
Functional interactions between P2X4 and P2X7 receptors from mouse salivary epithelia
Upload
independentCategory
view
1download
0
Ii
ACMa
b
c
d
e
f
g
ARRAA
KAMMPR
I
r
BHasW
m6f
(
0h
Immunobiology 218 (2013) 1104– 1112
Contents lists available at SciVerse ScienceDirect
Immunobiology
j o ur nal homep age: www.elsev ier .com/ locate / imbio
mplication of purinergic P2X7 receptor in M. tuberculosis infection and hostnteraction mechanisms: A mouse model study
ndré A. Santos Jr a,b, Valnês Rodrigues-Juniora,c, Rafael F. Zanind, Thiago J. Borgesd,ristina Bonorinod, Robson Coutinho-Silvag, Christina M. Takyiag, Diógenes S. Santosa,aria M. Campose,f, Fernanda B. Morroneb,e,∗
Instituto Nacional de Ciência e Tecnologia em Tuberculose (INCT-TB), Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre, RS, BrazilPrograma de Pós Graduac ão em Biologia Celular e Molecular, Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre, RS, BrazilPrograma de Pós Graduac ão em Medicina e Ciências da Saúde, Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre, RS, BrazilInstituto de Pesquisas Biomédicas, Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre, RS, BrazilInstituto de Toxicologia e Farmacologia, Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre, RS, BrazilFaculdade de Odontologia, Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre, RS, BrazilInstituto de Biofísica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro, Rio de Janeiro, RJ, Brazil
a r t i c l e i n f o
rticle history:eceived 29 March 2012eceived in revised form 6 February 2013ccepted 8 March 2013vailable online 16 March 2013
eywords:TPouseycobacterium tuberculosis
2X7 receptor
a b s t r a c t
In the present study, we analyzed the role of purinergic P2X7 receptor in Mycobacterium tuberculosis infec-tion and host interaction mechanisms in vitro and in vivo. For experimental procedures, a macrophagemurine cell line RAW 264.7, and male Swiss, wild-type C57BL/6 and P2X7 receptor knockout (P2X7R−/−)mice were used throughout this study. We have demonstrated that treatment of RAW 264.7 cells with ATP(3 and 5 mM) resulted in a statistically significant reduction of M. tuberculosis-colony-forming units. Thepurinergic P2X7 receptor expression was found significantly augmented in the lungs of mice infected withM. tuberculosis H37Rv. Infected wild-type mice showed a marked increase in the spleen weight, in compar-ison to non-infected animals. Furthermore, M. tuberculosis-infected P2X7R−/− mice showed an increaseof M. tuberculosis burden in lung tissue, when compared to infected wild-type mice. In P2X7R−/− spleens,we observed a significant decrease in the populations of Treg (CD4+Foxp3+), T cells (CD4+, CD8+CD25+ and
+ + + + +
AW 264.7 CD4 CD25 ), dendritic cells (CD11c ) and B220 cells. However, a significant increase in CD11b cells wasobserved in P2X7R−/− mice, when compared to wild-type animals. In the lungs, P2X7R−/− M. tuberculosis-infected mice exhibited pulmonary infiltrates containing an increase of Treg cells (CD4+Foxp3+), T cells(CD4+ and CD8+) and a decrease in the B220+ cells, when compared with wild-type M. tuberculosis-infectedmice. The findings observed in the present study provide novel evidence on the role of P2X7 receptors inculos
the pathogenesis of tuberntroduction
Tuberculosis (TB) is a worldwide public health priority andemains the leading cause of mortality due to a single bacterial
Abbreviations: APC, antigen-presenting cell; ATP, adenosine-5′-triphosphate;CG, bacille Calmette-Guerin; CFU, colony-forming unit; FBS, fetal bovine serum;IV, human immunodeficiency virus; MDR, multidrug-resistant; OADC, oleic acid,lbumin, dextrose, catalase; PBS, phosphate buffered saline; SDS, sodium dodecylulfate; TB, tuberculosis; TDR, totally drug-resistant; Treg, regulatory T cell; WHO,
orld Health Organization; XDR, extensively drug-resistant.∗ Corresponding author at: Faculty of Pharmacy/Institute of Toxicology and Phar-acology, Pontifícia Universidade Católica do Rio Grande do Sul, Avenida Ipiranga,
681 Partenon, 90619-900 Porto Alegre, RS, Brazil. Tel.: +55 51 3320 3512;ax: +55 51 3320 3612.
E-mail addresses: [email protected], [email protected]. Morrone).
171-2985/$ – see front matter © 2013 Elsevier GmbH. All rights reserved.ttp://dx.doi.org/10.1016/j.imbio.2013.03.003
is.© 2013 Elsevier GmbH. All rights reserved.
pathogen, Mycobacterium tuberculosis. The World Health Organi-zation (WHO) has estimated 8.8 million incident cases of TB, andapproximately 1.4 million people died of TB in 2010 (World HealthOrganization 2011). M. tuberculosis has been identified as the mostsuccessful pathogen of all times, because it remains silent and latentwithin the host, being able to escape from the immune defenses(Enserink 2001; Wickelgren 2000). By arresting phagosomal matu-ration (Malik et al. 2000; Pethe et al. 2004), it persists dormant untilthe host defenses are down, as seen in human immunodeficiencyvirus (HIV) infection. The growing rate of HIV–TB co-infection(Fatkenheuer et al. 1999; World Health Organization 2010, 2011),the emergence of multidrug-resistant (MDR), extensively drug-resistant (XDR), and more recently, totally drug-resistant (TDR)
strains of M. tuberculosis (World Health Organization 2010, 2011),increased the need for identifying different therapeutic options totreat TB, as well as to better understand the pathophysiology of thisinsidious disease.
obiolo
stdevT(2
F3sie2
o(aotiiTmpaswscteHe
ltiaaq2kmtmf
1fttrm(aaee
iaot
A.A. Santos Jr et al. / Immun
Extracellular purines, including ATP, display a series of roles ineveral physiological processes, such as vascular tonus, pain sensa-ion, neurotransmission, cell proliferation, differentiation, and celleath (Burnstock 2006). The purinergic receptors mediating theseffects are classified into P1 (A1, A2A, A2B, and A3), which are acti-ated by nucleosides, and P2 receptors that respond to nucleotides.hese receptors are further subdivided into metabotropic P2YP2Y1,2,4,6,11–14) and ionotropic P2X subtypes (P2X1–7) (Burnstock006).
The P2X7 receptor subtype presents a series of particularities.irst, its activation requires high concentrations of ATP (up to00 �M), while the other P2X receptors display a very high sen-itivity for ATP. Importantly, P2X7 receptor is highly expressed inmmune and inflammatory cells, throughout the central or periph-ral nervous systems (Burnstock 2011; Donnelly-Roberts and Jarvis007; Ferrari et al. 1997).
P2X7 receptor activation results in a rapid and reversiblepening of channels that are permeable to Na+, K+ and Ca2+
Donnelly-Roberts and Jarvis 2007). This receptor also serves as regulator of inflammation, and it is involved in the productionf pro-inflammatory cytokines such as TNF-� and IL-1 �, leadingo the induction of cyclooxygenase-2 (COX-2), metalloproteinases,nducible nitric oxide synthase (iNOS) and production of superox-de anion (Donnelly-Roberts and Jarvis 2007; Labasi et al. 2002).his receptor has been involved in the activation of peripheralacrophages and glia, neutrophil infiltration, and prostaglandin
roduction (Burnstock 2006; Di Virgilio 2007; Donnelly-Robertsnd Jarvis 2007; Goncalves et al. 2006; Labasi et al. 2002). Recenttudies have demonstrated the influence of P2X7 receptor in aide range of experimental models of disease, such as depres-
ion, epilepsy, Parkinson’s disease, arthritis, cancer, hemorrhagicystitis, and chronic pain, by using either selective P2X7 recep-or antagonists or knockout (KO) mice to P2X7 receptors (Bassot al. 2009; Chessell et al. 2005; Donnelly-Roberts and Jarvis 2007;onore et al. 2006; Li et al. 2006; Marcellino et al. 2010; Martinst al. 2012; Teixeira et al. 2010).
It was shown that absence of P2X7 receptor make epithelial cellsess apoptotic, suggesting that its activation might be involved inhe regulation of apoptosis (Goncalves et al. 2006). In this context,t has been proposed that prolonged exposure of P2X7 receptor togonists leads a formation of a cytolytic pore in the cell membrane,llowing the entry of larger particles up to 900 Da, and conse-uent cell death (Burnstock 2006; Donnelly-Roberts and Jarvis007). Noteworthy, ATP was found to induce both cell death andilling of intracellular mycobacteria within BCG-infected humanacrophages (Lammas et al. 1997; Molloy et al. 1994). The cyto-
oxic and mycobactericidal effects of ATP have been shown to beediated by the specific interaction of ATP with macrophage sur-
ace P2X7 receptors (Lammas et al. 1997).Recent studies have demonstrated that the polymorphism
513A→C of P2X7 receptor gene causes an amino acid changerom glutamic acid at amino acid position 496 to an alanine inhe C-terminus (Gu et al. 2001), impairing multiple P2X7 recep-or functions, including cation fluxes in a variety of cells, theelease of IL-1�, IL-18, and matrix metalloproteinase (MMP)-9 fromacrophages, or shedding of CD23 and CD62L from lymphocytes
Gu et al. 2001; Saunders et al. 2003; Sluyter et al. 2004a,b; Sluyternd Wiley 2002). This polymorphism is associated with impairedbility to kill Mycobacterium bovis BCG via ATP in vitro (Fernandot al. 2005; Saunders et al. 2003), and increased susceptibility toxtra pulmonary TB (Fernando et al. 2007).
In the present study, we analyzed the mycobactericidal activ-
ty of ATP in M. tuberculosis-infected murine macrophages, as wells the in vivo expression of P2X7 receptors in a murine modelf M. tuberculosis-infection. We have also assessed, for the firstime, the effects of P2X7 receptor gene deletion on splenomegalygy 218 (2013) 1104– 1112 1105
and colony-forming unit (CFU) counts in lungs and spleens of M.tuberculosis-infected mice. Attempts have also been made to eval-uate the immune profile of P2X7 receptor KO mice infected with thelaboratorial M. tuberculosis H37Rv strain, by means of an extensiveflow cytometry analysis.
Materials and methods
Murine cell line culture and macrophage infection
The macrophage murine cell line RAW 264.7 (obtained fromBanco de Células do Rio de Janeiro – BCRJ) was cultured in RPMI-1640 (Gibco) supplemented with 10% heat inactivated fetal bovineserum (FBS) and 1% penicillin–streptomycin at 37 ◦C with 5% CO2.For infection of macrophages, we used the method previouslydescribed by Mascarello et al. (2010) with some modifications.Briefly, M. tuberculosis H37Rv strain was grown in Middlebrook7H9 (Difco) containing 10% oleic acid/albumin/dextrose/catalase(OADC) enrichment at 37 ◦C. RAW 264.7 cells were seeded in 12-well tissue culture plates (Corning) at 1 × 105 cells per well inRPMI-1640 medium (supplemented with 10% FBS) for 24 h at 37 ◦Cwith 5% CO2. The cells were then washed three times with RPMI-1640 to remove non-adherent cells. Infection of RAW 264.7 cellswas performed at a multiplicity of infection (MOI) of 10:1 (bacte-ria/macrophage) for 3 h at 37 ◦C with 5% CO2. Infected RAW cellswere washed three times with RPMI-1640 to remove extracellularbacteria and replaced with 1 mL fresh RPMI (supplemented with10% FBS), with ATP in concentrations of 3 mM and 5 mM. This wasdefined as time 0 h. At 2 h and 20 h the infected macrophages werelysed with 0.025% SDS, and the number of viable bacteria was deter-mined by plating serial dilutions of each well on Middlebrook 7H10Agar supplement with 10% OADC. The plates were incubated at37 ◦C for 21 days before counting procedures. All experiments wereperformed in triplicates and the results are expressed in CFU perwell.
Determination of minimum inhibitory concentration (MIC) of ATPagainst M. tuberculosis H37Rv strain
MIC values for isoniazid (control drug) and ATP (test) weredetermined by resazurin microtiter assay (REMA) (Palomino et al.2002). Isoniazid and ATP growth inhibition activities were testedagainst the M. tuberculosis H37Rv strain. Mycobacteria was culti-vated in Middlebrook 7H9 (Difco) liquid medium supplementedwith 10% (v/v) OADC (Becton Dickinson) and 0.05% (m/v) Tween 80(Sigma). Culture was initially grown at 37 ◦C up to an optical densityat 600 nm (OD600) of 0.4. It was then diluted in Middlebrook 7H9to achieve an OD600 of 0.006, and 100 �L of M. tuberculosis inocu-lum was added to each well on the microplate, containing 100 �Lof compound or Middlebrook 7H9 (control inoculum) reaching anOD600 of 0.003. Final drug concentration ranged from 0.0078 to4 �g mL−1 for isoniazid and 0.019 to 10 mM for ATP containing 2%of dimethyl sulfoxide (DMSO). Plates were incubated for 7 days at37 ◦C. Sixty microliters of 0.01% resazurin (Sigma Chem. Co.) solu-tion was added to each well; plates were re-incubated for additional2 days. A change in color from blue to pink indicated the growthof bacteria, and the MIC was read as the minimum drug concen-tration that prevented the color change in resazurin solution. Theexperiment was carried out in duplicate.
Animals
Male Swiss, wild-type C57BL/6 (wild-type) and P2X7 receptorKO (P2X7R−/−) mice (25–30 g) were used throughout this study.Swiss and C57BL/6 mice were obtained from Universidade Federalde Pelotas (UFPEL; Pelotas, RS, Brazil), and P2X7R−/− mice were

1 obiolo
ddeICailA
E
m(rcii2osAiufl
I
b(iiswcpqtslabAbisstpRst1anblS
C
wb
106 A.A. Santos Jr et al. / Immun
onated by Dr Robson Coutinho-Silva, Federal University of Rioe Janeiro (UFRJ, Rio de Janeiro, Brazil). The KO mice were gen-rated by the method developed by Dr James Mobley (PGRD, Pfizernc, Groton, CT, USA). The KO mice used in the present study were57BL/6 inbred. The animals were housed in groups of six per cagend maintained in controlled temperature (22 ± 2 ◦C) and humid-ty (60–70%), under a 12 h light–dark cycle, with food and water adibitum. All the experimental protocols were approved by the Localnimal Ethics Committee (CEUA 10/00203-PUCRS).
xperimental infection in mice
Mice were anaesthetized by intraperitoneal (i.p.) injection of aixture containing ketamine (100 mg/kg) and xilazine (10 mg/kg)
Cristália, Itapira, SP, Brazil and Vetbrands, Jacareí, SP, Brazil,espectively), and subsequently infected with ∼107 M. tuber-ulosis H37Rv strain suspended in 0.2 mL of saline solutionntravenously (i.v.), through retro-orbital venous sinus, accord-ng to the technique described previously (Rodrigues-Junior et al.012). Non-infected control group received saline by i.v. retro-rbital route. The efficiency of infection was confirmed by acid-fasttaining of lung slices using the specific Ziehl-Neelsen staining.fter 4 weeks, the animals were euthanized under deep isoflurane
nhalation. The lungs and spleens were aseptically removed andsed to perform immunohistochemistry analysis, CFU counts andow cytometry analysis.
mmunohistochemistry
The expression of P2X7 receptors in lungs was assessedy immunostaining, according to method previously describedGoncalves et al. 2006), with some modifications. For these exper-ments, the lungs were collected 4 weeks post M. tuberculosisnfection and fixed in buffered neutral formalin. Paraffin-embeddedections (3 �m) were cut onto glass slides. The sections were de-axed in xylene three times for 15 min, rehydrated in decreasing
oncentrations of ethanol for 5 min and washed three times inhosphate-buffered saline for 10 min. Endogenous peroxidase wasuenched for 30 min with 0.3% hydrogen peroxide:methanol solu-ion (v/v). After washing in distilled water and phosphate-bufferedaline, a blocking step was included using either, extract of mouseiver or 5% bovine serum albumin in phosphate-buffered saline for
total of 30 min. Rabbit anti-P2X7 receptor-purified polyclonal anti-ody (1:200 in PBS; Alomone, Jerusalem, Israel, catalog numberPR-004) was incubated overnight at 4 ◦C in a humidified cham-er. Negative controls were carried out with bovine serum albumin
nstead of primary antibody. After achieving room temperature,ections were washed in 0.25% phosphate-buffered saline–tweenolution for 5 min. Universal immuno-peroxidase polymer for mouseissue sections anti-rabbit primary antibody (N-Histofine – Sim-le Stain Mouse MAX PO code 414341F), was incubated for 2 h.eactions were revealed using diaminobenzidine chromogen sub-trate (Liquid DAB, DAKO cat number K3466, USA). After washinghe slides, counterstaining was done with Harris hematoxylin for–2 min. The images were captured by Zeiss AxioImager M2, andnalyzed through the Image-Pro Plus 4.0 software (Media Cyber-etics L.P). The lung P2X7 receptor surface density was determinedy considering the percentage of stained areas in the total histo-
ogical fields. For this experimental set, we have used 8 wild-typewiss mice per group.
FU determination protocol
For evaluation of bacterial load, the right lungs and spleensere aseptically removed and homogenized in 3 ml of phosphate-
uffered saline (PBS). For each organ, the suspension was serially
gy 218 (2013) 1104– 1112
diluted 10-fold, and 0.1 ml of each appropriate dilution wascultured in duplicate by plating onto 7H10 Middlebrook agar sup-plemented with 10% OADC. The colonies were counted after 28 daysof incubation at 37 ◦C.
Spleen weight determination
The spleens were dissected free from connecting tissues. Theweight of each spleen was determined, and the result wasexpressed as milligram per gram of animal.
Flow cytometry analysis
After euthanasia, the spleens and lungs were excised and dis-rupted against a nylon screen in media containing CollagenaseD (Roche). Single cell suspensions were obtained, and the cellswere counted with Trypan blue. Subsequently, the cells werestained with antibodies against CD4-PECy5, CD25- PE, Foxp3-Alexa647, CD8a-FITC, Gr1-PE, CD11b-APC, CD11c-PECy7 e B220- FITC(BD Biosciences). We investigated the following population sub-sets of immune cells: CD4+, CD8+, CD4+Foxp3+, CD4+CD25+Foxp3+,CD4+CD25+, CD8+CD25+, CD11b+, CD11c+ and B220+. Cells wereanalyzed in a FACSCanto II flow cytometer (BD Biosciences). Datawas corrected using the correspondent (non-infected) controlgroup and expressed in fold variations between P2X7R−/− and wild-type mice.
Statistical analysis
To compare the viable M. tuberculosis cell counts recovered frominfected macrophages, or spleens and lungs of mice, the num-bers were firstly converted into logarithms of CFU (Log10 CFU).Data from spleen and lung CFU, and flow cytometry analysiswere evaluated using Student’s unpaired t-test. The spleen weightwas previously corrected with the body weight from each ani-mal (spleen weight/body weight). Data of spleen weight andmacrophages CFU, was evaluated by a one-way analysis of variance(ANOVA), followed by Dunnett’s post-test, using GraphPad Prism5.0 (GraphPad, San Diego, CA, USA). Differences were consideredsignificant at the 95% level of confidence.
Results
Effects of ATP on macrophage infection
In this study, we initially investigated whether the activation ofP2X7 receptors is related to colony formation in the macrophagemurine cell line RAW 264.7 infected with M. tuberculosis. Theincubation of M. tuberculosis-infected RAW 264.7 cells with highconcentrations of ATP (3 and 5 mM), which is known to selec-tively activate P2X7 receptors, resulted in a statistically significantreduction of CFU counts by 2.211 Log10 and 2.205 Log10 unitsrespectively, at 2 h (P < 0.001) and, by 0.832 Log10 and 1.084 Log10units, respectively, at 20 h post-treatment (P < 0.05) (Fig. 1).
Determination of minimum inhibitory concentration (MIC) of ATPagainst M. tuberculosis H37Rv strain
To investigate if ATP has a bactericidal effect per se or if theATP solution used in the experiments might be contaminated withother bactericidal chemicals, we tested the direct effect of ATP on
M. tuberculosis H37Rv strain, through determination of MIC. Theresults showed MIC values of 0.5 �g mL−1 and higher than 10 mMfor isoniazid and ATP, respectively, against M. tuberculosis H37Rvstrain (data not shown).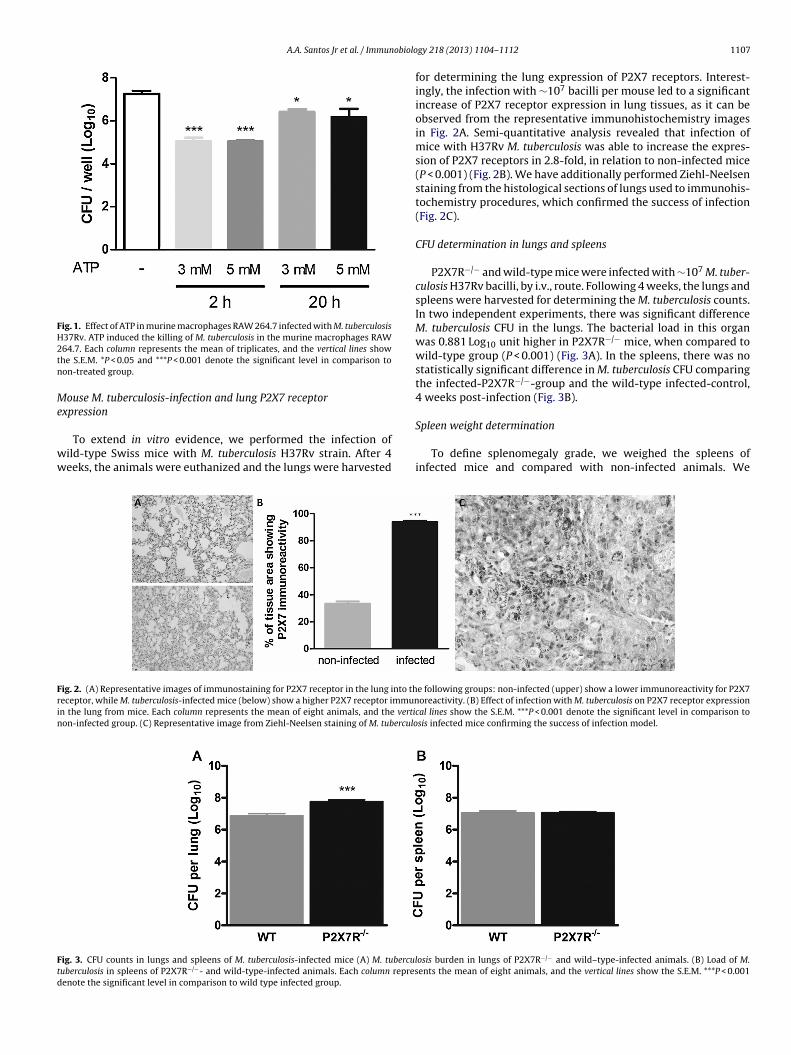
A.A. Santos Jr et al. / Immunobiolo
Fig. 1. Effect of ATP in murine macrophages RAW 264.7 infected with M. tuberculosisH37Rv. ATP induced the killing of M. tuberculosis in the murine macrophages RAW2tn
Me
ww
Frin
Ftd
64.7. Each column represents the mean of triplicates, and the vertical lines showhe S.E.M. *P < 0.05 and ***P < 0.001 denote the significant level in comparison toon-treated group.
ouse M. tuberculosis-infection and lung P2X7 receptorxpression
To extend in vitro evidence, we performed the infection ofild-type Swiss mice with M. tuberculosis H37Rv strain. After 4eeks, the animals were euthanized and the lungs were harvested
ig. 2. (A) Representative images of immunostaining for P2X7 receptor in the lung into theceptor, while M. tuberculosis-infected mice (below) show a higher P2X7 receptor immunn the lung from mice. Each column represents the mean of eight animals, and the vertion-infected group. (C) Representative image from Ziehl-Neelsen staining of M. tuberculo
ig. 3. CFU counts in lungs and spleens of M. tuberculosis-infected mice (A) M. tuberculuberculosis in spleens of P2X7R−/−- and wild-type-infected animals. Each column represenote the significant level in comparison to wild type infected group.
gy 218 (2013) 1104– 1112 1107
for determining the lung expression of P2X7 receptors. Interest-ingly, the infection with ∼107 bacilli per mouse led to a significantincrease of P2X7 receptor expression in lung tissues, as it can beobserved from the representative immunohistochemistry imagesin Fig. 2A. Semi-quantitative analysis revealed that infection ofmice with H37Rv M. tuberculosis was able to increase the expres-sion of P2X7 receptors in 2.8-fold, in relation to non-infected mice(P < 0.001) (Fig. 2B). We have additionally performed Ziehl-Neelsenstaining from the histological sections of lungs used to immunohis-tochemistry procedures, which confirmed the success of infection(Fig. 2C).
CFU determination in lungs and spleens
P2X7R−/− and wild-type mice were infected with ∼107 M. tuber-culosis H37Rv bacilli, by i.v., route. Following 4 weeks, the lungs andspleens were harvested for determining the M. tuberculosis counts.In two independent experiments, there was significant differenceM. tuberculosis CFU in the lungs. The bacterial load in this organwas 0.881 Log10 unit higher in P2X7R−/− mice, when compared towild-type group (P < 0.001) (Fig. 3A). In the spleens, there was nostatistically significant difference in M. tuberculosis CFU comparingthe infected-P2X7R−/−-group and the wild-type infected-control,4 weeks post-infection (Fig. 3B).
Spleen weight determination
To define splenomegaly grade, we weighed the spleens ofinfected mice and compared with non-infected animals. We
e following groups: non-infected (upper) show a lower immunoreactivity for P2X7oreactivity. (B) Effect of infection with M. tuberculosis on P2X7 receptor expression
cal lines show the S.E.M. ***P < 0.001 denote the significant level in comparison tosis infected mice confirming the success of infection model.
osis burden in lungs of P2X7R−/− and wild–type-infected animals. (B) Load of M.ents the mean of eight animals, and the vertical lines show the S.E.M. ***P < 0.001
1108 A.A. Santos Jr et al. / Immunobiolo
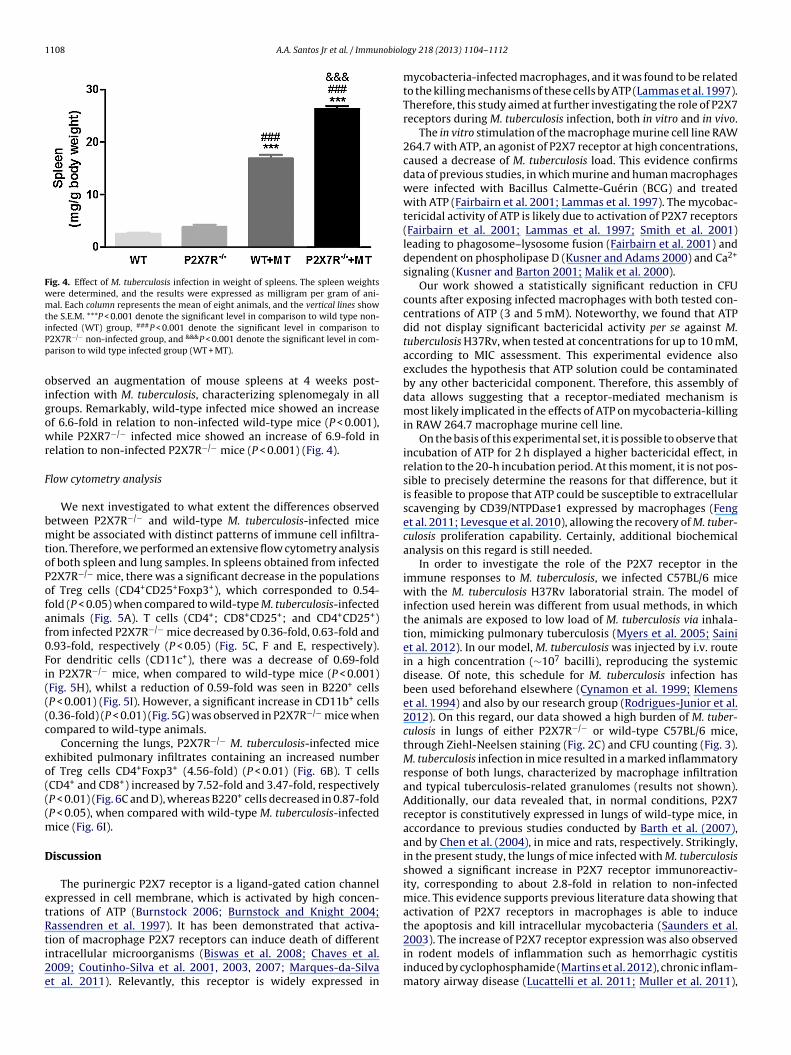
Fig. 4. Effect of M. tuberculosis infection in weight of spleens. The spleen weightswere determined, and the results were expressed as milligram per gram of ani-mal. Each column represents the mean of eight animals, and the vertical lines showthe S.E.M. ***P < 0.001 denote the significant level in comparison to wild type non-iPp
oigowr
F
bmtoPofaf0Fi(((c
eo(((m
D
etRti2e
nfected (WT) group, ###P < 0.001 denote the significant level in comparison to2X7R−/− non-infected group, and &&&P < 0.001 denote the significant level in com-arison to wild type infected group (WT + MT).
bserved an augmentation of mouse spleens at 4 weeks post-nfection with M. tuberculosis, characterizing splenomegaly in allroups. Remarkably, wild-type infected mice showed an increasef 6.6-fold in relation to non-infected wild-type mice (P < 0.001),hile P2XR7−/− infected mice showed an increase of 6.9-fold in
elation to non-infected P2X7R−/− mice (P < 0.001) (Fig. 4).
low cytometry analysis
We next investigated to what extent the differences observedetween P2X7R−/− and wild-type M. tuberculosis-infected miceight be associated with distinct patterns of immune cell infiltra-
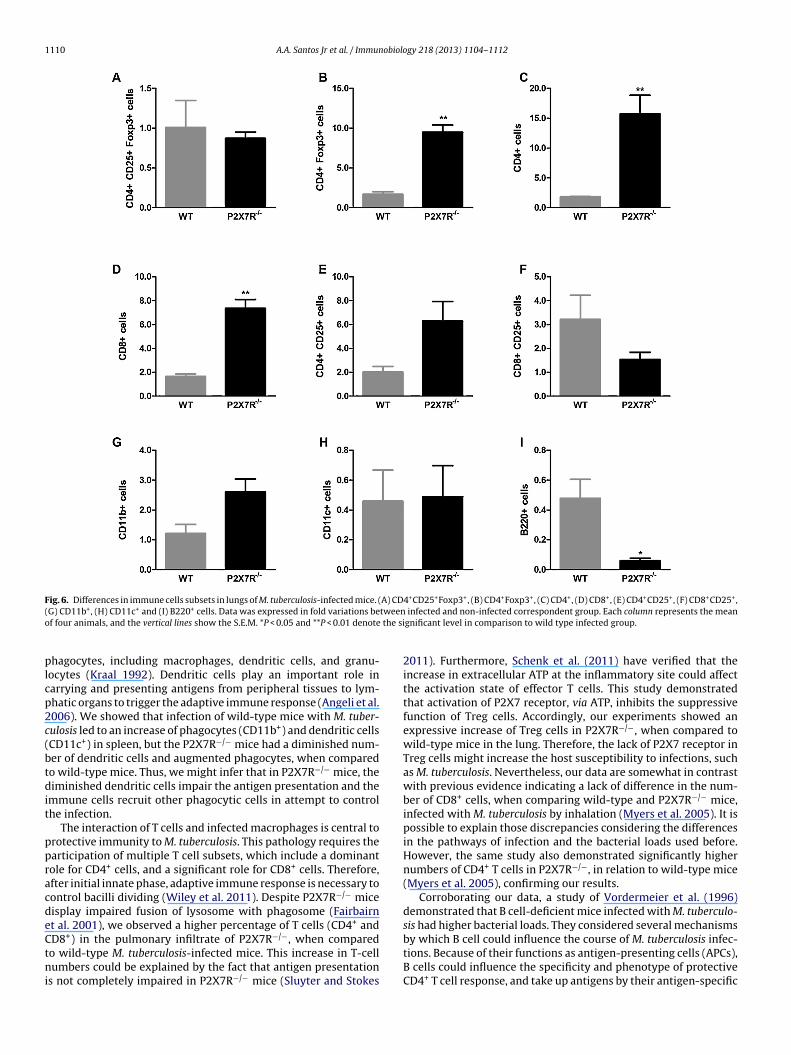
ion. Therefore, we performed an extensive flow cytometry analysisf both spleen and lung samples. In spleens obtained from infected2X7R−/− mice, there was a significant decrease in the populationsf Treg cells (CD4+CD25+Foxp3+), which corresponded to 0.54-old (P < 0.05) when compared to wild-type M. tuberculosis-infectednimals (Fig. 5A). T cells (CD4+; CD8+CD25+; and CD4+CD25+)rom infected P2X7R−/− mice decreased by 0.36-fold, 0.63-fold and.93-fold, respectively (P < 0.05) (Fig. 5C, F and E, respectively).or dendritic cells (CD11c+), there was a decrease of 0.69-foldn P2X7R−/− mice, when compared to wild-type mice (P < 0.001)Fig. 5H), whilst a reduction of 0.59-fold was seen in B220+ cellsP < 0.001) (Fig. 5I). However, a significant increase in CD11b+ cells0.36-fold) (P < 0.01) (Fig. 5G) was observed in P2X7R−/− mice whenompared to wild-type animals.
Concerning the lungs, P2X7R−/− M. tuberculosis-infected micexhibited pulmonary infiltrates containing an increased numberf Treg cells CD4+Foxp3+ (4.56-fold) (P < 0.01) (Fig. 6B). T cellsCD4+ and CD8+) increased by 7.52-fold and 3.47-fold, respectivelyP < 0.01) (Fig. 6C and D), whereas B220+ cells decreased in 0.87-foldP < 0.05), when compared with wild-type M. tuberculosis-infected
ice (Fig. 6I).
iscussion
The purinergic P2X7 receptor is a ligand-gated cation channelxpressed in cell membrane, which is activated by high concen-rations of ATP (Burnstock 2006; Burnstock and Knight 2004;assendren et al. 1997). It has been demonstrated that activa-
ion of macrophage P2X7 receptors can induce death of differentntracellular microorganisms (Biswas et al. 2008; Chaves et al.009; Coutinho-Silva et al. 2001, 2003, 2007; Marques-da-Silvat al. 2011). Relevantly, this receptor is widely expressed ingy 218 (2013) 1104– 1112
mycobacteria-infected macrophages, and it was found to be relatedto the killing mechanisms of these cells by ATP (Lammas et al. 1997).Therefore, this study aimed at further investigating the role of P2X7receptors during M. tuberculosis infection, both in vitro and in vivo.
The in vitro stimulation of the macrophage murine cell line RAW264.7 with ATP, an agonist of P2X7 receptor at high concentrations,caused a decrease of M. tuberculosis load. This evidence confirmsdata of previous studies, in which murine and human macrophageswere infected with Bacillus Calmette-Guérin (BCG) and treatedwith ATP (Fairbairn et al. 2001; Lammas et al. 1997). The mycobac-tericidal activity of ATP is likely due to activation of P2X7 receptors(Fairbairn et al. 2001; Lammas et al. 1997; Smith et al. 2001)leading to phagosome–lysosome fusion (Fairbairn et al. 2001) anddependent on phospholipase D (Kusner and Adams 2000) and Ca2+
signaling (Kusner and Barton 2001; Malik et al. 2000).Our work showed a statistically significant reduction in CFU
counts after exposing infected macrophages with both tested con-centrations of ATP (3 and 5 mM). Noteworthy, we found that ATPdid not display significant bactericidal activity per se against M.tuberculosis H37Rv, when tested at concentrations for up to 10 mM,according to MIC assessment. This experimental evidence alsoexcludes the hypothesis that ATP solution could be contaminatedby any other bactericidal component. Therefore, this assembly ofdata allows suggesting that a receptor-mediated mechanism ismost likely implicated in the effects of ATP on mycobacteria-killingin RAW 264.7 macrophage murine cell line.
On the basis of this experimental set, it is possible to observe thatincubation of ATP for 2 h displayed a higher bactericidal effect, inrelation to the 20-h incubation period. At this moment, it is not pos-sible to precisely determine the reasons for that difference, but itis feasible to propose that ATP could be susceptible to extracellularscavenging by CD39/NTPDase1 expressed by macrophages (Fenget al. 2011; Levesque et al. 2010), allowing the recovery of M. tuber-culosis proliferation capability. Certainly, additional biochemicalanalysis on this regard is still needed.
In order to investigate the role of the P2X7 receptor in theimmune responses to M. tuberculosis, we infected C57BL/6 micewith the M. tuberculosis H37Rv laboratorial strain. The model ofinfection used herein was different from usual methods, in whichthe animals are exposed to low load of M. tuberculosis via inhala-tion, mimicking pulmonary tuberculosis (Myers et al. 2005; Sainiet al. 2012). In our model, M. tuberculosis was injected by i.v. routein a high concentration (∼107 bacilli), reproducing the systemicdisease. Of note, this schedule for M. tuberculosis infection hasbeen used beforehand elsewhere (Cynamon et al. 1999; Klemenset al. 1994) and also by our research group (Rodrigues-Junior et al.2012). On this regard, our data showed a high burden of M. tuber-culosis in lungs of either P2X7R−/− or wild-type C57BL/6 mice,through Ziehl-Neelsen staining (Fig. 2C) and CFU counting (Fig. 3).M. tuberculosis infection in mice resulted in a marked inflammatoryresponse of both lungs, characterized by macrophage infiltrationand typical tuberculosis-related granulomes (results not shown).Additionally, our data revealed that, in normal conditions, P2X7receptor is constitutively expressed in lungs of wild-type mice, inaccordance to previous studies conducted by Barth et al. (2007),and by Chen et al. (2004), in mice and rats, respectively. Strikingly,in the present study, the lungs of mice infected with M. tuberculosisshowed a significant increase in P2X7 receptor immunoreactiv-ity, corresponding to about 2.8-fold in relation to non-infectedmice. This evidence supports previous literature data showing thatactivation of P2X7 receptors in macrophages is able to inducethe apoptosis and kill intracellular mycobacteria (Saunders et al.
2003). The increase of P2X7 receptor expression was also observedin rodent models of inflammation such as hemorrhagic cystitisinduced by cyclophosphamide (Martins et al. 2012), chronic inflam-matory airway disease (Lucattelli et al. 2011; Muller et al. 2011),
A.A. Santos Jr et al. / Immunobiology 218 (2013) 1104– 1112 1109
Fig. 5. Differences in immune cells subsets in spleen of M. tuberculosis-infected mice. (A) CD4+CD25+Foxp3+, (B) CD4+Foxp3+, (C) CD4+, (D) CD8+, (E) CD4+CD25+, (F)C fold vr **P < 0g
a2isMiiattP
ePPlPTri2a(
i
D8+CD25+, (G) CD11b+, (H) CD11c+ and (I) B220+ cells. Data was expressed in
epresents the mean of four animals, and the vertical lines show the S.E.M. *P < 0.05,
roup.
nd lipopolysaccharide-induced lung injury (Moncao-Ribeiro et al.011). Of high interest, the increase of P2X7 receptor immunopos-
tivity was localized especially around the granuloma region,trengthening the possible relevance of this receptor subtype for. tuberculosis infection mechanisms. Therefore, we might well
nfer that in the presence of M. tuberculosis, the immune cellsn lungs increase the membrane expression of P2X7 receptor inn attempt to control the infection spread. Nonetheless, addi-ional studies investigating the effects of ATP incubation on M.uberculosis-infected cultured primary macrophages obtained from2X7−/− mice remain to be performed in a near future.
In the present study, we showed the results of two independentxperiments, in which the burden of M. tuberculosis in the lungs of2X7R−/− were statistically different from the control group. The2X7R−/− mice showed an increase of M. tuberculosis load in theungs compared to infected wild-type mice. This data suggests that2X7 receptor has an important role in the control of pulmonaryB, corroborating the studies in which genetic variation in P2X7eceptor is associated with significant differences in the capac-ty of human macrophages to kill mycobacteria (Fernando et al.005), and also that P2X7 receptor gene polymorphisms in patients
re related to increased susceptibility for M. tuberculosis infectionAzad et al. 2012; Nino-Moreno et al. 2007; Singla et al. 2012).Notably, our results demonstrate that M. tuberculosisnfection induces severe splenomegaly. Of high interest,
ariations between infected and non-infected correspondent group. Each column.01 and ***P < 0.001 denote the significant level in comparison to wild type infected
M. tuberculosis-infected P2X7R−/− mice showed a significantincrease in the spleen weight when compared to infected wild-type mice. Martins de Sousa et al. (2010) previously demonstratedsplenomegaly in mice infected with Mycobacterium massiliense.Their results point out evidence indicating that splenomegalyin infected animals could be due to hemolysis or inflammation.Besides, a previous study showed that mycobacteria infectionexhibits an increase in the total number of cells in the liver andspleen, as a result of numerous focal granulomatous lesions and animmune inflammatory (Martins de Sousa et al. 2010). Of note, ithas been formerly shown that spleens from P2X2/P2X3 KO animalswere larger and weighed significantly more than the correspond-ing wild-type spleens (Coutinho-Silva et al. 2005). On the otherhand, our results did not show significant differences betweenthe spleen weights of wild-type and P2X7R−/− non-infected mice.Nevertheless, in our set of experiments the enlarged spleens of theM. tuberculosis-infected P2X7R−/− mice may be due to the chronicinfection and inflammation found in these animals. Prominently,flow cytometry analysis of P2X7R−/− spleens showed a significantdecrease in the populations of Treg (CD4+CD25+Foxp3+), T cells(CD4+, CD8+CD25+ and CD4+CD25+), dendritic cells (CD11c+) and
B220+ cells, when compared to wild-type animals.The spleen is a highly structured lymphoid organ (Mebiusand Kraal 2005). As microorganisms enter the marginal sinus ofthe spleen, they approach in contact with a variety of spleen
1110 A.A. Santos Jr et al. / Immunobiology 218 (2013) 1104– 1112
Fig. 6. Differences in immune cells subsets in lungs of M. tuberculosis-infected mice. (A) CD4+CD25+Foxp3+, (B) CD4+Foxp3+, (C) CD4+, (D) CD8+, (E) CD4+CD25+, (F) CD8+CD25+,( tweeno the si
plcp2c(btdit
ppracdeCtni
G) CD11b+, (H) CD11c+ and (I) B220+ cells. Data was expressed in fold variations bef four animals, and the vertical lines show the S.E.M. *P < 0.05 and **P < 0.01 denote
hagocytes, including macrophages, dendritic cells, and granu-ocytes (Kraal 1992). Dendritic cells play an important role inarrying and presenting antigens from peripheral tissues to lym-hatic organs to trigger the adaptive immune response (Angeli et al.006). We showed that infection of wild-type mice with M. tuber-ulosis led to an increase of phagocytes (CD11b+) and dendritic cellsCD11c+) in spleen, but the P2X7R−/− mice had a diminished num-er of dendritic cells and augmented phagocytes, when comparedo wild-type mice. Thus, we might infer that in P2X7R−/− mice, theiminished dendritic cells impair the antigen presentation and the
mmune cells recruit other phagocytic cells in attempt to controlhe infection.
The interaction of T cells and infected macrophages is central torotective immunity to M. tuberculosis. This pathology requires thearticipation of multiple T cell subsets, which include a dominantole for CD4+ cells, and a significant role for CD8+ cells. Therefore,fter initial innate phase, adaptive immune response is necessary toontrol bacilli dividing (Wiley et al. 2011). Despite P2X7R−/− miceisplay impaired fusion of lysosome with phagosome (Fairbairnt al. 2001), we observed a higher percentage of T cells (CD4+ and
D8+) in the pulmonary infiltrate of P2X7R−/−, when comparedo wild-type M. tuberculosis-infected mice. This increase in T-cellumbers could be explained by the fact that antigen presentations not completely impaired in P2X7R−/− mice (Sluyter and Stokes
infected and non-infected correspondent group. Each column represents the meangnificant level in comparison to wild type infected group.
2011). Furthermore, Schenk et al. (2011) have verified that theincrease in extracellular ATP at the inflammatory site could affectthe activation state of effector T cells. This study demonstratedthat activation of P2X7 receptor, via ATP, inhibits the suppressivefunction of Treg cells. Accordingly, our experiments showed anexpressive increase of Treg cells in P2X7R−/−, when compared towild-type mice in the lung. Therefore, the lack of P2X7 receptor inTreg cells might increase the host susceptibility to infections, suchas M. tuberculosis. Nevertheless, our data are somewhat in contrastwith previous evidence indicating a lack of difference in the num-ber of CD8+ cells, when comparing wild-type and P2X7R−/− mice,infected with M. tuberculosis by inhalation (Myers et al. 2005). It ispossible to explain those discrepancies considering the differencesin the pathways of infection and the bacterial loads used before.However, the same study also demonstrated significantly highernumbers of CD4+ T cells in P2X7R−/−, in relation to wild-type mice(Myers et al. 2005), confirming our results.
Corroborating our data, a study of Vordermeier et al. (1996)demonstrated that B cell-deficient mice infected with M. tuberculo-sis had higher bacterial loads. They considered several mechanisms
by which B cell could influence the course of M. tuberculosis infec-tions. Because of their functions as antigen-presenting cells (APCs),B cells could influence the specificity and phenotype of protectiveCD4+ T cell response, and take up antigens by their antigen-specific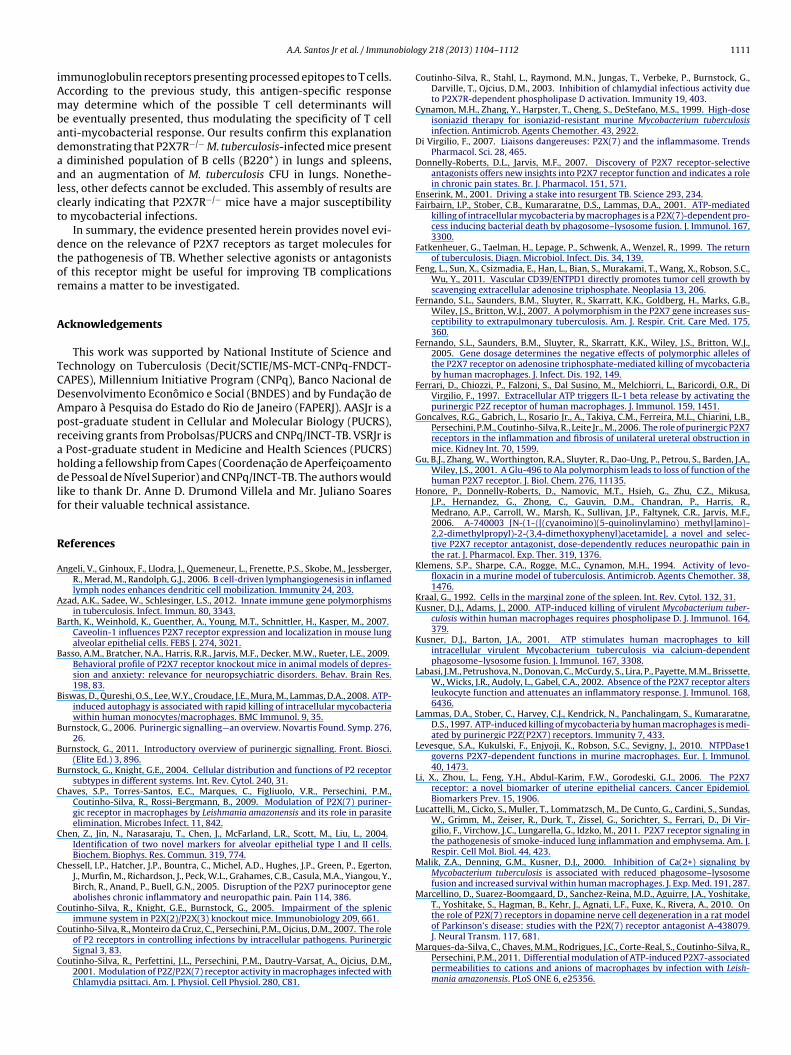
obiolo
iAmbadaalct
dtor
A
TCDAprahdlf
R
A
A
B
B
B
B
B
B
C
C
C
C
C
C
A.A. Santos Jr et al. / Immun
mmunoglobulin receptors presenting processed epitopes to T cells.ccording to the previous study, this antigen-specific responseay determine which of the possible T cell determinants will
e eventually presented, thus modulating the specificity of T cellnti-mycobacterial response. Our results confirm this explanationemonstrating that P2X7R−/− M. tuberculosis-infected mice present
diminished population of B cells (B220+) in lungs and spleens,nd an augmentation of M. tuberculosis CFU in lungs. Nonethe-ess, other defects cannot be excluded. This assembly of results arelearly indicating that P2X7R−/− mice have a major susceptibilityo mycobacterial infections.
In summary, the evidence presented herein provides novel evi-ence on the relevance of P2X7 receptors as target molecules forhe pathogenesis of TB. Whether selective agonists or antagonistsf this receptor might be useful for improving TB complicationsemains a matter to be investigated.
cknowledgements
This work was supported by National Institute of Science andechnology on Tuberculosis (Decit/SCTIE/MS-MCT-CNPq-FNDCT-APES), Millennium Initiative Program (CNPq), Banco Nacional deesenvolvimento Econômico e Social (BNDES) and by Fundac ão demparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ). AASJr is aost-graduate student in Cellular and Molecular Biology (PUCRS),eceiving grants from Probolsas/PUCRS and CNPq/INCT-TB. VSRJr is
Post-graduate student in Medicine and Health Sciences (PUCRS)olding a fellowship from Capes (Coordenac ão de Aperfeic oamentoe Pessoal de Nível Superior) and CNPq/INCT-TB. The authors would
ike to thank Dr. Anne D. Drumond Villela and Mr. Juliano Soaresor their valuable technical assistance.
eferences
ngeli, V., Ginhoux, F., Llodra, J., Quemeneur, L., Frenette, P.S., Skobe, M., Jessberger,R., Merad, M., Randolph, G.J., 2006. B cell-driven lymphangiogenesis in inflamedlymph nodes enhances dendritic cell mobilization. Immunity 24, 203.
zad, A.K., Sadee, W., Schlesinger, L.S., 2012. Innate immune gene polymorphismsin tuberculosis. Infect. Immun. 80, 3343.
arth, K., Weinhold, K., Guenther, A., Young, M.T., Schnittler, H., Kasper, M., 2007.Caveolin-1 influences P2X7 receptor expression and localization in mouse lungalveolar epithelial cells. FEBS J. 274, 3021.
asso, A.M., Bratcher, N.A., Harris, R.R., Jarvis, M.F., Decker, M.W., Rueter, L.E., 2009.Behavioral profile of P2X7 receptor knockout mice in animal models of depres-sion and anxiety: relevance for neuropsychiatric disorders. Behav. Brain Res.198, 83.
iswas, D., Qureshi, O.S., Lee, W.Y., Croudace, J.E., Mura, M., Lammas, D.A., 2008. ATP-induced autophagy is associated with rapid killing of intracellular mycobacteriawithin human monocytes/macrophages. BMC Immunol. 9, 35.
urnstock, G., 2006. Purinergic signalling—an overview. Novartis Found. Symp. 276,26.
urnstock, G., 2011. Introductory overview of purinergic signalling. Front. Biosci.(Elite Ed.) 3, 896.
urnstock, G., Knight, G.E., 2004. Cellular distribution and functions of P2 receptorsubtypes in different systems. Int. Rev. Cytol. 240, 31.
haves, S.P., Torres-Santos, E.C., Marques, C., Figliuolo, V.R., Persechini, P.M.,Coutinho-Silva, R., Rossi-Bergmann, B., 2009. Modulation of P2X(7) puriner-gic receptor in macrophages by Leishmania amazonensis and its role in parasiteelimination. Microbes Infect. 11, 842.
hen, Z., Jin, N., Narasaraju, T., Chen, J., McFarland, L.R., Scott, M., Liu, L., 2004.Identification of two novel markers for alveolar epithelial type I and II cells.Biochem. Biophys. Res. Commun. 319, 774.
hessell, I.P., Hatcher, J.P., Bountra, C., Michel, A.D., Hughes, J.P., Green, P., Egerton,J., Murfin, M., Richardson, J., Peck, W.L., Grahames, C.B., Casula, M.A., Yiangou, Y.,Birch, R., Anand, P., Buell, G.N., 2005. Disruption of the P2X7 purinoceptor geneabolishes chronic inflammatory and neuropathic pain. Pain 114, 386.
outinho-Silva, R., Knight, G.E., Burnstock, G., 2005. Impairment of the splenicimmune system in P2X(2)/P2X(3) knockout mice. Immunobiology 209, 661.
outinho-Silva, R., Monteiro da Cruz, C., Persechini, P.M., Ojcius, D.M., 2007. The role
of P2 receptors in controlling infections by intracellular pathogens. PurinergicSignal 3, 83.outinho-Silva, R., Perfettini, J.L., Persechini, P.M., Dautry-Varsat, A., Ojcius, D.M.,2001. Modulation of P2Z/P2X(7) receptor activity in macrophages infected withChlamydia psittaci. Am. J. Physiol. Cell Physiol. 280, C81.
gy 218 (2013) 1104– 1112 1111
Coutinho-Silva, R., Stahl, L., Raymond, M.N., Jungas, T., Verbeke, P., Burnstock, G.,Darville, T., Ojcius, D.M., 2003. Inhibition of chlamydial infectious activity dueto P2X7R-dependent phospholipase D activation. Immunity 19, 403.
Cynamon, M.H., Zhang, Y., Harpster, T., Cheng, S., DeStefano, M.S., 1999. High-doseisoniazid therapy for isoniazid-resistant murine Mycobacterium tuberculosisinfection. Antimicrob. Agents Chemother. 43, 2922.
Di Virgilio, F., 2007. Liaisons dangereuses: P2X(7) and the inflammasome. TrendsPharmacol. Sci. 28, 465.
Donnelly-Roberts, D.L., Jarvis, M.F., 2007. Discovery of P2X7 receptor-selectiveantagonists offers new insights into P2X7 receptor function and indicates a rolein chronic pain states. Br. J. Pharmacol. 151, 571.
Enserink, M., 2001. Driving a stake into resurgent TB. Science 293, 234.Fairbairn, I.P., Stober, C.B., Kumararatne, D.S., Lammas, D.A., 2001. ATP-mediated
killing of intracellular mycobacteria by macrophages is a P2X(7)-dependent pro-cess inducing bacterial death by phagosome–lysosome fusion. J. Immunol. 167,3300.
Fatkenheuer, G., Taelman, H., Lepage, P., Schwenk, A., Wenzel, R., 1999. The returnof tuberculosis. Diagn. Microbiol. Infect. Dis. 34, 139.
Feng, L., Sun, X., Csizmadia, E., Han, L., Bian, S., Murakami, T., Wang, X., Robson, S.C.,Wu, Y., 2011. Vascular CD39/ENTPD1 directly promotes tumor cell growth byscavenging extracellular adenosine triphosphate. Neoplasia 13, 206.
Fernando, S.L., Saunders, B.M., Sluyter, R., Skarratt, K.K., Goldberg, H., Marks, G.B.,Wiley, J.S., Britton, W.J., 2007. A polymorphism in the P2X7 gene increases sus-ceptibility to extrapulmonary tuberculosis. Am. J. Respir. Crit. Care Med. 175,360.
Fernando, S.L., Saunders, B.M., Sluyter, R., Skarratt, K.K., Wiley, J.S., Britton, W.J.,2005. Gene dosage determines the negative effects of polymorphic alleles ofthe P2X7 receptor on adenosine triphosphate-mediated killing of mycobacteriaby human macrophages. J. Infect. Dis. 192, 149.
Ferrari, D., Chiozzi, P., Falzoni, S., Dal Susino, M., Melchiorri, L., Baricordi, O.R., DiVirgilio, F., 1997. Extracellular ATP triggers IL-1 beta release by activating thepurinergic P2Z receptor of human macrophages. J. Immunol. 159, 1451.
Goncalves, R.G., Gabrich, L., Rosario Jr., A., Takiya, C.M., Ferreira, M.L., Chiarini, L.B.,Persechini, P.M., Coutinho-Silva, R., Leite Jr., M., 2006. The role of purinergic P2X7receptors in the inflammation and fibrosis of unilateral ureteral obstruction inmice. Kidney Int. 70, 1599.
Gu, B.J., Zhang, W., Worthington, R.A., Sluyter, R., Dao-Ung, P., Petrou, S., Barden, J.A.,Wiley, J.S., 2001. A Glu-496 to Ala polymorphism leads to loss of function of thehuman P2X7 receptor. J. Biol. Chem. 276, 11135.
Honore, P., Donnelly-Roberts, D., Namovic, M.T., Hsieh, G., Zhu, C.Z., Mikusa,J.P., Hernandez, G., Zhong, C., Gauvin, D.M., Chandran, P., Harris, R.,Medrano, A.P., Carroll, W., Marsh, K., Sullivan, J.P., Faltynek, C.R., Jarvis, M.F.,2006. A-740003 [N-(1-{[(cyanoimino)(5-quinolinylamino) methyl]amino}-2,2-dimethylpropyl)-2-(3,4-dimethoxyphenyl)acetamide], a novel and selec-tive P2X7 receptor antagonist, dose-dependently reduces neuropathic pain inthe rat. J. Pharmacol. Exp. Ther. 319, 1376.
Klemens, S.P., Sharpe, C.A., Rogge, M.C., Cynamon, M.H., 1994. Activity of levo-floxacin in a murine model of tuberculosis. Antimicrob. Agents Chemother. 38,1476.
Kraal, G., 1992. Cells in the marginal zone of the spleen. Int. Rev. Cytol. 132, 31.Kusner, D.J., Adams, J., 2000. ATP-induced killing of virulent Mycobacterium tuber-
culosis within human macrophages requires phospholipase D. J. Immunol. 164,379.
Kusner, D.J., Barton, J.A., 2001. ATP stimulates human macrophages to killintracellular virulent Mycobacterium tuberculosis via calcium-dependentphagosome–lysosome fusion. J. Immunol. 167, 3308.
Labasi, J.M., Petrushova, N., Donovan, C., McCurdy, S., Lira, P., Payette, M.M., Brissette,W., Wicks, J.R., Audoly, L., Gabel, C.A., 2002. Absence of the P2X7 receptor altersleukocyte function and attenuates an inflammatory response. J. Immunol. 168,6436.
Lammas, D.A., Stober, C., Harvey, C.J., Kendrick, N., Panchalingam, S., Kumararatne,D.S., 1997. ATP-induced killing of mycobacteria by human macrophages is medi-ated by purinergic P2Z(P2X7) receptors. Immunity 7, 433.
Levesque, S.A., Kukulski, F., Enjyoji, K., Robson, S.C., Sevigny, J., 2010. NTPDase1governs P2X7-dependent functions in murine macrophages. Eur. J. Immunol.40, 1473.
Li, X., Zhou, L., Feng, Y.H., Abdul-Karim, F.W., Gorodeski, G.I., 2006. The P2X7receptor: a novel biomarker of uterine epithelial cancers. Cancer Epidemiol.Biomarkers Prev. 15, 1906.
Lucattelli, M., Cicko, S., Muller, T., Lommatzsch, M., De Cunto, G., Cardini, S., Sundas,W., Grimm, M., Zeiser, R., Durk, T., Zissel, G., Sorichter, S., Ferrari, D., Di Vir-gilio, F., Virchow, J.C., Lungarella, G., Idzko, M., 2011. P2X7 receptor signaling inthe pathogenesis of smoke-induced lung inflammation and emphysema. Am. J.Respir. Cell Mol. Biol. 44, 423.
Malik, Z.A., Denning, G.M., Kusner, D.J., 2000. Inhibition of Ca(2+) signaling byMycobacterium tuberculosis is associated with reduced phagosome–lysosomefusion and increased survival within human macrophages. J. Exp. Med. 191, 287.
Marcellino, D., Suarez-Boomgaard, D., Sanchez-Reina, M.D., Aguirre, J.A., Yoshitake,T., Yoshitake, S., Hagman, B., Kehr, J., Agnati, L.F., Fuxe, K., Rivera, A., 2010. Onthe role of P2X(7) receptors in dopamine nerve cell degeneration in a rat modelof Parkinson’s disease: studies with the P2X(7) receptor antagonist A-438079.
J. Neural Transm. 117, 681.Marques-da-Silva, C., Chaves, M.M., Rodrigues, J.C., Corte-Real, S., Coutinho-Silva, R.,Persechini, P.M., 2011. Differential modulation of ATP-induced P2X7-associatedpermeabilities to cations and anions of macrophages by infection with Leish-mania amazonensis. PLoS ONE 6, e25356.

1 obiolo
M
M
M
M
M
M
M
M
N
P
P
R
112 A.A. Santos Jr et al. / Immun
artins de Sousa, E., Bonfim de Bortoli, F., Amaral, E.P., Batista, A.C., LibermanKipnis, T., Marques Cardoso, A., Kipnis, A., Junqueira-Kipnis, A.P., 2010. Acuteimmune response to Mycobacterium massiliense in C57BL/6 and BALB/c mice.Infect. Immun. 78, 1571.
artins, J.P., Silva, R.B., Coutinho-Silva, R., Takiya, C.M., Battastini, A.M., Morrone,F.B., Campos, M.M., 2012. The role of P2X7 purinergic receptors in inflammatoryand nociceptive changes accompanying cyclophosphamide-induced haemor-rhagic cystitis in mice. Br. J. Pharmacol. 165, 183.
ascarello, A., Chiaradia, L.D., Vernal, J., Villarino, A., Guido, R.V., Perizzolo, P., Poirier,V., Wong, D., Martins, P.G., Nunes, R.J., Yunes, R.A., Andricopulo, A.D., Av-Gay,Y., Terenzi, H., 2010. Inhibition of Mycobacterium tuberculosis tyrosine phos-phatase PtpA by synthetic chalcones: kinetics, molecular modeling, toxicity andeffect on growth. Bioorg. Med. Chem. 18, 3783.
ebius, R.E., Kraal, G., 2005. Structure and function of the spleen. Nat. Rev. Immunol.5, 606.
olloy, A., Laochumroonvorapong, P., Kaplan, G., 1994. Apoptosis, but not necrosis,of infected monocytes is coupled with killing of intracellular bacillus Calmette-Guerin. J. Exp. Med. 180, 1499.
oncao-Ribeiro, L.C., Cagido, V.R., Lima-Murad, G., Santana, P.T., Riva, D.R., Borojevic,R., Zin, W.A., Cavalcante, M.C., Rica, I., Brando-Lima, A.C., Takiya, C.M., Faffe, D.S.,Coutinho-Silva, R., 2011. Lipopolysaccharide-induced lung injury: role of P2X7receptor. Respir. Physiol. Neurobiol. 179, 314.
uller, T., Vieira, R.P., Grimm, M., Durk, T., Cicko, S., Zeiser, R., Jakob, T., Martin,S.F., Blumenthal, B., Sorichter, S., Ferrari, D., Di Virgillio, F., Idzko, M., 2011. Apotential role for P2X7R in allergic airway inflammation in mice and humans.Am. J. Respir. Cell Mol. Biol. 44, 456.
yers, A.J., Eilertson, B., Fulton, S.A., Flynn, J.L., Canaday, D.H., 2005. The purinergicP2X7 receptor is not required for control of pulmonary Mycobacterium tuber-culosis infection. Infect. Immun. 73, 3192.
ino-Moreno, P., Portales-Perez, D., Hernandez-Castro, B., Portales-Cervantes, L.,Flores-Meraz, V., Baranda, L., Gomez-Gomez, A., Acuna-Alonzo, V., Granados, J.,Gonzalez-Amaro, R., 2007. P2X7 and NRAMP1/SLC11 A1 gene polymorphismsin Mexican mestizo patients with pulmonary tuberculosis. Clin. Exp. Immunol.148, 469.
alomino, J.C., Martin, A., Camacho, M., Guerra, H., Swings, J., Portaels, F., 2002.Resazurin microtiter assay plate: simple and inexpensive method for detectionof drug resistance in Mycobacterium tuberculosis. Antimicrob. Agents Chemother.46, 2720.
ethe, K., Swenson, D.L., Alonso, S., Anderson, J., Wang, C., Russell, D.G., 2004. Isola-
tion of Mycobacterium tuberculosis mutants defective in the arrest of phagosomematuration. Proc. Natl. Acad. Sci. U. S. A. 101, 13642.assendren, F., Buell, G.N., Virginio, C., Collo, G., North, R.A., Surprenant, A., 1997. Thepermeabilizing ATP receptor, P2X7. Cloning and expression of a human cDNA.J. Biol. Chem. 272, 5482.
gy 218 (2013) 1104– 1112
Rodrigues-Junior, V.S., Dos Santos Junior, A., Dos Santos, A.J., Schneider, C.Z., Calixto,J.B., Sousa, E.H., de Franca Lopes, L.G., Souto, A.A., Basso, L.A., Santos, D.S., Campos,M.M., 2012. Activity of IQG-607, a new orally active compound, in a murinemodel of Mycobacterium tuberculosis infection. Int. J. Antimicrob. Agents 40, 182.
Saini, D., Hopkins, G.W., Seay, S.A., Chen, C.J., Perley, C.C., Click, E.M., Frothingham,R., 2012. Ultra-low dose of Mycobacterium tuberculosis aerosol creates partialinfection in mice. Tuberculosis (Edinburgh) 92, 160.
Saunders, B.M., Fernando, S.L., Sluyter, R., Britton, W.J., Wiley, J.S., 2003. A loss-of-function polymorphism in the human P2X7 receptor abolishes ATP-mediatedkilling of mycobacteria. J. Immunol. 171, 5442.
Schenk, U., Frascoli, M., Proietti, M., Geffers, R., Traggiai, E., Buer, J., Ricordi, C.,Westendorf, A.M., Grassi, F., 2011. ATP inhibits the generation and function ofregulatory T cells through the activation of purinergic P2X receptors. Sci. Signal.4, ra12.
Singla, N., Gupta, D., Joshi, A., Batra, N., Singh, J., 2012. Genetic polymorphisms in theP2X7 gene and its association with susceptibility to tuberculosis. Int. J. Tuberc.Lung Dis. 16, 224.
Sluyter, R., Dalitz, J.G., Wiley, J.S., 2004a. P2X7 receptor polymorphism impairs extra-cellular adenosine 5’-triphosphate-induced interleukin-18 release from humanmonocytes. Genes Immun. 5, 588.
Sluyter, R., Shemon, A.N., Wiley, J.S., 2004b. Glu496 to Ala polymorphism in theP2X7 receptor impairs ATP-induced IL-1 beta release from human monocytes.J. Immunol. 172, 3399.
Sluyter, R., Stokes, L., 2011. Significance of P2X7 receptor variants to human healthand disease. Recent Pat. DNA Gene Seq. 5, 41.
Sluyter, R., Wiley, J.S., 2002. Extracellular adenosine 5′-triphosphate induces a loss ofCD23 from human dendritic cells via activation of P2X7 receptors. Int. Immunol.14, 1415.
Smith, R.A., Alvarez, A.J., Estes, D.M., 2001. The P2X7 purinergic receptor on bovinemacrophages mediates mycobacterial death. Vet. Immunol. Immunopathol. 78,249.
Teixeira, J.M., Oliveira, M.C., Parada, C.A., Tambeli, C.H., 2010. Peripheral mecha-nisms underlying the essential role of P2X7 receptors in the development ofinflammatory hyperalgesia. Eur. J. Pharmacol. 644, 55.
Vordermeier, H.M., Venkataprasad, N., Harris, D.P., Ivanyi, J., 1996. Increase of tuber-culous infection in the organs of B cell-deficient mice. Clin. Exp. Immunol. 106,312.
Wickelgren, I., 2000. New clues to how the TB bacillus persists. Science 288, 1314.Wiley, J.S., Sluyter, R., Gu, B.J., Stokes, L., Fuller, S.J., 2011. The human P2X7 receptor
and its role in innate immunity. Tissue Antigens 78, 321.World Health Organization, 2010. Global Tuberculosis Control: WHO Report 2010.
WHO, Geneva.World Health Organization, 2011. Global Tuberculosis Control: WHO Report 2011.
WHO, Geneva.
Copyright © 2022 FDOKUMEN




















