
Mitochondrial dysfunction enhances Gal4-dependent transcription
7
Mitochondrial dysfunction enhances Gal4-dependent transcription Branka Jelic ˇic ´ 1 , Ana Traven 1,2 , Vedrana Filic ´, Mary Sopta * Department of Molecular Biology, Rud - er Bos ˇkovic ´ Institute, Bijenic ˇka 54, 10000 Zagreb, Croatia Received 4 April 2005; received in revised form 20 September 2005; accepted 23 September 2005 First published online 7 October 2005 Edited by M. Jacquet Abstract Mitochondrial dysfunction has been shown to elicit broad effects on nuclear gene expression. We show here that transcription dependent on the prototypical acidic activator Gal4 is responsive to mitochondrial dysfunction. In cells with no mitochondrial DNA, Gal4-dependent gene expression is elevated. A minimal Gal4 activator containing the DNA binding and activation domain is sufficient for this response. Transcription dependent on a fusion of Gal4 to a heterologous DNA binding domain is similarly ele- vated in a mitochondrial mutant. Analysis of different Gal4-dependent promoters and gel mobility shift assays suggest that the effect of mitochondrial dysfunction on Gal4 activity is related to increased DNA binding to the cognate Gal4 element. Given that fermen- tation is the only means to obtain energy in respiratory deficient cells, it is possible that higher Gal4 activity in cells with dysfunc- tional mitochondria works to promote more efficient fermentation of galactose. Ó 2005 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved. Keywords: Yeast; Mitochondrial dysfunction; Gal4; Transcription 1. Introduction Regulation of GAL gene expression in Saccharomyces cerevisiae is a paradigm for regulation of transcription by metabolic signals, reviewed in [1,2]. The protein products of the GAL genes are required for efficient galactose utilization and they are repressed by glucose and strongly activated by galactose. Transcription of the GAL genes is under the control of Gal4, a prototyp- ical acidic activator, reviewed in [1,2]. When galactose is absent, the activity of Gal4 is repressed by Gal80, which binds to the activation domain of Gal4 thereby prevent- ing its interactions with other components of the tran- scription machinery [3,4]. The galactose signal is transduced to the inactive Gal4–Gal80 complex by the inducer protein Gal3, which, in the presence of galac- tose, binds to Gal80 and relieves its inhibition of Gal4 [5–9]. In addition to being negatively regulated by Gal80, Gal4 is positively regulated by phosphorylation [10–13]. More recently it has been shown that regulation of Gal4 protein stability and transcriptional activation also occurs via the ubiquitin degradation system [14]. Mitochondria are the central metabolic organelle in the cell and their functional state is directly involved in regulation of transcription factors and gene expres- sion. Microarray analysis of whole genome expression in wild type versus mitochondrial mutants has shown a variety of coordinate changes in response to mitochon- drial dysfunction [15,16]. The best studied transcrip- tional response to loss of mitochondrial function is the retrograde signaling pathway, which is activated by 0378-1097/$22.00 Ó 2005 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.femsle.2005.09.033 * Corresponding author. Tel.: +385 1 4560 948; fax: +385 1 4561 177. E-mail address: [email protected] (M. Sopta). 1 These authors contributed equally to this work. 2 Present address: St. VincentÕs Institute, 9 Princes Street, Fitzroy, Vic. 3065, Australia. www.fems-microbiology.org FEMS Microbiology Letters 253 (2005) 207–213
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Mitochondrial dysfunction enhances Gal4-dependent transcription

www.fems-microbiology.org
FEMS Microbiology Letters 253 (2005) 207–213
Mitochondrial dysfunction enhances Gal4-dependent transcription
Branka Jelicic 1, Ana Traven 1,2, Vedrana Filic, Mary Sopta *
Department of Molecular Biology, Rud-er Boskovic Institute, Bijenicka 54, 10000 Zagreb, Croatia
Received 4 April 2005; received in revised form 20 September 2005; accepted 23 September 2005
First published online 7 October 2005
Edited by M. Jacquet
Abstract
Mitochondrial dysfunction has been shown to elicit broad effects on nuclear gene expression. We show here that transcriptiondependent on the prototypical acidic activator Gal4 is responsive to mitochondrial dysfunction. In cells with no mitochondrialDNA, Gal4-dependent gene expression is elevated. A minimal Gal4 activator containing the DNA binding and activation domainis sufficient for this response. Transcription dependent on a fusion of Gal4 to a heterologous DNA binding domain is similarly ele-vated in a mitochondrial mutant. Analysis of different Gal4-dependent promoters and gel mobility shift assays suggest that the effectof mitochondrial dysfunction on Gal4 activity is related to increased DNA binding to the cognate Gal4 element. Given that fermen-tation is the only means to obtain energy in respiratory deficient cells, it is possible that higher Gal4 activity in cells with dysfunc-tional mitochondria works to promote more efficient fermentation of galactose.� 2005 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved.
Keywords: Yeast; Mitochondrial dysfunction; Gal4; Transcription
1. Introduction
Regulation of GAL gene expression in Saccharomyces
cerevisiae is a paradigm for regulation of transcriptionby metabolic signals, reviewed in [1,2]. The proteinproducts of the GAL genes are required for efficientgalactose utilization and they are repressed by glucoseand strongly activated by galactose. Transcription ofthe GAL genes is under the control of Gal4, a prototyp-ical acidic activator, reviewed in [1,2]. When galactose isabsent, the activity of Gal4 is repressed by Gal80, whichbinds to the activation domain of Gal4 thereby prevent-
0378-1097/$22.00 � 2005 Federation of European Microbiological Societies
doi:10.1016/j.femsle.2005.09.033
* Corresponding author. Tel.: +385 1 4560 948; fax: +385 1 4561177.
E-mail address: [email protected] (M. Sopta).1 These authors contributed equally to this work.2 Present address: St. Vincent�s Institute, 9 Princes Street, Fitzroy,
Vic. 3065, Australia.
ing its interactions with other components of the tran-scription machinery [3,4]. The galactose signal istransduced to the inactive Gal4–Gal80 complex by theinducer protein Gal3, which, in the presence of galac-tose, binds to Gal80 and relieves its inhibition of Gal4[5–9]. In addition to being negatively regulated byGal80, Gal4 is positively regulated by phosphorylation[10–13]. More recently it has been shown that regulationof Gal4 protein stability and transcriptional activationalso occurs via the ubiquitin degradation system [14].
Mitochondria are the central metabolic organelle inthe cell and their functional state is directly involvedin regulation of transcription factors and gene expres-sion. Microarray analysis of whole genome expressionin wild type versus mitochondrial mutants has showna variety of coordinate changes in response to mitochon-drial dysfunction [15,16]. The best studied transcrip-tional response to loss of mitochondrial function is theretrograde signaling pathway, which is activated by
. Published by Elsevier B.V. All rights reserved.

208 B. Jelicic et al. / FEMS Microbiology Letters 253 (2005) 207–213
mitochondrial dysfunction and positively regulates thetranscription factors Rtg1 and Rtg3, whose target genesare required for the metabolic adaptation of the cell torespiratory deficiency [15,17–19]. Additionally, mito-chondrial dysfunction has been shown to change tran-scription of the rRNA genes and genes encodingenzymes of the TCA cycle [20,21], convert the corepres-sor Tup1/Ssn6 to a coactivator of transcription [22] andincrease the activity of Pdr3, a yeast transcription factorinvolved in pleiotropic drug resistance [23,24].
In this report, we were interested in analyzing moredirectly the influence of changes in mitochondrial statuson transcription factor activity. To that end, we chose toexamine the yeast Gal4 activator, since transcriptionalmechanisms of Gal4 action are well studied and it is asystem that is responsive to changes in metabolicrequirements.
2. Materials and methods
2.1. Yeast strains and plasmids
The yeast strains used for assaying b and a-galactosi-dase activity and electrophoretic mobility shifts areW3031B (MATa ade2-1 trp1 leu2-3 112 his3-11 15
ura3-1), GGY1 (Dgal4 Dgal80 tyr1 ade leu2 his3 ura3)and NLY2 (Dgal4 Dgal80 ura3 his3 leu2 trp1 lys2). Mu-tants that lack mitochondrial DNA (rho0) were inducedby treatment with ethidium bromide, as described [25]and respiratory deficiency was confirmed by their inabil-ity to use glycerol as the sole carbon source. ReportersSV15 [26] and RJR227 [4] were integrated into theURA3 locus of GGY1. The reporter pSH18-34 [27]was transformed into the strain W3031B and its respec-tive rho0 mutant. Plasmids expressing full length Gal4(amino acids 1–881), or its deletion derivative contain-ing the DNA binding domain (amino acids 1–100) fusedto the activation domain (amino acids 840–881) are de-scribed in [4]. The plasmids are HIS3, CEN/ARS andGal4 and its derivatives are expressed from the naturalGAL4 promoter. Lex–Gal4 [27] was expressed fromLEU2, CEN/ARS plasmids that contain LexA residues1–202 fused to Gal4 (amino acids 74–881). For EMSAexperiments a plasmid expressing short Gal4 under thecontrol of the natural Gal4 promoter was used [4].
2.2. b-Galactosidase assays
For assaying b-galactosidase activities, yeast strainswere transformed and grown in selective 2% galactose,2% sucrose media to logarithmic phase. Specific b-galac-tosidase activities were determined from yeast culturesby the method of [28]. The assays were performed in0.5 ml of reaction buffer and b-galactosidase units werecalculated as described [28]. The assay with Lex–Gal4 in
the rho0 mutant was performed in triplicate from twoindependent transformants. The standard deviationwas <20%, except for the full length Gal4 in rho+ cellsin Fig. 2(a), which was 22%. The activities are expressedas percentages of Gal4 activity in the wild type (rho+)strain, which is set to 100%.
2.3. Protein extracts and a-galactosidase assays
Cells from 10 ml of selective 2% galactose, 2% sucrosemedia were harvested at OD600 = 1.0–1.4 and resus-pended in 250 ll of breaking buffer (20 mM Tris–Cl,pH 8, 0.1 mM DTT, 10% glycerol). After addition ofan equal volume of glass beads and 12.5 ll of 40 mMphenylmethanesulfonyl fluoride (PMSF), samples werevigorously vortexed (6 · 30 s). Extracts were cleared bycentrifugation (15 min at 16,000 g, 4 �C) and 5–20 llof supernatants were used for a-galactosidase assaysperformed as described in [29]. Units were calculatedas: a-galactosidase units = 1000 · OD400/(0.0182 · lgof proteins · time (min)). All assays were performed intriplicate from at least four independent colonies andthe standard deviation was between 2% and 18%. Theactivity of the Gal4 construct in the wild type (rho+)strain is set to 100% and the other activities are ex-pressed as percentages compared to the 100% activityin rho+ cells.
2.4. Northern analysis
Poly A+ mRNA was isolated from yeast cells (NLY2harbouring a plasmid expressing full length Gal4) usingthe Qiagen Oligotex kit according to the manufacturer�sprotocol. 500 ng poly A+ mRNA was analyzed byNorthern blotting as described [30]. A PCR fragmentof the Gal4 gene was used as probe, generated withthe following primers, 5 0-CATGGCATCATTGAAA-CAGC-3 0 and 5 0-CAGGCAAAATATGGGGTGAC-3 0. Northern blot analysis of the ACT1 transcript (aPCR fragment used as a probe was generated with fol-lowing primers: 5 0-TTTCAACGTTCCAGCCTTCT-3 0
and 5 0-TTGGTCAATACCGGCAGATT-3 0) served asa loading control. Probes were labeled with [a-32P]dCTP using the Random primed DNA labeling kit(Roche). Unincorporated nucleotides were removedusing Nick columns (Amersham Biosciences).
2.5. Western blot analysis
Protein extracts were prepared as described previ-ously [31]. Western blots were performed as describedpreviously [30]. 100 lg of protein extract preparationwas analyzed. Proteins were resolved on a 7.5% SDS–polyacrylamide gel and transferred to a nitrocellulosemembrane. The nitrocellulose membrane was incubatedwith a 1:10,000 dilution of anti-lexA monoclonal anti-

B. Jelicic et al. / FEMS Microbiology Letters 253 (2005) 207–213 209
body (Santa Cruz Biotechnology), followed by a 1:5000dilution of goat anti-mouse horseradish peroxidase-conjugated secondary antibody (Novagen). Signals weredetected by enhanced chemiluminescence as per themanufacturer�s protocol (Pierce).
2.6. Electrophoretic mobility shift assays
Protein extracts for EMSA, from GGY1 transformedwith short Gal4 (1–100 + 840–881), were made in thesame way as for a-galactosidase assays except for thedifference in the breaking buffer (100 mM Tris-Cl, pH8.0, 1 mM DTT and 20% glycerol). A double-stranded32P-labeled oligonucleotide (5 0-TCCGGAGGACTGT-CCTCCGGT-3 0) containing the consensus Gal4 site [4]was used as a probe. The same unlabeled oligo was usedas a control cold competitor. A mutant oligo (5 0-TCCA-GAGGACTGTCCTCTGG-3 0) from [32] was also usedas a control competitor.
Gel electrophoretic mobility shift assays (EMSA)reactions were set up in 20 ll with 18 lg of protein ex-tract, 20 fmol of an end-labelled probe in a 2· buffercontaining 20 mM HEPES, pH 7.5, 300 mM NaCl,1 mM MgCl2, 0.1 mM ZnCl2, 10% glycerol and 200lg/ml bovine serum albumin. Binding reactions wereincubated for 10–20 min at 4 �C and resolved at 180Vfor 2 h on a native pre-run 4% polyacrylamide gel(29:1) containing 0.5· TBE and 1% glycerol. The gelwas dried and subjected to autoradiography.
3. Results
To address whether the functional state of mitochon-dria influences transcriptional activation by Gal4, the re-porter plasmid SV15 was integrated into the URA3
locus of wild type (rho+) or the rho0 mitochondrial mu-tant of GGY1. As shown in Fig. 1(a), the activity ofGal4 was approximately 2-fold higher in the mitochon-drial mutant compared to the wild-type strain.
The DNA binding and activation domains of Gal4are sufficient for regulation of this activator by a numberof signals [33]. We, therefore, analyzed whether theywere also sufficient for regulation of Gal4 by mitochon-drial dysfunction. As shown in Fig. 1a, the expression oflacZ in cells transformed with a plasmid expressing theDNA binding domain of Gal4 fused to the minimal acti-vation domain (amino acids 1–100 + 840–881) was ele-vated in the rho0 mitochondrial mutant as comparedto respiratory proficient cells (rho+). The constructexpressing just the DNA binding domain (amino acids1–100) showed no activity in either strain (data notshown).
We next tested whether this effect on Gal4 activitycould be seen using the endogenous MEL1 gene as areporter, where a single weak Gal4 binding site in the
promoter determines Gal4-dependent transcription. Forthose experiments a-galactosidase activity was assayedin strain GGY1 transformed with plasmids expressingdifferent versions of Gal4. As shown in Fig. 1b, both fulllength Gal4 (1–881) or the version containing just theDNA binding and activation domains (1–100 + 840–881) showed higher activity in rho0 compared to rho+
cells (1.7- and 2.7-fold, respectively). The control con-struct with the DNA binding domain alone (residues1–100) again exhibited no activity in either strain (datanot shown). This effect was observed when Gal4 wasexpressed either from its own natural promoter orexpressed from the heterologous ACT1 promoter (datanot shown).
To determine whether this upregulation was a func-tion of increased transcription from the Gal4 promoteritself we performed Northern blot analysis examiningthe level of Gal4 transcript. Fig. 1c shows that the levelof Gal4 transcript is unchanged in rho0 versus wild-typecells. We also tested for the ability of mitochondrial dys-function to affect Gal4-dependent transcription from areporter with multiple high affinity sites (reporterRJR227 with 5X 17 bp consensus Gal4 binding sites).No difference in transcriptional activity between rho+
and rho0 strains was observed for either the full lengthor short version of Gal4 (Table 1). In contrast, bothGal4 constructs showed higher activity in the mitochon-drial mutant when just one strong consensus binding sitewas present in the UAS (reporter SV15, Fig. 1a) or whentranscription of the MEL1 gene was assayed, which hasa single weak Gal4 binding site (Fig. 1b). Thus, the pres-ence of five high affinity consensus binding sites abro-gates the effect of mitochondrial dysfunction on Gal4transcription.
To test whether the Gal4 response to mitochondrialdysfunction is mediated via the activation domain orthe DNA binding domain, we used a lex-Gal4 fusionconstruct to assay transcriptional activity. As seen inFig. 2a the heterologous Gal4 fusion construct wasresponsive to mitochondrial dysfunction indicating thatthe response is specifically mediated through the Gal4activation domain. Western blot analysis showed thatthe protein levels were equivalent in the wild type andrho0 backgrounds (Fig. 2b).
Since experiments with reporter constructs with dif-ferent UAS structure indicated that mitochondrial dys-function could regulate Gal4 activity by influencingoccupancy of its binding sites, we sought to determineif Gal4 DNA binding activity increases in respiratorydeficient cells. We used gel electrophoretic mobility shiftassays with a 32P-labeled consensus Gal4 DNA bindingsite as a probe to examine the amount of DNA bindingactivity in wild type and rho0 strain backgrounds. ShortGal4 protein was expressed from its own natural pro-moter. As shown in Fig. 3, we observed that Gal4 bind-ing activity was increased in the rho0 strain relative to
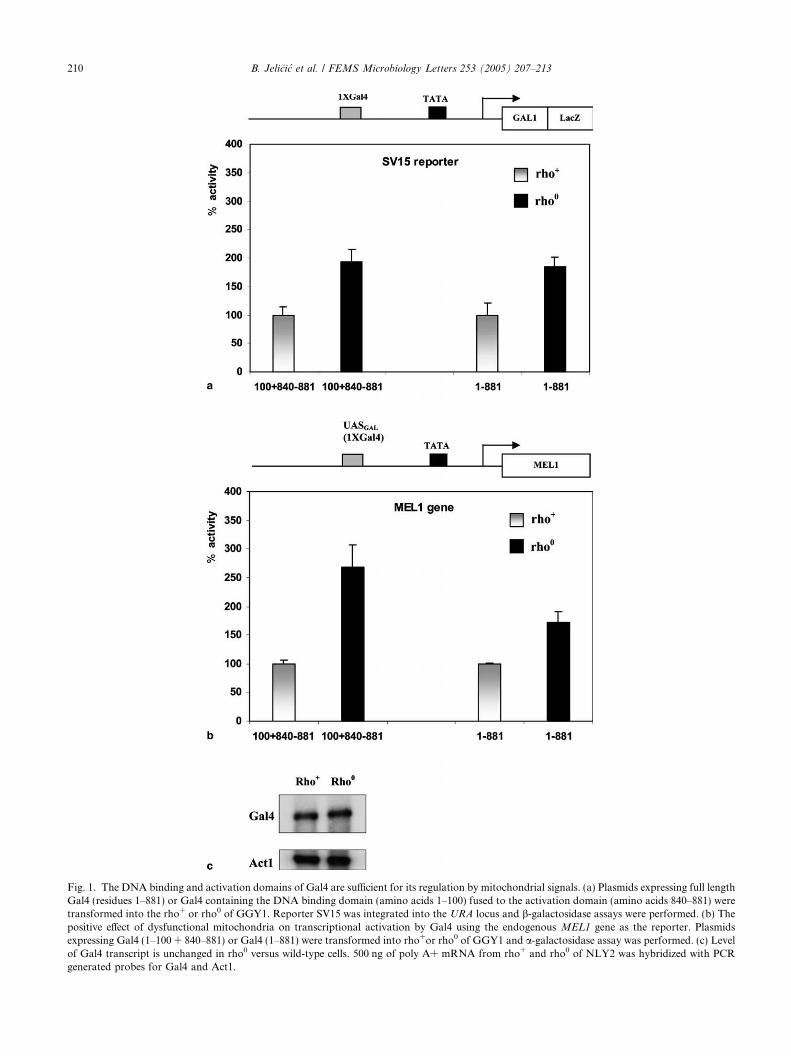
Fig. 1. The DNA binding and activation domains of Gal4 are sufficient for its regulation by mitochondrial signals. (a) Plasmids expressing full lengthGal4 (residues 1–881) or Gal4 containing the DNA binding domain (amino acids 1–100) fused to the activation domain (amino acids 840–881) weretransformed into the rho+ or rho0 of GGY1. Reporter SV15 was integrated into the URA locus and b-galactosidase assays were performed. (b) Thepositive effect of dysfunctional mitochondria on transcriptional activation by Gal4 using the endogenous MEL1 gene as the reporter. Plasmidsexpressing Gal4 (1–100 + 840–881) or Gal4 (1–881) were transformed into rho+or rho0 of GGY1 and a-galactosidase assay was performed. (c) Levelof Gal4 transcript is unchanged in rho0 versus wild-type cells. 500 ng of poly A+ mRNA from rho+ and rho0 of NLY2 was hybridized with PCRgenerated probes for Gal4 and Act1.
210 B. Jelicic et al. / FEMS Microbiology Letters 253 (2005) 207–213
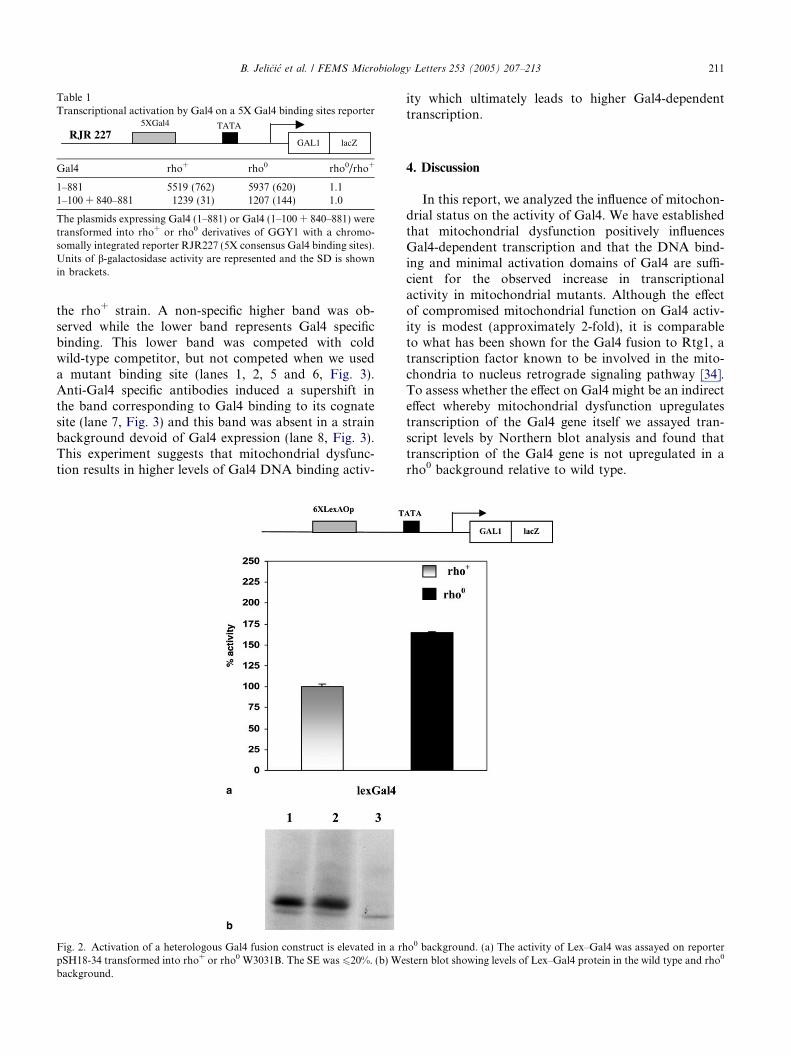
Table 1Transcriptional activation by Gal4 on a 5X Gal4 binding sites reporter
RJR 227 lacZGAL1
TATA5XGal4
Gal4 rho+ rho0 rho0/rho+
1–881 5519 (762) 5937 (620) 1.11–100 + 840–881 1239 (31) 1207 (144) 1.0
The plasmids expressing Gal4 (1–881) or Gal4 (1–100 + 840–881) weretransformed into rho+ or rho0 derivatives of GGY1 with a chromo-somally integrated reporter RJR227 (5X consensus Gal4 binding sites).Units of b-galactosidase activity are represented and the SD is shownin brackets.
B. Jelicic et al. / FEMS Microbiology Letters 253 (2005) 207–213 211
the rho+ strain. A non-specific higher band was ob-served while the lower band represents Gal4 specificbinding. This lower band was competed with coldwild-type competitor, but not competed when we useda mutant binding site (lanes 1, 2, 5 and 6, Fig. 3).Anti-Gal4 specific antibodies induced a supershift inthe band corresponding to Gal4 binding to its cognatesite (lane 7, Fig. 3) and this band was absent in a strainbackground devoid of Gal4 expression (lane 8, Fig. 3).This experiment suggests that mitochondrial dysfunc-tion results in higher levels of Gal4 DNA binding activ-
Fig. 2. Activation of a heterologous Gal4 fusion construct is elevated in a rhpSH18-34 transformed into rho+ or rho0 W3031B. The SE was 620%. (b) Webackground.
ity which ultimately leads to higher Gal4-dependenttranscription.
4. Discussion
In this report, we analyzed the influence of mitochon-drial status on the activity of Gal4. We have establishedthat mitochondrial dysfunction positively influencesGal4-dependent transcription and that the DNA bind-ing and minimal activation domains of Gal4 are suffi-cient for the observed increase in transcriptionalactivity in mitochondrial mutants. Although the effectof compromised mitochondrial function on Gal4 activ-ity is modest (approximately 2-fold), it is comparableto what has been shown for the Gal4 fusion to Rtg1, atranscription factor known to be involved in the mito-chondria to nucleus retrograde signaling pathway [34].To assess whether the effect on Gal4 might be an indirecteffect whereby mitochondrial dysfunction upregulatestranscription of the Gal4 gene itself we assayed tran-script levels by Northern blot analysis and found thattranscription of the Gal4 gene is not upregulated in arho0 background relative to wild type.
o0 background. (a) The activity of Lex–Gal4 was assayed on reporterstern blot showing levels of Lex–Gal4 protein in the wild type and rho0

Fig. 3. Mobility shift assay with short Gal4 (100 + 840–881). In eachlane 18 lg of protein extract from GGY1 transformed with short Gal4(1–100 + 840-881) was incubated with a 32P-labeled Gal4 consensusbinding site and Gal4–DNA complexes were resolved on a 4%polyacrilamide gel. Lanes are as follows: 1 – rho+ cells + cold mutantcompetitor, 2 – rho+ cells + cold wild-type competitor, 3 – rho+ cells, 4– rho0 cells, 5 – rho0 cells + cold wild-type competitor, 6 – rho0
cells + cold mutant competitor, 7 – rho0 cells + Gal4 antibody, 8 –negative control (strain lacking Gal4). The arrows indicate the positionof the Gal4–DNA complex.
212 B. Jelicic et al. / FEMS Microbiology Letters 253 (2005) 207–213
The effect of dysfunctional mitochondria on Gal4activity appears to depend on the number of bindingsites in the UAS: when reporters that have one Gal4binding site (a strong consensus site, as in SV15 or aweak site as in the UAS of MEL1) were assayed,Gal4-dependent transcription was higher in the mito-chondrial mutant relative to wild type. However, this ef-fect was not seen when Gal4 activity was assayed with areporter containing five strong consensus Gal4 bindingsites. We interpret this to mean that mitochondrial dys-function increases Gal4-dependent transcription by reg-ulating occupancy of the UAS by this activator. It hasbeen shown that the degree to which Gal4 activates tran-scription depends on the degree of occupancy of itsbinding sites and one consensus binding site or the weaksite from the MEL1 UAS are not completely saturatedin vivo [4,35]. Mitochondrial dysfunction might act toincrease binding site occupancy by Gal4 resulting inmore efficient activation. When multiple strong consen-sus sites are present (as in RJR227) Gal4 binds cooper-atively and saturates the UAS, resulting in maximalactivity in rho+ cells that cannot be further increasedby respiratory deficiency. This conclusion is supportedby our EMSA results, which show that the level ofGal4 DNA binding activity is higher in rho0 cells. Alter-
natively, increased DNA binding to the Gal4 consensussite as measured by EMSA may reflect an increase inGal4 protein levels. It remains to be determined whetherthe signaling of mitochondrial dysfunction to Gal4 in-volves post-translational events such as phosphorylationor ubiquitination which could potentially regulate pro-tein stability or DNA binding.
We also tested the ability of a fusion of the Gal4 acti-vation domain with the bacterial lexA DNA binding do-main for responsiveness to mitochondrial dysfunction.We observed that such a construct is responsive to mito-chondrial dysfunction indicating that the signaling path-way acts specifically via the Gal4 activation domain.The Gal4 activation domain mediates several functionsin the transcription process including recruitment ofcofactors and general initiation factors, regulation ofprotein stability and activation via ubiquitination, aswell as regulation of activation via phosphorylation. Itis possible that mitochondrial dysfunction impinges onany one or more of these processes to elicit increasedtranscriptional activation.
Our results suggest that Gal4 is another transcriptionfactor regulated by the functional state of mitochondriaand that the activation domain is the primary target ofthe signaling pathway. In the case of Gal4 this signalleads to increased protein stability and/or DNA bindingwhich ultimately results in increased Gal4-dependenttranscription. Studies of the Rtg1-3 pathway show thatmitochondrial dysfunction activates a transcriptional re-sponse that mainly acts to reprogram metabolic path-ways in order for the cells to maintain essentialcellular functions [36]. The reason for enhancing Gal4activity in cells in which respiration is compromised isnot obvious, but it could be to increase fermentationof galactose, since rho0 cells can obtain energy exclu-sively via non-respiratory pathways.
Acknowledgments
We thank Drs. M. Ptashne, R. Reece, K. Struhl andF. Winston for strains and plasmids and Dr.W. Schaff-ner for support. This work was supported by a grantto MS from the Croatian Ministry of Science and Tech-nology and a SCOPES grant from the Swiss NationalScience Foundation (7KRPJ065578).
References
[1] Lohr, D., Venkov, P. and Zlatanova, J. (1995) Transcriptionalregulation in the yeast GAL gene family: a complex geneticnetwork. FASEB J. 9, 777–787.
[2] Bhat, P.J. and Murthy, T.V.S. (2001) Transcriptional control ofthe GAL/MEL regulon of yeast Saccharomyces cerevisiae: mech-anism of galactose-mediated signal transduction. Mol. Microbiol.40, 1059–1066.

B. Jelicic et al. / FEMS Microbiology Letters 253 (2005) 207–213 213
[3] Ma, J. and Ptashne, M. (1987) The carboxy-terminal 30 aminoacids of GAL4 are recognized by GAL80. Cell 50, 137–142.
[4] Wu, Y., Reece, R.J. and Ptashne, M. (1996) Quantitation ofputative activator-target affinities predicts transcriptional activat-ing potentials. EMBO J. 15, 3951–3963.
[5] Suzuki-Fujimoto, T., Fukuma, M., Yano, K.I., Sakurai, H.,Vonika, A., Johnston, S.A. and Fukasawa, T. (1996) Analysis ofthe galactose signal transduction pathway in Saccharomyces
cerevisiae: interaction between Gal3p and Gal80p. Mol. Cell.Biol. 16, 2504–2508.
[6] Platt, A. and Reece, R.J. (1998) The yeast galactose genetic switchis mediated by the formation of a Gal4p–Gal80p–Gal3p complex.EMBO J. 17, 4086–4091.
[7] Sil, A.K., Alam, S., Xin, P., Ma, L., Morgan, M., Lebo, C.M.,Woods, M.P. and Hopper, J.E. (1999) The Gal3p–Gal80p–Gal4ptranscription switch of yeast: Gal3p destabilizes the Gal80p–Gal4p complex in response to galactose and ATP. Mol. Cell. Biol.19, 7828–7840.
[8] Peng, G. and Hopper, J.E. (2000) Evidence for Gal3p�s cytoplas-mic location and Gal80p�s dual cytoplasmic-nuclear locationimplicates new mechanisms for controlling Gal4p activity inSaccharomyces cerevisiae. Mol. Cell. Biol. 20, 5140–5148.
[9] Peng, G. and Hopper, J.E. (2002) Gene activation by interactionof an inhibitor with a cytoplasmic signaling protein. Proc. Natl.Acad. Sci. USA 99, 8548–8553.
[10] Hirst, M., Kobor, M.S., Kuriakose, N., Greenblatt, J. andSadowski, I. (1999) GAL4 is regulated by the RNA polymerase IIholoenzyme-associated cyclin-dependent protein kinase SRB10/CDK8. Mol. Cell 3, 673–678.
[11] Rohde, J.R., Trinh, J. and Sadowski, I. (2000) Multiple signalsregulateGAL transcription in yeast.Mol.Cell. Biol. 20, 3880–3886.
[12] Sadowski, I., Niedbala, D., Wood, K. and Ptashne, M. (1991)GAL4 is phosphorylated as a consequence of transcriptionalactivation. Proc. Natl. Acad. Sci. USA 88, 10510–10514.
[13] Sadowski, I., Costa, C. and Dhanawansa, R. (1996) Phosphor-ylation of Gal4p at a single C-terminal residue is necessary forgalactose-inducible transcription. Mol. Cell. Biol. 16, 4879–4887.
[14] Muratani, M., Kung, C., Shokat, K.M. and Tansey, W.P. (2005)The F Box Protein Dsg1/Mdm30 is a transcriptional coactivatorthat stimulates Gal4 turnover and cotranscriptional mRNAprocessing. Cell 120, 887–899.
[15] Epstein, C.B., Waddle, J.A., Hale 4th, W., Dave, V., Thornton, J.,Macatee, T.L., Garner, H.R. and Butow, R.A. (2001) Genome-wide responses to mitochondrial dysfunction. Mol. Biol. Cell 12,297–308.
[16] Traven, A., Wong, J.M.S., Xu, D., Sopta, M. and Ingles, C.J.(2001) Interorganellar communication. Altered nuclear geneexpression profiles in a yeast mitochondrial DNA mutant. J.Biol. Chem. 276, 4020–4027.
[17] Liao, X. and Butow, R.A. (1993) RTG1 and RTG2: two yeastgenes required for a novel path of communication from mito-chondria to the nucleus. Cell 72, 61–71.
[18] Sekito, T., Thornton, J. and Butow, R.A. (2000) Mitochondria-to-nuclear signaling is regulated by the subcellular localization ofthe transcription factors Rtg1p and Rtg3p. Mol. Biol. Cell 11,2103–2115.
[19] Rothermel, B.A., Thornton, J.L. and Butow, R.A. (1997) Rtg3p,a basic helix-loop-helix/leucine Zipper protein that functions in
mitochondrial-induced changes in gene expression, containsindependent activation domains. J. Biol. Chem. 272, 19801–19807.
[20] Conrad-Webb, H. and Butow, R.A. (1995) A polymerase switchin the synthesis of rRNA in Saccharomyces cerevisiae. Mol. Cell.Biol. 15, 2420–2428.
[21] Liu, Z. and Butow, R.A. (1999) A transcriptional switch in theexpression of yeast tricarboxylic acid cycle genes in response to areduction or loss of respiratory function. Mol. Cell. Biol. 19,6720–6728.
[22] Conlan, R.S., Gounalaki, N., Hatzis, P. and Tzamarias, D. (1999)The Tup1-Cyc8 protein complex can shift from a transcriptionalco-repressor to a transcriptional co-activator. J. Biol. Chem. 274,205–210.
[23] Hallstrom, T.C. and Moye-Rowley, W.S. (2000) Multiple signalsfrom dysfunctional mitochondria activate the pleiotropic drugresistance pathway in Saccharomyces cerevisiae. J. Biol. Chem.275, 37347–37356.
[24] Devaux, F., Carvajal, E., Moye-Rowley, S. and Jacq, C. (2002)Genome-wide studies on the nuclear PDR3-controlled response tomitochondrial dysfunction in yeast. FEBS Lett. 515, 25–28.
[25] Rickwood, D., Dujon, B. and Darley-Usmar, V.M. (1988) Yeastmitochondria In: Yeast: A Practical Approach (Campbel, I. andDuffus, J.H., Eds.), pp. 185–254. IRL Press Limited, Oxford.
[26] Giniger, E., Varnum, S.M. and Ptashne, M. (1985) Specific DNAbinding of GAL4, a positive regulatory protein of yeast. Cell 40,767–774.
[27] Zaman, Z., Ansari, A.Z., Koh, S.S., Young, R. and Ptashne, M.(2001) Interaction of a transcriptional repressor with the RNApolymerase II holoenzyme plays a crucial role in repression. Proc.Natl. Acad. Sci. USA 98, 2550–2554.
[28] Adams, A., Gottschling, D.E., Kaiser, C.A. and Stearns, T. (1997)Methods in yeast genetics. A Cold Spring Harbor LaboratoryCourse Manual. Cold spring Harbor Laboratory Press.
[29] Melcher, K., Sharma, B., Ding, W.V. and Nolden, M. (2000) Zerobackground yeast reporter plasmids. Gene 247, 53–61.
[30] Ausubel, F.M., Brent, R., Kingston, R., Moore, D., Seidman, J.,Smith, J. and Struhl, K. (1987) Current Protocols in MolecularBiology. Wiley, New York, NY, USA.
[31] Kushnirov, V.V. (2000) Rapid and reliable protein extractionfrom yeast. Yeast 16, 857–860.
[32] Liang, S.D., Marmorstein, R., Harrison, S.C. and Ptashne, M.(1996) DNA sequence preferences of GAL4 and PPR1: how asubset of Zn2 Cys6 binuclear cluster proteins recognizes DNA.Mol. Cell. Biol. 16, 3773–3780.
[33] Ding, W.V. and Johnston, S.A. (1997) The DNA binding andactivation domains of Gal4p are sufficient for conveying itsregulatory signals. Mol. Cell. Biol. 17, 2538–2549.
[34] Rothermel, B.A., Shyjan, A.W., Etheredge, J.L. and Butow,R.A. (1995) Transactivation by Rtg1p, a basic helix-loop-helixprotein that functions in communication between mitochon-dria and the nucleus in yeast. J. Biol. Chem. 270, 29476–29482.
[35] Xu, H.E., Kodadek, T. and Johnston, S.A. (1995) A Single GAL4dimer can maximally activate transcription under physiologicalconditions. Proc. Natl. Acad. Sci. USA 92, 7677–7680.
[36] Butow, R.A. and Avadhani, N.G. (2004) Mitochondrial signaling:the retrograde response. Mol. Cell 14, 1–15.




















