulation. J. Neurophysiol. 63, 82–104.
6. Recanzone, G. H., Merzenich, M. M., Jenkins, W. M., Grajski,K. A., and Dinse, H. R. (1992) Topographic reorganization ofthe hand representation in cortical area 3b of owl monkeystrained in a frequency-discrimination task. J. Neurophysiol. 67,1031–1057.
7. Wang, X., Merzenich, M. M., Sameshima, K., and Jenkins, W. M.(1995) Remodeling of hand representation in adult cortex deter-mined by timing of tactile stimulation.Nature 378, 71–75.
8. Calford, M. B., and Tweedale, R. (1991) Acute changes incutaneous receptive fields in primary somatosensory cortex afterdigit denervation in adult flying fox. J. Neurophysiol. 65, 178–187.
9. Nicolelis, M. A., Lin, R. C. S., Woodward, D. J., and Chapin, J. K.(1993) Induction of immediate spatiotemporal changes in tha-lamic networks by peripheral block of ascending cutaneousinformation.Nature 361, 533–536.
10. Silva, A. C., Rasey, S. K., Wu, X., and Wall, J. T. (1996) Initialcortical reactions to injury of the median and radial nerves to thehands of adult primates. J. Comp. Neurol. 366, 700–716.
11. Stent, G. S. (1973) A physiological mechanism for Hebb’spostulate of learning. Proc. Natl. Acad. Sci. USA 70, 997–1001.
12. Alloway, K., and Burton, H. (1991) Differential effects of GABAand bicuculline on rapidly and slowly-adapting neurons inprimary somatosensory cortex of primates. Exp. Brain Res. 85,598–610.
13. Castro-Alamancos, M. A., and Connors, B. W. (1996) Short-termplasticity of a thalamocortical pathway dynamically modulatedby behavioral state. Science 272, 274–277.
14. Recanzone, G. H., Merzenich, M. M., and Jenkins, W. M. (1992)Frequency discrimination training engaging a restricted skinsurface results in emergence of a cutaneous response zone incortical area 3a. J. Neurophysiol. 67, 1057–1070.
15. Kano, M., Lino, K., and Kano, M. (1991) Functional reorganiza-tion of adult cat somatosensory cortex is dependent on NMDAreceptors.NeuroReport 2, 77–80.
16. Garraghty, P. E., and Muja, N. (1996) NMDA receptors andplasticity in adult primate somatosensory cortex. J. Comp. Neurol.367, 319–326.
17. Garraghty, P. E., LaChica, E. A., and Kaas, J. H. (1991) Injury-induced reorganization of somatosensory cortex is accompaniedby reductions in GABA staining. Somat. Motor Res. 8, 347–354.
18. Weinberger, N. M. (1997) Learning effects in auditory cortex.Semin. Neurosci. 9.
19. Cusick, C. G. (1991) Nerve injury-induced depletion of tachyki-nin immunoreactivity in the somatosensory cortex of adultsquirrel monkeys. Brain Res. 568, 314–318.
20. Avendano, C., Umbriaco, D., Dykes, R. W., and Descarries, L.(1995) Decrease and long-term recovery of choline acetyltransfer-ase immunoreactivity in adult cat somatosensory cortex afterperipheral nerve transections. J. Comp. Neurol. 354, 321–332.
21. Mogliner, A., Grossman, J. A. I., Ribary, U., Joliot, M., Volkmann,J., Rapaport, D., Beasely, R. W., and Llinas, R. R. (1993) Somato-sensory cortical plasticity in adult humans revealed bymagneto-encephalography. Proc. Natl. Acad. Sci. USA 90, 3593–3597.
22. Dellon, A. L. (1981) Evaluation of sensibility and re-education ofsensation in the hand. Williams &Wilkins, Baltimore.
23. Paul, R. L., Goodman, H., and Merzenich, M. M. (1972) Alter-ations in mechanoreceptor input to Brodmann’s areas 1 and 3 ofthe postcentral hand area of Macaca mulatta after nerve sectionand regeneration. Brain Res. 39, 1–19.
24. Florence, S. L., Garraghty, P. E., Wall, J. T., and Kaas, J. H. (1994)Sensory afferent projections and area 3b somatotopy followingmedian nerve cut and repair in macaque monkeys. CerebralCortex 4, 391–407.
25. Florence, S. L., Jain, N., Pospichal, M. W., Beck, P. D., Sly, D. L.,and Kaas, J. H. (1996) Central reorganization of sensory path-ways following peripheral nerve regeneration in fetal monkeys.Nature 381, 69–71.
26. Sperry, R. W. (1959) The growth of nerve circuits. Sci. Am. 201,68–75.
27. Wall, P. D. (1981) The nature and origins of plasticity in adultspinal cord. In Spinal Cord Sensation (Brown, A. G., andRethelyi, M., Eds.), pp. 297–308, Scottish Academic Press, Edin-burgh.
28. Devor, M. (1983) Plasticity of spinal cord somatotopy in adultmammals: Involvement of relatively ineffective synapses. BirthDefects: Original Article Series, 19, 287–314.
29. Garraghty, P. E., Kaas, J. H., and Florence, S. L. (1994) Plasticity ofsensory and motor maps in adult and developing mammals. InAdvances in Neural and Behavioral Development (Casagrande,V. A., and Shinkman, P. G., Eds.), pp. 1–36, Ablex, Norwood.
30. Wilson, P., and Snow, P. J. (1987) Reorganization of the receptivefields of spinocervical tract neurons following denervation of asingle digit in the cat. J. Neurophysiol. 57, 803–818.
31. Koerber, H. R., and Brown, P. B. (1995) Quantitative analysis ofdorsal horn cell receptive fields following limited deafferenta-tion. J. Neurophysiol. 74, 2065–2076.
32. Dostrovsky, J. O., Millar, J., and Wall, P. D. (1976) The immediateshift of afferent drive of dorsal column nucleus cells followingdeafferentation: A comparison of acute and chronic deafferenta-tion in gracile nucleus and spinal cord. Exp. Neurol. 52, 480–495.
33. Millar, J., Basbaum, A. F., and Wall, P. D. (1976) Restructuring ofthe somatotopic map and appearance of abnormal neuronalactivity in the gracile nucleus after partial deafferentation. Exp.Neurol. 50, 658–672.
34. McMahon, S. B., and Wall, P. D. (1983) Plasticity in the nucleusgracilis of the rat. Exp. Neurol. 80, 195–207.
35. Pettit, M. J., and Schwark, H. D. (1993) Receptive field reorgani-zation in dorsal column nuclei during temporary denervation.Science 292, 2054–2056.
36. Northgrave, S. A., and Rasmusson, D. D. (1996) The immediateeffects of peripheral deafferentation on neurons of the cuneatenucleus in raccoons. Somat. Motor Res. 13, 103–113.
37. Pollin, B., and Albe-Fessard, D. (1979) Organization of somaticthalamus in monkeys with and without section of dorsal spinaltracts. Brain Res. 173, 431–449.
38. Garraghty, P. E., and Kaas, J. H. (1991) Functional reorganizationin adult monkey thalamus after peripheral nerve injury.NeuroRe-port 2, 747–750.
39. Rasmusson, D. D., Louw, D. R., and Northgrave, S. A. (1993) Theimmediate effects of peripheral denervation on inhibitorymecha-nisms in the somatosensory thalamus. Somat. Motor Res. 10,69–80.
40. Rasmusson, D. D. (1996) Changes in the organization of theventroposterior lateral thalamic nucleus after digit removal inadult raccoon. J. Comp. Neurol. 364, 92–103.
41. Rasmusson, D. D. (1996) Changes in the response properties ofneurons in the ventroposterior lateral thalamic nucleus of the
Plasticity of Somatosensory Cortex 11
Copyright r 1997 by Academic Press

raccoon after peripheral deafferentation. J. Neurophysiol. 75,2441–2450.
42. Shin, H.-C., Park, S., Son, J., and Sohn, J.-H. (1995) Responsesfrom new receptive fields of VPL neurones following deafferen-tation.NeuroReport 7, 33–36.
43. Garraghty, P. E., and Kaas, J. H. (1991) Large-scale functionalreorganization in adult monkey cortex after peripheral nerveinjury. Proc. Natl. Acad. Sci. USA 88, 6976–6980.
44. Garraghty, P. E., Pons, T. P., Sur, M., and Kaas, J. H. (1989) Thearbors of axons terminating in the middle cortical layers ofsomatosensory area 3b in owl monkeys. Somat. Motor Res. 6,401–411.
45. Florence, S. L., Wall, J. T., and Kaas, J. H. (1989) Somatotopicorganization of inputs from the hand to the spinal gray andcuneate nucleus of monkeys with observations on the cuneatenucleus of humans. J. Comp. Neurol. 286, 48–70.
46. Florence, S. S., Wall, J. T., and Kaas, J. H. (1991) Centralprojections from the skin of the hand in squirrel monkeys. J.Comp. Neurol. 311, 563–578.
47. Schroeder, C. E., Seto, S., Arezzo, J. C., and Garraghty, P. E. (1995)Electrophysiological evidence for overlapping dominant andlatent inputs to somatosensory cortex in squirrel monkeys. J.Neurophysiol. 74, 722–732.
48. Florence, S. L., and Kaas, J. H. (1995) Somatotopy: Plasticity ofsensory maps. In M. A. Arbib (Eds.), The Handbook of BrainTheory and Neural Networks (Arbib, M. A., Ed.), pp. 888–891,MIT Press, Cambridge.
49. Pons, T. P., Garraghty, P. E., Ommaya, A. K., Kaas, J. H., Taub, E.,and Mishkin, M. (1991) Massive cortical reorganization aftersensory deafferentation in adult macaques. Science 252, 1857–1860.
50. Florence, S. L., and Kaas, J. H. (1995) Large-scale reorganizationat multiple levels of the somatosensory pathway follows thera-peutic amputation of the hand in monkeys. J. Neurosci. 15,8083–8095.
51. Wall, P. D. (1970) The sensory and motor role of impulsestraveling in the dorsal columns towards cerebral cortex. Brain 93,505–524.
52. Vierck, C. J., Jr., Cohen, R. H., and Cooper, B. Y. (1985) Effects ofspinal lesions on temporal resolution of cutaneous sensations.Somat. Res. 3, 45–56.
53. Azulay, A., and Schwartz, A. S. (1975) The role of dorsalfuniculus of the primate in tactile discrimination. Exp. Neurol. 46,315–332.
54. Andersson, S. A., Finger, S., and Norrsell, U. (1975) Cerebralunits activated by tactile stimuli via a ventral spinal pathway inprimates. Acta Physiol. Scand. 93, 119–128.
55. Brinkman, J., Bush, B. M., and Porter, R. (1978) Deficientinfluences of peripheral stimuli on precentral neurons in mon-keys with dorsal column lesions. J. Physiol. 276, 27–48.
56. Schneider, R. J. (1990) Loss of information concerning hairdisplacement and other somatic stimuli in the first somaticsensory cortex of unanesthetizedMacaca mulattamonkeys follow-ing dorsal funiculus transections. Exp. Brain Res. 83, 105–114.
57. Jain, N., Catania, K. C., and Kaas, J. H. (1997) Deactivation andreactivation of somatosensory cortex after dorsal spinal cordinjury.Nature 386, 495–498.
58. McMahon, S. B., and Kett-White, R. (1991) Sprouting of periph-eral regenerating primary sensory neurones in the adult centralnervous system. J. Comp. Neurol. 304, 307–315.
59. Woolf, C. J., Shortland, P., and Coggeshall, R. E. (1992) Peripheralnerve injury triggers central sprouting of myelinated afferents.Nature 355, 75–78.
60. Skene, J. H. P. (1989) Axonal growth-associated proteins. Annu.Rev. Neurosci. 12, 127–156.
61. Smith, D. S., and Skene, J. H. P. (1997) A transcription-dependentswitch controls competence of adult neurons for distinct modesof axon growth. J. Neurosci. 17, 646–658.
62. Schwab, M. E. (1996) Bridging the gap in spinal cord regenera-tion.Nature Med. 2, 976.
63. Ramachandran, V. S. (1993) Behavioral and magnetoencephalo-graphic correlates of plasticity in the adult human brain. Proc.Natl. Acad. Sci. USA 90, 10413–10420.
64. Ramachandran, V. S., Rogers-Ramachandran, D., and Stewart,M. (1992) Perceptual correlates of massive cortical reorganiza-tion. Science 258, 1159–1160.
65. Sweet, W. H. (1975) ‘‘Phantom’’ sensations following intraspinalinjury.Neurochirurgia 18, 139–154.
66. Yang, T. T., Gallen, C., Schwartz, B., Bloom, F. E., Ramachandran,V. S., and Cobb, S. (1994) Sensory maps in the human brain.Nature 368, 592–593.
67. Elbert, T., Flor, H., Birbaumer, N., Knecht, S., Hampson, S.,Larbig, W., and Taub, E., (1994) Extensive reorganization of thesomatosensory cortex in adult humans after nervous systeminjury.NeuroReport 5, 2593–2597.
68. Flor, H., Elbert, T., Knecht, S., Wienbruch, C., Pantev, C.,Birbaumer, N., Larbig, W., and Taub, E. (1995) Phantom-limbpain as a perceptual correlate of cortical reorganization follow-ing arm amputation.Nature 375, 482–484.
69. Melzack, R. (1990) Phantom limbs and the concept of a neuroma-trix. Trends Neurosci. 13, 88–92.
70. Garraghty, P. E., and Sur, M. (1990) Morphology of singleintracellularly stained axons terminating in area 3b of macaquemonkeys. J. Comp. Neurol. 294, 583–593.
71. Kaas, J. H., and Florence, S. L. (1996) Brain reorganization andexperience. Peabody J. Ed. 71, 152–167.
12 Florence, Jain, and Kaas
Copyright r 1997 by Academic Press




















