
Endoplasmic reticulum dysfunction in neurological disease
14
www.thelancet.com/neurology Vol 12 January 2013 105 Review Endoplasmic reticulum dysfunction in neurological disease Benoit D Roussel, Antonina J Kruppa, Elena Miranda, Damian C Crowther, David A Lomas, Stefan J Marciniak Endoplasmic reticulum (ER) dysfunction might have an important part to play in a range of neurological disorders, including cerebral ischaemia, sleep apnoea, Alzheimer’s disease, multiple sclerosis, amyotrophic lateral sclerosis, the prion diseases, and familial encephalopathy with neuroserpin inclusion bodies. Protein misfolding in the ER initiates the well studied unfolded protein response in energy-starved neurons during stroke, which is relevant to the toxic effects of reperfusion. The toxic peptide amyloid β induces ER stress in Alzheimer’s disease, which leads to activation of similar pathways, whereas the accumulation of polymeric neuroserpin in the neuronal ER triggers a poorly understood ER-overload response. In other neurological disorders, such as Parkinson’s and Huntington’s diseases, ER dysfunction is well recognised but the mechanisms by which it contributes to pathogenesis remain unclear. By targeting components of these signalling responses, amelioration of their toxic effects and so the treatment of a range of neurodegenerative disorders might become possible. Introduction Most neurodegenerative disorders could arguably be viewed as arising from defective protein folding. This statement is clearly true for those rare disorders that are caused by the misfolding of mutated neuronal proteins, such as Huntington’s disease, but it is also the case for common disorders such as cerebral ischaemia, in which low energy can impair normal protein folding, 1 and sporadic Alzheimer’s disease, in which protein aggre- gation causes cellular stress. 2–4 Even some infectious neurodegenerative disorders such as the prion diseases arise from abnormal protein folding. 5–9 As such, the processes for disposing of misfolded proteins are the focus of much research. Although the subcellular lo- cation of protein misfolding can differ between these disorders, the interdependence of protein folding throughout the cell suggests that endoplasmic reticulum (ER) dysfunction could be the final common pathway for many neurological diseases (table). 1–7,9–60 The ER is necessary for the folding of all secreted and membrane proteins, and insults that impair its function induce a pathological state known as ER stress. 61 ER stress triggers an adaptive programme called the un- folded protein response, which combines the early inhibition of protein synthesis with a later upregulation of genes that promote protein folding or disposal. 61 Both the translational and transcriptional components of the response protect neurons from being overwhelmed by misfolded ER proteins; however, when the insult is too great, apoptotic cell death will often follow. Although many cells are fairly well protected from accumulation of misfolded protein through the continued dilution of the ER by cell replication, this process is unavailable to post-mitotic neurons, which depend exclusively on the unfolded protein response for survival of such insults. Even glial cells, which can replicate, are susceptible to ER stress because of their highly developed secretory pathways. The unfolded protein response should be distinguished from the less well understood ER-overload response. 47 The unfolded protein response is triggered by protein misfolding within the ER lumen, whereas the ER- overload response occurs when well folded or misfolded proteins accumulate and distend the ER. In this Review, we discuss the relevance of these forms of ER dysfunction to neurological disease, since an improved understanding of each might help us to identify new therapeutic strategies. We focus on ischaemic stroke, Alzheimer’s disease, and Parkinson’s disease to discuss ER stress, and use familial encephalopathy with neuroserpin inclusion bodies (FENIB) as an example of ER overload, since these are the diseases for which most evidence currently exists. Additionally, we discuss the evidence for ER dysfunction in multiple sclerosis, amyotrophic lateral sclerosis, and the prion diseases. Some neurological disorders provoke ER stress signalling despite being caused by the accumulation of protein within the cytoplasm—eg, Huntington’s disease. 35 Such disorders emphasise the interdependence of protein-folding networks in the neuron and so shall be discussed as examples of disordered protein homoeostasis (proteostasis). ER stress signalling ER stress and the unfolded protein response The complex signalling pathways triggered by ER stress constitute the so-called unfolded protein response (figure 1). 61 Many of the indicators used to detect and quantify ER stress are themselves components of the unfolded protein response. Some of these components are activated transiently, so their absence cannot be used as evidence for the absence of ER stress; 61,62 conversely, some components can be activated by stresses other than ER protein misfolding, so need to be interpreted with caution. We begin with a summary of the major mediators of ER stress signalling, with an emphasis on their role in neurons, before discussing some specific examples of neuropathology mediated by ER dysfunction. PERK Whenever misfolded protein accumulates in the ER lumen, new protein synthesis is rapidly inhibited. 63 This inhibition occurs because ER stress activates PERK (PKR-like endoplasmic reticulum eIF2α kinase; also known as eukaryotic translation initiation factor 2α Lancet Neurol 2013; 12: 105–18 Department of Medicine, Cambridge Institute for Medical Research (B D Roussel PhD, A J Kruppa PhD, Prof D A Lomas PhD, S J Marciniak PhD) and Department of Genetics (D C Crowther PhD), University of Cambridge, Cambridge, UK; Charles Darwin Department of Biology and Biotechnology and Institut Pasteur–Cenci Bolognetti Foundation, Sapienza University of Rome, Rome, Italy (E Miranda PhD); and INSERM U919, University of Caen Lower Normandy, Serine Proteases and Pathophysiology of the Neurovascular Unit, Public Interest Group CYCERON, Caen, France (B D Roussel) Correspondence to: Dr Stefan J Marciniak, Cambridge Institute for Medical Research, Wellcome Trust/MRC Building, Hills Road, Cambridge CB2 0XY, UK [email protected]
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Endoplasmic reticulum dysfunction in neurological disease

www.thelancet.com/neurology Vol 12 January 2013 105
Review
Endoplasmic reticulum dysfunction in neurological diseaseBenoit D Roussel, Antonina J Kruppa, Elena Miranda, Damian C Crowther, David A Lomas, Stefan J Marciniak
Endoplasmic reticulum (ER) dysfunction might have an important part to play in a range of neurological disorders, including cerebral ischaemia, sleep apnoea, Alzheimer’s disease, multiple sclerosis, amyotrophic lateral sclerosis, the prion diseases, and familial encephalopathy with neuroserpin inclusion bodies. Protein misfolding in the ER initiates the well studied unfolded protein response in energy-starved neurons during stroke, which is relevant to the toxic eff ects of reperfusion. The toxic peptide amyloid β induces ER stress in Alzheimer’s disease, which leads to activation of similar pathways, whereas the accumulation of polymeric neuroserpin in the neuronal ER triggers a poorly understood ER-overload response. In other neurological disorders, such as Parkinson’s and Huntington’s diseases, ER dysfunction is well recognised but the mechanisms by which it contributes to pathogenesis remain unclear. By targeting components of these signalling responses, amelioration of their toxic eff ects and so the treatment of a range of neurodegenerative disorders might become possible.
IntroductionMost neurodegenerative disorders could arguably be viewed as arising from defective protein folding. This statement is clearly true for those rare disorders that are caused by the misfolding of mutated neuronal proteins, such as Huntington’s disease, but it is also the case for common disorders such as cerebral ischaemia, in which low energy can impair normal protein folding,1 and sporadic Alzheimer’s disease, in which protein aggre-gation causes cellular stress.2–4 Even some infectious neurodegenerative disorders such as the prion diseases arise from abnormal protein folding.5–9 As such, the processes for disposing of misfolded proteins are the focus of much research. Although the subcellular lo-cation of protein misfolding can diff er between these disorders, the interdependence of protein folding throughout the cell suggests that endoplasmic reticulum (ER) dysfunction could be the fi nal common pathway for many neurological diseases (table).1–7,9–60
The ER is necessary for the folding of all secreted and membrane proteins, and insults that impair its function induce a pathological state known as ER stress.61 ER stress triggers an adaptive programme called the un-folded protein response, which combines the early inhibition of protein synthesis with a later upregulation of genes that promote protein folding or disposal.61 Both the translational and transcriptional components of the response protect neurons from being overwhelmed by misfolded ER proteins; however, when the insult is too great, apoptotic cell death will often follow. Although many cells are fairly well protected from accumulation of misfolded protein through the continued dilution of the ER by cell replication, this process is unavailable to post-mitotic neurons, which depend exclusively on the unfolded protein response for survival of such insults. Even glial cells, which can replicate, are susceptible to ER stress because of their highly developed secretory pathways.
The unfolded protein response should be distinguished from the less well understood ER-overload response.47 The unfolded protein response is triggered by protein misfolding within the ER lumen, whereas the ER-
overload response occurs when well folded or misfolded proteins accumulate and distend the ER. In this Review, we discuss the relevance of these forms of ER dysfunction to neurological disease, since an improved understanding of each might help us to identify new therapeutic strategies. We focus on ischaemic stroke, Alzheimer’s disease, and Parkinson’s disease to discuss ER stress, and use familial encephalopathy with neuroserpin inclusion bodies (FENIB) as an example of ER overload, since these are the diseases for which most evidence currently exists. Additionally, we discuss the evidence for ER dysfunction in multiple sclerosis, amyotrophic lateral sclerosis, and the prion diseases. Some neurological disorders provoke ER stress signalling despite being caused by the accumulation of protein within the cytoplasm—eg, Huntington’s disease.35 Such disorders emphasise the interdependence of protein-folding networks in the neuron and so shall be discussed as examples of disordered protein homoeostasis (proteostasis).
ER stress signallingER stress and the unfolded protein responseThe complex signalling pathways triggered by ER stress constitute the so-called unfolded protein response (fi gure 1).61 Many of the indicators used to detect and quantify ER stress are themselves components of the unfolded protein response. Some of these components are activated transiently, so their absence cannot be used as evidence for the absence of ER stress;61,62 conversely, some components can be activated by stresses other than ER protein misfolding, so need to be interpreted with caution. We begin with a summary of the major mediators of ER stress signalling, with an emphasis on their role in neurons, before discussing some specifi c examples of neuropathology mediated by ER dysfunction.
PERKWhenever misfolded protein accumulates in the ER lumen, new protein synthesis is rapidly inhibited.63 This inhibition occurs because ER stress activates PERK (PKR-like endoplasmic reticulum eIF2α kinase; also known as eukary otic translation initiation factor 2α
Lancet Neurol 2013; 12: 105–18
Department of Medicine, Cambridge Institute for Medical Research (B D Roussel PhD, A J Kruppa PhD, Prof D A Lomas PhD, S J Marciniak PhD) and Department of Genetics (D C Crowther PhD), University of Cambridge, Cambridge, UK; Charles Darwin Department of Biology and Biotechnology and Institut Pasteur–Cenci Bolognetti Foundation, Sapienza University of Rome, Rome, Italy (E Miranda PhD); and INSERM U919, University of Caen Lower Normandy, Serine Proteases and Pathophysiology of the Neurovascular Unit, Public Interest Group CYCERON, Caen, France (B D Roussel)
Correspondence to:Dr Stefan J Marciniak, Cambridge Institute for Medical Research, Wellcome Trust/MRC Building, Hills Road, Cambridge CB2 0XY, [email protected]

106 www.thelancet.com/neurology Vol 12 January 2013
Review
kinase 3 [EIF2AK3] or pancreatic eIF2α kinase [PEK], because of the tissue in which it was fi rst discovered).63 This kinase is ubiquitously expressed, but is most important in highly secretory tissues such as the brain, where it functions to match protein synthesis with the effi ciency of protein folding in the ER.64 When protein folding is progressing well, PERK is inactive, but when misfolded ER proteins accumulate, the kinase is activated and phosphorylates eukaryotic translation initiation
factor 2α (eIF2α; fi gure 1).65 This substrate is a subunit of the heterotrimeric GTP-binding complex eIF2, which regulates the initiation of protein synthesis.63 eIF2 recruits methionyl-tRNA to the ribosome at the onset of protein translation, but when its eIF2α subunit is phosphorylated this activity is lost and translation ceases. The load of proteins that enters the ER is thus reduced after PERK activation, which prevents the further accumulation of misfolded proteins.
Neurological phenotype Cause Proposed mechanism of ER dysfunction
CACH/VWM10–12 Cerebellar ataxia, central hypomyelinationAutosomal recessive
EIF2B mutations Enhanced ATF4 synthesis
Pelizaeus-Merzbacher disease13
Cerebellar ataxia, central hypomyelinationX-linked
PLP1 duplication Accumulation of PLP1 in the ER
Charcot-Marie-Tooth disease type 1A14
Peripheral sensory neuropathy, peripheral demyelinationAutosomal dominant
PMP22 mutations or duplication Accumulation of myelin protein PMP22 in the ER
Charcot-Marie-Tooth disease type 1B15
Peripheral sensory neuropathy, peripheral demyelinationAutosomal dominant
MPZ mutations Presumed accumulation of myelin protein P0 in the ER
Multiple sclerosis16,17 Progressive motor and cognitive impairment, patchy central demyelinationSporadic
Infl ammatory loss of myelination Infl ammation-driven ER stress
Marinesco-Sjögren syndrome18,19
Cerebellar ataxia, Purkinje cell lossAutosomal recessive
SIL1 mutations BiP dysfunction due to impaired SIL1 nucleotide exchange activity
Alzheimer’s disease2–4,20–26 Loss of cortical neurons, progressive cognitive impairmentSporadic and autosomal dominant
Poorly understood amyloid-β accumulation, hyperphosphorylation of tau
Impaired ER calcium homoeostasis, direct amyloid-β toxic eff ects
Parkinson’s disease27–34 Progressive motor and cognitive dysfunctionSporadic; autosomal recessive; autosomal dominant
Poorly understood loss of dopaminergic neurons from the substantia nigra; familial forms caused by mutations in PARK2, SNCA, and PINK1
Impaired mitochondrial ATP generation that causes secondary ER failure; accumulation of aggregated synuclein in the ER
Huntington’s disease35,38 Progressive motor and cognitive dysfunctionAutosomal dominant
Expansion of CAG repeats in HTT leads to selective neuronal loss in caudate and putamen
Failure of proteostasis, probably caused by impairment of proteasome function by cytosolic aggregates of HTT
Amyotrophic lateral sclerosis39–46
Progressive motor impairmentSporadic and autosomal dominant
Poorly understood loss of motor neurons; some familial forms caused by mutations in SOD1
Accumulation of mutant SOD1 in the ER
Prion diseases5–7,9 Spongiform encephalopathyInfective and autosomal dominant
Propagation of infective prion conformation of PrP Impaired cellular proteostasis; possible direct ER toxic eff ects
FENIB47,48 Progressive dementia and epilepsyAutosomal dominant
SERPINI1 mutations Polymerised neuroserpin accumulates in the ER and activates the ER-overload response
Tuberous sclerosis complex49
Central hamartomas cause epilepsy, often with cognitive and behavioural defectsAutosomal dominant
TSC1 and TSC2 mutations mTORC-mediated impairment of ER stress signalling
Epilepsy50 Progressive neuronal lossSporadic
Repeated seizure activity Incompletely understood; probably involves disturbed ER calcium homoeostasis
Wolfram syndrome 151 Diabetes insipidus, optic atrophy, deafnessAutosomal recessive
WFS1 mutations WFS1 encodes an ER ion channel, dysfunction of which induces ER stress
Torsion dystonia 152 Involuntary sustained muscle contractionAutosomal dominant
TOR1A mutations Impairment of torsin A, an AAA ATPase chaperone in the ER
Lipid storage disorders (eg, Gaucher’s disease)53
Progressive ataxia; mental and motor defi ciencyAutosomal recessive
GBA mutations Disturbed ER calcium homoeostasis and accumulation of mutated protein in the ER
Cerebral ischaemia1,54–58 Neuronal deathSporadic
Ischaemia causes impaired protein folding Energy defi ciency causes dysfunction of ER chaperones and enzymes
Sleep apnoea59 Neuronal deathSporadic
Ischaemia causes impaired protein folding Energy defi ciency causes dysfunction of ER chaperones and enzymes
Trauma60 Neuronal deathSporadic
Neuronal trauma and impaired perfusion Incompletely understood; probably involves disturbed energy metabolism and ER calcium homoeostasis
CACH=childhood ataxia with central hypomyelination. VWM=vanishing white matter disease. ER=endoplasmic reticulum. FENIB=familial encephalopathy with neuroserpin inclusion bodies. mTORC=mammalian target of rapamycin (sirolimus) complex. AAA=ATPases associated with diverse cellular activities.
Table: Endoplasmic reticulum stress in neurodegenerative diseases

www.thelancet.com/neurology Vol 12 January 2013 107
Review
ATF4Despite the inhibition of protein translation that accompanies phosphorylation of eIF2α, a small subset of mRNAs is translated more effi ciently under these conditions.66 The best characterised of these is activating transcription factor 4 (ATF4), which triggers a tran-scriptional programme of stress-responsive genes.66 Other members of the eIF2α kinase family can inhibit translation and activate these genes; for example, PKR (also known as EIF2AK2) responds to viral in fection, HRI (EIF2AK1) to iron defi ciency, and GCN2 (EIF2AK4) to aminoacid starvation.67 Because of this ability of eIF2α phosphorylation to mediate a response to several stresses, the pathway was named the integrated stress response.68 Surprisingly, within the brain this pathway can provoke specifi c behavioural responses. An absence of essential dietary aminoacids triggers the phosphorylation of eIF2α by GCN2 in the anterior piriform cortex, which changes feeding behaviour in rodents.69,70 Thus, phosphorylation of eIF2α is not synonymous with ER stress, and several lines of evi dence should always be sought when attributing phosphorylation of eIF2α to ER dysfunction.
Phosphorylation of eIF2α is especially important for the maintenance of axonal myelination in the human
CNS; indeed, mutations that disrupt the regulation of eIF2α result in white matter hypo myelination disorders such as childhood ataxia with central hypomyelination (CACH syndrome) and vanishing white matter disease (VWM).10 This family of autosomal recessive disorders encompasses a range of severity, from congenital to adult-onset, and involves the pro gressive loss of mental and motor function caused by a loss of brain white matter. In all cases, the causative mutations are found within subunits of eIF2B, a guanine nucleotide exchange factor that recharges the eIF2 complex with GTP (fi gure 1).11,12 These mutations disrupt the activity of eIF2B and enhance the synthesis of ATF4, which deregulates many target genes, including components of a negative feedback system that includes CHOP (also known as DNA-damage-inducible transcript 3 [DDIT3]), and GADD34 (protein phosphatase 1, regulatory subunit 15A [PPP1R15A]), which normally recover protein translation after its initial inhibition by PERK.62,71
CHOPThe transcription factor CHOP is induced by ATF4 and has widely been regarded as proapoptotic during ER stress, since Chop–/– animals are resistant to ER-stress-
Figure 1: Endoplasmic reticulum stress signallingIn health (left), newly synthesised secretory proteins are translocated into the endoplasmic reticulum (ER) lumen cotranslationally and fold with the help of chaperones—eg, BiP. When recognised as correctly folded, the proteins exit the ER in COPII-coated vesicles and traffi c along the secretory pathway. During ER stress (right), misfolded proteins accumulate in the ER lumen and sequester BiP from the sensor molecules PERK, IRE1, and ATF6. PERK phosphorylates eIF2α, which binds to and inhibits its guanine nucleotide exchange factor, eIF2B. This inhibition blocks new secretory protein synthesis and triggers the integrated stress response through raised ATF4. Activation of IRE1 leads to the generation of active XBP1, and cleavage of ATF6 (by S1P [MBTPS1] and S2P [MBTPS2], represented as scissors) generates ATF6c. These transcription factors cooperate to induce target genes of the unfolded protein response (UPR). CACH=childhood ataxia with central hypomyelination. VWM=vanishing white matter disease. ALS=amyotrophic lateral sclerosis. CMT=Charcot-Marie-Tooth disease.
PERKIRE1
ATF6
eIF2α
eIF2B
ATF6c
XBP1
ATF4
BiP
Ongoing translation
Attenuatedtranslation
Endoplasmic reticulum
Golgi
Nucleus UPR genetranscription
Health
P
P P PP
Nascentprotein
Foldedprotein
Unfoldedprotein BiP
Stroke
Alzheimer’sdisease
Parkinson’sdisease
Huntington’sdisease
CACH/VWM
ATP
ADP
Ca2+
Ca2+
Ca2+Ca2+
ADPATF6
GTP
GDP
ALSMultiplesclerosis
CMT 1Aand 1B
Endoplasmic reticulum stress

108 www.thelancet.com/neurology Vol 12 January 2013
Review
induced cell death.62 However, this belief is inconsistent with observations made in neurological diseases. For example, Chop expression is clearly antiapoptotic in mouse models of Pelizaeus-Merzbacher disease, an X-linked leukodystrophy caused by abnormalities of the proteolipid protein 1 (PLP1) gene.13 Indeed, identifi cation of the target genes of CHOP did not reveal a link with apoptosis, but instead identifi ed genes that promote protein secretion.62 This fi nding led to the suggestion that in models of severe ER stress, the promotion of protein secretion by CHOP might increase the intensity of stress and thus induce cell death.62 By contrast, during conditions of milder ER stress (eg, in Pelizaeus-Merzbacher disease), CHOP probably does not increase ER stress suffi ciently to trigger cell death.13 These fi ndings emphasise the need to model not only the nature of the stress, but also its intensity and duration, since diff erences in either can substantially aff ect the outcome and thus lead to benefi cial or deleterious eff ects depending on the context. Some consequences of CHOP expression might be cell-type specifi c. For example, although deletion of Chop worsens the central hypomyelination seen in a mouse model of Pelizaeus-Merzbacher disease,13 Chop–/– animals are protected against the peripheral hypomyelination caused by mutant myelin protein zero (P0) in mouse models of Charcot-Marie-Tooth disease type 1B.15
GADD34An important target gene of CHOP encodes GADD34, which is a regulatory subunit of protein phosphatase 1.62 With this subunit, protein phosphatase 1 de-phosphorylates eIF2α and so brings about the recovery of protein translation after its initial inhibition. Inactivation of Gadd34 has been shown in mice to protect against tissue damage during ER stress by reducing the accumulation of misfolded proteins.62 A small molecule called salubrinal has since been identifi ed that increases eIF2α phosphorylation and promotes the survival of ER-stressed cells.72 Some have suggested that salubrinal might be an inhibitor of the eIF2α phosphatases,72 but direct evidence for this suggestion is absent. However, a selective inhibitor of GADD34, guanabenz, has been identifi ed, which has been used to confi rm the benefi cial eff ects of GADD34 inhibition during ER stress, at least in cultured cells.73 Future therapies that involve the inhibition of GADD34 might therefore be able to modulate ER stress.
ATF6In addition to the integrated-stress-response genes regulated by PERK and ATF4, many additional genes are induced during the unfolded protein response by two further ER-stress sensors, inositol-requiring enzyme 1 (IRE1; also known as endoplasmic reticulum to nucleus signalling 1 [ERN1]) and activating transcription factor 6 (ATF6; fi gure 1).61 Like PERK, both are ER-membrane
proteins that are held inactive under normal conditions by the binding of the ER chaperone BiP (binding immunoglobulin protein; also known as heat shock 70kDa protein 5 [HSPA5] or GRP78). During stress, misfolded proteins sequester BiP, which frees ATF6 to traffi c to the Golgi apparatus, where it is cleaved to release a soluble transcription factor. Notably, astrocytes express an ATF6-related protein called OASIS (also known as CREB3L1), the functions of which seem to overlap partly with those of ATF6.74 This protein might enable astrocytes to respond to ER stress in a tissue-specifi c way.
Cleavage of ATF6 is strong evidence of ER stress, but is diffi cult to detect. As such, many studies do not measure this arm of the unfolded protein response. However, the use of sensitive reporters that measure the binding of ATF6 to specifi c DNA sequences can overcome this diffi culty in vitro.47
IRE1The activation of IRE1 is similar to that of PERK, but unlike PERK, IRE1 triggers splicing of the mRNA that encodes the transcription factor X-box binding protein 1 (XBP1).75 This splicing induces a frameshift that generates the active form of this protein. Like phosphorylation of eIF2α, the splicing of XBP1 mRNA is transient.62 After the alleviation of ER stress by the induction of unfolded-protein-response target genes, spliced XBP1 mRNA is rapidly lost from the cell, to be replaced by the unspliced form.62 In laboratory studies, supraphysiological ER stress can cause complete splicing of XBP1 mRNA, but in-vivo stresses are often more subtle, so observation of complete splicing is rare and careful quantifi cation is therefore necessary.76
If proteins do not fold in the ER, they are prevented from progressing along the secretory pathway and eventually are targeted for degradation.77 This ER-associated protein degradation (ERAD) returns ter-minally misfolded proteins to the cytosol, where they are ubiquitinated by ER-associated ubiquitin ligases for degradation by the proteasome.78 Many components of the ERAD machinery are target genes of XBP1, and so the capacity for ERAD is increased by activation of IRE1.79
Cerebral hypoxiaIn high-income countries, stroke is the third biggest killer and the leading cause of disability in adults. Acute ischaemia, the main cause of neuronal loss, has a complex pathology that seems to involve ER stress.1,64,80–82 During cerebral ischaemia, neuronal depolarisation due to energy depletion causes the uncontrolled release of glutamate. The consequent activation of NMDA receptors on nearby neurons generates further glutamate release and causes an ischaemic depolarisation wave to spread outwards from the initial site of damage, which leads to widespread disturbance of calcium homoeostasis.83 Unsurprisingly, many

www.thelancet.com/neurology Vol 12 January 2013 109
Review
studies implicate ER-calcium-store release in the resultant excitotoxic death.83
The ER is the main site for calcium storage within the cell and so its chaperones have evolved to function effi ciently in this high-calcium environment; indeed, many need a high calcium concentration to function.84 Calcium is pumped into the ER by the sarcoplasmic/endoplasmic reticulum calcium ATPase (SERCA pump) and released back into the cytosol by the inositol trisphosphate receptor (IP3R or ITPR) in response to extracellular signals, and by the ryanodine receptors during calcium-induced calcium release.83 Inhibition of the SERCA pump by thapsigargin induces apoptosis in many cell types, including neuroblastoma cells, and is often used to induce ER stress in experimental settings.85 During cerebral ischaemia, energy depletion leads to failure of the SERCA pump and thus redistribution of ER calcium into the cytosol, which delivers a double blow of combined toxic eff ects from uncontrolled cytosolic calcium and ER stress caused by chaperone dysfunction (fi gure 2).83
The unfolded protein response is activated in many rodent models of cerebral ischaemia.1,80 In these models, protein synthesis is rapidly inhibited and accompanies the phosphorylation of eIF2α. Although the inhibition of translation has been attributed to aggregation of components of the translation machinery,86 the primary mechanism seems to be through phosphorylation of eIF2α,87 which can remain phosphorylated for up to 12 h after ischaemia.88 PERK is the only eIF2α kinase to be activated by cerebral ischaemia and, predictably, Perk–/– mice do not show eIF2α phosphorylation after transient cerebral ischaemia or reduced protein trans-lation during the reperfusion period.64 Even with the far milder recurrent cerebral hypoxia seen in obstructive sleep apnoea, activation of PERK has been reported and has been implicated in the loss of upper-airway motor neurons in patients.59
IRE1 is also activated during cerebral ischaemia, which leads to the induction of unfolded-protein-response target genes.81,82 IRE1 seems to promote activation of the proapoptotic caspase-12 in cultured rodent cells, and this has also been detected in ischaemic rodent brains.89 However, caspase-12 is non-functional in most human beings, with the exception of people from sub-Saharan Africa, because of a common truncating mutation.90 For this reason, caspase-4 has been proposed to serve an analogous function during ER stress in humans to that of caspase-12 in rodents.91
Because of the diffi culties of detection, the role of ATF6 during ischaemia is unclear. ATF6 expression is raised after transient middle-cerebral-artery occlusion in rats,80 but its activation has not been detected in rat brain after cardiac arrest and resuscitation.92 OASIS is also upregulated and seems to have a neuroprotective role in response to kainate excitotoxicity.93
CHOP is induced after forebrain ischaemia in various rodent models, including bilateral common carotid
artery54,94 or middle-cerebral-artery occlusion in mice88 and global cerebral ischaemia in rats.95 The location in the brain of CHOP induction seems to be closely associated with subsequent cell death,55,95 and deletion of Chop protects mice during bilateral common-carotid-artery occlusion.94 Similarly, depletion of CHOP with RNA interference partly prevents the death of astrocyte cultures stressed by oxygen and glucose deprivation.96 Some of the toxic eff ects attributable to CHOP are caused by increased protein translation mediated by GADD34.62 Indeed, GADD34 induction is well recognised to accompany cerebral ischaemia97 (detected at the peri-infarct penumbra within 2–24 h)98 and accounts for the transient nature of eIF2α phos-phorylation during stroke.64 Attenuation of translation
Figure 2: Endoplasmic reticulum stress during cerebral ischaemiaDuring an ischaemic stroke, the poor supply of glucose and oxygen in neurons induces a substantial fall in ATP production. Without energy, BiP binds to unfolded proteins with reduced effi ciency, which results in the accumulation of misfolded endoplasmic reticulum (ER)-client proteins and subsequent activation of the unfolded protein response (UPR). In parallel, failure of the sarcoplasmic/endoplasmic reticulum calcium ATPase (SERCA pump) depletes ER calcium and worsens chaperone function still further. Extracellular tissue-type plasminogen activator (tPA) cleaves the NMDA receptor, which allows calcium infl ux into the cytosol. The resultant increase in cytosolic calcium triggers calcium-induced calcium release via ryanodine receptors located in the ER membrane, which leads to calcium depletion from the ER. In severely aff ected neurons, raised cytosolic calcium ultimately induces cell death. In surviving neurons, induction of the eIF2α phosphatase GADD34 by ATF4 in a CHOP-dependent way enables the eventual recovery of protein translation. This process can contribute to reperfusion injury through increased ER protein load.
ADP
NMDA receptor
Inefficient BiP(low Ca2+ and
low ATP)
SERCA pumpfailure
Calcium-induced calcium release
Reduced ATP(low glucoseand hypoxia)
GADD34 CHOP
ATF6c
XBP1
ATF4
UPR genetranscription
P
P P PP
Unfoldedprotein
?
Ca2+Ca2+
Ca2+
Golgi
ATF6
PERK
eIF2α
IRE4
Ca2+
tPA

110 www.thelancet.com/neurology Vol 12 January 2013
Review
protects the cell from accumulation of misfolded proteins. Thus, treatment with salubrinal (a drug that enhances eIF2α phosphorylation) reduces ER stress and promotes cellular survival in kainate-induced neuro-toxicity.99 Indeed, salubrinal limits infarct size in a mouse model of stroke.1
In addition to attenuation of protein translation, the unfolded protein response leads to upregulation of ER molecular chaperones, the most abundant of which is BiP.61 After transient forebrain ischaemia, BiP expression in the hippocampus and cortex peaks be tween 12 and 72 h after reperfusion,55,56 but can remain high for up to 2 weeks.80 This upregulation is protective, since suppression of BiP enhances apoptosis in hippocampal neurons exposed to excitotoxic and oxi dative insults,100 whereas its overexpression in primary astrocyte cultures is protective against oxygen and glucose deprivation.57 Indeed, increased induction of BiP is associated with neuronal protection.58,81
Induction of the ER chaperone HYOU1 (also known as ORP150) is also seen in ischaemic neurons, and overexpression of HYOU1 protects mice from cerebral ischaemia.101 HYOU1 modulates the activity of BiP, which exists in either an ATP-bound or ADP-bound form, each with diff erent substrate-binding affi nity. HYOU1 functions as a nucleotide exchange factor for BiP, to replace its bound ADP with ATP. Another BiP nucleotide exchange factor, SIL1, is mutated in the Marinesco-Sjögren syndrome of cerebellar ataxia, in which homozygous loss of SIL1 leads to ER stress and selective loss of cerebellar Purkinje cells.18 When ATP–BiP binds its substrate protein, a co-chaperone of the DNAJ family is needed to stimulate the ATPase activity of BiP and so promote high-affi nity substrate binding. The relative amounts of ADP–BiP and ATP–BiP seem to be important for neuronal survival, since the toxic eff ects of SIL1 loss can be ameliorated by the deletion of an ER luminal DNAJ co-chaperone called DNAJC3 (or p58IPK).19 Whereas loss of SIL1 would be predicted to reduce ATP–BiP through impaired nucleotide exchange, loss of DNAJC3 is likely to restore balance through reduced BiP ATPase activity.
When taken together, these fi ndings suggest that ER stress is induced by cerebral ischaemia and that the unfolded protein response limits cerebral infarct size. Good evidence exists to suggest that manipulation of ER stress signalling through increased phosphorylation of eIF2α could have important therapeutic eff ects as an acute intervention for cerebral ischaemia.
Alzheimer’s diseaseAlzheimer’s disease is a relentlessly progressive neuro-degenerative disorder and the most common cause of dementia in adults. The classic histopathological features consist of extracellular plaques of the amyloid-β peptide and intracellular neurofi brillary tangles of hyperphos-phorylated aggregates of microtubule-associated protein
tau. Studies of familial Alzheimer’s disease have shown that the disorder is caused by overproduction of the aggregation-prone amyloid β42 (fi gure 3).102 Despite decades of study, the molecular pathology of the disease remains unclear. However, increasing evidence suggests that ER stress might be relevant for understanding the disorder.
Studies of post-mortem tissue from the brains of individuals with Alzheimer’s disease have provided evidence for ER dysfunction. However, when such studies use only one marker of ER stress, they can generate confusing results. For example, when BiP is examined in isolation, it can show no change,3 a decrease,103 or even an increase in expression,2,104 although BiP does seem to be increased in post-mortem brains with histologically advanced dis ease.104,105 However, although it is often a useful indicator, the expression of BiP alone is inadequate to assess the activation of unfolded-protein-response signalling, especially in a chronic disorder in which histological features change with time. Inclusion of additional markers of the response has therefore proved useful in such studies. For example, protein disulfi de isomerase, another target of the unfolded protein response, is raised in the temporal lobes of the brains of individuals with Alzheimer’s disease,3 and is associated with neuro-fi brillary tangles in the hippocampus and frontal lobes and in dystrophic neurites of senile plaques.4 With respect to more proximal markers of unfolded-protein-response signalling, PERK is activated in the hippo-campus of patients with Alzheimer’s disease, colocalises with staining for phosphorylated tau, and is associated with histological staging.20,106 However, phosphorylated PERK is barely detectable in tangle-bearing neurons themselves, which suggests that PERK is activated at the pretangle stage in neurons of the hippocampus.20 Additionally, spliced XBP1 mRNA is raised in the temporal cortex of patients with the disease compared with age-matched controls.3 A substantial increase in the number of neurons that are immuno reactive for phosphorylated (active) IRE1α has also been detected in the hippocampus of individuals with the disease.20
Good evidence exists for activation of PERK in laboratory models of Alzheimer’s disease. Increased BiP expression seen on application of exogenous amyloid β to primary cortical neurons suggests that accumulation of the peptide could directly activate ER stress signalling.21,22 Moreover, exogenous amyloid β has been reported to increase the phosphorylation of PERK and eIF2α in cultured neuronal cells,22,107 whereas silencing of PERK by small interfering RNA in amyloid-β42-treated cells limits eIF2α phosphorylation and enhances cell death.22 Additionally, amyloid-β42 treatment induces CHOP expression both in cultured cells and in rabbit hippocampus,23,108 and previous treatment of cells with CHOP antisense RNA improves survival after exposure to amyloid β, which suggests a role for CHOP in amyloid-β-mediated cell death.109 In

www.thelancet.com/neurology Vol 12 January 2013 111
Review
Drosophila melanogaster neurons and mammalian PC12 cells, exposure to amyloid β induces XBP1 mRNA splicing, and in both models overexpression of spliced XBP1 is protective, whereas knockdown of XBP1 exacerbates amyloid-β toxicity.110
PC12 cells that overexpress mutant forms of presenilin 1 (PSEN1), a component of the γ-secretase that has been linked to familial Alzheimer’s disease, show increased phosphorylation of eIF2α, as do hippocampal homogenates from PSEN1 mutant knock-in mice.111 PC12 cells that express mutant PSEN1 have raised levels of CHOP, as do knock-in mice that express mutant PSEN1.111 The nature of the interaction between PSEN1 and ER stress has yet to be fully elucidated, although PSEN1 and the SERCA calcium pump do physically interact with one another.24 Disordered calcium homeo-stasis is a potent cause of ER stress, and much evidence suggests that calcium signalling is perturbed in Alz-heimer’s disease.25,112 ER stress induced by tunicamycin, a glycosylation inhibitor often used to induce ER stress in
vitro, increases PSEN1 expression in several cell lines, including neuroblastoma.24 Evidence suggests that phosphorylation of eIF2α by PERK during ER stress can increase levels of BACE1 (β-site APP cleaving enzyme 1) and thereby promote amyloid formation,26 which suggests that the interaction between Alzheimer’s disease and ER stress might prove to be bidirectional.
ER stress changes amyloid precursor protein (APP) localisation, processing, and degradation by the ERAD machinery (fi gure 4).113 During ER stress, immature forms of APP bind to BiP, and when BiP is overexpressed, amyloid-β generation is reduced because APP is retained in earlier compartments of the secretory pathway.113 The ERAD ubiquitin ligase HRD1 (also known as SYVN1) promotes ubiquitination and degradation of APP in human embryonic kidney 293 cells, which reduces the gener ation of both amyloid β40 and amyloid β42.
2 Additionally, an intronic polymorphism (IVS3–88A→G) of SEL1L, another component of the ERAD system, has been associated
Figure 3: Alzheimer’s disease and endoplasmic reticulum stressIn health (left), amyloid precursor protein (APP) is cleaved sequentially by β-secretase then γ-secretase in the trans-Golgi network (TGN) and early endosomes, which mainly generates non-toxic amyloid β40 (Aβ40). APP is recycled via the endosomal pathway. In Alzheimer’s disease (right), APP is cleaved by α, β and γ-secretases, most likely in the ER, to generate the toxic Aβ42 form (triangles), rather than the Aβ40 form (squares) generated by secretase activity within the TGN. Exogenous Aβ42 causes endoplasmic reticulum (ER) stress via a poorly understood mechanism. In some cases of familial Alzheimer’s disease, mutations in PSEN1 (which encodes presenilin 1) impair the traffi cking of APP, and thereby enhance secretion of Aβ42. Mutated presenilin 1 might also lead to APP processing within the ER.
Health
Exogenous Aβ42 induces ER stress (route of ER access is unclear)
PSEN1 mutations impair APP trafficking and enhance Aβ42 production in the TGN
Early endosomal and TGN-localised γ-secretase (PSEN1 and other components) co-operate with β-secretase to process APP to Aβ40
Aberrant secretase activity in ER might generate Aβ42
PPP
Alzheimer’s disease
Aβ40
Aβ42
APPSecretase
Endoplasmic reticulum
Golgi
Nucleus
PERK
eIF2α
Endosomes

112 www.thelancet.com/neurology Vol 12 January 2013
Review
with Alzheimer’s disease in an Italian popu lation.114 These hints that ERAD is important in the pathogenesis of Alzheimer’s disease need further valid ation. However, evidence for raised unfolded-protein-response signal-ling in the disease is accumulating and, combined with the support from model systems, provides a potential mechanism by which amyloid β and mutants of PSEN1 might induce ER stress. Such mechanisms point to new potential therapeutic targets for a currently incurable disease.
Parkinson’s diseaseAbout 2% of the population lose dopaminergic neurons in their nigrostriatal pathway with age and so develop Parkinson’s disease, which is characterised by hypokinesia, rigidity, and tremor. Post-mortem studies identify character-istic Lewy body inclusions in many aff ected brains, which suggests aberrant protein disposal as a possible cause, whereas toxins that cause parkinsonism, such as 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP), rotenone, and 6-hydroxydopamine (6-OHDA), impair mitochondrial function and promote ER stress.27,28,115–117
Several lines of evidence suggest a role for ER stress in the pathogenesis of Parkinson’s disease. For example,
excess dopamine is toxic to PC12 cells and is associated with the induction of ER chaperones.29 The brains of patients with sporadic Parkinson’s disease have raised levels of phosphorylated PERK and eIF2α compared with controls.118 Mouse Perk–/– sympathetic neurons are hypersensitive to 6-OHDA,118 and overexpression of spliced XBP1 is protective in cultured cells and animals treated with 1-methyl-4-phenylpyridinium or MPTP.27 CHOP ex pression is increased by both 6-OHDA and MPTP, and knockout of the Chop gene is protective in mice treated with 6-OHDA.28
In Japanese and European families with the juvenile-onset autosomal recessive variant of Parkinson’s disease, classic genetic mapping identifi ed the disease-associated gene PARK2.119 This gene encodes the ubiquitin ligase parkin, the activity of which is lost with disease-associated mutations.120 Parkin has been reported to localise to the ER and to be up regulated during the unfolded protein response;30 however, a major function of parkin seems to involve the targeting of defective mitochondria for destruction by autophagy.121 Impaired mitochondrial function in patients with PARK2 mutations could possibly manifest as ER stress because of defects in energy supply during ER protein folding.
However, another component of Lewy bodies, α-synuclein, can be found throughout the cell, including in the ER lumen, and might also be a target of parkin.122 Mutations of α-synuclein cause an autosomal dominant form of Parkinson’s disease,123 and the glycosylated ER form of α-synuclein is ubiquitinated by parkin and accumulates in a non-ubiquitinated form in the brains of individuals with parkin defi ciency.122 Overexpression of α-synuclein induces ER stress,124 perhaps through binding to BiP,31 and protein disulfi de isomerase seems to impair α-synuclein fi bril formation.124 Oligo mers of mutant α-synuclein accumulate within the ER in a mouse model of Parkinson’s disease and trigger the unfolded protein response, albeit with little PERK activation.32 When eIF2α phosphorylation was augmented with salubrinal, the accumulation of α-synuclein oligomers was ameli orated and disease onset was delayed. Modulation of ER stress could be benefi cial in Parkinson’s disease; 4-phenyl butyric acid, which has chemical chaperone activity that can protect cells from ER stress, rescues cells from the neurotoxicity of rotenone,33 and pre conditioning neuroblastoma cells with ER stress to upregulate chaperones can protect the cells from subsequent challenge with 6-OHDA.34
Huntington’s diseaseHuntington’s disease is an inherited autosomal dom-inant disorder that is characterised by motor dysfunction, psychiatric disturbances, and intellectual decline. It is caused by expansions of CAG repeats within the HTT gene that encodes huntingtin, and so is an archetypal polyQ disease. In health, the HTT gene contains 6–35 repeats, whereas patients with Huntington’s disease
Figure 4: Endoplasmic reticulum-associated protein degradation of amyloid precursor proteinNewly synthesised proteins, including amyloid precursor protein (APP), interact with endoplasmic reticulum (ER) chaperones during several folding cycles until they reach their native folded state, whereupon they are packaged into COPII vesicles for onward traffi cking. Overexpression of BiP can increase retention of APP in the ER. If protein folding fails, the terminally misfolded client is targeted for degradation via the ER-associated protein degradation (ERAD) pathway. The interaction of glycoprotein substrates with both the folding machinery and the ERAD machinery is governed by cycles of demannosylation and remannosylation. Eventually, low-mannose species can interact with ER degradation enhancer, mannosidase α-like (EDEM) and be targeted for dislocation from the ER lumen into the cytosol. This process is incompletely understood and shows substrate-specifi c diff erences, but for APP involves a complex that contains HRD1 and SEL1L. Once in the cytosol, the ERAD substrate is ubiquitinated and eventually degraded by the proteasome.
Successful folding
Endoplasmic reticulum
Unsuccessful folding
SEL1L HRD1
Ubiquitin
Polyubiquitin
COPII vesicles
APP
Proteasome
BiP

www.thelancet.com/neurology Vol 12 January 2013 113
Review
have more than 40, which generates a protein prone to aggregation. Although this aggregation takes place in the cytosol, it has been shown to provoke ER stress.125
That perturbations of protein folding within one organelle can be propagated to another is now generally accepted. This idea of cell-wide protein homoeostasis, or proteostasis, could be especially relevant to Hun tington’s disease and other polyQ disorders in which cytoplasmic aggregates trigger the unfolded protein response.36 The precise mechanism that links aggre gation in the cytosol with malfunction of the ER is poorly understood; however, accumulated polyQ proteins, including mutant huntingtin, have been shown to bind components of the ERAD machinery and impair their function.35,37 Huntingtin could be a target of the ERAD ubiquitin ligase HRD1, since HRD1 can ameliorate the toxic eff ects of the polyQ protein.37 Through sequestration of HRD1 and other components, huntingtin might reduce the capacity of a neuron to cope with misfolded proteins by ERAD, and thereby render it more vulnerable to ER stress.35 Additionally, whereas chronic ER stress induces cell death in some circumstances, it might also impair intracellular signalling in viable cells. Experimental data from 2010 suggest that ER calcium release via the IP3R channel is impaired by ER stress in Huntington’s disease, because of the loss of a stimulatory interaction between this channel and BiP.38
Amyotrophic lateral sclerosisAmyotrophic lateral sclerosis involves a progressive loss of motor neurons, which causes an irreversible decline of muscle power and is usually fatal within 5 years. The cause remains largely unknown, although 5–10% of cases show a dominant pattern of inheritance, which is suggestive of a toxic gain of function (so-called familial amyotrophic lateral sclerosis). Of these, about a fi fth are caused by mutations in the copper–zinc superoxide dismutase 1 (SOD1) that accumulates as fi brils in Lewy-body-like hyaline inclusions—the histological hallmark of familial amyotrophic lateral sclerosis.
Post-mortem brains from patients with amyotrophic lateral sclerosis show increased concentrations of mark-ers of ER stress.39 Furthermore, models of familial dis-ease implicate ER stress in its pathogenesis. Although SOD1 was thought to be an abundant cytosolic protein, it is also secreted by several cell types and colocalises with ER markers.126 When expressed in cultured cells or in animal neuronal tissue, mutant SOD1 accumulates within the ER and upregulates targets of the unfolded protein response.40–42
Activation of PERK can be detected in animals before the development of overt disease.43 Three models of familial amyotrophic lateral sclerosis have been used to show that a subset of motor neurons are more sensitive to the toxic eff ects of mutant SOD1 and show evidence of ER stress at a younger age.44 When one allele of Perk is deleted in mice that express a mutant form of SOD1,
onset of disease is accelerated, which suggests that phosphorylation of eIF2α is protective in this disorder.45 This idea is lent support by the protection of vulnerable neurons in animals treated with salubrinal.44
Surprisingly, deletion of neuronal XBP1 delays the onset of disease and extends life in a mouse model of familial amyotrophic lateral sclerosis.127 In cultured neurons, depletion of XBP1 reduces SOD1 aggregates and toxic eff ects through enhanced autophagy, which might plausibly account for this apparent neuroprotec tion through increased clearance of aggregated protein. Similarly, enhanced degradation of SOD1 by overex pression of the ERAD protein derlin-1 ameliorates SOD1-induced cell death.128 However, mutant SOD1 interacts with derlin-1 and so impairs ERAD and promotes ER stress.46
Multiple sclerosisRegulated phosphorylation of eIF2α is important for the health of myelinated neurons within the central nervous system, as shown by the CACH/VWD syndrome and Pelizaeus-Merzbacher disease. Regulated eIF2α phos-phorylation is also important in multiple sclerosis, in which the timing of phosphorylation relative to the onset of disease determines its consequences. In experimental autoimmune encephalitis, ER stress can hasten or delay disease progression if induced after of before disease onset, respectively16,17 (an occurrence previously seen with interferon gamma treatment). Indeed, both experimental autoimmune encephalitis and interferon gamma cause phosphor ylation of eIF2α by PERK. Only when animals are exposed to interferon gamma before induction of experimental autoimmune encephalitis is a protective preconditioning seen and, importantly, the protection is lost if one allele of Perk is deleted.16 By contrast, interferon gamma treatment of animals with estab lished experimental autoimmune encephalitis exacer bates their ER stress and worsens demyelination through impaired oligodendrocyte survival.
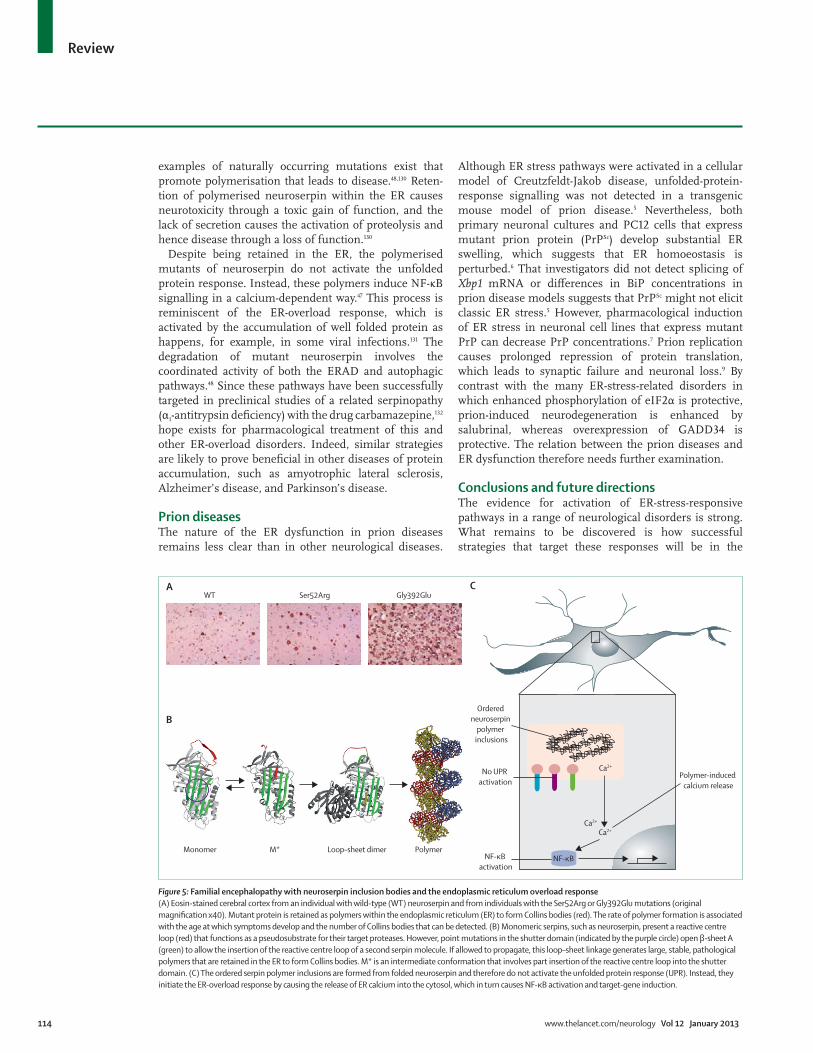
Familial encephalopathy with neuroserpin inclusion bodies10 years ago, the fi rst cases of a new familial encephalopathy with inclusion bodies, FENIB, were described in two families in the USA.129 The primary symptom of this autosomal dominant neurodegenera tive disease was presenile dementia. In each family, eosinophilic neuronal inclusions were seen in deep layers of the cerebral cortex and in the substantia nigra (fi gure 5). After isolation and sequencing of these inclusions, they were reported to be composed of a single protein, neuroserpin. When expressed in neurons, the mutants of neuroserpin accumulate to form acid-Schiff -positive, diastase-resistant inclusions in the ER (neuroserpin inclusion bodies or Collins bodies).129
Neuroserpin is a member of the serine protease inhibitor (serpin) superfamily. Serpins are capable of polymerisation under some circumstances, and many
114 www.thelancet.com/neurology Vol 12 January 2013
Review
examples of naturally occurring mutations exist that promote polymerisation that leads to disease.48,130 Reten-tion of polymerised neuroserpin within the ER causes neurotoxicity through a toxic gain of function, and the lack of secretion causes the activation of proteolysis and hence disease through a loss of function.130
Despite being retained in the ER, the polymerised mutants of neuroserpin do not activate the unfolded protein response. Instead, these polymers induce NF-κB signalling in a calcium-dependent way.47 This process is reminiscent of the ER-overload response, which is activated by the accumulation of well folded protein as happens, for example, in some viral infections.131 The degradation of mutant neuroserpin involves the coordinated activity of both the ERAD and autophagic pathways.48 Since these pathways have been successfully targeted in preclinical studies of a related serpinopathy (α1-antitrypsin defi ciency) with the drug carbamazepine,132 hope exists for pharmacological treat ment of this and other ER-overload disorders. Indeed, similar strategies are likely to prove benefi cial in other diseases of protein accumulation, such as amyotrophic lateral sclerosis, Alzheimer’s disease, and Parkinson’s disease.
Prion diseasesThe nature of the ER dysfunction in prion diseases remains less clear than in other neurological diseases.
Although ER stress pathways were activated in a cellular model of Creutzfeldt-Jakob disease, un folded-protein-response signalling was not detected in a transgenic mouse model of prion disease.5 Nevertheless, both primary neuronal cultures and PC12 cells that express mutant prion protein (PrPSc) develop substantial ER swelling, which suggests that ER homoeostasis is perturbed.6 That investigators did not detect splicing of Xbp1 mRNA or diff erences in BiP concentrations in prion disease models suggests that PrPSc might not elicit classic ER stress.5 However, pharmacological induction of ER stress in neuronal cell lines that express mutant PrP can decrease PrP concentrations.7 Prion replication causes prolonged repression of protein translation, which leads to synaptic failure and neuronal loss.9 By contrast with the many ER-stress-related disorders in which enhanced phosphorylation of eIF2α is protective, prion-induced neurodegeneration is enhanced by salubrinal, whereas overexpression of GADD34 is protective. The relation between the prion diseases and ER dysfunction therefore needs further examination.
Conclusions and future directionsThe evidence for activation of ER-stress-responsive pathways in a range of neurological disorders is strong. What remains to be discovered is how successful strategies that target these responses will be in the
Figure 5: Familial encephalopathy with neuroserpin inclusion bodies and the endoplasmic reticulum overload response(A) Eosin-stained cerebral cortex from an individual with wild-type (WT) neuroserpin and from individuals with the Ser52Arg or Gly392Glu mutations (original magnifi cation x40). Mutant protein is retained as polymers within the endoplasmic reticulum (ER) to form Collins bodies (red). The rate of polymer formation is associated with the age at which symptoms develop and the number of Collins bodies that can be detected. (B) Monomeric serpins, such as neuroserpin, present a reactive centre loop (red) that functions as a pseudosubstrate for their target proteases. However, point mutations in the shutter domain (indicated by the purple circle) open β-sheet A (green) to allow the insertion of the reactive centre loop of a second serpin molecule. If allowed to propagate, this loop–sheet linkage generates large, stable, pathological polymers that are retained in the ER to form Collins bodies. M* is an intermediate conformation that involves part insertion of the reactive centre loop into the shutter domain. (C) The ordered serpin polymer inclusions are formed from folded neuroserpin and therefore do not activate the unfolded protein response (UPR). Instead, they initiate the ER-overload response by causing the release of ER calcium into the cytosol, which in turn causes NF-κB activation and target-gene induction.
Ca2+
Ca2+Ca2+
NF-κB
No UPR activation
Ordered neuroserpin
polymer inclusions
Polymer-induced calcium release
NF-κB activation
A
B
CGly392GluSer52ArgWT
PolymerLoop–sheet dimerM*Monomer

www.thelancet.com/neurology Vol 12 January 2013 115
Review
treatment of neurological disorders. Research into unfolded-protein-response signalling has matured to a point at which small-molecule inhibitors of its com-ponents are under development.
The muscle relaxant dantrolene targets IP3R channels of the ER to prevent the release of calcium. This drug is neuroprotective in models of Huntington’s disease133 and in cerebral hypoxia and reperfusion injury, in which its action is associated with reduced ER stress.134 Prolongation of the phosphorylation of eIF2α and the resultant delay in the recovery of protein translation with agents such as salubrinal or guanabenz is another promising approach.72,73 Salubrinal has already been shown to reduce the accumulation of α-synuclein in models of amyotrophic lateral scler-osis,32,135 to reduce the toxicity of amyloid β to primary neurons,22 and to reduce ER stress in models of cerebral ischaemia–reperfusion and Huntington’s disease.1,136 Agents that inhibit IRE1 have also been developed, including quinotrierixin137 and 8-formyl-7-hydroxy-4-methylcoumarin (abbreviated to 4μ8C).138 Both agents selectively inhibit the splicing of XBP1 mRNA, and 4μ8C prevents expansion of the ER without obvious toxic eff ects, which might have therapeutic value in preventing the formation of protein inclusions in neurons.
The list of neurological disorders with impaired ER protein folding is already long and seems set to grow further for some time to come. The neurologist and neuroscientist alike will therefore benefi t from an appreciation of the signalling pathways that commu-nicate this failure of ER function to the rest of the cell, since these insights are likely to inform research, diagnosis, and ultimately therapy.
ContributorsBDR and SJM planned and wrote the Review. All authors commented
on the fi nal draft. BDR reviewed articles for and wrote the sections
about stroke, Parkinson’s disease, Huntington’s disease, multiple
sclerosis, and prion diseases and prepared fi gure 1. AJK reviewed
articles for and contributed to the section about Alzheimer’s disease
and contributed to fi gure 3. EM helped to draft the section about
familial encephalopathy with neuroserpin inclusion bodies (FENIB).
DCC helped to draft the section about Alzheimer’s disease. DAL
helped to draft the section about FENIB. SJM reviewed articles about
endoplasmic reticulum stress and supervised the preparation of
the manuscript.
Search strategy and selection criteria
We searched PubMed with the terms “endoplasmic reticulum” AND (“brain” OR “neurological”) AND “disease”. We included only articles published in English. Titles and abstracts were screened and relevant papers were selected for detailed assessment. The bibliographies of the selected articles were also screened for additional sources. Our search covered all indexed articles published up to Sept 24, 2012, although whenever possible priority has been given to articles published in the past 5 years.
Confl icts of interestWe declare that we have no confl icts of interest.
AcknowledgmentsThe laboratories of DCC, DAL, and SJM are funded by grants from the
UK Medical Research Council (MRC) and Alzheimer’s Research UK
(formerly Alzheimer’s Research Trust), and by Papworth NHS Trust. AJK
is a Wellcome Trust PhD student and SJM is an MRC Senior Clinical
Fellow (G1002610). DCC is an Alzheimer’s Research UK Senior Research
Fellow and is supported by the MRC (G0700990) and the UK
Engineering and Physical Sciences Research Council. EM is funded by
the Institut Pasteur–Cenci Bolognetti Foundation and by the Telethon
Fo undation.
References1 Nakka VP, Gusain A, Raghubir R. Endoplasmic reticulum stress
plays critical role in brain damage after cerebral ischemia/reperfusion in rats. Neurotox Res 2010; 17: 189–202.
2 Kaneko M, Koike H, Saito R, Kitamura Y, Okuma Y, Nomura Y. Loss of HRD1-mediated protein degradation causes amyloid precursor protein accumulation and amyloid-β generation. J Neurosci 2010; 30: 3924–32.
3 Lee JH, Won SM, Suh J, et al. Induction of the unfolded protein response and cell death pathway in Alzheimer’s disease, but not in aged Tg2576 mice. Exp Mol Med 2010; 42: 386–94.
4 Honjo Y, Ito H, Horibe T, Takahashi R, Kawakami K. Protein disulfi de isomerase-immunopositive inclusions in patients with Alzheimer disease. Brain Res 2010; 1349: 90–96.
5 Quaglio E, Restelli E, Garofoli A, et al. Expression of mutant or cytosolic PrP in transgenic mice and cells is not associated with endoplasmic reticulum stress or proteasome dysfunction. PLoS One 2011; 6: e19339.
6 Dossena S, Imeri L, Mangieri M, et al. Mutant prion protein expression causes motor and memory defi cits and abnormal sleep patterns in a transgenic mouse model. Neuron 2008; 60: 598–609.
7 Nunziante M, Ackermann K, Dietrich K, et al. Proteasomal dysfunction and endoplasmic reticulum stress enhance traffi cking of prion protein aggregates through the secretory pathway and increase accumulation of pathologic prion protein. J Biol Chem 2011; 286: 33942–53.
8 Rane NS, Kang SW, Chakrabarti O, Feigenbaum L, Hegde RS. Reduced translocation of nascent prion protein during ER stress contributes to neurodegeneration. Dev Cell 2008; 15: 359–70.
9 Moreno JA, Radford H, Peretti D, et al. Sustained translational repression by eIF2α-P mediates prion neurodegeneration. Nature 2012; 485: 507–11.
10 Kantor L, Harding HP, Ron D, et al. Heightened stress response in primary fi broblasts expressing mutant eIF2B genes from CACH/VWM leukodystrophy patients. Hum Genet 2005; 118: 99–106.
11 Leegwater PA, Vermeulen G, Konst AA, et al. Subunits of the translation initiation factor eIF2B are mutant in leukoencephalopathy with vanishing white matter. Nat Genet 2001; 29: 383–88.
12 Richardson JP, Mohammad SS, Pavitt GD. Mutations causing childhood ataxia with central nervous system hypomyelination reduce eukaryotic initiation factor 2B complex formation and activity. Mol Cell Biol 2004; 24: 2352–63.
13 Southwood CM, Garbern J, Jiang W, Gow A. The unfolded protein response modulates disease severity in Pelizaeus-Mezbacher disease. Neuron 2002; 36: 585–96.
14 D’Urso D, Prior R, Greiner-Petter R, Gabreels-Festen AA, Muller HW. Overloaded endoplasmic reticulum-Golgi compartments, a possible pathomechanism of peripheral neuropathies caused by mutations of the peripheral myelin protein PMP22. J Neurosci 1998; 18: 731–40.
15 Pennuto M, Tinelli E, Malaguti M, et al. Ablation of the UPR-mediator CHOP restores motor function and reduces demyelination in Charcot-Marie-Tooth 1B mice. Neuron 2008; 57: 393–405.
16 Lin W, Bailey SL, Ho H, et al. The integrated stress response prevents demyelination by protecting oligodendrocytes against immune-mediated damage. J Clin Invest 2007; 117: 448–56.
17 Lin W, Kemper A, Dupree JL, Harding HP, Ron D, Popko B. Interferon-γ inhibits central nervous system remyelination through a process modulated by endoplasmic reticulum stress. Brain 2006; 129: 1306–18.

116 www.thelancet.com/neurology Vol 12 January 2013
Review
18 Zhao L, Longo-Guess C, Harris BS, Lee JW, Ackerman SL. Protein accumulation and neurodegeneration in the woozy mutant mouse is caused by disruption of SIL1, a cochaperone of BiP. Nat Genet 2005; 37: 974–79.
19 Zhao L, Rosales C, Seburn K, Ron D, Ackerman SL. Alteration of the unfolded protein response modifi es neurodegeneration in a mouse model of Marinesco-Sjögren syndrome. Hum Mol Genet 2010; 19: 25–35.
20 Hoozemans JJ, van Haastert ES, Nijholt DA, Rozemuller AJ, Eikelenboom P, Scheper W. The unfolded protein response is activated in pretangle neurons in Alzheimer’s disease hippocampus. Am J Pathol 2009; 174: 1241–51.
21 Resende R, Ferreiro E, Pereira C, Oliveira CR. ER stress is involved in Aβ-induced GSK-3β activation and tau phosphorylation. J Neurosci Res 2008; 86: 2091–99.
22 Lee DY, Lee K-S, Lee HJ, et al. Activation of PERK signaling attenuates Aβ-mediated ER stress. PLoS One 2010; 5: e10489.
23 Song S, Lee H, Kam TI, et al. E2-25K/Hip-2 regulates caspase-12 in ER stress-mediated Aβ neurotoxicity. J Cell Biol 2008; 182: 675–84.
24 Jin H, Sanjo N, Uchihara T, et al. Presenilin-1 holoprotein is an interacting partner of sarco endoplasmic reticulum calcium-ATPase and confers resistance to endoplasmic reticulum stress. J Alzheimers Dis 2010; 20: 261–73.
25 Stutzmann GE, Mattson MP. Endoplasmic reticulum Ca²+ handling in excitable cells in health and disease. Pharmacol Rev 2011; 63: 700–27.
26 O’Connor T, Sadleir KR, Maus E, et al. Phosphorylation of the translation initiation factor eIF2α increases BACE1 levels and promotes amyloidogenesis. Neuron 2008; 60: 988–1009.
27 Sado M, Yamasaki Y, Iwanaga T, et al. Protective eff ect against Parkinson’s disease-related insults through the activation of XBP1. Brain Res 2009; 1257: 16–24.
28 Silva RM, Ries V, Oo TF, et al. CHOP/GADD153 is a mediator of apoptotic death in substantia nigra dopamine neurons in an in vivo neurotoxin model of parkinsonism. J Neurochem 2005; 95: 974–86.
29 Dukes AA, Van Laar VS, Cascio M, Hastings TG. Changes in endoplasmic reticulum stress proteins and aldolase A in cells exposed to dopamine. J Neurochem 2008; 106: 333–46.
30 Bouman L, Schlierf A, Lutz AK, et al. Parkin is transcriptionally regulated by ATF4: evidence for an interconnection between mitochondrial stress and ER stress. Cell Death Diff er 2011; 18: 769–82.
31 Bellucci A, Navarria L, Zaltieri M, et al. Induction of the unfolded protein response by α-synuclein in experimental models of Parkinson›s disease. J Neurochem 2011; 116: 588–605.
32 Colla E, Coune P, Liu Y, et al. Endoplasmic reticulum stress is important for the manifestations of α-synucleinopathy in vivo. J Neurosci 2012; 32: 3306–20.
33 Inden M, Kitamura Y, Takeuchi H, et al. Neurodegeneration of mouse nigrostriatal dopaminergic system induced by repeated oral administration of rotenone is prevented by 4-phenylbutyrate, a chemical chaperone. J Neurochem 2007; 101: 1491–504.
34 Hara H, Kamiya T, Adachi T. Endoplasmic reticulum stress inducers provide protection against 6-hydroxydopamine-induced cytotoxicity. Neurochem Int 2011; 58: 35–43.
35 Duennwald ML, Lindquist S. Impaired ERAD and ER stress are early and specifi c events in polyglutamine toxicity. Genes Dev 2008; 22: 3308–19.
36 Lajoie P, Snapp EL. Changes in BiP availability reveal hypersensitivity to acute endoplasmic reticulum stress in cells expressing mutant huntingtin. J Cell Sci 2011; 124: 3332–43.
37 Bennett EJ, Shaler TA, Woodman B, et al. Global changes to the ubiquitin system in Huntington’s disease. Nature 2007; 448: 704–08.
38 Higo T, Hamada K, Hisatsune C, et al. Mechanism of ER stress-induced brain damage by IP3 receptor. Neuron 2010; 68: 865–78.
39 Sasaki S. Endoplasmic reticulum stress in motor neurons of the spinal cord in sporadic amyotrophic lateral sclerosis. J Neuropathol Exp Neurol 2010; 69: 346–55.
40 Kikuchi H, Almer G, Yamashita S, et al. Spinal cord endoplasmic reticulum stress associated with a microsomal accumulation of mutant superoxide dismutase-1 in an ALS model. Proc Natl Acad Sci USA 2006; 103: 6025–30.
41 Kieran D, Woods I, Villunger A, Strasser A, Prehn JH. Deletion of the BH3-only protein puma protects motoneurons from ER stress-induced apoptosis and delays motoneuron loss in ALS mice. Proc Natl Acad Sci USA 2007; 104: 20606–11.
42 Walker AK, Farg MA, Bye CR, McLean CA, Horne MK, Atkin JD. Protein disulphide isomerase protects against protein aggregation and is S-nitrosylated in amyotrophic lateral sclerosis. Brain 2010; 133: 105–16.
43 Nagata T, Ilieva H, Murakami T, et al. Increased ER stress during motor neuron degeneration in a transgenic mouse model of amyotrophic lateral sclerosis. Neurol Res 2007; 29: 767–71.
44 Saxena S, Cabuy E, Caroni P. A role for motoneuron subtype-selective ER stress in disease manifestations of FALS mice. Nat Neurosci 2009; 12: 627–36.
45 Wang L, Popko B, Roos RP. The unfolded protein response in familial amyotrophic lateral sclerosis. Hum Mol Genet 2011; 20: 1008–15.
46 Nishitoh H, Kadowaki H, Nagai A, et al. ALS-linked mutant SOD1 induces ER stress- and ASK1-dependent motor neuron death by targeting Derlin-1. Genes Dev 2008; 22: 1451–64.
47 Davies MJ, Miranda E, Roussel BD, Kaufman RJ, Marciniak SJ, Lomas DA. Neuroserpin polymers activate NF-κB by a calcium signalling pathway that is independent of the unfolded protein response. J Biol Chem 2009; 284: 18202–09.
48 Kroeger H, Miranda E, MacLeod I, et al. Endoplasmic reticulum-associated degradation (ERAD) and autophagy cooperate to degrade polymerogenic mutant serpins. J Biol Chem 2009; 284: 22793–802.
49 Kang YJ, Lu MK, Guan KL. The TSC1 and TSC2 tumor suppressors are required for proper ER stress response and protect cells from ER stress-induced apoptosis. Cell Death Diff er 2011; 18: 133–44.
50 Liu G, Guo H, Guo C, Zhao S, Gong D, Zhao Y. Involvement of IRE1α signaling in the hippocampus in patients with mesial temporal lobe epilepsy. Brain Res Bull 2011; 84: 94–102.
51 Fonseca SG, Ishigaki S, Oslowski CM, et al. Wolfram syndrome 1 gene negatively regulates ER stress signaling in rodent and human cells. J Clin Invest 2010; 120: 744–55.
52 Chen P, Burdette AJ, Porter JC, et al. The early-onset torsion dystonia-associated protein, torsinA, is a homeostatic regulator of endoplasmic reticulum stress response. Hum Mol Genet 2010; 19: 3502–15.
53 Wang F, Agnello G, Sotolongo N, Segatori L. Ca2+ homeostasis modulation enhances the amenability of L444P glucosylcerebrosidase to proteostasis regulation in patient-derived fi broblasts. ACS Chem Biol 2011; 6: 158–68.
54 Osada N, Kosuge Y, Kihara T, Ishige K, Ito Y. Apolipoprotein E-defi cient mice are more vulnerable to ER stress after transient forebrain ischemia. Neurochem Int 2009; 54: 403–09.
55 Oida Y, Shimazawa M, Imaizumi K, Hara H. Involvement of endoplasmic reticulum stress in the neuronal death induced by transient forebrain ischemia in gerbil. Neuroscience 2008; 151: 111–09.
56 Osada N, Kosuge Y, Ishige K, Ito Y. Characterization of neuronal and astroglial responses to ER stress in the hippocampal CA1 area in mice following transient forebrain ischemia. Neurochem Int 2010; 57: 1–7.
57 Ouyang YB, Xu LJ, Emery JF, Lee AS, Giff ard RG. Overexpressing GRP78 infl uences Ca2+ handling and function of mitochondria in astrocytes after ischemia-like stress. Mitochondrion 2011; 11: 279–86.
58 Kudo T, Kanemoto S, Hara H, et al. A molecular chaperone inducer protects neurons from ER stress. Cell Death Diff er 2008; 15: 364–75.
59 Zhu Y, Fenik P, Zhan G, Sanfi llipo-Cohn B, Naidoo N, Veasey SC. Eif-2a protects brainstem motoneurons in a murine model of sleep apnea. J Neurosci 2008; 28: 2168–78.
60 Penas C, Verdu E, Asensio-Pinilla E, et al. Valproate reduces CHOP levels and preserves oligodendrocytes and axons after spinal cord injury. Neuroscience 2011; 178: 33–44.
61 Marciniak SJ, Ron D. Endoplasmic reticulum stress signaling in disease. Physiol Rev 2006; 86: 1133–49.
62 Marciniak SJ, Yun CY, Oyadomari S, et al. CHOP induces death by promoting protein synthesis and oxidation in the stressed endoplasmic reticulum. Genes Dev 2004; 18: 3066–77.
63 Harding H, Zhang Y, Ron D. Translation and protein folding are coupled by an endoplasmic reticulum resident kinase. Nature 1999; 397: 271–74.

www.thelancet.com/neurology Vol 12 January 2013 117
Review
64 Owen CR, Kumar R, Zhang P, McGrath BC, Cavener DR, Krause GS. PERK is responsible for the increased phosphorylation of eIF2α and the severe inhibition of protein synthesis after transient global brain ischemia. J Neurochem 2005; 94: 1235–42.
65 Marciniak SJ, Garcia-Bonilla L, Hu J, Harding HP, Ron D. Activation-dependent substrate recruitment by the eukaryotic translation initiation factor 2 kinase PERK. J Cell Biol 2006; 172: 201–09.
66 Harding H, Zhang Y, Zeng H, et al. An integrated stress response regulates amino acid metabolism and resistance to oxidative stress. Mol Cell 2003; 11: 619–33.
67 Dalton LE, Healey E, Irving J, Marciniak SJ. Phosphoproteins in stress-induced disease. Prog Mol Biol Transl Sci 2012; 106: 189–221.
68 Harding HP, Zhang Y, Zeng H, et al. An integrated stress response regulates amino acid metabolism and resistance to oxidative stress. Mol Cell 2003; 11: 619–33.
69 Hao S, Sharp JW, Ross-Inta CM, et al. Uncharged tRNA and sensing of amino acid defi ciency in mammalian piriform cortex. Science 2005; 307: 1776–78.
70 Maurin AC, Jousse C, Averous J, et al. The GCN2 kinase biases feeding behavior to maintain amino acid homeostasis in omnivores. Cell Metab 2005; 1: 273–77.
71 Harding HP, Calfon M, Urano F, Novoa I, Ron D. Transcriptional and translational control in the mammalian unfolded protein response. Annu Rev Cell Dev Biol 2002; 18: 575–99.
72 Boyce M, Bryant KF, Jousse C, et al. A selective inhibitor of eIF2α dephosphorylation protects cells from ER stress. Science 2005; 307: 935–39.
73 Tsaytler P, Harding HP, Ron D, Bertolotti A. Selective inhibition of a regulatory subunit of protein phosphatase 1 restores proteostasis. Science 2011; 332: 91–94.
74 Kondo S, Murakami T, Tatsumi K, et al. OASIS, a CREB/ATF-family member, modulates UPR signalling in astrocytes. Nat Cell Biol 2005; 7: 186–94.
75 Calfon M, Zeng H, Urano F, et al. IRE1 couples endoplasmic reticulum load to secretory capacity by processing the XBP-1 mRNA. Nature 2002; 415: 92–96.
76 Hosoi T, Ogawa K, Ozawa K. Homocysteine induces X-box-binding protein 1 splicing in the mice brain. Neurochem Int 2010; 56: 216–20.
77 Meusser B, Hirsch C, Jarosch E, Sommer T. ERAD: the long road to destruction. Nat Cell Biol 2005; 7: 766–72.
78 Haynes CM, Titus EA, Cooper AA. Degradation of misfolded proteins prevents ER-derived oxidative stress and cell death. Mol Cell 2004; 15: 767–76.
79 Yoshida H, Matsui T, Hosokawa N, Kaufman RJ, Nagata K, Mori K. A time-dependent phase shift in the Mammalian unfolded protein response. Dev Cell 2003; 4: 265–71.
80 Rissanen A, Sivenius J, Jolkkonen J. Prolonged bihemispheric alterations in unfolded protein response related gene expression after experimental stroke. Brain Res 2006; 1087: 60–66.
81 Morimoto N, Oida Y, Shimazawa M, et al. Involvement of endoplasmic reticulum stress after middle cerebral artery occlusion in mice. Neuroscience 2007; 147: 957–67.
82 Hayashi T, Hamakawa K, Nagotani S, et al. HMG CoA reductase inhibitors reduce ischemic brain injury of Wistar rats through decreasing oxidative stress on neurons. Brain Res 2005; 1037: 52–58.
83 Szydlowska K, Tymianski M. Calcium, ischemia and excitotoxicity. Cell Calcium 2010; 47: 122–29.
84 Coe H, Michalak M. Calcium binding chaperones of the endoplasmic reticulum. Gen Physiol Biophys 2009; 28: F96–103.
85 Nguyen HN, Wang C, Perry DC. Depletion of intracellular calcium stores is toxic to SH-SY5Y neuronal cells. Brain Res 2002; 924: 159–66.
86 Zhang F, Liu CL, Hu BR. Irreversible aggregation of protein synthesis machinery after focal brain ischemia. J Neurochem 2006; 98: 102–12.
87 Althausen S, Mengesdorf T, Mies G, et al. Changes in the phosphorylation of initiation factor eIF-2α, elongation factor eEF-2 and p70 S6 kinase after transient focal cerebral ischaemia in mice. J Neurochem 2001; 78: 779–87.
88 Qi X, Okuma Y, Hosoi T, Nomura Y. Edaravone protects against hypoxia/ischemia-induced endoplasmic reticulum dysfunction. J Pharmacol Exp Ther 2004; 311: 388–93.
89 Shibata M, Hattori H, Sasaki T, Gotoh J, Hamada J, Fukuuchi Y. Activation of caspase-12 by endoplasmic reticulum stress induced by transient middle cerebral artery occlusion in mice. Neuroscience 2003; 118: 491–99.
90 Xue Y, Daly A, Yngvadottir B, et al. Spread of an inactive form of caspase-12 in humans is due to recent positive selection. Am J Hum Genet 2006; 78: 659–70.
91 Hitomi J, Katayama T, Eguchi Y, et al. Involvement of caspase-4 in endoplasmic reticulum stress-induced apoptosis and Aβ-induced cell death. J Cell Biol 2004; 165: 347–56.
92 Kumar R, Krause GS, Yoshida H, Mori K, DeGracia DJ. Dysfunction of the unfolded protein response during global brain ischemia and reperfusion. J Cereb Blood Flow Metab 2003; 23: 462–71.
93 Chihara K, Saito A, Murakami T, et al. Increased vulnerability of hippocampal pyramidal neurons to the toxicity of kainic acid in OASIS-defi cient mice. J Neurochem 2009; 110: 956–65.
94 Tajiri S, Oyadomari S, Yano S, et al. Ischemia-induced neuronal cell death is mediated by the endoplasmic reticulum stress pathway involving CHOP. Cell Death Diff er 2004; 11: 403–15.
95 Hayashi T, Saito A, Okuno S, Ferrand-Drake M, Dodd RL, Chan PH. Damage to the endoplasmic reticulum and activation of apoptotic machinery by oxidative stress in ischemic neurons. J Cereb Blood Flow Metab 2005; 25: 41–53.
96 Benavides A, Pastor D, Santos P, Tranque P, Calvo S. CHOP plays a pivotal role in the astrocyte death induced by oxygen and glucose deprivation. Glia 2005; 52: 261–75.
97 Paschen W, Hayashi T, Saito A, Chan PH. GADD34 protein levels increase after transient ischemia in the cortex but not in the CA1 subfi eld: implications for post-ischemic recovery of protein synthesis in ischemia-resistant cells. J Neurochem 2004; 90: 694–701.
98 McCaig D, Imai H, Gallagher L, et al. Evolution of GADD34 expression after focal cerebral ischaemia. Brain Res 2005; 1034: 51–61.
99 Sokka AL, Putkonen N, Mudo G, et al. Endoplasmic reticulum stress inhibition protects against excitotoxic neuronal injury in the rat brain. J Neurosci 2007; 27: 901–08.
100 Yu Z, Luo H, Fu W, Mattson MP. The endoplasmic reticulum stress-responsive protein GRP78 protects neurons against excitotoxicity and apoptosis: suppression of oxidative stress and stabilization of calcium homeostasis. Exp Neurol 1999; 155: 302–14.
101 Tamatani M, Matsuyama T, Yamaguchi A, et al. ORP150 protects against hypoxia/ischemia-induced neuronal death. Nat Med 2001; 7: 317–23.
102 Luheshi LM, Tartaglia GG, Brorsson AC, et al. Systematic in vivo analysis of the intrinsic determinants of amyloid β pathogenicity. PLoS Biol 2007; 5: e290.
103 Katayama T, Imaizumi K, Sato N, et al. Presenilin-1 mutations downregulate the signalling pathway of the unfolded-protein response. Nat Cell Biol 1999; 1: 479–85.
104 Hoozemans JJ, Veerhuis R, Van Haastert ES, et al. The unfolded protein response is activated in Alzheimer’s disease. Acta Neuropathol 2005; 110: 165–72.
105 Hoozemans JJ, Stieler J, van Haastert ES, et al. The unfolded protein response aff ects neuronal cell cycle protein expression: implications for Alzheimer’s disease pathogenesis. Exp Gerontol 2006; 41: 380–86.
106 Unterberger U, Hoftberger R, Gelpi E, Flicker H, Budka H, Voigtlander T. Endoplasmic reticulum stress features are prominent in Alzheimer disease but not in prion diseases in vivo. J Neuropathol Exp Neurol 2006; 65: 348–57.
107 Seyb KI, Ansar S, Bean J, Michaelis ML. β-amyloid and endoplasmic reticulum stress responses in primary neurons: eff ects of drugs that interact with the cytoskeleton. J Mol Neurosci 2006; 28: 111–23.
108 Ghribi O, Herman MM, DeWitt DA, Forbes MS, Savory J. Aβ(1-42) and aluminum induce stress in the endoplasmic reticulum in rabbit hippocampus, involving nuclear translocation of gadd 153 and NF-κB. Brain Res Mol Brain Res 2001; 96: 30–38.
109 Schapansky J, Olson K, Van Der Ploeg R, Glazner G. NF-κB activated by ER calcium release inhibits Aβ-mediated expression of CHOP protein: enhancement by AD-linked mutant presenilin 1. Exp Neurol 2007; 208: 169–76.
110 Casas-Tinto S, Zhang Y, Sanchez-Garcia J, Gomez-Velazquez M, Rincon-Limas DE, Fernandez-Funez P. The ER stress factor XBP1s prevents amyloid-β neurotoxicity. Hum Mol Genet 2011; 20: 2144–60.

118 www.thelancet.com/neurology Vol 12 January 2013
Review
111 Milhavet O, Martindale JL, Camandola S, et al. Involvement of Gadd153 in the pathogenic action of presenilin-1 mutations. J Neurochem 2002; 83: 673–81.
112 Bezprozvanny I, Mattson MP. Neuronal calcium mishandling and the pathogenesis of Alzheimer’s disease. Trends Neurosci 2008; 31: 454–63.
113 Kudo T, Okumura M, Imaizumi K, et al. Altered localization of amyloid precursor protein under endoplasmic reticulum stress. Biochem Biophys Res Commun 2006; 344: 525–30.
114 Saltini G, Dominici R, Lovati C, et al. A novel polymorphism in SEL1L confers susceptibility to Alzheimer’s disease. Neurosci Lett 2006; 398: 53–58.
115 Bauereis B, Haskins WE, Lebaron RG, Renthal R. Proteomic insights into the protective mechanisms of an in vitro oxidative stress model of early stage Parkinson’s disease. Neurosci Lett 2011; 488: 11–16.
116 Egawa N, Yamamoto K, Inoue H, et al. The endoplasmic reticulum stress sensor, ATF6α, protects against neurotoxin-induced dopaminergic neuronal death. J Biol Chem 2011; 286: 7947–57.
117 Chigurupati S, Wei Z, Belal C, et al. The homocysteine-inducible endoplasmic reticulum stress protein counteracts calcium store depletion and induction of CCAAT enhancer-binding protein homologous protein in a neurotoxin model of Parkinson disease. J Biol Chem 2009; 284: 18323–33.
118 Hoozemans JJ, van Haastert ES, Eikelenboom P, de Vos RA, Rozemuller JM, Scheper W. Activation of the unfolded protein response in Parkinson’s disease. Biochem Biophys Res Commun 2007; 354: 707–11.
119 Kitada T, Asakawa S, Hattori N, et al. Mutations in the parkin gene cause autosomal recessive juvenile parkinsonism. Nature 1998; 392: 605–08.
120 Shimura H, Hattori N, Kubo S, et al. Familial Parkinson disease gene product, parkin, is a ubiquitin-protein ligase. Nat Genet 2000; 25: 302–05.
121 Narendra D, Tanaka A, Suen DF, Youle RJ. Parkin is recruited selectively to impaired mitochondria and promotes their autophagy. J Cell Biol 2008; 183: 795–803.
122 Shimura H, Schlossmacher MG, Hattori N, et al. Ubiquitination of a new form of α-synuclein by parkin from human brain: implications for Parkinson›s disease. Science 2001; 293: 263–69.
123 Kruger R, Kuhn W, Muller T, et al. Ala30Pro mutation in the gene encoding α-synuclein in Parkinson’s disease. Nat Genet 1998; 18: 106–08.
124 Smith WW, Jiang H, Pei Z, et al. Endoplasmic reticulum stress and mitochondrial cell death pathways mediate A53T mutant alpha-synuclein-induced toxicity. Hum Mol Genet 2005; 14: 3801–11.
125 Carnemolla A, Fossale E, Agostoni E, et al. Rrs1 is involved in endoplasmic reticulum stress response in Huntington disease. J Biol Chem 2009; 284: 18167–73.
126 Urushitani M, Sik A, Sakurai T, Nukina N, Takahashi R, Julien JP. Chromogranin-mediated secretion of mutant superoxide dismutase proteins linked to amyotrophic lateral sclerosis. Nat Neurosci 2006; 9: 108–18.
127 Hetz C, Thielen P, Matus S, et al. XBP-1 defi ciency in the nervous system protects against amyotrophic lateral sclerosis by increasing autophagy. Genes Dev 2009; 23: 2294–306.
128 Mori A, Yamashita S, Uchino K, et al. Derlin-1 overexpression ameliorates mutant SOD1-induced endoplasmic reticulum stress by reducing mutant SOD1 accumulation. Neurochem Int 2011; 58: 344–53.
129 Davis RL, Shrimpton AE, Holohan PD, et al. Familial dementia caused by polymerization of mutant neuroserpin. Nature 1999; 401: 376–79.
130 Miranda E, MacLeod I, Davies MJ, et al. The intracellular accumulation of polymeric neuroserpin explains the severity of the dementia FENIB. Hum Mol Genet 2008; 17: 1527–39.
131 Pahl HL, Baeuerle PA. The ER-overload response: activation of NF-κB. Trends Biochem Sci 1997; 22: 63–67.
132 Hidvegi T, Ewing M, Hale P, et al. An autophagy-enhancing drug promotes degradation of mutant α1-antitrypsin Z and reduces hepatic fi brosis. Science 2010; 329: 229–32.
133 Chen X, Wu J, Lvovskaya S, Herndon E, Supnet C, Bezprozvanny I. Dantrolene is neuroprotective in Huntington’s disease transgenic mouse model. Mol Neurodegener 2011; 6: 81.
134 Li F, Hayashi T, Jin G, et al. The protective eff ect of dantrolene on ischemic neuronal cell death is associated with reduced expression of endoplasmic reticulum stress markers. Brain Res 2005; 1048: 59–68.
135 Jiang P, Gan M, Ebrahim AS, Lin WL, Melrose HL, Yen SH. ER stress response plays an important role in aggregation of α-synuclein. Mol Neurodegener 2010; 5: 56.
136 Reijonen S, Putkonen N, Norremolle A, Lindholm D, Korhonen L. Inhibition of endoplasmic reticulum stress counteracts neuronal cell death and protein aggregation caused by N-terminal mutant huntingtin proteins. Exp Cell Res 2008; 314: 950–60.
137 Yamamoto K, Tashiro E, Imoto M. Quinotrierixin inhibited ER stress-induced XBP1 mRNA splicing through inhibition of protein synthesis. Biosci Biotechnol Biochem 2011; 75: 284–88.
138 Cross BC, Bond PJ, Sadowski PG, et al. The molecular basis for selective inhibition of unconventional mRNA splicing by an IRE1-binding small molecule. Proc Natl Acad Sci USA 2012; 109: E869–78.




















