
Ontology - Based Context - Dependent Personalization Technology

Ca2+/calmodulin-dependent kinase II signalling cascademediates P2X7 receptor-dependent inhibition ofneuritogenesis in neuroblastoma cellsRosa Gomez-Villafuertes1, Ana del Puerto2,3,4, Miguel Dıaz-Hernandez1,4, Diego Bustillo5,Juan I. Dıaz-Hernandez1, Paula G. Huerta1, Antonio R. Artalejo5, Juan J. Garrido2,3,4
and Ma Teresa Miras-Portugal1
1 Departamento de Bioquımica y Biologıa Molecular, Facultad de Veterinaria, Universidad Complutense de Madrid, Spain
2 Centro de Biologıa Molecular ‘Severo Ochoa’, CSIC-UAM, Madrid, Spain
3 Departamento de Neurobiologıa Celular Molecular y del Desarrollo, Instituto Cajal, CSIC, Madrid, Spain
4 Centro de Investigacion Biomedica en Red sobre Enfermedades Neurodegenerativas (CIBERNED), Spain
5 Departamento de Toxicologıa y Farmacologıa, Facultad de Veterinaria, Universidad Complutense de Madrid, Spain
Keywords
brilliant blue G; Ca2+ ⁄ calmodulin-dependent
kinase II; neuritogenesis; neuroblastoma;
P2X7 receptor
Correspondence
M. T. Miras-Portugal, Departamento de
Bioquımica y Biologıa Molecular IV, Facultad
de Veterinaria, Universidad Complutense de
Madrid, Av. Puerta de Hierro s ⁄ n, 28040
Madrid, Spain
Fax: +34 91 3943909
Tel: +34 91 3943894
E-mail: [email protected]
(Received 13 May 2009, revised 25 June
2009, accepted 22 July 2009)
doi:10.1111/j.1742-4658.2009.07228.x
ATP, via purinergic P2X receptors, acts as a neurotransmitter and modula-
tor in both the central and peripheral nervous systems, and is also involved
in many biological processes, including cell proliferation, differentiation
and apoptosis. Previously, we have reported that P2X7 receptor inhibition
promotes axonal growth and branching in cultured hippocampal neurons.
In this article, we demonstrate that the P2X7 receptor negatively regulates
neurite formation in mouse Neuro-2a neuroblastoma cells through a
Ca2+ ⁄ calmodulin-dependent kinase II-related mechanism. Using both
molecular and immunocytochemical techniques, we characterized the pres-
ence of endogenous P2X1, P2X3, P2X4 and P2X7 subunits in these cells.
Of these, the P2X7 receptor was the only functional receptor, as its activa-
tion induced intracellular calcium increments similar to those observed in
primary neuronal cultures, exhibiting pharmacological properties character-
istic of homomeric P2X7 receptors. Patch-clamp experiments were also
conducted to fully demonstrate that ionotropic P2X7 receptors mediate
nonselective cation currents in this cell line. Pharmacological inhibition of
the P2X7 receptor and its knockdown by small hairpin RNA interference
resulted in increased neuritogenesis in cells cultured in low serum-contain-
ing medium, whereas P2X7 overexpression significantly reduced the forma-
tion of neurites. Interestingly, P2X7 receptor inhibition also modified the
phosphorylation state of focal adhesion kinase, Akt and glycogen synthase
kinase 3, protein kinases that participate in the Ca2+ ⁄ calmodulin-depen-
dent kinase II signalling cascade and that have been related to neuronal
differentiation and axonal growth. Taken together, our results provide the
first mechanistic insight into P2X7 receptor-triggered signalling pathways
that regulate neurite formation in neuroblastoma cells.
Abbreviations
[Ca2+]i, intracellular free calcium concentration; BBG, brilliant blue G; BzATP, 2¢,3¢-O-(4-benzoyl)-benzoyl ATP; CaMKII, Ca2+ ⁄ calmodulin-
dependent kinase II; CNS, central nervous system; DiBucAMP, dibutyryl-cAMP; FAK, focal adhesion kinase; Fura-2 AM, fura-2
acetoxymethyl ester; GFP, green fluorescent protein; GSK3, glycogen synthase kinase 3; N2a, Neuro-2a; PFA, paraformaldehyde, PI3K,
phosphatidylinositol 3-kinase; shRNA, small hairpin RNA; a,b-meATP, a,b-methylene-ATP.
FEBS Journal 276 (2009) 5307–5325 ª 2009 The Authors Journal compilation ª 2009 FEBS 5307

Introduction
Purinergic signal transduction mechanisms constitute a
complex intra- and intercellular signalling network that
plays an important regulatory role in both the central
(CNS) and peripheral nervous systems. Nucleotides
exert their extracellular effects by acting on specific P2
receptors, which are subdivided into ionotropic P2X
and metabotropic P2Y subtypes [1]. To date, seven
mammalian P2X receptor subtypes (P2X1–7) and eight
mammalian P2Y receptor subtypes (P2Y1, 2, 4, 6, 11–
14) have been cloned and functionally characterized
[2–5].
P2X receptors are nonselective cation channels
formed by the association of three P2X subunits [6].
Each P2X subunit consists of two transmembrane-
spanning segments separated by an N-glycosylated
extracellular loop containing 10 conserved cysteine res-
idues [7,8]. The N- and C-termini are intracellular. The
distribution and relative abundance of P2X receptors
have been studied in different brain areas and cell
types [9]. P2X receptor activation results in Na+ and
Ca2+influx across the cell membrane, which leads to
depolarization of the plasma membrane that can, in
turn, activate voltage-gated channels. Using phar-
macological tools and gene knockout ⁄knockdownapproaches, it has become clear that P2X receptors
are involved in a wide and growing range of physio-
logical processes [10]. In the CNS, ATP directly medi-
ates fast excitatory synaptic transmission, acting via
P2X receptors in mammalian neurons [11–13]. Fur-
thermore, presynaptic P2X receptors can modulate
neurotransmitter release: ATP enhances glutamate
release in the hippocampus [14,15], spinal cord [16],
and midbrain [17]; 4-aminobutyrate release is facili-
tated by P2X activation in cultured spinal cord dorsal
horn neurons [18], midbrain synaptosomes [19] and
lateral hypothalamic neurons [20]; and glycine release
is enhanced by ATP from interneurons in the dor-
sal horn region [21], trigeminal nucleus pars caudalis
neurons [22] and spinal cord substantia gelatinosa
neurons [23].
Interestingly, purinergic signalling is already present
at the early stages of embryogenesis, being involved in
cell proliferation, migration and the differentiation of
a wide variety of structures [24–26]. Moreover, ATP
promotes cell proliferation, acting through P2X3 and
P2X4 receptors in murine embryonic stem cells [27],
and participates in the neuronal maturation of P19
mouse embryonic carcinoma cells via P2X2 and P2X6
receptors, suggesting the presence of purinergic signal-
ling in the initiation and direction of cell differentia-
tion [28]. A trophic effect of purines in regeneration
and neurite outgrowth has also been reported in vari-
ous cell systems, such as PC12 phaeochromocytoma
cells [29,30], retinal ganglion cells [31] and striatal neu-
rons [32]. P2X receptors also participate in the forma-
tion of neuronal networks during hippocampal
development [33,34]. Recently, our group has reported
that the inhibition of P2X7 receptors promotes axonal
growth and branching in cultured hippocampal neu-
rons [35]. Furthermore, ATP, acting via P2X7 recep-
tors, sustains the growth of human neuroblastoma
cells through a substance P-dependent mechanism [36],
whereas a functional decrease in P2X7 receptors seems
to be associated with retinoic acid-induced differentia-
tion of neuroblastoma cells [37]. However, little is
known about the signalling pathways that lead to the
differentiation of neuroblastoma cells following P2X7
receptor inhibition.
In this study, the neurotrophic effect of P2X7 recep-
tors was further examined in mouse Neuro-2a (N2a)
neuroblastoma cells. Pharmacological inhibition and
interference with P2X7 receptor expression were asso-
ciated with neuritogenesis in N2a cells cultured in low
serum-containing medium, whereas P2X7 overexpres-
sion significantly reduced neurite formation. Moreover,
we linked the activation of the P2X7 receptor with the
regulation of Ca2+ ⁄ calmodulin-dependent kinase II
(CaMKII) activity and some of its downstream effec-
tors, including focal adhesion kinase (FAK), phospha-
tidylinositol 3-kinase (PI3K), Akt and glycogen
synthase kinase 3 (GSK3). Together, our results
support a role of P2X7 receptors in the regulation of
neuritogenesis and in the fine control of the balance
between proliferation ⁄differentiation of neuroblastoma
cells.
Results
Molecular characterization of purinergic P2X
receptors in N2a cells
In order to investigate the presence of native puriner-
gic receptors in N2a cells, the expression of mouse
P2X subunits was analysed at both the transcriptional
and protein levels. RT-PCR experiments demonstrated
that most P2X subunits are expressed in N2a cells
(Fig. 1A). To characterize the relative amount of each
P2X transcript, quantitative real-time PCR assays
were performed. Four main P2X mRNAs coding for
P2X1, P2X3, P2X4 and P2X7 subunits were found in
this cell line, with P2X1 and P2X4 transcripts being
the most abundant. The expression of P2X2, P2X5
P2X7 negatively controls neuritogenesis via CaMKII R. Gomez-Villafuertes et al.
5308 FEBS Journal 276 (2009) 5307–5325 ª 2009 The Authors Journal compilation ª 2009 FEBS

and P2X6 was practically undetectable (Fig. 1B). Wes-
tern blot analyses were performed on total protein
extracts obtained from N2a cells using commercial
subunit-specific antibodies. Specific neutralizing pep-
tides were employed to guarantee the specificity of
these bands (data not shown). As expected, the expres-
sion of P2X proteins correlated well with the results
obtained by quantitative PCR (Fig. 1C). Thus, bands
corresponding to monomeric P2X1, P2X3, P2X4,
P2X5 and P2X7 proteins were immunodetected, the
strongest corresponding to P2X3 and P2X7 subunits.
A high level of heterogeneity in the size of P2X su-
bunits has been reported in the literature, probably
caused by differences in glycosylation and ⁄or oligo-
merization. The predicted sizes for unglycosylated
P2X subunits have been reported to range between 43
and 68 kDa, whereas the molecular masses of glycosy-
lated forms are between 50 and 79 kDa (reviewed in
[3]). In this study, the bands obtained for P2X1
(59 kDa), P2X3 (64 kDa), P2X5 (63 kDa) and P2X7
(78 kDa) subunits are consistent with the molecular
masses reported for the glycosylated monomers [6,38–
40]. The low-molecular-mass bands observed for P2X1
(48 kDa) and P2X4 (42 kDa) subunits probably repre-
sent unglycosylated proteins [6,41]. A band detected at
45 kDa with the P2X7 antibody may correspond to a
truncated splice variant of the P2X7 receptor, as
described in humans [42]. High-molecular-mass oligo-
meric forms (dimers or trimers) of P2X subunits were
not observed.
Pharmacological characterization of P2X7
receptors in N2a cells
The pharmacological characterization of P2X receptors
was carried out by analysing the change in intracellular
free calcium concentration ([Ca2+]i) elicited by a vari-
ety of purinergic compounds, in combination with spe-
cific P2X7 antagonists, in N2a cells loaded with the
calcium indicator fura-2 acetoxymethyl ester (Fura-2
AM). First, we tested the ability of several purinergic
agonists, including a,b-methylene-ATP (a,b-meATP),
CTP, 2¢,3¢-O-(4-benzoyl)-benzoyl ATP (BzATP) and
ATP, to increase [Ca2+]i in N2a cells (Fig. 2A). Nei-
ther a,b-meATP (100 lm) nor CTP (300 lm) modified
[Ca2+]i, whereas BzATP (100 lm), which is known to
induce Ca2+influx in cells expressing P2X7 receptors,
elicited a discrete [Ca2+]i increase in the presence of
Mg2+ ions in the superfusion medium. Last, ATP
(100 lm) triggered a substantial calcium response in
N2a cells, even in the absence of Ca2+ in the extracel-
lular medium (data not shown), indicating the partici-
pation of functional metabotropic P2Y receptors in
this cell line, as reported previously [43].
Decreases in extracellular Ca2+ and Mg2+ concen-
tration are known to modulate P2X7 receptor-medi-
ated responses [44–46]. Based on these findings, N2a
cells were challenged with BzATP in Locke’s solution
with or without 1.2 mm Mg2+. As shown in Fig. 2B,
the potency of BzATP increased by two-fold when an
Mg2+-free medium was employed. Moreover, BzATP
P2X1
P2X2
P2X3
P2X4
P2X5
P2X6
P2X7
0
20
40
60
80
No
rmal
ized
to
β-a
ctin
(R
.U.)
M H2O H2O H2O H2ON B
P2X1 P2X2 P2X5P2X3 P2X4 P2X7 P2X6 β-Actin
N B N B N B M H2O H2O H2O H2ON B N B N B N B
75 kDa
50 kDa
35 kDa
P2X
1P
2X2
P2X
3P
2X4
P2X
5P
2X6
P2X
7
α-tubulin
A
B C
Fig. 1. Several P2X receptors are simultaneously expressed in nondifferentiated murine N2a neuroblastoma cells. (A) RT-PCR expression of
P2X receptors. The bands for P2X1–7 receptors were around 100 bp and were amplified from both N2a cells (N) and adult whole mouse
brain mRNA extracts (B). No amplification products were observed in parallel assays carried out without template (H2O). M, DirectLoad�Wide Range DNA marker. (B) Quantitative real-time PCR showing the expression levels of P2X subunits in N2a cells. Values were normal-
ized by the content of b-actin transcript. (C) Total protein isolated from N2a cells was subjected to SDS-PAGE, transferred onto poly(vinyli-
dene difluoride) membrane and incubated with specific antibodies against P2X1–7 subunits. Immunodetection of a-tubulin was used as a
loading control.
R. Gomez-Villafuertes et al. P2X7 negatively controls neuritogenesis via CaMKII
FEBS Journal 276 (2009) 5307–5325 ª 2009 The Authors Journal compilation ª 2009 FEBS 5309
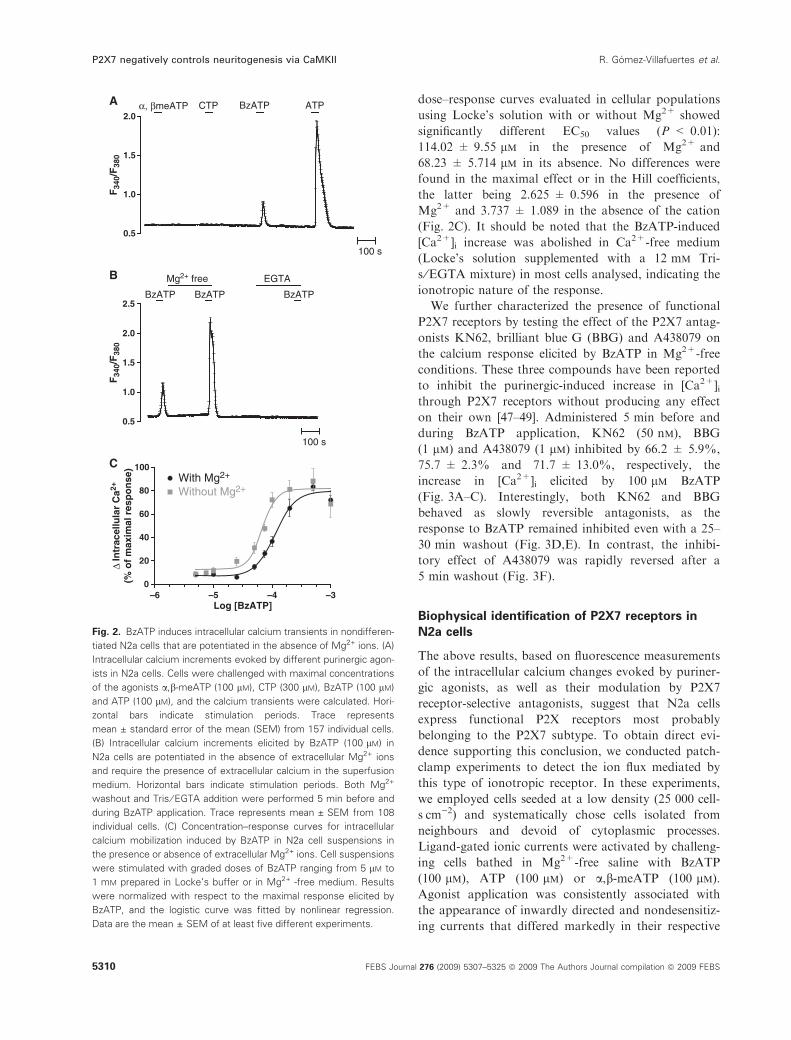
dose–response curves evaluated in cellular populations
using Locke’s solution with or without Mg2+ showed
significantly different EC50 values (P < 0.01):
114.02 ± 9.55 lm in the presence of Mg2+ and
68.23 ± 5.714 lm in its absence. No differences were
found in the maximal effect or in the Hill coefficients,
the latter being 2.625 ± 0.596 in the presence of
Mg2+ and 3.737 ± 1.089 in the absence of the cation
(Fig. 2C). It should be noted that the BzATP-induced
[Ca2+]i increase was abolished in Ca2+-free medium
(Locke’s solution supplemented with a 12 mm Tri-
s ⁄EGTA mixture) in most cells analysed, indicating the
ionotropic nature of the response.
We further characterized the presence of functional
P2X7 receptors by testing the effect of the P2X7 antag-
onists KN62, brilliant blue G (BBG) and A438079 on
the calcium response elicited by BzATP in Mg2+-free
conditions. These three compounds have been reported
to inhibit the purinergic-induced increase in [Ca2+]ithrough P2X7 receptors without producing any effect
on their own [47–49]. Administered 5 min before and
during BzATP application, KN62 (50 nm), BBG
(1 lm) and A438079 (1 lm) inhibited by 66.2 ± 5.9%,
75.7 ± 2.3% and 71.7 ± 13.0%, respectively, the
increase in [Ca2+]i elicited by 100 lm BzATP
(Fig. 3A–C). Interestingly, both KN62 and BBG
behaved as slowly reversible antagonists, as the
response to BzATP remained inhibited even with a 25–
30 min washout (Fig. 3D,E). In contrast, the inhibi-
tory effect of A438079 was rapidly reversed after a
5 min washout (Fig. 3F).
Biophysical identification of P2X7 receptors in
N2a cells
The above results, based on fluorescence measurements
of the intracellular calcium changes evoked by puriner-
gic agonists, as well as their modulation by P2X7
receptor-selective antagonists, suggest that N2a cells
express functional P2X receptors most probably
belonging to the P2X7 subtype. To obtain direct evi-
dence supporting this conclusion, we conducted patch-
clamp experiments to detect the ion flux mediated by
this type of ionotropic receptor. In these experiments,
we employed cells seeded at a low density (25 000 cell-
sÆcm)2) and systematically chose cells isolated from
neighbours and devoid of cytoplasmic processes.
Ligand-gated ionic currents were activated by challeng-
ing cells bathed in Mg2+-free saline with BzATP
(100 lm), ATP (100 lm) or a,b-meATP (100 lm).
Agonist application was consistently associated with
the appearance of inwardly directed and nondesensitiz-
ing currents that differed markedly in their respective
0.5
1.0
1.5
2.0
100 s
α, βmeATP CTPF
340/
F38
0F
340/
F38
0BzATP ATP
100 s
0.5
1.0
1.5
2.0
2.5BzATP BzATP BzATP
Mg2+ free EGTA
–6 –5 –4 –30
20
40
60
80
100With Mg2+
Without Mg2+
Log [BzATP]
Δ In
trac
ellu
lar
Ca2+
(% o
f m
axim
al r
esp
on
se)
A
B
C
Fig. 2. BzATP induces intracellular calcium transients in nondifferen-
tiated N2a cells that are potentiated in the absence of Mg2+ ions. (A)
Intracellular calcium increments evoked by different purinergic agon-
ists in N2a cells. Cells were challenged with maximal concentrations
of the agonists a,b-meATP (100 lM), CTP (300 lM), BzATP (100 lM)
and ATP (100 lM), and the calcium transients were calculated. Hori-
zontal bars indicate stimulation periods. Trace represents
mean ± standard error of the mean (SEM) from 157 individual cells.
(B) Intracellular calcium increments elicited by BzATP (100 lM) in
N2a cells are potentiated in the absence of extracellular Mg2+ ions
and require the presence of extracellular calcium in the superfusion
medium. Horizontal bars indicate stimulation periods. Both Mg2+
washout and Tris ⁄ EGTA addition were performed 5 min before and
during BzATP application. Trace represents mean ± SEM from 108
individual cells. (C) Concentration–response curves for intracellular
calcium mobilization induced by BzATP in N2a cell suspensions in
the presence or absence of extracellular Mg2+ ions. Cell suspensions
were stimulated with graded doses of BzATP ranging from 5 lM to
1 mM prepared in Locke’s buffer or in Mg2+ -free medium. Results
were normalized with respect to the maximal response elicited by
BzATP, and the logistic curve was fitted by nonlinear regression.
Data are the mean ± SEM of at least five different experiments.
P2X7 negatively controls neuritogenesis via CaMKII R. Gomez-Villafuertes et al.
5310 FEBS Journal 276 (2009) 5307–5325 ª 2009 The Authors Journal compilation ª 2009 FEBS
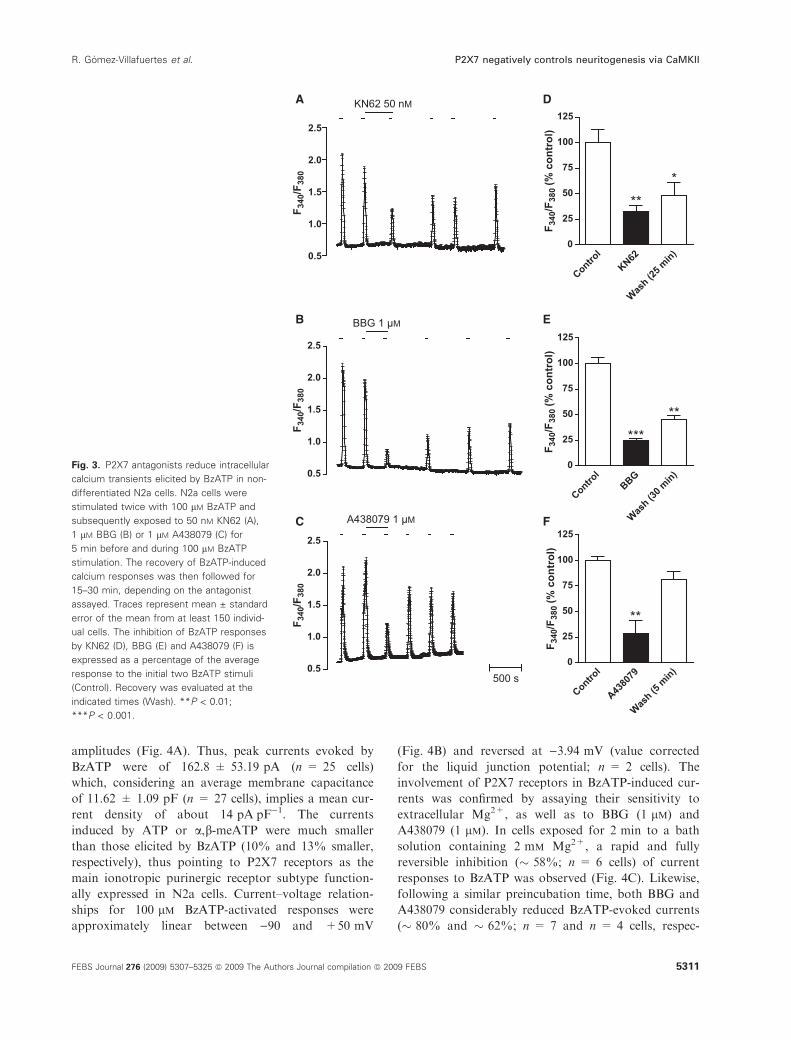
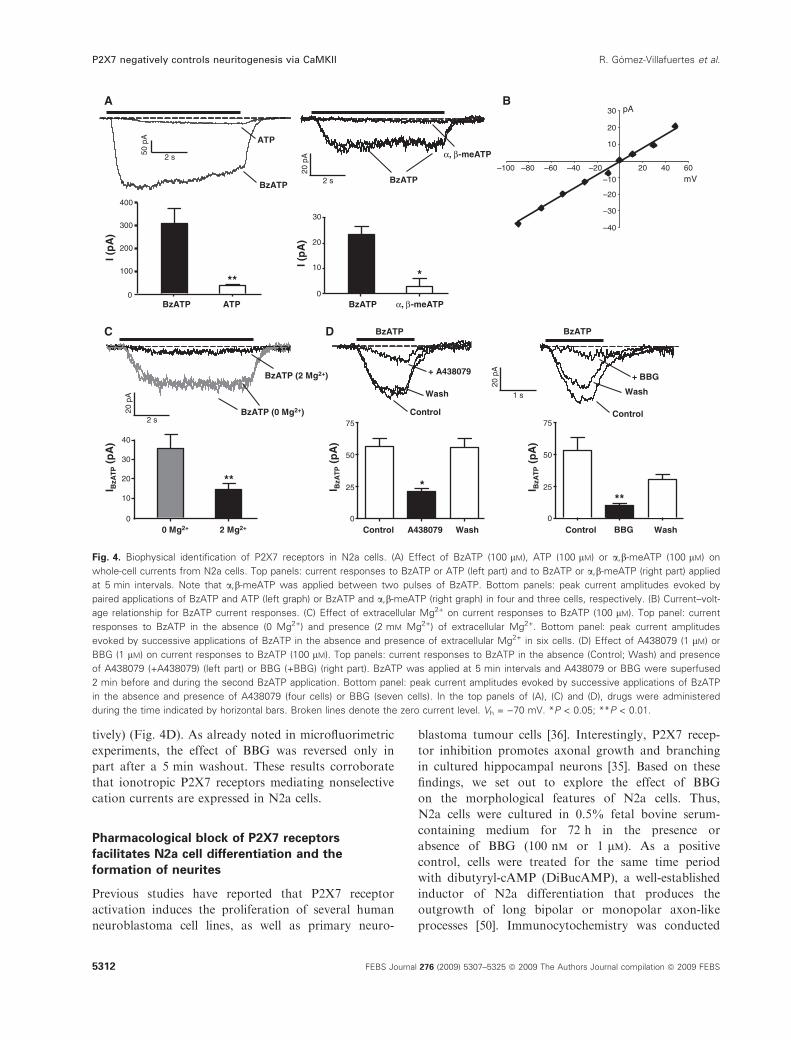
amplitudes (Fig. 4A). Thus, peak currents evoked by
BzATP were of 162.8 ± 53.19 pA (n = 25 cells)
which, considering an average membrane capacitance
of 11.62 ± 1.09 pF (n = 27 cells), implies a mean cur-
rent density of about 14 pAÆpF)1. The currents
induced by ATP or a,b-meATP were much smaller
than those elicited by BzATP (10% and 13% smaller,
respectively), thus pointing to P2X7 receptors as the
main ionotropic purinergic receptor subtype function-
ally expressed in N2a cells. Current–voltage relation-
ships for 100 lm BzATP-activated responses were
approximately linear between )90 and +50 mV
(Fig. 4B) and reversed at )3.94 mV (value corrected
for the liquid junction potential; n = 2 cells). The
involvement of P2X7 receptors in BzATP-induced cur-
rents was confirmed by assaying their sensitivity to
extracellular Mg2+, as well as to BBG (1 lm) and
A438079 (1 lm). In cells exposed for 2 min to a bath
solution containing 2 mm Mg2+, a rapid and fully
reversible inhibition (� 58%; n = 6 cells) of current
responses to BzATP was observed (Fig. 4C). Likewise,
following a similar preincubation time, both BBG and
A438079 considerably reduced BzATP-evoked currents
(� 80% and � 62%; n = 7 and n = 4 cells, respec-
Fig. 3. P2X7 antagonists reduce intracellular
calcium transients elicited by BzATP in non-
differentiated N2a cells. N2a cells were
stimulated twice with 100 lM BzATP and
subsequently exposed to 50 nM KN62 (A),
1 lM BBG (B) or 1 lM A438079 (C) for
5 min before and during 100 lM BzATP
stimulation. The recovery of BzATP-induced
calcium responses was then followed for
15–30 min, depending on the antagonist
assayed. Traces represent mean ± standard
error of the mean from at least 150 individ-
ual cells. The inhibition of BzATP responses
by KN62 (D), BBG (E) and A438079 (F) is
expressed as a percentage of the average
response to the initial two BzATP stimuli
(Control). Recovery was evaluated at the
indicated times (Wash). **P < 0.01;
***P < 0.001.
R. Gomez-Villafuertes et al. P2X7 negatively controls neuritogenesis via CaMKII
FEBS Journal 276 (2009) 5307–5325 ª 2009 The Authors Journal compilation ª 2009 FEBS 5311
tively) (Fig. 4D). As already noted in microfluorimetric
experiments, the effect of BBG was reversed only in
part after a 5 min washout. These results corroborate
that ionotropic P2X7 receptors mediating nonselective
cation currents are expressed in N2a cells.
Pharmacological block of P2X7 receptors
facilitates N2a cell differentiation and the
formation of neurites
Previous studies have reported that P2X7 receptor
activation induces the proliferation of several human
neuroblastoma cell lines, as well as primary neuro-
blastoma tumour cells [36]. Interestingly, P2X7 recep-
tor inhibition promotes axonal growth and branching
in cultured hippocampal neurons [35]. Based on these
findings, we set out to explore the effect of BBG
on the morphological features of N2a cells. Thus,
N2a cells were cultured in 0.5% fetal bovine serum-
containing medium for 72 h in the presence or
absence of BBG (100 nm or 1 lm). As a positive
control, cells were treated for the same time period
with dibutyryl-cAMP (DiBucAMP), a well-established
inductor of N2a differentiation that produces the
outgrowth of long bipolar or monopolar axon-like
processes [50]. Immunocytochemistry was conducted
0
10
20
30
40
0 Mg2+ 2 Mg2+
**
I BzA
TP
(pA
)
–40
–30
–20
–10
10
20
30
–100 –80 –60 –40 –20 20 40 60
mV
pA
BzATP
20 p
A
1 s
BzATP
I BzA
TP
(pA
)Control BBG Wash
0
25
50
75
**
I BzA
TP
(pA
)
*
Control A438079 Wash0
25
50
75
0
100
200
300
400
I (p
A)
BzATP ATP
**
Control
Wash
+ A438079
Control
Wash
+ BBG
50 p
A
2 s
BzATP
ATP
BzATP (0 Mg2+)20 p
A
2 s
α, β-meA
BA
C D
TP
BzATP
20 p
A
2 s
BzATP (2 Mg2+)
0
10
20
30
BzATP
*
I (p
A)
α, β-meATP
Fig. 4. Biophysical identification of P2X7 receptors in N2a cells. (A) Effect of BzATP (100 lM), ATP (100 lM) or a,b-meATP (100 lM) on
whole-cell currents from N2a cells. Top panels: current responses to BzATP or ATP (left part) and to BzATP or a,b-meATP (right part) applied
at 5 min intervals. Note that a,b-meATP was applied between two pulses of BzATP. Bottom panels: peak current amplitudes evoked by
paired applications of BzATP and ATP (left graph) or BzATP and a,b-meATP (right graph) in four and three cells, respectively. (B) Current–volt-
age relationship for BzATP current responses. (C) Effect of extracellular Mg2+ on current responses to BzATP (100 lM). Top panel: current
responses to BzATP in the absence (0 Mg2+) and presence (2 mM Mg2+) of extracellular Mg2+. Bottom panel: peak current amplitudes
evoked by successive applications of BzATP in the absence and presence of extracellular Mg2+ in six cells. (D) Effect of A438079 (1 lM) or
BBG (1 lM) on current responses to BzATP (100 lM). Top panels: current responses to BzATP in the absence (Control; Wash) and presence
of A438079 (+A438079) (left part) or BBG (+BBG) (right part). BzATP was applied at 5 min intervals and A438079 or BBG were superfused
2 min before and during the second BzATP application. Bottom panel: peak current amplitudes evoked by successive applications of BzATP
in the absence and presence of A438079 (four cells) or BBG (seven cells). In the top panels of (A), (C) and (D), drugs were administered
during the time indicated by horizontal bars. Broken lines denote the zero current level. Vh = )70 mV. *P < 0.05; **P < 0.01.
P2X7 negatively controls neuritogenesis via CaMKII R. Gomez-Villafuertes et al.
5312 FEBS Journal 276 (2009) 5307–5325 ª 2009 The Authors Journal compilation ª 2009 FEBS
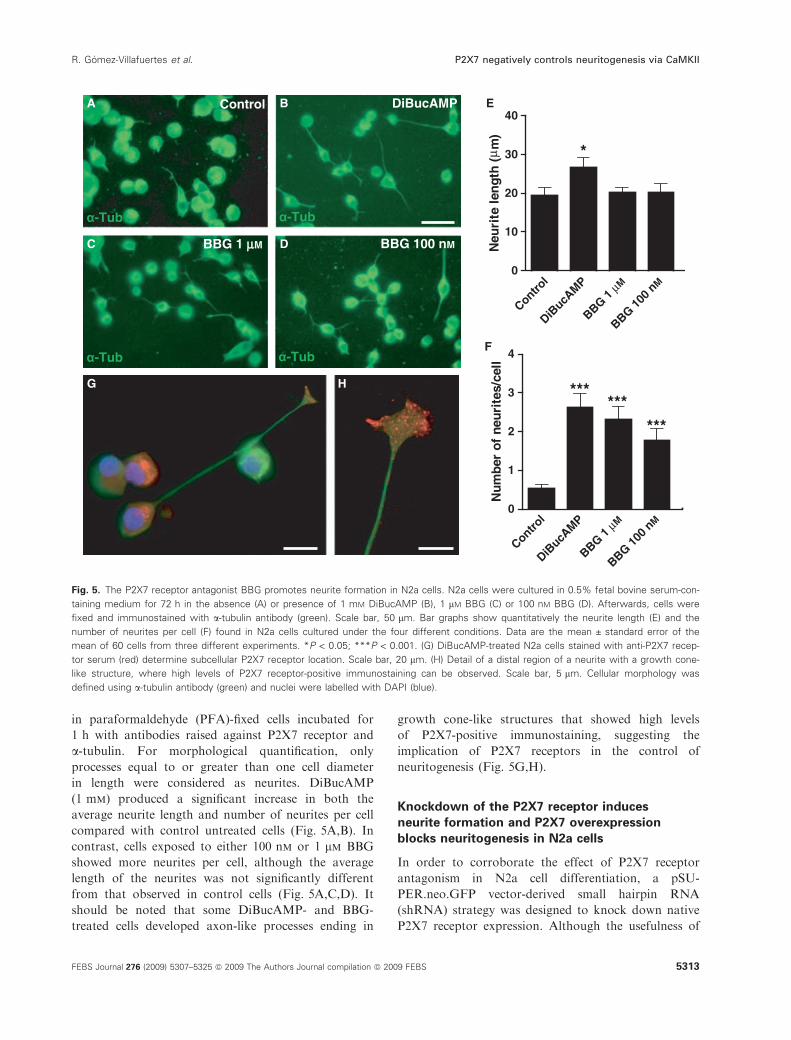
in paraformaldehyde (PFA)-fixed cells incubated for
1 h with antibodies raised against P2X7 receptor and
a-tubulin. For morphological quantification, only
processes equal to or greater than one cell diameter
in length were considered as neurites. DiBucAMP
(1 mm) produced a significant increase in both the
average neurite length and number of neurites per cell
compared with control untreated cells (Fig. 5A,B). In
contrast, cells exposed to either 100 nm or 1 lm BBG
showed more neurites per cell, although the average
length of the neurites was not significantly different
from that observed in control cells (Fig. 5A,C,D). It
should be noted that some DiBucAMP- and BBG-
treated cells developed axon-like processes ending in
growth cone-like structures that showed high levels
of P2X7-positive immunostaining, suggesting the
implication of P2X7 receptors in the control of
neuritogenesis (Fig. 5G,H).
Knockdown of the P2X7 receptor induces
neurite formation and P2X7 overexpression
blocks neuritogenesis in N2a cells
In order to corroborate the effect of P2X7 receptor
antagonism in N2a cell differentiation, a pSU-
PER.neo.GFP vector-derived small hairpin RNA
(shRNA) strategy was designed to knock down native
P2X7 receptor expression. Although the usefulness of
Control
BBG 1 μμM
α -Tub
0
10
20
30
40
* ( h
t g
n
e l e t i r u
e N
μ
) m
Control
DiBucA
MP
BBG 1 μM
BBG 100 n
M
Control
DiBucA
MP
BBG 1 μM
BBG 100 n
M
0
1
2
3
4
*** ***
***
l l e c / s e t i r u
e n
f o
r e
b
m
u
N
α -Tub
DiBucAMP
BBG 100 nM
α -Tub
α -Tub
A B E
F
C D
G H
Fig. 5. The P2X7 receptor antagonist BBG promotes neurite formation in N2a cells. N2a cells were cultured in 0.5% fetal bovine serum-con-
taining medium for 72 h in the absence (A) or presence of 1 mM DiBucAMP (B), 1 lM BBG (C) or 100 nM BBG (D). Afterwards, cells were
fixed and immunostained with a-tubulin antibody (green). Scale bar, 50 lm. Bar graphs show quantitatively the neurite length (E) and the
number of neurites per cell (F) found in N2a cells cultured under the four different conditions. Data are the mean ± standard error of the
mean of 60 cells from three different experiments. *P < 0.05; ***P < 0.001. (G) DiBucAMP-treated N2a cells stained with anti-P2X7 recep-
tor serum (red) determine subcellular P2X7 receptor location. Scale bar, 20 lm. (H) Detail of a distal region of a neurite with a growth cone-
like structure, where high levels of P2X7 receptor-positive immunostaining can be observed. Scale bar, 5 lm. Cellular morphology was
defined using a-tubulin antibody (green) and nuclei were labelled with DAPI (blue).
R. Gomez-Villafuertes et al. P2X7 negatively controls neuritogenesis via CaMKII
FEBS Journal 276 (2009) 5307–5325 ª 2009 The Authors Journal compilation ª 2009 FEBS 5313
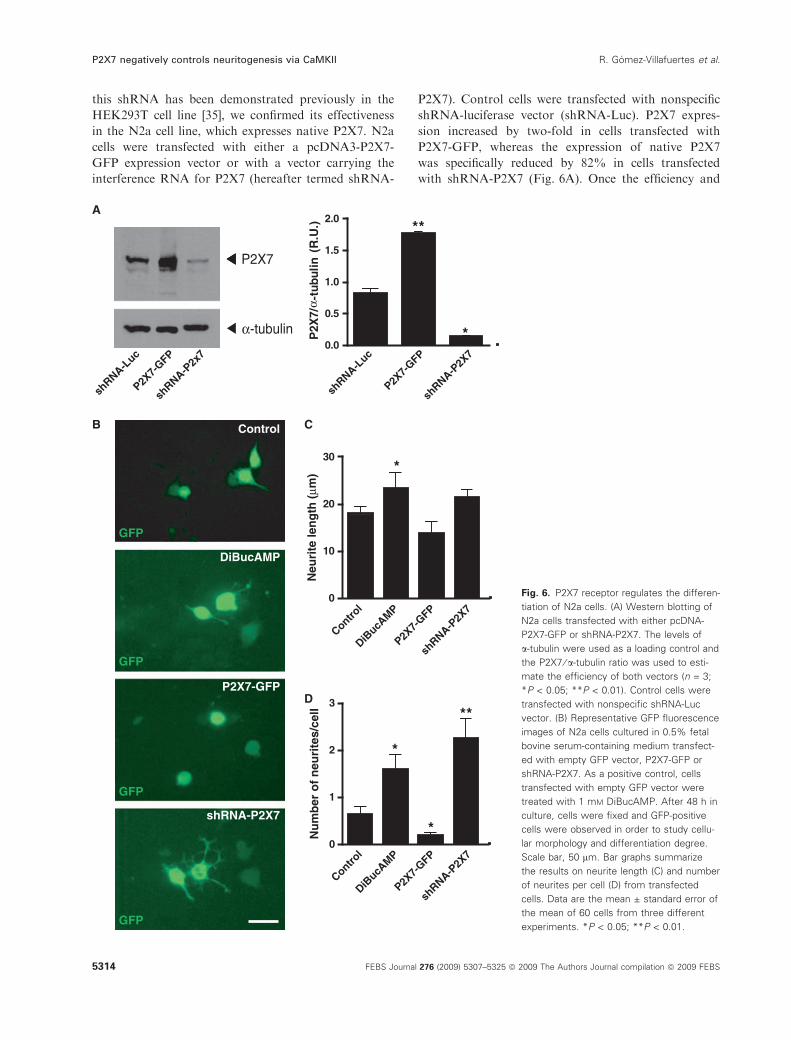
this shRNA has been demonstrated previously in the
HEK293T cell line [35], we confirmed its effectiveness
in the N2a cell line, which expresses native P2X7. N2a
cells were transfected with either a pcDNA3-P2X7-
GFP expression vector or with a vector carrying the
interference RNA for P2X7 (hereafter termed shRNA-
P2X7). Control cells were transfected with nonspecific
shRNA-luciferase vector (shRNA-Luc). P2X7 expres-
sion increased by two-fold in cells transfected with
P2X7-GFP, whereas the expression of native P2X7
was specifically reduced by 82% in cells transfected
with shRNA-P2X7 (Fig. 6A). Once the efficiency and
Control
DiBucAMP
shRNA-P2X7
GFP
GFP
P2X7-GFP
0
10
20
30
Neu
rite
len
gth
(μm
)
P2X7-
GFP
DiBucA
MP
Control
A N
R h
s
7 X 2 P
-
P2X7-
GFP
DiBucA
MP
Control
A N
R h
s
7 X 2 P
-
0
1
2
3
*
*
**
Nu
mb
er o
f n
euri
tes/
cell
GFP
GFP
*
P2X7
α -tubulin /7
X2P
α).
U.R(
nilubut-
c u L - A
N R
h s
P F
G - 7 X
2 P
7 x 2 P - A
N R
h s
c u L - A
N R
h s
P F
G - 7 X
2 P
7 X 2 P
- A N
R h
s
0.0
0.5
1.0
1.5
2.0 **
*
A
B C
D
Fig. 6. P2X7 receptor regulates the differen-
tiation of N2a cells. (A) Western blotting of
N2a cells transfected with either pcDNA-
P2X7-GFP or shRNA-P2X7. The levels of
a-tubulin were used as a loading control and
the P2X7 ⁄ a-tubulin ratio was used to esti-
mate the efficiency of both vectors (n = 3;
*P < 0.05; **P < 0.01). Control cells were
transfected with nonspecific shRNA-Luc
vector. (B) Representative GFP fluorescence
images of N2a cells cultured in 0.5% fetal
bovine serum-containing medium transfect-
ed with empty GFP vector, P2X7-GFP or
shRNA-P2X7. As a positive control, cells
transfected with empty GFP vector were
treated with 1 mM DiBucAMP. After 48 h in
culture, cells were fixed and GFP-positive
cells were observed in order to study cellu-
lar morphology and differentiation degree.
Scale bar, 50 lm. Bar graphs summarize
the results on neurite length (C) and number
of neurites per cell (D) from transfected
cells. Data are the mean ± standard error of
the mean of 60 cells from three different
experiments. *P < 0.05; **P < 0.01.
P2X7 negatively controls neuritogenesis via CaMKII R. Gomez-Villafuertes et al.
5314 FEBS Journal 276 (2009) 5307–5325 ª 2009 The Authors Journal compilation ª 2009 FEBS

specificity of both vectors had been confirmed, N2a
cells cultured in 0.5% fetal bovine serum-containing
medium were transfected with either P2X7-GFP or
shRNA-P2X7 and, 48 h later, were PFA-fixed and
analysed. Control cells were transfected with empty
green fluorescent protein (GFP) vector and, as a posi-
tive control, cells were transfected with empty GFP
vector and treated with 1 mm DiBucAMP for 48 h.
N2a cells that overexpressed the P2X7 receptor showed
a rounded morphology and were practically devoid of
visible neurites, hence indicating that P2X7 receptor
activation is involved in the maintenance of N2a cells
in a nondifferentiated state (Fig. 6B–D). As occurred
after BBG treatment, P2X7 receptor knockdown
promoted neurite formation in GFP-positive cells,
although, once again, differences in neurite average
length were only found in DiBucAMP-treated cells
(Fig. 6B–D). Interestingly, the silencing of P2X7 recep-
tors produced alterations in N2a cell morphology,
showing a broad increase in the number of neurites
that displayed lamellipodia-like morphology.
CaMKII-dependent signalling is modulated by
P2X7 receptors in N2a cells
Intracellular calcium is a critical regulator of neuronal
differentiation, neurite outgrowth and synaptic plastic-
ity (for a review, see [51]). It has been reported that
Ca2+ influx and the resulting transient elevation of
[Ca2+]i in growth cones decrease the rate of axon out-
growth, whereas the suppression of Ca2+ transients
accelerates axon extension [52]. CaMKII is a calcium-
dependent serine ⁄ threonine kinase that also possesses
the ability to become calcium independent through
autophosphorylation [53]. Moreover, once it is acti-
vated, it remains functional for at least several hours
[54]. Previously, we have reported that P2X7 receptor
activation induces CaMKII phosphorylation in cere-
bellar granule neurons [55], whereas P2X7 receptor
inhibition reduces its phosphorylation in cultured hip-
pocampal neurons [35]. As CaMKII and its down-
stream targets could be good effectors in N2a cell
differentiation induced by P2X7 antagonists, we analy-
sed the phosphorylation state of CaMKII in N2a cells
exposed to 100 nm BBG. This antagonist produced a
significant decrease in the ratio between phospho-
CaMKII (active form of the enzyme) and total CaM-
KII after 60 min of treatment (Fig. 7A–D), suggesting
that CaMKII participates in the negative regulation of
neuritogenesis exerted by P2X7 receptors. To corro-
borate this hypothesis, N2a cells were cultured in 0.5%
fetal bovine serum-containing medium for 72 h in the
presence or absence of 1 lm KN93, a well-known
CaMKII inhibitor. As positive controls, cells were
treated for the same time period with either 1 mm
DiBucAMP or 1 lm BBG. Immunocytochemistry was
performed in PFA-fixed cells incubated for 1 h with
a-tubulin antibodies. As reported previously, both
DiBucAMP and BBG produced a significant increase
in the number of neurites per cell compared with
untreated cells. As expected, cells exposed to KN93
also developed more neurites than control cells, point-
ing to a key role of this kinase in the control of
neuritogenesis in N2a cells (Fig. 5E).
Involvement of FAK in BBG-mediated neurite
formation in N2a cells
As shown previously, silencing of P2X7 receptors with
shRNAs produces a significant increase in the number
of neurites exhibiting a lamellipodia-like morphology.
This phenomenon may be explained by alterations in
the actin cytoskeleton, as reported previously in hippo-
campal neurons [35]. FAK is an actin-interacting pro-
tein that regulates focal adhesion contact formation
and lamellipodia stability [56]. It has been reported
that phospho-CaMKIIa is able to phosphorylate FAK
at Ser843 (pFAKS846), which results in a decrease in
FAK activity. In contrast, when the Ser843 residue is
dephosphorylated, FAK is autophosphorylated at
Tyr397 (pFAKT397) and becomes active [57]. In this
line, we examined whether the relative amount of
active pFAKT397 was modified in N2a cells following
BBG treatment. In agreement with a decrease in
CaMKII activity, Tyr397 phosphorylation of FAK
was significantly higher in N2a cells exposed to BBG,
indicating that P2X7 receptor inhibition produces an
increase in the activity of FAK (Fig. 7F,G).
PI3K-dependent signalling is modulated by P2X7
receptors in N2a cells
Previous studies have reported that one of the down-
stream targets of FAK is PI3K [58,59]. It is generally
accepted that PI3K plays a central role in cell signal-
ling, mediating neurite extension [60,61], cell prolifera-
tion, survival and motility [62,63]. Based on these
findings, we studied whether PI3K could participate in
the signalling cascade regulated by P2X7 receptors.
N2a cells cultured in 0.5% fetal bovine serum-contain-
ing medium were treated with 1 lm BBG for 72 h in
the presence or absence of the PI3K inhibitor
LY-294002 (50 lm). Cells were treated for the same
time period with 1 mm DiBucAMP as a positive
control. Immunocytochemical labelling was performed
in PFA-fixed cells incubated for 1 h with a-tubulin
R. Gomez-Villafuertes et al. P2X7 negatively controls neuritogenesis via CaMKII
FEBS Journal 276 (2009) 5307–5325 ª 2009 The Authors Journal compilation ª 2009 FEBS 5315
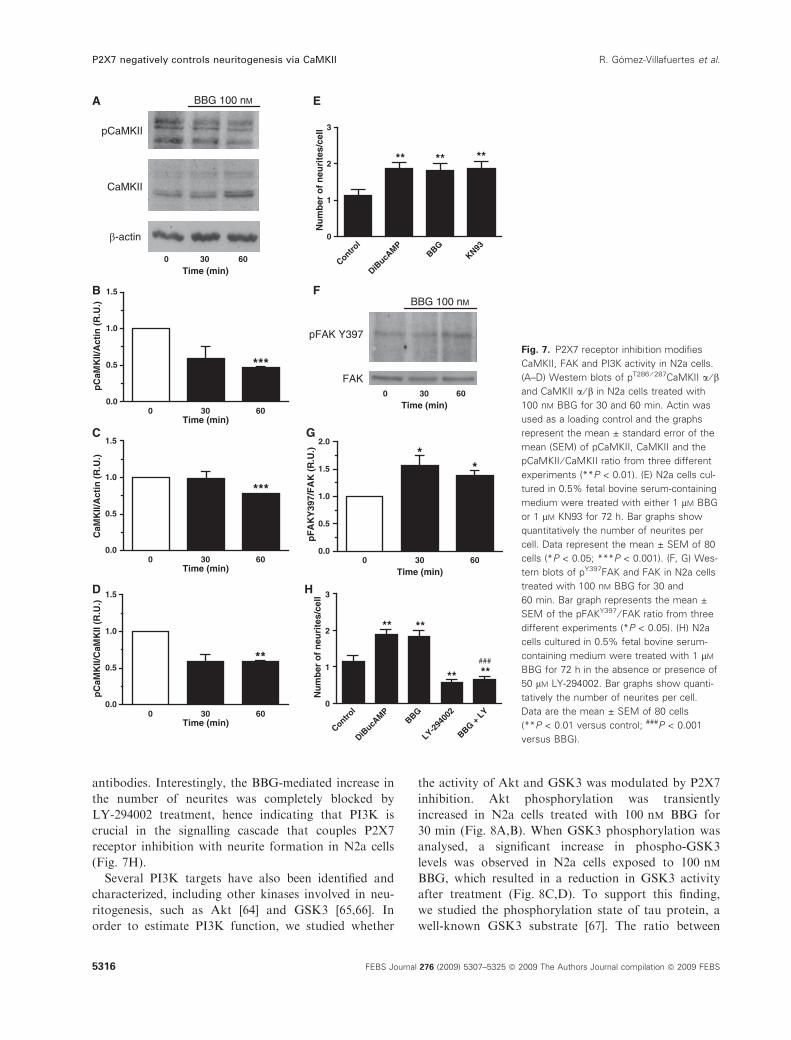
antibodies. Interestingly, the BBG-mediated increase in
the number of neurites was completely blocked by
LY-294002 treatment, hence indicating that PI3K is
crucial in the signalling cascade that couples P2X7
receptor inhibition with neurite formation in N2a cells
(Fig. 7H).
Several PI3K targets have also been identified and
characterized, including other kinases involved in neu-
ritogenesis, such as Akt [64] and GSK3 [65,66]. In
order to estimate PI3K function, we studied whether
the activity of Akt and GSK3 was modulated by P2X7
inhibition. Akt phosphorylation was transiently
increased in N2a cells treated with 100 nm BBG for
30 min (Fig. 8A,B). When GSK3 phosphorylation was
analysed, a significant increase in phospho-GSK3
levels was observed in N2a cells exposed to 100 nm
BBG, which resulted in a reduction in GSK3 activity
after treatment (Fig. 8C,D). To support this finding,
we studied the phosphorylation state of tau protein, a
well-known GSK3 substrate [67]. The ratio between
0.0
0.5
1.0
1.5
0.0
0.5
1.0
1.5
**
0 30 60Time (min)
0 30 60Time (min)
0 30 60Time (min)
pC
aMK
II/C
aMK
II (R
.U.)
0.0
0.5
1.0
1.5
2.0
**
Time (min)
pF
AK
Y39
7/F
AK
(R
.U.)
***
CaM
KII/
Act
in (
R.U
.)
0 30 60
0.0
0.5
1.0
1.5
***
pC
aMK
II/A
ctin
(R
.U.)
BBG 100 nMA E
B F
C G
D H
pCaMKII
CaMKII
-actin
0 30 60Time (min)
pFAK Y397
FAK
BBG 100 nM
0 30 60Time (min)
Control
DiBucA
MPBBG
KN930
1
2
3
** ****
Nu
mb
er o
f n
euri
tes/
cell
Control
DiBucA
MPBBG
LY-294
002
BBG + L
Y0
1
2
3
**
**###
**
**
Nu
mb
er o
f n
euri
tes/
cell
Fig. 7. P2X7 receptor inhibition modifies
CaMKII, FAK and PI3K activity in N2a cells.
(A–D) Western blots of pT286 ⁄ 287CaMKII a ⁄ band CaMKII a ⁄ b in N2a cells treated with
100 nM BBG for 30 and 60 min. Actin was
used as a loading control and the graphs
represent the mean ± standard error of the
mean (SEM) of pCaMKII, CaMKII and the
pCaMKII ⁄ CaMKII ratio from three different
experiments (**P < 0.01). (E) N2a cells cul-
tured in 0.5% fetal bovine serum-containing
medium were treated with either 1 lM BBG
or 1 lM KN93 for 72 h. Bar graphs show
quantitatively the number of neurites per
cell. Data represent the mean ± SEM of 80
cells (*P < 0.05; ***P < 0.001). (F, G) Wes-
tern blots of pY397FAK and FAK in N2a cells
treated with 100 nM BBG for 30 and
60 min. Bar graph represents the mean ±
SEM of the pFAKY397 ⁄ FAK ratio from three
different experiments (*P < 0.05). (H) N2a
cells cultured in 0.5% fetal bovine serum-
containing medium were treated with 1 lM
BBG for 72 h in the absence or presence of
50 lM LY-294002. Bar graphs show quanti-
tatively the number of neurites per cell.
Data are the mean ± SEM of 80 cells
(**P < 0.01 versus control; ###P < 0.001
versus BBG).
P2X7 negatively controls neuritogenesis via CaMKII R. Gomez-Villafuertes et al.
5316 FEBS Journal 276 (2009) 5307–5325 ª 2009 The Authors Journal compilation ª 2009 FEBS
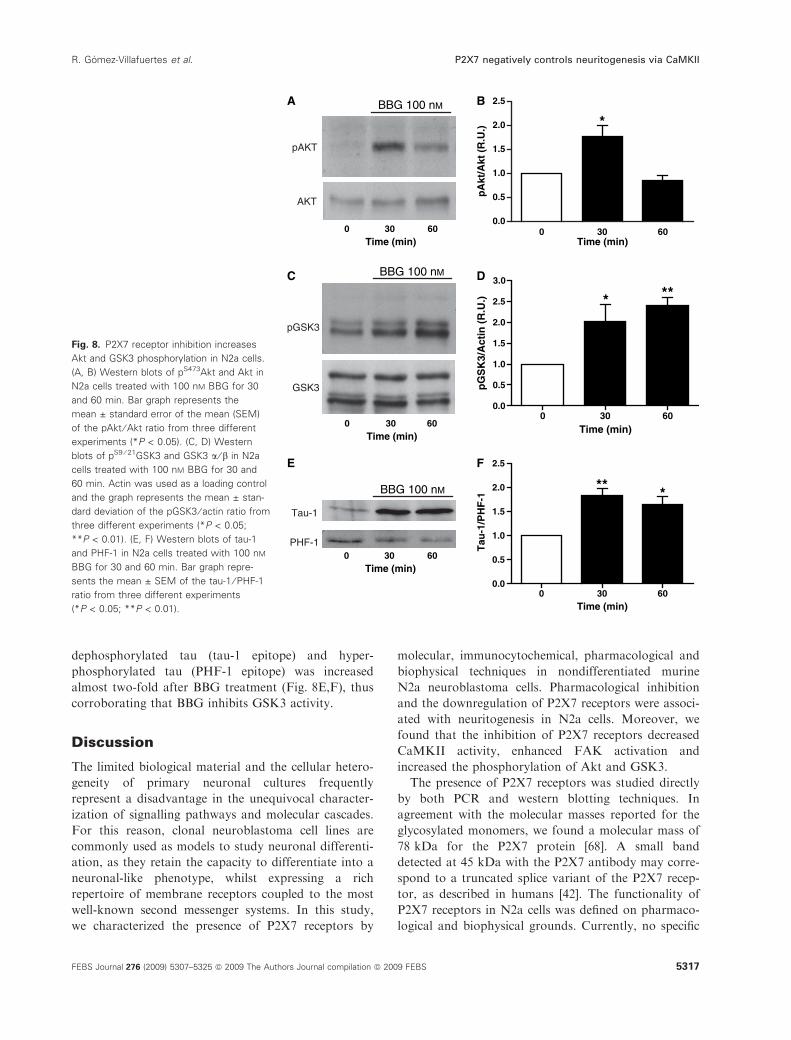
dephosphorylated tau (tau-1 epitope) and hyper-
phosphorylated tau (PHF-1 epitope) was increased
almost two-fold after BBG treatment (Fig. 8E,F), thus
corroborating that BBG inhibits GSK3 activity.
Discussion
The limited biological material and the cellular hetero-
geneity of primary neuronal cultures frequently
represent a disadvantage in the unequivocal character-
ization of signalling pathways and molecular cascades.
For this reason, clonal neuroblastoma cell lines are
commonly used as models to study neuronal differenti-
ation, as they retain the capacity to differentiate into a
neuronal-like phenotype, whilst expressing a rich
repertoire of membrane receptors coupled to the most
well-known second messenger systems. In this study,
we characterized the presence of P2X7 receptors by
molecular, immunocytochemical, pharmacological and
biophysical techniques in nondifferentiated murine
N2a neuroblastoma cells. Pharmacological inhibition
and the downregulation of P2X7 receptors were associ-
ated with neuritogenesis in N2a cells. Moreover, we
found that the inhibition of P2X7 receptors decreased
CaMKII activity, enhanced FAK activation and
increased the phosphorylation of Akt and GSK3.
The presence of P2X7 receptors was studied directly
by both PCR and western blotting techniques. In
agreement with the molecular masses reported for the
glycosylated monomers, we found a molecular mass of
78 kDa for the P2X7 protein [68]. A small band
detected at 45 kDa with the P2X7 antibody may corre-
spond to a truncated splice variant of the P2X7 recep-
tor, as described in humans [42]. The functionality of
P2X7 receptors in N2a cells was defined on pharmaco-
logical and biophysical grounds. Currently, no specific
GSK3
pGSK3
0 30 60Time (min)
0 30 60Time (min)
Tau-1
PHF-1
BBG 100 nM
BBG 100 nM
0 30 60
***
Time (min)
pG
SK
3/A
ctin
(R
.U.)
0 30 600.0
0.5
1.0
1.5
2.0
2.5
0.0
0.5
1.0
1.5
2.0
2.5
0.0
0.5
1.0
1.5
2.0
2.5
3.0
** *
Time (min)
Tau
-1/P
HF
-1
0 30 60
*
Time (min)
pA
kt/A
kt (
R.U
.)
AKT
pAKT
0 30 60Time (min)
BBG 100 nMA B
C D
E F
Fig. 8. P2X7 receptor inhibition increases
Akt and GSK3 phosphorylation in N2a cells.
(A, B) Western blots of pS473Akt and Akt in
N2a cells treated with 100 nM BBG for 30
and 60 min. Bar graph represents the
mean ± standard error of the mean (SEM)
of the pAkt ⁄ Akt ratio from three different
experiments (*P < 0.05). (C, D) Western
blots of pS9 ⁄ 21GSK3 and GSK3 a ⁄ b in N2a
cells treated with 100 nM BBG for 30 and
60 min. Actin was used as a loading control
and the graph represents the mean ± stan-
dard deviation of the pGSK3 ⁄ actin ratio from
three different experiments (*P < 0.05;
**P < 0.01). (E, F) Western blots of tau-1
and PHF-1 in N2a cells treated with 100 nM
BBG for 30 and 60 min. Bar graph repre-
sents the mean ± SEM of the tau-1 ⁄ PHF-1
ratio from three different experiments
(*P < 0.05; **P < 0.01).
R. Gomez-Villafuertes et al. P2X7 negatively controls neuritogenesis via CaMKII
FEBS Journal 276 (2009) 5307–5325 ª 2009 The Authors Journal compilation ª 2009 FEBS 5317

agonists or antagonists exist for all of the P2X recep-
tors, and therefore their pharmacological identification
relies on the effects of a number of compounds (for a
review, see [69]). ATP and BzATP were able to induce
an increase in [Ca2+]i, whereas a,b-meATP and CTP
were ineffective. The potency for BzATP was increased
by two-fold following the omission of Mg2+ from the
extracellular medium, suggesting that ATP4) is the
active ligand. Likewise, the fact that an increase in
[Ca2+]i could not be evoked by a,b-meATP clearly
demonstrated that, if present, P2X1 and P2X3 recep-
tors are not functionally prominent in N2a cells [70].
The lack of CTP-mediated calcium responses also
excludes the presence of functional P2X4 receptors in
these cells [41]. In contrast, both the substantial
increase in [Ca2+]i and the small ionic current evoked
by micromolar concentrations of ATP indicate that, in
this cell model, the response to micromolar ATP con-
centration is mainly mediated by metabotropic P2Y
receptor activation, as reported previously [43]. The
P2X7 receptor antagonists KN62, BBG and A438079
inhibited BzATP-evoked calcium responses, thus dem-
onstrating the presence of P2X7 receptors [47–49].
Interestingly, both KN62 and BBG behaved as slowly
reversible antagonists, whereas the inhibitory effect of
A438079 was rapidly reversed. In addition, voltage-
clamp studies showed that BzATP elicits nondesensitiz-
ing inward currents in N2a cells, with current–voltage
relationships that do not display rectification. More-
over, the fact that BzATP-activated currents reversed
at about 0 mV and were sensitive to BBG and
A438079 indicates that they are mediated by genuine
P2X7 receptors. Together, these data point to the exis-
tence of native functional P2X7 receptors in N2a cells.
Calcium is an essential second messenger involved in
neuronal remodelling, this term referring to neuronal
differentiation, neurite outgrowth and synaptic plastic-
ity. Our results demonstrate that BzATP induces an
increase in [Ca2+]i in N2a cells that can be prevented
by the addition of specific P2X7 receptor antagonists
(BBG, KN62 and A438079) and is associated with
opposite changes (inhibition or facilitation, respec-
tively) in neuritogenesis. A trophic effect of ATP in
regeneration and neurite development has been
reported previously in various cell systems, such as
phaeochromocytoma cells (PC12), retinal ganglion cells
and striatal neurons [29–32]. It is worth mentioning
that ATP, acting via metabotropic P2Y receptors, can
also stimulate neurite outgrowth in N2a cells, indepen-
dent of calcium entry and other neurotrophic factors
[43]. With regard to P2X7 receptors, previous studies
have shown that ATP, via P2X7 receptors, sustains the
growth of human neuroblastoma cells through a sub-
stance P-dependent mechanism [36], and that a func-
tional decrease in P2X7 receptors seems to be
associated with retinoic acid-induced differentiation of
N2a cells [37]. In this context, the current study shows
that pharmacological inhibition of P2X7 receptors and
their downregulation with shRNAs promote neurite
formation in N2a cells, whereas P2X7 overexpression
significantly reduces neuritogenesis. Furthermore, a
local control of neurite formation by P2X7 receptors is
supported by the strong P2X7-positive immunolabel-
ling detected at the axon-like processes and growth
cone-like structures observed in a fraction of N2a cells
treated with DiBucAMP or BBG. It is obvious that
the effects caused by the functional inhibition of P2X7
receptors (either by pharmacological or molecular pro-
cedures) must be derived from the removal of an extra-
cellular nucleotidic tone. Interestingly, it has been
reported that neuroblastoma cells maintain a steady
ATP concentration in the incubation medium close to
100–200 nmolÆL)1 [36]. Moreover, in some physiologi-
cal fluids, such as aqueous humour, an ATP concen-
tration close to 1 lm can be reached [71]. An open
question is the source of extracellular ATP. It is well
known that ATP does not cross the plasma membrane
because of its negative charge, and cells release ATP
either alone or together with other mediators by vesic-
ular or granule secretion. In neuronal cells, ATP is
packaged within vesicles close to the presynaptic mem-
brane, and the stimulation of exocytosis leads to the
release of ATP into the extracellular medium [72,73].
Concerning other mechanisms implicated in nonvesicu-
lar ATP secretion, there is clear evidence to support
the release of ATP in a regulated manner through
connexons, also known as hemichannels [74].
Previous studies have linked increases in [Ca2+]iwith microtubule depolymerization [75]. Indeed, the
transient receptor potential vanilloid receptor 1, a non-
selective cation channel, plays an inhibitory role in sen-
sory neuronal extension and motility by regulating the
disassembly of microtubules [76]. In a similar way,
P2X7 receptors could be mediating cytoskeleton reor-
ganization in a Ca2+ -dependent manner and, conse-
quently, the repression of neurite outgrowth. It should
be noted that P2X7 receptors can interact directly with
structural proteins such as b-actin, a-actinin, lami-
nin a3 and integrin b2 in HEK cells, some of which
might initiate cytoskeletal rearrangements following
receptor activation [77]. We have reported previously
that P2X7 receptor activation is coupled to CaMKII
phosphorylation in cerebellar granule neurons [55] and
hippocampal neurons in culture [35]. In neurons,
CaMKII inhibition induces the reorganization of
F-actin and the formation of growth cones with a
P2X7 negatively controls neuritogenesis via CaMKII R. Gomez-Villafuertes et al.
5318 FEBS Journal 276 (2009) 5307–5325 ª 2009 The Authors Journal compilation ª 2009 FEBS

filopodial structure [78]. Interestingly, our data show
that P2X7 receptor inhibition by BBG reduces the
amount of phospho-CaMKII, the active form of the
enzyme, thus pointing towards a common biochemical
mechanism in the control of neuritogenesis by P2X7
receptors in both primary neurons and neuron-like
tumour cells.
In human neuroblastoma cells, insulin-like growth
factor-I produces morphological changes, accompanied
by actin cytoskeletal rearrangement, followed by neu-
ronal differentiation through the activation of FAK
[79]. FAK is an integrin adaptor protein crucial for
focal adhesion formation, tumour migration [80], axo-
nal branching and synapse configuration [81]. FAK
activity is reduced by CaMKII-mediated phosphoryla-
tion at Ser843, whereas, in the absence of this process,
FAK is autophosphorylated at Tyr397, thereby
increasing its activity [57]. This mechanism underlies
the ability of local [Ca2+]i transients to reduce
pFAKY397, and thus to promote focal complex
removal and deadhesion of neuronal growth cones and
fibroblasts [82]. In our studies, phosphorylation of
Tyr397 is augmented after P2X7 receptor inhibition,
indicating that neurite formation induced by BBG is
probably a result of a decrease in calcium influx
through P2X7 receptors.
In an attempt to go further in the signalling path-
way regulated by P2X7 receptors in N2a cells, we eval-
uated the activity of PI3K, Akt and GSK3, all protein
kinases involved in neuritogenesis [60,61,64–66]. Both
Akt and GSK3 are phosphorylated by PI3K, a down-
stream target of pFAKY397 [58,59]. In a previous
study, we characterized the participation of PI3K in
the control of axonal growth and branching in hippo-
campal neurons [35]. The present study also demon-
strates that PI3K plays a key role in the formation of
neurites induced by P2X7 receptor inhibition. More-
over, our results show that P2X7 receptor inhibition
increases the phosphorylation of two PI3K targets,
Akt and GSK3, resulting in Akt activation and GSK3
inhibition. It should be noted that increased Akt activ-
ity can also induce the inhibition of GSK3, which has
been proposed to promote axonal growth and branch-
ing in neurons [83,84] and neurite formation in N2a
cells [85]. The inhibition of GSK3 activity by BBG was
confirmed by the augmentation of dephosphorylated
tau, a well-established GSK3 substrate [67].
Taken together, our results provide the first mecha-
nistic insight into the P2X7-triggered signalling path-
ways regulating neurite formation in N2a cells. P2X7
receptor inhibition leads to increased neurite formation
in N2a cells, this effect occurring in parallel with a
modification of the phosphorylation state of CaMKII,
FAK, Akt and GSK3, all kinases related to axonal
growth and cellular differentiation. In addition, our
results indicate that N2a cells may be considered to be
a reliable and convenient model for the initial charac-
terization of the signalling cascades coupled to P2X
receptors in the nervous system. This cellular model
may also be an excellent system to assay the effect of a
large number of compounds that may be potential
modulators of axonal growth and regeneration via
P2X7 receptors. Moreover, our data support the
involvement of P2X7 receptors in the maintenance of
neuroblastoma cells in a nondifferentiated state, and
point to P2X7 receptor antagonists as future potential
therapeutic agents in the treatment of neuroblastoma
tumour cells.
Experimental procedures
Chemicals and antibodies
ATP, UTP, CTP, a,b-meATP, BzATP, BBG, BSA, DiBu-
cAMP and DirectLoad� Wide Range DNA marker were
purchased from Sigma (St Louis, MO, USA), Fura-2 AM
was obtained from Molecular Probes (Leiden, the Nether-
lands), and KN62, KN93, LY-294002 and A438079 were
obtained from Tocris Bioscience (Bristol, UK). The com-
mercial antibodies used in this study were raised against
mouse P2X1, P2X2, P2X3, P2X4, P2X5, P2X6 and P2X7
(intracellular epitope) receptors, and all were purchased
from Alomone Labs (Jerusalem, Israel); phospho-CaM-
KII a ⁄ b (pT286 ⁄ 287) antibody was obtained from Upstate
Cell Signalling Solutions (Lake Placid, NY, USA); CaM-
KII a ⁄ b, phospho-FAK (pY397), FAK and GSK3 a ⁄ bantibodies were purchased from Invitrogen (San Francisco,
CA, USA); phospho-GSK3 (pS9 ⁄ 21) and phospho-Akt
(pS473) were obtained from Cell Signalling (Beverly, MA,
USA); Akt was obtained from Santa Cruz Biotechnology
(Santa Cruz, CA, USA); and tau-1 was purchased from
Chemicon (Temecula, CA, USA). PHF-1 antibody was a
kind gift from Dr Jesus Avila (CBM, Madrid, Spain).
Cell culture
N2a cells were cultured in DMEM (Sigma) supplemented
with glutamax� (Invitrogen), penicillin ⁄ streptomycin (Invi-
trogen) and 10% heat-inactivated fetal bovine serum (Euro-
Clone, Padova, Italy). Cells were grown at 37 �C in a
humidified atmosphere containing 5% CO2.
Transfections
N2a cells were plated at 4 · 104 cellsÆcm)2 and transiently
transfected using Lipofectamine� 2000 (Life Technologies,
Milan, Italy) following the manufacturer’s instructions.
R. Gomez-Villafuertes et al. P2X7 negatively controls neuritogenesis via CaMKII
FEBS Journal 276 (2009) 5307–5325 ª 2009 The Authors Journal compilation ª 2009 FEBS 5319

After 6 h, the medium was removed and the cells were fur-
ther incubated for the indicated time periods in culture
medium. Expression vectors and shRNAs were obtained as
reported previously [35]. Briefly, human P2X7 full-length
cDNA was cloned into the pd2EGFP-N1 vector (Clontech,
Mountain View, CA, USA), and the ligation product was
confirmed by sequencing. P2X7 knockdown was achieved
by RNA interference (RNAi) using a vector-based shRNA
approach (pSUPER.neo.GFP vector, OligoEngine, Seattle,
WA, USA). The concomitant expression of GFP allowed
transfected cells to be identified by fluorescence microscopy.
RT-PCR and quantitative real-time PCR
Total RNA was extracted from cultured N2a cells and from
whole adult mice brain (strain C57BL ⁄ 6J) using an
Rneasy� plus mini kit (Qiagen, Hilden, Germany), follow-
ing the manufacturer’s instructions. After digestion with
TURBO DNase (Ambion, Austin, TX, USA), total RNA
was quantified and reversed transcribed using M-MLV
reverse transcriptase, 6 lg of random primers and 350 lm
dNTPs (all from Invitrogen). PCRs were carried out using
AmpliTaq Gold� PCR Master Mix (Applied Biosystems,
Foster City, CA, USA), 5 lL of the RT product and spe-
cific commercial oligonucleotide primers for mouse P2X1–5
and P2X7 receptors, as well as for b-actin (Applied Biosys-
tems). P2X6-specific primers were designed using Primer
Express 3.0 Software (Applied Biosystems): forward primer,
5¢-AGGGCAGATGTCCAGAGCAT-3¢; reverse primer,
5¢-GTCTTCATCAGCCCAGCAGTT-3¢. PCRs involved an
initial denaturation step at 94 �C for 5 min, followed by 40
cycles of amplification (94 �C for 30 s, 60 �C for 30 s and
72 �C for 30 s) conducted with a thermocycler GeneAmp
PCR System 2400 (Applied Biosystems). Control reactions
were carried out in the absence of RT product (template)
to avoid cross-contamination. Amplified PCR products
were electrophoresed on a 1% agarose gel and visualized
by SYBR� Safe DNA gel stain (Invitrogen).
Quantitative real-time PCRs were performed using gene-
specific primers and TaqMan MGB probes for mouse
P2X1–5 and P2X7 receptors, and b-actin (all from Applied
Biosystems). For the P2X6 receptor, the primers used have
been described previously and the probe design was FAM-
5¢-CTTCCGTTCCTCTGGC-3¢-MGB. Fast thermal cycling
was performed using a StepOnePlus� Real-Time PCR Sys-
tem (Applied Biosystems) as follows: denaturation, one
cycle of 95 �C for 20 s, followed by 40 cycles each of 95 �Cfor 1 s and 60 �C for 20 s. The results were normalized as
indicated by parallel amplification of the endogenous
control b-actin.
Western blotting
N2a cells were lysed and homogenized for 1 h at 4 �C in
lysis buffer containing 50 mm Tris ⁄HCl, 150 mm NaCl,
1% Nonidet P40 and Complete� Protease Inhibitor Cock-
tail Tablets (Roche Diagnostics GmbH, Mannheim,
Germany), pH 7.4. Separation of the proteins was per-
formed on 10% SDS-PAGE gels. Proteins were transferred
to poly(vinylidene difluoride) (Hybond�-P, Amersham GE
Healthcare, Madrid, Spain) or nitrocellulose membranes,
saturated for 1 h at room temperature with 5% nonfat
dried milk in Tris-buffered saline (10 mm Tris ⁄HCl,
100 mm NaCl and 0.1% Tween; pH 7.5) (TBS-T) and
incubated overnight at 4 �C with the following antisera at
the dilutions specified in parentheses: P2X1 (1 : 200), P2X2
(1 : 200), P2X3 (1 : 500), P2X4 (1 : 500), P2X5 (1 : 400),
P2X6 (1 : 200), P2X7 (1 : 250), pT286 ⁄ 287CaMKII a ⁄ b(1 : 1000), Akt (1 : 1000), pS473Akt (1 : 1000), pS9 ⁄ 21GSK3
(1 : 1000), pY397FAK (1 : 1000) and FAK (1 : 1000). The
monoclonal antibodies used in this study were as follows
(dilution specified in parentheses): a-tubulin (1 : 10 000),
CaMKII a ⁄ b (1 : 1000), GSK3 a ⁄ b (1 : 1000), tau-1
(1 : 5000) and PHF-1 (1 : 100). Blots were then washed in
TBS-T, and incubated for 1 h at room temperature with
goat anti-rabbit or goat anti-mouse IgGs coupled to horse-
radish peroxidase (HRP, Amersham GE Healthcare), both
used at 1 : 5000 dilution. Protein bands were detected by
ECL chemiluminescence (Amersham GE Healthcare).
Calcium microfluorimetric analysis in single cells
N2a cells cultured on coverslips placed in 35 mm dishes
(250 000 cells per well) were washed with Locke’s solution
(composition in mm: NaCl, 140; KCl, 4.5; CaCl2, 2.5;
KH2PO4, 1.2; MgSO4, 1.2; glucose, 5.5; Hepes, 10; pH 7.4),
and loaded with the calcium dye Fura-2 AM (5 lm) for
45 min at 37 �C. Subsequently, the coverslips were washed
and placed in a superfusion chamber on a Nikon Eclipse
TE-2000 microscope (Nikon, Barcelona, Spain), where the
cells were continuously superfused with Locke’s medium at
a rate of 1.5 mLÆmin)1. N2a cells were stimulated for 40 s
with a variety of purinergic receptor agonists at near-maxi-
mal effective concentration: 100 lm a,b-meATP, 300 lm
CTP, 100 lm BzATP and 100 lm ATP. In other studies,
the P2X7 antagonists KN62 (50 nm), BBG (1 lm) and
A438079 (1 lm) were preincubated for 5 min before BzATP
(100 lm) superfusion. In some experiments, an Mg2+-free
Locke’s solution was used, replacing MgSO4 by glucose at
a concentration that conserved the solution osmolarity.
Cells were visualized using a Plan Fluor 20· ⁄ 0.5 lens on a
Nikon Eclipse TE-2000-E microscope. The wavelength of
the incoming light (340 or 380 nm) was selected with the
aid of an Optoscan monochromator (10 nm bandwidth),
from Cairn Research (Faversham, UK); 12-bit images were
acquired with an ORCA-ER C 47 42–98 CCD camera
(Hamamatsu, Barcelona, Spain) controlled by Metafluor
6.3r6 PC software (Universal Imaging Corp., Cambridge,
UK). The exposure time was 250 ms for each wavelength
and the changing time was less than 5 ms. The images were
P2X7 negatively controls neuritogenesis via CaMKII R. Gomez-Villafuertes et al.
5320 FEBS Journal 276 (2009) 5307–5325 ª 2009 The Authors Journal compilation ª 2009 FEBS

acquired continuously and buffered in a fast SCSI disk.
The time course data represent the average light intensity in
a small elliptical region within each cell. The background
was subtracted at each wavelength and the 340 ⁄ 380 ratio
was calculated. The data are represented as the normalized
F340 ⁄F380 fluorescence ratio, which increases as [Ca2+]iincreases.
Intracellular calcium fluorimetric analysis in
cellular suspensions
Population-averaged intracellular Ca2+ changes were also
determined with the fluorescent indicator Fura-2 AM. N2a
cells were collected by trypsinization from confluent cul-
tures in 75 cm2 flasks, washed and resuspended in Locke’s
solution. The cells were then loaded by incubation with
5 lm Fura-2 AM for 45 min at 37 �C in Locke’s solution
containing 1 mgÆmL)1 BSA. After the loading period, the
cells were washed in fresh Locke’s solution and resuspended
at a density of 106 cellsÆmL)1. Recordings were made on
1.5 mL samples in thermostatically controlled, stirred
cuvettes in a LS-50B fluorometer (Perkin Elmer, Waltham,
MA, USA). The fluorescence intensity was determined with
an excitation wavelength of 340 nm and an emission wave-
length of 510 nm. The results were normalized with respect
to the maximal response elicited by BzATP. Agonists and
other compounds were added to the cuvettes from at least
100-fold concentrated stock solutions to avoid large volume
variations. Fluorescence traces were calibrated individually
by releasing intracellular dye contents with Triton X-100
(0.3%) and determining dye fluorescence in the presence of
a mixture of EGTA ⁄Tris ([Ca2+] < 0.2 nm) and 2.5 mm
Ca2+ to calculate Fmin and Fmax, respectively. [Ca2+]i was
calculated from the fluorescence traces using the equation
of Grynkiewicz [86].
Electrophysiological recordings
Electrophysiological recordings were performed with an
EPC9 patch-clamp amplifier using pulse software (HEKA
Electronic, Lambrecht, Germany). Pipettes were pulled
from Kimax borosilicate glass (Witz Scientific, Holland,
OH, USA) and subsequently wax-coated and fire-polished
to obtain a final resistance of 2–3 MX when filled with
standard solutions. The standard extracellular solution
(bath) contained (in mm): NaCl, 140; KCl, 2.8; CaCl2, 2;
Hepes, 10; glucose, 10 (pH 7.2, adjusted with NaOH;
298 mOsm). Recording pipettes were filled with a solution
containing (in mm): KCl, 140; MgCl2, 1; Hepes, 10
(pH 7.2, adjusted with KOH; 292 mOsm). Cells attached to
glass coverslips were transferred to a recording chamber
placed in the stage of an inverted Zeiss Axiovert 100 micro-
scope (Zeiss, Barcelona, Spain) and continuously super-
fused with bath solution (perfusion rate of 1 mLÆmin)1).
Membrane currents were measured in the whole-cell config-
uration of the patch-clamp technique, filtered at 3 kHz and
sampled at 10 kHz. Once electrical access to the cytoplasm
had been established, cells were held at a voltage (Vh) of
)70 mV and those with holding currents larger than 20 pA
were rejected. Series resistance was compensated by 80%
and monitored throughout the experiment, together with
the cell membrane capacitance.
Ligand-gated currents were activated by P2X receptor
agonists applied onto the cell under investigation by means
of a gravity-driven perfusion system with five independent
lines controlled by electronic valves (The Lee Company,
Westbrook, CO, USA). This system allowed the exchange
of the medium surrounding a cell in < 200 ms. Stock solu-
tions of drugs were diluted daily in extracellular saline and
incorporated into the perfusion system a few minutes before
starting the experiments. Current–voltage relations for
BzATP-evoked responses were obtained by stepping the
holding voltage to the indicated values, 2 s prior to and
during agonist application. Reversal potential values from
individual cells were corrected for the junction potential
between the pipette’s solution and the extracellular solution,
which was calculated to be +4.42 mV using the Patcher’s
Tools module included in igor-pro software (WaveMetrics,
Inc., Lake Oswego, OR, USA). All recordings were
obtained at room temperature (21–24 �C).
Immunocytochemistry
N2a cells cultured on coverslips placed in 35 mm dishes
(250 000 cells per well) were washed with NaCl ⁄Pi and fixed
with 4% PFA for 15 min. After washing with NaCl ⁄Pi, the
cells were permeabilized with 0.1% Triton X-100 and
blocked with 5% goat serum and 10% fetal bovine serum
for 1 h at room temperature. After washing with 3% BSA
in NaCl ⁄Pi, the cells were incubated for 1 h with primary
antibodies against P2X7 (1 : 100) and a-tubulin (1 : 1000).
Subsequently, cells were washed with NaCl ⁄Pi and incu-
bated for 1 h with Cy3�-conjugated donkey anti-rabbit
IgG (Jackson InmunoResearch, Westgrove, PA, USA) or
fluorescein isothiocyanate-goat anti-mouse IgG (Sigma).
Nuclei were counterstained with 4¢,6-diamidino-2-phenylin-
dole (DAPI, from Invitrogen), a fluorescent stain that binds
strongly to DNA. Coverslips were mounted on glass slides
using FluoroSave� Reagent (Calbiochem, Nottingham,
UK). Images were acquired using a Nikon Eclipse TE-200
microscope coupled to a CCD camera (Kappa ACC-1).
The analysis of neurite length and ramifications was carried
out using Neuron J freeware.
Statistical analysis
Results were analysed by unpaired t-test using graph pad
prism 5 (Graph Pad Software Inc., San Diego, CA, USA)
and expressed as the mean ± standard error of the mean.
Differences were considered to be significant at P £ 0.05.
R. Gomez-Villafuertes et al. P2X7 negatively controls neuritogenesis via CaMKII
FEBS Journal 276 (2009) 5307–5325 ª 2009 The Authors Journal compilation ª 2009 FEBS 5321

Acknowledgements
PHF-1 antibody was a generous gift from Dr Jesus
Avila (CBM, Madrid, Spain). This work was sup-
ported by research grants from MICINN (BFU2008-
02699, BFU2005-06034 and SAF2006-00906), CAM
(S-SAL-0253-2006), ‘The Spanish Ion Channel Initia-
tive (SICI)’ (CSD2008-00005) and Fundacion Marceli-
no Botın. RGV was supported by SICI. AP was
supported by CIBERNED. MDH and JIDH were sup-
ported by Juan de la Cierva and Ramon y Cajal
Programs, respectively. DB was a research fellow of
the Basque Government.
References
1 Burnstock G & Kennedy C (1985) Is there a basis for
distinguishing two types of P2-purinoceptor? Gen Phar-
macol 16, 433–440.
2 Khakh BS, Burnstock G, Kennedy C, King BF, North
RA, Seguela P, Voigt M & Humphrey PP (2001) Inter-
national union of pharmacology XXIV. Current status
of the nomenclature and properties of P2X receptors
and their subunits. Pharmacol Rev 53, 107–118.
3 North RA (2002) Molecular physiology of P2X recep-
tors. Physiol Rev 82, 1013–1067.
4 Von Kugelgen I & Wetter A (2000) Molecular pharma-
cology of P2Y-receptors. Naunyn Schmiedebergs Arch
Pharmacol 362, 310–323.
5 Abbracchio MP, Boeynaems JM, Barnard EA, Boyer
JL, Kennedy C, Miras-Portugal MT, King BF, Gachet
C, Jacobson KA, Weisman GA et al. (2003) Character-
ization of the UDP-glucose receptor (re-named here the
P2Y14 receptor) adds diversity to the P2Y receptor
family. Trends Pharmacol Sci 24, 52–55.
6 Nicke A, Baumert HG, Rettinger J, Eichele A, Lambr-
echt G, Mutschler E & Schmalzing G (1998) P2X1 and
P2X3 receptors form stable trimers: a novel structural
motif of ligand-gated ion channels. EMBO J 17, 3016–
3028.
7 Clyne JD, Wang LF & Hume RI (2002) Mutational
analysis of the conserved cysteines of the rat P2X2
purinoceptor. J Neurosci 22, 3873–3880.
8 Ennion SJ & Evans RJ (2002) Conserved cysteine resi-
dues in the extracellular loop of the human P2X(1)
receptor form disulfide bonds and are involved in
receptor trafficking to the cell surface. Mol Pharmacol
61, 303–311.
9 Franke H & Illes P (2006) Involvement of P2 receptors
in the growth and survival of neurons in the CNS.
Pharmacol Ther 109, 297–324.
10 Khakh BS & North RA (2006) P2X receptors as cell-
surface ATP sensors in health and disease. Nature 442,
527–532.
11 Evans RJ, Derkach V & Surprenant A (1992) ATP
mediates fast synaptic transmission in mammalian
neurons. Nature 357, 503–505.
12 Silinsky EM, Gerzanich V & Vanner SM (1992) ATP
mediates excitatory synaptic transmission in mammalian
neurones. Br J Pharmacol 106, 762–763.
13 Edwards FA, Gibb AJ & Colquhoun D (1992) ATP
receptor-mediated synaptic currents in the central
nervous system. Nature 359, 144–147.
14 Sperlagh B, Kofalvi A, Deuchars J, Atkinson L,
Milligan CJ, Buckley NJ & Vizi ES (2002) Involvement
of P2X7 receptors in the regulation of neurotransmitter
release in the rat hippocampus. J Neurochem 81,
1196–1211.
15 Khakh BS, Gittermann D, Cockayne DA & Jones A
(2003) ATP modulation of excitatory synapses onto
interneurons. J Neurosci 23, 7426–7437.
16 Nakatsuka T, Tsuzuki K, Ling JX, Sonobe H & Gu
JG (2003) Distinct roles of P2X receptors in modulat-
ing glutamate release at different primary sensory
synapses in rat spinal cord. J Neurophysiol 89, 3243–
3252.
17 Gualix J, Gomez-Villafuertes R, Diaz-Hernandez M &
Miras-Portugal MT (2003) Presence of functional ATP
and dinucleotide receptors in glutamatergic synaptic ter-
minals from rat midbrain. J Neurochem 87, 160–171.
18 Jo YH & Schlichter R (1999) Synaptic corelease of
ATP and GABA in cultured spinal neurons. Nat Neuro-
sci 2, 241–245.
19 Gomez-Villafuertes R, Gualix J & Miras-Portugal MT
(2001) Single GABAergic synaptic terminals from rat
midbrain exhibit functional P2X and dinucleotide recep-
tors, able to induce GABA secretion. J Neurochem 77,
84–93.
20 Jo YH & Role LW (2002) Coordinate release of ATP
and GABA at in vitro synapses of lateral hypothalamic
neurons. J Neurosci 22, 4794–4804.
21 Rhee JS, Wang ZM, Nabekura J, Inoue K & Akaike N
(2000) ATP facilitates spontaneous glycinergic IPSC fre-
quency at dissociated rat dorsal horn interneuron syn-
apses. J Physiol 524 (Pt 2), 471–483.
22 Wang ZM, Katsurabayashi S, Rhee JS, Brodwick M &
Akaike N (2001) Substance P abolishes the facilitatory
effect of ATP on spontaneous glycine release in neurons
of the trigeminal nucleus pars caudalis. J Neurosci 21,
2983–2991.
23 Jang IS, Rhee JS, Kubota H, Akaike N & Akaike N
(2001) Developmental changes in P2X purinoceptors
on glycinergic presynaptic nerve terminals projecting
to rat substantia gelatinosa neurones. J Physiol 536,
505–519.
24 Brandle U, Zenner HP & Ruppersberg JP (1999) Gene
expression of P2X-receptors in the developing inner ear
of the rat. Neurosci Lett 273, 105–108.
P2X7 negatively controls neuritogenesis via CaMKII R. Gomez-Villafuertes et al.
5322 FEBS Journal 276 (2009) 5307–5325 ª 2009 The Authors Journal compilation ª 2009 FEBS

25 Adrian K, Bernhard MK, Breitinger HG & Ogilvie A
(2000) Expression of purinergic receptors (ionotropic
P2X1–7 and metabotropic P2Y1–11) during myeloid
differentiation of HL60 cells. Biochim Biophys Acta
1492, 127–138.
26 Huang N, Wang DJ & Heppel LA (1989) Extracellular
ATP is a mitogen for 3T3, 3T6, and A431 cells and acts
synergistically with other growth factors. Proc Natl
Acad Sci USA 86, 7904–7908.
27 Heo JS & Han HJ (2006) ATP stimulates mouse embry-
onic stem cell proliferation via protein kinase C, phos-
phatidylinositol 3-kinase ⁄Akt, and mitogen-activated
protein kinase signaling pathways. Stem Cells 24, 2637–
2648.
28 Resende RR, Majumder P, Gomes KN, Britto LR &
Ulrich H (2007) P19 embryonal carcinoma cells as
in vitro model for studying purinergic receptor expres-
sion and modulation of N-methyl-D-aspartate-gluta-
mate and acetylcholine receptors during neuronal
differentiation. Neuroscience 146, 1169–1181.
29 Heilbronn A, Maienschein V, Carstensen K, Gann W &
Zimmermann H (1995) Crucial role of ecto-5¢-nucleotid-ase in differentiation and survival of developing neural
cells. Neuroreport 7, 257–261.
30 Gysbers JW & Rathbone MP (1996) Neurite outgrowth
in PC12 cells is enhanced by guanosine through both
cAMP-dependent and -independent mechanisms. Neuro-
sci Lett 220, 175–178.
31 Benowitz LI, Jing Y, Tabibiazar R, Jo SA, Petrausch
B, Stuermer CA, Rosenberg PA & Irwin N (1998) Axon
outgrowth is regulated by an intracellular purine-sensi-
tive mechanism in retinal ganglion cells. J Biol Chem
273, 29626–29634.
32 Hopker VH, Saffrey MJ & Burnstock G (1996) Neurite
outgrowth of striatal neurons in vitro: involvement of
purines in the growth-promoting effect of myenteric
plexus explants. Int J Dev Neurosci 14, 439–451.
33 Rathbone MP, Middlemiss PJ, Gysbers JW, Andrew C,
Herman MA, Reed JK, Ciccarelli R, Di Iorio P &
Caciagli F (1999) Trophic effects of purines in neurons
and glial cells. Prog Neurobiol 59, 663–690.
34 Heine C, Heimrich B, Vogt J, Wegner A, Illes P &
Franke H (2006) P2 receptor-stimulation influences axo-
nal outgrowth in the developing hippocampus in vitro.
Neuroscience 138, 303–311.
35 Diaz-Hernandez M, Del Puerto A, Diaz-Hernandez
JI, Diez-Zaera M, Lucas JJ, Garrido JJ & Miras-
Portugal MT (2008) Inhibition of the ATP-gated
P2X7 receptor promotes axonal growth and branching
in cultured hippocampal neurons. J Cell Sci 121,
3717–3728.
36 Raffaghello L, Chiozzi P, Falzoni S, Di Virgilio F &
Pistoia V (2006) The P2X7 receptor sustains the growth
of human neuroblastoma cells through a substance
P-dependent mechanism. Cancer Res 66, 907–914.
37 Wu PY, Lin YC, Chang CL, Lu HT, Chin CH, Hsu
TT, Chu D & Sun SH (2009) Functional decreases in
P2X7 receptors are associated with retinoic acid-induced
neuronal differentiation of Neuro-2a neuroblastoma
cells. Cell Signal 21, 881–891.
38 Vulchanova L, Riedl MS, Shuster SJ, Buell G,
Surprenant A, North RA & Elde R (1997) Immuno-
histochemical study of the P2X2 and P2X3 receptor
subunits in rat and monkey sensory neurons and their
central terminals. Neuropharmacology 36, 1229–1242.
39 Bo X, Kim M, Nori SL, Schoepfer R, Burnstock G &
North RA (2003) Tissue distribution of P2X4 receptors
studied with an ectodomain antibody. Cell Tissue Res
313, 159–165.
40 Roberts JA, Vial C, Digby HR, Agboh KC, Wen H,
Atterbury-Thomas A & Evans RJ (2006) Molecular
properties of P2X receptors. Pflugers Arch 452, 486–
500.
41 Soto F, Garcia-Guzman M, Gomez-Hernandez JM,
Hollmann M, Karschin C & Stuhmer W (1996) P2X4:
an ATP-activated ionotropic receptor cloned from rat
brain. Proc Natl Acad Sci USA 93, 3684–3688.
42 Cheewatrakoolpong B, Gilchrest H, Anthes JC &
Greenfeder S (2005) Identification and characterization
of splice variants of the human P2X7 ATP channel.
Biochem Biophys Res Commun 332, 17–27.
43 Lakshmi S & Joshi PG (2006) Activation of
Src ⁄ kinase ⁄phospholipase C ⁄mitogen-activated protein
kinase and induction of neurite expression by ATP,
independent of nerve growth factor. Neuroscience 141,
179–189.
44 Surprenant A, Rassendren F, Kawashima E, North RA
& Buell G (1996) The cytolytic P2Z receptor for
extracellular ATP identified as a P2X receptor (P2X7).
Science 272, 735–738.
45 Virginio C, Church D, North RA & Surprenant A
(1997) Effects of divalent cations, protons and calmi-
dazolium at the rat P2X7 receptor. Neuropharmacology
36, 1285–1294.
46 Sun SH, Lin LB, Hung AC & Kuo JS (1999) ATP-stim-
ulated Ca2+ influx and phospholipase D activities of a
rat brain-derived type-2 astrocyte cell line, RBA-2, are
mediated through P2X7 receptors. J Neurochem 73,
334–343.
47 Jiang LH, Mackenzie AB, North RA & Surprenant A
(2000) Brilliant blue G selectively blocks ATP-gated rat
P2X(7) receptors. Mol Pharmacol 58, 82–88.
48 Donnelly-Roberts DL & Jarvis MF (2007) Discovery of
P2X7 receptor-selective antagonists offers new insights
into P2X7 receptor function and indicates a role in
chronic pain states. Br J Pharmacol 151, 571–579.
49 Romagnoli R, Baraldi PG, Cruz-Lopez O, Lopez-Cara
C, Preti D, Borea PA & Gessi S (2008) The P2X7
receptor as a therapeutic target. Expert Opin Ther
Targets 12, 647–661.
R. Gomez-Villafuertes et al. P2X7 negatively controls neuritogenesis via CaMKII
FEBS Journal 276 (2009) 5307–5325 ª 2009 The Authors Journal compilation ª 2009 FEBS 5323

50 Fischer I, Shea TB, Sapirstein VS & Kosik KS (1986)
Expression and distribution of microtubule-associated
protein 2 (MAP2) in neuroblastoma and primary neuro-
nal cells. Brain Res 390, 99–109.
51 Berridge MJ (1998) Neuronal calcium signaling. Neuron
21, 13–26.
52 Gomez TM & Spitzer NC (1999) In vivo regulation of
axon extension and pathfinding by growth-cone calcium
transients. Nature 397, 350–355.
53 Saitoh T & Schwartz JH (1985) Phosphorylation-depen-
dent subcellular translocation of a Ca2+ ⁄ calmodulin-
dependent protein kinase produces an autonomous
enzyme in Aplysia neurons. J Cell Biol 100, 835–842.
54 Ahmed T & Frey JU (2005) Plasticity-specific phos-
phorylation of CaMKII, MAP-kinases and CREB
during late-LTP in rat hippocampal slices in vitro.
Neuropharmacology 49, 477–492.
55 Leon D, Hervas C & Miras-Portugal MT (2006) P2Y1
and P2X7 receptors induce calcium ⁄ calmodulin-depen-
dent protein kinase II phosphorylation in cerebellar
granule neurons. Eur J Neurosci 23, 2999–3013.
56 Robles E & Gomez TM (2006) Focal adhesion kinase
signaling at sites of integrin-mediated adhesion controls
axon pathfinding. Nat Neurosci 9, 1274–1283.
57 Fan RS, Jacamo RO, Jiang X, Sinnett-Smith J &
Rozengurt E (2005) G protein-coupled receptor activa-
tion rapidly stimulates focal adhesion kinase phosphor-
ylation at Ser-843. Mediation by Ca2+, calmodulin,
and Ca2+ ⁄ calmodulin-dependent kinase II. J Biol Chem
280, 24212–24220.
58 Chen HC, Appeddu PA, Isoda H & Guan JL (1996)
Phosphorylation of tyrosine 397 in focal adhesion
kinase is required for binding phosphatidylinositol
3-kinase. J Biol Chem 271, 26329–26334.
59 Xia H, Nho RS, Kahm J, Kleidon J & Henke CA
(2004) Focal adhesion kinase is upstream of phosphati-
dylinositol 3-kinase ⁄Akt in regulating fibroblast survival
in response to contraction of type I collagen matrices
via a beta 1 integrin viability signaling pathway. J Biol
Chem 279, 33024–33034.
60 Kimura K, Hattori S, Kabuyama Y, Shizawa Y, Taka-
yanagi J, Nakamura S, Toki S, Matsuda Y, Onodera K
& Fukui Y (1994) Neurite outgrowth of PC12 cells is
suppressed by wortmannin, a specific inhibitor of phos-
phatidylinositol 3-kinase. J Biol Chem 269, 18961–
18967.
61 Jackson TR, Blader IJ, Hammonds-Odie LP, Burga
CR, Cooke F, Hawkins PT, Wolf AG, Heldman KA &
Theibert AB (1996) Initiation and maintenance of
NGF-stimulated neurite outgrowth requires activation
of a phosphoinositide 3-kinase. J Cell Sci 109 (Pt 2),
289–300.
62 Yao R & Cooper GM (1995) Requirement for phospha-
tidylinositol-3 kinase in the prevention of apoptosis by
nerve growth factor. Science 267, 2003–2006.
63 Roche S, Downward J, Raynal P & Courtneidge SA
(1998) A function for phosphatidylinositol 3-kinase beta
(p85alpha-p110beta) in fibroblasts during mitogenesis:
requirement for insulin- and lysophosphatidic acid-med-
iated signal transduction. Mol Cell Biol 18, 7119–7129.
64 Yang H, Shaw G & Raizada MK (2002) ANG II stim-
ulation of neuritogenesis involves protein kinase B in
brain neurons. Am J Physiol Regul Integr Comp Physiol
283, R107–R114.
65 Munoz-Montano JR, Lim F, Moreno FJ, Avila J &
Diaz-Nido J (1999) Glycogen synthase kinase-3 modu-
lates neurite outgrowth in cultured neurons: possible
implications for neurite pathology in Alzheimer’s dis-
ease. J Alzheimers Dis 1, 361–378.
66 Sanchez S, Sayas CL, Lim F, Diaz-Nido J, Avila J &
Wandosell F (2001) The inhibition of phosphatidylinosi-
tol-3-kinase induces neurite retraction and activates
GSK3. J Neurochem 78, 468–481.
67 Sperber BR, Leight S, Goedert M & Lee VM (1995)
Glycogen synthase kinase-3 beta phosphorylates tau
protein at multiple sites in intact cells. Neurosci Lett
197, 149–153.
68 Roberts VH, Greenwood SL, Elliott AC, Sibley CP &
Waters LH (2006) Purinergic receptors in human pla-
centa: evidence for functionally active P2X4, P2X7,
P2Y2, and P2Y6. Am J Physiol Regul Integr Comp
Physiol 290, R1374–R1386.
69 Jarvis MF & Khakh BS (2009) ATP-gated P2X cation-
channels. Neuropharmacology 56, 208–215.
70 Evans RJ (1996) The molecular biology of P2X recep-
tors. J Auton Pharmacol 16, 309–310.
71 Pintor J, Peral A, Pelaez T, Martin S & Hoyle CH
(2003) Presence of diadenosine polyphosphates in the
aqueous humor: their effect on intraocular pressure.
J Pharmacol Exp Ther 304, 342–348.
72 Gualix J, Pintor J & Miras-Portugal MT (1999) Charac-
terization of nucleotide transport into rat brain synaptic
vesicles. J Neurochem 73, 1098–1104.
73 Pintor J, Diaz-Rey MA, Torres M & Miras-Portugal
MT (1992) Presence of diadenosine polyphosphates –
Ap4A and Ap5A – in rat brain synaptic terminals.
Ca2+ dependent release evoked by 4-aminopyridine
and veratridine. Neurosci Lett 136, 141–144.
74 Bahima L, Aleu J, Elias M, Martin-Satue M, Muhaisen
A, Blasi J, Marsal J & Solsona C (2006) Endogenous
hemichannels play a role in the release of ATP from
Xenopus oocytes. J Cell Physiol 206, 95–102.
75 Keith C, DiPaola M, Maxfield FR & Shelanski ML
(1983) Microinjection of Ca++-calmodulin causes a
localized depolymerization of microtubules. J Cell Biol
97, 1918–1924.
76 Goswami C, Schmidt H & Hucho F (2007) TRPV1 at
nerve endings regulates growth cone morphology and
movement through cytoskeleton reorganization. Febs J
274, 760–772.
P2X7 negatively controls neuritogenesis via CaMKII R. Gomez-Villafuertes et al.
5324 FEBS Journal 276 (2009) 5307–5325 ª 2009 The Authors Journal compilation ª 2009 FEBS

77 Kim M, Jiang LH, Wilson HL, North RA & Surpre-
nant A (2001) Proteomic and functional evidence for a
P2X7 receptor signalling complex. EMBO J 20, 6347–
6358.
78 Easley CA, Faison MO, Kirsch TL, Lee JA, Seward
ME & Tombes RM (2006) Laminin activates CaMK-II
to stabilize nascent embryonic axons. Brain Res 1092,
59–68.
79 Kim B, van GolenCM & Feldman EL (2004) Insulin-
like growth factor-I signaling in human neuroblastoma
cells. Oncogene 23, 130–141.
80 Sieg DJ, Hauck CR, Ilic D, Klingbeil CK, Schaefer E,
Damsky CH & Schlaepfer DD (2000) FAK integrates
growth-factor and integrin signals to promote cell
migration. Nat Cell Biol 2, 249–256.
81 Rico B, Beggs HE, Schahin-Reed D, Kimes N, Schmidt
A & Reichardt LF (2004) Control of axonal branching
and synapse formation by focal adhesion kinase. Nat
Neurosci 7, 1059–1069.
82 Conklin MW, Lin MS & Spitzer NC (2005) Local cal-
cium transients contribute to disappearance of pFAK,
focal complex removal and deadhesion of neuronal
growth cones and fibroblasts. Dev Biol 287, 201–212.
83 Zhou FQ, Zhou J, Dedhar S, Wu YH & Snider WD
(2004) NGF-induced axon growth is mediated by local-
ized inactivation of GSK-3beta and functions of the
microtubule plus end binding protein APC. Neuron 42,
897–912.
84 Garrido JJ, Simon D, Varea O & Wandosell F (2007)
GSK3 alpha and GSK3 beta are necessary for axon
formation. FEBS Lett 581, 1579–1586.
85 Chen G, Bower KA, Xu M, Ding M, Shi X, Ke ZJ &
Luo J (2009) Cyanidin-3-glucoside reverses ethanol-
induced inhibition of neurite outgrowth: role of glyco-
gen synthase kinase 3 beta. Neurotox Res 15, 321–331.
86 Grynkiewicz G, Poenie M & Tsien RY (1985) A new
generation of Ca2+ indicators with greatly improved
fluorescence properties. J Biol Chem 260, 3440–3450.
R. Gomez-Villafuertes et al. P2X7 negatively controls neuritogenesis via CaMKII
FEBS Journal 276 (2009) 5307–5325 ª 2009 The Authors Journal compilation ª 2009 FEBS 5325
Copyright © 2022 FDOKUMEN




















