
Water-soluble stimuli responsive star-shaped segmented macromolecules
Upload
independentCategory
view
0download
0
International Journal of Psychophysiology xxx (2012) xxx–xxx
INTPSY-10532; No of Pages 9
Contents lists available at SciVerse ScienceDirect
International Journal of Psychophysiology
j ourna l homepage: www.e lsev ie r .com/ locate / i jpsycho
Working memory of somatosensory stimuli: An fMRI study
Nicoletta Savini a,⁎, Marcella Brunetti a,b, Claudio Babiloni c,d, Antonio Ferretti a,b
a Department of Neuroscience and Imaging, University “G. d'Annunzio” of Chieti, Italyb ITAB — Institute for Advanced Biomedical Technologies, University “G. d'Annunzio” Foundation, Chieti, Italyc Department of Clinical and Experimental Medicine, University of Foggia, Foggia, Italyd IRCCS San Raffaele Pisana, Roma, Italy
⁎ Corresponding author at: Institute for Advanced Biom“G. D'Annunzio” of Chieti, Via dei Vestini, 33, 66013 Ch3556952; fax: +39 0871 3556930.
E-mail address: [email protected] (N. Savini).
0167-8760/$ – see front matter © 2012 Elsevier B.V. Allhttp://dx.doi.org/10.1016/j.ijpsycho.2012.09.007
Please cite this article as: Savini, N., et al., Wo(2012), http://dx.doi.org/10.1016/j.ijpsycho.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 27 April 2012Received in revised form 16 August 2012Accepted 14 September 2012Available online xxxx
Keywords:Working memoryTactilefMRIN-BACK taskFronto-parietal circuit
In a previous study, we have shown that passive recognition of tactile geometrical shapes (i.e. no exploratorymovement) engages prefrontal and premotor areas in addition to somatosensory regions (Savini et al., 2010).In the present study we tested the hypothesis that these regions are involved not only in the perception butalso during working memory of such somatic information.We performed functional magnetic resonance imaging (fMRI) during the execution of N-BACK tasks, with 2Dgeometrical shapes blindly pressed on the subjects' right hand palm. Three conditions with increasing memoryload (0-BACK, 1-BACK, 2-BACK)were used. Results showed that primary somatosensory area (SI), secondary so-matosensory area (SII) and bilateral Insula were active in all conditions, confirming their importance in codingsomatosensory stimuli. Activation of fronto-parietal circuit in supplementary motor area (SMA), right superiorparietal lobe (rSPL), bilateral middle frontal gyrus, left inferior frontal gyrus, and right superior frontal sulcuswas significantly larger during 1-BACK and 2-BACK than 0-BACK. Left superior parietal lobe and right frontaleye field showed a higher activation during the 2-BACK than 0-BACK. Finally, SMA and rSPL were characterizedby a statistically significant higher activation during 2-BACK than 1-BACK, revealing their sensitivity to themem-ory load. These results suggest that working memory of tactile geometrical shapes (no exploratory movement)involves a complex circuit of modal and supramodal fronto-parietal areas.
© 2012 Elsevier B.V. All rights reserved.
1. Introduction
Working memory (WM) is defined as the ability to transiently storeand manipulate information held “on-line” to be used for cognition orfor behavioral guidance (Baddeley, 1996, 2003).WM is typicallymodeledas a “central executive” attentive system supported by a second, moreperipherally-based storage system, interacting together for informationencoding and manipulation, in order to subserve planning, reasoningand problem solving (Baddeley, 1996, 2003; Goldman-Rakic, 1996).
Neuropsychological studies have used several paradigms to disentan-gle different aspects of working memory in humans, such as continuousupdating of information (i.e. on-linemanipulation),maintenance of infor-mation, and information storage. One of the most used paradigms is theN-BACK task, which combines maintenance and encoding, as well asmonitoring of a continuous sequence of stimuli (Fletcher and Henson,2001). The majority of neuroimaging studies of human working mem-ory mainly employed visuo-spatial and verbal N-BACK tasks. A meta-analysis of 24 neuroimaging (positron emission tomography andfunctional magnetic resonance imaging, fMRI) studies using N-BACK
edical Technologies, Universityieti (CH), Italy. Tel.: +39 0871
rights reserved.
rking memory of somatosens2012.09.007
tasks of verbal and visuospatial information has emphasized the roleof a fronto-parietal network including lateral premotor cortex, dorsalcingulate and medial premotor cortex, dorsolateral and ventrolateralprefrontal cortex, frontal poles,medial and lateral posterior parietal cor-tex (Owen et al., 2005).
Nevertheless, the cortical circuits underlying the working memoryload elicited by N-BACK tasks using tactile inputs are still poorly under-stood. Several studies have used 1-BACK condition (i.e. subject has tomaintain inWMthe stimulus before the target) to investigate a sequentialdiscrimination of haptically explored objects (Kaas et al., 2006) and visualand/or tactile stimuli (Stoeckel et al., 2003; Ricciardi et al., 2006). The re-sults showed that during visual and tactile spatialWMa supramodal fron-tal or fronto-parietal network was recruited in all sensory modalities.
Noteworthy, the lack of studies performing a tactile 2-BACK tasklimits a detailed analysis ofmemory load effects on tactile input process-ing. In particular, it is not clear if the premotor and parietal networks sen-sitive to the verbal and visualmemory load (Ragland et al., 2002; Carlsonet al., 1998; Owen et al., 2005) are involved even in themanipulation andmaintenance in memory of somatosensory information.
In a previous study, we have shown that passive recognition oftactile geometrical shapes (i.e. no exploratory movement) engagesprefrontal and premotor areas in addition to somatosensory regions(Savini et al., 2010). In the present study we tested the hypothesisthat these regions are involved not only in the perception but also
ory stimuli: An fMRI study, International Journal of Psychophysiology

2 N. Savini et al. / International Journal of Psychophysiology xxx (2012) xxx–xxx
during working memory of such somatic information. We performedfMRI during the execution of N-BACK tasks, with 2D geometrical shapesblindly pressed on the subjects' right hand palm. Two conditions withincreasing memory load (1-BACK, 2-BACK) were used in addition to arecognition condition with nomemory load (0-BACK). In order to iden-tify regions underlyingWMof somatosensory stimuli we compared thedifferent memory loads to the recognition condition. For the explorato-ry nature of the present study, we focused on the well known generalfeatures of the N-BACK task. This is a well known reference task thatcombines “maintenance of task instructions and previous target senso-ry information in a short term buffer” and “inhibition of impulse re-sponses related to the sensory information of the distracters”. This iswhy the N-BACK is considered a WM task but not merely a short termmemory task. For this reason, we designed a variant of this task forthe specific and original evaluation of the cortical correlates of somaticWM for 2D tactile shapes in the present study.
2. Methods
2.1. Subjects and stimulation procedures
We used the same stimuli and experimental procedures validatedin a previous reference study (Savini et al., 2010). Briefly, stimuliwere delivered to the subjects right hand palm by means of fourwooden pieces, each having one face with a geometrical shape: a cir-cle (area=3.14 cm2), a square (area=2.89 cm2), a triangle (area=2.71 cm2) and an ellipse (area=3.07 cm2) (Fig. 1a). In order to stan-dardize the force applied on the hand, the pieces were mounted onflexible bars with a graduate scale (Fig. 1b). This device allowed theinvestigator to manually present the shapes with the same force(4 N) across subjects and trials. A group of 12 healthy male volunteers,ranging in age from 19 to 32 years (mean age of 23.9±3.5 SD), was en-rolled for the fMRI experiment with these stimuli. All subjects wereright handed according to the Edinburgh Inventory (Oldfield, 1971),gave their written informed consent according to the Declaration ofHelsinki (World Medical Association Declaration of Helsinki, 1997),and could request an interruption of the investigation at any time; thegeneral procedures were approved by the Ethics Committee for Bio-medical Research of University “G. d'Annunzio”, Chieti (IT).We enrolledonly male subjects to maintain a more homogeneous sample, avoidingintersubject variability effects due to e.g. different phases of the men-strual cycle in females.
Fig. 1. a) Bi-dimensional shapes ofwoodenpieces pressed on the righthandpalm; b) devicewithwoodenflexible bars, with a reference scale indicating the applied force. In the picture,the device bearing the triangular shape is shown; c) schematic of the block paradigm: 30 sof rest, 30 s of stimulation, 6 shapes presented per block, and 4 stimulation blocks, for a totalof 24 shapes.
Please cite this article as: Savini, N., et al., Working memory of somatosens(2012), http://dx.doi.org/10.1016/j.ijpsycho.2012.09.007
A training session was performed following the same procedure ofour previous study (Savini et al., 2010) in order to allow the subjectsto recognize the geometrical shapes using the tactile presentation.Then all the subjects were trained to memorize the shape and theywere able to do the N-BACK task.
2.2. Experimental design
Three tasks were performed: i) 0-BACK in which the subjectswere asked to press (with the left hand) the button correspondingto the shape of the wooden surface pressed on their right handpalm; ii) 1-BACK in which the subjects were asked to press the buttoncorresponding to the shape presented one time before (maintaining inmemory the preceding shape); 2-BACK in which the subjects wereasked to press the button corresponding to the shape presented twotimes before (maintaining in memory the preceding two shapes). Sub-jects were instructed to keep their gaze on a fixation cross while doingthe task.
The fMRI session was performed in three different runs (corre-sponding to the three memory load tasks: 0-BACK, 1-BACK, 2-BACK)according to a block paradigm alternating 30 s of stimulation with30 s of a control state. For each run, 4 stimulation blocks and 4 controlblocks were used starting with a control state. In the control blockssubjects were required to rest and keep their gaze on the fixationcross. During each stimulation block, 6 trials were presented. Eachtrial consisted of 2 s, during which a shape was pressed – in a blindcondition – on the subject right hand palm, and 3 s of release. The pre-sentation order of the geometrical shapes was pseudorandom and atotal of 24 shapes were presented per run (Fig. 1c).
The tactile stimuli were delivered by an experimenter following in-structions transmitted via fMRI compatible headphones. To allow a stan-dardized administration of stimuli, the pressure levelwas controlled by areference on the flexible bars (Fig. 1b), whereas the pressure durationand the pseudo-random order of presentation of the different shapeswere controlled by means of a Matlab® (Mathworks, Natick, MA, USA)code. The program transmitted to the experimenter a registered voice,indicating the shape to press, followed by a 2 s of beep indicating thepressure duration. The button press (MRI-compatible response pad Lu-mina LSC-400 controller, Cedrus, California, USA) was recorded by thesame Matlab code, allowing to record reaction times and behavioral re-sponses (correct/incorrect shape recognition).
The three fMRI tasks (0-BACK, 1-BACK, 2-BACK) were completelyrandomized across subjects to avoid any habituation or order effect.
2.3. fMRI recordings
BOLD contrast functional imaging was performed with a SiemensMagnetom Vision scanner at 1.5 T by means of T2*-weighted echoplanar imaging (EPI) free induction decay (FID) sequences with thefollowing parameters: TR 3 s, TE 60 ms, matrix size 64×64, FOV256 mm, inplane voxel size 4 mm×4 mm, flip angle 90°, slice thick-ness 4 mm and no gap. A standard head coil was used and the subjecthead was fixed with foam pads to reduce involuntary movement.Subject right hand was blocked on a soft pad by means of tape toavoid any exploratory movement (the experimenter giving the stim-ulation continuously checked it). 83 functional volumes consisting of28 transaxial slices parallel to the AC–PC line were acquired per run.
A high-resolution structural volume was acquired at the end of thesession via a 3DMPRAGE sequencewith the following features: sagittal,matrix 256×256, FOV 256 mm, slice thickness 1 mm, no gap, in-planevoxel size 1 mm×1 mm, flip angle 12°, TR=9.7 ms, TE=4 ms.
2.4. Data analysis
Rawdatawere analyzed bymeans of the Brain Voyager QX software(Brain Innovation, The Netherlands). Due to T1 saturation effects, the
ory stimuli: An fMRI study, International Journal of Psychophysiology

3N. Savini et al. / International Journal of Psychophysiology xxx (2012) xxx–xxx
first three scans of each run were discarded from the analysis. Prepro-cessing of functional scans included slice scan time correction, motioncorrection, spatial smoothing with a Gaussian isotropic kernel (fullwidth half maximum of 6 mm), and the application of a temporalhigh-pass filter with cut-off set to 3 cycles per time-course.
A three-dimensional motion correction was performed by meansof a rigid body transformation to match each functional volume tothe reference volume (the fourth volume, since the first three vol-umes were discarded to avoid the T1 saturation effect), estimatingthree translation and three rotation parameters. These parameterswere stored in log files and inspected to check that (i) estimatedmovement was not larger than 2 mm or 2° (approximately half avoxel) for each functional run and (ii) no stimulus-correlated move-ment occurred (Friston et al., 1995; Hajnal et al., 1994).
Spatial normalization was performed for structural and function-al data sets. The spatial normalization of the structural volumes wasperformed in two steps. The first step consisted in aligning the 3DMPRAGE data set of each subject with the stereotaxic axes. For thisstep the location of the anterior commissure (AC), the posterior com-missure (PC) and two rotation parameters for midsagittal alignmenthad to be specifiedmanually in the 3D data set. In the second step theextreme points of the cerebrum were specified. These points togeth-er with the AC and PC coordinates were then used to scale the 3Ddata sets into the dimensions of the standard brain of the Talairachand Tournaux atlas (Talairach and Tournoux, 1998) using a piece-wise affine and continuous transformation. To transform the func-tional data into Talairach space, the preprocessed functional timeseries were first resampled at a voxel size of 3 mm×3 mm×3 mmand coregistered with the corresponding structural data set. Thecoregistration transformation in BrainVoyager QX was determinedconcatenating an initial alignment matrix obtained using the Siemensposition parameters of the functional and structural images with a finetuning alignmentmatrix obtained bymeans of an intensity-driven align-ment algorithm. The alignment between functional and anatomicalscans was finally checked by means of an accurate visual inspection.Then the rigid body AC–PC transformation matrix and the piecewise af-fine Talairach grid scaling transformation performed for the 3D anatom-ical data set were applied to coregistered functional data. This procedureresulted in a normalized four-dimensional data representation (volumetime course — VTC) for each functional run. In order to avoid qualityloss due to successive data sampling, the spatial normalizationwas actu-ally performedusing a single transformationmatrix obtainedby combin-ing the described spatial transformations.
The voxel-wise statistical group analysis was performed accordingto the following steps. First, the voxel time series from each subjectwere z-normalized. Such a transformation normalizes the mean of avoxel time course to zero and the variance to one. This normalizationis useful in order to take into account baseline signal variations inmulti-study general linear models, and it is particularly important inour design where the experimental conditions are separated in differ-ent runs. However, one potential disadvantage is that it diminishesreal differences in the effect size of different runs from different sub-jects. To avoid this effect, we used a variant of z-normalization whichnormalizes using only the time course values corresponding to thebaseline condition. This results in a z normalization based solely onthe noise variance and leads to effect estimations which are expressedrelative to the noise variance.
Statistical analysis was then performed on normalized VTCs of in-dividual subjects using the general linear model (GLM; Friston et al.,1995) with correction for temporal autocorrelation (Bullmore et al.,1996; Woolrich et al., 2001), considering three predictors, corre-sponding to the 0-BACK, 1-BACK and 2-BACK tasks. To account forthe hemodynamic delay, the boxcar waveform representing the restand task conditions was convolved with an empirically founded he-modynamic response function (Boynton et al., 1996). Second, theGLM parameters (normalized beta values) estimated in individual
Please cite this article as: Savini, N., et al., Working memory of somatosens(2012), http://dx.doi.org/10.1016/j.ijpsycho.2012.09.007
subject analysis were entered in a second level voxel-wise random ef-fect analysis, in order to search for activated areas that were consis-tent for the whole group (N=12).
The group statistical maps were thresholded at pb0.05, correctedfor multiple comparisons. The correction for multiple comparisonswas performed using a cluster-size thresholding algorithm (Formanet al., 1995) based on Monte Carlo simulations and implemented inthe BrainVoyager QX software. A threshold of pb0.0009 at the voxellevel and an estimate of the spatial correlation of voxels were used asinput in the simulations, yielding a minimum cluster-size of sevenvoxels to obtain statistical maps thresholded at a standard alpha level(pb0.05, corrected for multiple comparisons).
2.5. Statistical analysis of BOLD activation in regions of interest (ROIs)
First, regions of interest (ROIs) were defined in functional terms byconsidering, in the group statistical maps, voxels obtained contrastingthe task conditionswith the rest condition. In order to avoid bias towardone of the conditions, ROIs were delineated by considering the maskobtained pooling voxels activated during any task condition, i.e. includ-ing voxels activated in at least one of the three conditions, following thesameprocedure used in the previous papers (Ferretti et al., 2004; Saviniet al., 2010). These ROIs were then used for the analysis of fMRI activa-tion across subjects and conditions. Specifically, the mean time courseof the fMRI signal from voxels belonging to a given ROI was analyzedfor each subject and the individual BOLD responses to the differenttask conditions were characterized by the fitted parameters (nor-malized beta values) of the GLM. For each ROI, activation across the0-BACK, 1-BACK and 2-BACK conditions was compared by means ofthe analysis of variance (ANOVA) for repeated measures. The depen-dent variable of the ANOVA analysis was the BOLD response (nor-malized beta values) and the ANOVA factor was the task condition(0-BACK, 1-BACK and 2- BACK). For all the ANOVA calculations,Mauchley's test was used to evaluate the sphericity assumption.The number of degrees of freedom was corrected by means of theGreenhouse–Geisser procedure and the Duncan's test was used forpost hoc comparisons. This ANOVA analysis allowed to reveal, amongall activated areas, those ROIs showing a significant modulation of acti-vation with the memory load.
3. Results
3.1. Behavioral data
The 12 subjects participating in the experiment were able to performthe task during the fMRI session with a mean accuracy of 62.5% (±24.7%standard deviation, SD) and a reaction time of 1633 ms (±157 ms stan-dard deviation, SD) in the 0-BACK task, 81.8% (±16.2% standard devia-tion, SD) and a reaction time of 1862 ms (±244 ms standard deviation,SD) in the 1-BACK task, 71.8% (±14.9% standard deviation, SD) and areaction time of 1747 ms (±231 ms standard deviation, SD) in the2-BACK task. In each experimental condition a negative correlationtrend was observed across subjects between reaction time and hit rate(r=−0.52 p=0.08 N=12 in 0-BACK; r=−0.31 p=0.3 N=12 in1-BACK; r=−0.53 p=0.07 N=12 in 2-BACK). Even if not statisticallysignificant (p>0.05), this trend indicated that, within each task, the ex-ecution of a correct performance occurs with a short reaction time. Thelow mean accuracy in the 0-BACK condition was due to three subjectswith low hit rate for this condition. However, they showed an averageperformance in the higher memory loads. Given that the comparison2-BACK vs. 1-BACK is a major focus of the study, we did not excludethese subjects from the fMRI analysis. Anyway, within each condition,the negative correlation trend of the performance with the responsetime in our data confirmed that the execution of a correct performanceoccurs with a short reaction time.
ory stimuli: An fMRI study, International Journal of Psychophysiology

Table 1Talairach coordinates (center of gravity) and Z scores of activated areas during 0-BACK, 1-BACK and 2-BACK conditions.SI: primary somatosensory area; SII: secondary somatosensory area; DLPFC: dorsolateral prefrontal cortex.
Talairach coord. Cluster size z-score
x y Z (mm3) 0-back 1-back 2-back
Cortical areas of somatosensory network and motor cortexLeft SI −40 −37 50 3213 4.4 4.1 4Contralateral SII −55 −18 18 1377 4.6 5.6 4Right supramarginal gyrus 51 −20 28 1728 3 4.1 3.5Right primary motor area M1 38 −29 50 2862 4.2 4.3 4.5
Cortical areas of fronto-parietal networkCingulate cortex 4 20 37 1944 4.1 3.8 3.1Supplementary motor area (SMA) −2 −3 59 3564 3.7 4.5 5.6Left DLPFC BA9 posterior −45 4 30 1917 4 4 5Right DLPFC BA9 posterior 43 2 36 2322 4.2 5 4.3Right DLPFC BA9/BA46 42 27 32 3024 3 4.5 3.1Right DLPFC BA9 anterior 24 32 30 1080 2.7 2.9 4.2Left DLPFC BA9 anterior −29 41 30 621 1.1 2.5 4.8Right middle temporal gyrus 48 −30 −4 864 3.1 1.6 4Right superior temporal gyrus 50 −39 27 1053 4.3 4.1 5.3Left frontal eye fields −30 −11 53 2862 3.2 3.3 4.7Right superior frontal gyrus 26 10 50 567 1.9 3 4.3Right frontal eye fields 31 −11 54 3159 4.3 3.4 5.1Left insula −40 −4 11 351 3.4 3.7 4.4Left anterior insula/frontal operculus −35 19 5 1917 3.7 4.9 3.8Right anterior insula/frontal operculus 35 17 4 1674 4.7 4.1 4Left Broca's area −42 16 8 1431 2.7 4.3 4.4Right inferior parietal lobe 40 −39 45 3915 4.9 4.9 4.4Left inferior parietal lobe −42 −37 43 3969 5.8 4.4 4.5Right superior parietal lobe 6 −64 58 189 – 1.7 3.7Left superior parietal lobe −28 −63 48 1215 2.5 2.9 3.9
4 N. Savini et al. / International Journal of Psychophysiology xxx (2012) xxx–xxx
In the 1-BACK and 2-BACK conditions, behavioral results show thatthe subjects memorized a reasonable amount of tactile stimuli, takinginto account that the task was performed inside the fMRI scanner. Sur-prisingly, the performance in the 0-BACK conditionwas lower than thatof the 1-BACK and 2-BACK conditions. It can be speculated that in the0-BACK condition, some subjects underrated the difficulty of the recog-nition task with respect to the 1-BACK and 2-BACK conditions.
3.2. Common activation in 0-BACK, 1-BACK, 2-BACK conditions
A significant activation (pb0.05, corrected for multiple compari-sons) when contrasting the 0-BACK, 1-BACK and 2-BACK conditionswith the rest condition was observed in the contralateral SII (cSII),and contralateral SI (cSI). Furthermore, fronto-parietal activation wasobserved in bilateral inferior parietal lobe (IPL), bilateral superior pari-etal lobe (SPL), bilateral insular cortex, SMA, bilateral dorsolateral pre-frontal cortex (DLPFC), left inferior frontal gyrus (Broca's area), right
Table 2Talairach coordinates (center of gravity) in areas modulating during 0-BACK, 1-BACK and 2
Cortical areas of fronto-parietal network Talairach coord.
x y z
Supplementary motor area (SMA) 0 −1 55Right superior parietal lobe 6 −64 58Left anterior DLPFC BA9 −29 41 30Right anterior DLPFC BA9 24 32 30Left posterior DLPFC BA9 −45 4 30Left Broca's area −42 16 8Right superior frontal sulcus 26 10 50Right frontal eye fields 31 −11 54Left superior parietal lobe −28 −63 48
Please cite this article as: Savini, N., et al., Working memory of somatosens(2012), http://dx.doi.org/10.1016/j.ijpsycho.2012.09.007
superior frontal sulcus (SFS) and bilaterally in the frontal eye field(FEF). A detailed list of activated areas is reported in Table 1.
3.3. Statistical comparison of the fMRI responses across different tasks
A one-way ANOVA design was used to compare, for each ROI, theBOLD response (dependent variable) during the 0-BACK, 1-BACK, and2-BACK conditions across subjects. A significant main effect was ob-served for the factor condition in the areas shown in Table 2. DetailedF-values and significance p-values for main effect and post-hoc compar-isons are reported as well. Bilateral anterior DLPFC (BA9), left posteriorDLPFC (BA9), right SFS (BA6), and Broca's area showed a statistically sig-nificant larger activation during 1-BACK and 2-BACK conditions with re-spect to 0-BACK condition.
Statistically significant differences in activation when comparing2-BACK vs. 0-BACK, were observed in the left SPL and in the right FEF.Moreover, SMA and right SPL showed a statistically significant largeractivation in 2-BACK vs. 1-BACK and at the same time a statistically
-BACK conditions, and ANOVA results.
Anova results Duncan's post hoc comparisons
F (2,22) p 1-BACKvs.0-BACK
2-BACKvs.0-BACK
2-BACKvs.1-BACK
14.443 .00010 .020 .000093 .00917.168 .000032 .030 .000076 .0025.9872 .00839 .008 .007 n.s.6.4013 .00644 .021 .003 n.s.3.6923 .04143 .048 .023 n.s.6.3184 .00679 .033 .002 n.s.4.9975 .01625 .010 .019 n.s.4.0911 .03086 n.s. .012 n.s.5.2767 .01343 n.s. .005 n.s.
ory stimuli: An fMRI study, International Journal of Psychophysiology

Fig. 2. Activated areas during the 0-BACK, 1-BACK, and 2-BACK conditions in somatosensory cortices (secondary somatosensory area— SII) and insula. For visualization purpose, thestatistical activation maps were obtained pooling the three conditions and thresholded at pb0.05, corrected for multiple comparisons (see main text). Bar graphs represent nor-malized beta values and standard errors for the three conditions, extracted for each area from the corresponding ROI defined as described in the main text.
5N. Savini et al. / International Journal of Psychophysiology xxx (2012) xxx–xxx
significant larger activation in 1-BACK vs. 0-BACK. Therewere no statis-tically significant differences in activation in the somatosensory areas(cSI, cSII, iSII) and insula across the three conditions. The right primarymotor area (M1)was equally activated during all conditions aswell. Theactivation maps and BOLD responses of the principal areas discussedbelow are reported in Figs. 2–4.
4. Discussion
The present exploratory fMRI study aimed to clarify the neural sys-tems underlying the working memory of tactile 2D geometrical shapesin a relatively small group of healthy subjects. FMRI was performedwhile wooden 2D geometrical shapes were blindly pressed on subjects'passive right hand palm, without exploratory movements in three ex-perimental conditions (0-BACK, 1-BACK and 2-BACK). The 0-BACK con-dition corresponded to the recognition (identification) of the shape,whereas the 1-BACK and 2-BACK conditions were the memory condi-tions. In order to disentangle the functional network underlying WMof somatosensory stimuli, we compared the recognition condition tothe two memory load conditions.
As mentioned in the Introduction, the concept of WM includes notonly the maintenance function but also other cognitive processes in-cluding the inhibition of the interference effects of the distracters.Therefore, the functional network that we observed globally refers tothe cognitive functions involved in the concept of WM, in line withthe literature on fMRI andWM in humans. Indeed, there is a consensus
Please cite this article as: Savini, N., et al., Working memory of somatosens(2012), http://dx.doi.org/10.1016/j.ijpsycho.2012.09.007
that theWM load is higher in the 2-BACK than in the 1-BACK conditionand in the 2-BACK and 1-BACK than in the 0-BACK.
4.1. Somatosensory system and insula activation
We reported activation in SI and SII in all experimental conditions(0-BACK, 1-BACK, 2-BACK), in line with the well known functional roleperformed by the somatosensory system during processing of somaticstimuli (Burton and Fabri, 1995; Ferretti et al., 2004; Diamond et al.,2002). No significant differences in activation were observed across thedifferent memory loads, suggesting a preliminary and steady-state fea-ture extraction processing along the increasing memory load process, asa possible contribution of the somatosensory system in WM of tactilegeometrical shapes.
A lot of studies have demonstrated the involvement of “modal” brainareas during the “type of material” information storage. Visual presenta-tion of linguistic contents has induced an activation of primary visualareas during different memory loads in N-BACK conditions (Smith andJonides, 1997). Similar results were shown during an N-BACK with vi-suospatial contents, inwhich activation of primary visual areaswas com-parable across different memory loads (Carlson et al., 1998). The samewas true for the activation of auditory areas when auditory stimuliwere used in N-BACK tasks (Martinkauppi et al., 2000).
Activity in bilateral insula showed a similar pattern, with no sig-nificant differences in activation observed across the different memo-ry loads. However, a proper interpretation of insular activation would
ory stimuli: An fMRI study, International Journal of Psychophysiology
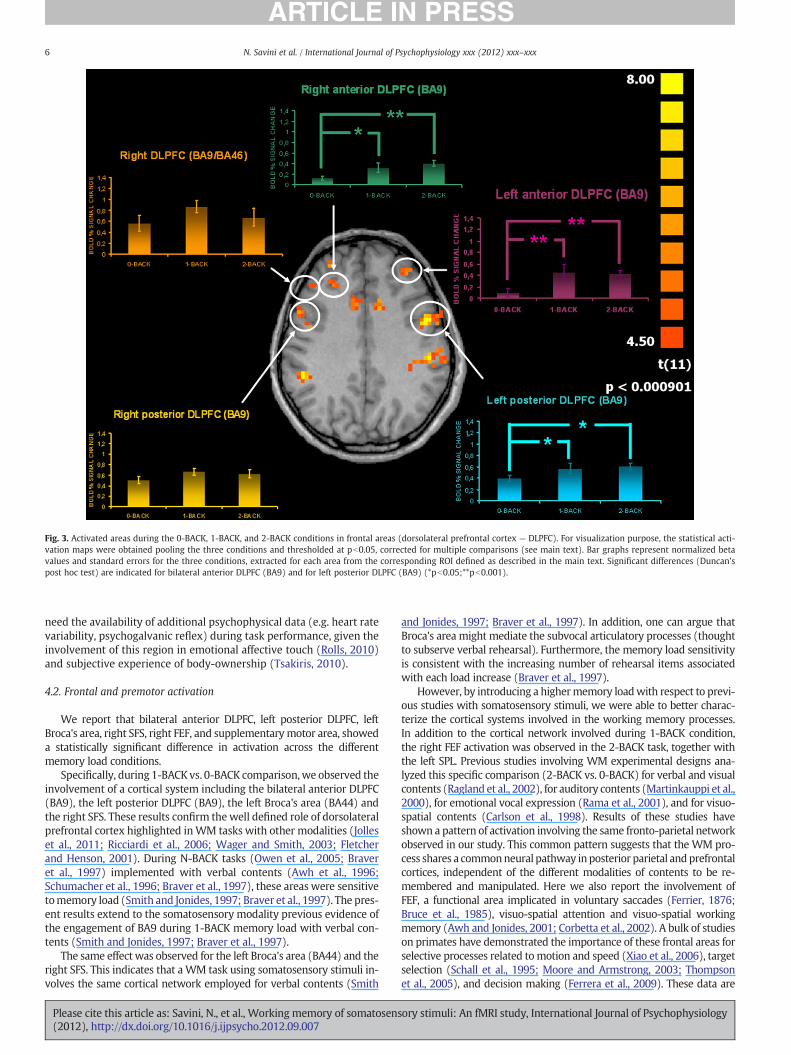
Fig. 3. Activated areas during the 0-BACK, 1-BACK, and 2-BACK conditions in frontal areas (dorsolateral prefrontal cortex — DLPFC). For visualization purpose, the statistical acti-vation maps were obtained pooling the three conditions and thresholded at pb0.05, corrected for multiple comparisons (see main text). Bar graphs represent normalized betavalues and standard errors for the three conditions, extracted for each area from the corresponding ROI defined as described in the main text. Significant differences (Duncan'spost hoc test) are indicated for bilateral anterior DLPFC (BA9) and for left posterior DLPFC (BA9) (*pb0.05;**pb0.001).
6 N. Savini et al. / International Journal of Psychophysiology xxx (2012) xxx–xxx
need the availability of additional psychophysical data (e.g. heart ratevariability, psychogalvanic reflex) during task performance, given theinvolvement of this region in emotional affective touch (Rolls, 2010)and subjective experience of body-ownership (Tsakiris, 2010).
4.2. Frontal and premotor activation
We report that bilateral anterior DLPFC, left posterior DLPFC, leftBroca's area, right SFS, right FEF, and supplementarymotor area, showeda statistically significant difference in activation across the differentmemory load conditions.
Specifically, during 1-BACK vs. 0-BACK comparison,we observed theinvolvement of a cortical system including the bilateral anterior DLPFC(BA9), the left posterior DLPFC (BA9), the left Broca's area (BA44) andthe right SFS. These results confirm thewell defined role of dorsolateralprefrontal cortex highlighted inWM tasks with other modalities (Jolleset al., 2011; Ricciardi et al., 2006; Wager and Smith, 2003; Fletcherand Henson, 2001). During N-BACK tasks (Owen et al., 2005; Braveret al., 1997) implemented with verbal contents (Awh et al., 1996;Schumacher et al., 1996; Braver et al., 1997), these areas were sensitivetomemory load (Smith and Jonides, 1997; Braver et al., 1997). The pres-ent results extend to the somatosensory modality previous evidence ofthe engagement of BA9 during 1-BACK memory load with verbal con-tents (Smith and Jonides, 1997; Braver et al., 1997).
The same effect was observed for the left Broca's area (BA44) and theright SFS. This indicates that a WM task using somatosensory stimuli in-volves the same cortical network employed for verbal contents (Smith
Please cite this article as: Savini, N., et al., Working memory of somatosens(2012), http://dx.doi.org/10.1016/j.ijpsycho.2012.09.007
and Jonides, 1997; Braver et al., 1997). In addition, one can argue thatBroca's area might mediate the subvocal articulatory processes (thoughtto subserve verbal rehearsal). Furthermore, the memory load sensitivityis consistent with the increasing number of rehearsal items associatedwith each load increase (Braver et al., 1997).
However, by introducing a highermemory loadwith respect to previ-ous studies with somatosensory stimuli, we were able to better charac-terize the cortical systems involved in the working memory processes.In addition to the cortical network involved during 1-BACK condition,the right FEF activation was observed in the 2-BACK task, together withthe left SPL. Previous studies involving WM experimental designs ana-lyzed this specific comparison (2-BACK vs. 0-BACK) for verbal and visualcontents (Ragland et al., 2002), for auditory contents (Martinkauppi et al.,2000), for emotional vocal expression (Rama et al., 2001), and for visuo-spatial contents (Carlson et al., 1998). Results of these studies haveshown a pattern of activation involving the same fronto-parietal networkobserved in our study. This common pattern suggests that the WM pro-cess shares a commonneural pathway inposterior parietal andprefrontalcortices, independent of the different modalities of contents to be re-membered and manipulated. Here we also report the involvement ofFEF, a functional area implicated in voluntary saccades (Ferrier, 1876;Bruce et al., 1985), visuo-spatial attention and visuo-spatial workingmemory (Awh and Jonides, 2001; Corbetta et al., 2002). A bulk of studieson primates have demonstrated the importance of these frontal areas forselective processes related tomotion and speed (Xiao et al., 2006), targetselection (Schall et al., 1995; Moore and Armstrong, 2003; Thompsonet al., 2005), and decision making (Ferrera et al., 2009). These data are
ory stimuli: An fMRI study, International Journal of Psychophysiology
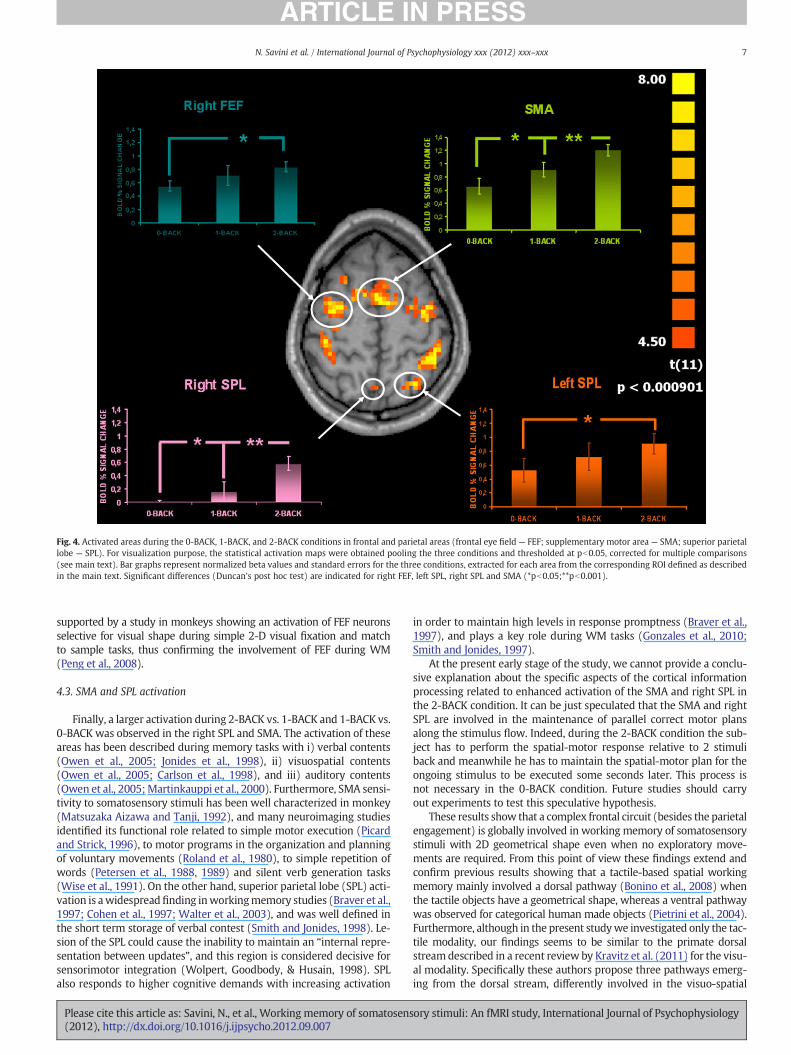
Fig. 4. Activated areas during the 0-BACK, 1-BACK, and 2-BACK conditions in frontal and parietal areas (frontal eye field — FEF; supplementary motor area — SMA; superior parietallobe — SPL). For visualization purpose, the statistical activation maps were obtained pooling the three conditions and thresholded at pb0.05, corrected for multiple comparisons(see main text). Bar graphs represent normalized beta values and standard errors for the three conditions, extracted for each area from the corresponding ROI defined as describedin the main text. Significant differences (Duncan's post hoc test) are indicated for right FEF, left SPL, right SPL and SMA (*pb0.05;**pb0.001).
7N. Savini et al. / International Journal of Psychophysiology xxx (2012) xxx–xxx
supported by a study in monkeys showing an activation of FEF neuronsselective for visual shape during simple 2-D visual fixation and matchto sample tasks, thus confirming the involvement of FEF during WM(Peng et al., 2008).
4.3. SMA and SPL activation
Finally, a larger activation during 2-BACK vs. 1-BACK and 1-BACK vs.0-BACK was observed in the right SPL and SMA. The activation of theseareas has been described during memory tasks with i) verbal contents(Owen et al., 2005; Jonides et al., 1998), ii) visuospatial contents(Owen et al., 2005; Carlson et al., 1998), and iii) auditory contents(Owen et al., 2005;Martinkauppi et al., 2000). Furthermore, SMA sensi-tivity to somatosensory stimuli has been well characterized in monkey(Matsuzaka Aizawa and Tanji, 1992), and many neuroimaging studiesidentified its functional role related to simple motor execution (Picardand Strick, 1996), to motor programs in the organization and planningof voluntary movements (Roland et al., 1980), to simple repetition ofwords (Petersen et al., 1988, 1989) and silent verb generation tasks(Wise et al., 1991). On the other hand, superior parietal lobe (SPL) acti-vation is awidespreadfinding inworkingmemory studies (Braver et al.,1997; Cohen et al., 1997; Walter et al., 2003), and was well defined inthe short term storage of verbal contest (Smith and Jonides, 1998). Le-sion of the SPL could cause the inability to maintain an “internal repre-sentation between updates”, and this region is considered decisive forsensorimotor integration (Wolpert, Goodbody, & Husain, 1998). SPLalso responds to higher cognitive demands with increasing activation
Please cite this article as: Savini, N., et al., Working memory of somatosens(2012), http://dx.doi.org/10.1016/j.ijpsycho.2012.09.007
in order to maintain high levels in response promptness (Braver et al.,1997), and plays a key role during WM tasks (Gonzales et al., 2010;Smith and Jonides, 1997).
At the present early stage of the study, we cannot provide a conclu-sive explanation about the specific aspects of the cortical informationprocessing related to enhanced activation of the SMA and right SPL inthe 2-BACK condition. It can be just speculated that the SMA and rightSPL are involved in the maintenance of parallel correct motor plansalong the stimulus flow. Indeed, during the 2-BACK condition the sub-ject has to perform the spatial-motor response relative to 2 stimuliback and meanwhile he has to maintain the spatial-motor plan for theongoing stimulus to be executed some seconds later. This process isnot necessary in the 0-BACK condition. Future studies should carryout experiments to test this speculative hypothesis.
These results show that a complex frontal circuit (besides the parietalengagement) is globally involved in working memory of somatosensorystimuli with 2D geometrical shape even when no exploratory move-ments are required. From this point of view these findings extend andconfirm previous results showing that a tactile-based spatial workingmemory mainly involved a dorsal pathway (Bonino et al., 2008) whenthe tactile objects have a geometrical shape, whereas a ventral pathwaywas observed for categorical human made objects (Pietrini et al., 2004).Furthermore, although in the present studywe investigated only the tac-tile modality, our findings seems to be similar to the primate dorsalstreamdescribed in a recent review by Kravitz et al. (2011) for the visu-al modality. Specifically these authors propose three pathways emerg-ing from the dorsal stream, differently involved in the visuo-spatial
ory stimuli: An fMRI study, International Journal of Psychophysiology

8 N. Savini et al. / International Journal of Psychophysiology xxx (2012) xxx–xxx
processing. Future studies should be performed in order to investigate ifthis segregation is partially shared with the tactile modality.
5. Conclusions
In a previous study, we have shown that passive recognition of tactilegeometrical shapes (i.e. no exploratory movement) engages prefrontaland premotor areas in addition to somatosensory regions (Savini et al.,2010). In the present exploratory fMRI study, we tested the hypothesisthat these regions are involved not only in the perception but also dur-ing WM of such somatic information. The results showed that the in-volvement of the somatosensory system is extended over the meretactile recognition process and is maintained during storage of geomet-ric shapes. Furthermore, the results suggest that WM of 2D tactile geo-metrical shapes (with no exploratorymovement) involves primary andsecondary somatosensory areas and, in addition, a fronto-parietal cir-cuit similar to that involved in theWMof visual, visuospatial and verbalcontents.
Since WM includes not only short term memory of sensory infor-mation but also other cognitive processes including the inhibition of“interference” processing related to distracters, the heuristic valueof the present results is to motivate further investigations aimed attesting specific hypotheses on the functional meaning of the activa-tion of the regions of interest characterizing the 2-BACK conditionwith respect to the 1-BACK and 0-BACK conditions.
Acknowledgments
Wewant to thank Mr. Mario Savini for wooden objects realization.
References
Awh, E., Jonides, J., 2001. Overlapping mechanisms of attention and spatial workingmemory. Trends in Cognitive Sciences 5 (3).
Awh, E., Jonides, J., Smith, E.E., Schumacher, E.H., Koeppe, R.A., Katz, S., 1996. Dissocia-tion of storage and rehearsal in verbal working memory: evidence from PET. Psy-chological Science 7, 25–31.
Baddeley, A., 1996. The fractionation of working memory. Proceedings of the NationalAcademy of Sciences of the United States of America 93, 13468–13472.
Baddeley, A., 2003. Working memory: looking back and looking forward. Nature Re-views. Neuroscience 4, 829–839.
Bonino, D., Ricciardi, E., Sani, L., Gentili, C., Vanello, N., Guazzelli, M., Vecchi, T., Pietrini,P., 2008. Tactile spatial working memory activates the dorsal extrastriate corticalpathway in congenitally blind individuals. Archives Italiennes de Biologie 146,133–146.
Boynton, G.M., Engel, S.A., Glover, G.H., Heeger, D.J., 1996. Linear systems analysis offunctional magnetic resonance imaging in human V1. The Journal of Neuroscience16, 4207–4241.
Braver, T.S., Cohen, J.D., Nystrom, L.E., Jonides, J., Smith, E.E., Noll, D.C., 1997. A parametricstudy of prefrontal cortex involvement in human working memory. NeuroImage 5,49–62.
Bruce, C.J., Goldberg, M.E., Bushnell, M.C., Stanton, G.B., 1985. Primate frontal eye fields.II. Physiological and anatomical correlates of electrically evoked eye movements.Journal of Neurophysiology 54, 714–734.
Bullmore, E., Brammer, M., Williams, S., Rabe-Hesketh, S., Janot, N., David, A., Mellers, J.,Howard, R., Sham, P., 1996. Statistical methods of estimation and inference forfunctional MR image analysis. Magnetic Resonance in Medicine 35, 261–277.
Burton, H., Fabri, M., 1995. Ipsilateral intracortical connections of physiologically de-fined cutaneous representations in areas 3b and 1 of macaque monkeys: projec-tions in the vicinity of the central sulcus. The Journal of Comparative Neurology355, 508–538.
Carlson, S., Martinkauppi, S., Rama, P., Salli, E., Korvenoja, A., Aronen, H.J., 1998. Distri-bution of cortical activation during visuospatial n-back tasks as revealed by func-tional magnetic resonance imaging. Cerebral Cortex 8, 743–752.
Cohen, J.D., Perlstein, W.M., Braver, T.S., Nystrom, L.E., Noll, D.C., Jonides, J., 1997. Temporaldynamics of brain activation during a working memory task. Nature 386, 604–608.
Corbetta, M., Kincade, M.J., Shulman, G., 2002. Neural systems for visual orienting andtheir relationships to spatial working memory. Journal of Cognitive Neuroscience14, 508–523.
Diamond, M.E., Harris, J.A., Petersen, R.S., 2002. Sensory Learning and the Brain's BodyMap. In: Nelson, R.J. (Ed.), The somatosensory system. CRC Press, Boca Raton, USA,pp. 183–195.
Ferrera, V.P., Yanike, M., Cassanello, C., 2009. Frontal eye field neurons signal changesin decision criteria. Nature Neuroscience 12, 1458–1462.
Ferretti, A., Del Gratta, C., Babiloni, C., Caulo, M., Arienzo, D., Tartaro, A., Rossini, P.M.,Romani, Gian Luca, 2004. Functional topography of the secondary somatosensory
Please cite this article as: Savini, N., et al., Working memory of somatosens(2012), http://dx.doi.org/10.1016/j.ijpsycho.2012.09.007
cortex for nonpainful and painful stimulation of median and tibial nerve: an fMRIstudy. NeuroImage 23, 1217–1225.
Ferrier, D., 1876. The Functions of the Brain. G. P. Putnam's Sons, New York.Fletcher, P.C., Henson, R.N., 2001. Frontal lobes and human memory: insights from
functional neuroimaging. Brain 124, 849–881.Forman, S.D., Cohen, J.D., Fitzgerald, M., Eddy,W.F., Mintun, M.A., Noll, D.C., 1995. Improved
assessment of significant activation in functional magnetic resonance imaging (fMRI):use of a cluster-size threshold. Magnetic Resonance in Medicine 33, 636–647.
Friston, K.J., Holmes, A.P., Worsley, K.J., Poline, J.P., Frith, C.D., Frackowiak, R.S.J., 1995.Statistical parametric maps in functional imaging: a general linear approach.Human Brain Mapping 2, 173–181.
Goldman-Rakic, P.S., 1996. Regional and cellular fractionation of working memory.Proceedings of the National Academy of Sciences of the United States of America93, 13473–13480 (Review).
Gonzales, M.M., Tarumi, T., Tanaka, H., Sugawara, J., Swann-Sternberg, T., Goudarzi, K.,Haley, A.P., 2010. Functional imaging of working memory and peripheral endothe-lial function in middleaged adults. Brain and Cognition 73, 146–151.
Hajnal, J.V., Myers, R., Oatridge, A., Schwieso, J.E., Young, I.R., Bydder, G.M., 1994. Arti-facts due to stimulus correlated motion in functional imaging of the brain. Magnet-ic Resonance in Medicine 31, 283–291.
Jolles, D.D., van Buchem, M.A., Crone, E.A., Rombouts, S.A., 2011. Functional brain con-nectivity at rest changes after working memory training. Human Brain Mapping.http://dx.doi.org/10.1002/hbm.21444 (Nov 11, Epub ahead of print).
Jonides, J., Schumacher, E.H., Smith, E.E., Koeppe, R.A., Awh, E., Reuter-Lorenz, P.A.,Marshuetz, C., Willis, C.R., 1998. The role of parietal cortex in verbal working mem-ory. The Journal of Neuroscience 18, 5026–5034.
Kaas, A.L., van Mier, H., Goebel, R., 2006. The neural correlates of human working mem-ory for haptically explored object orientations. Cerebral Cortex 17, 1637–1649.
Kravitz, D.J., Saleem, K.S., Baker, C.I., Mishkin, M., 2011. A new neural framework for vi-suospatial processing. Nature Reviews. Neuroscience 12, 217–230.
Martinkauppi, S., Rama, P., Aronen, H.J., Korvenoja, A., Carlson, S., 2000. Working mem-ory of auditory localization. Cerebral Cortex 10, 889–898.
Matsuzaka Aizawa, H., Tanji, J., 1992. A motor area rostral to the supplementary motorarea (presupplementary motor area) in the monkey: neuronal activity during alearned motor task. Journal of Neurophysiology 68, 653–662.
Moore, T., Armstrong, K.M., 2003. Selective gating of visual signals by microstimulationof frontal cortex. Nature 421, 370–373.
Oldfield, R.C., 1971. The assessment and analysis of handedness: the Edinburgh inven-tory. Neuropsychologia 9, 97–113.
Owen, A.M., McMillan, K.M., Laird, A.R., Bullmore, E., 2005. N-back working memoryparadigm: a meta-analysis of normative functional neuroimaging studies. HumanBrain Mapping 25, 46–59.
Peng, X., Sereno, M.E., Sila, A.K., Lehky, S.R., Sereno, A.B., 2008. Shape selectivity in pri-mate FEF. Journal of Neurophysiology 100, 796–814.
Petersen, S.E., Fox, P.T., Posner, M.I., Mintun, M., Raichle, M.E., 1988. Positron emissiontomographic studies of the cortical anatomy of single-word processing. Nature331, 585–589.
Petersen, S.E., Robinson, D.L., Currie, J.N., 1989. Influences of lesions of parietal cortexon visual spatial attention in humans. Experimental Brain Research 76, 267–280.
Picard, N., Strick, P.L., 1996. Motor areas of the medial wall: a review of their locationand functional activation. Cerebral Cortex 6, 342–353.
Pietrini, P., Furey, M.L., Ricciardi, E., Gobbini, M.I., Wu, W.H., Cohen, L., Guazzelli, M.,Haxby, J.V., 2004. Beyond sensory images: object-based representation in thehuman ventral pathway. Proceedings of the National Academy of Sciences of theUnited States of America 101, 5658–5663.
Ragland, J.D., Turetsky, B.I., Gur, R.C., Gunning-Dixon, F., Turner, T., Schroeder, L., Chan,R., Gur, R.E., 2002. Working memory for complex figures: an fMRI comparison ofletter and fractal n-back tasks. Neuropsychology 16, 370–379.
Rama, P., Martinkauppi, S., Linnankoski, I., Koivisto, J., Aronen, H.J., Crlson, S., 2001.Working memory of identification of emotional vocal expression: an fMRI study.NeuroImage 13, 1090–1101.
Ricciardi, E., Bonino, D., Gentili, C., Sani, L., Pietrini, P., Vecchi, T., 2006. Neural correlatesof spatial working memory in humans: a functional magnetic resonance imagingstudy comparing visual and tactile processes. Neuroscience 139, 339–349.
Roland, P.E., Larsen, B., Lassen, N.A., Skinhøj, E., 1980. Supplementary motor area andother cortical areas in organization of voluntary movements in man. Journal ofNeurophysiology 43, 118–136.
Rolls, E.T., 2010. The affective and cognitive processing of touch, oral texture, and tem-perature in the brain. Neuroscience and Biobehavioral Reviews 34 (2), 237–245(Review).
Savini, N., Babiloni, C., Brunetti, M., Caulo, M., Del Gratta, C., Perrucci, M.G., Rossini, P.M.,Romani, G.L., Ferretti, A., 2010. Passive tactile recognition of geometrical shape inhumans: an fMRI study. Brain Research Bulletin 83, 223–231.
Schall, J.D., Hanes, D.P., Thompson, K.G., King, D.J., 1995. Saccade target selection infrontal eye field of macaque. I Visual and premovement activation. The Journal ofNeuroscience 15, 6905–6918.
Schumacher, E.H., Lauber, E., Jonides, J., Smith, E.E., Koeppe, R.A., 1996. PET evidence foran amodal verbal working memory system. NeuroImage 3, 79–88.
Smith, E.E., Jonides, J., 1997. Working memory: a view from neuroimaging. CognitivePsychology 33, 5–42.
Smith, E.E., Jonides, J., 1998. Neuroimaging analyses of human working memory. Pro-ceedings of the National Academy of Sciences of the United States of America 95,12061–12068.
Stoeckel, M.C., Weder, B., Binkofski, F., Buccino, G., Shah, N.J., Seitz, R.J., 2003. A fronto-parietal circuit for tactile object discrimination: an event-related fMRI study.NeuroImage 19, 1103–1114.
ory stimuli: An fMRI study, International Journal of Psychophysiology

9N. Savini et al. / International Journal of Psychophysiology xxx (2012) xxx–xxx
Talairach, J., Tournoux, P., 1998. Co-planar Stereotaxic Atlas of the human Brain.Thieme Medical Publisher, New York.
Thompson, K.G., Biscoe, J.D., Sato, T.R., 2005. Neuronal basis of covert spatial attentionin the frontal eye field. The Journal of Neuroscience 25, 9479–9487.
Tsakiris, M., 2010. My body in the brain: a neurocognitive model of body-ownership.Neuropsychologia 48 (3), 703–712.
Wager, T.D., Smith, E.E., 2003. Neuroimaging studies of working memory: a meta-analysis. Cognitive, Affective, & Behavioral Neuroscience 3, 255–274.
Walter, H., Bretschneider, V., Grön, G., Zurowski, B., Wunderlich, A.P., Tomczak, R.,2003. Special issue evidence for quantitative domain dominance for verbal andspatial working memory in the frontal and parietal cortex. Cortex 39, 897–911.
Please cite this article as: Savini, N., et al., Working memory of somatosens(2012), http://dx.doi.org/10.1016/j.ijpsycho.2012.09.007
Wise, R., Chollet, F., Hadar, U., Friston, K., Hoffner, E., Frackowiak, R., 1991. Distributionof cortical neural networks involved in word comprehension and word retrieval.Brain 114, 1803–1817.
Woolrich, M.W., Ripley, B.D., Brady, M., Smith, S.M., 2001. Temporal autocorrelation inunivariate linear modeling of fMRI data. NeuroImage 14, 1370–1386.
Xiao, Q., Barborica, A., Ferrera, V.P., 2006. Radial motion bias in macaque frontal eyefield. Visual Neuroscience 23, 49–60.
ory stimuli: An fMRI study, International Journal of Psychophysiology
Copyright © 2022 FDOKUMEN




















