
ASXL1 Mutations Promote Myeloid Transformation through Loss of PRC2-Mediated Gene Repression
Upload
khangminh22Category
view
0download
0
HAL Id: hal-03239787https://hal.archives-ouvertes.fr/hal-03239787
Submitted on 27 May 2021
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinée au dépôt et à la diffusion de documentsscientifiques de niveau recherche, publiés ou non,émanant des établissements d’enseignement et derecherche français ou étrangers, des laboratoirespublics ou privés.
Gene expression plasticity and frontloading promotethermotolerance in Pocillopora corals
K. Brener-Raffalli, Jeremie Vidal-Dupiol, M. Adjeroud, O. Rey, P. Romans, F.Bonhomme, M. Pratlong, A. Haguenauer, R. Pillot, L. Feuillassier, et al.
To cite this version:K. Brener-Raffalli, Jeremie Vidal-Dupiol, M. Adjeroud, O. Rey, P. Romans, et al.. Gene expressionplasticity and frontloading promote thermotolerance in Pocillopora corals. Peer Community Journal,Peer Community In/Centre Mersenne, 2022, 2, pp.e13. �10.24072/pcjournal.79�. �hal-03239787�

PEERCOMMUNITYINECOLOGY 1
RESEARCHARTICLE
Citeas:Brener-RaffalliK,Vidal-DupiolJ,AdjeroudM,ReyO,RomansP,BonhommeF,PratlongM,HaguenauerA,PillotR,FeuillassierL,ClaereboudtM,MagalonH,GélinP,PontarottiP,AurelleD,MittaGandToulzaE.GeneexpressionplasticityandfrontloadingpromotethermotoleranceinPocilloporacorals.bioRxiv398602,ver.4peer-reviewedandrecommendedbyPCIEcology(2019).Posted:4thSeptember2019Recommender:StaffanJacobReviewers:XavierPichonandMarSobralCorrespondence:[email protected]
GeneexpressionplasticityandfrontloadingpromotethermotoleranceinPocilloporacoralsBrener-RaffalliK.1,Vidal-DupiolJ.2,AdjeroudM.3,ReyO.1,RomansP.4,BonhommeF.5,PratlongM.6,8,HaguenauerA.8,PillotR.4,FeuillassierL.4,ClaereboudtM.9,MagalonH.10,GélinP.10,PontarottiP.6,7,AurelleD.8,11,MittaG.1andToulzaE.11 IHPE,Univ.Montpellier,CNRS, Ifremer,Univ.PerpignanviaDomitia,PerpignanFrance.2IHPE,Univ.Montpellier,CNRS,Ifremer,Univ.PerpignanviaDomitia,MontpellierFrance.3InstitutdeRecherchepourleDéveloppement,UMR9220ENTROPIE,Laboratoired'Excellence"CORAIL",UPVD52AvenuePaulAlduy,66860Perpignan,France.4 Sorbonne Universités, UPMC Univ Paris 06, UMS 2348, Centre de RessourcesBiologiquesMarines,ObservatoireOcéanologique,F-66650Banyuls/Mer,France.5UniversitédeMontpellier, InstitutdesSciencesde l'Évolution,CNRSUMR5554IRD-EPHE,Sète,France.6 Aix Marseille Univ, IRD, APHM, Microbe, Evolution, PHylogénie, InfectionIHUMéditerranéeInfection,MarseilleFrance.EvolutionaryBiologyteam.7CNRS8Aix-MarseilleUniversité,AvignonUniversité,CNRS,IRD,IMBE,Marseille,France9DepartmentofMarineScienceandFisheries,CollegeofAgriculturalandMarineSciences,SultanQaboosUniversity,Al-Khod,123,SultanateofOman.10 UMR 9220 ENTROPIE (Université de La Réunion, IRD, CNRS), Laboratoired’excellence"CORAIL",UniversitédeLaRéunion,StDenis,LaRéunion,France.11 Aix-Marseille Université, Université de Toulon CNRS, IRD, MIO, Marseille,France.
Thisarticlehasbeenpeer-reviewedandrecommendedbyPeerCommunityinEcology
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 2
Introduction
Earthisundergoingunprecedentedglobalenvironmentalchangeswithmajoreffectsonbiodiversity (Barnoskyet al. 2011). Theongoing erosionof themost vulnerableecosystemsduetocurrentenvironmentaldegradationisparticularlyworryingandisonlyapremisetowhatscientistshavecalledthesixthmassextinction(Barnoskyetal. 2011). In particular, climate change, ocean acidification and extreme climaticevents have already resulted in the irreversible degradation of more than 20% ofcoral reefs worldwide (Bellwood et al. 2004; Hoegh-Guldberg et al. 2007).Scleractinian corals constitute the biological and physical framework for a largediversityofmarineorganisms[c.a.~600coral,~2000fish,and~5000molluskspecies(Veron & Stafford-Smith 2000; Reaka-Kudla 2005)]. Hence, the extinction or evenmajor decrease of corals would have dramatic repercussions on the overallassociatedcommunities(Hughesetal.2017a).Naturalvariationinthermaltoleranceexists among coral populations (Oliver & Palumbi 2010; Palumbi et al. 2014),
ABSTRACTEcosystemsworldwideare suffering fromclimate change.Coral reef ecosystemsare globally threatened by increasing sea surface temperatures. However, geneexpressionplasticityprovides thepotentialfororganismstorespondrapidlyandeffectively to environmental changes, and would be favored in variableenvironments. In this study, we investigated the thermal stress response inPocilloporacoralcoloniesfromtwocontrastingenvironmentsbyexposingthemtoheat stress. We compared the physiological state, bacterial and Symbionaceaecommunities (using 16S and ITS2 metabarcoding), and gene expression levels(usingRNA-Seq)betweencontrolconditionsandheatstress(thetemperaturejustbelowthefirstsignsofcompromisedhealth).Coloniesfromboththermalregimesremainedapparentlynormalandpresentedopenandcoloredpolypsduringheatstress,withnochangeinbacterialandSymbionaceaecommunitycomposition.Incontrast,theydifferedintheirtranscriptomicresponses.ThecoloniesfromOmandisplayed a more plastic transcriptome, but some genes had a higher basalexpressionlevel(frontloading)comparedtothelessthermotolerantcoloniesfromNewCaledonia. In termsofbiological functions,weobservedan increase in theexpressionofstressresponsegenes(includinginductionoftumornecrosisfactorreceptors, heat shock proteins, and detoxification of reactive oxygen species),together with a decrease in the expression of genes involved in morpho-anatomical functions. Gene regulation (transcription factors, mobile elements,histonemodifications andDNAmethylation) appeared tobeoverrepresented intheOman colonies, indicatingpossibleepigenetic regulation.Theseresultsshowthat transcriptomic plasticity and frontloading can be co-occurring processes incoralsconfrontedtohighlyvariablethermalregimes.Keywords:Geneexpressionplasticity,Frontloading,Coralholobiont,Pocillopora,Heatstress.
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 3
especially alonga latitudinal gradient (Polatoetal. 2010;Dixonetal. 2015),henceproviding some hope for coral survival based on their capacity to cope with heatstress.Morespecifically,ithasbeenshownthatpopulationsinhabitinginzoneswithmorevariabletemperatureregimesdisplaybettertolerancetoheatstressfromlocal(Kenkeletal.2013)togeographicalscales(Hughesetal.2003;Riegletal.2011;Coles&Riegl2013).
It nevertheless remains unclear whether thermo-tolerance is acquired viaacclimation (i.e. intra-generational geneexpressionplasticity (Barnoskyetal. 2011;Kenkel & Matz 2016)) and/or through genetic adaptation (i.e. inter-generationalmicroevolution (Barnosky et al. 2011; Dixon et al. 2015)). Actually, some studiesstronglysuggestthatbothprocessesare likelytoco-occur inwildcoralpopulations(Hoegh-Guldbergetal.2007;Reusch2013;Palumbietal.2014;Hughesetal.2017a;Tordaetal.2017).
With the recent advancesofhigh throughputmolecularmethods, it is nowpossibletoprovideamorepreciseaccountofthemolecularmechanismsunderlyingcoralresponsetoheatstress.Inparticular,recentstudiesclearlydemonstratedthatcoralresponsestoheatstress involvethefine-tunedregulationofexpression levelsofsomegenes/proteinsinvolvedinseveralmolecularpathwayssuchasmetabolism,stress-responseandapoptosis(Brownetal.2002;Weis2008;Ainsworthetal.2011;Barnoskyetal.2011;Bellantuonoetal.2012a;Barshisetal.2013;Kenkeletal.2013;Palumbi et al. 2014). In this regard, twomainmolecular patterns having differenttemporalities have been put forward: (1) “transcriptional plasticity”, i.e. extensivechanges ingeneexpression levelsaccordingtotheoccurringthermalconditionand(2) “transcriptional frontloading”, i.e. theelevationof stress relatedgenesbaselineexpression thatpreconditionsorganisms tosubsequent (recurrent) stresses (Reaka-Kudla2005;Mayfieldetal.2011;Barnoskyetal.2011;Barshisetal.2013;Palumbietal. 2014; Hughes et al. 2017a). While such elevated constitutive gene expressionlevels could reflect local adaptation (i.e. genetically fixed gene expression level(Bellwoodetal.2004;Hoegh-Guldbergetal.2007;Oliver&Palumbi2010;Palumbietal. 2014), it could also reflect an acclimation via epigenetic processes leading toconstitutivegeneexpression(Veron&Stafford-Smith2000;Reaka-Kudla2005;Tordaet al. 2017). Epigenetic changes through environmental priming (i.e. translation ofenvironmentalcues)maybeinvolvedinadaptiveevolutionatsuchshorttimescales,eventually enabling transgenerational plasticity (Hughes et al. 2017a; Torda et al.2017;Jablonka2017).
Surprisingly, frontloading and gene expression plasticity were generallydiscussedasmutuallyexclusivepatterns(Oliver&Palumbi2010;Barshisetal.2013;Palumbi et al. 2014; Dixon et al. 2015; Kenkel & Matz 2016) although these twomolecular processes most likely co-occur during coral responses to heat stress. Inparticular, onemightexpect that the regulation strategyof genes (plasticity versus
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 4
frontloading) will greatly depend on the molecular pathways in which they areinvolvedandtheenergetic,physiological,andultimatelyfitnesscostassociatedwithgene expression. So far, frontloading has been detected for stress response genessuch as Heat Shock Proteins (HSPs), apoptosis and tumour suppression factors inresilient coral populations under experimentally simulated heat stress inducingbleachinginthecommonreef-buildingcoralAcroporahyacinthus(Polatoetal.2010;Barshisetal.2013;Dixonetal.2015;Kenkel&Matz2016)andformetabolicgenesinpopulationspre-exposedtowarmtemperaturesinresponsetolong-termheatstressinPoritesastreoides(Kenkeletal.2013;Palumbietal.2014).Conversely,inthelatterspecies,plasticitywasobserved in theexpressionofenvironmental stress responsegenes(Hughesetal.2003;Riegletal.2011;Coles&Riegl2013;Kenkel&Matz2016),hence challenging the patterns observed in A. hyacinthus (Barnosky et al. 2011;Barshis et al. 2013; Coles & Riegl 2013; Kenkel & Matz 2016). Although bothstrategies (i.e. constitutive frontloading versus expression plasticity) undoubtedlyexistinwildcoralpopulations,thepre-exposureconditionsthatfostertheirinductionand their relativeeffectsoncoral resistance toheat stress still remainunclear (butsee (Hughes et al. 2003; Barnosky et al. 2011; Dixon et al. 2015; Kenkel & Matz2016)).
Importantly, scleractinian corals are composed of several symbioticorganismsincludingthecnidarianhost,themutualistphotosyntheticalgae(formerlydefined as belonging to the genus Symbiodinium but now considered as differentgenerawithinthefamilySymbionaceae(Bellwoodetal.2004;Barnoskyetal.2011;Dixonetal.2015;LaJeunesseetal.2018))andbacterialcommunities.Allsymbiontsinvolved in a stable symbiosis effectively form the entire organism, and constitutewhat is referred to the holobiont (Margulis & Fester 1991; Hoegh-Guldberg et al.2007;Reusch2013;Palumbietal.2014;Tordaetal.2017).Adecadeafterthistermwasdefined,itsusehasbeenparticularlypopularizedinreferencetocorals(Rohweretal.2002),andsubsequentresearchhasledtothehologenometheoryofevolution(Rosenberg et al. 2007; Zilber-Rosenberg & Rosenberg 2008). In this context, thehologenome is defıned as the sum of the genetic information of the host and itssymbiotic microorganisms. Phenotypes are thus the product of the collectivegenomes of the holobiont partners in interaction with the environment, whichconstitutetheunitofbiologicalorganizationandthustheobjectofnaturalselection(Zilber-Rosenberg&Rosenberg2008;Guerreroetal.2013;McFall-Ngaietal.2013;Bordenstein & Theis 2015; Theis et al. 2016). Additionally to the cnidarian hostresponse,thegenotype-orassociationofgenotypes-ofthephotosyntheticmutualistSymbionaceae symbionts plays a key role in the thermotolerance of the holobiont(Humeetal.2013;Mayfieldetal.2014;Suggettetal.2017).There is lesscertaintyabouttheimportanceofthecoralbacterialcommunityinparticipatingtothefitnessof the holobiont, although accruing evidences strongly suggest their implication in
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 5
coral response to environmental conditions (Li et al. 2014; Pantos et al. 2015;Hernandez-Agredaet al. 2016), and in the resistance to diseases (Satoet al. 2009;Cróquer et al. 2013; Meyer et al. 2016). Finally, the role of the coral-associatedmicroorganismsandtheirpotentialtomodifyholobiontresponsetostressremainsofaroverlooked(butsee(Ziegleretal.2017;Tordaetal.2017).Hence,studyinghowcorals respondtostress impliesan integrativeapproach toanalyze theresponseofeachsymbioticprotagonist.
With this aim, we investigated the molecular mechanisms underlyingthermo-toleranceofcoralholobionts.Weanalyzedtheholobiontresponsetostressof two coral populations originating from environments with contrasting thermalregimes.Weused scleractinian corals from the genusPocilloporaasmodel speciesbecausetheyhaveabroadspatialdistributionthroughout the Indo-Pacific (Veron&Stafford-Smith 2000). The genus Pocillopora is considered to be one of the mostenvironmentally sensitive (vanWoesik et al. 2011) but its widespread distributionclearlysuggestspotentialforacclimationand/oradaptationwhichmaybecorrelatedto specific differences (i.e. different cryptic lineages may be adapted to differentenvironmental conditions). In particular, we focused on Pocillopora damicornis-likecolonies from two localities with contrasting thermal regimes: colonies from NewCaledonia (NC) are exposed to temperate and stable temperatures over the year,while those from Oman are exposed to globally warmer and more seasonalfluctuating temperatures. As the corallummacromorphology is not a discriminantcharacterinPocilloporaandasthetaxonomicrevisionofthisgenususingmoleculardatarevealsthatsomeofthePocilloporaspecies(Schmidt-Roachetal.2014;Gélinetal. 2017b) are actually species complexes,we identifieda posteriori the species ofthe sampled colonies (mitochondrial ORF sequencing and individual clustering) inorder to interpret the results in a precise evolutionary context. To avoid biasesinherent in transplantation-based field experiments resulting from environmentalfactorsotherthantemperature,weundertookourcomparativestudyinacontrolledenvironmentinwhichwemimickedecologicallyrealisticheatstresstocomparetheresponsesofcoloniesfrombothlocalities.WecombinedaspecificRNA-seqapproachto study the cnidarian host response, andmetabarcoding analyses using ITS-2 and16S amplicon sequencing to study the dynamics of the associated algal(Symbionaceae) and bacterial community compositions, respectively. According tothe literature we first expected to detect changes in both symbiotic algal andbacterial communities in corals from both localities when exposed to heat stress.Moreover, since variable environments are expected to select for plasticity, wepredicted that the cnidarian hosts from Oman may display more gene expressionplasticity than those fromNewCaledonia.However, because frontloadingwasalsofoundtobeanalternativeresponsetorecurrentchangingconditions,wemightalso
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 6
expectsomedegreesofconstitutivehighlevelsofgeneexpressionatleastforsomemolecularpathwaysandmoreparticularlyinOmancorals.
MaterialandMethods
CoralsamplingandmaintenancePocilloporadamicornis-likecoloniesoriginating fromenvironmentscharacterizedbycontrasting thermal regimes were sampled during the warmer month in twodifferentlocalities:(1)inOman,GulfofOman,NorthwesternIndianOcean(Om;June2014;localseawatertemperatureduringsampling30.8°C),wherecoralsareexposedto a globallywarmerand variable thermal environment, and (2) inNewCaledonia,SouthwesternPacificOcean(NC;November2014;localseawatertemperatureduringsampling27.1°C),wherecoralsaresubjecttomoremitigateandstabletemperatures(seeTable1forthesamplingsitesandTable2fortemperatureregimeoflocality).
Table1.SamplingsitesLocality Colony GPS
NewCaledonia
NC1 22°18’919S166°26’333E
NC2 22°17’768S166°26’209E
NC3 22°20’886S166°21’952E
Oman
Om1 23°30’806N58°45,340E
Om2 23°31’287N58°44’995E
Om3 23°37’258N058°36’003E
Table2.SeaSurfaceTemperatures(SST)towhichthecoloniessampledinthisstudyareexposedintheirnaturalenvironments.Thermalregimedescriptorswere compiled from weekly mean sea surface temperature data collectedfrom the IntegratedGlobalOceanServices SystemProductsBulletin (IGOSS:http://iridl.ldeo.columbia.edu/SOURCES/.IGOSS/)forquadratsof1°longitudex1°latitudefrom1982totheyearofsampling(2014).
NewCaledonia Oman
MeanSST(°C) 24.8 27.9
Variance(°C) 2.7 9.5
MinSST(°C) 22.6 22.1
MaxSST(°C) 27.1 33.2
MeanSSTof3warmermonths(°C) 26.8 31.3
MeanSSTof3coolermonths(°C) 22.8 23.8
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 7
From each location, we thus sampled coloniesmorphologically similar andoccupying the same water depth niche. To account for possible intra-populationdiversity, three colonies (>20cm in diameter) were collected in each locality, andseparated by at least 10m to decrease the probability to collect members of thesame genet, as some Pocillopora species are able to propagate by asexualreproduction(Adjeroudetal.2013;Gélinetal.2017a;2018).
Immediately following collection, a 1cm branch tip of each colony wasexcised, rinsed three times in filtered seawater (0.22µm), and placed in RNAlatersolution(SigmaAldrich)fortheinsitumicrobiotaanalysis.Therestofthecolonywasfragmented into 20branches eachof 10cm length andphysiologically stabilized inopenwater system for oneweek before shipping (Al-Hail field stationof the SultanQaboos University and the Public aquarium of Noumea for OM and NC localitiesrespectively).Forshipping,individualbrancheswereplacedinplasticbagscontainingoxygenated seawater (800mL seawater and 1600mL of medical oxygen), andtransported by aircraft to the research aquarium of the Banyuls-sur-Meroceanographicobservatory(France).Thecoralbranchesweremaintainedinartificialseawater (Seachem Reef Salt) at 26°C, and supplied daily with Artemia nauplii tosatisfy their heterotrophic demand. The conditions in the maintenance tank werecontrolled tomimic thenatural physicochemical parameters of coral reefs (pH:8.2;salinity:36psu; light intensity:150to250µmolofphotons/m²/s;photoperiod:12hnight/12h day;kH: 6–7.5dkH; calcium concentration: 410–450mg/L; inorganicphosphate concentration: <0.1mg/L; magnesium concentration: 1300–1400mg/L;nitrate concentration: <5mg/L). After 3 and 7 months of acclimatization to thelaboratory condition (marked by growth resumption) for Om and NC colonies,respectively,coralswerefragmentedtoproduceatotalof~15to20clones(nubbins)fromeachcolony(~3cm).Thesewere individually fixedtoasupport (hereap1000tip) using an epoxy adhesive. We waited for complete healing (evident as tissueextendingtocovertheepoxyadhesive)priortoruntheexperiment.EcologicallyrealisticheatstressTheaimof thisexperimentwastocomparetheresponsetoheatstressofcoloniesfromtwolocalitieshavingthesamephysiologicalstate,toinvestigatethepatternsofexpression of themolecular pathways involvedduring the stress exposure and theputativemodificationsofthecoralmicrobiota.
The experimental design comprised four tanks of 53L per locality inwhichtheseawaterwascontinuouslyrecycled.ThewaterwassterilizedusingUV(rate3200L/h) and renewed twice per hour in each tank (recirculation rate: 100L/h in eachtank). The eight tanks shared the same seawater but their temperature wasmonitored individually (HOBBY BiothermPro, model 10892; 500W Aqua Medictitaniumheater;HOBOTidbiTv2logger)(SupplementaryFigureS1).Foreachlocality,
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 8
5 to8nubbinspermothercolonieswere randomlyplaced ineach tank (four tanksperlocality)fortwoweeksatthecontroltemperatureandthefollowingprotocolwasapplied: three tankswere thensubjected toagradual temperature increase (stresstreatment)whilethefourth(control)wasmaintainedatthecontroltemperaturetoverify that the stress observed in the stressful treatment was not due to otherpotential confounding effects orwater cues (Figure 1). Both the control and stresstemperatures were specific for each sampling locality to mimic their respectivenatural environment. In particular, we set the control temperature as the meanwatertemperatureforthethreewarmermonthsmeasuredatthecoralsamplingsitelocality(Table1):31°CforthecoloniesfromOm,and27°CforthecoloniesfromNC.The stress treatment was ecologically realistic, i.e. reflecting a naturally occurringwarming anomaly, and consisted in increasing the temperature gradually by 1°C(over 5 consecutive hours) each week until physiological collapse of the coralsbecameevident(polypsclosure,bleachingornecrosis),asdescribedby(Vidal-Dupioletal.2009).Samplingwasperformedinthethreesamplingtanksjustbeforethefirsttemperature increase (control condition) as well as each week before the nexttemperatureincrease.Thebeginningofpolypclosurewasconsistentlyobservedforthe different colonies of the same locality at the same temperature threshold.Samples for subsequent genetic and transcriptomic analyses were chosen aposteriori. They corresponded to those sampled in each tank just before the firstincrease of temperature (control samples), and just before the temperature thatproduced the first signs of physiological collapse and before bleaching (stresstemperature samples). Thus, for each condition (control and stress) we obtainedthree biological replicates of each colony from the three different tanks (threecolonies per locality) to reach a total of 36 samples (2 localities × 3 colonies× 2experimentalconditions×3replicates/tanks).Thegeneralhealthofthenubbinswasassessedviadailyphotographicmonitoring(atnoonpriortofeeding)throughouttheperiodoftheexperiment.
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 9
Figure 1. The ecologically realistic heat stress experiment: from meantemperatures of the warmer months in natura to a pre-bleachingphysiological state. Nubbins were collected at each time point and arrowsrepresent points at which nubbins were chosen for analyzing themicrobialcompositionandthetranscriptomicresponseofthehost.DNAextractionDNAwasextracted fromeach36samplesaswellascoral tipsdirectlycollectedonthesixcolonies innatura for the insitu condition (three inOm, three inNC),usingtheDNeasyBloodandTissuekit(Qiagen)followingthemanufacturer’s instructions.DNAwasquantifiedbyspectrophotometry(NanoDrop).HostspeciesandclonematesidentificationAsthecorallummacromorphology isnotadiagnosticcriterion inPocilloporagenus,thehostspecieswasthusidentifiedmolecularly.Thuseachcolonywassequencedforthemitochondrial variable open reading frame (ORF) andwas genotyped using 13specificmicrosatellites,asinGélinetal.(Gélinetal.2017b).Theneachcolonyusedin theexperimentwasassigned toPrimaryandSecondarySpeciesHypothesis (PSHand SSH; sensu Pante et al.) (Pante et al. 2015) following the nomenclature fromGélin et al. (Gélin et al. 2017b). Indeed, sampling Pocillopora colonies presentingvariousmorphs fromdifferent locations from the Indo-Pacific,Gélin et al classifiedthese colonies,withouta prioribased on corallummacromorphology, into SpeciesHypotheses(sensuPanteetal.,i.e.thespeciesarehypothesesthatcanbeconfirmedor refuted while new data are added) (Pante et al. 2015) using sequence-basedspecies delimitation methods, a first sorting allowed to define Primary SpeciesHypotheses (PSH) and then individual clustering based onmicrosatellitemultilocus
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 10
genotypesallowedasecondsortingdelimitingSecondarySpeciesHypotheses(SSH).ThuscomparingtheORFsequencesobtainedinthisstudytothosefrom(Gélinetal.2017b),thesampledcolonieswereassignedtoaPSH.Then,ifrelevant,thecolonieswereassignedtoSSHperformingclusteringanalysisusingStructure2.3.4 (Pritchardetal.2000),asin(Gélinetal.2017b).Meanwhile,theidenticalmulti-locusgenotypes(i.e. clonemates if any) were identified by microsatellite analysis using GenClone(Arnaud-Haond&Belkhir2006)asinGélinetal.(Gélinetal.2017a).MicrobialcommunityanalysisusingMiSeq16SandITS2metabarcodingTheaimofthisanalysiswastoinvestigatethecompositionandthedynamicsofthetwoprincipalsymbioticcoralcommunities(i.e.bacterialandalgal)insituandduringheatstress.AmpliconSequencingAbacterial16SrDNAampliconlibrarywasgeneratedforeachofthe42samples(onein situ condition, three control conditions and three stress conditions per colony,threecoloniesperlocality,twolocalities),usingthe341F(CCTACGGGNGGCWGCAG)and 805R (GACTACHVGGGTATCTAATCC) primers, which target the variable V3/V4loops (Klindworth et al. 2012). The Symbiodiniaceae assemblages were analyzedusingITS2(internaltranscribedspaceroftheribosomalRNAgene)ampliconlibrariesand specific primers targeting a sequence of approximately 350bp (ITS2-FGTGAATTGCAGAACTCCGTG; ITS2-R CCTCCGCTTACTTATATGCTT) (Lajeunesse &Trench2000;Quigleyetal.2014).Forbothmarkers,paired-endsequencingusinga250bp read length was performed on the MiSeq system (Illumina) using the v2chemistry, according to the manufacturer’s protocol at the Centre d'InnovationGénomeQuébecandMcGillUniversity,Montreal,Canada.BioinformaticanalysisThe FROGS pipeline (Find Rapidly OTU with Galaxy Solution) implemented on aGalaxy platform (http://sigenae-workbench.toulouse.inra.fr/galaxy/) was used fordataprocessing(Escudiéetal.2017).Inbrief,pairedreadsweremergedusingFLASH(Magoč & Salzberg 2011). After cleaning and removal of primer/adapters usingcutadapt (Martin2011),denovo clusteringwasperformedusingSWARM (Mahéetal.2014).Thisusesalocalclusteringthresholdwithanaggregationdistance(d)of3.ChimeraswereremovedusingVSEARCH(Rognesetal.2016).Wefilteredthedatasetfor singletons and performed affiliation using Blast+ against the Silva database(release 128, September 2016) for 16S amplicons (Altschul et al. 1990). For ITS2metabarcoding, the Symbiodiniaceae typewas assessed using Blast+ against an in-house database of Symbiodiniaceae reference sequences built from sequences
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 11
publiclyavailable.AnOTUtable instandardBIOMformatwithtaxonomicaffiliationwasproducedforsubsequentanalyses.ForcommunitycompositionanalysisweusedthephyloseqRpackage(McMurdie&Holmes2013)toinferalphadiversitymetricsattheOTUlevel(i.e.ShannonandChaoindexes), and beta diversity (between sample similarity) from the OTU table.CommunitysimilaritywasassessedbyPrincipalCoordinateAnalysis(PCoA)usingtheBray-Curtisdistancematrices.Totestforpossibleeffectofcorals’localityoforiginandfortemperaturetreatmentson the twoalphadiversitymetrics (i.e. ShannonandChao)we rana seriesof fourgeneralizedmixedmodelsincludingthesetwofixedfactorsandtheirinteractionsasexplanatoryvariables.The fourmodels consisted in consideringall factors, the twofixed factorswithoutaccounting for theirpossible interactions,andthetwofactorsindependently.Inallmodels,wealsoaccountedforpossibleeffectofgenotypesthatwereconsideredasrandomfactorsnestedwithinlocality.Allmodelswererunusingthe lme4 package implemented in R (Bates et al. 2015) and considering a Gammadistribution.ThebestmodelforeachalphadiversitymetricswasidentifiedbasedontheAkaike InformationCriterion (AIC (Akaike1974)). Concerningbetadiversity,weperformed MANOVAs to compare beta diversity metrics among the groups ofsamplesbysamplinglocality,genotypeortreatment.Corrections based on multiple testing were performed using FDR (Benjamini &Hochberg1995).Forallanalyses,thethresholdsignificancelevelwassetat0.05.TranscriptomeanalysisTheaimofthisanalysiswastostudythetranscriptomesofthesampledcolonies inresponsetoheatstresscomparedwithcontrolledconditions.RNAextractionTotal RNAwas extracted from each coral sample using TRIzol reagent (Invitrogen),accordingtothemanufacturer'sprotocol.ThequantityandintegrityofthetotalRNAextracted was checked using an Agilent 2100 Bioanalyzer (AgilentTechnologies)(mean RIN=7.5). Paired-end fragment libraries (2×100bp) wereconstructed and sequenced on an Illumina HiSeq 2000 platform at the Centred'InnovationGénomeQuébecatMcGillUniversity,Montreal,Canada.BioinformaticanalysesFastq read files were processed on the Galaxy instance of the IHPE(http://bioinfo.univ-perp.fr)(Giardineetal.2005).Qualitycontrolandinitialcleaningofthereadswereperformedusingthefilterbyqualityprogram(version1.0.0)basedontheFASTX-toolkit(Blankenbergetal.2010).Readshavingfewerthan90%ofbaseshaving a Phredquality score ≤26werediscarded (probability of 2.5 incorrect base
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 12
calloutof1000,andabasecallaccuracyof99.75%).Adaptorsusedforsequencingwereremovedusingthecutadaptprogramversion1.6(Martin2011).Allpaired-endreadswerealignedusingRNAstar softwareunderdefaultparameters,withat least66%of the bases being required to align to the reference, and amaximumof tenmismatches per read (Dobin et al. 2013). The Pocillopora damicornis sensu latoreference genomeused in this study (Vidal-Dupiolet al. 2019) consistedof a draftassembly of 25,553 contigs (352Mb total) and N50=171,375bp. The resultingtranscriptomeservedasthereferenceforreadsmapping,andaGTFannotationfilewas constructedusing cufflink/cuffmerge (Trapnelletal. 2010).HTseqwasused toproducecountfilesforgenes(Andersetal.2015).TheDESeq2packagewasusedtoestimatethenormalizedabundancesofthetranscripts,andtocalculatedifferentialgene expression for samples between the control temperature and the stresstemperatureforeachlocality(Loveetal.2014),consideringthedifferentgenotypes(threebiologicalreplicatesforeachgenotype)andusingdefaultparameters.Wenextanalyzedgenesaccordingto theirexpressionpatternsamongthedifferentcoloniesand temperature treatments. Genes were clustered manually into six groupsaccordingtotheirdifferentialexpressionlevels:commonover-expressedgenes,NC-specific over-expressed genes, Om-specific over-expressed genes, common under-expressed genes, NC-specific under-expressed genes, and Om-specific under-expressed genes. Cluster 3.0 (de Hoon et al. 2004) and Treeview (Saldanha 2004)wereusedtobuildtheheatmap.Discriminantanalysisofprincipalcomponents(DAPC)Our aim was to quantify and compare the level of genome-wide transcriptomeplasticitybetweencolonies fromOmandNC in responsetoheatstress.Toachievethisweperformedadiscriminantanalysisofprincipalcomponents(DAPC)basedonalog-transformed transcript abundance matrix (containing 26,600 genes) obtainedfrom the 36 samples (i.e. 9 control and 9 stressed replicates per locality), asdescribedpreviously(Kenkel&Matz2016).Specifically,weranaDAPCanalysisusingthe resulting log2 transformeddataset for the colonies fromNC andOm reared incontrolledconditionsaspredefinedgroupsintheadegenetpackageimplementedinR(Jombartetal.2010).Twoprincipalcomponentsandasinglediscriminantfunctionwere retained. We then predicted the position of stressed colonies from bothlocalities(OmandNC)ontotheuniquediscriminantfunctionoftheDAPC.We next ran a general linear model (GLM) using the DAPC scores as dependentvariable,andaccountedforthelocalityoforigin(NCandOm),thetreatment(controlandheatstress),andtheir interactionasexplanatoryvariables.Wealsoconsideredthe effect of individual colonies nested within localities as random effects in themodel. Our final objective was to test for a potential significant effect of theinteractionbetweenthe localityandthetreatmenteffects,asaproxyofsignificant
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 13
differences in thegenome-widegeneexpression reactionnorms (i.e. differences inDAPCscoresbetweenthecontrolandtheheatstresstreatments)betweenOmandNCcolonies.GOenrichmentofdifferentiallyexpressedgenesThetranscriptomewasannotateddenovousingatranslatednucleotidequery(blastx(Altschuletal.1990))againstthenon-redundantproteinsequencedatabase(nr).The25best hitswere thenused to search for geneontology termsusing theBlast2Goprogram (Conesa et al. 2016). To identify the biological functions significantlyenrichedwithinupordown-regulatedgenes,aGeneOntology(GO)termenrichmentanalysiswasperformed.ListsofGOtermsbelongingtosignificantlyup-regulatedanddown-regulatedgeneswerecomparedtotheGOtermsofthewholeexpressedgenesetusingaFischerexacttestandaFDRvalueof0.05.WeusedREVIGOtovisualizetheenrichedbiologicalprocesses(Supeketal.2011).
Results
HostidentificationAmong the three colonies from New Caledonia, colonies NC2 and NC3 presentedhaplotypeORF18andwereassignedtoPrimarySpeciesHypothesisPSH05andmorepreciselytoSecondarySpeciesHypothesisSSH05a(Gélinetal.2017b),correspondingtoP.damicornis typeβ (Schmidt-Roachetal.2014)or type5a (Pinzónetal.2013),whilecolonyNC1presentedORF09andwasassignedtoPSH04,P.damicornistypeα,P.damicornisortype4a,respectively.AsforcoloniesfromOman,theyallpresentedORF34andwereassignedtoPSH12(Gélinetal.2017a)ortype7a(Pinzónetal.2013)(Supplementary Table S2), that is not part of the P.damicornis sensu lato speciescomplex. Thus NC colonies are phylogenetically closer from each other than fromcoloniesfromOman.ThesethreePSHsrepresentthreedifferentspecies.
Furthermore,NC2andNC3multi-locusgenotypes(MLGs)differedonlyfromonealleleover26genecopies,andwerethuspartofthesameclonallineage(genet),i.e. the entity that groups together colonies whose multi-locus genotypes slightlydiffer due to somaticmutations or scoring errors. All the other colonies presentedMLG that differed enough not to be considered as clonemates ormembers of thesameclonallineage(genet).EcologicallyrealisticheatstressOurgoalwastoensurethatourexperimentalheatstressfaithfullyreflectsarealisticheatstressinnatura.Followingcollectionfromthefield,thecoralsfromthedifferentlocalitieswerefirstmaintainedinthesamecontrolledconditionsat26°Cpriortotheexperiment. During this period no mortality or signs of degradation/stress were
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 14
observed for any of the coral colonies. Two weeks before the experiment, a firstacclimatization to the control temperatures (27°C or 31°C for NC and Omrespectively) was performed. During the experimental heat stress (i.e. gradualtemperatureincrease),visualandphotographicmonitoringclearlyindicatedthatthefirst sign of coral stress (i.e. the closure of polyps) occurred at day 30 for bothsampling localities, corresponding to 30°C and 34°C for the NC and Om colonies,respectively. These temperatures perfectly match the warmest temperatureexperiencedbythesecoloniesinthefield(Table1).Nosignofphysiologicalcollapsewere observed in control corals throughout the experiment indicating that all theotherparametersweremaintainedoptimalforcoralcolonies.BacterialcommunitiesAmong the overall 42 samples analyzed, a total of 5,308,761 16S rDNA ampliconsequences were obtained after cleaning and singleton filtering corresponding to15,211OTUs. InallsamplestheclassGammaproteobacteriawasdominant (77.7%),particularlythegenusEndozoicomonas(44.7%ofthesequences);thisgenusisknownto be an endosymbiont of numerous scleractinians (Neave et al. 2016b) (SeeSupplementary Figure S3 for complete bacterial composition in each colony andreplicate).
Concerningthetwoalphadiversitymetrics(i.e.ShannonandChaoindexes),thebeststatisticalmodelstoexplaintheoverallvarianceincludedthecoral’slocalityof origin only (AIC = 101.9). According to these models, colonies from Omandisplayed a higher Shannon index than those from New Caledonia (P = 0.04).Conversely, the effect of localitywas not significant for the Chao index (P > 0.05).ThePCoAofBray-Curtisdistancesforallcoloniesshowednoevidentclustersbasedontheexperimentaltreatments(Figure2).Weobserveda loosegroupingbasedonlocalities and colonies, except for colony NC1, which appeared to have a morespecificmicrobiota composition, as it had a different grouping associatedwith thefirst axis,whichexplained22.3%of the variance. This couldbe correlatedwith thedifferentspecieshypothesesforNC1comparedtoNC2andNC3(seeabove).Theseobservations were confirmed by statistical analyses (MANOVA between localities:P=0.003; between genotypes:P=0.001; between temperature treatments:P=0.761;SupplementaryTableS4).Thus, thebacterialcompositionappearedtoberelativelyspecifictoeachcolonywithineach locality,butnomajorshiftwasobservedduringthe transition from the natural environment to artificial seawater, nor during heatstressexposure.
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 15
Figure 2. Principal coordinate analysis plot for Bray-Curtis distances of thebacterial composition of each colony in each experimental condition.Different colors represent different colonies, the stars represent the in situconditions,theopencirclesrepresentthecontrolconditions,andthesquaresrepresentthestressconditions.SymbiodiniaceaeassemblagesAnalysis of the Symbiodiniaceae composition was performed based on an ITS2metabarcoding,whichallowedintra-claderesolution.
Removal ofOTUshaving an abundanceof <1% left only 4OTUs among allsamples.TwoofthesecorrespondedtotypeC1,whiletheothertwocorrespondedtotypeD1aaccordingto(Baker2003).TypeD1awashighlydominantinthecoloniesoriginatingfromOman,whereastypeC1wasalmostexclusivetothecoralsfromNewCaledonia(Figure3).TheSymbiodiniaceaecommunitycompositionwasveryspecificto each locality, but remained stable during the transition from the naturalenvironmenttoartificialseawater,andduringheatstressexposure.
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;
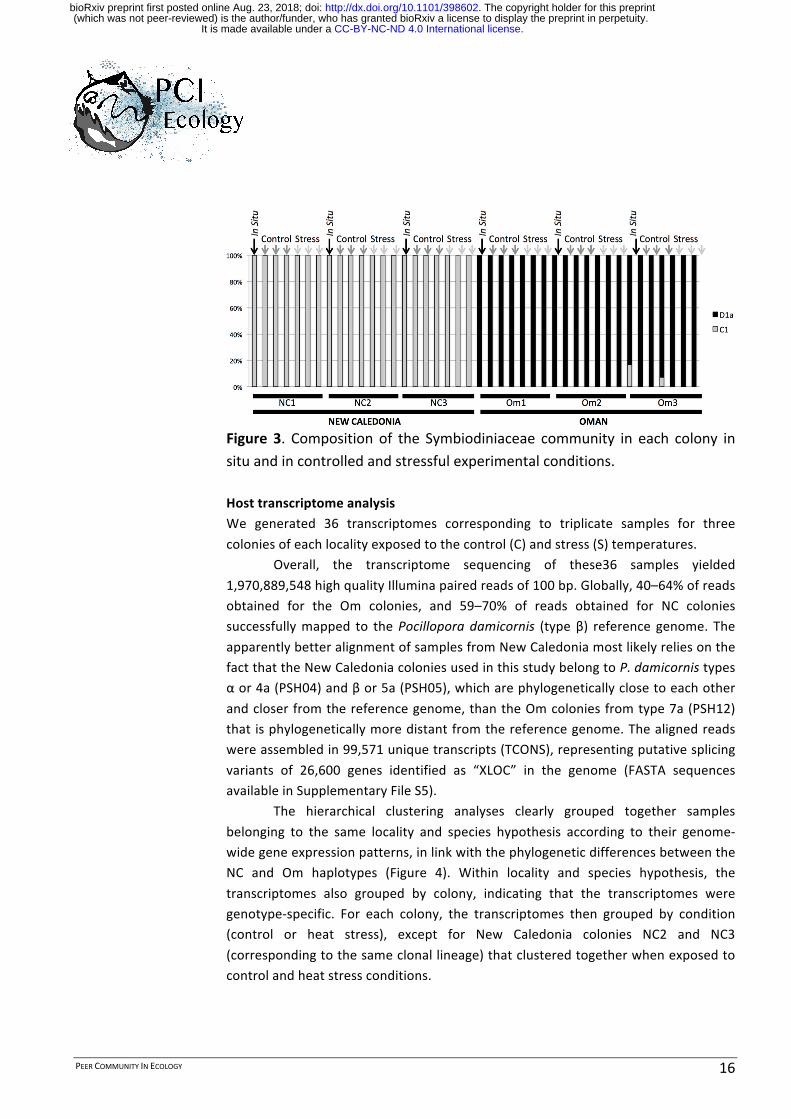
PEERCOMMUNITYINECOLOGY 16
Figure 3. Composition of the Symbiodiniaceae community in each colony insituandincontrolledandstressfulexperimentalconditions.HosttranscriptomeanalysisWe generated 36 transcriptomes corresponding to triplicate samples for threecoloniesofeachlocalityexposedtothecontrol(C)andstress(S)temperatures.
Overall, the transcriptome sequencing of these36 samples yielded1,970,889,548highqualityIlluminapairedreadsof100bp.Globally,40–64%ofreadsobtained for the Om colonies, and 59–70% of reads obtained for NC coloniessuccessfullymapped to thePocillopora damicornis (typeβ) reference genome. TheapparentlybetteralignmentofsamplesfromNewCaledoniamostlikelyreliesonthefactthattheNewCaledoniacoloniesusedinthisstudybelongtoP.damicornistypesαor4a(PSH04)andβor5a(PSH05),whicharephylogeneticallyclosetoeachotherandcloserfromthereferencegenome,thantheOmcoloniesfromtype7a(PSH12)that isphylogeneticallymoredistantfromthereferencegenome.Thealignedreadswereassembledin99,571uniquetranscripts(TCONS),representingputativesplicingvariants of 26,600 genes identified as “XLOC” in the genome (FASTA sequencesavailableinSupplementaryFileS5).
The hierarchical clustering analyses clearly grouped together samplesbelonging to the same locality and species hypothesis according to their genome-widegeneexpressionpatterns,inlinkwiththephylogeneticdifferencesbetweentheNC and Om haplotypes (Figure 4). Within locality and species hypothesis, thetranscriptomes also grouped by colony, indicating that the transcriptomes weregenotype-specific. For each colony, the transcriptomes then grouped by condition(control or heat stress), except for New Caledonia colonies NC2 and NC3(correspondingtothesameclonallineage)thatclusteredtogetherwhenexposedtocontrolandheatstressconditions.
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 17
Figure 4. Hierarchical clustering analyses performed using DESeq2 rlog-normalizedRNA-seqdata for the36 transcriptomes: twoconditions (controlandheatstress);threereplicatesperconditionforeachcolony;threecoloniesper locality; and two localities [Oman (Om) and New Caledonia (NC)]. Thecolor (from white to dark blue) indicates the distance metric used forclustering(darkbluecorrespondstothemaximumcorrelationvalues).
Despite clustering of the transcriptomes by locality, as the sampling of
Pocillopora damicornis-like colonies actually corresponded to different species weperformed differential gene expression analysis for each colony independently(comparing the biological triplicates for the control condition vs. triplicates for theheat stress conditions). For each locality, the different colonies displayed similarpatterns of differential gene expression with in any case a higher number ofdifferentially expressed genes and higher fold-changes between control and heatstressconditioninOmcomparedtoNC(SupplementaryFigureS6).Wedetected673,479 and 265 differentially expressed genes forNC1,NC2 andNC3 respectively, vs.2870,2216and959 forOm1,Om2andOm3.Sampleswere thusgrouped foreachlocality (nine control nubbins + nine heat stress nubbins) for subsequent analyses(fullresultsofthecomparisonsbetweenstressedandcontrols(log2-foldchangeandadjusted p-values) for each colony or between the two localities are provided inSupplementary File S7). ForOmcolonies, a total of 5,287 genesweredifferentiallyexpressedbetweencontrolandstressconditions(Figure5).ThisnumberwasmuchlowerforNCcolonieswith1,460differentiallyexpressedgenes(adjustedP<0.05).
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 18
Figure 5. Heatmap and clustering of significantly differentially expressedgenes between the control and the heat stress condition for colonies fromeachlocality.Eachgeneisrepresentedbyarow.
Among differentially expressed genes, 848 were differentially expressed in
thesamedirectioninbothlocalities(498over-expressedand350under-expressed).In both cases, the differential expression level was significantly higher for the Omcoralswithameanlog2-foldchangeof0.9forsharedover-expressedgenesinOmvs.0.6inNC(Wilcoxontest;P<0.0001),and-1.2forthesharedunder-expressedgenesinOmvs. -0.8 inNC(Wilcoxontest;P<0.0001) (Figure6andSupplementaryTableS8).
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 19
Figure6.Scatterplotofthelog2-foldchangesingeneexpressioninresponsetoheatstress in theOmcolonies (y-axis)vs. theNCcolonies (x-axis) for the848 genes that were over-expressed (498 genes) or under-expressed (350genes) in colonies from both localities. The line represents the y=x linedepictingsimilarresponsesbetweencolonies.
Additionally, colonies from the two localities also responded specifically to heatstress. Inparticular,272geneswereover-expressedand294wereunder-expressedonly in the NC corals, whereas 2,082were over-expressed and 2,311were under-expressed only in the Om ones when exposed to heat stress. Finally, the coloniesfrombothlocalitiesdisplayedantagonistictranscriptomicresponsestoheatstressforasmallsubsetofgenes(24over-expressedinNCbutunder-expressedinOm,and22under-expressedinNCbutover-expressedinOm).
Altogether these results revealedagreater transcriptomic response toheatstress in colonies originating from Oman compared to those from New Caledonia(4,393 differentially expressed genes for the Om corals vs. 566 genes for the NCones).
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 20
DiscriminantAnalysisofPrincipalComponents(DAPC)At the overall gene expression level, our DAPC analysis clearly discriminated thecolonies from both localities (Figure 7). More interestingly, the GLM revealed asignificant interaction term between the locality and condition (control or heatstress) effects (P = 0.04), hence indicating that the slopeof the reactionnormwasdifferent between localities. More particularly, the Om colonies responded to agreater extent than the NC ones, and thus showed significantly higher geneexpressionplasticityinresponsetoheatstress.
Figure 7. Colony level gene expression variation in response to heat stress,based on DAPC analysis. The x-axis is the first discriminant function of theDAPCalongwhichtheoverallgeneexpressiondifferencebetweencoloniesatbothexperimentalconditions(stressandcontrol)andfrombothlocalities(NCandOm)wasmaximized.This indicatesthedegreeofsimilaritybetweenthetranscriptomes. The density plots obtained for NC and Om colonies arerepresented in blue and green, respectively. Dark and light density plotscorrespond to samples from thecontroland stressexperimental conditions.The arrows above the density plots represent the direction of the meanchangeinthegeneexpressionprofiles.
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 21
It is worth stressing that colonies having experienced the heat stressdisplayed more similar genome-wide expression profiles than controlled colonies(Figure 7). Such pattern was also observed in colonies of mustard hill coral P.astreoides experiencing a heat stress compared to controls (Kenkel &Matz 2016).This apparent convergence in the functional response of colonies from differenthabitats to heat stressmight be at least partly explain by the fact some commonmolecular pathways are turned-on when colonies are facing stressful conditionsalthoughthemagnitudeofsuchresponsesisdifferent.AnalysisofgenefunctionTo investigate the functions associated with the differentially expressed genes weperformed a BLASTX annotation of transcripts followed by a Gene Ontology (GO)term(biologicalprocess,molecularfunction,andcellcompartment)(SupplementaryFileS9).
For the 498 common over-expressed genes, 139 biological processes wereenriched compared to the entire set of annotated genes. The most significantbiologicalprocess identified intheREVIGOanalysis(i.e.with lowestFDR=2.1×10-68)was response to stress (Figure 8). Following this sequentially, were cellularmetabolism (FDR=3.7×10-49), positive regulation of biological processes (FDR =2.4×10-43), cell death (FDR = 2.5×10-33), cellular localization (FDR = 8.4×10-25), andpigmentmetabolism(FDR=2.1×10-21).Amongthe272genesover-expressed intheNC but not in the Om colonies in response to heat stress, 38 biological processeswere enriched: organic acid catabolism (FDR = 1.6×10-22), protein transport (FDR =1.8×10-16),stressresponse(FDR=4.8×10-13),andcellularmetabolism(FDR=3×10-12)werethefourmostsignificantlyenrichedbiologicalprocesses(Figure8).Amongthe2,082genesover-expressedintheOmbutnotintheNCcoloniesinresponsetoheatstress, 160 biological processes were enriched, the most significant being ncRNAmetabolism (FDR= 8.9×10-303), cellularmetabolism (FDR= 4.4×10-70), carbohydratederivative biosynthesis (FDR = 5.9×10-64), and organic substance transport (FDR =2×10-44).
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 22
Figure8.SummaryoftheGOenrichmentanalysisfollowingREVIGOsynthesis.Eachenrichedbiologicalprocess is representedbyabarproportional to thelog10(FDR).Thecolorscorrespondtothethreecategoriesofgenes(common:black; Om-specific: grey; NC-specific: white) that were over-expressed (leftpanel)orunder-expressed(rightpanel).
For the 350 genes that were under-expressed following heat stress
irrespective to the locality of origin (Om or NC), 48 biological processes wereenriched and grouped into five biological processes: nitrogen compound transport(FDR=5.4×10-89),localization(FDR=8.1×10-10),regulationofneurotransmitterlevels(FDR=1.2×10-8),systemdevelopment(FDR=8.8×10-6),andsingleorganismprocess(FDR=4×10-4).Among theunder-expressedgenes in theNCcoloniesonly,a singlebiological process (anatomical structure/morphogenesis)was found to be enriched(FDR=9×10-3).Amongtheunder-expressedgenesintheOmcolonies,139biologicalprocesses were enriched, with the most significant being ion transmembranetransport(FDR=7.6×10-104),singlemulticellularorganismprocess(FDR=7.5×10-53),regulationof biological quality (FDR= 6×10-48), cell–cell signaling (FDR= 1.5×10-23),single organism process (FDR = 1.1×10-18), multicellular organism process (FDR =1.5×10-16),biologicalregulation(FDR=2.3×10-15),responsetoabioticstimulus(FDR=
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 23
6.2×10-13), and localization (FDR = 4.6×10-12). The complete results for all GO termcategoriesincludingmolecularfunctionareavailableinSupplementaryFileS9.
Regarding cellular compartments, the mitochondria was the mostsignificantlyenrichedinthecommonover-expressedgenes(FDR=1.5×10-180),aswellas among genes over-expressed in NC (FDR = 2.5×10-82) while in genes over-expressed inOmcorals the intracellularorganelle lumenwas themost significantlyenriched(FDR=1×10-560).
To investigatewhether thepresumablymore thermotolerant colonies fromOman displayed a frontloading strategy (i.e. a higher expression for some genescompared to the colonies from NC) as previously described in scleractinian corals(Barshisetal.2013),wecompared thegeneexpression levels incontrol conditionsbetween Om and NC colonies for those genes that were over-expressed in NCcolonies (Supplementary File S10). This comparison revealed that the constitutiveexpression levelwas often greater in theOm colonies. Among the 770 genes thatwere over-expressed in NC colonies in response to thermal stress (272 specificallyand498incommonwithOm),484wereconstitutively(i.e.inthecontrolcondition)more expressed inOm. Among these genes, 163were not differentially expressedbetween the control and stress temperatures reflecting true frontloadingbasedonthe definition of Barshis et al. (2013), while 301were over-expressed and only 20wereunder-expressedduringheat stress.These484geneswithhigher constitutiveexpression in Om were submitted to GO term enrichment analysis. No significantresults were found for the under-expressed genes. The frontloaded genes wereenriched in the biological processes cellular respiration (FDR = 4.4×10-23), cellularcomponentorganization (FDR=0.002), homeostaticprocess (FDR=0.005), cellularcomponent organization or biogenesis (FDR = 0.007), cofactor metabolism (FDR =0.009), and stress response (FDR = 0.009), and in themitochondrion for themostsignificant cellular compartment (FDR = 1.6×10-66). Most interestingly, for genesassociatedwithahigherbasalexpressionleveltogetherwithover-expressionintheOm colonies, the most enriched biological processes were stress response (FDR =1.2×10-26), pigment metabolism (FDR = 5.1×10-24), regulation of phosphatemetabolism (FDR = 3.2×10-15), cellular metabolism (FDR = 2.7×10-11), and proteinfolding(FDR=7.3×10-6).
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 24
Discussion
SpecificcontextofadaptationOur aim was to compare the phenotypic plasticity in terms of transcriptomicresponse to heat stress of coral colonies originating from different localitiesdisplayingcontrastedthermalregimes.Asmorphologycanbemisleadingforspeciesidentification inscleractinians,notably inPocilloporagenus(Gélinetal.,2017a),weused a molecular approach to test the species relationships of our samples. Theanalysis ofmitochondrial sequences and clustering analyses indicated that, despitesimilar morphologies, our samples corresponded to different species. This agreeswell with previous works showing the importance of cryptic lineages andmorphologicalplasticity in thePocillopora genus (Gélinetal. 2017aand referencesherein). Oman colonies corresponded to species hypothesis PSH12 of (Gélin et al.2017b), which is restricted to the Northwestern Indian Ocean. Regarding the twospecies hypotheses from NC, SSH05a (P.damicornis type β SSH05a or P.acuta) isfound in the PacificOcean andPSH04 (P.damicornis typeα orP.damicornis sensustricto)isnearlyexclusivelyfoundinthePacificOcean(veryrareintheIndianOcean,andnot foundyet inRedSea) (Gélinetal.2017b). Itwouldbe interesting tostudywhether inside each species hypothesis, different thermotolerance phenotypes arepresent.Conversely, theobservationof a similar response to thermal stress in twodifferentspecies inNC,as revealedbydifferentialgeneexpressionaswellasDAPCanalyses,couldindicateeitheraconservedstrategyoraconvergenceunderthesameecologicalconditions.AnecologicallyrealisticheatstressTheheat stress applied in this studywas ecologically realistic, since the first visualresponse (i.e. polyp closure) was observed for all colonies when the graduallyincreasing experimental temperature reached the upper temperature they aresubjected to in natura (30°C and34°C forNC andOmcorals, respectively). Fromabiological point of view this first result hence clearly supports that these coloniesfrom two localities that are experiencing two different thermal regimes in naturadisplaydifferentialabilitytodealwithheatstress.Moreover,theaccuratecontrolofallotherseawaterparametersallowsustoconsider that theholobiontresponsetothe thermal treatment is specific to heat stress and not to other possibleconfoundingeffects.Last,asweanalyzedthesamplesbeforethefirstvisiblesignsofstress (polyp closure), any change in the holobiont would therefore reflect theresponsetotheheatstressandnothomeostasisbreakdownafterdisruptionofthecoralintegrity.
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 25
Symbioticcommunity:bacterialandSymbionaceaecompositionForthebacterialcommunity,weidentifiedsignificantdifferencesbetweenlocalitiesandcolonies.Themicrobiotacompositionofallsampleswasconsistentwithpreviousstudies, showingahighproportionofGammaproteobacteriaanddominanceof thesymbioticEndozoicomonasgenus(Bourne&Munn2005;Neaveetal.2016a;Peixotoetal. 2017).However,our results clearlydemonstrate thatneithermaintenance inthe experimental structure nor experimental heat stress induced major bacterialcommunity changes in coral colonies irrespective to their locality of origin. For theSymbionaceae community, the ITS2 metabarcoding analysis enabled inter-claderesolution(Quigleyetal.2014).TwodistincttypesofD1aandC1cladesdominated,representingmostofthesequencesintheOmandNCcorals,respectively.NineITStypes (Ato I)havebeen identified intheformergenusSymbiodinium (Baker2003).Some Symbiodiniaceae strains strongly participate to the overall holobiont fitness,with typeD providing tolerance to higher temperatures (Berkelmans& vanOppen2006)andC1enhancingcoralgrowthrates(Littleetal.2004).Interestingly,wefoundthat the type D1a is dominant in the more thermotolerant Om corals, which isconsistent with the results of previous works (Berkelmans & van Oppen 2006),howeverrecentresultsshowsthatsuchanassociationisratherlinkedwithminimaltemperatures than annual amplitudeof temperature changes (Brener-Raffalliet al.2018).
Although the microbial community (both bacterial and Symbiodiniaceae)differed between the NC and Om corals, the composition did not change duringtransition from the field to the artificial seawater conditions, and remained similarduringtheexperimentaltemperatureincrease.Thus,thecoralholobiontassemblageremainedsimilarafter thecourseof theexperiment.Suchstabilityof themicrobialcommunity during experimental heat stress was previously observed in thescleractinianAcroporamillepora(Bellantuonoetal.2012b)andA.tenuis(Littmanetal. 2010). Thus, our study conforms to the idea that microbial communitiesassociated with scleractinian corals remain unchanged when the holobionts areexposedtostressfultemperatures(butsee(Ziegleretal.2017))butfurtheranalysesofgeneexpressionlevelwouldbeneededtoassesstheirfunctionalresponses.RNA-sequencing of eukaryotic poly-adenylated mRNA would allow in principle dualanalysis of Symbiodiniaceae and coral host transcripts (Mayfield et al. 2014), butsince our RNA extraction method resulted in very few algal transcripts, we onlyfocusedonthehosttranscriptomicresponse.
Basedon these results,we investigatedchanges inhostgeneexpressionasthemainmechanismofresponsetoheatstressinourexperimentaldesign.
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 26
HosttranscriptomicresponseGiventheobservedstabilityofthemicrobialsymbioticcommunityduringheatstress,wefocusedmorespecificallyontheresponsesattributabletothecoralhost.Wethuscomparedgeneexpressionpatternsatthequalitativeandquantitative levels inOmand NC colonies in response to heat stress compared to the control condition.Altogether, our results clearly highlight that the Oman colonies exposed to morevariable thermal conditions in natura also display, in response to heat stress, agreater plasticity in gene expression levels than the NC colonies. In particular, thetranscriptomic response of the Oman colonies involved a larger number of geneswith 73% of commonly differentially expressed genes having higher fold changescompared to the NC colonies. These findings are consistent with the theoreticalexpectationsthatamorevariableenvironmentpromotestheevolutionofagreaterplasticity (Lande 2009). Accordingly, a recent transplantation study conducted innaturaalsoidentifiedgreatertranscriptomicplasticityinamorethermotolerant(in-shore)populationcomparedwithan(off-shore)populationinhabitingamorestablethermalhabitatinthemustardhillcoralP.astreoides(Kenkel&Matz2016).
Importantly however,we also identified several geneswhose expression isconstitutivelyhigherintheOmcoloniescomparedtotheNCcoloniesbycomparingthe expression levels in the control condition. This process recently called“frontloading” (Barshis et al. 2013) reflects the preemptive expression of stress-response genes, hence predisposing organisms to better respond to stress. It hasbeen proposed that the occurrence of plasticity vs. frontloading strategies maydepend on the frequency of stresses relative to the typical response time oforganisms, with frequent stresses promoting frontloading strategies whereas lessfrequentperturbationswouldresultinanincreasedplasticity(Kenkel&Matz2016).Other conceptual considerations especially in regards to the predictability ofenvironmental variation through generations should also be taken into account(Danchin 2013; Herman et al. 2014). The frontloading is by definitionmore costlythan plasticity since it transforms a response to the environment in a constitutivefunction. Frontloading is therefore a strategy that would be more efficient whenoffspring’s habitat is highly predictable. On the contrary, an unpredictable or lesspredictableoffspringenvironmentmaypromoteplasticitytoenabletheexplorationofawiderphenotypiclandscapeatalessercost.Plasticityandfrontloadingareoftendiscussedasmutuallyexclusiveresponses(Barshisetal.2013;Kenkel&Matz2016).However,coralsareknowntodisplayahigh levelofvariation intheir reproductionstrategies(broodervs.broadcastspawner)(Whitaker2006;Bairdetal.2009),timing(Fan et al. 2006) and pelagic larval duration (Harrison & Wallace 1990).Environmental predictability in terms of stress frequency and annual temperaturevariation should be therefore limited andwe hypothesized that, rather than being
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 27
exclusive, plasticity and frontloading often co-occur especially in the populationexperiencingextremeenvironments.
Our results clearly support that plasticity and frontloading indeed co-occurspecificallyinthethermotolerantOmcoloniesexperiencingamorevariablethermalenvironmentinnatura.Toteaseapartthebiologicalprocessesthatareregulatedviaplasticity or frontloading in Pocillopora response to heat stress, we conducted anenrichmentanalysis.Keepinginmindthatcongruencybetweengeneexpressionandprotein levels should be cautious (Mayfield et al. 2016), we propose a detaileddiscussion of the response of coral colonies at the molecular level for each mainbiologicalprocessidentified(SupplementaryFileS11).Notably,wefounddifferencesin gene expression levels in response to temperature increase between the twolocalitiesforgenesinvolvedinresponsetoheatstress(suchasHSPs),detoxificationof reactive oxygen species, apoptosis, mitochondria energetic functioning, andsymbiontmaintenancewithhighernumberofdifferentiallyexpressedgenesfortheOmcoralsassociatedtohigherfoldchanges. Incorals,Hsp70whichdisplayedmoreintense overexpression in Om colonies, is one of the most documented proteinchaperonesassociatedtoheatstressresponse(Barshisetal.2013;Haguenaueretal.2013).Severalgenes involved inoxidativestresshadgreaterdifferentialexpressionlevels in Om such as calmodulin, quinone oxidoreductases and thioredoxin(SupplementaryFileS11).Reactiveoxygenspecies(ROS),generatedinconsequencetoheatstress,playacentralroleinhost-symbiontinteractionsultimatelyleadingtobleachingasahostinnateimmuneresponsetoacompromisedsymbiont(see(Weis2008)forareview).Wealso identifiedmanygenes involvedinapoptosis,aprocessthathasbeenrecurrentlyassociatedwithcoralresponsestoheatstress(Ainsworthetal.2011;Barshisetal.2013;Pratlongetal.2015;Maor-Landaw&Levy2016).
Our results also suggest that allocatingenergy inheat stress response is atthe expense of other crucial biological processes such as growth and reproductivefunctionsasalreadyshowninPocillopora(Vidal-Dupioletal.2009;2014),evenifwecouldnottestexperimentallyfitnesseffectoftheexperimentalheatstress.However,themolecularmechanismsunderlying such overall response toheat stress are stillpartly unresolved. Interestingly, we also found specific gene expression patternslinked with epigenetic regulation such as histone modifying enzymes as well asenzymes involved in DNA methylation and dsRNA regulation (Supplementary FileS11). We also identified expression of many retro-transposons in Om. Theseprocesses could be involved in rapid epigenomemodifications and thus fuel rapidadaptiveevolution(Maumusetal.2009;Tordaetal.2017;Jablonka2017).
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 28
Conclusion
Comparisonoftheresponsetoanecologicallyrealisticheatstressofcoralsfromthesamegenusbutpertainingtodifferentspecieshypothesesthrivingintwocontrastingthermal environments sheds light on the molecular basis of thermotolerance.Wefound that after heat exposure, the symbiotic community composition remainedsimilar in colonies fromboth localities, butwe identifiedmajordifferences in generegulationprocessesinthecoral,therebyunderliningtheroleofthecoralhostintheresponse toheat stress.Thecolonies fromthe localitydisplaying themostvariableenvironment displayed (i) a more plastic transcriptome response involving moredifferentially expressed genes and higher fold expression changes; as well as (ii) aconstitutivehigherlevelofexpressionforanothersetofgenes(frontloading).Inthecontext of climate change, which is predicted to cause abnormal and rapidtemperature increase (IPCC 2014), phenotypic plasticity and the capacity for rapidadaptationthroughepigeneticregulationand/orgeneticassimilationwouldincreasetheprobabilityofcoralsurvival.Previousstudieshighlightedtheimportanceofreefmanagementsmeasures(Rogersetal.2015)andassistedevolution(vanOppenetal.2015),butalsounderlinedtheimportanceofpreservingstandinggenetic/epigeneticvariation in wild coral populations (Matz et al. 2017). Although the molecularmechanisms we described are most likely largely shared in this group ofscleractinians, the question remains of the determinism of this thermotolerantphenotypeandoftheheritabilityofthischaracter.Itishoweveressentialtokeepinmind that even themost thermotolerant coralsmay bleach if they are exposed totemperaturesignificantlyhighertotheirownnorm(Hughesetal.2017b;LeNohaïcetal.2017).
Dataaccessibility
ThedatasetsgeneratedandanalyzedduringthecurrentstudyhavebeensubmittedtotheSRArepositoryunderbioprojectnumberPRJNA399069(https://www.ncbi.nlm.nih.gov/bioproject/PRJNA399069).
Supplementarymaterial
RScriptsareavailableonline:https://osf.io/s5y34
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 29
Acknowledgements
WearethankfultoDr.MadjidDelghandifromtheCenterofMarineBiotechnologyatSultanQaboosUniversity, theAl-Hail field stationand thecentral laboratoryof theCollege of Agricultural and Marine Sciences for providing us with necessaryequipmentduringfieldworkinOman.WeacknowledgeDr.GillesLeMoullacandDr.YannickGueguenfromtheCentreIfremerduPacifiqueforprovidingusaquaculturefacilitiesduringsampling.WeacknowledgethePlateformeGentyaneof the InstitutNational de la Recherche Agronomique (INRA, Clermont-Ferrand, France) formicrosatellitegenotyping.WealsothanktheGenotoulbioinformaticsplatform,andthe Toulouse Midi-Pyrenees and Sigenae group for providing help and computingresources(Galaxy instance;http://sigenae-workbench.toulouse.inra.fraswellastheBio-Environnement platform (University of Perpignan) for sequencing andbioinformaticsservice.ThisprojectwasfundedbytheADACNIprogramoftheFrenchnational research agency (ANR) (project no. ANR-12-ADAP-0016;http://adacni.imbe.fr), the Campus France PHC programMaïmonide-Israel and theDHOF program of theUMR5244 IHPE (http://ihpe.univ-perp.fr/en/ihpe-transversal-holobiont/).ThisworkisacontributiontotheLabexOT-Med(n°ANR-11-LABX-0061)funded by the ANR “Investissements d’Avenir” program through the A*MIDEXproject (n° ANR-11-IDEX-0001-02). This study is set within the framework of the“Laboratoired’Excellence(LABEX)”TULIP(ANR-10-LABX-41).WewouldliketothankStaffan Jacob, Xavier Pichon and Mar Sobral for their constructive and helpfulcomments.Version4ofthispreprinthasbeenreviewedandrecommendedbyPeerCommunityInEcology(https://doi.org/10.24072/pci.ecology.100028).
Conflictofinterestdisclosure
The authors of this preprint declare that they have no financial conflict of interestwiththecontentofthisarticle.GMisrecommenderforPCIEcology.
References
AdjeroudM,GuérécheauA,Vidal-DupiolJetal.(2013)Geneticdiversity,clonalityandconnectivityinthescleractiniancoralPocilloporadamicornis:amulti-scaleanalysisinaninsular,fragmentedreefsystem.MarineBiology,161,531–541.
AinsworthTD,WasmundK,UkaniLetal.(2011)Definingthetippingpoint:acomplexcellularlife/deathbalanceincoralsinresponsetostress.ScientificReports,1,160.
AkaikeH(1974)ANewLookattheStatisticalModelIdentification.IEEETransactionsonAutomaticControl,6,716–723.
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 30
AltschulSF,GishW,MillerW,MyersEW,LipmanDJ(1990)Basiclocalalignmentsearchtool.Journalofmolecularbiology,215,403–410.
AndersS,PylPT,HuberW(2015)HTSeq--aPythonframeworktoworkwithhigh-throughputsequencingdata.Bioinformatics,31,166–169.
Arnaud-HaondS,BelkhirK(2006)genclone:acomputerprogramtoanalysegenotypicdata,testforclonalityanddescribespatialclonalorganization.MolecularEcologyNotes,7,15–17.
BairdAH,GuestJR,WillisBL(2009)SystematicandBiogeographicalPatternsintheReproductiveBiologyofScleractinianCorals.AnnualReviewofEcology,Evolution,andSystematics,40,551–571.
BakerAC(2003)FlexibilityandSpecificityinCoral-AlgalSymbiosis:Diversity,Ecology,andBiogeographyofSymbiodinium.AnnualReviewofEcology,Evolution,andSystematics,34,661–689.
BarnoskyAD,MatzkeN,TomiyaSetal.(2011)HastheEarth’ssixthmassextinctionalreadyarrived?Nature,471,51–57.
BarshisDJ,BarshisDJ,LadnerJTetal.(2013)FromtheCover:Genomicbasisforcoralresiliencetoclimatechange.ProceedingsoftheNationalAcademyofSciences,110,1387–1392.
BatesD,MächlerM,BolkerB,WalkerS(2015)FittingLinearMixed-EffectsModelsUsinglme4.JournalofStatisticalSoftware,67,1-48.
BellantuonoAJ,BellantuonoAJ,Granados-CifuentesCetal.(2012a)CoralThermalTolerance:TuningGeneExpressiontoResistThermalStress.PloSone,7,e50685.
BellantuonoAJ,Hoegh-GuldbergO,Rodriguez-LanettyM(2012b)Resistancetothermalstressincoralswithoutchangesinsymbiontcomposition.Proceedings.Biologicalsciences,279,1100–1107.
BellwoodDR,HughesTP,FolkeC,NyströmM(2004)Confrontingthecoralreefcrisis.Nature,429,827–833.
BenjaminiY,HochbergY(1995)Controllingthefalsediscoveryrate:apracticalandpowerfulapproachtomultipletesting.JournaloftheroyalstatisticalsocietySeriesB,57,289–300.
BerkelmansR,vanOppenMJH(2006)Theroleofzooxanthellaeinthethermaltoleranceofcorals:a“nuggetofhope”forcoralreefsinaneraofclimatechange.ProceedingsoftheRoyalSocietyB-BiologicalSciences,273,2305–2312.
BlankenbergD,GordonA,KusterVonGetal.(2010)ManipulationofFASTQdatawithGalaxy.Bioinformatics,26,1783–1785.
BordensteinSR,TheisKR(2015)HostBiologyinLightoftheMicrobiome:TenPrinciplesofHolobiontsandHologenomes(MKWaldor,Ed,).PLoSbiology,13,e1002226.
BourneDG,MunnCB(2005)DiversityofbacteriaassociatedwiththecoralPocilloporadamicornisfromtheGreatBarrierReef.Environmentalmicrobiology,7,1162–1174.
Brener-RaffalliK,ClerissiC,Vidal-DupiolJetal.(2018)Thermalregimeandhostclade,ratherthangeography,driveSymbiodiniumandbacterialassemblagesinthescleractiniancoralPocilloporadamicornissensulato.Microbiome,6,2.
BrownBE,DownsCA,DunneRP,GibbSW(2002)ExploringthebasisofthermotoleranceinthereefcoralGoniastreaaspera.MarineEcologyProgress
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 31
Series,242,119–129.ColesSL,RieglBM(2013)ThermaltolerancesofreefcoralsintheGulf:areviewof
thepotentialforincreasingcoralsurvivalandadaptationtoclimatechangethroughassistedtranslocation.MarinePollutionBulletin,72,323–332.
ConesaA,MadrigalP,TarazonaSetal.(2016)AsurveyofbestpracticesforRNA-seqdataanalysis.Genomebiology,17,1.
CróquerA,BastidasC,ElliottA,SweetM(2013)BacterialassemblagesshiftsfromhealthytoyellowbanddiseasestatesinthedominantreefcoralMontastraeafaveolata.EnvironmentalMicrobiologyReports,5,90–96.
DanchinE(2013)Avatarsofinformation:towardsaninclusiveevolutionarysynthesis.TrendsinEcology&Evolution,28,351–358.
deHoonMJL,ImotoS,NolanJ,MiyanoS(2004)Opensourceclusteringsoftware.Bioinformatics,20,1453–1454.
DixonGB,DaviesSW,AglyamovaGAetal.(2015)CORALREEFS.Genomicdeterminantsofcoralheattoleranceacrosslatitudes.Science,348,1460–1462.
DobinA,DavisCA,SchlesingerFetal.(2013)STAR:ultrafastuniversalRNA-seqaligner.Bioinformatics,29,15–21.
EscudiéF,AuerL,BernardMetal.(2017)FROGS:Find,Rapidly,OTUswithGalaxySolution.Bioinformatics,34,1287-1294.
FanTY,LinKH,KuoFW,SoongK,LiuLL(2006)Dielpatternsoflarvalreleasebyfivebroodingscleractiniancorals.MarineEcologyProgressSeries,321,133–142.
GélinP,FauvelotC,MehnVetal.(2017a)SupercloneExpansion,Long-DistanceClonalDispersalandLocalGeneticStructuringintheCoralPocilloporadamicornisTypeβinReunionIsland,SouthWesternIndianOcean.PloSone,12,e0169692.
GélinP,PirogA,FauvelotC,MagalonH(2018)HighgeneticdifferentiationandlowconnectivityinthecoralPocilloporadamicornistypeβatdifferentspatialscalesintheSouthwesternIndianOceanandtheTropicalSouthwesternPacific.MarineBiology,165,531.
GélinP,PostaireB,FauvelotC,MagalonH(2017b)Reevaluatingspeciesnumber,distributionandendemismofthecoralgenusPocilloporaLamarck,1816usingspeciesdelimitationmethodsandmicrosatellites.MolecularPhylogeneticsandEvolution,109,430–446.
GiardineB,GiardineB,RiemerCetal.(2005)Galaxy:Aplatformforinteractivelarge-scalegenomeanalysis.GenomeResearch,15,1451–1455.
GuerreroR,MargulisL,BerlangaM(2013)Symbiogenesis:theholobiontasaunitofevolution.InternationalMicrobiology,16,133–143.
HaguenauerA,ZubererF,LedouxJ-B,AurelleD(2013)AdaptiveabilitiesoftheMediterraneanredcoralCoralliumrubruminaheterogeneousandchangingenvironment:frompopulationtofunctionalgenetics.JournalofExperimentalMarineBiologyandEcology,449,349–357.
HarrisonPL,WallaceCC(1990)Reproduction,dispersalandrecruitmentofscleractiniancorals.Ecosystemsoftheworld.
HermanJJ,SpencerHG,DonohueK,SultanSE(2014)Howstable“should”epigeneticmodificationsbe?Insightsfromadaptiveplasticityandbethedging.Evolution,68,632–643.
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 32
Hernandez-AgredaA,GatesRD,AinsworthTD(2016)25.TrendsinMicrobiology,25,125–140.
Hoegh-GuldbergO,MumbyPJ,HootenAJetal.(2007)Coralreefsunderrapidclimatechangeandoceanacidification.Science,318,1737–1742.
HughesTP,BairdAH,BellwoodDRetal.(2003)Climatechange,humanimpacts,andtheresilienceofcoralreefs.Science,301,929–933.
HughesTP,BarnesML,BellwoodDRetal.(2017a)CoralreefsintheAnthropocene.Nature,546,82–90.
HughesTP,KerryJT,Álvarez-NoriegaMetal.(2017b)Globalwarmingandrecurrentmassbleachingofcorals.Nature,543,373–377.
HumeB,D'AngeloC,BurtJetal.(2013)CoralsfromthePersian/ArabianGulfasmodelsforthermotolerantreef-builders:PrevalenceofcladeC3Symbiodinium,hostfluorescenceandexsitutemperaturetolerance.MarinePollutionBulletin,72,313–322.
JablonkaE(2017)Theevolutionaryimplicationsofepigeneticinheritance.Interfacefocus,7,20160135.
JombartT,DevillardS,BallouxF(2010)Discriminantanalysisofprincipalcomponents:anewmethodfortheanalysisofgeneticallystructuredpopulations.BMCgenetics,11,94.
KenkelCD,MatzMV(2016)Geneexpressionplasticityasamechanismofcoraladaptationtoavariableenvironment.NatureEcology&Evolution,1,0014.
KenkelCD,MeyerE,MatzMV(2013)Geneexpressionunderchronicheatstressinpopulationsofthemustardhillcoral(Poritesastreoides)fromdifferentthermalenvironments.MolecularEcology,22,4322–4334.
KlindworthA,PruesseE,SchweerTetal.(2012)Evaluationofgeneral16SribosomalRNAgenePCRprimersforclassicalandnext-generationsequencing-baseddiversitystudies.NucleicAcidsResearch,41,e1–e1.
LajeunesseTC,TrenchRK(2000)BiogeographyoftwospeciesofSymbiodinium(Freudenthal)inhabitingtheintertidalseaanemoneAnthopleuraelegantissima(Brandt).TheBiologicalbulletin,199,126–134.
LaJeunesseTC,ParkinsonJE,GabrielsonPWetal.(2018)SystematicRevisionofSymbiodiniaceaeHighlightstheAntiquityandDiversityofCoralEndosymbionts.CurrentBiology,28,2570–2580.e6.
LandeR(2009)Adaptationtoanextraordinaryenvironmentbyevolutionofphenotypicplasticityandgeneticassimilation.JournalofEvolutionaryBiology,22,1435–1446.
LeNohaïcM,RossCL,CornwallCEetal.(2017)MarineheatwavecausesunprecedentedregionalmassbleachingofthermallyresistantcoralsinnorthwesternAustralia.ScientificReports,7,14999.
LiJ,ChenQ,LongL-Jetal.(2014)Bacterialdynamicswithinthemucus,tissueandskeletonofthecoralPoritesluteaduringdifferentseasons.ScientificReports,4,7320.
LittleAF,vanOppenMJH,WillisBL(2004)Flexibilityinalgalendosymbiosesshapesgrowthinreefcorals.Science,304,1492–1494.
LittmanR,BourneDG,WillisBL(2010)Responsesofcoral-associatedbacterialcommunitiestoheatstressdifferwithSymbiodiniumtypeonthesamecoral
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 33
host.MolecularEcology,19,1978–1990.LoveMI,HuberW,AndersS(2014)Moderatedestimationoffoldchangeand
dispersionforRNA-seqdatawithDESeq2.Genomebiology,15,31.MagočT,SalzbergSL(2011)FLASH:fastlengthadjustmentofshortreadstoimprove
genomeassemblies.Bioinformatics,27,2957-63.MahéF,RognesT,QuinceC,DeVargasC,DunthornM(2014)Swarm:robustandfast
clusteringmethodforamplicon-basedstudies.PeerJ,2,e593.Maor-LandawK,LevyO(2016)Geneexpressionprofilesduringshort-termheat
stress;branchingvs.massiveScleractiniancoralsoftheRedSea.PeerJ,4,e1814.MargulisL,FesterR(1991)SymbiosisasaSourceofEvolutionaryInnovation.MIT
Press.MartinM(2011)Cutadaptremovesadaptersequencesfromhigh-throughput
sequencingreads.EMBnetjournal,17,10.MatzMV,TremlEA,AglyamovaGA,vanOppenMJH,BayLK(2017)Potentialforrapid
geneticadaptationtowarminginaGreatBarrierReefcoral.PLoSGenet.,19,e1007220.
MaumusF,AllenAE,MhiriCetal.(2009)Potentialimpactofstressactivatedretrotransposonsongenomeevolutioninamarinediatom.BMCGenomics,10,624.
MayfieldAB,WangL-H,TangP-Cetal.(2011)Assessingtheimpactsofexperimentallyelevatedtemperatureonthebiologicalcompositionandmolecularchaperonegeneexpressionofareefcoral.PloSone,6,e26529.
MayfieldAB,WangY-B,ChenC-S,ChenS-H,LinC-Y(2016)Dual-compartmentaltranscriptomic+proteomicanalysisofamarineendosymbiosisexposedtoenvironmentalchange.MolecularEcology,25,5944–5958.
MayfieldAB,WangY-B,ChenC-S,LinC-Y,ChenS-H(2014)Compartment-specifictranscriptomicsinareef-buildingcoralexposedtoelevatedtemperatures.MolecularEcology,23,5816–5830.
McFall-NgaiM,HadfieldMG,BoschTCetal.(2013)Animalsinabacterialworld,anewimperativeforthelifesciences.ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,110,3229–3236.
McMurdiePJ,HolmesS(2013)phyloseq:AnRPackageforReproducibleInteractiveAnalysisandGraphicsofMicrobiomeCensusData(MWatson,Ed,).PloSone,8,e61217.
MeyerJL,RodgersJM,DillardBA,PaulVJ,TeplitskiM(2016)EpimicrobiotaAssociatedwiththeDecayandRecoveryofOrbicellaCoralsExhibitingDarkSpotSyndrome.FrontiersinMicrobiology,7,893.
NeaveMJ,ApprillA,Ferrier-PagesC,VoolstraCR(2016a)DiversityandfunctionofprevalentsymbioticmarinebacteriainthegenusEndozoicomonas.Appliedmicrobiologyandbiotechnology,100,8315–8324.
NeaveMJ,RachmawatiR,XunLetal.(2016b)DifferentialspecificitybetweencloselyrelatedcoralsandabundantEndozoicomonasendosymbiontsacrossglobalscales.TheISMEJournal,11,186–200.
OliverTA,PalumbiSR(2010)Manycoralshostthermallyresistantsymbiontsinhigh-temperaturehabitat.CoralReefs,30,241–250.
PalumbiSR,BarshisDJ,Traylor-KnowlesN,BayRA(2014)Mechanismsofreefcoral
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 34
resistancetofutureclimatechange.Science,344,895–898.PanteE,PuillandreN,ViricelAetal.(2015)Speciesarehypotheses:avoid
connectivityassessmentsbasedonpillarsofsand.MolecularEcology,24,525–544.
PantosO,BongaertsP,DennisPG,TysonGW,Hoegh-GuldbergO(2015)Habitat-specificenvironmentalconditionsprimarilycontrolthemicrobiomesofthecoralSeriatoporahystrix.TheISMEJournal,9,1916–1927.
PeixotoRS,RosadoPM,LeiteDCdeA,RosadoAS,BourneDG(2017)BeneficialMicroorganismsforCorals(BMC):ProposedMechanismsforCoralHealthandResilience.FrontiersinMicrobiology,8,100.
PinzónJH,SampayoE,CoxEetal.(2013)Blindtomorphology:geneticsidentifiesseveralwidespreadecologicallycommonspeciesandfewendemicsamongIndo-Pacificcauliflowercorals(Pocillopora,Scleractinia)(CMaggs,Ed,).JournalofBiogeography,40,1595–1608.
PolatoNR,VoolstraCR,SchnetzerJetal.(2010)Location-specificresponsestothermalstressinlarvaeofthereef-buildingcoralMontastraeafaveolata.PloSone,5,e11221.
PratlongM,HaguenauerA,ChabrolOetal.(2015)Theredcoral(Coralliumrubrum)transcriptome:anewresourceforpopulationgeneticsandlocaladaptationstudies.Molecularecologyresources,15,1205–1215.
PritchardJK,StephensM,DonnellyP(2000)Inferenceofpopulationstructureusingmultilocusgenotypedata.Genetics,155,945–959.
QuigleyKM,QuigleyKM,DaviesSWetal.(2014)Deep-SequencingMethodforQuantifyingBackgroundAbundancesofSymbiodiniumTypes:ExploringtheRareSymbiodiniumBiosphereinReef-BuildingCorals.PloSone,9,e94297.
Reaka-KudlaML(2005)Theglobalbiodiversityofcoralreefs:acomparisonwithrainforests.
ReuschTBH(2013)Climatechangeintheoceans:evolutionaryversusphenotypicallyplasticresponsesofmarineanimalsandplants.EvolutionaryApplications,7,104–122.
RieglBM,PurkisSJ,Al-CibahyAS,Abdel-MoatiMA,Hoegh-GuldbergO(2011)PresentLimitstoHeat-AdaptabilityinCoralsandPopulation-LevelResponsestoClimateExtremes.PloSone,6,e24802.
RogersA,HarborneAR,BrownCJetal.(2015)Anticipativemanagementforcoralreefecosystemservicesinthe21stcentury.GlobalChangeBiology,21,504–514.
RognesT,FlouriT,NicholsB,QuinceC,MahéF(2016)VSEARCH:aversatileopensourcetoolformetagenomics.PeerJ,18,e2584.
RohwerF,SeguritanV,AzamF(2002)Diversityanddistributionofcoral-associatedbacteria.MarineEcologyProgressSeries,243,1-10.
RosenbergE,KorenO,ReshefL,EfronyR,Zilber-RosenbergI(2007)Theroleofmicroorganismsincoralhealth,diseaseandevolution.NatureReviewsMicrobiology,5,355–362.
SaldanhaAJ(2004)JavaTreeview--extensiblevisualizationofmicroarraydata.Bioinformatics,20,3246–3248.
SatoY,WillisBL,BourneDG(2009)Successionalchangesinbacterialcommunitiesduringthedevelopmentofblackbanddiseaseonthereefcoral,Montipora
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 35
hispida.TheISMEJournal,4,203–214.Schmidt-RoachS,MillerKJ,LundgrenP,AndreakisN(2014)Witheyeswideopen:a
revisionofspecieswithinandcloselyrelatedtothePocilloporadamicornisspeciescomplex(Scleractinia;Pocilloporidae)usingmorphologyandgenetics.ZoologicalJournaloftheLinneanSociety,170,1–33.
SuggettDJ,WarnerME,LeggatW(2017)SymbioticDinoflagellateFunctionalDiversityMediatesCoralSurvivalunderEcologicalCrisis.TrendsinEcology&Evolution,32,735–745.
SupekF,BošnjakM,ŠkuncaN,ŠmucT(2011)REVIGOsummarizesandvisualizeslonglistsofgeneontologyterms.PloSone,6,e21800.
TheisKR,DheillyNM,KlassenJLetal.(2016)GettingtheHologenomeConceptRight:anEco-EvolutionaryFrameworkforHostsandTheirMicrobiomes(JAGilbert,Ed,).mSystems,1,e00028–16.
TordaG,DonelsonJM,ArandaMetal.(2017)Rapidadaptiveresponsestoclimatechangeincorals.NatureClimateChange,7,627–636.
TrapnellC,WilliamsBA,PerteaGetal.(2010)TranscriptassemblyandquantificationbyRNA-Seqrevealsunannotatedtranscriptsandisoformswitchingduringcelldifferentiation.NatureBiotechnology,28,511–515.
vanOppenMJH,OliverJK,PutnamHM,GatesRD(2015)Buildingcoralreefresiliencethroughassistedevolution.ProceedingsoftheNationalAcademyofSciences,112,2307–2313.
vanWoesikR,SakaiK,GanaseA,LoyaY(2011)Revisitingthewinnersandthelosersadecadeaftercoralbleaching.MarineEcologyProgressSeries,434,67–76.
VeronJEN,Stafford-SmithM(2000)Coralsoftheworld.SeaChallengers.Vidal-Dupiol,J.,Chaparro,C.,Pratlong,M.,Pontaroti,P.,Grunau,C.,&Mitta,G.
(2019).Sequencing,denovoassemblyandannotationofthegenomeofthescleractiniancoral,Pocilloporaacuta.bioRxiv,698688.
Vidal-DupiolJ,DheillyNM,RondonRetal.(2014)ThermalstresstriggersbroadPocilloporadamicornistranscriptomicremodeling,whileVibriocoralliilyticusinfectioninducesamoretargetedimmuno-suppressionresponse.PloSone,9,e107672.
Vidal-DupiolJ,AdjeroudMetal.(2009)Coralbleachingunderthermalstress:putativeinvolvementofhost/symbiontrecognitionmechanisms.BMCphysiology,9,14.
WeisVM(2008)CellularmechanismsofCnidarianbleaching:stresscausesthecollapseofsymbiosis.TheJournalofexperimentalbiology,211,3059–3066.
WhitakerK(2006)GeneticevidenceformixedmodesofreproductioninthecoralPocilloporadamicornisanditseffectonpopulationstructure.MarineEcologyProgressSeries,306,115–124.
ZieglerM,SenecaFO,YumLK,PalumbiSR,VoolstraCR(2017)Bacterialcommunitydynamicsarelinkedtopatternsofcoralheattolerance.NatureCommunications,8,14213.
Zilber-RosenbergI,RosenbergE(2008)Roleofmicroorganismsintheevolutionofanimalsandplants:thehologenometheoryofevolution.FEMSMicrobiologyReviews,32,723–735.
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;

PEERCOMMUNITYINECOLOGY 36
Appendix
Supplementary Figure S1. Experimental setup. Four tanks were used for eachlocality,3tankscontainingthesampledcolonies(onereplicatepertimepointandpertank)andoneadditionaltankasacontrolofcoralhealthatthecontroltemperatureduringtheexperiment.Supplementary Table S2. Haplotype analysis of the six sampled colonies withmicrosatellitegenotypingforthecoloniesfromNewCaledonia.Supplementary Figure S3. Bacterial class composition (for the 24 most abundant)withineachreplicatefortheOmandNCcolonies,thethreecoloniesofeachlocality,and three experimental conditions per colony. In situ (dark arrows); controltemperature(greenarrows);stresstemperature(redarrows).Supplementary Table S4.MANOVA results for beta diversity (Bray-Curtis distance)betweenlocalities,colonies,orexperimentalconditions.Supplementary File S5. List and sequences of the 26,600 genes (XLOC) generatedduringRNAseqalignment.Supplementary Figure S6. Heatmap and clustering of significantly differentiallyexpressedgenesbetweenthecontrolandtheheatstressconditionforeachcolonyfromthetwolocalities.Eachgeneisrepresentedbyaline.Supplementary File S7. DESeq2 results for the log2-fold changes, and adjusted pvaluesbetweenstressandcontrolconditionsforeachlocality(sheet1)andforeachcolony(sheet2).Supplementary Table S8. Comparison between the log2-foldchange inOm andNCcoloniesforgenesdifferentiallyunder-expressedorover-expressedinthesamewayincoloniesfrombothlocalities.Supplementary File S9. GO enrichment results for biological processes, molecularfunctions,andcellularcompartmentsforcommon,NewCaledonia-specific,orOman-specificover-expressedandunder-expressedgenes.Supplementary File S10. Frontloaded genes in Oman corals among genes over-expressedinNewCaledoniacorals.Supplementary File S11. Description of the functional analysis of genes, biologicalfunctionsandcellcompartmentinvolvedintheresponsetostress.
.CC-BY-NC-ND 4.0 International licenseIt is made available under a (which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
The copyright holder for this preprint. http://dx.doi.org/10.1101/398602doi: bioRxiv preprint first posted online Aug. 23, 2018;
Copyright © 2022 FDOKUMEN




















