
Disruption of the P2X7 purinoceptor gene abolishes chronic inflammatory and neuropathic pain
11
Disruption of the P2X 7 purinoceptor gene abolishes chronic inflammatory and neuropathic pain Iain P. Chessell a, * , Jonathan P. Hatcher a , Chas Bountra a , Anton D. Michel a , Jane P. Hughes b , Paula Green b , Julie Egerton b , Melanie Murfin a , Jill Richardson b , Wendy L. Peck b , Caroline B.A. Grahames c,1 , Maria Anna Casula d , Yiangos Yiangou d , Rolfe Birch e , Praveen Anand d , Gary N. Buell f,2 a Pain Research, N&GI CEDD, GlaxoSmithKline, Third Avenue, Harlow, Essex CM19 5AW, UK b Neuro-Cell Sciences, GlaxoSmithKline, Third Avenue, Harlow CM19 5AW, UK c Glaxo Institute of Applied Pharmacology, Tennis Court Road, Cambridge CB2 5DH, UK d Peripheral Neuropathy Unit, Imperial College School of Medicine, Hammersmith Hospital, Du Cane Road, London W12 0NN, UK e Peripheral Nerve Injury Unit, Royal National Orthopaedic Hospital, Stanmore HA7 4LP, UK f Glaxo Institute of Molecular Biology, 14 Chemin Aulx, Plan Les Ouates, Geneva CH-1228, Switzerland Received 20 August 2004; received in revised form 24 November 2004; accepted 5 January 2005 Abstract The P2X 7 purinoceptor is a ligand-gated cation channel, expressed predominantly by cells of immune origin, with a unique phenotype which includes release of biologically active inflammatory cytokine, interleukin (IL)-1b following activation, and unique ion channel biophysics observed only in this receptor family. Here we demonstrate that in mice lacking this receptor, inflammatory (in an adjuvant- induced model) and neuropathic (in a partial nerve ligation model) hypersensitivity is completely absent to both mechanical and thermal stimuli, whilst normal nociceptive processing is preserved. The knockout animals were unimpaired in their ability to produce mRNA for pro- IL-1b, and cytometric analysis of paw and systemic cytokines from knockout and wild-type animals following adjuvant insult suggests a selective effect of the gene deletion on release of IL-1b and IL-10, with systemic reductions in adjuvant-induced increases in IL-6 and MCP-1. In addition, we show that this receptor is upregulated in human dorsal root ganglia and injured nerves obtained from chronic neuropathic pain patients. We hypothesise that the P2X 7 receptor, via regulation of mature IL-1b production, plays a common upstream transductional role in the development of pain of neuropathic and inflammatory origin. Drugs which block this target may have the potential to deliver broad-spectrum analgesia. q 2005 International Association for the Study of Pain. Published by Elsevier B.V. All rights reserved. Keywords: P2X 7 ; Purinoceptor; Interleukin 1 beta; Neuropathic; Taqman 1. Introduction The relationships governing expression and regulation of key inflammatory and sensitising mediators are highly complex with multiple redundancies. Identification of upstream processes for intervention has proved to be extremely challenging, either because discovery of suitable molecular tools has hampered progress (e.g. interleukin-1b antagonists) or because disruption of a signalling process has revealed multiple divergent roles (e.g. inducible nitric oxide) (Aley et al., 1998; Budzinski et al., 2000). Chronic pain is a huge global issue, affecting up to 80% of the population (Clark, 2003), and the difficulty in identifying pivotal processes manifests as significant shortcomings in Pain 114 (2005) 386–396 www.elsevier.com/locate/pain 0304-3959/$20.00 q 2005 International Association for the Study of Pain. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.pain.2005.01.002 * Corresponding author. Tel.: C44 1279875362. E-mail address: [email protected] (I.P. Chessell). 1 Present address: AstraZeneca R&D Charnwood, Bakewell Road, Loughborough, Leicestershire LE11 5RH, UK. 2 Present address: Serono International, 15bis, Chemin des Mines, CH-1211 Geneva 20, Switzerland.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Disruption of the P2X7 purinoceptor gene abolishes chronic inflammatory and neuropathic pain

Disruption of the P2X7 purinoceptor gene abolishes chronic inflammatory
and neuropathic pain
Iain P. Chessella,*, Jonathan P. Hatchera, Chas Bountraa, Anton D. Michela, Jane P. Hughesb,Paula Greenb, Julie Egertonb, Melanie Murfina, Jill Richardsonb, Wendy L. Peckb,Caroline B.A. Grahamesc,1, Maria Anna Casulad, Yiangos Yiangoud, Rolfe Birche,
Praveen Anandd, Gary N. Buellf,2
aPain Research, N&GI CEDD, GlaxoSmithKline, Third Avenue, Harlow, Essex CM19 5AW, UKbNeuro-Cell Sciences, GlaxoSmithKline, Third Avenue, Harlow CM19 5AW, UK
cGlaxo Institute of Applied Pharmacology, Tennis Court Road, Cambridge CB2 5DH, UKdPeripheral Neuropathy Unit, Imperial College School of Medicine, Hammersmith Hospital, Du Cane Road, London W12 0NN, UK
ePeripheral Nerve Injury Unit, Royal National Orthopaedic Hospital, Stanmore HA7 4LP, UKfGlaxo Institute of Molecular Biology, 14 Chemin Aulx, Plan Les Ouates, Geneva CH-1228, Switzerland
Received 20 August 2004; received in revised form 24 November 2004; accepted 5 January 2005
Abstract
The P2X7 purinoceptor is a ligand-gated cation channel, expressed predominantly by cells of immune origin, with a unique phenotype
which includes release of biologically active inflammatory cytokine, interleukin (IL)-1b following activation, and unique ion channel
biophysics observed only in this receptor family. Here we demonstrate that in mice lacking this receptor, inflammatory (in an adjuvant-
induced model) and neuropathic (in a partial nerve ligation model) hypersensitivity is completely absent to both mechanical and thermal
stimuli, whilst normal nociceptive processing is preserved. The knockout animals were unimpaired in their ability to produce mRNA for pro-
IL-1b, and cytometric analysis of paw and systemic cytokines from knockout and wild-type animals following adjuvant insult suggests a
selective effect of the gene deletion on release of IL-1b and IL-10, with systemic reductions in adjuvant-induced increases in IL-6 and
MCP-1. In addition, we show that this receptor is upregulated in human dorsal root ganglia and injured nerves obtained from chronic
neuropathic pain patients. We hypothesise that the P2X7 receptor, via regulation of mature IL-1b production, plays a common upstream
transductional role in the development of pain of neuropathic and inflammatory origin. Drugs which block this target may have the potential
to deliver broad-spectrum analgesia.
q 2005 International Association for the Study of Pain. Published by Elsevier B.V. All rights reserved.
Keywords: P2X7; Purinoceptor; Interleukin 1 beta; Neuropathic; Taqman
1. Introduction
The relationships governing expression and regulation of
key inflammatory and sensitising mediators are highly
0304-3959/$20.00 q 2005 International Association for the Study of Pain. Publi
doi:10.1016/j.pain.2005.01.002
* Corresponding author. Tel.: C44 1279875362.
E-mail address: [email protected] (I.P. Chessell).1 Present address: AstraZeneca R&D Charnwood, Bakewell Road,
Loughborough, Leicestershire LE11 5RH, UK.2 Present address: Serono International, 15bis, Chemin des Mines,
CH-1211 Geneva 20, Switzerland.
complex with multiple redundancies. Identification of
upstream processes for intervention has proved to be
extremely challenging, either because discovery of suitable
molecular tools has hampered progress (e.g. interleukin-1bantagonists) or because disruption of a signalling process
has revealed multiple divergent roles (e.g. inducible nitric
oxide) (Aley et al., 1998; Budzinski et al., 2000). Chronic
pain is a huge global issue, affecting up to 80% of the
population (Clark, 2003), and the difficulty in identifying
pivotal processes manifests as significant shortcomings in
Pain 114 (2005) 386–396
www.elsevier.com/locate/pain
shed by Elsevier B.V. All rights reserved.

I.P. Chessell et al. / Pain 114 (2005) 386–396 387
the treatment of pain; currently no drug is capable of
adequately treating both inflammatory and neuropathic
pain, and drugs which are currently used to treat these pain
states suffer from poor responder rates, whilst more
efficacious therapies are often limited by tolerability and
safety concerns.
The P2X7 purinoceptor for ATP is a ligand-gated ion
channel with several unusual phenotypic features (Chessell
et al., 1998; Surprenant et al., 1996). Expressed predomi-
nantly on immune cells (Collo et al., 1997; see also Sim
et al., 2004), the receptor functions as a non-selective
cationic channel following brief activation, but prolonged or
repeated activation yields a pore dilation phenomenon.
Uniquely, activation of the P2X7 receptor is also associated
with release of mature, biologically active interleukin-1b(IL-1b; Perregaux and Gabel, 1994; Solle et al., 2001), a
potent inflammatory cytokine whose downstream effects
include induction of nitric oxide synthase (iNOS), cyclo-
oxygenase 2 (COX-2; Samad et al., 2001) and production of
superoxide products (Parvathenani et al., 2003) and tumour
necrosis factor a (TNF-a; Woolf et al., 1997), all of which
have well described roles in the generation or maintenance
of pain. Whilst currently used clinical pain therapies address
some of these targets individually (e.g. COX-2 inhibitors),
interruption of the pain cascade upstream may deliver a
broader analgesic profile than that achieved using current
therapies. Investigation of such a role of the P2X7 receptor
has been difficult as available antagonists lack selectivity,
although more selective molecules are now being developed
(Alcarez et al., 2004). Here we have used homologous
recombination to generate mice with a disruption of exon 1
of the mouse P2X7 gene to investigate the role of this
receptor in inflammatory and neuropathic hypersensitivity,
using recognised animal models of these pain states
(intraplantar Freund’s Complete Adjuvant (FCA), and
partial ligation of the sciatic nerve). Whilst genetically
modified animals may be subject to compensatory changes
in signalling or transduction, such phenotypic analysis
obtained in the absence of selective pharmacological tools
can be a useful predictor of potential drug action. We have
examined the effect of the knockout on the regulation of the
production of a number of inflammatory cytokines to further
elucidate the mechanism underlying the observed pheno-
type in the animals with the gene deletion. We have also
studied the expression of P2X7-like immunoreactivity in
painful injured human nerves and avulsed human dorsal root
ganglia to indicate whether this expression of this receptor is
changed in a human chronic pain state.
2. Methods
2.1. Targeting of P2X7 gene and generation of mutant mice
Mice carrying a targeted null mutation of the P2X7 gene were
generated according to published protocols (Conquet, 1995).
The P2X7 gene was isolated from a genomic library obtained
from 129/Sv mice. Partial sequencing of the 5 0 exons permitted
ligation of two fragments into the neomycin resistant ‘knock-out
vector’ pGN, a plasmid described previously (Le Mouellic et al.,
1990). Homologous recombination of the resulting plasmid DNA
into embryonic stem cells resulted in a disrupted P2X7 gene.
Germline chimaeras were crossed with C57Bl/6J females to
generate heterozygotes, and then intercrossed giving rise to overtly
healthy mutant offspring in the expected Mendelian ratio.
Successful targetting and transmission was confirmed by Southern
and Western analysis (Collo et al., 1997), by the polymerase chain
reaction, and by monitoring loss of channel function, using ATP-
activated YO-PRO-1 permeation of peritoneal macrophages
(Steinberg et al., 1987). As described for similar P2X7 mutants
(Solle et al., 2001), mice displayed altered cytokine production in
response to ATP and lipopolysaccharide. A further 6 backcrosses
onto the C57Bl/6J strain were performed before producing
homozygotes for study. Where animal numbers were limiting,
some studies were performed in one sex only.
2.2. Quantification of dorsal root ganglia neuronal populations
L4 and L5 dorsal root ganglia (DRG) were removed from
P2X7K/K and wild type litter mate controls (nZ5 animals per
group). Following fixation of DRG in 4% paraformaldehyde, DRG
were sectioned (12 mm) and stained for neurofilament L (1:50;
Novocastra, Newcastle, UK) and peripherin (1:1000; Chemicon,
Temicula, USA) to provide a total neuronal stain. Antibody
labelling was detected using standard fluorescent techniques. For
each animal, 4 whole DRG sections were analysed using KS300
image analysis software. The total numbers of neurones were
counted, and cell size determined for each neurone. The total
counts for the 5 animals in each group were then summed, and
distribution histograms generated and analysed using Stastistica
(Statsoft, Tulsa, USA).
2.3. Hot plate test
All animal studies were conducted under a UK Home Office
Licence in accordance with regulated guidelines. Mice of each sex
and genotype (nZ15) were tested in a random and blind fashion
for thermal withdrawal nociception using a hot plate (Harvard
Analgesia Meter, Harvard Instruments, USA) maintained at 50,
52.5 or 55 8C. Mice were observed for signs of nociception, i.e.
rapid fanning or licking of the paws. The response latency was
recorded and results analysed using 2-way analysis of variance in
Statistica (Statsoft Inc., Tulsa, USA) with sex, genotype and hot
plate temperature being used as independent variables. Follow up
analysis was carried out using Duncan’s test.
2.4. FCA-induced inflammation
Female mice (nZ15) were employed for these studies. Animals
were tested for changes in mechanical hypersensitivity using a
weight bearing averager. Following the establishment of baseline
latencies all mice received a sub plantar injection of 30 ml of
1 mg/ml FCA. Mice were re-tested 1 and 7 days later for changes
in weight bearing. Results were expressed as mean ipsilateral/con-
tralateral ratio for each test day and analysed using 2-way analysis
of variance in Statistica with genotype and days post FCA being

I.P. Chessell et al. / Pain 114 (2005) 386–396388
used as independent variables. Follow up analysis was carried out
using Duncan’s test.
2.5. Partial nerve ligation
Two separate studies were performed. For measurement of
thermal hyperalgesia, 20 mice of each sex and genotype were used,
whilst for investigation of mechanical hyperalgesia, 15 female
mice of each genotype were employed. For thermal studies, mice
were tested for paw withdrawal latency by aiming a localised heat
source (Plantar Test Apparatus, Ugo Basile, Italy) at the plantar
surface of the foot. For mechanical studies, withdrawal latencies
were assessed using a hand held electronic Von Frey hair (Life
Science Instruments, UK) onto the plantar surface of each hind
paw in turn until a withdrawal response was observed. On day 0 all
mice underwent surgery to partially ligate the sciatic nerve using a
method based on that described by Seltzer et al. (2000). After 3
days recovery, mice were tested for paw withdrawal latency, and
testing was repeated up to 28 days post operation. For the thermal
readout, results were expressed as mean withdrawal latency for the
ipsilateral paw and expressed as percentage of baseline reading.
For the mechanical readout, results were expressed as mean
percentage ipsilateral/contralateral ratio for each test day. Results
were analysed using 2-way analysis of variance in Statistica with
sex, genotype and days post op being used as independent
variables. Follow up analysis was carried out using Duncan’s test.
2.6. Profiling pro-IL1b gene expression
Tissues were isolated, dissected and stored at K80 8C prior to
extraction. RNA was then isolated from the animal tissues using
RNeasyw Lipid Tissue Mini Kit (Qiagen), following manufac-
turers’ instructions.
Reverse transcription and Taqmanw RT-PCR was performed as
described previously (MacDonald et al., 2001). The primer and
probe set (synthesized by Promega) were designed using Primer3
software and primer specificity was verified using FASTA
searches. Primer and probe sequences (forward primer, reverse
primer, Taqman probe): 5 0-TTGGGCCTCAAAGGAAAGAAT,
5 0-TCTCCAGCTGCAGGGTGG, 5-TATACCTGTCCTGTG-
TAATGAAAGACGGCACA. Analysis of covariance was per-
formed using Microsoftw Excel 97 (Microsoft, Redmond, USA)
with an add-in toolkit v2.1.2 for the analysis of real-time PCR data
(PRISM Training and Consultancy Ltd, Cambridge, UK).
Cyclophilin was used as covariate for normalization.
2.7. Cytokine analysis in P2X7C/C and P2X7
K/K animals
Studies to examine ex vivo, cellular regulation of IL-1b were
performed as previously described with modification (Grahames
et al., 1999). Briefly, in experiments using P2X7K/K animals,
peritoneal macrophages were elicited by injection of 1.5 ml
sterilised 4% thioglycolate into the peritoneal cavity of male F2
P2X7K/K mice (nZ6) or male C57Bl/6-DBA/1 F2 hybrid control
mice (nZ6). Macrophages were lavaged from the cavity 3 days
later using 5 ml DMEM media. Cells were plated in 24 well plates
and left for at least 24 h to adhere. Cells were maintained in
DMEM/10% foetal bovine serum/penicillin (100 U/ml) and
streptomycin (0.1 mg/ml) (Gibco, Paisley, UK). Similarly, follow-
ing seeding, THP-1 cells were maintained in RPMI/10% foetal
bovine serum (Gibco, Paisley, UK). Two days prior to experimen-
tation, THP-1 cells were differentiated using 0.5 mM phorbol-12-
myristate-13-acetate (PMA; Sigma, Poole, UK) for 3 h. Following
washing, all cells were replated at a density of 4!105 cells/well,
left for 24 h and treated with 10 mg/ml LPS for 24 h. Cells were
washed again in complete media immediately prior to experimen-
tation. For experiments examining peritoneal macrophage IL-1brelease, cells were incubated with various concentrations of 2 0- and
3 0-O-(4-benzoyl-benzoyl)-ATP (Bz-ATP; Sigma, Poole, UK) in a
sucrose-based buffer (Michel et al., 1999) at 37 8C and super-
natants removed 30 min following Bz-ATP application. For
experiments examining THP-1 IL-1b release, cells were incubated
in complete media with various concentrations of Bz-ATP and
reactions were terminated at 0, 4, 32 or 64 min. In some
experiments, apyrase (0.4 U/ml; Sigma, Poole, UK) was added to
the wells at 4 min following agonist application. Experiments were
performed in triplicate on at least 3 separate occasions.
Concentrations of IL-1b were performed using a Medgenix IL-
1b EASIA kit, according to the manufacturers instructions. The
monoclonal antibody supplied with this kit does not cross-react
with pro-IL-1b.
For cytometric analysis of concentrations of IL-10, IL-1b, IL-
6, MCP-1 and TNF-a following adjuvant treatment in P2X7K/K
and P2X7C/C animals, blood was collected into EDTA tubes and
subjected to centrifugation (13,000 rpm for 5 min), the resulting
plasma supernatants were harvested and their cytokine content
analysed by Luminex (Luminex Corp.) using a Bio-Plex
cytokine assay (Bio-Rad Laboratories, Inc.) as per manufacturers
instructions. Samples were collected at day 0, 1 and 7 following
FCA injection (nZ5 per group).
Paw samples were dissected and homogenised in PBS
containing ‘complete’ protease inhibitor cocktail (1 tablet/50 ml,
Boehringer Mannheim) using a Mixer Mill MM300 (Qiagen). The
homogenised samples were subjected to centrifugation
(13,000 rpm for 5 min) with the resulting supernatants analysed
for their cytokine content by Luminex (Luminex Corp.) using a
Bio-Plex cytokine assay (Bio-Rad Laboratories, Inc.) as per
manufacturers instructions. Samples were collected at day 0, 1
and 7 following FCA injection (nZ5 per group).
2.8. Immunochemical studies
Fully informed consent was obtained for all human tissues
collected, with approval of the Local Ethics Committee. Peripheral
nerves proximal to injury were obtained from patients with
associated maximum pain greater than 4/10 on a visual analogue
scale in the 24 h prior to surgery (nZ11; 5 male; age range 26–63,
delay from 7 weeks up to 8 y between injury and surgery): these
were used for immunohistochemical (IHC) studies. Extracts from
painful injured nerves (nZ15; 12 male; age range 15–62 y; delay
6 weeks-1 y) were available for Western blotting (WB). Control
nerves (nZ21; 19 male; age range 6–77 y; 13 for IHC and 8 for
WB) were obtained during limb amputation for a non-neurological
reason. Avulsed (central rhizotomy) DRG (nZ14; 13 male; age
range 15–44 y, delay 2 days up to 4.5 months) were obtained from
patients during brachial plexus reconstruction, 12 for IHC (11 male;
age range 15–44 y) and 8 for WB (8 male; age range 18–37 y).
Control post-mortem DRG (nZ13; 4 male; age range 34–98 yrs;
10 for IHC and 5 for WB) were obtained via the Netherlands Brain
Bank. Frozen post-fixed tissue sections (10 mm) were
I.P. Chessell et al. / Pain 114 (2005) 386–396 389
immunostained with primary antibodies to P2X7 (GlaxoSmithK-
line, Harlow, UK), GFAP (glial fibrillary acidic protein; Dako
Cytomation, High Wycombe, Bucks, UK), and neurofilaments
(Dako Cytomation, High Wycombe, Bucks, UK). Specificity of the
P2X7 antibody used has been demonstrated previously (Buell et al.,
1998). Sites of antibody attachment were revealed using a nickel-
enhanced ABC (immunoperoxidase; Vector Laboratories, High
Wycombe, Bucks., UK) method (Shu et al., 1988) or immunoalka-
line phosphatase (APAAP; DAKO Ltd, Ely, Cambs., UK) for
double staining co-localisation studies.
Computerized image analysis (Seescan, Cambs., UK) was used
to assess changes of immunoreactivity. Images were collected
from five microscopic fields (objective !40) from each specimen.
A fixed detection threshold was used for P2X7 and GFAP in
peripheral nerves, and the P2X7:GFAP ratio determined. For DRG,
in order to examine any changes in satellite cells, P2X7-
immunoreactivity was related to neuronal density, since neurons
and their associated satellite cells are not randomly distributed—all
neuronal profiles in each field were outlined and image analysed to
determine P2X7:neuronal area ratio.
For Western blotting, computerised image analysis and optical
density analysis of the P2X7 bands was performed as previously
described using 6% polyacrylamide gels (Yiangou et al., 2001).
3. Results
3.1. Quantification of DRG cell numbers
No significant differences (ANOVA comparison of
log transformed distribution data) were observed in
total cell counts, or distribution of cell size between the
P2X7K/K and P2X7
C/C animals (Fig. 1).
3.2. Neuropathic hypersensitivity
The partial nerve ligation model (Seltzer et al., 2000) is
an established preclinical model of neuropathic hypersensi-
tivity. Here, we have shown profound differences in both
thermal and mechanical sensitivity between P2X7C/C and
Fig. 1. Size distribution histogram for neuronal count against cross sectional
area for whole DRG sections from P2X7K/K (open bars) and P2X7
C/C (filled
bars) animals. No significant differences were observed between the cell
numbers or cross sectional area between the knockout and wild type
animals. NZ5 per group.
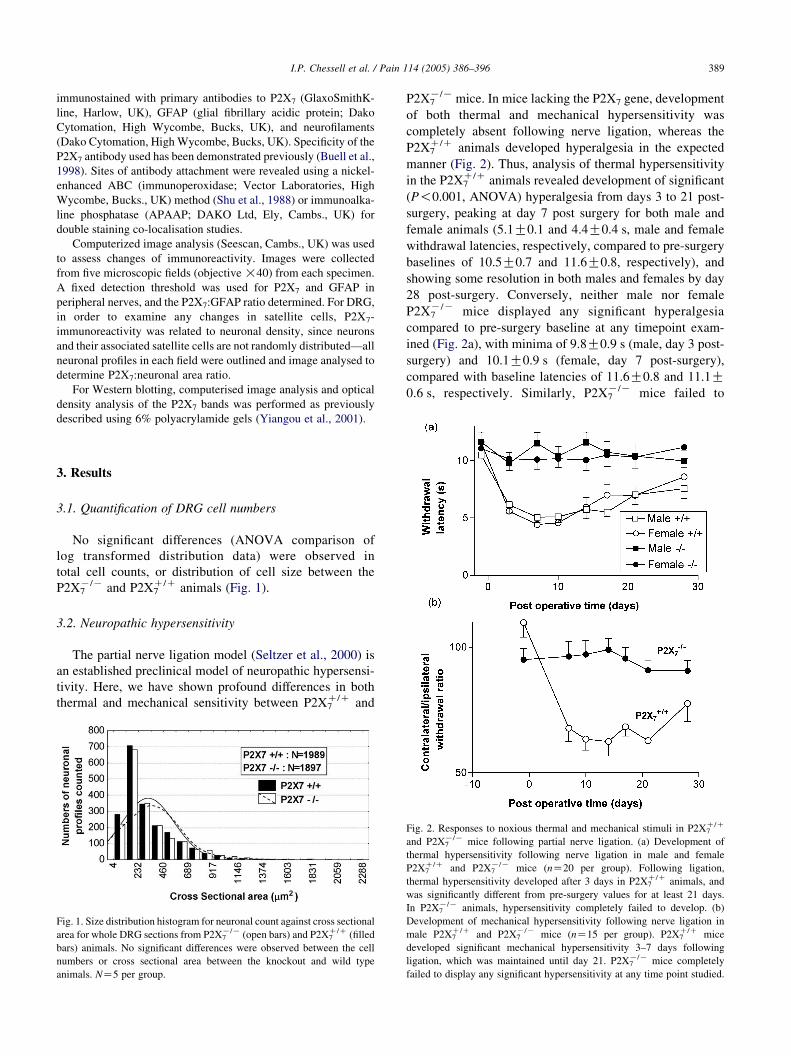
P2X7K/K mice. In mice lacking the P2X7 gene, development
of both thermal and mechanical hypersensitivity was
completely absent following nerve ligation, whereas the
P2X7C/C animals developed hyperalgesia in the expected
manner (Fig. 2). Thus, analysis of thermal hypersensitivity
in the P2X7C/C animals revealed development of significant
(P!0.001, ANOVA) hyperalgesia from days 3 to 21 post-
surgery, peaking at day 7 post surgery for both male and
female animals (5.1G0.1 and 4.4G0.4 s, male and female
withdrawal latencies, respectively, compared to pre-surgery
baselines of 10.5G0.7 and 11.6G0.8, respectively), and
showing some resolution in both males and females by day
28 post-surgery. Conversely, neither male nor female
P2X7K/K mice displayed any significant hyperalgesia
compared to pre-surgery baseline at any timepoint exam-
ined (Fig. 2a), with minima of 9.8G0.9 s (male, day 3 post-
surgery) and 10.1G0.9 s (female, day 7 post-surgery),
compared with baseline latencies of 11.6G0.8 and 11.1G0.6 s, respectively. Similarly, P2X7
K/K mice failed to
Fig. 2. Responses to noxious thermal and mechanical stimuli in P2X7C/C
and P2X7K/K mice following partial nerve ligation. (a) Development of
thermal hypersensitivity following nerve ligation in male and female
P2X7C/C and P2X7
K/K mice (nZ20 per group). Following ligation,
thermal hypersensitivity developed after 3 days in P2X7C/C animals, and
was significantly different from pre-surgery values for at least 21 days.
In P2X7K/K animals, hypersensitivity completely failed to develop. (b)
Development of mechanical hypersensitivity following nerve ligation in
male P2X7C/C and P2X7
K/K mice (nZ15 per group). P2X7C/C mice
developed significant mechanical hypersensitivity 3–7 days following
ligation, which was maintained until day 21. P2X7K/K mice completely
failed to display any significant hypersensitivity at any time point studied.
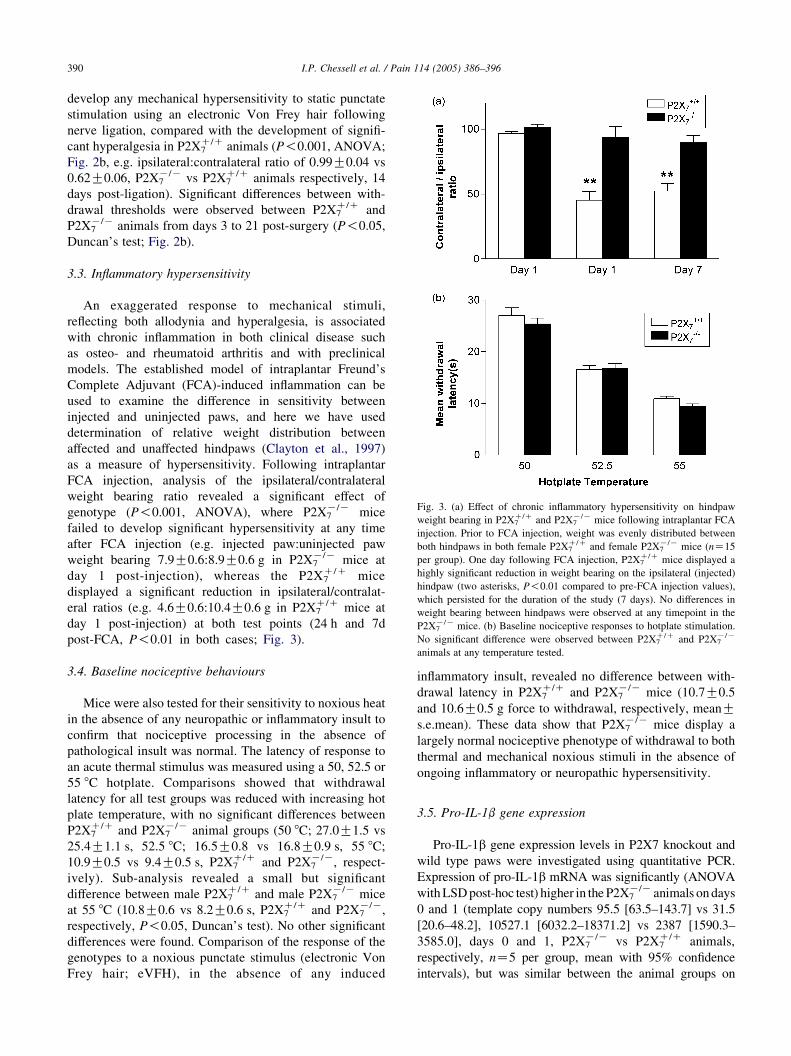
Fig. 3. (a) Effect of chronic inflammatory hypersensitivity on hindpaw
weight bearing in P2X7C/C and P2X7
K/K mice following intraplantar FCA
injection. Prior to FCA injection, weight was evenly distributed between
both hindpaws in both female P2X7C/C and female P2X7
K/K mice (nZ15
per group). One day following FCA injection, P2X7C/C mice displayed a
highly significant reduction in weight bearing on the ipsilateral (injected)
hindpaw (two asterisks, P!0.01 compared to pre-FCA injection values),
which persisted for the duration of the study (7 days). No differences in
weight bearing between hindpaws were observed at any timepoint in the
P2X7K/K mice. (b) Baseline nociceptive responses to hotplate stimulation.
No significant difference were observed between P2X7C/C and P2X7
K/K
animals at any temperature tested.
I.P. Chessell et al. / Pain 114 (2005) 386–396390
develop any mechanical hypersensitivity to static punctate
stimulation using an electronic Von Frey hair following
nerve ligation, compared with the development of signifi-
cant hyperalgesia in P2X7C/C animals (P!0.001, ANOVA;
Fig. 2b, e.g. ipsilateral:contralateral ratio of 0.99G0.04 vs
0.62G0.06, P2X7K/K vs P2X7
C/C animals respectively, 14
days post-ligation). Significant differences between with-
drawal thresholds were observed between P2X7C/C and
P2X7K/K animals from days 3 to 21 post-surgery (P!0.05,
Duncan’s test; Fig. 2b).
3.3. Inflammatory hypersensitivity
An exaggerated response to mechanical stimuli,
reflecting both allodynia and hyperalgesia, is associated
with chronic inflammation in both clinical disease such
as osteo- and rheumatoid arthritis and with preclinical
models. The established model of intraplantar Freund’s
Complete Adjuvant (FCA)-induced inflammation can be
used to examine the difference in sensitivity between
injected and uninjected paws, and here we have used
determination of relative weight distribution between
affected and unaffected hindpaws (Clayton et al., 1997)
as a measure of hypersensitivity. Following intraplantar
FCA injection, analysis of the ipsilateral/contralateral
weight bearing ratio revealed a significant effect of
genotype (P!0.001, ANOVA), where P2X7K/K mice
failed to develop significant hypersensitivity at any time
after FCA injection (e.g. injected paw:uninjected paw
weight bearing 7.9G0.6:8.9G0.6 g in P2X7K/K mice at
day 1 post-injection), whereas the P2X7C/C mice
displayed a significant reduction in ipsilateral/contralat-
eral ratios (e.g. 4.6G0.6:10.4G0.6 g in P2X7C/C mice at
day 1 post-injection) at both test points (24 h and 7d
post-FCA, P!0.01 in both cases; Fig. 3).
3.4. Baseline nociceptive behaviours
Mice were also tested for their sensitivity to noxious heat
in the absence of any neuropathic or inflammatory insult to
confirm that nociceptive processing in the absence of
pathological insult was normal. The latency of response to
an acute thermal stimulus was measured using a 50, 52.5 or
55 8C hotplate. Comparisons showed that withdrawal
latency for all test groups was reduced with increasing hot
plate temperature, with no significant differences between
P2X7C/C and P2X7
K/K animal groups (50 8C; 27.0G1.5 vs
25.4G1.1 s, 52.5 8C; 16.5G0.8 vs 16.8G0.9 s, 55 8C;
10.9G0.5 vs 9.4G0.5 s, P2X7C/C and P2X7
K/K, respect-
ively). Sub-analysis revealed a small but significant
difference between male P2X7C/C and male P2X7
K/K mice
at 55 8C (10.8G0.6 vs 8.2G0.6 s, P2X7C/C and P2X7
K/K,
respectively, P!0.05, Duncan’s test). No other significant
differences were found. Comparison of the response of the
genotypes to a noxious punctate stimulus (electronic Von
Frey hair; eVFH), in the absence of any induced
inflammatory insult, revealed no difference between with-
drawal latency in P2X7C/C and P2X7
K/K mice (10.7G0.5
and 10.6G0.5 g force to withdrawal, respectively, meanGs.e.mean). These data show that P2X7
K/K mice display a
largely normal nociceptive phenotype of withdrawal to both
thermal and mechanical noxious stimuli in the absence of
ongoing inflammatory or neuropathic hypersensitivity.
3.5. Pro-IL-1b gene expression
Pro-IL-1b gene expression levels in P2X7 knockout and
wild type paws were investigated using quantitative PCR.
Expression of pro-IL-1b mRNA was significantly (ANOVA
with LSD post-hoc test) higher in the P2X7K/K animals on days
0 and 1 (template copy numbers 95.5 [63.5–143.7] vs 31.5
[20.6–48.2], 10527.1 [6032.2–18371.2] vs 2387 [1590.3–
3585.0], days 0 and 1, P2X7K/K vs P2X7
C/C animals,
respectively, nZ5 per group, mean with 95% confidence
intervals), but was similar between the animal groups on
00
G6
.4
4.7
G2
71
.9
5.6
G6
4.9
*
G2
5.4
G3
.3
G4
.8
I.P. Chessell et al. / Pain 114 (2005) 386–396 391
day 7 post-FCA (1501.8 [998.9–2257.8] vs 2124.3 [1384.2–
3260.2] P2X7K/K vs P2X7
C/C animals, respectively, nZ5 per
group, mean with 95% confidence intervals).
TN
Fa
KO
wt
KO
0G
15
.31
00
G1
5.0
10
0G
3.1
1
7G
94
2.1
11
00
5.7
G1
03
0.3
99
9.4
G1
19
.01
42
5G
16
25.7
88
96
.1G
77
1.0
*1
22
4.8
G9
6.9
59
inb
old
ital
ics.
CP
-1T
NF
a
tK
Ow
tK
O
10
0G
8.5
10
0G
4.1
10
0G
12
.71
00
65
.0G
4.9
78
4.1
G6
.79
9.7
G2
.09
8.2
10
.6G
12
.08
9.7
G2
.91
05
.3G
3.0
84
.9
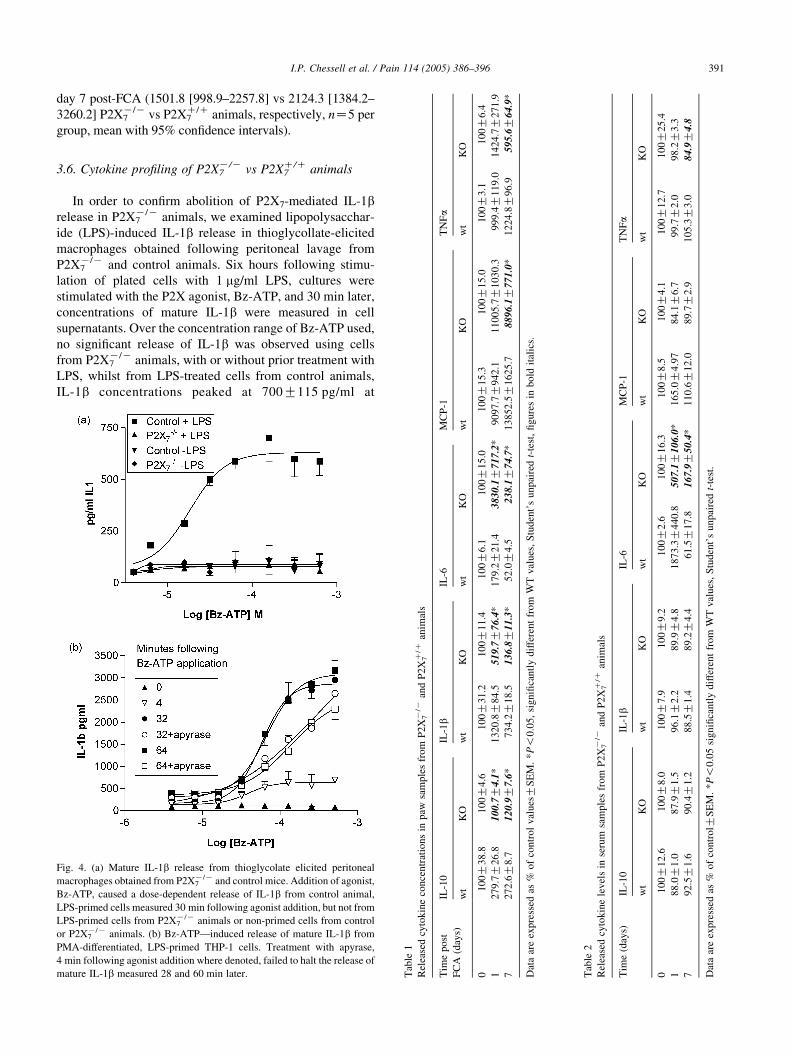
3.6. Cytokine profiling of P2X7K/K vs P2X7
C/C animals
In order to confirm abolition of P2X7-mediated IL-1brelease in P2X7
K/K animals, we examined lipopolysacchar-
ide (LPS)-induced IL-1b release in thioglycollate-elicited
macrophages obtained following peritoneal lavage from
P2X7K/K and control animals. Six hours following stimu-
lation of plated cells with 1 mg/ml LPS, cultures were
stimulated with the P2X agonist, Bz-ATP, and 30 min later,
concentrations of mature IL-1b were measured in cell
supernatants. Over the concentration range of Bz-ATP used,
no significant release of IL-1b was observed using cells
from P2X7K/K animals, with or without prior treatment with
LPS, whilst from LPS-treated cells from control animals,
IL-1b concentrations peaked at 700G115 pg/ml at
Fig. 4. (a) Mature IL-1b release from thioglycolate elicited peritoneal
macrophages obtained from P2X7K/K and control mice. Addition of agonist,
Bz-ATP, caused a dose-dependent release of IL-1b from control animal,
LPS-primed cells measured 30 min following agonist addition, but not from
LPS-primed cells from P2X7K/K animals or non-primed cells from control
or P2X7K/K animals. (b) Bz-ATP—induced release of mature IL-1b from
PMA-differentiated, LPS-primed THP-1 cells. Treatment with apyrase,
4 min following agonist addition where denoted, failed to halt the release of
mature IL-1b measured 28 and 60 min later. Tab
le1
Rel
ease
dcy
tok
ine
con
cen
trat
ion
sin
paw
sam
ple
sfr
om
P2
X7K
/Kan
dP
2X
7C/C
anim
als
Tim
ep
ost
FC
A(d
ays)
IL-1
0IL
-1b
IL-6
MC
P-1
wt
KO
wt
KO
wt
KO
wt
01
00
G3
8.8
10
0G
4.6
10
0G
31
.21
00
G1
1.4
10
0G
6.1
10
0G
15
.01
0
12
79
.7G
26
.81
00
.7G
4.1
*1
32
0.8
G8
4.5
51
9.7
G7
6.4
*1
79
.2G
21
.43
83
0.1
G7
17
.2*
90
97.
72
72
.6G
8.7
12
0.9
G7
.6*
73
4.2
G1
8.5
13
6.8
G1
1.3
*5
2.0
G4
.52
38
.1G
74
.7*
13
852
.
Dat
aar
eex
pre
ssed
as%
of
con
tro
lv
alues
GS
EM
.*
P!
0.0
5,
sig
nifi
can
tly
dif
fere
nt
fro
mW
Tv
alues
,S
tud
ent’
su
np
aire
dt-
test
,fi
gure
s
Tab
le2
Rel
ease
dcy
tok
ine
lev
els
inse
rum
sam
ple
sfr
om
P2
X7K
/Kan
dP
2X
7C/C
anim
als
Tim
e(d
ays)
IL-1
0IL
-1b
IL-6
M
wt
KO
wt
KO
wt
KO
w
01
00
G1
2.6
10
0G
8.0
10
0G
7.9
10
0G
9.2
10
0G
2.6
10
0G
16
.3
18
8.0
G1
.08
7.9
G1
.59
6.1
G2
.28
9.9
G4
.81
87
3.3
G4
40
.85
07
.1G
10
6.0
*1
79
2.5
G1
.69
0.4
G1
.28
8.5
G1
.48
9.2
G4
.46
1.5
G1
7.8
16
7.9
G5
0.4
*1
Dat
aar
eex
pre
ssed
as%
of
con
tro
lGS
EM
.*P
!0
.05
sig
nifi
can
tly
dif
fere
nt
from
WT
val
ues
,S
tud
ent’
su
np
aire
dt-
test
.

I.P. Chessell et al. / Pain 114 (2005) 386–396392
a Bz-ATP concentration of 160 mM. No significant IL-1brelease was observed from cells from normal animals which
were not treated with LPS (Fig. 4).
The natural ligand for the P2X7 receptor is ATP, and
whilst it has been determined that activation of this receptor
mediates release of IL-1b, experiments are often based on
prolonged contact of the ligand with the receptor; we wished
to confirm that a single, brief activation of the receptor
commits cells to release IL-1b at times after removal of
agonist, as this may be more representative of the pulsatile
nature of P2X7 receptor activation in vivo. Here we have
used LPS-treated, differentiated human monocytic cells,
THP-1, to examine Bz-ATP-induced IL-1b release, and the
agonist degrading enzyme, apyrase (Ferrari et al., 1997;
Grahames et al., 1999) to limit receptor activation time
(applied !4 min following agonist administration). In the
absence of apyrase addition, Bz-ATP (40 mM to 5 mM)
caused a concentration and time dependent release of IL-1b,
which reached a maximum at 32 min following Bz-ATP
addition (Fig. 4). Comparison of data obtained 4 min after
Bz-ATP addition with that obtained from cells treated with
apyrase after 4 min shows that removal of the agonist by
addition of the enzyme fails to halt the release of mature
IL-1b from the cells (Fig. 4).
In order to further elucidate a potential mechanism by
which deletion of the P2X7 gene abolishes FCA-induced
hypersensitivity, we profiled both local and systemic
released cytokine levels in P2X7K/K vs P2X7
C/C animals
0, 1 and 7 days post FCA-challenge using cytometric
analysis.
Profiling of the P2X7C/C paw samples revealed a sustained
increase of IL-1b levels in the P2X7C/C animals following
intraplantar FCA injection. This increase was significantly
reduced in the P2X7K/K animals (Table 1). Additional
cytokines upregulated following intraplanter FCA injection
included IL-10, MCP-1, TNF-a and in IL-6 (Table 1).
Interestingly, the increases in IL-10 following intraplantar
FCA injection are also significantly attenuated in the
P2X7K/K animals across the whole timecourse. In contrast,
the IL-6 response was enhanced in the P2X7K/K animals
suggesting potential compensatory changes.
Following FCA treatment, a transient increase (peaking
at 1 day post-FCA) in IL-6 and MCP-1 levels was observed
in the sera from the mice. The increase in IL-6 concentration
was significantly reduced in the P2X7K/K animals compared
to the wild-types on day 1, suggesting that this early
systemic inflammatory process following FCA-challenge is
attenuated in P2X7K/K animals. No significant increase was
observed in the levels of IL-10, TNFa or IL-1b in sera from
P2X7C/C mice post-FCA (Table 2).
3.7. P2X7 immunolocalisation in human tissue samples
To determine the significance of these findings in
human pain states, we have used a monoclonal antibody
raised to the extracellular loop of the human P2X7 receptor
(Buell et al., 1998) to examine its expression in tissues
obtained from patients suffering from persistent neuropathic
pain and hypersensitivity following trauma. In serial tissue
sections of painful injured nerves and matched control
nerves, P2X7 immunostaining (Fig. 5a) was observed and
correlated well with immunoreactivity for the glial/Schwann
cell marker, GFAP (Fig. 5b), but not with axonal marker,
neurofilament (Fig. 5c); co-localisation of P2X7 and GFAP
confirmed the presence of P2X7 in Schwann cells (Inset
Fig. 5a). Comparison of P2X7 and GFAP immunoreactivities
(P2X7:GFAP) by image analysis showed a significantly
increased ratio of P2X7:neurofilament immunostaining in
painful nerves [nZ11; median (range) 0.58 (0.26–1.78)]
compared to controls [nZ13; median (range) 0.45 (0.06–
0.58); Mann–Whitney U test; PZ0.032]. P2X7 immunos-
taining was not detected in sensory neurons of human dorsal
root ganglia (DRG) in both post-mortem controls and after
spinal root avulsion injury (i.e. central axotomy; Fig. 6a and
b), but was present in satellite cells surrounding neurons
(Inset Fig. 6a satellite cells red; neurons black), with
markedly increased density of these in the avulsed DRG
preparations. Image analysis of P2X7 immunoreactivity in
satellite cells of human DRG, expressed as a ratio to neuronal
cell body area (nZ10 control, nZ12 avulsed), showed a
significantly increased area of staining in avulsed DRG
[nZ12; median (range) 0.60 (0.17–0.76)], compared to
controls [nZ10; median (range) 0.38 (0.21–0.54); Mann–
Whitney U test; PZ0.045]. In control DRG, weak GFAP
immunostaining was observed in some fibres and cells
surrounding neurons (Fig. 6c). However, in avulsed DRG
there was a marked increase of both intensity and frequency
of GFAP-immunoreactive structures (Fig. 6d), in a very
similar fashion to that seen with P2X7. These findings in
DRG are similar to those reported in an animal model of
sciatic nerve injury, where neurotrophins such as NGF
expressed by reactive satellite cells contribute to sprouting of
sympathetic fibres. In our study, NGF immunoreactivity was
also detected in satellite cells of DRG, and appeared to be
increased after avulsion injury (data not shown). There was
no correlation between delay before surgery and P2X7
immunostaining in injured nerves or DRG. No P2X7
immunostaining was observed in human sympathetic
ganglion neurons, or in motoneurons in post-mortem
human spinal cord (data not shown).
3.8. P2X7 western blots from human tissue samples
Western blotting with the P2X7 monoclonal antibody in
nerve extracts showed a 70 kDa band in all samples
(Fig. 5d), and in some extracts a 65 kDa and larger,
approximately 140 kDa band. The relative optical density of
the 70 kDa band was significantly elevated in extracts of
painful injured nerve [median (range)Z36.0 (4.0–120)]
compared to controls [median (range)Z11.0 (6.0–34.0);
PZ0.018, Mann–Whitney U test]. There was a trend for
increased optical densities of the 70 kDa band in injured

I.P. Chessell et al. / Pain 114 (2005) 386–396 393
DRG [median (range), 18.5 (11.0–36.0)] compared to
controls [15.0 (7.0–36.0)]. The 65 kDa band may be an
unglycosylated form of the 70 kDa band and the larger
140 kDa band may be a 70 kDa dimer (Glass and Burnstock,
2001; Kim et al., 2001).
4. Discussion
These data indicate a clear effect of P2X7 gene deletion
upon responsiveness to noxious thermal or mechanical
stimuli in conditions of both chronic neuropathic and
inflammatory hypersensitivity. The gene deletion did not
affect DRG total cell numbers, the distribution of cell size,
or normal nociceptive responses to mechanical or thermal
stimuli. Given the role of the P2X7 receptor in triggering
post-translational processing of IL-1b, with the resulting
activation of an inflammatory cascade which includes
induction or release of cyclo-oxygenase 2 (Samad et al.,
2001), metalloproteinases (Flannery et al., 2003), inducible
nitric oxide synthase, superoxide products (Parvathenani
et al., 2003), and other cytokines, coupled with degenerative
changes that these mediators can induce, it appears that the
P2X7 receptor plays a pivotal role in initiating or
maintaining pathological changes as a result of inflamma-
tory or neuropathic insults. The locus of action may include
peripheral nerves, sensory neuronal cell bodies (Parvathe-
nani et al., 2003), or central pathways. Indeed, a cascade,
initiated by release of ATP following cellular perturbation
can be proposed in both the periphery and in the CNS. Thus,
in the periphery, it is likely that inflammation or a
neuropathic insult causes P2X7-dependent IL-1b release
Fig. 5. Serial sections of nerve immunostained with antibodies to P2X7 (a), GFA
GFAP (black) in Schwann cell]. (d) Representative P2X7 western blot using an an
polyacrylamide gel. Lanes 1–4 are extracts of human control nerve and lanes 5–8
from mononuclear, mast and Schwann cells (Colomar et al.,
2003), and in turn induces COX-2, nerve growth factor, and
other algogens (Woolf et al., 1996, 1997). Similarly, in the
CNS, maturation and release of IL-1b is able to induce
COX-2 (Samad et al., 2001) and other mediators, many of
which are thought to underlie increased neuronal excit-
ability, particularly in the spinal cord. ATP itself may play
a role in recruitment (Honda et al., 2001) and activation
(Verderio and Matteoli, 2001) of microglia or macrophages
which mediate plastic and degenerative changes, suggesting
that this transmitter is able to play a role in initiation of these
disease pathologies. Recent studies support initial findings
that P2X7 is expressed predominantly on cells of immune
origin (Sim et al., 2004), and functional studies suggest that
P2X7 is not expressed on neuronal structures (Kukley et al.,
2004), and thus P2X7 appears to play a critical role in glia-
neuronal signalling. The clinical relevance of these
proposals is supported by our findings of increased levels
of P2X7 receptor in injured nerve tissues from patients
suffering from chronic neuropathic pain. However, the
human tissue data needs to be interpreted with caution.
Further studies in greater numbers of patients with a range
of severity of pain symptoms, and in injured nerves from
patients who do not suffer from neuropathic pain, are
necessary to support a role for P2X7 in clinical pain
symptomatology. Moreover the lack of immunostaining in
sensory cell bodies, while consistent with current hypoth-
eses and finding, must be interpretated with caution, as this
may reflect a lack of sensitivity of the method.
In order to support the hypothesis that key actions of
activation of P2X7 are transduced via release of mature IL-1b,
we have also performed experiments in P2X7K/K animals to
P (b) or neurofilaments (c). [Inset in (a): Co-localisation of P2X7 (red) and
tibody dilution of 1:100,000 with human nerve extracts separated on a 6%
are extracts from painful injured nerve. Scale bars (a–c), 100 mm.
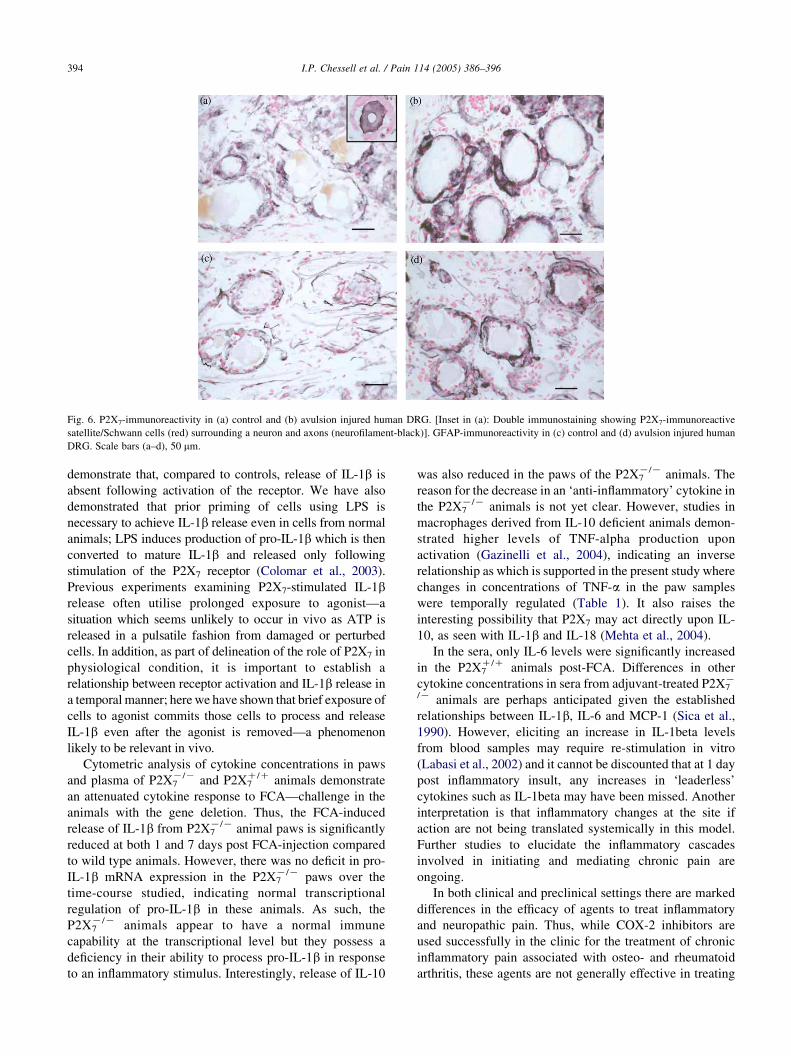
Fig. 6. P2X7-immunoreactivity in (a) control and (b) avulsion injured human DRG. [Inset in (a): Double immunostaining showing P2X7-immunoreactive
satellite/Schwann cells (red) surrounding a neuron and axons (neurofilament-black)]. GFAP-immunoreactivity in (c) control and (d) avulsion injured human
DRG. Scale bars (a–d), 50 mm.
I.P. Chessell et al. / Pain 114 (2005) 386–396394
demonstrate that, compared to controls, release of IL-1b is
absent following activation of the receptor. We have also
demonstrated that prior priming of cells using LPS is
necessary to achieve IL-1b release even in cells from normal
animals; LPS induces production of pro-IL-1b which is then
converted to mature IL-1b and released only following
stimulation of the P2X7 receptor (Colomar et al., 2003).
Previous experiments examining P2X7-stimulated IL-1brelease often utilise prolonged exposure to agonist—a
situation which seems unlikely to occur in vivo as ATP is
released in a pulsatile fashion from damaged or perturbed
cells. In addition, as part of delineation of the role of P2X7 in
physiological condition, it is important to establish a
relationship between receptor activation and IL-1b release in
a temporal manner; here we have shown that brief exposure of
cells to agonist commits those cells to process and release
IL-1b even after the agonist is removed—a phenomenon
likely to be relevant in vivo.
Cytometric analysis of cytokine concentrations in paws
and plasma of P2X7K/K and P2X7
C/C animals demonstrate
an attenuated cytokine response to FCA—challenge in the
animals with the gene deletion. Thus, the FCA-induced
release of IL-1b from P2X7K/K animal paws is significantly
reduced at both 1 and 7 days post FCA-injection compared
to wild type animals. However, there was no deficit in pro-
IL-1b mRNA expression in the P2X7K/K paws over the
time-course studied, indicating normal transcriptional
regulation of pro-IL-1b in these animals. As such, the
P2X7K/K animals appear to have a normal immune
capability at the transcriptional level but they possess a
deficiency in their ability to process pro-IL-1b in response
to an inflammatory stimulus. Interestingly, release of IL-10
was also reduced in the paws of the P2X7K/K animals. The
reason for the decrease in an ‘anti-inflammatory’ cytokine in
the P2X7K/K animals is not yet clear. However, studies in
macrophages derived from IL-10 deficient animals demon-
strated higher levels of TNF-alpha production upon
activation (Gazinelli et al., 2004), indicating an inverse
relationship as which is supported in the present study where
changes in concentrations of TNF-a in the paw samples
were temporally regulated (Table 1). It also raises the
interesting possibility that P2X7 may act directly upon IL-
10, as seen with IL-1b and IL-18 (Mehta et al., 2004).
In the sera, only IL-6 levels were significantly increased
in the P2X7C/C animals post-FCA. Differences in other
cytokine concentrations in sera from adjuvant-treated P2X7K
/K animals are perhaps anticipated given the established
relationships between IL-1b, IL-6 and MCP-1 (Sica et al.,
1990). However, eliciting an increase in IL-1beta levels
from blood samples may require re-stimulation in vitro
(Labasi et al., 2002) and it cannot be discounted that at 1 day
post inflammatory insult, any increases in ‘leaderless’
cytokines such as IL-1beta may have been missed. Another
interpretation is that inflammatory changes at the site if
action are not being translated systemically in this model.
Further studies to elucidate the inflammatory cascades
involved in initiating and mediating chronic pain are
ongoing.
In both clinical and preclinical settings there are marked
differences in the efficacy of agents to treat inflammatory
and neuropathic pain. Thus, while COX-2 inhibitors are
used successfully in the clinic for the treatment of chronic
inflammatory pain associated with osteo- and rheumatoid
arthritis, these agents are not generally effective in treating

I.P. Chessell et al. / Pain 114 (2005) 386–396 395
clinical neuropathic pain (Wallace, 2001). Similarly, drugs
used for the treatment of neuropathic pain, such as anti-
epileptic drugs and antidepressants, are ineffective in
adequately treating inflammatory pain (Jain, 2000).
Opioids, widely regarded as the most powerful analgesic
agents available, are effective in a broad range of pain states,
but chronic opiate use is associated with tolerability, side-
effect and addiction issues, which limits their use. Whilst
deletion of genes in animals provides important phenotypic
information, the possibility cannot be discounted that
compensatory changes, caused by persistent absence of a
receptor, may contribute to the phenotype in a way that
antagonism of the receptor after completion of development
may not. Efficacy of small molecule antagonists of P2X7
must be examined in these models of pain. However, the
data reported here strongly suggests that therapies based on
antagonism of this receptor may be capable of producing
broad-spectrum analgesia.
Acknowledgements
We thank Paul Facer for help with immunohistological
studies, Mr David Elliot for providing some of the nerve
specimens, and Ms Sarah Friel for assistance with the DRG
cell analysis.
References
Alcarez L, Baxter A, Bent J, Bowers K, Braddock M, Cladingboel D,
Donald D, Fagura M, Furber M, Laurent C, Lawson M, Martimore M,
McCormick M, Roberts N, Robertson M. Novel P2X7 receptor
antagonists. Bioorg Med Chem Lett 2004;13:4043–6.
Aley KO, McCarter G, Levine JD. Nitric oxide signaling in pain and
nociceptor sensitization in the rat. J Neurosci 1998;18:7008–14.
Budzinski M, Misterek K, Gumulka W, Dorociak A. Inhibition of inducible
nitric oxide synthase in persistant pain. Life Sci 2000;66:301–5.
Buell G, Chessell IP, Michel AD, Collo G, Salazzo M, Herren S,
Gretener D, Grahames CBA, Kaur R, Kosco-Vilbois M,
Humphrey PPA. Blockade of human P2X7 receptor function with a
monoclonal antibody. Blood 1998;92:3521–8.
Chessell IP, Simon J, Hibell AD, Michel AD, Barnard EA, Humphrey PPA.
Cloning and functional characterisation of the mouse P2X7 receptor.
Fed Eur Biolchem Soc Lett 1998;439:26–30.
Clark JD. Chronic pain prevalence and analgesic prescribing in a general
medical population. J Pain Sympt Manage 2003;23:131–7.
Clayton NM, Oakley I, Thompson S, Wheeldon A, Sargent RS, Bountra C.
Validation of the dual channel weight averager as an instrument for the
measurement of inflammatory pain. Br J Pharmacol 1997;120:219.
Collo G, Neidhart S, Kawashima E, Kosco-Vilbois M, North RA, Buell G.
Tissue distribution of the P2X7 receptor. Neuropharmacology 1997;36:
1277–83.
Colomar A, Marty V, Medina C, Combe C, Parnet P, Amedee T.
Maturation and release of interleukin-1b by lipopolysaccharide-primed
mouse Schwann cells require the stimulation of P2X7 receptors. J Biol
Chem 2003;278:30732–40.
Conquet F. Inactivation in vivo of metabotropic glutamate receptor 1 by
specific chromosomal insertion of a reported gene lacZ. Neurophar-
macology 1995;34:865–70.
Ferrari D, Chiozzi P, Falzoni S, Hanau S, Di Virgilio F. Purinergic
modulation of interleukin-1-beta release from microglial cells stimu-
lated with bacterial endotoxin. J Exp Med 1997;185:579–82.
Flannery CR, Little CB, Caterson B, Hughes CE. Effects of culture
conditions and exposure to catabolic stimulators (IL-1 and retinoic acid)
on the expression of matrix metalloproteinases (MMPs) and disintegrin
metalloproteinases (ADAMs) by articular cartilage chondrocytes.
Matrix Biol 2003;18:225–37.
Gazinelli RT, Wysocka M, Hieny S, Scharton-Kersten T, Cheever A, Kuhn R,
Muller W, Trinchieri G, Sher A. In the absence of endogenous IL-10, mice
acutely infected with Toxoplasma gondii succumb to a lethal immune
response dependent onCD4C T cells and accompanied by overproduction
of IL-12, IFN-gamma and TNF-alpha. J Immunol 2004;157:798–805.
Glass R, Burnstock G. Immunohistological identification of cells expres-
sing ATP-gated cation channels (P2X receptors) in the adult rat thyroid.
J Anat 2001;198:569–79.
Grahames CBA, Michel AD, Chessell IP, Humphrey PPA. Pharmacologi-
cal characterization of ATP- and LPS-induced IL-1b release in human
monocytes. Br J Pharmacol 1999;127:1915–21.
Honda S, Sasaki Y, Oshawa K, Imai Y, Nakamura Y, Inoue K, Kohsaka S.
Extracellular ATP or ADO induce chemotaxis of cultured microglia
through Gi/o-coupled P2Y receptors. J Neurosci 2001;21:1975–82.
Jain KK. A guide to drug evaluation for chronic pain. Emerg Drugs 2000;5:
241–57.
Kim M, Spelta V, North RA, Surprenant A. Differential assembly of rat
purinergic P2X7 receptor in immune cells of the brain and periphery.
J Biol Chem 2001;276:23232–67.
Kukley M, Stausberg P, Adelmann G, Chessell IP, Dietrich D. Ectonu-
cleotidases and nucleoside transporters mediate activation of adenosine
receptors by P2X7 receptor agonist 2 0-30-O-(4-benzoylbenzoyl)-ATP.
J Neurosci 2004;24:7128–39.
Labasi JM, Petrushova N, Donovan C, McCurdy S, Lira P, Payette MM,
Brissette W, Wicks JR, Audoly L, Gabel CA. Absence of the P2X7
receptor alters leukocyte function and attenuates an inflammatory
response. J Immunol 2002;168:6436–45.
Le Mouellic H, Lallemand Y, Brulet P. Targeted replacement of the
homeobox gene hox-3.1 by the escherichia-coli lacZ in mouse chimeric
embryos. Proc Natl Acad Sci USA 1990;87:4712–6.
MacDonald R, Bingham S, Bond BC, Parsons AA, Philpott KL. Determi-
nation of changes in mRNA expression in a rat model of neuropathic pain
by Taqman quantitative RT-PCR. Brain Res 2001;90:48–56.
Mehta VB, Hart J, Wewers MD. ATP-stimulated release of interleukin
(IL)-1beta and IL-18 requires priming by lipopolysaccharide and is
independent of caspase-1 cleavage. J Biol Chem 2004;276:3820–6.
MichelAD,Chessell IP,HumphreyPPA. Ioniceffectsonhumanrecombinant
P2X7 receptor function. N-S Arch Pharmacol 1999;359:102–9.
Parvathenani LK, Tertyshnikova S, Greco CR, Roberts SB, Robertson B,
Posmantur R. P2X7 mediates superoxide production in primary
microglia and is up-regulated in a transgenic mouse model of
Alzheimer’s disease. J Biol Chem 2003;278:13309–17.
Perregaux DG, Gabel CA. Interleukin-1b maturation and release in
response to ATP and nigericin. J Biol Chem 1994;269:15195–203.
Samad TA, Moore KA, Sapirstein A, Billet S, Allchorne A, Poole S,
Bonventre JV, Woolf CJ. Interleukin-1b-mediated induction of Cox-2
in the CNS contributes to inflammatory pain hypersensitivity. Nature
2001;410:471–5.
Seltzer Z, Dubner R, Shir Y. A novel behavioral model of neuropathic pain
disordersproducedinratsbypartial sciaticnerve injury.Pain2000;43:205–18.
Shu SY, Ju G, Fan LZ. The glucose oxidase-DAB-nickel method in
peroxidase histochemistry of the nervous system. Neurosci Lett 1988;
85:169–71.
Sica A, Wang JM, Colotta F, Dejana E, Mantovani A, Oppenheim JJ,
Larsen CG, Zachariae CO, Matsushima K. Monocyte chemotactic and
activating factor gene expression induced in endothelial cells by IL-1
and tumor necrosis factor. J Immunol 1990;144:3034–8.
Sim JA, Young MT, Sung H-Y, North RA, Surprenant A. Reanalysis of
P2X7 receptor expression in rodent brain. J Neurosci 2004;24:6307–14.

I.P. Chessell et al. / Pain 114 (2005) 386–396396
Solle M, Labasi J, Perregaux DG, Stam E, Petrushova N, Koller BH,
Griffiths RJ, Gabel CA. Altered cytokine production in mice lacking
P2X7 receptors. J Biol Chem 2001;276:125–32.
Steinberg TH, Newman AS, Swanson JA, Silverstein SC. ATP4K
permeabilizes the plasma membrane of mouse macrophages to
flourescent dyes. J Biol Chem 1987;262:8884–8.
Surprenant A, Rassendren F, Kawashima E, North RA, Buell G. The
cytolytic P2Z receptor for extracellular ATP identified as a P2X receptor
(P2X7). Science 1996;272:735–8.
Verderio C, Matteoli M. ATP mediates calcium signalling between
astrocytes and microglia cells: modulation by IFN-g. J Immunol
2001;166:6383–91.
Wallace MS. Emerging drugs for neuropathic pain. Expert Opin Emerg
Drugs 2001;6:249–59.
Woolf CJ, Ma Q, Allchorne A, Poole S. Peripheral cell types contributing to
the hyperalgesic action of nerve growth factor in inflammation.
J Neurosci 1996;16:2716–23.
Woolf CJ, Allchorne A, Safieh-Garabedian B, Poole S. Cytokines,
nerve growth factor and inflammatory hyperlagesia: the
contribution of tumour necrosis factor a. Br J Pharmacol 1997;
121:417–24.
Yiangou Y, Facer P, Dyer NH, Chan CL, Knowles C, Williams NS,
Anand P. Vannilloid receptor 1 immunoreactivity in inflammed human
bowel. Lancet 2001;357:1338–9.




















