
identification of putative functional motifs in - UNSWorks
154
IDENTIFICATION OF PUTATIVE FUNCTIONAL MOTIFS IN VIRAL PROTEINS ESSENTIAL FOR HUMAN CYTOMEGALOVIRUS (HCMV) DNA REPLICATION Heng Giap Woon Master of Science (Research) 2008
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of identification of putative functional motifs in - UNSWorks

IDENTIFICATION OF PUTATIVE FUNCTIONAL MOTIFS IN
VIRAL PROTEINS ESSENTIAL FOR HUMAN
CYTOMEGALOVIRUS (HCMV) DNA REPLICATION
Heng Giap Woon
Master of Science (Research)
2008

I
ABSTRACT
Human cytomegalovirus (HCMV) is a ubiquitous virus that causes significant morbidity
and mortality in immunocompromised individuals. Although there are prophylactic
treatments available, all current antiviral drugs ultimately target the DNA polymerase,
resulting in the increasing emergence of antiviral resistant strains in the clinical setting.
There is a fundamental need for understanding the role of other essential genes in DNA
replication as a foundation for developing new antiviral treatments that are safe and
which utilize a mechanism of action different to existing therapies. In this study we
looked at six HCMV replication genes encoding for the DNA polymerase accessory
protein (UL44), single stranded DNA binding protein (UL57), primase (UL70), helicase
(UL105), primase-helicase associated protein (UL102), and the putative initiator protein
(UL84) in order to increase our understanding of their role in DNA replication. The aim
of this project was to identify variation within these genes as well as to predict putative
domains and motifs in order to ultimately express and study the functional properties of
the HCMV primase (UL70) through the use of recombinant mutants. Sequencing of
these genes revealed a high degree of conservation between the isolates with amino acid
sequence identity of >97% for all genes. Using ScanProsite software from the Expert
Protein Analysis System (ExPASy) proteomics server, we have mapped putative motifs
throughout these HCMV replication genes. In particular, highly conserved putative N-
linked glycosylation sites were identified in UL105 that were also conserved across 33
homologues as well as several unique motifs including casein kinase II phosphorylation
sites (CKII) in UL105 and UL84, a microbodies signal motif in UL57 and an integrin
binding site in the UL102 helicase-primase associated protein. Our investigations have
also elucidated motif-rich regions of the UL44 DNA polymerase accessory protein,
mapped functionally important domains of the UL105 helicase and identified cysteine
motifs that have implications for folding of the UL70 primase. Taken together, these
findings provide insights to regions of these HCMV replication proteins that are
important for post-translation modification, activation and overall function, and this
information can be utilized to target further research into these proteins and advance the
development of novel antiviral agents that target these processes.

II
ACKNOWLEDGEMENTS
I would like to thank my supervisor, Dr Gillian Scott, for her guidance and
encouragement throughout the project, and to my co-supervisor, Professor Bill
Rawlinson, for his advice and for giving me the opportunity to work in research. I
would also like to extend my thanks to Professor Andrew Lloyd for his advice and
guidance.
I would also like to thank my colleagues at the Virology Research Lab for their
company, friendship, encouragement, advice and humor. In particular, thanks to Sharon
and Min for the countless conversations over lunch.
Thanks to the people at the UNSW Ramaciotti Centre for their technical aid in this
project.
I would like to give a special thanks to Professor Patrick Tam for his insights and advice
on science research in general and to my parents who have supported me throughout.
Finally I would like to give a very special thanks to Ying, who kept me company on the
long nights in the lab and while writing this thesis.

III
TABLE OF CONTENTS
ABSTRACT………………………………………………………………………….....I
ACKNOWLEDGEMENTS………………………………………………………......II
TABLE OF CONTENTS…………………………………………………………….III
LIST OF TABLES…………………………………………………………………....VI
LIST OF FIGURES……………………………………………………………….....VII
COMMONLY USED ABBREVIATIONS………………………………………..VIII
1 INTRODUCTION............................................................................................... 1
1.1 BIOLOGY....................................................................................................... 1
1.1.1 The herpesviridae .................................................................................. 1
1.1.2 The Cytomegaloviruses.......................................................................... 2
1.2 HCMV EPIDEMIOLOGY................................................................................ 3
1.2.1 Transmission ......................................................................................... 4
1.2.2 Congenital infection............................................................................... 5
1.2.3 Immunocompromised infection and disease outcomes............................ 6
1.2.4 Antiviral treatment................................................................................. 6
1.2.5 Antiviral resistance................................................................................ 8
1.3 HCMV GENOME STRUCTURE AND ORGANIZATION ..................................... 13
1.3.1 Open reading frame organization ........................................................ 13
1.4 CMV GROWTH CYCLE ................................................................................ 14
1.4.1 Cell attachment and viral entry............................................................ 15
1.4.2 HCMV replication genes...................................................................... 16
1.4.3 DNA replication................................................................................... 24
1.4.4 Capsid assembly, maturation and egress.............................................. 25
1.4.5 Latency and reactivation...................................................................... 26
1.5 AIMS AND OBJECTIVES ............................................................................... 26

IV
2 REAGENTS AND EQUIPMENT .................................................................... 28
2.1 VIRAL STOCKS ............................................................................................ 28
2.2 DNA EXTRACTION ...................................................................................... 28
2.2.1 Buffers ................................................................................................. 28
2.2.2 Chenicals............................................................................................. 28
2.2.3 Kits...................................................................................................... 29
2.3 POLYMERASE CHAIN REACTION (PCR) ...................................................... 29
2.3.1 Buffers ................................................................................................. 29
2.3.2 Enzymes............................................................................................... 29
2.3.3 Equipment ........................................................................................... 30
2.3.4 Reagents .............................................................................................. 30
2.4 ELECTROPHORESIS ..................................................................................... 30
2.4.1 Buffers ................................................................................................. 30
2.4.2 DNA Markers ...................................................................................... 31
2.4.3 Equipment ........................................................................................... 31
2.4.4 Reagents .............................................................................................. 31
2.5 DNA SEQUENCING ...................................................................................... 32
2.5.1 Equipment ........................................................................................... 32
2.5.2 Reagents .............................................................................................. 32
2.6 RESTRICTION ENZYMES .............................................................................. 32
2.6.1 Buffers ................................................................................................. 32
2.6.2 Enzymes............................................................................................... 33
2.6.3 Reagents .............................................................................................. 33
2.7 CLONING REAGENTS ................................................................................... 33
2.7.1 Antibiotics ........................................................................................... 33
2.7.2 Cells .................................................................................................... 34
2.7.3 Equipment ........................................................................................... 34
2.7.4 Kits...................................................................................................... 35
2.7.5 Media .................................................................................................. 36
2.7.6 Reagents .............................................................................................. 36
2.7.7 Vectors ................................................................................................ 36
2.8 PROTEIN EXPRESSION AND DETECTION ....................................................... 37
2.8.1 Antibodies............................................................................................ 37

V
2.8.2 Buffers ................................................................................................. 37
2.8.3 Equipment ........................................................................................... 38
2.8.4 Kits...................................................................................................... 38
2.8.5 Protein Markers................................................................................... 38
2.8.6 Reagents .............................................................................................. 39
2.8.7 Solutions.............................................................................................. 39
3 IDENTIFICATION OF PUTATIVE FUNCTIONAL MOTIFS IN VIRAL
PROTEINS ESSENTIAL FOR HUMAN CYTOMEGALOVIRUS DNA
REPLICATION ........................................................................................................ 40
3.1 INTRODUCTION ........................................................................................... 40
3.2 METHODS ................................................................................................... 41
3.2.1 Viral stocks and DNA extraction.......................................................... 41
3.2.2 Polymerase chain reaction (PCR) amplification .................................. 41
3.2.3 DNA sequencing and analysis.............................................................. 44
3.2.4 Phylogenetic analysis .......................................................................... 44
3.2.5 Genbank Accession Numbers ............................................................... 44
3.3 RESULTS ..................................................................................................... 45
3.3.1 Strain variation amongst HCMV isolates ............................................. 45
3.3.2 Prediction of potential motifs ............................................................... 45
3.3.3 Strain variation and identification of protein motifs in UL44 ............... 46
3.3.4 Strain variation and identification of protein motifs in UL57 ............... 51
3.3.5 Strain variation and identification of protein motifs in UL70 ............... 60
3.3.6 Strain variation and identification of protein motifs in UL102 ............. 67
3.3.7 Strain variation and identification of protein motifs in UL105 ............. 73
3.3.8 Strain variation and identification of protein motifs in UL84 ............... 80
3.4 DISCUSSION................................................................................................. 85
4 HCMV PRIMASE EXPRESSION ................................................................... 93
4.1 INTRODUCTION ........................................................................................... 93
4.2 METHODS ................................................................................................... 95
4.2.1 Cloning of UL70 into high-copy pBluescript II primase vector............. 95
4.2.2 Construction of baculovirus entry clone for HCMV primase expression
99

VI
4.2.3 In vitro UL70 primase expression using TNT Quick Coupled
Transcription/Translation ................................................................................. 102
4.3 RESULTS ................................................................................................... 108
4.3.1 UL70 was cloned into pBluescript II high-copy vector ....................... 108
4.3.2 Generating Baculovirus entry clones via BP recombination............... 110
4.3.3 UL70 was cloned into pCITE expression vector ................................. 114
4.3.4 UL70 in vitro expression using the TNT system.................................. 115
4.4 DISCUSSION............................................................................................... 117
5 CONCLUDING DISCUSSION AND FUTURE DIRECTIONS................... 119
6 REFERENCES................................................................................................ 121

VII
LIST OF TABLES
Table 1 Classification of Human herpesviruses and pathological outcomes................... 2
Table 2 Sequenced cytomegalovirus (CMV) genomes................................................... 3
Table 3 Epidemiological seroprevalence of HCMV infection........................................ 4
Table 4 Mutations of UL97 and UL27 antiviral resistance .......................................... 11
Table 5 Mutations of UL54 associated with antiviral resistance .................................. 12
Table 6 Essential DNA replication proteins in HCMV and HSV-1.............................. 40
Table 7 Primers used for PCR amplification of HCMV DNA fragments ..................... 43
Table 8 Sequence variation within HCMV replication genes....................................... 45
Table 9 Summary of the putative motifs identified in the HCMV replication genes..... 46
Table 10 Nucleotide primers used for UL70/pBluescript cloning ................................ 96
Table 11 Nucleotides primers used for UL70/pDONR cloning.................................. 100
Table 12 Nucleotide primers used for UL70/pCITE cloning...................................... 103

VIII
LIST OF FIGURES
Figure 1 Arrangement of the HCMV genome.............................................................. 13
Figure 2 CMV growth cycle........................................................................................ 15
Figure 3 Relative positions of genes investigated in this study .................................... 27
Figure 4 HCMV strain alignments of UL44 ................................................................ 50
Figure 5 HCMV strain alignments of UL57 ................................................................ 58
Figure 6 Putative zinc finger domain of UL57............................................................. 59
Figure 7 HCMV strain alignments of UL70 ................................................................ 65
Figure 8 Conserved domains between UL70 homologues. .......................................... 66
Figure 9 HCMV strain alignments for UL102 ............................................................. 72
Figure 10 HCMV strain alignments for UL105 ........................................................... 78
Figure 11 Putative motifs conserved across UL105 homologues ................................. 79
Figure 12 HCMV strain alignments for UL84 ............................................................. 83
Figure 13 Overlapping putative casein kinase-2 (CKII) phosphorylation sites of UL8484
Figure 14 Isolate variation in the HCMV primase-helicase complex. .......................... 91
Figure 15 Insert to vector ratio formula ....................................................................... 96
Figure 16 Overview of cloning UL70 into pBluescript ................................................ 98
Figure 17 pDONR/Zeocin Vector ............................................................................. 100
Figure 18 converting femtomoles to nanograms ........................................................ 101
Figure 19 Overview of UL70/pDONR recombination ............................................... 102
Figure 20 Overview of cloning UL70 into pCITE expression vector ......................... 105
Figure 21 Overview of UL70 in vitro protein expression........................................... 107
Figure 22 pBluescript UL70a .................................................................................... 108
Figure 23 pBluescript UL70b.................................................................................... 109
Figure 24 Screening of entry clone BPR4 ................................................................. 111
Figure 25 The BPR4 truncation removes key motifs from UL70 ............................... 111
Figure 26 Colony PCR screening for entry clone BPR5 ............................................ 112
Figure 27 The BPR5 truncation of UL70................................................................... 113
Figure 28 The BPR8 truncation removes the putative zinc finger from UL70............ 113
Figure 29 UL70/pCITE expression vector................................................................. 114
Figure 30 Expression of UL70 in TNT system .......................................................... 115
Figure 31 Time course expression of UL70 with TNT system................................... 116
Figure 32 Sequence analysis of BPR4 and BPR8 ...................................................... 118

IX
COMMONLY USED ABBREVIATIONS
AIDS Acquired immunodeficiency syndrome
ANGIS Australian National Genomic Information Service
ATP Adenosine triphosphate
bp Base pairs
CDV Cidofovir
CMV Cytomegalovirus
DNA Deoxyribonucleic acid
dNTP Deoxyribonucleoside triphosphate
EBV Epstein-Barr virus
FOS Foscarnet
GCV Ganciclovir
HCMV Human cytomegalovirus
HHV Human Herpesvirus
HIV Human immunodeficiency virus
HSV-1/2 Herpes simplex virus type 1/2
IE Immediate early gene
kbp Kilo-base pairs
MIE Major immediate early gene
mRNA Messenger RNA
NLS Nuclear localization signal

X
OBP Origin binding protein
ORF Open reading frame
PCR Polymerase chain reaction
PEG Polyethylene glycol
RNA Ribonucleic acid
UL
Long unique genome region
US Short unique genome region
vGCV Valganciclovir
VZV Varicella-zoster virus

1
1 Introduction
1.1 Biology
1.1.1 The herpesviridae
The herpesviridae family consists of a large group of host-specific animal viruses that
are ubiquitous to the vertebrate species and in at least one invertebrate (Davison, 2002).
Herpesviruses are defined by their virion morphology, which consists of the core, capsid,
tegument, and envelope (Arvin, 2007). The core consists of a single, linear double-
stranded DNA genome of 125–240 kilo-base pairs (kbp) packaged within an icosahedral
capsid that is imbedded in an inner proteinaceous tegument matrix and an outer
glycoprotein-rich lipid membrane (Davison, 2002). Initial phylogenetic analysis of fish
and mammalian herpesviruses dated the divergence of the Alpha-, Beta- and
Gammaherpesvirinae subfamilies to approximately 180-200 million years ago
(McGeoch, 1995), but the inclusion of avian and reptilian herpesviruses have pushed
back the common ancestor of the herpesvirinae subfamilies to about 400 million years
ago (McGeoch, 2005). Classification into Alpha-, Beta- and Gammaherpesvirinae
subfamilies were based on biological properties such as host range and ability to
establish latent infection in certain cell types. The Alphaherpesvirinae include four
genera (Simplexvirus, Varicellovirus, Mardivirus, and Iltovirus) as well as the reptilian
herpesviruses and are characterized by their relatively rapid replication and ability to
maintain latent infections in sensory ganglia as well as productively infect mucous
membranes and skin tissue (Arvin, 2007). The Betaherpesvirinae include the
Cytomegalovirus, Muromegalovirus, and Roseolovirus lineages and are characterized by
their slower replication rate in vitro relative to other herpesviruses (Arvin, 2007). The
Gammaherpesvirinae are divided into two genera, the Lymphocryptoviridae and
Rhadinoviridae, which primarily infect mammals and are known to induce
lymphoproliferation and tumors in endothelial cells (Arvin, 2007). In addition, there is a
genus (Ictalurivirus) that is unattached to any subfamily as well as a large number of
species not assigned to genera (Arvin, 2007). Currently, there are eight herpesviruses
known to infect humans with pathological consequences (Table 1).

2
The herpesviridae subfamilies share many homologous ‘core genes’ which were
presumably inherited from a common ancestor and are associated with DNA replication,
processing and packaging, capsid assembly and egress, as well as a host of tegument
proteins and surface glycoproteins (Davison, 2002).
Table 1 Classification of Human herpesviruses and pathological outcomes
Common Name Genome Subfamily Pathological Outcomes
(Classification) size (kbp)*
Herpes Simplex Virus 1 (HHV-1) 152 α Predominantly oro-facial lesions,
keratoconjunctivitis
Herpes Simplex Virus 2 (HHV-2) 155 α Predominantly genital lesions
Varicella Zoster Virus (HHV-3) 125 α Chickenpox, shingles
Epstein-Barr Virus (HHV-4) 184 γ Burkitt’s and Hodgkin’s lymphoma
infectious mononucleosis, tumors
Human Cytomegalovirus (HHV-5) ~230 β Vasculature and end-organ disease,
congenital abnormalities, retinitis,
Human Herpesvirus 6 (HHV-6A/B) 159/162 β Exanthem subitum (HHV6A)
Human Herpesvirus 7 (HHV-7) ~149 β Exanthem subitum
Human Herpesvirus 8 (HHV-8) ~141 γ Kaposi’s sarcoma
*in kilobase pairs (Arvin, 2007; Davison, 2002)
1.1.2 The Cytomegaloviruses
The cytomegalovirus genus infects a wide range of mammalian species including
rodents (Rawlinson, 1996; Vink, 2000), primates (Davison, 2003; Hansen, 2003), and
humans (Chee, 1990; Dolan, 2004; Dunn, 2003; Murphy, 2003) with several sequenced
genomes available on public databases (Table 2). The human cytomegalovirus (HCMV)
is the prototype of the betaherpesvirinae subfamily and is the most complex of the eight
human herpesvirus species (Davison, 2003). The first HCMV strains were isolated 50
years ago (Craig, 1957; Rowe, 1956; Smith, 1956) while the genetic content of the virus
has been available for almost two decades (Chee, 1990). However, the mechanism of
HCMV DNA replication and its regulation is not completely understood, with the
function of several genes inferred from studies on the herpes simplex virus (HSV). In
addition, current anti-viral treatments exclusively target only one aspect of the CMV
replication cycle, by competitive inhibition of the DNA polymerase, and with the advent

3
of anti-viral resistance strains, the efficacy of treatment is greatly reduced. Hence other
aspects of the HCMV replication machinery need to be studied in greater detail as a
basis of identifying a broader range of potential anti-viral targets in the future.
Table 2 Sequenced cytomegalovirus (CMV) genomes
Common Name Strain Accession size (kbp) Reference
Human CMV AD169 X17403 229 Chee et al. (1990)
Towne AY315197 231 Dunn et al. (2003)
Merlin AY446894 236 Dolan et al. (2004)
Toledo AC146905 227 Murphy et al. (2003)
Chimpanzee CMV - AF480884 241 Davison et al. (2003)
Rhesus CMV 68-1 AY186194 221 Hansen et al. (2003)
Mouse CMV Smith U68299 230 Rawlinson et al. (1996)
Rat CMV Maastricht AF232689 230 Vink et al. (2000)
1.2 HCMV Epidemiology
Human cytomegalovirus (HCMV) is a ubiquitous pathogen that is acquired early in life
in most populations and generally increases in seroprevalence with age. However, the
patterns of acquisition vary greatly based on geographic and socioeconomic
backgrounds such that in developing nations, acquisition of HCMV is nearly universal
in early childhood (Kubo, 1991; Miles, 2007; Prabhakar, 1992). HCMV infection is
endemic in the human population (Table 3) and is not influenced by seasonal variations
(Malm, 2007; Pass, 2005). The seroprevalence of HCMV infection within the adult
population is very common ranging from 60% in developed countries, to 100% in
developing countries (Munro, 2005). In particular, countries in Africa (Adjei, 2006;
Pultoo, 2001), Asia (Kangro, 1994; Kothari, 2002; Liu, 1990; Shen, 1992; Urwijitaroon,
1993; Wong, 2000), and the Eurasian border (Hizel, 1999) tend to have a higher
incidence of HCMV infection with less significant differences between age groups than
the European countries (Alanen, 2005; Andersson-Ellström, 1995; de Ory, 2004;
Hecker, 2004; Kangro, 1994; Knowles, 2005; Natali, 1997), Australia (Seale, 2006),
New Zealand (Beresford, 1988), and the United States (Staras, 2007).

4
Table 3 Epidemiological seroprevalence of HCMV infection
Location Population Age Seropositivity Reference
Study (years) (%)
Europe
Germany Blood donor 18+ 46 Hecker et al. (2004)
Parma, Italy General 2-54 72 Natali et al. (1997)
Finland Women 16-45 56 Alanen et al. (2005)
Madrid, Spain Women 2-40 66 de Ory et al. (2004)
United Kingdom General 12-21 19 Kangro et al. (1993)
Ankara, Turkey Women 15-49 99 Hizel et al. (1999)
Ireland Women 15-46 30 Knowles et al. (2005)
Sweden Women 16 45 Andersson-Ellström et al. (1995)
Asia
Chengdu, China General 4-7 60 Liu et al. (1990)
Taipei, Taiwan General 4-12 58 Shen et al. (1992)
Delhi, India Blood donor 95 Kothari et al. (2002)
Thailand Blood donor 17-59 93 Urwijitaroon et al. (1993)
Hong Kong General 12-21 80 Kangro et al. (1993)
Nepal General 1+ 100 Kubo et al. (1991)
Singapore Pregnant women ≤30 77 Wong et al. (2000)
North America
USA General 6+ 59 Staras et al. (2006)
Cuernavaca, Mexico Women 13-44 92 Echániz-Avilés et al. (1993)
South America
Jamaica General 15-25 90+ Prabhakar et al. (1992)
Santiago, Chile General <30 60 Abarca et al. (1997)
Buenos Aires, Argentina Socioeconomic* <15 46 Damilano et al. (1992)
Africa
Ghana Blood donor 20-69 93 Adjei et al. (2006)
Gambia General 1 85 Miles et al. (2007)
Mauritius Blood donor 18-60 94 Pultoo et al. (2001)
Australia General 1-59 57 Seale et al. (2006)
New Zealand Blood donor 56-65 65 Beresford et al. (1988)
*Population study based on middle-socioeconomic classes
1.2.1 Transmission
HCMV infection is acquired through direct contact with body fluids from an infected
person and is present in saliva, urine, breast milk, blood products, semen, cervical and
vaginal secretions, as well as allograft tissues (Arvin, 2007). In a hospital environment,
allograft recipients are among the highest at risk to HCMV infection, with over 75% of

5
solid organ transplant patients contracting or reactivating latent CMV after
transplantation (Fishman, 1998). Transfusion associated HCMV infections were first
described 40 years ago (Kääriäinen, 1966) and is attributed to the virus’s ability to
establish latency in cells of the myeloid lineage (Cervia, 2007; Miller, 1991). In a child-
care environment, horizontal transmission of HCMV is common amongst young
children, parents and other care-givers from the community (Adler, 1988; Adler, 1989;
Bale, 1999; de Mello, 1996; Kiss, 2002; Pass, 1990; Pass, 1984) while in young adult
populations, sexual activity is the major mode of transmission for CMV (Handsfield,
1985). The presence of infectious virus is common in salivary glands and cervical
secretions (Britt, 1996; Shen, 1994). HCMV has also been cultured from semen despite
being urine, blood, and saliva negative for infectious virus (Biggar, 1983; Lang, 1975).
In addition, artificial insemination therapy has been suggested as another route of
transmission (Mansat, 1997; Prior, 1994), with HCMV detected in 0.02% to 34% of
donor semen depending on the population study (Mansat, 1997; McGowan, 1983; Shen,
1994a; Tjiam, 1987; Yang, 1995).
1.2.2 Congenital infection
HCMV is the leading cause of congenital viral infection, occurring in 0.15-3.0% of
newborns worldwide (Hassan, 2007; Malm, 2007). Infection can occur via viral
transmission through the placenta, during delivery via cervical secretions and blood or
from the mother via breast milk (Malm, 2007). The risk for viral transmission is higher
in primary infected mothers than in mothers with reactivated disease (Gaytant, 2002).
Primary CMV infections are reported in 1-4% of seronegative women during pregnancy
and the risk for viral transmission to the fetus is 30-40% (Stagno, 1986; Stagno, 1982)
while reactivation of a CMV infection during pregnancy is reported in 10-30% of
seropositive women but the risk of transmitting the virus is only 1-3% (Stagno, 1982).
Approximately 30% of those infected will develop symptomatic disease (Ahlfors, 1999),
although only 10-15% of children with congenital CMV are symptomatic during the
neonatal period (Malm, 2007). Hearing loss is the most common sequela of congenital
CMV infection, occurring in 10–15% of infected children (Dahle, 2000; Dollard, 2007;
Pass, 2005; Ross, 2006) while other neurological deficits such as mental retardation,
autism, learning disabilities, cerebral palsy, epilepsy, visual impairment, microcephaly,
encephalitis, seizures, and upper motor neuron disorders may also develop (Gandhi,

6
2004; Ross, 2005). The most severely affected infants have a mortality rate of about
30% with death occurring as a result of hepatic dysfunction, bleeding, disseminated
intravascular coagulation or secondary bacterial infections (Malm, 2007).
1.2.3 Immunocompromised infection and disease outcomes
HCMV infection often remains asymptomatic within an immunocompetent host,
whereas it may cause significant morbidity and mortality in immunocompromised
individuals such as transplant patients and HIV/AIDS patients (Arvin, 2007). Over 75%
of solid organ transplant patients are newly infected or reactivate latent CMV after
transplantation (Fishman, 1998), often within the first 3 months when
immunosuppression is most intense (Dummer, 1983). CMV disease manifests in the
vast majority of transplant recipients as a viral syndrome that includes fever, malaise,
muscle pain or headache (Steininger, 2007) and has been associated with diminished
graft survival and allograft rejection (Grattan, 1989; Pouteil-Noble, 1993; Ricart, 2005).
In HIV-infected patients, retinitis is the single most common manifestation of CMV
disease, accounting for 85% of all cases (Yust, 2004). CMV infection has also been
linked to the onset of diabetes mellitus in kidney transplant recipients (Hjelmesaeth,
2004) as well as accelerated cardiac allograft vasculopathy in heart transplant recipients
(Lemström, 1995).
1.2.4 Antiviral treatment
Currently, only five compounds have been licensed to treat established HCMV
infections: ganciclovir (GCV), its oral pro-drug, valganciclovir (vGCV), Foscarnet
(FOS), cidofovir (CDV) and fomivirsen (Mercorelli, 2007). GCV is a nucleoside analog
commonly used to treat CMV disease and in particular, CMV retinitis in the
immunocompromised (De Clercq, 2001). GCV requires intracellular phosphorylation,
firstly by the UL97 viral kinase and then by cellular kinases to create a triphosphate
active form that acts as a competitive inhibitor of deoxyguanosine triphosphate,
incorporating itself into the viral DNA via the DNA polymerase during elongation to
disrupt DNA synthesis (Abdel-Haq, 2006). Valganciclovir is an inactive pro-drug form
of GCV that is rapidly converted to active GCV in the plasma (Abdel-Haq, 2006) and
has a ten-fold higher bioavailability than GCV (Cocohoba, 2002; Pescovitz, 2000),

7
which is normally administered intravenously. Hematologic toxicities such as
neutropenia, anemia, and thrombocytopenia are among the most frequently reported
adverse side effects (Abdel-Haq, 2006; Mercorelli, 2007) while nausea, diarrhea,
vomiting, fever, and abdominal pain have also been reported during GCV and
valganciclovir treatment (Cocohoba, 2002). CDV is an acyclic nucleoside phosphonate
with a mechanism of action similar to that of other nucleoside analogs, but unlike GCV,
is not dependent on activation by a viral encoded enzyme (Mercorelli, 2007). Instead,
host kinases convert CDV to the active diphosphoryl form, and CDV disphosphate then
acts as a competitive inhibitor of the viral DNA polymerase, causing premature chain
termination in viral DNA synthesis (Biron, 2006). Although CDV is approved for use to
treat CMV retinitis in AIDS patients, severe nephrotoxicity is a major limitation of its
use (Biron, 2006; Ho, 2000; Mercorelli, 2007) and thus remains a second-line therapy
option (Biron, 2006). In addition, it has low oral bioavailability and is only available as
an intravenous treatment (Mercorelli, 2007). Neutropenia is another toxicity associated
with CDV, and CDV was shown to be both carcinogenic and teratogenic in preclinical
toxicological studies (Biron, 2006). FOS is a pyrophosphate analogue that inhibits the
viral DNA polymerase by binding to the pyrophosphate binding site and blocking
cleavage of pyrophosphate from the terminal nucleoside triphosphate added to the
growing DNA chain (Biron, 2006; Wagstaff, 1994). FOS is also considered a second-
line therapy due to its nephrotoxic potential (Biron, 2006; Mercorelli, 2007), but is the
preferred drug for patients who are failing GCV therapy due to viral resistance, or those
who cannot be treated with GCV due to dose-limiting neutropenia or leucopenia
(Razonable, 2004). Fomivirsen is a 21-nucleotide anti-sense RNA (5’-GCG TTT GCT
CTT CTT CTT GCG-3‘) (De Clercq, 2001), specifically targeted against the major
immediate-early (MIE) transactivator gene of CMV (Geary, 2002). As it is administered
only as an intraocular injection, it is an end-organ treatment that does not affect
systemic CMV (Arvin, 2007). The most commonly reported adverse effect associated
with fomivirsen is ocular inflammation, which occurs in 25% patients (Group., 2002),
with other less common side-effects including abnormal vision, cataracts, bleeding in
and around the eye, reduced color vision, eye pain, retinal detachment, stomach pain,
low blood count, weakness, dehydration, cough, flu-like symptoms, and chest pain
(Group., 2002).

8
In addition to these licensed drugs, several anti-CMV drugs are also currently in clinical
development. Maribavir (MBV), (1-(β-L-ribofuranosyl)-2-isopropylamino-5, 6-
dichlorobenzimidazole), is a riboside analog with a novel mechanism of action that is
being developed to inhibit viral DNA synthesis in CMV and EBV (Biron, 2002; Zacny,
1999). In CMV-infected cells, Maribavir targets the HCMV protein kinase gene (UL97)
that has been shown to play a role in viral nucleocapsid egress from the nucleus, thereby
reducing the yield of infectious CMV (Evers, 2004; Krosky, 2003a). Its mechanism of
action is not fully understood although it has been hypothesized that inhibition of viral
DNA synthesis is a consequence of preventing the phosphorylation of the DNA
polymerase accessory protein (pUL44)by the protein kinase (Krosky, 2003; Marschall,
2003). In addition, laboratory-generated resistant mutations have been mapped to the
UL27 gene, a protein of unknown function (Chou, 2004; Komazin, 2003). Maribavir
pre-clinically shows advantages over existing anti-CMV drugs in its in vitro potency,
bioavailability, safety profile in acute, chronic and genetic toxicology testing, and the
lack of cross-resistance with GCV, FOS, and CDV resistant strains (Biron, 2006; Drew,
2006).
BAY 38-4766 represents a novel class of non-nucleoside antiviral agents that are highly
selective inhibitors of CMV in vitro (Reefschlaeger, 2001). The compound
demonstrates antiviral activity similar to that of GCV (Weber, 2001) and showed a
favorable safety profile in healthy male volunteers at single oral doses up to 2000 mg
(Biron, 2006). The mechanism of action is largely unknown although resistance
mutations have been mapped to the UL89 and UL56 genes (Krosky, 1998;
Reefschlaeger, 2001), which encode the two terminase subunit proteins involved in
DNA cleavage and packaging (Bogner, 2001). It is also active against strains that are
resistant to GCV (McSharry, 2001) and does not have cross-resistance with other
current antiviral drugs (Reefschlaeger, 2001).
1.2.5 Antiviral resistance
Resistance to antiviral treatment mainly occurs under selective pressure from lengthy
periods of antiviral treatment, although other risk factors including host immune
competence and levels of ongoing viral replication are also involved (Chou, 2001). In
addition, currently approved drugs for treatment share a similar mechanism of action by

9
targeting the DNA polymerase, hence increasing the risk of mutations that confer
resistance to multiple anti-viral drugs. Antiviral resistant CMV strains may emerge
between 6 weeks to 2 months of treatment depending on the patient group (Boivin, 2001;
Eckle, 2002; Limaye, 2000; Springer, 2005) although low levels of resistant genotypes
are present in patients even before antiviral treatment commences (Emery, 2000).
Antiviral resistance can be determined by genotypic or phenotypic assays. Phenotypic
assays measure the concentration of an antiviral agent necessary to inhibit viral
replication while genotypic assays, screen for key resistance mutations in the UL97 and
UL54 genes (Springer, 2005). These two gene products encode for the CMV protein
kinase and DNA polymerase respectively. Point mutations or deletions of portions of
UL97 can lead to GCV resistance (Baldanti, 1995; Baldanti, 1998; Chou, 1995; Chou,
1997; Chou, 2000a; Chou, 2005; Chou, 2002; Faizi-Khan, 1998; Hanson, 1995; Hantz,
2005; Ijichi, 2002; Lurain, 1994; Marfori, 2007; Sullivan, 1992; Wolf, 1995) (Table 4),
while UL54 mutations, depending on their locations, can confer resistance to one or
more anti-viral drugs (Chou, 2003; Chou, 1997; Chou, 1998; Chou, 2007a; Cihlar, 1998;
Cihlar, 1998a; Ducancelle, 2006; Gilbert, 2005a; Marfori, 2007; Mousavi-Jazi, 2001;
Mousavi-Jazi, 2003; Scott, 2007; Springer, 2005; Weinberg, 2003) (Table 5).
UL97 GCV resistance mutations are clustered at codons 460, 520, and 590-607 (Chou,
1999), but the degree of resistance varies depending on the mutation or combination of
mutations present. For example, the most commonly encountered mutations in UL97,
A594V, L595S, and M460V, confer a 5-10 fold increase resistance to GCV while
C592G and A594T confer only a 2-3 fold increase resistance to GCV (Chou, 1999;
Chou, 2005). Maribavir resistance mutations have also been mapped to the UL97 gene
(Biron, 2002; Chou, 2007), although the affected codons are clustered upstream of the
known GCV resistance mutations and share no cross-resistance with GCV (Chou, 2007).
In addition to GCV, UL54 mutations are known to confer varying degrees of resistance
to FOS and/or CDV as well. Among the most frequent mutations associated with
resistances to these drugs are V715M, V781I, and L802M, which confer resistance to
FOS (Cihlar, 1998), F412C, L501I, and P522S, which confer resistance to GCV and
CDV (Chou, 1997; Cihlar, 1998), and A809V, which confer resistance to GCV and
FOS (Chou, 1998). In addition, an increasing number of GCV and FOS cross-resistant
strains have been observed in several laboratory and clinical isolates (Biron, 2006;
Mercorelli, 2007) while there have also been cases of mutations that confer resistance to

10
all three anti-CMV drugs (Chou, 2000; Scott, 2007). The issue of cross-resistance
arising from the above mentioned factors poses a need to develop drugs with a novel
mechanism of action to use with patients not responding to current treatment or in
conjunction with existing anti-viral drugs to reduce the incidence of resistance.

11
Table 4 Mutations of UL97 and UL27 antiviral resistance
Gene Mutation Domain Resistance* Reference
UL97 V353A --- MBV Biron 2002, Chou 2007
L397R --- MBV Biron 2002
T409M --- MBV Biron 2002, Chou 2007
H411 --- MBV Chou 2008
M460V VI GCV Chou 1995
M460I VI GCV Lurain 1994
H520Q --- GCV Hanson 1995
590-93 deletion IX GCV Sullivan 1992
A591V IX GCV Chou 2002
591-607 deletion IX GCV Chou 2002
C592G IX GCV Chou 2005
A594V IX GCV Chou 1995
A594P IX GCV Ijichi 2002
A594T IX GCV Chou 2002
L595S IX GCV Chou 1995
L595F IX GCV Wolf 1995
L595W IX GCV Chou 2002
L595 deletion IX GCV Baldanti 1995
595-603 deletion IX GCV Chou 2000a
E596G --- GCV Chou 2002
K599T --- GCV Faizi-Khan 1998
601 deletion --- GCV Hantz 2005
601-03 deletion --- GCV Marfori 2007
C603W/R/S --- GCV Chou 1997
C607Y --- GCV Baldanti 1998
C607F --- GCV Chou 2002
UL27 L355P --- MBV Komazin 2003
R233S --- MBV Chou 2004
W362R --- MBV Chou 2004
A406V/415Stop --- MBV Chou 2004
*GCV = Gancyclovir, MBV = Maribavir

12
Table 5 Mutations of UL54 associated with antiviral resistance
Gene Mutation Domain Resistance* Reference
UL54 D301N EXOI GCV Chou 2003
N408D IV GCV, CDV Cihlar 1998
N408K IV GCV, CDV Scott 2007
N410K IV GCV, CDV Chou 2003
F412C/V IV GCV, CDV Chou 1997, Cihlar 1998
D413E or A IV GCV, CDV Chou 2003
D413A IV GCV, CDV Marfori 2007
T419M IV FOS Mousavi 2003
N495K δ-C FOS Ducancelle 2006
L501I δ-C GCV, CDV Cihlar 1998
T503I δ-C GCV, CDV Chou 2003
K513N or E δ-C GCV, CDV Cihlar 1998a
L516R δ-C GCV, CDV Chou 2003
P522S δ-C GCV, CDV Cihlar 1998
L545S δ-C GCV, CDV Cihlar 1998
Q578H δ-C FOS Mousavi 2003
D588N or E δ-C FOS Mousavi 2001, Cihlar 1998
A692S --- FOS Chou 2003
T700A II FOS Cihlar 1998
V715M II FOS Cihlar 1998
E756K --- GCV, CDV, FOS Mousavi 2001, Chou 2003
E756D or Q --- FOS Chou 2003, Weinberg 2003
L773V VI FOS Mousavi 2003
V781I VI GCV, FOS Cihlar 1998, Mousavi 2001
V787L VI FOS Weinberg 2003
L802M III FOS Cihlar 1998, Chou 1997
K805Q III CDV Cihlar 1998
A809V III GCV, FOS Chou 1998
V812L III GCV, CDV, FOS Chou 1997, Cihlar 1998a
T813S III GCV, FOS Chou 2007a
T821I III GCV, CDV, FOS Cihlar 1998
P829S III FOS Gilbert 2005a
A834P III GCV, CDV, FOS Scott 2007
T838A III FOS Springer 2005
G841A III GCV, FOS Chou 2007a
981-82 deletion V GCV, CDV, FOS Chou 2000
A987G V GCV, CDV Cihlar 1998
*GCV = Ganciclovir, CDV = Cidofovir, FOS = Foscarnet

13
1.3 HCMV genome structure and organization
Human cytomegalovirus has the largest genome of all the herpesviruses with a linear
double stranded DNA genome of 230-240kbp. It is also the only known betaherpesvirus
to have a class E genome structure that consists of a long unique (UL) and short unique
(US) sequence capable of inverting to give four sequence isomers (Gibson, 1999). Each
unique region is flanked by inverted repeat sequences (TRL and IRL, TRS and IRS), such
that the overall structure of the HCMV genome is TRL-UL-IRL-IRS-US-TRS (Mocarski,
2001) (Figure 1). Large scale sequencing analysis has revealed the existence of seven
conserved sequence blocks present in all herpesviruses, but arranged in different orders
(Chee, 1990). These conserved regions were found to have a functional role in DNA
replication, DNA repair, nucleotide metabolism, and virion structure (Mocarski, 2001).
Figure 1 Arrangement of the HCMV genome
1.3.1 Open reading frame organization
Initial analysis of the fully sequenced AD169 laboratory strain predicted the existence
of 208 open reading frames (ORFs) (Chee, 1990), including 33 ORFs that have
substantial sequence similarity to HSV-1, varicella zoster virus (VZV), and Epstein-
Barr virus (EBV) (Chee, 1990), as well as 41 ORFs that are dispensable for replication
(Spaete, 1987). Several studies have proposed their models for a consensus genome
(Chee, 1990; Davison, 2003; Murphy, 2003), while various interpretations to the coding
potential of AD169 (Chee, 1990) has led to several revisions of its genomic content.
Sequence comparisons with the closely related chimpanzee cytomegalovirus (CCMV)
discounted 51 previously identified ORFs, re-interpreted 24 ORFs, as well as predicting
the existence of 10 novel genes, including five in AD169 (UL15A, UL21A, UL128,
UL131A, and US34A) (Davison, 2003). Another study, using the Bio-Dictionary-based
Gene Finder (BDGF) analysis tool, found that 37 ORFs from the original AD169
sequence can be discarded as non-polypeptide encoding as well as the addition of up to

14
12 ORFs that had not been previously identified (Murphy, 2003). In addition, sequence
differences have been pointed out in ORF UL102 (Smith, 1995a), US28 (Neote, 1993),
and UL15 (Davison, 2003). Certain stocks of AD169 were also found to contain an
additional 929 base pairs that were not in the original sequence of AD169 and which
resulted in alterations to the UL42 and UL43 genes (Dargan, 1997; Mocarski, 1997) as
well as a reinterpretation of the UL41 gene (Dargan, 1997). Several ORFs have also
been modified to account for splicing events, including UL111A, UL118, UL119, UL22,
US3 (Rawlinson, 1993), and UL33 (Davis-Poynter, 1997). Furthermore, various regions
of the genome are known to be hypervariable among clinical isolates (Prichard, 2001).
These include: three major genotype variations in UL11 (Hitomi, 1997), UL144 (Lurain,
1999), four major genotype variations in US9, US28 (Rasmussen, 2003), UL4 (Bar,
2001), UL73 (Dal Monte, 2004; Pignatelli, 2001), UL55 (Chou, 1991; Meyer-König,
1998), and five major genotype variations in UL37 (Hayajneh, 2001), as well as several
associated with UL146 (Hassan-Walker, 2004). Nevertheless, laboratory strain AD169
is still widely regarded as the consensus strain for interpretation of coding.
1.4 CMV growth cycle
HCMV generally replicates slowly in cell culture, taking days for cytopathic effects to
appear (Britt, 1996). However, in vivo, CMV has been observed to have a doubling time
of approximately 1-2 days, depending on the methods used (Emery, 1999). The HCMV
growth cycle consists of cell attachment and viral entry, transport to the nucleus,
transcription and translation of viral proteins, DNA replication, capsid assembly, and
maturation and egress from the cell (Figure 2). The CMV replication cycle has not been
completely elucidated, although it shares numerous homologies to HSV-1 replication
(Boehmer, 2003).

15
Figure 2 CMV growth cycle
1. Cell attachment and viral entry; 2. Transport to the nucleus; 3. Transcription/translation of viral
proteins; 4. DNA replication; 5. Capsid assembly; and 6. Maturation and egress
1.4.1 Cell attachment and viral entry
HCMV cell attachment and penetration of the host cell occurs by fusion with the plasma
membrane (Compton, 2004) and is mainly mediated by envelope glycoproteins (Arvin,
2007; Mach, 2005). Entry of HCMV involves the interaction of these envelope
glycoproteins with a number of distinct receptors, initiated by the use of the cell surface
proteoglycan heparan sulfate by glycoprotein B (gB) (Compton, 1993). HCMV gB
(UL55) and the gH:gL (UL75:UL115) complex have been the most extensively studied
(Baldwin, 2000; Feire, 2004; Lopper, 2004; Navarro, 1993) while recent evidence
suggests core envelope glycoproteins gM (UL100) and gN (UL73) may also be
important for viral entry (Mach, 2005). HCMV and other herpesviruses exploit normal
cytoplasmic transport systems of the host (Dohner, 2005; Ogawa-Goto, 2003) to control
nucleocapsid transit through the cytoplasm. It relies on microtubules to gain access to
the nucleus and nuclear pores where un-coating is completed and the viral genome is
released into the nucleoplasm (Dohner, 2005). The importance of essential protein-

16
protein interactions in receptor recognition and signaling for viral attachment and entry
underscores the potential for developing drugs which target other aspects of the CMV
replication cycle.
1.4.2 HCMV replication genes
HCMV replication is associated with eleven loci that have been identified as necessary
for transient complementation of oriLyt-dependent DNA replication (Pari, 1993). Six of
these were found to encode homologs of herpes simplex virus (HSV-1) DNA replication
proteins (Anders, 1996; Pari, 1993), including: a two-subunit DNA polymerase (UL54
and UL44), a single-stranded DNA binding protein (UL57), a primase (UL70), a
helicase (UL105), and a primase-associated factor (UL102). In addition to these six core
replication genes, five other loci supplemented in regulatory and transactivation roles.
The UL36-38 locus is involved in the up-regulation of viral transcription, inhibition of
apoptosis, and growth in human cells (Colberg-Poley, 1996; Goldmacher, 1999; Pari,
1993a; Smith, 1995; Terhune, 2007), while the four spliced variants of UL112-113
cooperate with each other to relocate UL44 and possibly other core replication proteins
to the pre-replication foci (Iskenderian, 1996; Park, 2006). The TRS1/IRS1 locus is
involved in the disruption of host cell antiviral response pathways, the production of
viable virus, as well as having a regulatory function for the other immediate early genes
(Adamo, 2004; Blankenship, 2002; Child, 2004; Hakki, 2005; Romanowski, 1997). The
major immediate-early region (IE1/IE2) plays a role in regulating the cell cycle,
metabolism and apoptosis (Chiou, 2006; Lukac, 1999; Yu, 2002; Zhu, 1995). Although
not considered to be part of the core set of replication machinery proteins, UL84 has
been shown to have an essential role in oriLyt-dependent DNA replication (Sarisky,
1996; Xu, 2004). It is also involved in promoting the formation of replication
compartments (Sarisky, 1996; Xu, 2002) as well as interacting with IE2 to regulate
transactivation activity in the UL112-113 locus (Gebert, 1997; Spector, 1994).
1.4.2.1 DNA polymerase (UL54)
HCMV encodes a two-subunit DNA polymerase which comprises a catalytic subunit,
UL54 (Heilbronn, 1987) and an accessory protein, UL44, which acts as a processivity
factor (Ertl, 1992). Initial studies revealed that UL54 possesses DNA-dependent DNA

17
polymerase activity (Mar, 1981) as well as 3’-5’ exonuclease activity (Nishiyama, 1983)
and was highly homologous to the UL30 protein of HSV-1 (Gottlieb, 1990; Hernandez,
1990). UL54 shares a series of conserved regions and structural domains with the α-
family of DNA polymerases. These conserved regions are located within amino acid
residues 379–1100 and include conserved domains I–VII, which are involved in the
DNA polymerisation reaction (Hwang, 1992; Wong, 1988), the δ-region C (Zhang,
1991) and three motifs designated Exo I–III which are located within conserved
domains IV and δ-region C (Bernad, 1989). Mutations conferring resistance to GCV,
CDV, and FOS map to several of these conserved motifs (Lurain, 1992; Sullivan, 1993)
while mutations induced in these regions leading to a loss of function (Ye, 1993)
demonstrate the importance of these conserved domains. In particular, domains I, II and
III have been shown to directly participate in binding to deoxynucleoside triphosphates,
in chelating Mg2+
ions and in interacting with primer and template (Ye, 1993) while it
has been suggested that domains III, V, and δ -region C can also form part of the dNTP-
binding site (Ye, 1993). The region involved with interacting with the UL44 subunit has
been mapped to C terminal residues 1221-1242 of UL54 and appears to be dependent on
hydrophobic interactions (Loregian, 2003).
1.4.2.2 DNA polymerase accessory protein (UL44)
Initial studies on the HCMV DNA polymerase (UL54) and its accessory subunit (UL44)
were based on the HSV-1 equivalent, UL30 and UL42. UL54 is a functional homolog of
the HSV-1 UL30 DNA polymerase, both sharing polymerase activity (Mar, 1981) as
well as 3’-5’ exonuclease activity (Nishiyama, 1983) that are dependent on salt
concentration (Cihlar, 1997; Weiland, 1994). UL44 is analogous to the HSV-1
processivity factor UL42 (Gottlieb, 1990), functioning to bind double-stranded DNA
and stimulating DNA synthesis by the main catalytic subunit UL54 (Appleton, 2004;
Ertl, 1992; Loregian, 2004a; Weiland, 1994). The observation that both UL54 and UL44
are required for DNA synthesis and that disruption to either subunit severely inhibits
viral DNA replication (Digard, 1993; Pari, 1993a; Ripalti, 1995) provides a strong basis
to study the UL54/UL44 interaction. The UL44 binding site on UL54 had previously
been elucidated to the C-terminal 22 amino-acids of UL54 with two cysteine residues
suggested to play a key role in binding or stabilizing the UL54/UL44 interaction
(Loregian, 2004; Loregian, 2004a; Loregian, 2003). Alanine mutants of the C-terminal

18
region in UL54 further identified Leu1227 and Phe1231 as key residues for subunit
interaction (Loregian, 2004). Although the HSV-1 homolog interaction between
UL30/UL42 had been mapped to an analogous region earlier (Monahan, 1993; Zuccola,
2000), neither UL30 nor UL54 shared any sequence similarity with each other
(Loregian, 2005).
Analysis of residues 1-290 from the UL44 crystal structure revealed an overall fold that
was similar to other processivity factors including UL42 of HSV-1 and the proliferating
cell nuclear antigen (PCNA) of eukaryotic DNA polymerases, despite sharing no
obvious sequence similarity (Appleton, 2004). Crystallization of the N-terminal two-
thirds of UL44 revealed a C-clamp-shaped head-to-head homodimer (Appleton, 2004)
in contrast to UL42, which carries out its functions as a monomer (Gottlieb, 1990;
Randell, 2004; Zuccola, 2000). Analysis of the dimer binding region revealed specific
residue interactions involving six main-chain-to-main-chain hydrogen bonds and
extensive packaging of hydrophobic side chains at the interface (Appleton, 2004). At
the binding interface, F121 of each monomer is buried against a hydrophobic loop
composed of P85, L86 and L87 of the other monomer. L86 and L87 also pack against
M123 and L93 of the opposite monomer (Appleton, 2004). Furthermore, L86 and L87
are conserved as hydrophobic residues in UL44 homologs of other β-herpesvirus
(Appleton, 2004), suggesting an importance in specific binding mechanisms. Alanine
substitutions of the two lysine residues in the hydrophobic loop disrupted DNA binding
by up to 100 fold (Appleton, 2004; Loregian, 2005).
In addition, UL44 shares a ‘connector loop’ structure similar to UL42 which was shown
to connect two topologically similar domains of UL42 (Zuccola, 2000) as well as
playing a crucial role in interacting with UL30 (Bridges, 2001). Indeed, recent studies
have shown that the connector loop in UL44 has functional similarities to its homolog
in HSV-1 (Appleton, 2004; Loregian, 2004). Constructed alanine mutants of the
connector loop region showed that substitutions that affected residues 133-136 severely
reduced physical and functional interaction with UL54 while a substitution at residue
I135 disrupted UL54/UL44 binding and long-chain DNA synthesis completely
(Appleton, 2004; Loregian, 2004). Similar studies done with the UL42 connector loop
identified the glutamine residue at position 171 as crucial for subunit interaction
(Bridges, 2001). However, subtle differences between the two systems have also been

19
described, suggesting that despite the similarities in function and analogous positioning,
the mechanism of binding is probably different. Firstly, the residues important for
binding the two HCMV subunits are hydrophobic while their counterparts in the HSV-1
interaction are hydrophilic (Bridges, 2001; Loregian, 2004; Zuccola, 2000). This is
further supported by findings that the accessory subunit of HSV-1 cannot stimulate the
catalytic subunit of HCMV, and vice-versa (Loregian, 2005). Secondly, despite the
substitution of residue Q171 in UL42, weak binding could still be detected in maltose
binding protein (MBP)-pulldown assays (Bridges, 2001), whereas substitution of the
I135 residue in UL44 abolished binding completely (Loregian, 2004). The observation
that even a single substitution can disrupt key interactions to such an extent has
provided some insight into possible anti-viral treatments such as the use of small
inhibitory molecules (Loregian, 2004; Pilger, 2004).
Although the crucial functions of UL44 had been mapped to the N-terminal two-thirds
of the gene, the region beyond residue 290 has been less well defined functionally.
Earlier studies had determined that the C-terminus of the gene was dispensable for
UL44 activity (Loregian, 2004a; Weiland, 1994), despite the presence of salient
glycine-rich strings (Ertl, 1992). However, more recent studies suggest the C-terminus
to be involved in nuclear transport with the key nuclear localization site (NLS) located
at residues 425-431 of the C-terminus (Alvisi, 2005). This is consistent with similar
studies on nuclear localization sites flanked by protein kinase phosphorylation sites of
other homologous herpesviruses (Loh, 1999; Takeda, 2000). Apart from its function as
the polymerase processivity factor, UL44 has also been associated with mediating cell
adhesion via a putative integrin binding RGD motif that has been studied in murine
CMV (Loh, 2000).
1.4.2.3 Single-stranded DNA binding protein (UL57)
HCMV UL57 encodes an early single-stranded DNA-binding protein that is
homologous to the HSV-1 major DNA-binding protein, ICP8 (Anders, 1996; Pari,
1993). ICP8 has been studied more extensively than UL57 and as such, much of our
understanding of UL57 is based on observations derived from studies on ICP8. Studies
in the HSV-1 major DNA-binding protein ICP8 have been linked to multiple functions
within the HSV replication cycle. Its ability to modulate activity of the HSV-1 DNA

20
polymerase (Hernandez, 1990), affect genome recombination and processing (Bortner,
1993; Dutch, 1993; Nimonkar, 2003), regulate late viral gene expression (Chen, 1996;
Gao, 1989; Gao, 1991), organize DNA replication enzymes into nuclear replication
compartments (Bush, 1991; de Bruyn Kops, 1988) as well as interactions with the
helicase-primase complex (Boehmer, 1998; Boehmer, 1993; Crute, 1991; Falkenberg,
1998; Hamatake, 1997; He, 2001; Lee, 1997; Makhov, 1996) provides an insight into
potential functional parallels that may be found with its HCMV counterpart. However, it
has already been established that like ICP8, UL57 serves as an essential component for
transient complementation in HCMV oriLyt-mediated DNA replication (Pari, 1993;
Sarisky, 1996). Physical aspects of the ICP8 protein have also been characterized to a
certain extent. Cooperative DNA binding has been mapped to the C-terminus of ICP8
(Dudas, 1998; Mapelli, 2000) with the possible involvement of two cysteine residues at
245 and 455 playing a role in modulation (Dudas, 1998). The DNA binding region has
been harder to elucidate with studies mapping regions which range from residues 564-
1160 (Gao, 1989), 300-849 (Wang, 1990), and 368-902 (White, 1999) as potential
active sites. Several other functional regions have been mapped including: a nuclear
localization signal at the C-terminal 28 amino acid residues (Gao, 1992), an intranuclear
localization site located in the region between residues 1080-1135 (Taylor, 2003), and a
putative zinc binding motif between residues 499-512 (Gao, 1988; Gupte, 1991).
UL57 is located about 1kb downstream of oriLyt, raising the possibility that UL57
promoter elements contribute to oriLyt function (Kiehl, 2003). Indeed, mapping the 5’
ends of the UL57 transcripts elucidated a 42 base-pair sequence spanning the oriLyt-
proximal start site for UL57 that was found to play a role for both UL57 transcription
and oriLyt function, albeit not an essential one for the latter (Kiehl, 2003). Furthermore,
the 1kb non-coding region between oriLyt and UL57 consists of numerous potential
transcriptional control signals, including transcription factor binding sites and
polyadenylation signals, all of which have been shown to participate in replicator
functions in other herpesvirus systems (Nguyen-Huynh, 1998; Schepers, 1993).
1.4.2.4 Primase-helicase complex (UL70, UL102, and UL105)
Despite several studies detailing the functionality of the helicase-primase complex, most
of them were done in the context of the herpes simplex virus model (Barnard, 1997;

21
Biswas, 1999; Dracheva, 1995; Graves-Woodward, 1996; Graves-Woodward, 1997;
Klinedinst, 1994). Apart from the initial characterization of the HCMV helicase-primase
proteins (Pari, 1993; Smith, 1995a; Smith, 1996), there have been relatively few
publications on the specific interactions of these proteins. Although studies have shown
that the UL70, UL102, and UL105 proteins interact with each other to form a
heterotrimeric complex (McMahon, 2002), the specific regions that bind the complex
together have not been studied. Based on sequence and positional similarities with the
herpes simplex virus counterparts, the helicase protein (UL105) is predicted to be
involved in unwinding the DNA helix, while the HCMV primase protein (UL70) is
predicted to be involved in initiating DNA synthesis. However, the primase-helicase
associated protein (UL102) is not specifically involved in the activity of either, yet it
interacts with both UL70 and UL105 to enhance their activities. By elucidating the
binding relationship of these three proteins, we hope to understand more about the role
of UL102.
The HCMV primase ORF (UL70) encodes a 947 amino acid protein that shares 27%
homology with the HSV-1 primase, UL52 (Chee, 1990). Similar to the HSV-1 primase,
pUL70 contains several conserved regions including a putative DXD motif that is
associated with primase catalysis (Dracheva, 1995; Klinedinst, 1994). Furthermore,
substitutions of the aspartate residues in the motif have been shown to abolish primase
but not helicase or ATPase activity in vitro (Dracheva, 1995; Klinedinst, 1994). The
HSV-1 UL52 protein contains a putative zinc-finger motif at its C terminus that is
highly conserved among herpesviruses as well as other prokaryotic, and eukaryotic
primases (Ilyina, 1992; Mendelman, 1994). Although the role of the zinc finger is not
well defined, alanine substitutions of the third and fourth conserved cysteines resulted in
the loss of primase, helicase, ATPase, and DNA-binding activities of the HSV helicase-
primase complex (Biswas, 1999). UL70 is predicted to engage in primer synthesis
during DNA replication on the basis of its relative homology to the HSV-1 primase,
which has been shown to produce short oligoribonucleotide primers up to 10-13
nucleotides long on ssDNA (Crute, 1991; Ramirez-Aguilar, 1995), allowing the DNA
polymerase to begin replicating DNA via dNTP polymerization (Gottlieb, 1994;
Nimonkar, 2003). In addition, the HSV-1 primase was found to initiate primer synthesis
with a purine, at the second nucleotide of a 3'-deoxyguanylate-pyrimidine-pyrimidine

22
(3'-G-pyr-pyr) template sequence (Ramirez-Aguilar, 1995), which may be similar in the
HCMV primase.
HCMV also encodes a helicase-primase associated protein (UL102) that shares little
sequence homology to its HSV counterpart, UL8 (Chee, 1990), but instead is
recognized as a positional homolog (McGeoch, 1988; Pari, 1993). Initial identification
of the UL102 ORF revealed a 798 amino acid protein that was essential for origin-
dependent DNA replication (Chee, 1990; Pari, 1993). In addition, the UL101 ORF
located upstream of UL102 was also shown to be an essential component in replication
(Pari, 1993). However, comparison against cDNA and subsequent re-sequencing of
UL102 in Towne and AD169 (Smith, 1995a) revealed that the genomic stop codon for
UL101 was an error in the original published sequence (Chee, 1990). As a result,
UL102 has been redefined as a 2.7kb un-spliced transcript encoding an 873 amino acid
protein that has the capacity to encode several smaller proteins, all within frame of the
UL102 stop codon (Smith, 1995a). Although further elucidations of the biochemical
activities of UL102 have yet to be published, its positional and functional homology to
HSV UL8 may provide an insight into potential functional parallels derived from
studies on UL8. Firstly, studies looking at primase activity have demonstrated that UL8
may serve a key role in stimulating the synthesis of RNA primers (Tenney, 1994) as
well as increasing the efficiency of primer utilization by DNA polymerase (Sherman,
1992). A central segment of UL8 has been attributed to interact with both the HSV
helicase (UL5) and primase (UL52) subunits (Barnard, 1997). However, in the absence
of UL8, the HSV primase and helicase protein form a sub-complex that is enzymatically
indistinguishable from a complex formed by all three proteins (Calder, 1992; Crute,
1991), suggesting that UL8 may play a role in promoting nuclear localization of the
complex (Calder, 1992). In addition, UL8 also interacts directly with the origin binding
protein (UL9) and at least functionally with the single-stranded DNA binding protein
(ICP8) (Hamatake, 1997; McLean, 1994). UL8 has also been hypothesized to help
direct the polymerase to the initiation complex, where it coordinates polymerase and
helicase-primase activities (Marsden, 1997).
The HCMV helicase ORF (UL105) encodes a 982 amino acid protein that shares 34%
sequence homology with the HSV-1 helicase, UL5 (Chee, 1990), and is present in
infected cells as early as 24 hrs post-infection (Smith, 1996). Like the HSV-1 helicase,

23
the HCMV helicase protein (pUL105) contains six conserved helicase motifs common
to superfamily-1 DNA helicases (Gorbalenya, 1989; Zhu, 1992). Motif I consists of a
GxxGxGKT/S Walker A motif associated with binding the di- or triphosphate moiety of
the nucleotide cofactor (Walker, 1982) while Motif II consists of a Walker B motif
containing a group of hydrophobic residues terminated by an Asp and a Glu residue and
is associated with the stabilization of the coordinated Mg2+
ion (Walker, 1982). In HSV,
motif I has been shown to be directly involved in ATP binding and/or hydrolysis while
motif II appears to be required for coupling of DNA binding to ATP hydrolysis
(Graves-Woodward, 1997). The functional significance of motifs III, IV, V, and VI are
yet to be fully elucidated but the strong conservation of these six motifs suggests that
they may be important for helicase activity. However, mutations generated in motifs III,
IV, V, and VI did not eliminate ATP hydrolysis nor affect DNA binding and therefore is
postulated to be involved in the coupling of these two activities to the process of DNA
unwinding (Graves-Woodward, 1997). Other studies on the HSV helicase have shown
that single amino acid substitutions in the most conserved residues of these motifs
abolish the ability of the HSV helicase to support DNA replication in vivo suggesting
that these conserved residues are essential to DNA replication (Zhu, 1992).
1.4.2.5 Putative Initiator Protein (UL84)
HCMV UL84 encodes a 586 amino acid protein that is detected in infected cells as early
as 2.5 hrs post-infection (He, 1992). Currently, unlike the other replication genes, UL84
shares no functional or positional homology to any HSV replication gene. Analysis of
the UL84 amino acid sequence has revealed the presence of two potential leucine zipper
domain in the N-terminal half of the protein, at amino acids 114–135 and at 325-373
(He, 1992). The leucine zipper domain between residues 114-135 has been implicated in
the interaction of UL84 with the IE2 protein (Colletti, 2004), an immediate-early protein,
that has been identified as one of the auxiliary components required for origin-
dependent DNA replication in human fibroblasts (Pari, 1993). Substitutions of the
leucine residues in this domain rendered UL84 incapable of complementing oriLyt-
dependent DNA replication (Colletti, 2004) while further studies on the functional role
of the UL84–IE2 interaction revealed that an over-expression of UL84 interfered with
the IE2-mediated transactivation of the UL112/113 promoter, decreasing IE2-mediated
transient transactivation (Gebert, 1997). UL84 has also been shown to undergo

24
oligomerization with key interaction domain mapped to residues 151-201 (Colletti,
2004). Initial studies identified UL84 as utilizing a nuclear localization signal similar to
that of the simian virus 40 large T antigen and IE2 nuclear localization signals (Xu,
2002), although subsequent studies report that UL84 interacts with at least four
members of the α importin protein family in vitro and in vivo to utilize a classic
importin-mediated pathway for nuclear import (Lischka, 2003). The region of UL84
responsible for the interaction with α importin has been identified as a complex domain
spanning residues 226-508 and containing a cluster of basic amino acids similar to that
of the classical nuclear localization signals (NLS), albeit lacking classical NLS activity
(Lischka, 2003). This domain also contains two motifs homologous to leucine-rich
nuclear export signals (NES) which have been shown to enable UL84 to shuttle between
the nucleus and the cytoplasm (Lischka, 2006). There are also evidence to suggest that
UL84 may have enzymatic properties that is similar to that of a helicase (Colletti, 2005).
Studies have demonstrated UTPase activity in UL84 that has been suggested to be part
of an energy-generating system for helicase activity (Colletti, 2005; Davison, 2005). In
addition, the amino acid sequence of UL84 displays some structural homology to the
DExD/H box family of RNA helicases (Colletti, 2005). However, most importantly are
recent implications that UL84 provides the role as an initiator of DNA replication, with
several studies demonstrating that UL84 interacts with DNA replication genes UL57
and UL44 (Gao, 2007).
1.4.3 DNA replication
Herpesvirus DNA replication is thought to begin with genome circularization and theta
form replication which then proceeds to a rolling circle form which has been well
documented in HSV (Boehmer, 2003). Initiation of CMV DNA synthesis has yet to be
elucidated. However, in general, initiation of DNA synthesis involves targeted
unwinding of particular DNA sequences to enable assembly of a replisome. It has been
proposed that DNA synthesis in herpesvirus initially proceeds by a theta type
mechanism in which DNA replication proceeds bi-directionally from the lytic origin of
replication, although this has yet to be seen in vitro (Boehmer, 2003). HCMV has a
highly complex oriLyt region that spans over a kilo-base of DNA upstream of the UL57
ORF (Kiehl, 2003). However, unlike HSV, it is the only lytic-phase replicator identified
in HCMV (Anders, 1992). Herpesviruses encode a virion-associated transcript that

25
associates with oriLyt to form a three-stranded structure whose precise role in DNA
synthesis is yet to be elucidated (Prichard, 1998). In general, binding of the Origin
Binding Protein (OBP) with specific sites in OriLyt is followed by an interaction with
single stranded DNA binding proteins which lead to localized unwinding and access of
replication fork proteins. Here, the replication fork machinery consisting of the DNA
polymerase catalytic and accessory subunits, and the hetero-trimeric helicase-primase
complex direct continuous, leading strand viral DNA replication in a rolling circle
mechanism (Boehmer, 2003).
1.4.4 Capsid assembly, maturation and egress
Rolling circle replication generates long head-to-tail concatemers consisting of multiple
unit-length genomes that accumulate in the nuclei. These concatemers are cleaved at
regular intervals and packaged into individual, pre-formed capsids by a mechanism that
is conserved throughout the entire herpesviridae family (Homa, 1997). Structural studies
of the HSV capsid have indicated that structural and functional protein homologues of
HSV capsid proteins are present in the capsid of HCMV (Gibson, 1996). In addition,
cryo-electron microscopic analysis of the capsids has revealed that HCMV and HSV
have near identical structures (Butcher, 1998; Chen, 1999) suggesting that the assembly
of the HCMV capsid follows a very similar assembly pathway as that of HSV
(Grunewald, 2003). The HCMV capsid is composed of 162 capsomeres consisting of
150 hexons and 12 pentons (Butcher, 1998; Chen, 1999) of which the most abundant
protein components consist of the major capsid protein (MCP, UL86) and the smallest
capsid protein (SCP, UL48–49). Two copies of the minor capsid protein (MnCP, UL85)
combined with a single copy of the minor capsid binding protein (MnCP-BP, UL46)
form the triplexes that are located between adjacent pentons and hexons (Butcher, 1998;
Chen, 1999). The process of capsid assembly involves UL86 and UL48-49 interacting
in the cytoplasm in the presence of assembly protein precursor gene UL80a before
translocating to the nucleus. UL85 and UL46 are also translocated to the nucleus, albeit
separately from the UL86/UL48-49 complex. Once in the nucleus, self-interaction
domains in the products of the UL80a lead to formation of pentons and hexons and the
generation of the capsid scaffold. The UL86/UL48-49 complex then interacts with
UL85 and UL46 to form the immature shell of the capsid (Gibson, 1996). In HCMV,
the packaging of viral DNA involves a portal protein, proposed to be encoded by

26
UL104 (Komazin, 2004), and is mediated through virus encoded protein recognition of
two conserved sequence motifs, the pac-1 and pac-2 sequences, that are located at each
end of the viral genome (Mocarski, 2001). In HSV, this process is also coupled with
capsid maturation (Heymann, 2003), which involves the proteolytic cleavage of the
carboxyl terminal of the MCP binding domain (Gibson, 1996). The process of HCMV
egress is not well known, but is thought to involve an initial envelope obtained at the
inner nuclear membrane that is lost by a fusion event at the outer nuclear membrane,
releasing free nucleocapsids into the cytoplasm, where egress continues through a
second and final envelopment step as the virion leaves the host cell (Mettenleiter, 2004).
1.4.5 Latency and reactivation
A unique biological property common to all herpesviruses, including HCMV, is the
ability of the virus to establish lifelong persistence within the host following initial
infection. In general, HCMV reactivation from latency is observed in 13% of healthy
adults (Ling, 2003), and though episodes of sporadic reactivation may occur, they are
generally well-controlled by cell-mediated immuno-surveillance. However, as with
primary infection, reactivation within an immunocompromised individual can lead to
high levels of morbidity and mortality. Recent studies have implicated endothelial cells
and specific cell types of the myeloid lineage as sites of HCMV persistence and latency
(Jarvis, 2002). The factors leading to reactivation from latency are not completely
understood, although studies have shown that myeloid differentiation can stimulate the
production of viral gene expression and in some cases, the production of infectious virus
(Reeves, 2005; Soderberg-Naucler, 2001; Taylor-Wiedeman, 1994). In addition, the
production of the stress hormones cortisol, adrenocorticotrophic hormone, epinephrine
and norepinephrine has been implicated in the reactivation and shedding of CMV in
urine (Mehta, 2000).
1.5 Aims and Objectives
There is a fundamental need for understanding the role of other HCMV genes essential
for DNA replication as a foundation for developing new antiviral treatments. In this
study we looked at six HCMV replication genes encoding for the DNA polymerase
accessory protein (UL44), single stranded DNA binding protein (UL57), primase

27
(UL70), helicase (UL105), primase-helicase associated protein (UL102), and the
putative initiator protein (UL84) in order to increase our understanding of their role in
DNA replication (Figure 3). In particular, UL70 is an essential component of the
primase-helicase complex and elucidation of its functional domains is essential for
understanding its role in DNA replication and subsequently, ways to inhibit its activity
as a potential drug target. Hence, the aims of this project are to:
1. Identify strain variation within the UL44, UL57, UL70, UL102, UL105,
and UL84 genes.
2. Predict putative functional domains and motifs for these genes using the
ScanProsite software, and
3. Express and study the UL70 (Primase) gene through the use of
recombinant mutants.
Figure 3 Relative positions of genes investigated in this study

28
2 Reagents and equipment
All reagents and equipment used in the study are detailed in this section. Individual
experimental methods are outlined in their corresponding chapters. In general, most
chemicals used were molecular biology grade and solutions sterilized by autoclaving or
filtration. Processing of biological specimens was carried out in Class II cabinets using
sterile techniques. In addition all purchased reagents were tested sterile and pyrogen-
free by the manufacturers. Biological waste was sterilized by autoclaving prior to
disposal.
2.1 Viral stocks
HCMV low-
passage strains
Clinical isolates 44A, 70A, 77A, 90A, 91A, 16B, 21B, 80B, 4E,
and 30E were obtained from the Prince of Wales Hospital.
2.2 DNA extraction
2.2.1 Buffers
T10E50
buffer
T10E50 buffer consisted of 10mM Tris-HCl (pH 8.0), 50mM EDTA in
MilliQ water and autoclaved before use.
TE buffer TE buffer consisted of 10mM Tris-HCl (pH 8.0), 1mM EDTA in MilliQ
water and autoclaved before use.
2.2.2 Chenicals
Chloroform Chloroform was obtained from Sigma-Aldrich (USA).
Ethanol 99.5% ethanol was obtained from Sigma-Aldrich (USA).
NaCl >98% (titration) sodium chloride for molecular biology was obtained
from Sigma-Aldrich (USA).
Phenol Phenol was obtained from Sigma-Aldrich (USA).

29
2.2.3 Kits
QIAamp DNA
Mini Kit
QIAamp DNA mini kits (Qiagen, USA) were used to extract DNA
from frozen cell-associated viral stock using a QIAamp spin-column
procedure.
2.3 Polymerase chain reaction (PCR)
2.3.1 Buffers
ExpandLong
template reaction
buffer 1
10X Expand Long Template Buffer 1 was obtained from Roche
(Germany) and is supplemented with 17.5mM MgCl2.
GoTaq green reaction
buffer
5X GoTaq green reaction buffer was obtained from
Promega (USA) and is supplemented with 7.5mM MgCl2.
Platinum Taq High
Fidelity PCR buffer
10X Platinum Taq high fidelity PCR buffer was obtained from
Invitrogen (USA) and consisted of 600mM Tris-SO4 buffer (pH
8.9), 180mM ammonium sulfate.
Red Hot reaction
buffer
10X Red Hot reaction buffer was obtained from AB gene (UK)
and consisted of 750mM Tris-HCl (pH 8.8), 200mM
(NH4)2SO4, 0.1% (v/v) Tween 20.
2.3.2 Enzymes
ExpandLong template
enzyme
Expand Long Template enzyme mix was obtained from
Roche (Germany) and stored at -20°C before use.
GoTaq Flexi DNA
polymerase
GoTaq Flexi DNA polymerase was obtained from
Promega (USA) and stored at -20°C before use.
Platinum Taq High Fidelity
DNA Polymerase
Platinum Taq DNA polymerase was obtained from
Invitrogen (USA) and stored at -20°C before use.
Red Hot DNA polymerase Red Hot DNA polymerase was obtained from ABgene
(UK) and stored at -20°C before use.

30
2.3.3 Equipment
Thermal
cycler
PTC 200 Peltier Thermal cyclers (GMI, USA) were used for
amplification of DNA fragments.
2.3.4 Reagents
dNTP Deoxynucleoside triphosphates, dATP, dCTP, dGTP and dTTP, each at stock
concentrations of 100mM (Promega, USA) were diluted 1:10 and 1:100 with
sterile water (Baxter, Australia) to a final concentration of 10mM and 1mM
respectively. Diluted dNTP stocks were stored at -20°C.
MgCl2 25mM magnesium chloride was obtained from AB gene (UK) or Promega
(USA) and stored at -20°C before use.
MgSO4 50mM magnesium sulfate was obtained from Invitrogen (USA) and stored at -
20°C before use.
Primers Primers were synthesized by Sigma Genosys (Australia) as 400µM stock
solutions. These were further diluted to 10µM with sterile water (Baxter,
Australia) and stored at -20°C before use.
2.4 Electrophoresis
2.4.1 Buffers
TBE
buffer
10X TBE consisted of Trizma base (Sigma-Aldrich, USA), boric acid (Sigma-
Aldrich, USA) and EDTA (Sigma-Aldrich, USA) in MilliQ water and
autoclaved. 0.5X working stock made by diluting 1:20 in MilliQ water.
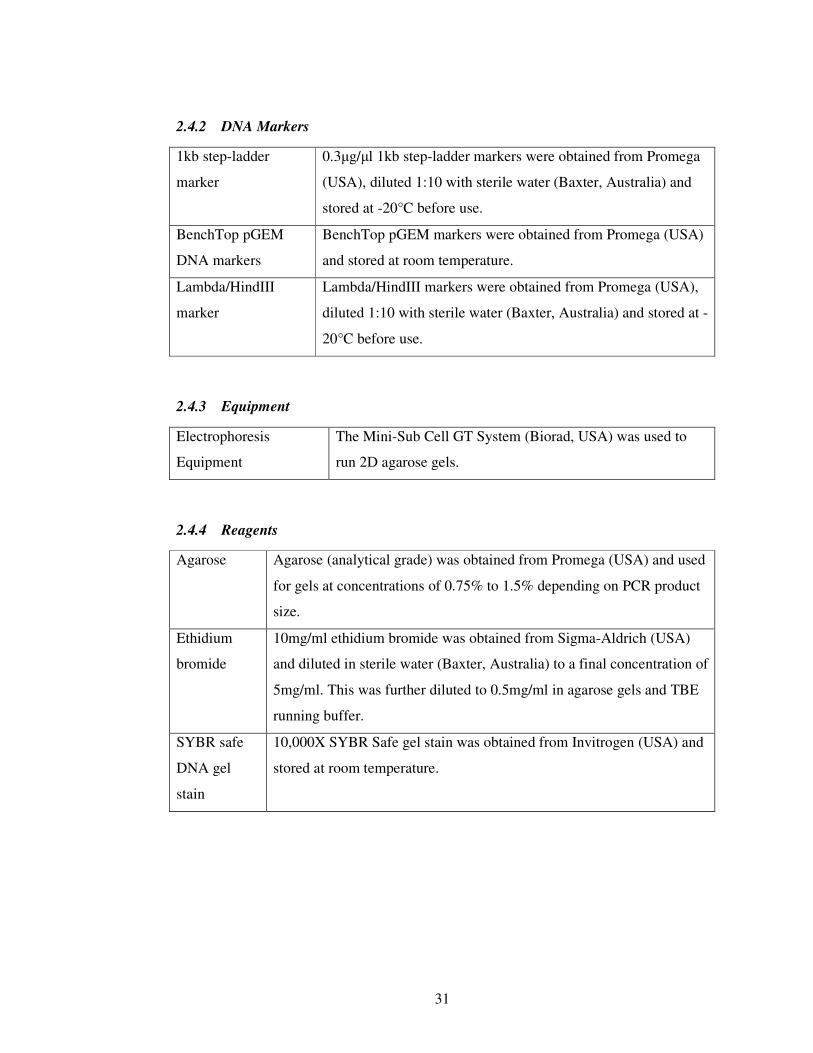
31
2.4.2 DNA Markers
1kb step-ladder
marker
0.3µg/µl 1kb step-ladder markers were obtained from Promega
(USA), diluted 1:10 with sterile water (Baxter, Australia) and
stored at -20°C before use.
BenchTop pGEM
DNA markers
BenchTop pGEM markers were obtained from Promega (USA)
and stored at room temperature.
Lambda/HindIII
marker
Lambda/HindIII markers were obtained from Promega (USA),
diluted 1:10 with sterile water (Baxter, Australia) and stored at -
20°C before use.
2.4.3 Equipment
Electrophoresis
Equipment
The Mini-Sub Cell GT System (Biorad, USA) was used to
run 2D agarose gels.
2.4.4 Reagents
Agarose Agarose (analytical grade) was obtained from Promega (USA) and used
for gels at concentrations of 0.75% to 1.5% depending on PCR product
size.
Ethidium
bromide
10mg/ml ethidium bromide was obtained from Sigma-Aldrich (USA)
and diluted in sterile water (Baxter, Australia) to a final concentration of
5mg/ml. This was further diluted to 0.5mg/ml in agarose gels and TBE
running buffer.
SYBR safe
DNA gel
stain
10,000X SYBR Safe gel stain was obtained from Invitrogen (USA) and
stored at room temperature.

32
2.5 DNA sequencing
2.5.1 Equipment
DNA
sequencer
A 3730 DNA Analyzer (Applied Biosystems, USA) was used for
analysis of sequencing products at the Ramaciotti Centre, University of
New South Wales.
2.5.2 Reagents
PEG PCR
mix
Polyethylene glycol PCR mix consisted of 26.7% (w/v) polyethylene
glycol (PEG) 8000 (Sigma-Aldrich, USA), 0.6M NaOAc (pH 5.2),
6.5mM MgCl2.
Primers Primers were synthesized by Sigma Genosys (Australia) as 400µM
stock solutions. These were further diluted to 10µM with sterile water
(Baxter, Australia) and stored at -20°C before use.
Sequencing
mix
BigDye Terminator v3.1 was obtained from Applied Biosystems (USA)
and stored at -20°C before use.
Sequencing
buffer
5X sequencing buffer was obtained from Applied Biosystems (USA)
and stored at -20°C before use.
2.6 Restriction enzymes
2.6.1 Buffers
CIAP stop
buffer
CIAP stop buffer consisted of 10mM Tris-HCl (pH 7.5), 1mM EDTA
(pH 7.5), 200mM NaCl, and 0.5% (w/v) SDS.
Multi-core
buffer
10X multi-core buffer was obtained from Promega (USA) and consisted
of 10mM Tris-HCl (pH 7.5), 100mM NaCl, 50% glycerol.

33
2.6.2 Enzymes
BamHI BamHI was obtained from New England BioLabs (USA) with NEBuffer 3
and stored at -20°C before use.
HindIII HindIII was obtained from Promega (USA) with buffer B and stored at -20°C
before use.
SalI SalI was obtained from New England BioLabs (USA) with NEBuffer 3 and
stored at -20°C before use.
SpeI SpeI was obtained from Promega (USA) with buffer B and stored at -20°C
before use.
2.6.3 Reagents
BSA 1µg/µl Bovine serum albumin (BSA) was obtained from Promega (USA) and
stored at -20°C before use.
CIAP Calf intestinal alkaline phosphatase (CIAP) was obtained from Promega (USA)
and stored at -20°C before use.
TSAP Thermosensitive alkaline phosphatase (TSAP) was obtained from Promega
(USA) and stored at -20°C before use.
2.7 Cloning reagents
2.7.1 Antibiotics
Ampicillin 91.0-100.5% Ampicillin was obtained from Sigma-Aldrich (USA),
diluted with sterile water (Baxter, USA) to a working concentration of
10mg/ml, filtered and stored at -20°C before use.
Tetracycline 100mg/ml Tetracycline was obtained from Invitrogen (USA) and stored
at -20°C before use.
Zeocin 100mg/ml Zeocin was obtained from Invitrogen (USA) and stored at -
20°C before use.

34
2.7.2 Cells
Library efficiency DH5α
competent cells
DH5α competent cells were obtained from Invitrogen
(USA) and stored at -80°C before use.
XL10 Gold Ultracompetent
cells
XL10 Gold ultracompetent cells were obtained from
Stratagene (USA) and stored at -80°C before use.
2.7.3 Equipment
Low-salt LB
zeocin plates
Low salt LB consisted of 1% (w/v) tryptone (Oxoid, Australia), 0.5%
NaCl (Sigma-Aldrich, USA), 0.5% yeast (Oxoid, Australia), 1.5%
(w/v) agar (Promega, USA), adjusted to pH 7.5 and autoclaved.
Allowed to cool to 55°C and zeocin (Invitrogen, USA) added to final
concentration of 50µg/ml before pouring.
LB
Ampicillin
plates
2% (w/v) agar (Promega, USA) added to LB broth and autoclaved.
Allowed to cool to 55°C and Ampicillin (Sigma-Aldrich, USA) added
to final concentration of 100µg/ml before pouring.
LB
tetracycline
plates
2% (w/v) agar (Promega, USA) added to LB broth and autoclaved.
Allowed to cool to 55°C and tetracycline (Sigma-Aldrich, USA) added
to final concentration of 125µg/ml before pouring.

35
2.7.4 Kits
DNA ligation kit DNA ligation kit was obtained from Stratagene (USA) and stored at
-20°C before use. Includes pUC18 (BamH I digested) and cI857
wild-type lambda (Hind III digested) control DNA, 10mM rATP
(pH 7.5), T4 DNA ligase, and 10X ligase buffer consisting of
500mM Tris-HCl (pH 7.5), 70mM MgCl2, 10mM dithiothreitol
(DTT).
Gateway BP
Clonase II
enzyme mix
BP Clonase II kit was obtained from Invitrogen (USA) and stored
at -20°C before use.
Wizard Plus
Midipreps DNA
purification
system
Wizard Plus midiprep kits (Promega, USA) were used to extract
plasmid DNA using a column and vacuum procedure. Reagents
were stored at room temperature.
Wizard Plus
Minipreps DNA
purification
system
Wizard Plus miniprep kits (Promega, USA) were used to extract
plasmid DNA using a column and vacuum procedure. Reagents
were stored at room temperature.
Wizard SV
Gel/PCR Clean-
up System
Wizard PCR clean-up kits (Promega, USA) were used to purify
restriction digested DNA. Reagents were stored at room
temperature.

36
2.7.5 Media
Luria
broth
Luria broth (LB) consisted of 1% (w/v) NaCl (Sigma-Aldrich, USA), 1%
(w/v) tryptone (Oxoid, Australia), 0.5% (w/v) yeast (Oxoid, Australia),
adjusted to pH 7.0, and autoclaved prior to use.
NZY
broth
NZY broth consisted of 1% (w/v) casein hydrolysate (Merck, USA), 0.5%
(w/v) yeast (Oxoid, Australia), and 0.5% (w/v) NaCl (Sigma-Aldrich, USA),
adjusted to pH 7.5 and autoclaved. Add 12.5ml of 1M MgCl2 (Sigma, USA),
12.5ml of 1M MgSO4 (BDH, UK), and 10ml of 2M glucose (Gibco BRL,
USA) per liter of broth prior to use.
S.O.C
media
S.O.C media consisted of 2% (w/v) tryptone (Oxoid, Australia), 0.5% (w/v)
yeast (Oxoid, Australia), 10mM NaCl (Sigma-Aldrich, USA), 2.5mM KCl
(BDH, UK), 10mM MgCl2 (Sigma, USA), 10mM MgSO4 (BDH, UK), 20mM
glucose (Gibco BRL, USA), and autoclaved before use.
2.7.6 Reagents
IPTG 100mM Isopropyl B-D-Thiogalactopyranoside was obtained from
Promega (USA).
Proteinase
K
2µg/µl Proteinase K was obtained from Invitrogen (USA) and stored at -
20°C before use.
X-gal 50mg/ml 5-bromo-4-chloro-3-indolyl-B-D-galactosidase was obtained
from Promega (USA).
2.7.7 Vectors
pBluescript II
SK(+)
pBluescript II SK(+) vectors were obtained from Stratagene (USA)
and stored at -20°C before use.
pCITE-4A(+) pCITE-4A(+) vectors were obtained from Novagen (Germany) and
stored at -20°C before use.
pDONR/Zeo pDONR/Zeo vectors were obtained from Invitrogen (USA) and
stored at -20°C before use.

37
2.8 Protein expression and detection
2.8.1 Antibodies
Anti-His G Antibody Anti-His G antibodies (Invitrogen, USA) were used for
protein detection and stored at -20°C before use.
S-Protein alkaline
phosphatase conjugate
S-Protein alkaline phosphatase conjugate (S-Tag) was
obtained from Novagen (Germany) and stored at -20°C
before use.
2.8.2 Buffers
Loading
buffer
4X loading buffer consisted of 0.25M Tris-Cl (Sigma-Aldrich, USA), 40%
(w/v) glycerol, 8% (w/v) SDS, 0.4% (w/v) Bromophenol Blue (BDH, UK),
8% (v/v) β-mercaptoethanol (Sigma-Aldrich, USA)
PBS 10X phosphate buffered solution (PBS) were obtained from Gibco (USA).
Running
buffer
1X running buffer consisted of 25mM Trizma base (Sigma-Aldrich, USA),
250mM (pH 8.3) glycine (Sigma-Aldrich, USA), and 0.1% (w/v) SDS in
MilliQ water.
Transfer
buffer
1X transfer buffer consisted of 25mM Trisma base (Sigma-Aldrich, USA),
250mM (pH 8.3) glycine (Sigma-Aldrich, USA), and 20% methanol
(Mallinckrodt Chemical, UK) in MilliQ water.

38
2.8.3 Equipment
2-D gel
electrophoresis
equipment
The Mini-PROTEAN 3 Cell electrophoresis system (Biorad,
USA) was used to run 2D poly-acrylamide gels.
Nitrocellulose Hybond-ECL nitrocellulose was obtained from Amersham
Biosciences.
Resolving gel 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis
(SDS-PAGE) resolving gels consisted of 34% (v/v) 30%
Bis/Acrylamide, 0.39M Tris (pH 8.8), 0.1% SDS, 0.1% APS, and
0.04% TEMED.
Stacking gel 5% sodium dodecyl sulfate polyacrylamide gel electrophoresis
(SDS-PAGE) stacking gels consisted of 16.5% (v/v) 30%
Bis/Acrylamide, 0.125M Tris (pH 6.8), 0.1% SDS, 0.1% APS,
and 0.1% TEMED.
2.8.4 Kits
TNT Quick Coupled
Transcription/Translation
System
TNT Quick Coupled Transcription/Translation
System (Promega, USA) was used to express protein
utilizing the pCITE expression vector.
2.8.5 Protein Markers
Kaleidoscope marker Precision Plus Protein Kaleidoscope Standards were obtained
from Biorad (USA) and stored at -20°C before use.
Perfect Protein
Western markers
Perfect protein western markers were obtained from Novagen
(Germany) and stored at -20°C before use.

39
2.8.6 Reagents
APS >98% ammonium persulfate (APS) was obtained from
Sigma-Aldrich (USA) and stored at room temperature.
Bis/Acrylamide 30% Acrylamide/Bis Solution (37.5:1) was obtained from
Biorad (USA) and stored at 4°C before use.
Gelcode blue stain Gelcode blue stain (Pierce, USA) was used to stain
PAGE gels for total protein.
Glycerol >99% glycerol was obtained from Sigma-Aldrich (USA)
Ponceau S 0.1% (w/v) Ponceau S in 5% acetic acid (Sigma-Aldrich,
USA) was used to stain nitrocellulose membranes for
total protein.
SDS >98.5% Sodium dodecyl sulfate (for molecular
biology, .98.5%) was obtained from Sigma-Adrich
(USA).
SuperSignal West Pico
chemiluminescent substrate
SuperSignal West Pico Chemiluminescent Substrate kit
was obtained from Pierce (USA) and stored at 4°C before
use.
TEMED >99% N, N, N’, N’-Tetramethylethylenediamine was
obtained from Sigma-Aldrich (USA) and stored at room
temperature.
2.8.7 Solutions
Blocking
solution
Blocking solution consisted of 5% skim milk powder (Diploma,
Australia) in PBST.
PBST
solution
Phosphate buffered solution with Tween (PBST) consisted of 1X PBS,
and 0.1% Tween (Sigma-Aldrich, USA)

40
3 Identification of putative functional motifs in viral proteins
essential for human cytomegalovirus DNA replication
3.1 Introduction
HCMV DNA replication is associated with eleven loci necessary for transient
complementation of oriLyt-dependent DNA replication (Pari, 1993). These include six
core genes homologous to six of the seven essential replication genes in Herpes Simplex
Virus (HSV) (Anders, 1996) (Table 6). In HCMV, they consist of a two-subunit DNA
polymerase (UL54 and UL44), single-stranded DNA binding protein (UL57), primase
(UL70), helicase (UL105), and the primase-helicase associated factor (UL102). In
addition, five other loci (TRS1/IRS1, IE1/IE2, UL112-113, UL84, and UL36-38) have
supporting regulatory and trans-activation roles in HCMV replication (Iskenderian,
1996; Pari, 1993). DNA replication in HCMV is considered to be analogous to that in
HSV, with the function of several HCMV proteins being predicted on the basis of
sequence and functional homology with their HSV counterparts (Anders, 1996; Chee,
1990).
Table 6 Essential DNA replication proteins in HCMV and HSV-1
Function HCMV HSV1
DNA Polymerase UL54 UL30
DNA Polymerase accessory protein UL44 UL42
Single-Stranded DNA Binding Protein UL57 UL29
Primase UL70 UL52
Helicase UL105 UL5
Primase-Helicase Associated Protein UL102 UL8
Initiator Protein UL84 UL9
The role of the HCMV DNA polymerase accessory protein (UL44) has been well
documented over the years with the elucidation of key functional motifs and residues
(Appleton, 2004; Loregian, 2004) as well as a partial crystal structure of the functional
N terminal half of the protein (Appleton, 2006; Appleton, 2004). In contrast, the HCMV
single stranded DNA binding protein (UL57) has been less well elucidated although
early studies established biochemical properties that were comparable to that of the

41
HSV major DNA-binding protein, ICP8 (Anders, 1987). The HCMV primase (UL70),
helicase (UL105) and primase-helicase associated factor (UL102) form a trimeric
complex (McMahon, 2002) that shares biologically important motifs with the HSV
primase-helicase complex (Dracheva, 1995; Klinedinst, 1994; Zhu, 1992). The UL84
ORF encodes the only non-core replication protein that is required for origin-dependent
DNA replication in human fibroblasts (Sarisky, 1996). Although initially classed as a
regulatory protein for HCMV replication (Pari, 1993), recent functional studies (Colletti,
2004; Xu, 2004) have suggested that the UL84 ORF encodes the initiator protein for
HCMV DNA synthesis (Xu, 2004a).
The residues comprising a protein are not all equally important. Some serve as
indispensable mediators of protein function while others can be readily replaced without
impacting on functionality. As the sequences of biologically significant residues are
evolutionarily conserved, conservation analysis has become one of the most widely used
techniques for predicting protein function. In this study we report our findings on the
variation of the HCMV replication proteins amongst isolate strains.
3.2 Methods
3.2.1 Viral stocks and DNA extraction
Viral DNA from 10 low-passage isolates (44A, 70A, 77A, 90A, 91A, 16B, 21B, 80B,
4E, and 30E) were extracted from frozen cell-associated viral stock using the QIAamp
DNA Mini Kit (Qiagen, USA) as per manufacturer’s instructions, with DNA eluted in
50µl of buffer AE after 5 min incubation instead of the recommended 200µl.
3.2.2 Polymerase chain reaction (PCR) amplification
Full-length ORFs encoding the accessory protein (UL44), helicase-primase complex,
(UL70, UL102, and UL105), single-stranded DNA-binding protein (UL57), and
putative initiator protein (UL84) were amplified in overlapping segments from viral
DNA by single rounds of PCR. PCR reaction mixes consisted of 1x buffer, 2mM MgCl2,
0.25mM deoxynucleoside triphosphates (dNTP), 0.2µM forward and reverse primers
(Table 7), 1 U/µl Red Hot DNA polymerase (AB Gene), and 5µl of extracted DNA
template (approximately 100ng). Single round cycling conditions were 94°C for 5 min

42
followed by 30 cycles of 94°C for 1 min, 55°C for 30 sec, and 72°C for 2 min. PCR
products were visualized following electrophoresis on 1.5% agarose gels.

43
Table 7 Primers used for PCR amplification of HCMV DNA fragments
Gene Primer** Region Amplified* Sequence (5' to 3')
UL44 UL44.1 [F] 172843 - 173547 ATGGATCGCAAGACGCGCCT
UL44.2 [R] GACGGCGCAATTGAGCAGCG
UL44.3 [F] 173438 - 174199 GCGAGCTGGAATTCACGGCC
UL44.4 [R] TTTCTCCATCACGGGACCGCG
UL57 UL57.1 [F] 138815 - 140062 CATCACGCTATTTTCGCGGGC
UL57.2 [R] CGGTGATCGGTTGCGTTGGTC
UL57.3 [F] 139960 - 141135 TGGCCTTCAGTTTTGCCTCGG
UL57.4 [R] AATAGGCGTAGGCTAGCAGCG
UL57.5 [F] 141020 - 142123 TTCGGAGGTGCGCCTCAAGAG
UL57.6 [R] CCGTTGCCCGAAAAGTAGCCG
UL57.7 [F] 141978 - 142993 ACTGTGGCGCGTGAATCGTTG
UL57.8 [R] TTCGGGGCTGTTGCTGCTGTT
UL70 UL70.1 [F] 125951 - 126606 GCTGGGCCAGCTGCATCGTGCCGGCGCGACG
UL70.2 [R] GCGAGCCCAGTAGCAGACGCGCGAA
UL70.3 [F] 126497 - 128309 CATCTCCACCGTGGAGGAGTACGTG
UL70.4 [R] GGGGCAGATTTTGTCGCGTACGCTG
UL70.5 [F] 128221 - 128853 CCCAACTGGATCTGCGTAACCTGCT
UL70.6 [R] GGCGGGTCACCGGCGCCGTGGAAAGTGAGGC
UL102 UL102.1 [F] 147750 - 148466 GCTCACGACGCGGTTTGAGCA
UL102.2 [R] ATACTTGTGAGTGCCGACGCG
UL102.3 [F] 148341 - 148966 CGTGTGGTCACGCATGCCGCGGAAC
UL102.4 [R] GCCGTGGCATTGAGACGCACGGCGC
UL102.5 [F] 148896 - 149960 GTGCCCGAGGATGAGTGGCAGGTCT
UL102.6 [R] GAGGATCTCGCGATACACGGCTTCC
UL102.7 [F] 149870 - 150480 CGCGGCCGGTGGCGACTGGCTCTCG
UL102.8 [R] TACCCACGTAGTTCCCCTACGTGACTCG
UL105 UL105.1 [F] 153111 - 153909 AGAGCAGCTGCTTGCGCAGCA
UL105.2 [R] CATTTGTCCACGATGTCGGCG
UL105.3 [F] 153748 - 155593 CTTCGTCAGCAAGCACGTGCCGCTG
UL105.4 [R] CTGCGAGCTGATCTGACAGTTGGCC
UL105.5 [F] 155478 - 156130 TCAAGCGCTACCAGCTCATGCAGCG
UL105.6 [R] GTCACGTTTTCCTTACACGGTGTTGTG
UL84 UL84.1 [F] 108076 - 109175 AGAAGGGCGACGCGCTATGCG
UL84.2 [R] TCTTCTTGCGACGTCGCGGGG
UL84.3 [F] 109101 - 110188 TCTGCTCTCTACGCCGCTGCA
UL84.4 [R] ACGTGGCGCCATTCTCGTCGC
UL84.5 [F] 110115 - 111151 GCCCGTTACGGTCTGGCTACC
UL84.6 [R] CCGACGGGTAGTGGTGCACGT
*Given in relation to AD169 for UL44, UL57, and UL70, Merlin for UL102 and UL105,
and Towne for UL84.
**[F] denotes forward primer and [R] denotes reverse primer

44
3.2.3 DNA sequencing and analysis
All PCR products were precipitated from solution with polyethylene glycol as
previously described (Craxton, 1991). Sequencing was carried out using a reaction mix
consisting of Big Dye Terminator (Applied Biosystems, USA), premix buffer, 0.25µM
forward or reverse primer (Table 7), and approximately 50ng/300bp of DNA. Cycling
conditions consisted of 25 cycles of 96°C for 10 sec, 50°C for 5 sec, and 60°C for 4 min.
Sequencing products were purified and analyzed at the University of New South Wales
DNA analysis facility. Forward and reverse sequences were compared for sequencing
accuracy using GAP (GCG) (Accelrys).
3.2.4 Phylogenetic analysis
Full length nucleotide sequences for UL44, UL70, UL102, UL105, UL57, and UL84
were compiled for each individual strain, translated to protein using Translate (GCG)
(Accelrys) and aligned with three laboratory strains (AD169, Towne, and Merlin) using
ClustalW (Thompson, 1994). Genetic distances between pairs of nucleotide and amino
acid sequences were calculated with DNAdist and Protdist programs (Felsenstein, 1989)
respectively. Homologous proteins from other virus or cellular systems were also
identified using BLASTP (Altschul, 1997) and the results of the searches aligned to
their respective CMV counterpart using ClustalW (fast) (Thompson, 1994). All of these
programs were accessed through the Australian National Genomic Information Service
(ANGIS). The PROSITE database of biologically meaningful motif descriptors derived
from multiple alignments and ProRule (Hulo, 2006; Sigrist, 2005) were used to scan the
sequences of each HCMV replication protein. Potential motifs were identified using the
ScanProsite program (de Castro, 2006) from the Expert Protein Analysis System
(ExPASy) proteomics server of the Institute of Bioinformatics.
3.2.5 Genbank Accession Numbers
Sequences derived from each of the HCMV clinical isolates were submitted to Genbank
and have the accession numbers: UL44 (EU294431 - EU294440), UL57 (EU294441 -
EU294450), UL70 (EU294451 - EU294460), UL84 (EU294461 - EU294469), UL102
(EU294470 - EU294479), and UL105 (EU294480 - EU294489).

45
3.3 Results
3.3.1 Strain variation amongst HCMV isolates
The DNA polymerase accessory protein (UL44), single-stranded DNA binding protein
(UL57), primase (UL70), helicase (UL105), primase-helicase associated factor (UL102),
and putative initiator protein (UL84) are highly conserved among the ten HCMV
clinical isolates and three reference strains (AD169, Towne, and Merlin). All six genes
studied were found to be highly conserved with nucleotide (nt) identity ranging from
97.9% to 100% and amino acid (aa) identity ranging from 97.46% to 100% (Table 8).
Major alterations of the coding sequence, such as frame-shifts or premature stop codons,
were not observed. Amino acid polymorphisms in UL44, UL57, UL70, UL102, UL105,
and UL84 are summarized in Table 8. Polymorphisms existed outside of known
functional domains with the exceptions of a K430R substitution in the nuclear
localization signal (NLS) of UL44 as well as various polymorphisms in the NLS of
UL84. Results of phylogenetic analysis showed no preferential clustering of isolates
although certain amino acid changes were linked in UL84 and UL44. In particular, a
S296G substitution was consistently associated with an I319L substitution in UL44
(AD169, Towne, 44A, 70A, 77A, and 4E) while three isolates (16B, 21B, and 30E) had
a S27T substitution in UL84 which correlated with A390T substitutions further towards
the carboxy terminal end of the protein.
Table 8 Sequence variation within HCMV replication genes
Gene Protein Nucleotide Amino Acid Amino Acid
Length Identity (%) Identity (%) Variations
UL44 433 98.1 - 100 98.6 - 100 18
UL57 1235 99.1 - 100 99.7 - 100 16
UL70 946 98.6 - 100 99.1 - 100 36
UL105 956 99.0 - 100 99.0 - 100 31
UL102 875 98.6 - 100 99.2 - 100 39
UL84 587 97.9 - 100 97.5 - 100 73
3.3.2 Prediction of potential motifs
No motifs were detected in the HCMV replication genes when ScanProsite was
designated to exclude motifs with a high probability of occurrence. However, this also
tended to omit documented motifs which had previously been shown to be biologically
significant (Alvisi, 2005; Colletti, 2004; Graves-Woodward, 1997; Lischka, 2006). By

46
contrast, the inclusion of motifs with a high probability of occurrence generated a range
of motifs associated with post-translational modifications as well as several domains
unique to specific genes (Table 9). In addition, several compositionally biased regions
(regions consisting of a high proportion of one amino acid type) that are often abundant
in certain motifs were also observed. For example, glycine-rich domains were evident
for UL44, UL57 and UL102. A number of conserved functionally significant motifs
were identified by comparison of the HCMV proteins with other Herpesvirus
homologues, and related viral and cellular proteins discovered by the BLASTP search.
Excluding HCMV, we were able to identify 12 homologues for UL44, 34 for UL57, 29
for UL70, 12 for UL102, 32 for UL105, and 4 for UL84.
Table 9 Summary of the putative motifs identified in the HCMV replication genes
Gene
Motif UL44 UL57 UL70 UL102 UL105 UL84
N-linked glycosylation site 4 3 3 1 9 4
cAMP-dependent protein kinase phosphorylation site 1 0 1 0 2 1
Casein kinase II phosphorylation site 5 14 20 8 15 12
Protein kinase C phosphorylation site 10 15 10 7 10 12
Amidation site 1 1 1 0 0 0
N-myristoylation site 27 42 5 22 11 9
Tyrosine kinase phosphorylation site 0 1 2 2 2 0
Cell attachment sequence 1 0 0 1 0 0
Leucine zipper 0 0 0 0 0 1
Microbodies targeting signal 0 1 0 0 0 0
ATP/GTP-binding site motif A (P-loop) 0 0 0 0 1 0
Total 49 77 42 41 50 39
3.3.3 Strain variation and identification of protein motifs in UL44
Sequence alignments of the ten HCMV strains show a highly conserved protein with
variations outside of essential UL44 domains such as the DNA polymerase connector
loop (Loregian, 2004) and dimer binding domains (Appleton, 2004). A lysine to
arginine substitution was observed at residue 430 of 4E in a region that was previously
identified as a nuclear localization signal (NLS) (Alvisi, 2005). However, the
significance of the substitution is unknown although both amino acids share similar
basic properties. Interestingly, a serine to glycine substitution at residue 296 of AD169,
Towne, 44A, 70A, 77A, and 4E correlated with an isoleucine to leucine substitution in
the same isolates (Figure 4). An unusual integrin binding motif was identified at residue

47
274 for HCMV, but was not present in other homologues except pongine Herpesvirus 4.
More than half of the potential motifs identified for UL44 by ScanProsite (Table 9)
were found in the carboxy-terminal third of the gene (codons 290-433), including a
putative casein kinase-2 (CKII) signal thought to enhance the carboxy-terminal NLS of
UL44 (Alvisi, 2005). In addition, half of the motifs identified by ScanProsite also
occurred in the glycine rich region of UL44 (codons 284-397).

48
α δ α δ α δ γ UL44 1 11 21 31 41 51 61 71 81 91 Consensus MDRKTRLSEPPTLALRLKPYKTAIQQLRSVIRALKENTTVTFLPTPSLILQTVRSHCVSKITFNSSCLYITDKSFQPKTINNSTPLLGNFMYLTSSKDLT
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ............V.......................................................................................
77A ....................................................................................................
90A .................................................................................................H..
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ...Q................................................................................................
30E ....................................................................................................
δ α 101 111 121 131 141 151 161 171 181 191
Consensus KFYVQDISDLSAKISMCAPDFNMEFSSACVHGQDIVRESENSAVHVDLDFGVVADLLKWIGPHTRVKRNVKKAPCPTGTVQILVHAGPPAIKFILTNGSE
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................

49
γ δ γ ε δ η 201 211 221 231 241 251 261 271 281 291
Consensus LEFTANNRVSFHGVKNMRINVQLKNFYQTLLNCAVTKLPCTLRIVTEHDTLLYVASRNGLFAVENFLTEEPFQRGDPFDKNYVGNSGKSRGGGGGSGSLS
AD169 ...............................................................................................G....
Merlin ....................................................................................................
Towne ...............................................................................................G....
44A ................I..............................................................................G....
70A ...............................................................................................G....
77A ...............................................................................................G....
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ...............................................................................................G....
30E ....................................................................................................
η η η ζ η δ β η δ γ δ η η η 301 311 321 331 341 351 361 371 381 391
Consensus SLANAGGLHDDGPGLDNDIMNEPMGLGGLGGGGGGGGKKHDRGGGGGSGTRKMSSGGGGGDHDHGLSSKEKYEQHKITSYLTSKGGSGGGGGGGGGGLDR
AD169 ..................L.................................................................................
Merlin ....................................................................................................
Towne ..................L.................................................................................
44A ..................L.................................................................................
70A ..................L.................................................................................
77A ..................L.................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ..................L......................................S..........................................
30E ....................................................................................................

50
η γ δ 401 411 421 431
Consensus NSGNYFNDAKEESDSEDSVTFEFVPNTKKQKCG
AD169 .................................
Merlin .................................
Towne .................................
44A .................................
70A .................................
77A .................................
90A .................................
91A .................................
16B .................................
21B .................................
80B .................................
4E ..............................R..
30E .................................
Figure 4 HCMV strain alignments of UL44
Residues highlighted in yellow indicate functionally significant amino acids (Alvisi, 2005; Appleton, 2004; Loregian, 2004) while underlined sequences represent putative
motifs identified with ScanProsite. α = N-linked glycosylation site (N-x-T/S-x), β = cAMP-dependent protein kinase phosphorylation site (RR/KK/RK-x-S/T), γ = Casein
kinase II phosphorylation site (S/T-xx-D/E), δ = Protein kinase C phosphorylation site (S/T-x-R/K), ε = Cell attachment sequence (R-G-D), ζ = Amidation site (x-G-
RR/KK/RK), η = N-myristoylation site (G-A/N/C/Q/G/I/L/M/S/T/V-xx-all except P).

51
3.3.4 Strain variation and identification of protein motifs in UL57
Although no well defined functional domains exist in UL57, sequence alignments
indicate a highly conserved protein with very little variation between strains (Figure 5).
An unusual micro-bodies targeting signal was identified at the extreme C-terminus of
UL57 (residues 1233-1235) while two short glycine-rich domains spanning residues
536-591 (score = 18.78) and 1167–1226 (score =13.29) were also observed. An
alignment of the UL57 protein and its 34 homologues identified several highly
conserved cysteine residues at amino acid positions Cys107, Cys483, Cys472, and
Cys475, with the latter two forming a putative canonical disulfide bond motif that was
conserved in 97% of the homologues. Three of the conserved cysteine residues (the
disulfide bond residues Cys472 and Cys475, as well as Cys483) are homologous to a
putative zinc binding motif identified in the HSV1 homolog, ICP8 (Wang, 1990). In this
study, we observed that this domain was retained in 16 of the 35 (46%) UL57
homologues (Figure 6).

52
γ γ δ η UL57 1 11 21 31 41 51 61 71 81 91
Consensus MSHEELTALAPVGPAAFLYFSRLNAETQEILATLSLCDRSSSVVIAPLLAGLTVEADFGVSVRTPVLCYDGGVLTKVTSFCPFALYFHHTQGIVAFTEDH
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ...................................................................................T................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................
η δ 101 111 121 131 141 151 161 171 181 191
Consensus GDVHRLCEDARQKYALEAYTPEADRVPTDLAALCAAVGCQASETTVHVVVGNGLKEFLFAGQLIPCVEEATTVRLHGGEAVRVPLYPPTLFNSLQLDAEA
AD169 ...................M................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ..............................V.....................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................

53
δ δ δ γ 201 211 221 231 241 251 261 271 281 291
Consensus DEVSLDARSAFVEARGLYVPAVSETLFYYVYTSWCQSLRFSEPRVLIEAALRQFVHDSQQSVKLAPHKRYLGYMSQRLSSLEKDHLMLSDAVVCELAFSF
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ................F...................................................................................
70A ................F...................................................................................
77A ....................................................................................................
90A ................F...................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ................F...................................................................................
4E ................F...................................................................................
30E ....................................................................................................
α δ η η 301 311 321 331 341 351 361 371 381 391
Consensus ASVFFDSAYQPAESMLFSEWPLVTNATDHRDLIRALTELKLHLSTHVAALVFSANSVLYQHRLVYLQSSARHPSAGGTASQETLLKAIQFTNGLSAACED
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................

54
η η η 401 411 421 431 441 451 461 471 481 491
Consensus VYNDARKVLKFQGAPLKDERYGPQHLALVCGTCPQLVSGFVWYLNRVSVYNTGLSGSSTLTNHLVGCAAGLCEACGGTCCHTCYQTAFVRVRTRLPVVPK
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................
ζ η η η η η η δ 501 511 521 531 541 551 561 571 581 591
Consensus QPKKEPCVITVQSRFLNDVDILGSFGRRYNVDAKDGGLDGKGDDGVPGGGAGGGGGRDVSGGPSDGLGGGRGGGGGGDSGGMMGRGGRMLGASVDRTYRL
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................

55
γ δ γ η 601 611 621 631 641 651 661 671 681 691
Consensus NRILDYCRKMRLIDPVTGEDTFSAHGKSDFVAVFSALNKFVDDEALGFVSEVRLKSSRDEVAGATQAFNLDLNPYAVAFQPLLAYAYFRSVFYVIQNVAL
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................
η δ δ γ β 701 711 721 731 741 751 761 771 781 791
Consensus ITATSYIVDNPLTTNLVSKWMTQHFQSIHGAFSTTSSRKGFLFTKQIKSSKNSDHDRLLDFRLYAQGTYAVVPMEIKLSRLSVPTLIMVRVKNRPIYRAG
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....A...............................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................

56
η γ 801 811 821 831 841 851 861 871 881 891
Consensus KGNAGSVFFRRDHVPRRNPAKGCLGFLLYRHHERLFPECGLPCLQFWQKVCSNALPKNVPIGDMGEFNAFVKFLVAVTADYQEHDLLDVAPDCVLSYVES
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................
η δ δ γ η γ η α 901 911 921 931 941 951 961 971 981 991
Consensus RFHNKFLCYYGFKDYIGSLHGLTTRLTTQNHAQFPHVLGASPRFSSPAEFALHVKGLKTAGVPAPMAATVARESLVRSVFEHRSLVTVPVSVEKYAGINN
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A .......................................................S............................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................

57
η α γ γ δ δ γ 1001 1011 1021 1031 1041 1051 1061 1071 1081 1091
Consensus SKEIYQFGQIGYFSGNGVERSLNVSSMSGQDYRFMRQRYLLATRLADVLIKRSRRENVLFDADLIKNRVMLALDAENLDCDPEVMAVYEILSVREEIPAS
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................
γ η η η 1101 1111 1121 1131 1141 1151 1161 1171 1181 1191
Consensus DDVLFFVDGCEALAASLMDKFAALQEQGVEDFSLENLRRVLDADAQRLTDAAGGEVHDLSALFAPSGVGAASGVGGGGLLLGESVAGNSICFGVPGETGG
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne .....................................................................................T..............
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ..................................D.................................................................
4E ....................................................................................................
30E ....................................................................................................

58
η η ε 1201 1211 1221 1231
Consensus GCFLVNAGEDEAGGVGGSSGGGGGSGLLPAKRSRL
AD169 ...................................
Merlin ...................................
Towne ...................................
44A ...................................
70A .......S...........................
77A ...............S...................
90A .......S...........................
91A ...................................
16B ...................................
21B ...................................
80B ...................................
4E .......S...........................
30E ...................................
Figure 5 HCMV strain alignments of UL57
Underlined sequences represent putative motifs identified with ScanProsite. α = N-linked glycosylation site (N-x-T/S-x), β = Tyrosine kinase phosphorylation site (R/K-xx-
D/E-xxx-Y/R/K-xxx-D/E-xxx-Y), γ = Casein kinase II phosphorylation site (S/T-xx-D/E), δ = Protein kinase C phosphorylation site (S/T-x-R/K), ε = Microbodies C-terminal
targeting signal (S/T/A/G/C/N-R/K/H-L/I/V/M/A/F/Y) ζ = Amidation site (x-G-RR/KK/RK), η = N-myristoylation site (G-A/N/C/Q/G/I/L/M/S/T/V-xx-all except P).

59
AHV1 (O36360) YNITEYVGSAANSP-VCSLCSGQCPCVCINTLF
AtHV3 YSVAQYVGTVAVSE-LCELCQGKCPAACIHTLF
BHV1 (Q89549) DALKYVASTLEGDV-PCGLCSRDDRHACAHTTL
BHV4 YNISNYVGTAASSD-MCNLCQGKKPAVCINTLF
BHV5 (Q6X241) DALKYVASTLEGDV-PCGLCSRDDRHACAHTTL
CHV3 (Q993K9) HEIGSYVAGAATSS-VCSLCEGSTPAVCLNTLF
EBV (U75698) PETGSYVAGAAASP-MCSLCEGRAPAVCLNTLF
EHV1 DALRYLANTLESDV-PCGLCNQATRPACAHTTL
EHV2 (Q66611) YNVPTYVGTAANTP-MCELCRGSCPASCVNTLF
GHV1 (Q9QH63) SISNSGKAPCTGAVPECRWCNDESRNHCIRYTM
GTH (Q6VQA1) TLLQYLNAKTEEGA--CDLCDVETRHVCPATTF
HGTH (Q5ZR67) TLLQYLNAKTEEGA--CDLCDVETRHVCPATTF
HSV1 (X14112) DVFRYVADSNQTDV-PCNLCTFDTRHACVHTTL
HSV2 DVFRYVADSGQTDV-PCNLCTFETRHACAHTTL
HHV6 TEIYNHLVNCSAN--LCEFCDGKCCQSCIGTAM
HHV7 SEIYNHIVNCSSN--LCEFCEGKCCHSCIGTAL
HHV8 (P88904) YNVVDYVGTAAPSQ-MCDLCQGQCPAVCINTLF
HCMV STLTNHLVGCAAG--LCEACGGTCCHTCYQTAF
LHV1 (Q82172) YNIAQYISTA-----------------------
MCMV DTVFSHIVNAGSK--LCGACGGRCCHTCYATSF
PorcHV4 (Q8QS31) STLSNHIVGCASS--LCEACGGTCCHTCYNTAF
PorcHV1 (Q8JYD5) YNISQYVGSAAVSN-VCQQCKGSYPCVCINTLF
PorcHV2 (Q8B424) YNICQYVGSAAVSN-VCQQCKGNYPCVCINTLF
PorcHV3 (Q8B408) YSISQYVGSVAGSN-VCQLCHGTCPCTCLNTLF
PHV1 (Q6UDK2) NTLKFVAAETTMAA-ECRWCTETTRQYCVRHTL
RCMV HAVYQHVVHSVGN--LCEACGGRCCHTCYATPF
Rhadinovirus (Q9WRU1) FNMVHYVGTAANSE-MCTLCHGNTPATCLNTLF
RhCMV STLSNHLIGCSSS--LCGACGGTCCHTCYNTAF
SaimHV2 YSVPQYVGTAAASD-LCELCQGTCPASCIHTLF
SCMV STLSNHLIGCSSS--LCGACGGTCCHTCYNTAF
SuidHV1 (Q5PPC5) DAVRYVAGSLDAEV-PCSLCDRASRPACAHTTL
THV1 SALYEHLVHCAVN--LCPACRGRCCQSCYQTAF
THV2 SALYEHLVHCAVN--LCPACRGRCCQSCYQTAF
Turtle HV (Q5Y969) TLLQYLNAKTEEGA--CDLCDVETRHVCPATTF
VZV DALKYVTGTFDSEI-PCSLCEKHTRPVCAHTTV
Figure 6 Putative zinc finger domain of UL57.
The DNA binding protein from 35 homologues are aligned to show conservation of the cysteines residues
(grey) in a region that is otherwise poorly conserved. Accession numbers (where available) are denoted
in brackets. Species abbreviations are as follows: AHV1: Alcelaphine herpesvirus 1 (wildebeest
herpesvirus); AtHV3: Ateline herpesvirus 3; BHV1/4/5: Bovine herpesvirus 1/4/5; CHV3: Callitrichine
herpesvirus 3 (marmoset lymphocryptovirus); EBV: Epstein-Barr virus; EHV1/2: Equine herpesvirus 1/2;
FATP: Fibropapilloma-associated turtle herpesvirus; GHV1: Gallid herpesvirus 1; GTH: Green turtle
herpesvirus; HGTH: Hawaiian green turtle herpesvirus; HCMV: Human cytomegalovirus; HHV6/7/8:
Human herpesvirus 6/7/8; HSV1/2: Herpes simplex virus 1/2; LHV1: Leporid herpesvirus 1; MCMV:
Murine cytomegalovirus; MMR: Macca Mulatta rhadinovirus; PongHV4: Pongine herpesvirus 4;
PorcHV1/2/3: Porcine herpesvirus 1/2/3; PHV1: Psittacid herpesvirus 1; RCMV: Rat cytomegalovirus;
RhCMV: Rhesus cytomegalovirus; SaimHV2: Saimiriine herpesvirus 2; SCMV: Simian cytomegalovirus;
SuidHV1: Pseudorabies virus; THV1/2: Tupaiid herpesvirus 1/2; VZV: Varicella-Zoster virus

60
3.3.5 Strain variation and identification of protein motifs in UL70
Sequence alignments of the ten HCMV strains show that most variations occur in the C-
terminal third of the protein (Figure 7) and that key functional motifs predicted in HSV
are also highly conserved in HCMV (Biswas, 1999; Dracheva, 1995) (Figure 8A). In
addition, alignment of the UL70 homologues reaffirms the importance of the highly
conserved DxD catalytic motif (Dracheva, 1995) in a region that is otherwise poorly
conserved. The HCMV primase also retains a putative zinc finger motif originally
identified in HSV (Biswas, 1999). The zinc finger motif, spanning residues 881-920,
remained highly conserved amongst the HCMV isolates studied and was present in 90%
of the aligned homologues. In addition, a key phenylalanine residue involved in zinc
finger structural stability (Michael, 1992) was present in all HCMV strains at position
896 and conserved in 22 of the 30 (73%) homologues (Figure 8D). Further analysis also
revealed a pair of cysteines conserved across all 30 homologues (Figure 8C) while a
previously uncharacterized domain spanning amino acids 566-572 contained several
highly conserved aromatic residues (Figure 8B).

61
γ γ δ δ δ α γ UL70 1 11 21 31 41 51 61 71 81 91
Consensus MTLVLFATEYDSAHIVANVLSQTPTDHCVFPLLVKHQVSRRVYFCLQTQKCSDSRRVAPVFAVNNETLQLSRYLAARQPIPLSALIASLDEAETQPLYRH
AD169 ....................................................................................................
Merlin ..............................................................................................R.....
Towne ....................................................................................................
44A ..............................................................................................R.....
70A ..............................................................................................R.....
77A ....................................................................................................
90A ....................................................................................................
91A ..............................................................................................R.....
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................
η γ γ 101 111 121 131 141 151 161 171 181 191
Consensus LFRTPVLSPEHGGEVREFKHLVYFHHAAVLRHLNQVFLCPTSPSWFISVFGHTEGQVLLTMAYYLFEGQYSTISTVEEYVRSFCTRDLGTIIPTHASMGE
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................

62
γ ζ 201 211 221 231 241 251 261 271 281 291
Consensus FARLLLGSPFRQRVSAFVAYAVARNRRDYTELEQVDTQINAFRERARLPDTVCVHYVYLAYRTALARARLLEYRRVVAYDADAAPEAQCTREPGFLGRRL
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................
β η η γ γ δ 301 311 321 331 341 351 361 371 381 391
Consensus STELLDVMQKYFSLDNFLHDYVETHLLRLDESPHSATSPHGLGLAGYGGRIDGTHLAGFFGTSTQLARQLERINTLSESVFSPLERSLSGLLRLCASLRT
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................

63
δ γ 401 411 421 431 441 451 461 471 481 491
Consensus AQTYTTGTLTRYSQRRYLLPEPALAPLLERPLPVYRVHLPNDQHVFCAVASETWHRSLFPRDLLRHVPDSRFSDEALTETVWLHDDDVASTSPETQFYYT
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................
γ γ γ γ 501 511 521 531 541 551 561 571 581 591
Consensus RHEVFNERLPVFNFVADFDLRLRDGVSGLARHTVFELCRGLRRVWMTVWASLFGYTHPDRHPVYFFKSACPPNSVPVDAAGAPFDDDDYLDYRDERDTEE
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A .................................I................................................S.................
70A ....................................................................................................
77A ....................................................................................................
90A ..................................................................................S.................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ..................................................................................S.................
30E ....................................................................................................

64
γ γ γ γ δ 601 611 621 631 641 651 661 671 681 691
Consensus DEDGKEDKNNVPDNGVFQKTTSSVDTSPPYCRCKGKLGLRIITPFPACTVAVHPSVLRAVAQVLNHAVCLDAELHTLLDPISHPESSLDTGIYHHGRSVR
AD169 ....................................................................................................
Merlin ............G....................................I..................................................
Towne ......N.............................................................................................
44A ....................................................................................................
70A ............G....................................I..................................................
77A .......................I............................................................................
90A ...................P..T..........................I..................................................
91A ............G....................................I..................................................
16B ..........................L.........................................................................
21B ..........................L.........................................................................
80B ............G.......................................................................................
4E ...................P..T.............................................................................
30E ..........................L.........................................................................
ε γ δ α ε 701 711 721 731 741 751 761 771 781 791
Consensus LPYMYKMDQDDGYFMHRRLLPLFIVPDAYREHPLGFVRAQLDLRNLLHHHPPHDLPALPLSPPPRVILSVRDKICPSTEANFIETRSLNVTRYRRRGLTE
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ......................................................................H.............................
21B ......................................................................H.............................
80B .............................................................................S......................
4E .............................................................................................Q......
30E ......................................................................H.............................

65
η γ α δ 801 811 821 831 841 851 861 871 881 891
Consensus VLAYHLYGGDGATAAAISDTDLQRLVVTRVWPPLLEHLTQHYEPHVSEQFTAPHVLLFQPHGACCVAVKRRDGARTRDFRCLNYTHRNPQETVQVFIDLR
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................
δ δ 901 911 921 931 941
Consensus TEHSYALWASLWSRCFTKKCHSNAKNVHISIKIRPPDAPMPPATAV
AD169 .......................................V......
Merlin ..............................................
Towne ..............................................
44A ..............................................
70A .......................................V......
77A .......................................V......
90A ..............................................
91A ..............................................
16B .......................................V......
21B .......................................V......
80B ..............................................
4E ..............................................
30E .......................................V......
Figure 7 HCMV strain alignments of UL70
Residues highlighted in yellow indicate functionally significant amino acids (Biswas, 1999; Dracheva, 1995) while underlined sequences represent putative motifs identified
with ScanProsite. α = N-linked glycosylation site (N-x-T/S-x), β = cAMP-dependent protein kinase phosphorylation site (RR/KK/RK-x-S/T), γ = Casein kinase II
phosphorylation site (S/T-xx-D/E), δ = Protein kinase C phosphorylation site (S/T-x-R/K), ε = Tyrosine kinase phosphorylation site (R/K-xx-D/E-xxx-Y/R/K-xxx-D/E-xxx-
Y), ζ = Amidation site (x-G-RR/KK/RK), η = N-myristoylation site (G-A/N/C/Q/G/I/L/M/S/T/V-xx-all except P).

66
518 A 526 562 B 566 633 C 644 883 D 924
HCMV FVADFDLRL PVYFFKSACP PYCRCKGKLGLRI FRCLNYTHRN.24.SRCFTKKCHS
AHV1 WVLDLDLPI EVFFFKSACI TFCTCTEKLGMRV FKCLRYNHRG.24.SQCFANKCQS
AtHV3 LVLDLDLHI PVYFFKSSCE KFCYCTKKLGFRI FSCLNFKHKL.24.SQCFASKCNN
BHV1 LVMDFDLKV PVYFFKSAC- RFCNCNSKIGLRI FRCLTFNHRG.24.SQCFANKCNS
BHV4 IVLDVDFRL PCFFYKSACP AACGCEDKMGFRV FGCLRAAHGR.25.QQCFATKCGS
BHV5 IVLDVDFRL PCFFYKSACP TACGCDDKMGFRV FGCLRAAHGR.25.QQCFATKCGS
CercoHV1 IILDLDVAL PCYFFKSACS PRCTCAKKIGLRV LSCLRFKHGR.25.QQCFASKCDS
CercoHV9 IILDVDFHV PCYFYKSSC- NTCSCNKNLGFRV FTCLRYPH-R.25.QQCFATKCDS
CHV3 LVLDMDVKI PVYFFKSACP PFCICREKLGLRV FKCLRYQHRN.24.SQCFSGRCGS
EBV LVLDLDLKI PVYFFKSACP PFCICTGKLGFRV FACLRHTHRA.24.SQCFAGRCGA
EHV1 IILDVDFGI PCYFYKTSCP DACECTEKMGFRV FSCLRAKHLR.25.QQCFATKCGN
EHV2 LVLDFDLPL PVYFFKSACP PFCTCDAKLGLRI FVCISSNHRN.24.SQCFAHKCNS
EHV4 IILDVDFDI PCYFYKTQCP NACQCTEKMGFRI FSCLRAKHLR.25.QQCFATKCGN
GHV1 LILDIDIPL PVYFYKSHCS TLCKCKEKLGFRV FTCLKHSHRS.25.D---------
HHV6 YIGDLDLPL PIFFFKTQCD QFCVCRKKIGLRI FSCLNRQHRG.24.STCFATKCQS
HHV7 YIGDLDLPL PIFFFKTTCS AFCVCKKKIGLRI FSCLTRNHKG.24.SKCFTTKCKS
HHV8 WVLDFDLPV PVYFFKSACP SFCRCHDKLGMRI FLCINHNHKN.24.SQCFASKCNN
HSV1 IILDLDIAL PCYFFKSACR PVCSCTDKIGLRV LSCLRFKHGR.25.QQCFAAKCDS
HSV2 IILDLDIAL PCYFFKSACR VVCSCADKIGLRV LSCLRFKHGR.25.QQCFATKCDN
MCMV FVGDVDLKL PVFFFKSAC- PFCVCRRKLGLRV FSCLARETYT.24.SRCFTRRCNS
MDHV1 IVLDVDIHL PCYFYKSSCK KPCGCHDKIGLRV FTCVRFKHAR.25.QQCFAAKCGN
MMR WVLDFDLKV PVYFFKSACP AFCHCDAKIGMRI FFCINHKHRN.24.SQCFAAKCNH
PhoHV2 LVLDFDLPL QVYFFKSSCP MFCNCSEKLGLRV ------------------------
PHV1 LPLDLDITL PVYFYKTQCD RFCRCERKIGFRI FTCLKYQHRG.25.TRCFATKCGS
PongHV4 FVADMDLRL PVYFFKSACR LYCRCTEKLGLRI FRCLNYVHRN.24.SRCFTKKCHS
RCMV FVGDLDLKL PVYFFKSACD -FCTCRRKIGMRI FRCLTTEHHF.25.SRCFANKCQS
RhCMV FIADFDLRL PVYFFKSACK DYCKCSEKLGLRI FRCLNYTHRN.24.SRCFTKKCHS
SaimHV2 LVLDLDLHI HVYFFKSACE KFCYCTKKLGFRI FSCLNFKHKL.24.SQCFASKCNS
THV1 LALDVDLAL PVYFFKSACP DFCACRAKLGFRV FRCLSRAHRI.24.SRCFATKCRS
VZV LILDVDFHI PCYFYKTACP LPCNCKEKIGFRV FTCLRYPH-R.25.QQCFATKCDS
* * * * * * * * * * * * ** *
Figure 8 Conserved domains between UL70 homologues.
(A) The DxD catalytic motif. (B) Domain consisting of several conserved aromatic residues. (C)
Conserved cysteines. (D) Putative zinc finger domain. Species abbreviations are as follows: AHV1:
Alcelaphine herpesvirus 1 (wildebeest herpesvirus); AtHV3: Ateline herpesvirus 3; BHV1/4/5: Bovine
herpesvirus 1/4/5; CercoHV1: Cercopothecine herpesvirus 1 (Simian herpes B virus); CercoHV9:
Cercopothecine herpesvirus 9 (Simian varicella virus); CHV3: Callitrichine herpesvirus 3 (marmoset
lymphocryptovirus); EBV: Epstein-Barr virus; EHV1/2/4: Equine herpesvirus 1/2/4; GHV1: Gallid
herpesvirus 1; HCMV: Human cytomegalovirus; HHV6/7/8: Human herpesvirus 6/7/8; HSV1/2: Herpes
simplex virus 1/2; MCMV: Murine cytomegalovirus; MDHV1: Meleagrid herpesvirus 1; MMR: Macca
Mulatta rhadinovirus; PongHV4: Pongine herpesvirus 4; PorcHV1/2/3: Porince herpesvirus 1/2/3;
PhoHV2: Phocid herpesvirus 2; PHV1: Psittacid herpesvirus 1; RCMV: Rat cytomegalovirus; RhCMV:
Rhesus cytomegalovirus; SaimHV2: Saimiriine herpesvirus 2; THV1: Tupaiid herpesvirus 1; VZV:
Varicella-Zoster virus. * denotes highly conserved residues

67
3.3.6 Strain variation and identification of protein motifs in UL102
Sequence alignments of the UL102 isolates show a highly conserved protein, although
alignments with homologues yielded a low degree of conservation for any region. As
with UL44, a putative integrin binding motif was also detected in UL102 at amino acid
residue 58. An alanine to glycine substitution at residue 76 in AD169 allowed the
formation of another N-myristoylation site while the insertion of additional glycines in
isolates 44A and 80B also increased the number of N-myristoylation sites formed
(Figure 9). ScanProsite detected a glycine-rich domain (score = 12.88) between residues
553-687 and a short serine-rich domain spanning residues 795-836 (score = 12.8;
normative score = 7.22). Similar to UL44, the glycine-rich domain in UL102 accounted
for a significant number of the motifs identified. Isolate variation within the glycine-rich
region spanning residues 629-633 was observed, with some isolates containing
additional and deleted glycine residues. Similar variations were not observed in the
glycine rich regions of UL44 or UL57.

68
η η ζ η γ UL102 1 11 21 31 41 51 61 71 81 91
Consensus MTAQPPLHHRHHPYTLFGTSCHLSWYGLLEASVPIVQCLFLDLGGGRAEPRLHTFVVRGDRLPPAEVRAVHRASYAALASAVTTDADERRRGLEQRSAVL
AD169 ..........................................................................-G........................
Merlin ....................................................................................................
Towne ..............A.....................................................................................
44A .....S..............................................................................................
70A .............C......................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................V...............................
30E ....................................................................................................
γ η γ 101 111 121 131 141 151 161 171 181 191
Consensus ARVLLEGSALIRVLARTFTPVQIQTDASGVEILEAAPALGVETAALSNALSLFHVAKLVVIGSYPEVHEPRVVTHAAERVSEEYGTHAHKKLRRGYYAYD
AD169 ...........................................................................T........................
Merlin ...........................................T........................................................
Towne .....................................................................S..............................
44A ............................................................................T.......................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ...........................................T........................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................

69
δ δ η 201 211 221 231 241 251 261 271 281 291
Consensus LAMSFRVGTHKYVLERDDEAVLARLFEVREVCFLRTCLRLVTPVGFVAVAVTDEQCCLLLQSAWTHLYDVLFRGFAGQPPLRDYLGPDLFETGAARSFFF
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................
γ η α 301 311 321 331 341 351 361 371 381 391
Consensus PGFPPVPVYAVHGLHTLMRETALDAAAEVLSWCGLPDIVGSAGKLEVEPCALSLGVPEDEWQVFGTEAGGGAVRLNATAFRERPAGGDRRWLLPPLPRDD
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ......................................................................................S.............
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................

70
β δ δ 401 411 421 431 441 451 461 471 481 491
Consensus GDGENNVVEVSSSTGGAHPPSDDATFTVHVRDATLHRVLIVDLVERVLAKCVRARDFNPYVRYSHRLHTYAVCEKFIENLRFRSRRAFWQIQSLLGYISE
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ...........................................L........................................................
21B ...........................................L........................................................
80B ....................................................................................................
4E ....................................................................................................
30E ...........................................L........................................................
β α δ η 501 511 521 531 541 551 561 571 581 591
Consensus HVTSACASAGLLWVLSRGHREFYVYDGYSGHGPVSAEVCVRTVVDCYWRKLFGGDDPGPTCRVQESAPGVLLVWGDERLVGPFNFFYGNGGAGGSPLHGV
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ..........................................................L.........................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ................................................................................................F...
30E ....................................................................................................
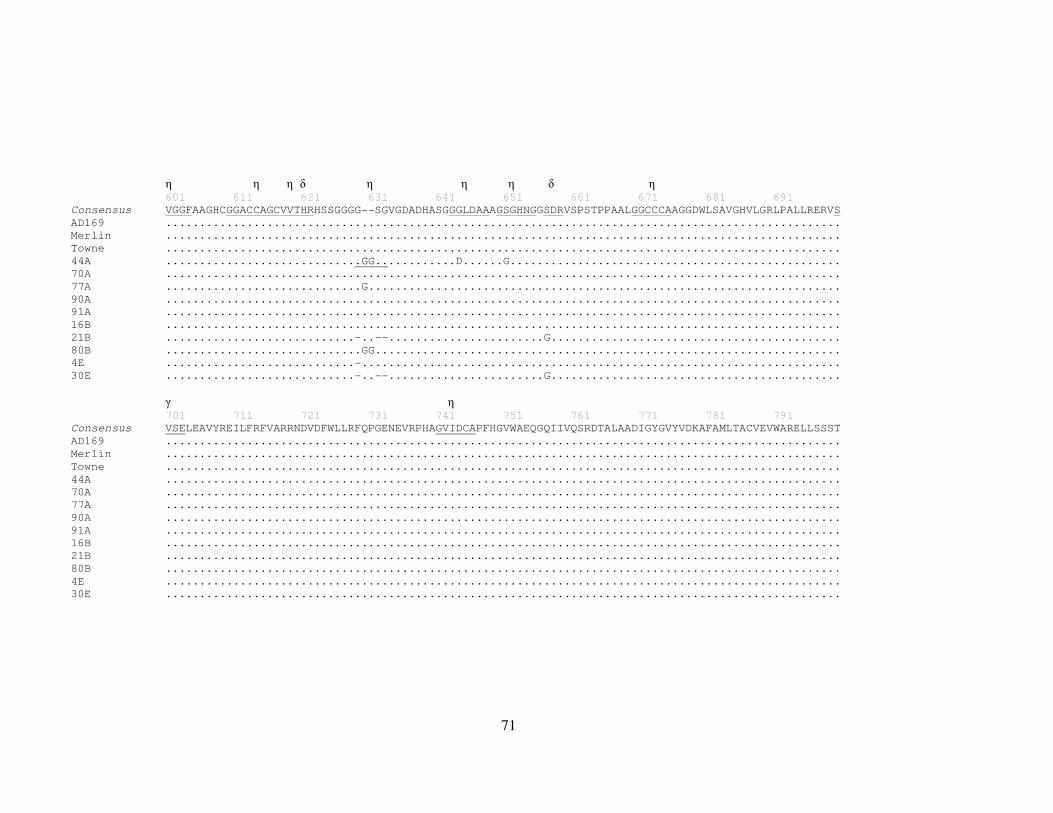
71
η η η δ η η η δ η 601 611 621 631 641 651 661 671 681 691
Consensus VGGFAAGHCGGACCAGCVVTHRHSSGGGG--SGVGDADHASGGGLDAAAGSGHNGGSDRVSPSTPPAALGGCCCAAGGDWLSAVGHVLGRLPALLRERVS
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A .............................GG............D......G.................................................
70A ....................................................................................................
77A .............................G......................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ............................-..--.......................G...........................................
80B .............................GG.....................................................................
4E ............................-.......................................................................
30E ............................-..--.......................G...........................................
γ η 701 711 721 731 741 751 761 771 781 791
Consensus VSELEAVYREILFRFVARRNDVDFWLLRFQPGENEVRPHAGVIDCAPFHGVWAEQGQIIVQSRDTALAADIGYGVYVDKAFAMLTACVEVWARELLSSST
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................

72
γ 801 811 821 831 841 851 861 871
Consensus ASTTACSSSSVLSSALPSVTSSSSGTATVSPPSCSSSSATWLEERDEWVRSLAVDAQHAAKRVASEGLRFFRLNA
AD169 ...........................................................................
Merlin ....T......................................................................
Towne ...........................................................................
44A ...........................................................................
70A .........-.................................................................
77A ...........................................................................
90A ...........................................................................
91A ....T......................................................................
16B ...........................................................................
21B ...........................................................................
80B ....T......................................................................
4E ....T....-.................................................................
30E ...........................................................................
Figure 9 HCMV strain alignments for UL102
Underlined sequences represent putative motifs identified with ScanProsite. α = N-linked glycosylation site (N-x-T/S-x), β = Tyrosine kinase phosphorylation site (R/K-xx-
D/E-xxx-Y/R/K-xxx-D/E-xxx-Y), γ = Casein kinase II phosphorylation site (S/T-xx-D/E), δ = Protein kinase C phosphorylation site (S/T-x-R/K), ζ = Cell attachment site (R-
G-D), η = N-myristoylation site (G-A/N/C/Q/G/I/L/M/S/T/V-xx-all except P).

73
3.3.7 Strain variation and identification of protein motifs in UL105
Sequence alignments of the UL105 isolates show a highly conserved protein with sparse
variation occurring outside of predicted functional domains (Figure 10). UL105 is
postulated to contain six motifs typical of superfamily-1 class of helicase proteins (Zhu,
1992) based on studies of the HSV helicase, UL5 (Graves-Woodward, 1997).
ScanProsite detected an ATP/GTP-binding site in motif I as well as a putative
cAMP/cGMP-dependent protein kinase phosphorylation site in Motif IV. Protein
alignments of the UL105 homologues predicted 2 potential N-linked glycosylation sites
as well as a potential CKII phosphorylation site based on conservation across 33
homologues (Figure 11).

74
δ δ α γ UL105 1 11 21 31 41 51 61 71 81 91
Consensus MSMTASSSTPRPTPKYDDALILNLSSAAKIERIVDKVKSLSRERFAPEDFSFQWFRSISRVERTTDNNPSAATTAAATTTVHSSASSSAAAAASSEAGGT
AD169 ....................................................................................................
Merlin ..............................................................................A.....V...............
Towne ....................................................................................................
44A ....................................................................................................
70A ..I..........S......................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ..............................................................................A.....V...............
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ..............................................................................A.....................
30E ....................................................................................................
ζ α α 101 111 121 131 141 151 161 171 181 191
Consensus RVPCVDRWPFFPFRALLVTGTAGAGKTSSIQVLAANLDCVITGTTVIAAQNLSAILNRTRSAQVKTIYRVFGFVSKHVPLADSAVSHETLERYRVCEPHE
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................

75
201 211 221 231 241 251 261 271 281 291
Consensus ETTIQRLQINDLLAYWPVIADIVDKCLNMWERKAASASAAAAAAACEDLSELCESNIIVIDECGLMLRYMLQVVVFFYYFYNALGDTRLYRERRVPCIIC
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B .............................G.......................................VVG..L.........................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................
γ γ α δ β δ 301 311 321 331 341 351 361 371 381 391
Consensus VGSPTQTEALESRYDHYTQNKSVRKGVDVLSALIQNEVLINYCDIADNWVMFIHNKRCTDLDFGDLLKYMEFGIPLKEEHVAYVDRFVRPPSSIRNPSYA
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................

76
γ δ ε α α 401 411 421 431 441 451 461 471 481 491
Consensus AEMTRLFLSHVEVQAYFKRLHEQIRLSERHRLFDLPVYCVVNNRAYQELCELADPLGDSPQPVELWFRQNLARIINYSQFVDHNLSSEITKEALRPAADV
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B D.............................C.....................................................................
80B ................................................................F...................................
4E ....................................................................................................
30E ....................................................................................................
α η η δ δ η γ γ ε 501 511 521 531 541 551 561 571 581 591
Consensus VATNNSSVQAHGGGGSVIGSTGGNDETAFFQDDDTTTAPDSRETLLTLRITYIKGSSVGVNSKVRACVIGYQGTVERFVDILQKDTFIERTPCEQAAYAY
AD169 ....................................................................................................
Merlin .....P..............................................................................................
Towne ....................................................................................................
44A .........T..........................................................................................
70A ....................................................................................................
77A .....P..............................................................................................
90A ..................................I.................................................................
91A .....P..............................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ..N.-P..K...........................................................................................
4E .....P..............................................................................................
30E ....................................................................................................
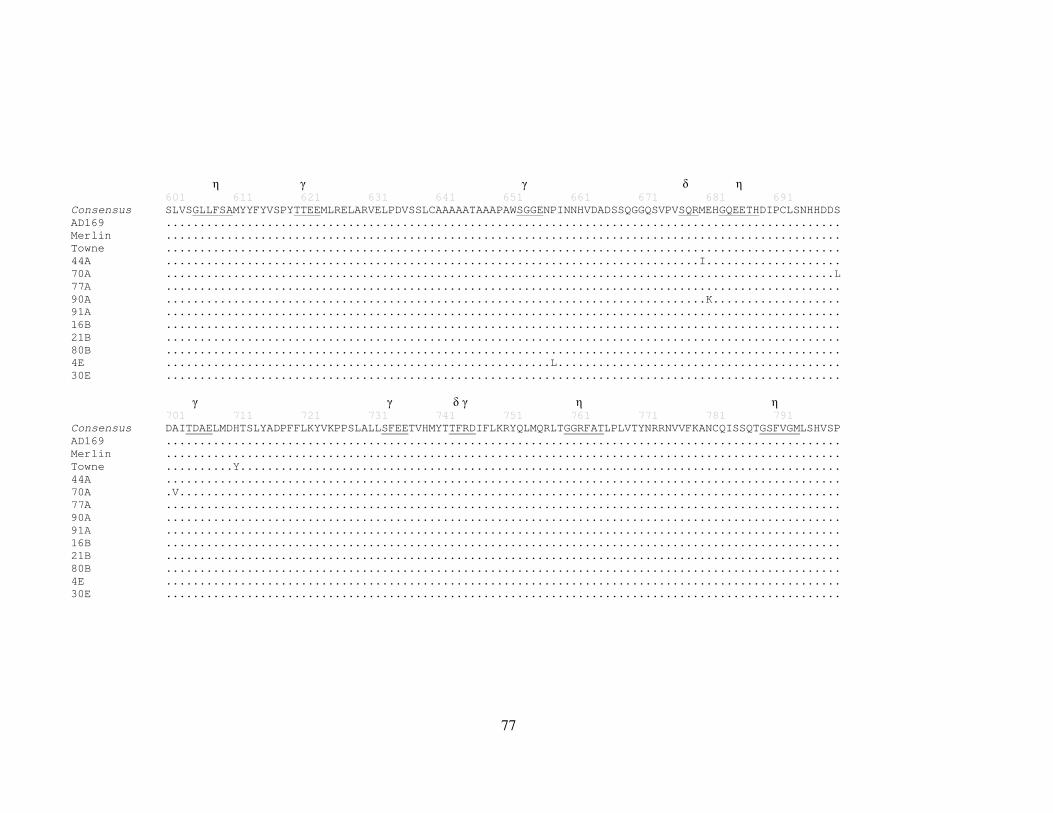
77
η γ γ δ η 601 611 621 631 641 651 661 671 681 691
Consensus SLVSGLLFSAMYYFYVSPYTTEEMLRELARVELPDVSSLCAAAAATAAAPAWSGGENPINNHVDADSSQGGQSVPVSQRMEHGQEETHDIPCLSNHHDDS
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ...............................................................................I....................
70A ...................................................................................................L
77A ....................................................................................................
90A ................................................................................K...................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E .........................................................L..........................................
30E ....................................................................................................
γ γ δ γ η η 701 711 721 731 741 751 761 771 781 791
Consensus DAITDAELMDHTSLYADPFFLKYVKPPSLALLSFEETVHMYTTFRDIFLKRYQLMQRLTGGRFATLPLVTYNRRNVVFKANCQISSQTGSFVGMLSHVSP
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ..........Y.........................................................................................
44A ....................................................................................................
70A .V..................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................

78
δ α γ δ 801 811 821 831 841 851 861 871 881 891
Consensus AQTYTLEGYTSDNVLSLPSDRHRIHPEVVQRGLSRLVLRDALGFLFVLDVNVSRFVESAQGKSLHVCTTVDYGLTSRTAMTIAKSQGLSLEKVAVDFGDH
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ....................................................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
80B ....................................................................................................
4E ....................................................................................................
30E ....................................................................................................
γ β 901 911 921 931 941 951
Consensus PKNLKMSHIYVAMSRVTDPEHLMMNVNPLRLPYEKNTAITPYICRALKDKRTTLIF
AD169 ........................................................
Merlin ........................................................
Towne ........................................................
44A ........................................................
70A ........................................................
77A ........................................................
90A ........................................................
91A ........................................................
16B ........................................................
21B ........................................................
80B ........................................................
4E ........................................................
30E ........................................................
Figure 10 HCMV strain alignments for UL105
Residues highlighted in yellow indicate functionally significant motifs (Graves-Woodward, 1996; Graves-Woodward, 1997) while underlined sequences represent putative
motifs identified with ScanProsite. α = N-linked glycosylation site (N-x-T/S-x), β = cAMP-dependent protein kinase phosphorylation site (RR/KK/RK-x-S/T), γ = Casein
kinase II phosphorylation site (S/T-xx-D/E), δ = Protein kinase C phosphorylation site (S/T-x-R/K), ε = Tyrosine kinase phosphorylation site (R/K-xx-D/E-xxx-Y/R/K-xxx-
D/E-xxx-Y), ζ = ATP/GTP-binding site motif A (A/G-xxxx-G-K-S/T), η = N-myristoylation site (G-A/N/C/Q/G/I/L/M/S/T/V-xx-all except P).

79
20 A 27 409 B 415 473 C 481
HCMV LILNLSSAA MTRLFLSHVEVQA RIINYSQFV
AHV1 FILNMTSEA WTRLFLSHSEVKQ RLSNYSQFI
AtHV3 FILNMTSDA WTRLFLSHAEVKS RLSNYSQFI
BHV1 FILNMTSDA WTRLFLSHNEVKA RLSNYSQFI
BHV4 VFLNFTSMH WTRLYSSHREVSA RITNYSQSR
BHV5 VFLNFTSMH WTRLYSSHREVSA RITNYSQSR
CHV3 FMLNMTSDA WTRLFLSHAEVKQ RLGNYSQFA
CercoHV1 VYLNFTSMH WTRLYSSHKEVSA RLNNYSQSR
CercoHV9 AFLNFTSMH WTRLFSSHKEVSA RITNYSQSQ
EBV FMLNMTSDA WTRLFLSHAEVKT RLGNYSQFA
EHV1 VYLNFTSMH WTRLYSSHKEVSA RVSNYSQSR
EHV2 FILNMTSDS WTRLFVSHREVKA RLSNYSQFI
EHV4 VYLNFTSMH WTRLYSSHKEVSA RVSNYSQSR
GHV1 TYLNFTAMH WTRLYSSHKEVSA RLSNWSQSR
GHV3 VYLNFSAMQ WTRLFSSHEEVKE RLGNYSQSR
HSV1 AFLNFTSMH WTRLFSSHKEVSA RITNYSQSQ
HSV2 AFLNFTSMH WTRLFSSHKEVSA RITNYSQSQ
HHV6 FLLNMSSAP TTRLFLSHNEVKN RLNTYSQFA
HHV7 FLLNMSSAA MTRLFLSHYEVKS RLNTYSQFA
HHV8 FILNMTSDA WTRLFISHQEVKS RLSNYSQFA
MCMV FVLNMSSAS VTRLFISHAEVKR RISNYSQFT
MDHV1 TYLNFTAMH WTRLYSSHKEVST RLSNWSQSR
MD TYLNFTAMH WTRLYSSHKEVSA RLSNWSQSR
MMR FILNMTSDA WTRLFLSHSEVKA KLSNYSQFV
PorcHV1 FFLNMTSDS WTRLFLSHIEVKN RLSNYSQFV
PorcHV2 FFLNMTSDA WTRLFLSHAEVKN RLSNYSQFV
PongHV4 LILNLSSAA MTRLFLSHAEVQL RIINYSQFV
Pseudorabies TYLNFTSMH WTRLYSSHREVSA RITNYSQSR
RCMV FVLNTSSAL ATRLFVSHREVKD RIGNYSQFT
RhCMV LILNLSSAA MTRLFLSHVEVQA RIINYSQFV
SuidHV1 TYLNFTSMH WTRLYSSHREVSA RITNYSQSR
THV1 FILNMSSAV MTRLFISHAEVKR RIINYSQFV
VZV VYLNFTSMH WTRLYSSHKEVSA RLHNYSQSR
*** * * *
Figure 11 Putative motifs conserved across UL105 homologues
(A) and (C) Conserved putative N-linked glycosylation sites. (B) Conserved putative CK2
phosphorylation site. Species abbreviations are as follows: AHV1: Alcelaphine herpesvirus 1 (wildebeest
herpesvirus); AtHV3: Ateline herpesvirus 3; BHV1/4/5: Bovine herpesvirus 1/4/5; CercoHV1:
Cercopothecine herpesvirus 1 (Simian herpes B virus); CercoHV9: Cercopothecine herpesvirus 9 (Simian
varicella virus); CHV3: Callitrichine herpesvirus 3 (marmoset lymphocryptovirus); EBV: Epstein-Barr
virus; EHV1/2/4: Equine herpesvirus 1/2/4; GHV1/3: Gallid herpesvirus 1/3; HCMV: Human
cytomegalovirus; HHV6/7/8: Human herpesvirus 6/7/8; HSV1/2: Herpes simplex virus 1/2; MCMV:
Murine cytomegalovirus; MDHV1: Meleagrid herpesvirus 1; MD: Marek’s disease; MMR: Macca
Mulatta rhadinovirus; PongHV4: Pongine herpesvirus 4; PorcHV1/2: Porince herpesvirus 1/2; RCMV:
Rat cytomegalovirus; RhCMV: Rhesus cytomegalovirus; SuidHV1: Pseudorabies virus; THV1: Tupaiid
herpesvirus 1; VZV: Varicella-Zoster virus.

80
3.3.8 Strain variation and identification of protein motifs in UL84
Sequence alignments revealed a high degree of variation amongst the isolates (Figure
12), especially in a region that has been associated with oligomerization of the UL84
protein (Colletti, 2004). In particular, deletions of glutamine residues in three clinical
isolates (16B, 21B and 30E) were observed in a region that was previously associated
with nuclear import (Xu, 2002) and is part of a previously described hydrophobic region
(He, 1992). Two overlapping CKII phosphorylation sites (consensus site = S/TxxD/E)
(Pinna, 1990) TLQE and SQEE, were detected flanking this region (Figure 13) and
interestingly, three strains contained only a single CKII site; two strains (AD169 and
70A) had substitutions which abolished the second motif while another isolate (77A)
abolished the first motif. ScanProsite also detected a previously described leucine zipper
domain (He, 1992) at the amino-terminus of UL84.

81
η η η η δ α δ η γ γ γ η γ δ UL84 1 11 21 31 41 51 61 71 81 91
consensus MPRVDPNLRNRARRPRARRGGGGGVGSNSSRHSGKCRRQRRALSAPPLTFLATTTTTTMMGVASTDDDSLLLKTPDELDKHSGSPQTILTLTDKHDIRQP
AD169 ................................................................................Y...................
Merlin ............................................T.......................................................
Towne ...A................................................................................................
44A ....................................................................................................
70A ................................................................................Y...................
77A ............................................T.......................................................
90A ............................................T.......................................................
91A ............................................T.......................................................
16B ..........................T.........................................................................
21B ..........................T.........................................................................
4E ........................................................R...........................................
30E ..........................T.........................................................................
ε γ γ γ 101 111 121 131 141 151 161 171 181 191
consensus RVHRGTYHLIQLHLDLRPEELRDPFQILLSTPLQLGEANGESQTAPATSQEEETAASHEPEKKKEKEE-KKEEDEDDRNDDRERGILCVVSNEDSDVRPA
AD169 .......................................D........L.................Q.................................
Merlin ....................................................................K.E.............................
Towne ...........................................................L........K.E.............................
44A ............................................DL.....................K................................
70A ................................................L.................Q.................................
77A ..............................................TM....................K.E.............................
90A ..............................................T.....................K.E.............................
91A ....................................................................K.E.............................
16B ...................................................................-................................
21B ...................................................................-................................
4E ....................................................................................................
30E ...................................................................-................................

82
γ α γ γ 201 211 221 231 241 251 261 271 281 291
consensus FSLFPARPGCHILRSVIDQQLTRMAIVRLSLNLFALRIITPLLKRLPLRRKAAHHTALHDCLALHLPELTFEPTLDINNVTENAASVAD AESTDADLTP
AD169 .........................................................................................T..........
Merlin .........................................................................................T..........
Towne .........................................P...V...............M..........S................A..........
44A .................................................................................................... 70A .........................................................................................T..........
77A ....................................................................................V.----.....T....
90A .........................................................................................T..........
91A .........................................................................................T..........
16B ......................................................................................A..A..........
21B ......................................................................................A..A..........
4E .....................................................................................TA..A..........
30E ......................................................................................A..A..........
δ δ δ δ γ α β 301 311 321 331 341 351 361 371 381 391
consensus TLTVRVRHALCWHRVEGGISGPRGLTSRISARLSETTAKTLGPSVFGRLELDPNESPPDLTLSSLTLYQDGILRFNVTCDRTEAPADPVAFRLRLRRETV
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne .........V.............................................................M............................
44A .......................................................................M............................
70A ....................................................................................................
77A .......................................................................M............................
90A ....................................................................................................
91A ....................................................................................................
16B .........................................................................................T..........
21B .........................................................................................T..........
4E ....................................................................................................
30E .........................................................................................T..........

83
δ δ δ γ 401 411 421 431 441 451 461 471 481 491
consensus RRPFFSDAPLPYFVPPRSGAADEGLEVRVPYELTLKNSHTLRIYRRFYGPYLGVFVPHNRQGLKMPVTVWLPRSWLELTVLVSDENGATFPRDALLGRLY
AD169 ....................................................................................................
Merlin ....................................................................................................
Towne ....................................................................................................
44A ....................................................................................................
70A ....................................................................................................
77A ...........................H........................................................................
90A ....................................................................................................
91A ....................................................................................................
16B ....................................................................................................
21B ....................................................................................................
4E ...........................................................C........................................
30E ....................................................................................................
δ η η α δ 501 511 521 531 541 551 561 571 581
consensus FISSKHTLNRGCLSAMTHQVKSTLHSRSTSHSPSQQQLSVLGASIALEDLLPMRLASPETEPQDCKLTENTTEKTSPVTLAMVCGDL
AD169 .......................................................................................
Merlin .......................................................................................
Towne .......................................................................................
44A .......................................................................................
70A ..............................Y........................................................
77A .......................................................................................
90A .......................................................................................
91A .......................................................................................
16B .......................................................................................
21B .......................................................................................
4E ...................................................................M....G..............
30E .......................................................................................
Figure 12 HCMV strain alignments for UL84
Residues highlighted in yellow indicate functionally significant motifs (Colletti, 2004; He, 1992; Lischka, 2006) while underlined sequences represent putative motifs
identified with ScanProsite. α = N-linked glycosylation site (N-x-T/S-x), β = cAMP-dependent protein kinase phosphorylation site (RR/KK/RK-x-S/T), γ = Casein kinase II
phosphorylation site (S/T-xx-D/E), δ = Protein kinase C phosphorylation site (S/T-x-R/K), ε = Leucine zipper (L-xxxxxx-L-xxxxxx-L-xxxxxx-L), η = N-myristoylation site
(G-A/N/C/Q/G/I/L/M/S/T/V-xx-all except P).

84
Consensus PATSQEEETAASHEPEKKKEKEE-KKEEDEDDRNDDRERGILCVVSNEDSDVRPA
AD169 PATLQEEETAASHEPEKKKEKQEKKEE-DEDDRNDDRERGILCVVSNEDSDVRPA
Merlin PATSQEEETAASHEPEKKKEKEEKKEEEDEDDRNDDRERGILCVVSNEDSDVRPA
Towne PATSQEEETAASHELEKKKEKEEKKEEEDEDDRNDDRERGILCVVSNEDSDVRPA
44A LATSQEEETAASHEPEKKKEKEKKKEE-DEDDRNDDRERGILCVVSNEDSDVRPA
70A PATLQEEETAASHEPEKKKEKQEKKEE-DEDDRNDDRERGILCVVSNEDSDVRPA
77A PTMSQEEETAASHEPEKKKEKEEKKEEEDEDDRNDDRERGILCVVSNEDSDVRPA
90A PTTSQEEETAASHEPEKKKEKEEKKEEEDEDDRNDDRERGILCVVSNEDSDVRPA
91A PATSQEEETAASHEPEKKKEKEEKKEEEDEDDRNDDRERGILCVVSNEDSDVRPA
16B PATSQEEETAASHEPEKKKEKE-KKEE-DEDDRNDDRERGILCVVSNEDSDVRPA
21B PATSQEEETAASHEPEKKKEKE-KKEE-DEDDRNDDRERGILCVVSNEDSDVRPA
4E PATSQEEETAASHEPEKKKEKEEKKEE-DEDDRNDDRERGILCVVSNEDSDVRPA
30E PATSQEEETAASHEPEKKKEKE-KKEE-DEDDRNDDRERGILCVVSNEDSDVRPA
**** ************* *** **** **********************
Figure 13 Overlapping putative casein kinase-2 (CKII) phosphorylation sites of UL84
CKII

85
3.4 Discussion
Several putative motifs were identified using protein scanning software and although
the core HCMV replication genes have already been characterized (Anders, 1988; Ertl,
1992; Pari, 1993; Smith, 1995a; Smith, 1996; Weiland, 1994), their significance in
replication was mainly predicted from similarities with the HSV counterparts (Anders,
1988; Pari, 1993). By combining the novel approach of motif scanning and phylogenetic
analysis between isolates and across species homologues, we have identified several
putative functional motifs in HCMV that may contribute to a complete understanding of
the CMV replication cycle.
One particular class of motifs has gained much attention in recent times. CKII is a
cellular serine-threonine kinase that has multiple target substrates involved in nucleic
acid replication, transcription, and protein synthesis among other biological processes
(Meggio, 2003). Active CKII is incorporated into the HCMV virion resulting in
immediate post-infection phosphorylation of cellular proteins that enhance HCMV
replication (Nogalski, 2007). There is also increasing evidence that CKII enhances
HCMV replication through phosphorylation of several HCMV proteins involved in
DNA replication and other processes (Alvisi, 2005; Barrasa, 2005; Jarvis, 2004). This
includes the UL44 DNA polymerase accessory protein that contains a (S/T)xx(D/E)
consensus CKII phosphorylation motif (also confirmed here) and is phosphorylated at
serine and/or threonine residues (Alvisi, 2005). In this study, we have identified
interesting and potential CKII phosphorylation motifs in the HCMV helicase (UL105)
and putative initiator protein (UL84), the latter recently shown to be a substrate of CKII
in pull-down assays (Gao, 2007). Although the previously described putative NLS
spanning residues 160-171 (Xu, 2002) has been disproved of its role in nuclear import
(Lischka, 2003), the position of the CKII phosphorylation site relative to this putative
NLS in UL84 (Xu, 2002) resemble a pattern shared by the CKII enhancing the NLS of
UL44 (Alvisi, 2005), which also binds with high affinity to the importin α/β
heterodimer (Alvisi, 2005). In addition, both these motifs fit the ‘CcN’ profile described
for phosphorylation regulated nuclear localization (Jans, 1996).
UL84 is an essential regulatory protein that is required for oriLyt-dependent DNA
replication (Lischka, 2003; Xu, 2004; Yu, 2003). Of the six replication genes studied, it

86
also had the lowest amino acid identity when compared against clinical isolates with a
score ranging from 97.46% to 100% identity. Initial characterization of UL84 identified
two potential leucine zippers between amino acids 114 to 135 and 325 to 373 (He,
1992), as well as a highly charged region starting at amino acid 161 (He, 1992).
Coincidentally, this region was initially associated with nuclear import (Xu, 2002),
although subsequent studies later mapped nuclear localization to a complex domain
downstream of this region (Lischka, 2003). In addition, two nuclear export signals (NES)
starting at amino acids 229 and 360 were also found to act as a non-conventional NLS
which allowed pUL84 to shuttle to the cytoplasm (Lischka, 2006). Recent studies have
shown that several regions of pUL84 share similar homology to the DExD/H box family
helicases (Colletti, 2005) while another bioinformatic study has shown homology to
dUTPases (Davison, 2005), both of which support the hypothesis that pUL84 acts as an
initiator protein for viral-DNA synthesis of HCMV (Xu, 2004a). PROSITE scanning for
potential motifs yielded three potential N-linked glycosylation sites while in earlier
studies, five were detected (He, 1992). However, on closer inspection one of the
original five N-linked glycosylation sites, beginning at amino acid 354, would have
been discounted as having functional significance on the basis of proline being present
within the consensus sequence (Bause, 1983; Gavel, 1990). BLAST searching revealed
only four sequence homologues, though when aligned showed a high degree of
conservation for both of the NES as well as a putative CKII phosphorylation site at
UL84 amino acid residue 474 identified in our studies. In addition, the putative CKII
phosphorylation site lies upstream of a motif suggested to be associated with helicase
function (Colletti, 2005) and may have a regulatory role. The putative leucine zipper
starting at amino acid 114 in HCMV was also detected in our scanning programs, while
the putative zipper starting at residue 325 was not, albeit neither were conserved
structures when aligned against the BLAST result homologues.
Among the ten HCMV isolates studied for UL44, variations in peptide sequence
occurred outside of regions currently known to have a functional role with the exception
of a K431R substitution in the previously identified NLS (Alvisi, 2005). Both the
HCMV UL44 and the HSV-1 homolog (UL42) share a “connector loop” structure that
plays a crucial role in binding to the DNA polymerase (Appleton, 2004; Bridges, 2001;
Loregian, 2004a). However, the sequence structure for the HCMV connector loop is
better conserved across the species, with more than 50% of the sequence alignments

87
having a similar structure of at least 57% sequence homology in that region. Although
the crucial functions of UL44 has previously been mapped to the N-terminal two-thirds
of the gene (Weiland, 1994), more than half of the potential motifs identified for UL44
in this study were found in the C terminal third of the gene, which has been determined
to be functionally dispensable for replication (Ertl, 1992). However, a recent study
elucidating the presence of a nuclear localization signal located at residues 425-431 of
the C-terminus (Alvisi, 2005) suggests that further study into this region needs to be
done, considering that the putative casein kinase-2 (CK2) site enhancing the NLS
(Alvisi, 2005) was also identified in our motif scans. Further analysis revealed an
unusual integrin binding motif in UL44 that was later shown to be conserved in pongine
Herpesvirus 4 as well. Integrins are known to mediate cell-cell association as well as
interactions between cells and extra-cellular proteins (Haynes, 2002). In addition, a
previous study on the murine CMV M44 homologue protein identified a putative
integrin binding RGD motif that was capable of mediating cell adhesion (Loh, 2000),
suggesting that UL44 could play a novel role in CMV infection.
Structure-function prediction for the UL57 single stranded DNA binding protein in
HCMV were largely based on studies of the HSV-1 homolog ICP8, which has so far
mapped the cooperative DNA binding region as well as the nuclear localization signal
to the C-terminus (Dudas, 1998; Gao, 1992; Mapelli, 2000), while an intranuclear
localization site was found to be located between residues 1080-1135 (Taylor, 2003). A
putative zinc binding motif between amino acids 499-512 in ICP8 (Gao, 1988; Gupte,
1991) was also found to be conserved in 16 of the 35 (46%) homolog sequences for the
single stranded DNA binding protein, albeit it was found to be non-essential for DNA
binding in HSV-1 (Wang, 1990). Although formal elucidation of its function in other
species of herpesvirus is yet to be validated, the structural nature of the motif is missing
key elements including a conserved phenylalanine and leucine involved in stabilizing
the motif (Jasanoff, 1993; Michael, 1992). However, the N terminal fragment of the
putative zinc finger beginning at amino acid 472 of UL57 showed a potential disulfide
bond that was conserved in 34 of the 35 (97%) homolog alignments. The conservation
of this putative disulfide bond motif across the species indicates an evolutionarily
conserved functional domain. Currently, the exact DNA binding region has yet to be
elucidated with numerous studies detailing a range of potential active sites (Gao, 1989;
Wang, 1990; White, 1999). The presence of a micro-bodies targeting signal observed in

88
the C-terminus of UL57, while yet to be functionally elucidated, suggests this protein
may be involved with the non-viral, non-lysosomal microbodies which have been
observed in CMV infected cells for decades (Craighead, 1972; Ruebner, 1966).
The primase-helicase complex of HCMV consisting of UL70, UL102, and UL105 is
considered to be homologous to the HSV-1 primase-helicase complex consisting of
UL52, UL8, and UL5 respectively (McMahon, 2002; Pari, 1993; Smith, 1996). All
three genes were highly conserved amongst the clinical isolates ranging from 98% to
100% sequence identity and 99 to 100% amino acid identity. Although the HCMV
primase (UL70) has not been formally characterized, it shares two functionally distinct
motifs that have been documented in its HSV-1 counterpart, UL52. The DxD catalytic
motif is essential for initiating DNA synthesis via the primer synthesis (Dracheva, 1995;
Klinedinst, 1994) while substitutions of the third and fourth conserved cysteines in the
zinc-finger at the C terminal end resulted in the loss of primase, helicase, ATPase, and
DNA-binding activities of the HSV helicase-primase complex (Biswas, 1999). The zinc
binding residues CHCC were found to be conserved in 26 of the 30 (87%) herpesvirus
homologues, while a key phenylalanine involved in structural stability (Michael, 1992)
was conserved in 22 of the 30 (73%) sequences. Interestingly, putative zinc fingers
lacking the conserved phenylalanine or an ‘aromatic swap’ (Kochoyan, 1991)
equivalent have been shown to be non-essential for key biological functions such as
DNA binding and replication (Hui, 2003; Wang, 1990). Analysis of the UL70
homologues show a highly conserved cluster of aromatic residues preceding a
conserved cysteine which has been associated with conferring resistance to a compound
which currently inhibits the HCMV primase (Chen, 2007). Aromatic interactions have
been attributed to involve up to 60% of the aromatic residues in a protein of which 80%
of these contribute to tertiary structure (Burley, 1985) and may provide some insight
into the structure-function relationship of that domain. Further analysis also revealed a
pair of cysteines conserved across all 30 homologues while a previously
uncharacterized domain spanning amino acids 631-646 contained several highly
conserved residues amongst the isolates and homologues in an otherwise poorly
conserved region (Figure 14B). Protein disulfide isomerase (PDI), with the canonical
CxxC motif, is the most efficient known catalyst of oxidative protein folding
(Weissman, 1993). Our phylogenetic analyses uncovered two highly conserved cysteine
residues forming a CxC motif in a previously uncharacterized domain of UL70 primase

89
and 30 primase homologues. The conservation of this CxC motif suggest that the CMV
primase may utilize a rare thiol-disulfide exchange mechanism (Woycechowsky, 2003)
that is present in only a handful of organisms including the Mengo encephalomyelitis
virus coat protein (Krishnaswamy, 1990), Bacillus Ak.1 protease (Smith, 1999) and the
E.Coli chaperone, Hsp33 (Jakob, 1999). The CxC motif is known to mediate disulfide
bond interactions between other conserved cysteines to form the well defined tertiary
structure for the chemokine class of proteins (Fernandez, 2002), and hence may have a
similar role in stabilizing the UL70 tertiary structure of HCMV here. In addition, an
earlier study of the HSV homologue demonstrated that substitutions of the arginine
residue seven amino acids downstream of the putative domain decreased the replication
efficiency of HSV by more than 50% (Klinedinst, 1994), suggesting that functionally
significant residues lie in close proximity to the CxC motif.
UL102 is a positional homolog of the HSV-1 UL8 primase associated factor (PAF) and
is shown to be involved in modulating primase-helicase activity (Barnard, 1997). Apart
from it’s initial characterization (Smith, 1995a), there have been few studies on UL102,
which, along with UL44 is the least well conserved of the core replication proteins in
HCMV, with BLAST searches yielding only 12 and 14 homologues respectively.
However, the functional domains of HSV-1 UL8 has been mapped to the regions
spanning amino acids 6–23 at the N-terminus, 718–750 at the C-terminus, and 78–339
internally where deletion mutants inhibit the ability of UL8 to support origin-dependent
DNA synthesis (Barnard, 1997). Several variations involving the distribution of glycine
residues were observed between residues 625-635 (Figure 14E) amongst the isolates
while the short serine-rich domain spanning residues 795-836 (Figure 14F) may have
implications for phosphorylation (Barrasa, 2005). Although UL102 was highly
conserved amongst the clinical isolates, alignments with the BLAST results showed no
distinguishable motifs conserved across the homologues. However, similar to UL44, an
unusual integrin binding motif was identified in the N-terminal of UL102 (Figure 14D),
although whether it plays a role in infection has yet to be seen.
UL105 is predicted to encode the HCMV helicase protein, based on sequence homology
to its HSV-1 UL5 counterpart which also contains six motifs typical of superfamily-1 of
helicase proteins (Gorbalenya, 1989; Graves-Woodward, 1997). However, of the six
motifs found to be essential for helicase activity in HSV-1 (Zhu, 1992), only two have

90
been functionally characterized. Motif I (also called the Walker A motif) follows the
consensus sequence GxxGxGKT/S and is believed to be involved in binding ATP
(Walker, 1982) while motif II (also called the Walker B motif) contains a series of
hydrophobic residues terminated by an aspartic and glutamic acid residue and is
believed to be involved in stabilizing Mg2+ (Walker, 1982). The functional significance
of Motifs III, IV, V, and VI is yet to be elucidated although a G815A substitution in
motif V of HSV-1 UL5 altered its ability to bind ATP, but otherwise left primase and
helicase activities intact (Graves-Woodward, 1996). In this study, a putative
cAMP/cGMP-dependent protein kinase phosphorylation site was detected in Motif IV
suggesting that this domain may interact with the cAMP signaling pathway as part of its
function. These six motifs were found to be highly conserved amongst the HCMV
clinical isolates. Comparisons with homologues revealed a high degree of conservation
for the six motifs amongst all 33 sequence homologues, emphasizing the strict structure-
function relationship of this class of proteins. In addition to the six helicase motifs, two
potential N-linked glycosylation sites as well as a potential casein-kinase 2
phosphorylation site was found to be highly conserved in the helicase homologues. All
three motifs were previously identified from the results of PROSITE scanning (Figure
14M/N/O). N-linked glycosylation sites follow a distinct Nx(T/S)x motif and have been
associated with aiding correct protein folding as well as cell-cell adhesion while casein
kinase 2 phosphorylation sites follow a (S/T)xx(D/E) consensus motif (Pinna, 1990) and
has already been shown to be involved with several biological processes of HCMV
replication (Alvisi, 2005; Barrasa, 2005; Jarvis, 2004).

91
Figure 14 Isolate variation in the HCMV primase-helicase complex.
Green lines represent amino acid substitutions amongst isolates while pink segments represent known
functional domains and tan segments represent interesting regions identified by phylogenetic analysis: A)
Catalytic domain (Dracheva, 1995), B) Novel conserved domain, C) Putative zinc finger motif (Biswas,
1999) D) Putative integrin binding motif, E) Glycine-rich domain, F) Serine-rich domain, G-L) Helicase
motifs I-VI (Zhu, 1992), M-N) Putative N-linked glycosylation sites, O) Putative CKII phosphorylation
sit
The functional domains of a protein are often found to be highly conserved amongst
homologues (Dracheva, 1995; Gorbalenya, 1988), with the implication that these
domains remained unchanged over time and during species differentiation due to the
crucial role they play in propagating the organism. In this study we have identified
nearly 300 potential motifs spanning five core and one non-core replication gene of
HCMV, many of which are ubiquitous commonly-occurring sequences. Motif numbers
can be overestimated by ScanProsite in glycine rich regions, where a string of glycines
can give rise to the same motif repeated several times. The 27 N-myristoylation sites
identified for UL44, for example, may be an overestimation, and caution should be
applied when using ScanProsite to determine a definitive number of particular motifs
within a given gene. However, as we have demonstrated, motif scanning programs

92
combined with phylogenetic analysis are useful tools for the discovery of previously
unidentified motifs likely to be important for overall protein activity. This provides
focus areas for future studies of these important HCMV replication proteins including
investigations of potential antiviral inhibitors.

93
4 HCMV primase expression
4.1 Introduction
Despite the efficacy of HCMV anti-viral therapy in treating HCMV-related disease
(Goldberg, 2003; Salzberger, 2005), long-term administration of ganciclovir (GCV) or
foscarnet (FOS) can lead to severe toxicity or the emergence of drug-resistant virus
strains (Baldanti, 1995). Therefore, there is a need to develop new therapeutic drugs
that combine efficacy and safety with a novel mechanism of action, thus excluding the
possibility of cross-resistance with existing therapeutics (Biron, 2006; Wathen, 2002).
Among the new anti-viral therapies currently being developed, Maribivir targets the
CMV protein kinase gene (Evers, 2004) and has been shown to be effective against
strains resistant to GCV, FOS, and Cidofovir (CDV) (Drew 2006). However, resistant
strains have already begun to emerge, with mutations conferring resistance identified in
the UL27 and UL97 genes (Chou, 2007; Komazin, 2003).
The HCMV primase, encoded by the UL70 ORF, shares 27% homology with the HSV-
1 primase, UL52 (Chee, 1990) and is an essential component of the viral DNA
replication apparatus (Pari, 1993). All known DNA polymerases are unable to initiate
synthesis of a new DNA strand de novo, while almost all require the activity of a DNA
primase to prime the template (Arezi, 2000). However, unlike eukaryotic primases,
which initiate primer synthesis at preferred but nonspecific template sequences,
prokaryotic and viral primases usually have specific template sequence requirements for
primer initiation (Frick, 2001). The HCMV primase is part of a trimeric complex which
also includes the helicase (UL105) and the primase-associated protein (UL102)
(McMahon, 2002). The complex is thought to unwind and prime the DNA template for
replication, which has been shown to be the case for homologous proteins of the HSV-1
(Graves-Woodward, 1997; Ramirez-Aguilar, 2004). Currently, high-throughput
screening has identified a series imidazolyl-pyrimidine compounds that can inhibit the
primase (Cushing, 2006) Although the mechanism of action of these compounds are
largely unknown, resistance conferred by a P570S point mutation in UL70 suggests that
the primase is inactivated via irreversible binding of the primase with the compound
(Chen, 2007). The importance of the primase-helicase complex in CMV DNA
replication presents as a logical target for future anti-viral therapeutics. Compounds

94
which can target either the primase activity or its ability to associate with complement
members of the essential complex can provide a novel mechanism of action that is
unique from existing treatments. In addition, they may be used in conjunction with
existing anti-viral therapies to provide effective treatment and reduce the incidence of
specific resistance mutants that often emerge with long term single course treatments.
By elucidating the functional aspects of the primase, we can increase our understanding
of the complex CMV replication cycle and provide the groundwork for developing new
anti-viral strategies in the future.
Due to the large genome size and prolonged replication cycle of CMV, generating
recombinant HCMV has been difficult and time-consuming (Wang, 2004), while
previous efforts to study the HCMV helicase–primase proteins in particular have been
hampered by their low abundance in HCMV-infected cells (McMahon, 2002). However,
the advent of Bacterial Artificial Chromosome (BAC) technology solves these problems
as well as providing a robust genetic system for constructing viral mutants (Jarvis 2007).
BAC technology has already been used in several studies ranging from systematic
profiling of the CMV genome (Dunn, 2003; Yu, 2003) to elucidating mutations
conferring anti-viral resistance (Chou, 2007a; Martin, 2006). We proposed using the
BAC system for the purpose of creating recombinant mutants to study the importance of
the HCMV primase for viral replication. Furthermore, we pursued the expression of
wild-type and mutant UL70 from baculovirus constructs in insect cells, as well as in
vitro expression using a streamlined TNT Quick Coupled Transcription/Translation Kit
(Promega, USA), with a view towards functional and protein-protein interaction
analyses.

95
4.2 Methods
4.2.1 Cloning of UL70 into high-copy pBluescript II primase vector
4.2.1.1 Viral stocks and DNA extraction
Viral DNA from high-passage HCMV isolates 29A and 5B were extracted from frozen
cell-associated viral stock using the QIAamp DNA Mini Kit (Qiagen, USA) as per
manufacturer’s instructions. Briefly, specimens were incubated in lysis buffer and
Proteinase K to release cellular DNA. Ethanol was used to precipitate the DNA which
was subsequently captured in microfuge spin columns (Qiagen, USA). The DNA was
then washed and eluted in 50µl of buffer AE after 5 min incubation instead of the
recommended 200µl to increase DNA concentration.
4.2.1.2 Polymerase chain reaction and purification of UL70 for pBluescript
The full length ORF encoding the HCMV primase (UL70) was amplified from the DNA
of high-passage isolates using primers containing the SpeI and HindIII restriction sites
in the 5’ and 3’ ends respectively (Table 10). Two sets of primers were created such that
UL70.SpeI and UL70.HindIII produced a ~3kb fragment containing only the UL70
ORF (UL70a) while UL70.3SpeI and UL70.4HindIII produced a ~5kb fragment that
contained the UL70 ORF as well as approximately 1kb genomic material flanking the 5’
and 3’ ends (UL70b) (Table 10). A single round PCR was carried out in a 50µl reaction
consisting of 1x buffer (supplemented with 1.75mM MgCl2), 400µM dNTPs, 300nM
primers, 100µg/ml bovine serum albumin (BSA) (Promega, US), 3.75U ExpandLong
template enzyme (Roche) and 5µl DNA template. The single round cycling conditions
consisted of denaturation at 94°C for 3 min; 10 cycles of 94°C for 10 sec, 65°C for 30
sec, 68°C for 4 min; 20 cycles of 94°C for 15 sec, 65°C for 30 sec, 68°C for 4 min
(increasing by 20 sec for each subsequent cycle); and a final elongation at 68°C for 7
min. PCR products were visualized following electrophoresis on 0.75% agarose gels
and purified using polyethylene glycol (PEG). Briefly, this involved mixing the PCR
product with 2x PEG (26.7% PEG 8000, 0.6 M NaOAc (pH 5.2), 6.5 mM MgCl2) in a
1:1 ratio. The mixture is incubated at room temperature for 10 min and centrifuged at

96
8000xg, with the supernatant removed afterwards. The DNA pellet was then washed
with 96% ethanol, air-dried and re-suspended in 20µl distilled water (Baxter, Australia).
Table 10 Nucleotide primers used for UL70/pBluescript cloning
Primer Name Primer sequence*
UL70.SpeI 5' - GCACTAGTTGACGTCGGTCCGAAACCTCC
UL70.HindIII 5' - CGCAAGCTTATCGTGCGGGTTGACGGGTAG
UL70.3SpeI 5' - TAGAGGACTAGTCAGCACGTAATCGGCCACAGG
UL70.4HindIII 5' - CGATGAAAGCTTTATAGATGAGACCGCTGCCGG
UL70.4 5' – CCCAACTGGATCTGCGTAACCTGCT
T3 5' - AATTAACCCTCACTAAAGGG
T7 5' - GTAATACGACTCACTATAGGGC
*Underlined sequences denote restriction sites
4.2.1.3 Molecular cloning of UL70 into pBluescript II
Both the 3kb and 5kb UL70 PCR products as well as the pBluescript II vector were
double digested with SpeI and HindIII restriction enzymes in separate 20µl reaction
consisting of 1x buffer B (Promega, US), 2 µg/µl acetylated BSA (Promega, US), 5U
SpeI (Promega, US), 5U HindIII (Promega, US), and ~1µg DNA, or a 5U/µg over-
digest. Restriction digest reactions were incubated at 37°C for 1.5hrs followed by heat
inactivation at 74°C for 15 min. Ligation was performed immediately after in a 10µl
reaction consisting of 1x ligase buffer (Stratagene, USA), 1mM rATP (Stratagene,
USA), 2U T4 DNA ligase (Stratagene, USA), and DNA in an insert to vector ratio of
3:1 as calculated by the following formula:
Figure 15 Insert to vector ratio formula
A reaction consisting of pUC18 plasmid in place of the sample was used as a ligation
control. The ligation reactions were incubated at room temperature for 1 hr followed by
transformation to XL10-Gold Ultracompetent Cells (Stratagene, USA) as per

97
manufacturer’s protocol. Briefly, this involved thawing a 100µl aliquot of cells on ice
before adding 4µl β-Mercaptoethanol and 2µl ligation mixture. The reaction was heat
pulsed at 42°C for 30 sec followed by the addition of 900µl NZY broth. The cells were
grown in a shaking incubator at 37°C and 225rpm for 1 hr prior to plating on LB-
Ampicillin agar plates supplemented with 100µl 1M IPTG and 20µl X-Gal and
incubated at 37°C for 16hrs. Plates were then placed in a 4°C fridge for 1 hr to enhance
the blue/white screening process.
Single white colonies were re-streaked and screened via colony PCR using an internal
UL70 primer and a pBluescript T7 primer (Table 10). A single round colony PCR was
carried out in a 50µl reaction consisting of 1x GoTaq Flexi buffer (Promega), 4mM
MgCl2, 200µM dNTPs, 0.2µM primers, 1.25U GoTaq DNA polymerase (Promega), and
5µl DNA template. The cycling conditions consisted of an initial denaturation at 95°C
for 2 min followed by 30 cycles at 95°C for 1 min, 65°C for 1 min, 72°C for 2 min and
a final elongation at 72°C for 5 min. PCR products were visualized following
electrophoresis on 1.5% agarose gels. PCR positive colonies were grown overnight in
2ml NZY broth supplemented with 100µg/ml Ampicillin at 37°C with 225rpm shaking.
Plasmid DNA was extracted using a Wizard Plus Minipreps DNA Purification System
(Promega, USA) as per manufacturer’s instruction and sequenced to check for genomic
integrity of the vector/insert construct. Sequencing reactions consisted of Big Dye
Terminator (Applied Biosystems, USA), premix buffer, 0.25µM primer (Table 7 and
Table 10), and approximately 75ng/300bp of DNA. Cycling conditions consisted of 25
cycles of 96°C for 10 sec, 50°C for 5 sec, and 60°C for 4 min. Sequencing products
were purified and analyzed at the University of New South Wales DNA analysis facility
(Figure 16).

98
Figure 16 Overview of cloning UL70 into pBluescript

99
4.2.2 Construction of baculovirus entry clone for HCMV primase expression
4.2.2.1 Viral stocks and DNA extraction
Plasmids containing the UL70 ORF were extracted from the UL70a pBluescript II stock
using the Wizard Plus Midipreps DNA Purification System (Promega, USA) as per
manufacturer’s instruction. Briefly, plasmids were inoculated and grown overnight in
5ml NZY broth supplemented with 10mg/ml Ampicillin at 37°C with 225rpm shaking.
The cells are centrifuged at 10,000xg for 10 min and the supernatant is discarded. Cell
re-suspension, lysis, and neutralization solutions are added to lyse the cells and
precipitate the protein from solution. The lysate was centrifuged at 14,000xg for 15 min
and the supernatant is decanted to a new microfuge tube for purification. The DNA is
transferred to a Wizard midi-column (Promega, USA) and purified under vacuum with
diatomaceous resin and washed with ethanol before a final elution with 100µl hot (65°C
-70°C) distilled water (Baxter, Australia).
4.2.2.2 Polymerase chain reaction and purification of UL70 for pDONR
The full length UL70 ORF was amplified from midipreps of the UL70a pBluescript
stock with primers which incorporated attachment sites into the product (Table 11). A
single round PCR to amplify the ~3kb product was carried out in a 50µl reaction
consisting of 1x High Fidelity buffer (Invitrogen, USA), 0.2mM dNTPs, 2mM MgSO4,
400nM primers, 1U Platinum Taq High Fidelity enzyme (Invitrogen, USA) and 5µl
DNA template. The single round cycling conditions consisted of denaturation at 94°C
for 3 min followed by 35 cycles of 94°C for 30 sec, 55°C for 30 sec, and 68°C for 3 min.
PCR products were visualized following electrophoresis on 1.0% agarose gels and
purified using a 30% PEG 8000/30mM MgCl2 Solution (Invitrogen, USA). Briefly, this
involved diluting the PCR product 4-fold with TE buffer, followed by adding 0.5 times
volume of 30% PEG 8000/30mM MgCl2 solution. The mixture is centrifuged for 15
min at 8000xg, with the supernatant removed afterwards and the clear pellet suspended
in 20µl TE.

100
Table 11 Nucleotides primers used for UL70/pDONR cloning
Primer Name Primer Sequence*
UL70attb1 5' - GGGGACAAGTTTGTACAAAAAAGCAGGCTTCCTGCATCGTGCCGGCGCGACG
UL70attb2 5' - CCCCTGGTGAAACATGTTCTTTCGACCCAGCGCCGTGGAAAGTGAGGCTAG
UL70.3 5' - GCGAGCCCAGTAGCAGACGCGCGAA
UL70.4 5' - CCCAACTGGATCTGCGTAACCTGCT
M13 Forward 5' - GTAAAACGACGGCCAG
M13 Reverse 5' - CAGGAAACAGCTATGAC
* Underlined sequences denote attachment sites
4.2.2.3 pDONR recombination and molecular cloning of UL70
The UL70 PCR product was recombined with the pDONR donor vector (Invitrogen,
USA) (Figure 17) using a Gateway BP Clonase II enzyme (Invitrogen, USA).
Figure 17 pDONR/Zeocin Vector
pDONR/Zeocin vector with key features including: pUC Ori: pUC Origin; Zeocin: Zeocin resistance gene;
T1/T2: Transcription termination sequence; attp1/2: Recombination sites; ccdB: ccdB gene (negative

101
selection of plasmid); CmR: Chloramphenicol resistance gene; M13 Forward/Reverse: Sequencing
priming sites.
A 10µl BP reaction consisting of 20-50fmol PCR product (Figure 18), 150ng pDONR
vector, TE Buffer, and 2µl BP Clonase II enzyme mix (Invitrogen, USA) was incubated
overnight at 25°C, followed by the addition of 1µl Proteinase K (Invitrogen, USA) and a
further incubation of 37°C for 10 min. A pEXP7-tet positive control was used in
conjunction with each experiment. A 1-2µl sample of the BP reaction was then
immediately transformed to Library Efficient DH5α competent cells (Invitrogen, USA).
Briefly, this involved thawing a 50µl aliquot of cells on ice before adding the sample
and heat shocking the reaction at 42°C for 30 sec followed by the addition of 250µl
SOC media pre-warmed to 37°C. The cells were grown in a shaking incubator at 37°C
and 225rpm for 1 hr prior to plating on LB-Zeocin agar plates while positive controls
were plated on LB-tetracycline plates. All plates were incubated overnight at 37°C.
Figure 18 converting femtomoles to nanograms
Single colonies were re-streaked and screened via colony PCR using an internal UL70
primer and a pDONR M13 primer (Table 11). A single round colony PCR as outlined in
section 4.2.1.3 was carried out to check for the insert. PCR positive colonies were
grown overnight in 5ml SOC media supplemented with 50µg/ml Zeocin at 37°C with
225rpm shaking. Plasmid DNA was extracted using a Wizard Plus Midipreps DNA
Purification System (Promega, USA) as per manufacturer’s instruction and sequenced
to check for genomic integrity of the construct. Sequencing reactions and cycling
conditions were previously outlined in section 4.2.1.3. A brief overview of the process
is outlined in Figure 19.

102
Figure 19 Overview of UL70/pDONR recombination
4.2.3 In vitro UL70 primase expression using TNT Quick Coupled
Transcription/Translation
4.2.3.1 Viral stocks and DNA extraction
Plasmids containing the UL70 ORF were extracted from the UL70a pBluescript II stock
using the Wizard Plus Midipreps DNA Purification System (Promega, USA) as per
manufacturer’s instruction and as outlined in previous sections.

103
4.2.3.2 Polymerase chain reaction and purification of UL70
The full length UL70 ORF was amplified from midipreps of the UL70a pBluescript
stock using primers which incorporated the BamHI and SalI restriction sites at the 5’
and 3’ ends of UL70 respectively (Table 12). A single round PCR to amplify the ~2.9kb
product was carried out in a 50µl reaction consisting of 1x High Fidelity buffer
(Invitrogen, USA), 0.2mM dNTPs, 2mM MgSO4, 400nM primers, 1U Platinum Taq
High Fidelity enzyme (Invitrogen, USA) and 5µl DNA template. The single round
cycling conditions consisted of denaturation at 94°C for 3 min followed by 35 cycles of
94°C for 30 sec, 60°C for 30 sec, and 68°C for 3 min. PCR products were visualized
following electrophoresis on 1.0% agarose gels and purified using a Wizard SV Gel and
PCR Clean-Up Kit (Promega, USA) as per manufacturer’s protocols. Briefly, the PCR
products were mixed in a 1:1 ratio with Membrane Binding Solution (Promega, USA),
incubated at room temperature and transferred to a SV mini-column (Promega, USA)
where it is centrifuged at 16,000xg to collect the DNA in the membrane. The membrane
is washed several times with Membrane Wash Solution (Promega, USA) before a final
elution of 30µl with nuclease-free water (Promega, USA).
Table 12 Nucleotide primers used for UL70/pCITE cloning
Primer Name Primer Sequence*
UL70pCITE.3 5' - ATTGGATCCCTGCATCGTGCCGGCGCGACG
UL70pCITE.4 5' - ATTGTCGACTGACGGCGGTCGCCGGCGGCAT
UL70.3 5' – GCGAGCCCAGTAGCAGACGCGCGAA
T7 5' - GTAATACGACTCACTATAGGGC
* Underlined sequences denote restriction sites
4.2.3.3 Molecular cloning of UL70 into pCITE 4A
Both the UL70 PCR product and pCITE vector were sequentially digested, initially with
SalI followed by BamHI. Restriction digests with SalI were carried out in a 20µl
reaction consisting of 1x NEBuffer 3 (New England BioLabs (NEB), USA), 100µg/ml
BSA (NEB, USA), 30U SalI (NEB, USA), and ~3µg DNA, or a 10U/µg DNA over-
digest. Digest reactions were incubated at 37°C for 3 hrs followed by heat inactivation
at 65°C for 20 min. Digest products were then purified using the Wizard SV Gel and

104
PCR Clean-Up Kit (Promega, USA) as per manufacturer’s protocols outlined in the
previous section prior to the second digest. Restriction digests with BamHI were also
carried out in 20µl reaction consisting of 1x NEBuffer 3 (NEB, USA), 100µg/ml BSA
(NEB, USA), 20U BamHI (NEB, USA), and ~2µg DNA, or a 10U/µg DNA over-digest.
Digest reactions were incubated at 37°C for 2 hrs and immediately followed by
processing with the Wizard SV Gel and PCR Clean-Up Kit (Promega, USA) as a means
to stop the digest reaction and purify the restriction products. Products were ligated in a
20µl reaction consisting of 1x ligase duffer (Stratagene, USA), 1mM rATP (Stratagene,
USA), 4U T4 DNA ligase (Stratagene, USA), and DNA in an insert to vector ratio of
3:1 as calculated by the formula in section 4.2.1.3. A reaction consisting of pUC18
plasmid in place of the sample was used as a ligation control. The ligation reactions
were incubated at room temperature for 16 hrs followed by transformation to Library
Efficient DH5α Competent Cells (Invitrogen, USA) as per manufacturer’s protocol,
albeit omitting the dilution steps. Briefly, this involved thawing a 100µl aliquot of cells
on wet ice before adding 1-10ng DNA and incubated on ice for 30 min. The mixture
was heat shocked at 42°C for 45 sec followed by the addition of 900µl SOC media. The
cells were grown in a shaking incubator at 37°C and 225rpm for 2 hrs prior to plating on
LB-Ampicillin agar plates.
Single colonies were re-streaked and screened via colony PCR using an internal UL70
primer and a pCITE T7 primer (Table 12). A single round colony PCR as outlined in
section 4.2.1.3 was carried out to check for the insert. PCR positive colonies were
grown overnight in 5ml SOC media supplemented with 100µg/ml Ampicillin at 37°C
with 225rpm shaking. Plasmid DNA was extracted using a Wizard Plus Midipreps
DNA Purification System (Promega, USA) as per manufacturer’s instruction and
sequenced to check for genomic integrity of the construct. The sequencing reaction and
cycling conditions used were previously outlined in section 4.2.1.3. An overview of the
process is outlined in Figure 20.

105
Figure 20 Overview of cloning UL70 into pCITE expression vector

106
4.2.3.4 Protein expression and detection
UL70 expression was carried out using a TNT Quick Coupled Transcription/Translation
Kit (Promega, USA) designed to express protein in an in vitro environment. Protein
expression was carried out in 50µl reactions consisting of 40µl TNT Quick Master-Mix
(Promega, USA), 20µM methionine (Promega, USA), and ~1ng DNA. The reaction was
incubated at 30°C for 90 min, followed by a 1:4 dilution with 4X loading dye and
finally denatured at 100°C for 5 min. A T7 luciferase which produced a 61kDa
monomeric protein was used as a translation control. Translation products were
visualized with a 10% SDS-PAGE resolving gel using a Mini-PROTEAN 3 Cell
electrophoresis system (Biorad, USA). The protein samples were loaded alongside 10µl
Kaleidoscope markers (Biorad, USA) and Magic Mark protein markers (Novagen,
Germany). All samples were run through the 5% stacking gel at a constant voltage of
80V for 30 min prior to running the samples through the resolving gel at a constant
voltage of 110V until the dye front left the gel. Protein bands were transferred from the
gel to nitrocellulose membrane using Western blotting techniques. The transfer was run
overnight at a constant current of 90mA (30V) using a magnetic stirrer and ice-pack to
keep the system from overheating. The gel was stained with 20ml Gelcode Blue Stain
(Pierce, USA) post-transfer to check for total protein. The S-Tag and His Tag antibodies
were used to detect for the UL70 protein product. Briefly, the nitrocellulose membrane
was incubated with gentle shaking in blocking solution for 1 hr, followed by thorough
washing with PBST solution. For membranes being detected with the S-Tag, the S-
Protein Alkaline Phosphatase Conjugate (Novagen, Germany) was diluted 1:5000 with
PBST and incubated with the membrane at room temperature for 30 min with gentle
shaking. Detection with the S-Tag did not require a secondary antibody. For membranes
being detected with the His-Tag, c-terminal Anti-His antibodies (Invitrogen, USA) were
diluted 1:5000 with PBST and incubated with the membrane at room temperature for 1-
2 hrs with gentle shaking. The membrane was washed thoroughly with PBST prior to
incubation with the secondary antibody. Immun-Star goat anti-mouse IgG-HRP
(Australian Laboratory Services, Australia) was diluted 1:4000 with blocking solution
and incubated with the membrane at room temperature with gentle shaking for 1 hr. The
membranes are thoroughly washed again with PBST after antibody incubation and
prepared for chemiluminescence visualization with SuperSignal West Pico
Chemiluminescent Substrate (Pierce, USA), as per manufacturer’s instructions. Briefly,
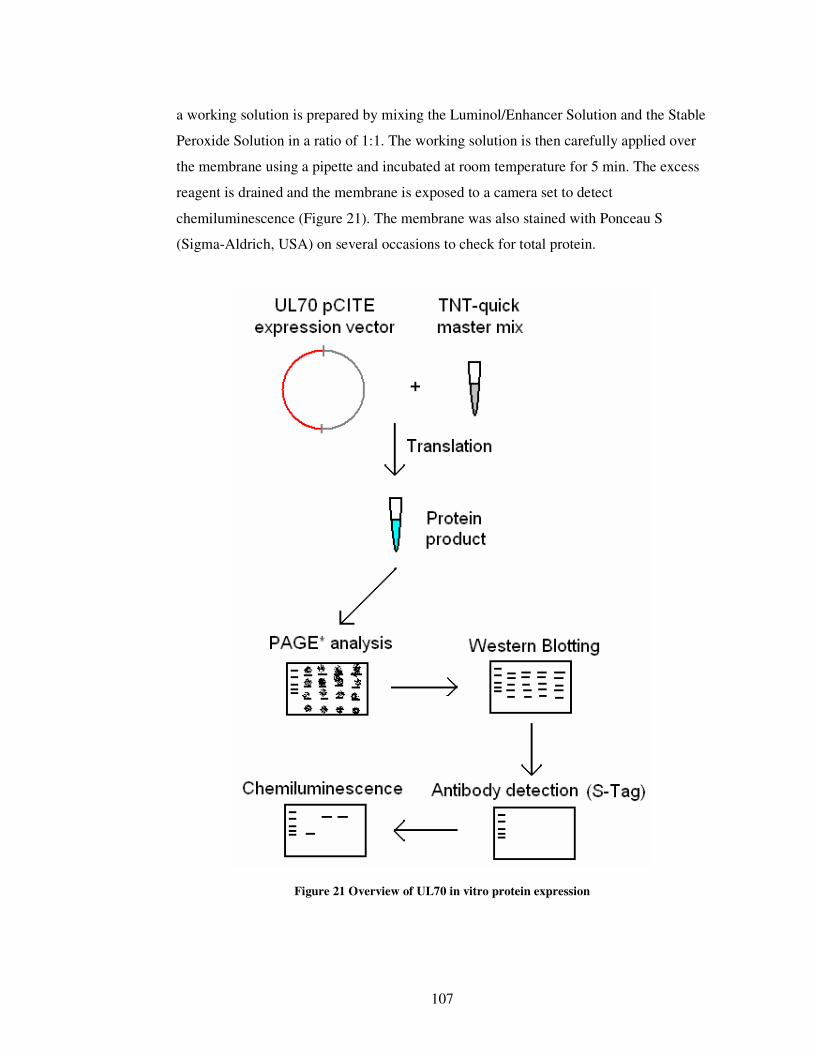
107
a working solution is prepared by mixing the Luminol/Enhancer Solution and the Stable
Peroxide Solution in a ratio of 1:1. The working solution is then carefully applied over
the membrane using a pipette and incubated at room temperature for 5 min. The excess
reagent is drained and the membrane is exposed to a camera set to detect
chemiluminescence (Figure 21). The membrane was also stained with Ponceau S
(Sigma-Aldrich, USA) on several occasions to check for total protein.
Figure 21 Overview of UL70 in vitro protein expression

108
4.3 Results
4.3.1 UL70 was cloned into pBluescript II high-copy vector
Both the UL70 ORF spanning 2,985bp from high-passage isolate 5B and the 5,250bp
fragment from high-passage isolate 29A were successfully cloned into pBluescript and
designated as UL70a and UL70b respectively. UL70a consisting of the 3kb pBluescript
vector and the 3kb UL70 insert (Figure 22A) produces a 6kb band when extracted from
a plasmid preparation. In addition, we also detected an extra band several kilo-bases
heavier than our anticipated product indicating that nicked DNA may also be present
(Figure 22B). However, sequencing of the plasmid confirmed the insert ligated correctly
into the vector without mutations.
A B
Figure 22 pBluescript UL70a
(A) UL70a plasmid map indicating position of UL70 (red) relative to key vector domains (orange). (B)
UL70a produces two products bands, the anticipated plasmid (II) and nicked DNA (I). Lambda DNA
HIndIII markers (Promega) on the left are indicated in base-pairs.
UL70b consists of the 3kb pBluescript vector and the 3kb UL70 ORF as well as
approximately 1kb of flanking genomic region either end of the UL70 ORF (Figure
23A). Plasmid preparations produces an anticipated band of approximately 8kb as well

109
as a high molecular weight product predicted to be nicked DNA (Figure 23B).
Sequencing of the plasmid confirmed that the 5kb insert ligated into pBluescript and
without mutations.
A B
Figure 23 pBluescript UL70b
(A) UL70b plasmid map indicating position of UL70 (red) relative to key vector domains (orange). (B)
UL70b produces two products bands, the anticipated plasmid (II) and nicked DNA (I). Lambda DNA
HIndIII markers (Promega) Markers on the right are indicated in base-pairs.

110
4.3.2 Generating Baculovirus entry clones via BP recombination
Experiments to create a UL70/pDONR recombinant have been unsuccessful with
several instances of the PCR incorporated attachment site failing to bind to their correct
complement on the pDONR vector, resulting in recombinants which truncated the UL70
ORF. As a result, subsequent experiments to recombine the pDONR/UL70 entry clone
with the baculovirus expression DNA have not been pursued. The length of the excised
regions varied, with designate samples BPR4, BPR5, and BPR8 missing 1,919bp, 36bp,
and 505bp respectively, all from the 3’ end of UL70. However, the 5’ end of UL70
recombined with pDONR without problems. Other attempts to generate an entry clone
failed to produce colonies at the screening stage despite the transformation controls
producing visible colonies. Experiment BPR1 and BPR2 both used BP Clonase I
(Invitrogen, USA) in the BP recombination reaction (c.f. BP Clonase II in later
experiments), with the former transformed into One Shot TOP10 competent cells
(Invitrogen, USA) while latter experiments used the DH5α competent cells. BPR3 was
carried out as a time-course experiment to determine the optimal BP reaction time, and
though colonies were generated at the longest incubation, all samples were found
negative for UL70. Samples BPR4 and BPR5 were generated using increasing amounts
of total DNA during the BP reaction, with BPR5 grown for an additional hour during
the transformation step to increase the number of cells prior to plating. BPR4 was
selected from several colonies of which only one produced a band at the expected size
of 780bp when screened using M13 forward and UL70.3 reverse primers (Figure 24A).
However, plasmid preparations of the construct revealed a prominent band at 2.3kb, the
expected band of 4.9kb, as well as a band exceeding 9kbp predicted to be nicked DNA
(Figure 24B).

111
A B
Figure 24 Screening of entry clone BPR4
(A) PCR screening of BPR4 generated the expected 780bp fragment. (B) Plasmid preparation of BPR4
revealed 3 products: a band >9kbp predicted to be nicked DNA (I), band at ~6kbp predicted to be the
expected entry clone, and an unknown prominent band at 2.3kbp (III). Lambda DNA HIndIII Markers
(Promega) (right) and BenchTop pGEM DNA markers (Promega) (left) sizes are indicated in base-pairs.
Sequence analysis of the BPR4 plasmid preparation revealed a truncated 3kb product
with UL70 missing the C-terminal 1,919bp, albeit retaining frame with the rest of
pDONR. The initial PCR screening was a false positive as the region amplified did not
involve the truncation. In addition, the truncation removes the catalytic motif and the
putative zinc finger binding domain (Figure 25).
Figure 25 The BPR4 truncation removes key motifs from UL70
Key UL70 motifs highlighted in purple while red box denotes section of gene truncated during
recombination. Orientation and length of te UL70 protein is denoted by numbers.

112
Despite the unfavorable recombination in BPR4, another attempt was carried out on the
assumption that increasing the overall number of cells prior to plating would increase all
populations universally, including the constructs which contained our desired product.
In addition, PCR screening of the colonies involved the use of both the M13
forward/UL70.3 reverse primers as well as a set of internal UL70 primers such that
colonies containing the truncation in BPR4 would be selected out. BPR5 was selected
on the basis of colony PCR (Figure 26).
Figure 26 Colony PCR screening for entry clone BPR5
Internal UL70 primers were used to amplify a 1.2kbp region as well as intrinsically select out
recombinants carrying the BPR4 truncation. BenchTop pGEM DNA marker (Promega) sizes on the right
are indicated in base-pairs.
However, after sequencing several BPR5 samples, a design flaw was identified at the 3’
end of the UL70 insert product, which caused a short truncation and amplification of
non-specific DNA (Figure 27). An inverted attachment sequence was identified for the
3’ primer, resulting in complications during the amplification as well as the subsequent
BP recombination.

113
UL70 junk Primer attb1 pDONR
-----------------------|----------|------------------------|------|---------------------
Figure 27 The BPR5 truncation of UL70
A primer design flaw caused the 36bp truncation of UL70 as well as the amplification of non-specific
sequences (green). Key UL70 motifs highlighted in purple while red box denotes section of gene
truncated during recombination. Orientation and length of the UL70 protein is denoted by numbers.
Subsequent experiments, including BPR6, BPR7 and BPR8 were carried out using a
modified primer with the correct attachment sequence. BPR6 and BPR7 failed to
generate colonies despite using the previously optimized protocol while BPR8,
transformed in a new batch of DH5α cells produced several colonies. PCR positive
colonies were sequenced, revealing another novel truncation at the 3’ end of UL70. A
505bp fragment at the c-terminal end of UL70 was shown to be excised, removing a
putative zinc finger domain in the process (Figure 28).
Figure 28 The BPR8 truncation removes the putative zinc finger from UL70
Key UL70 motifs highlighted in purple while red box denotes section of gene truncated during
recombination. Orientation and length of the UL70 protein is denoted by numbers.

114
4.3.3 UL70 was cloned into pCITE expression vector
The full length UL70 ORF was successfully cloned into expression vector pCITE 4A
with a functional S-Tag and c-terminal His-tag (Figure 29A). The original UL70 stop
codon, TGA, was replaced with TCA to ensure that subsequent translation incorporated
the down-stream His-tag (Figure 29A). Plasmid preparations of the DNA gave a distinct
band at the expected size of 6.7kbp (Figure 29B) while sequencing confirmed that the
3kbp insert ligated into pCITE without mutations.
A B
Figure 29 UL70/pCITE expression vector
(A) UL70/pCITE plasmid map indicating position of UL70 (red) relative to key vector domains (orange).
(B) UL70/pCITE produces one distinct band at 6.7kbp. Lambda DNA HIndIII markers (Promega) on the
right are indicated in base-pairs.

115
4.3.4 UL70 in vitro expression using the TNT system
The expression vector created in 4.3.3 was used with the TNT Quick Coupled
Transcription/Translation Kit (Promega, USA) to express the HCMV primase. However,
attempts at protein expression failed to produce a protein of the predicted 107kDa size.
In addition, an unknown product of ~55kDa was detected in all lanes (Figure 30). The
method of detection was by using the S Tag protein, which is present in the pCITE
expression vector, but not in the luciferase control DNA, hence the control reaction was
not carried out.
Figure 30 Expression of UL70 in TNT system
Typical protein expression produces an unknown product of ~55kDa (I) in both the UL70 sample (A) and
the negative control (B). Precision Plus Kaleidoscope markers (Biorad) on the left are given in Kilo-
Daltons (kDa).
Since previous attempts were based on a single incubation time of 90 min as
recommended per manufacturer’s instructions, we carried out a time course experiment
to determine the incubation time that would yield the optimal amount of protein.

116
However, samples taken at 30 min intervals up to duration of 3 hrs yielded similar
results to incubations at 90 min (Figure 31).
Figure 31 Time course expression of UL70 with TNT system
Samples were taken at 30 min intervals starting at 1hr (A) to 3 hrs (E). The negative control was taken at
3 hrs (F). Markers on the left are given in kDa.

117
4.4 Discussion
The ability to replicate the UL70 ORF at high yields using a high copy vector as
opposed to traditional cell culturing is an invaluable tool for elucidating the functions of
the primase in future expression studies. UL70b, which contains flanking regions either
side of the 5’ and 3’ ends of the UL70 ORF, was designed as a precursor to express the
UL70 gene in a Bacterial Artificial Chromosome (BAC) system. The BAC system has
been shown to be an effective tool for studying the importance of various genes in
HCMV (AuCoin, 2006; Hahn, 2002; Hahn, 2004; Jiang, 2008; Komazin, 2004; Lorz,
2006; Martin, 2006; Michel, 2005; Stropes, 2008; Xu, 2004) and the creation of a UL70
precursor may serve as a contribution to future studies on the HCMV helicase-primase
complex, which has yet to be studied in detail.
Protein expression involving baculovirus requires the creation of an expression virus
through recombination between an entry clone and the baculovirus linear DNA. The
entry clone is created through another recombination reaction involving an entry vector
and a PCR generated DNA fragment utilizing attachment sites. In this study, attempts to
generate entry clones for this process resulted in recombinants that truncated UL70 and
although initial results reflected the consequences of a faulty primer, later experiments
persisted with the truncation of the 3’ end. The original 3’ primer sequence was found to
have an inverted attachment sequence, which resulted in the PCR fragment failing to
recognize the complement sequence on pDONR. Initially this was thought to have
caused the truncations as demonstrated in BPR4 and BPR5. However, closer inspection
of the plasmid sequences suggests that the truncations may have been caused by the
recognition sequences binding to domains within UL70 that were similar to the
attachment sites (Figure 32). During sequence analysis we noticed that the
recombination site remained continuous between UL70 and pDONR, indicating that the
plasmid was circularized. In addition, BPR4 remained in frame with pDONR,
suggesting that expression of the partial UL70 protein should still be possible, and
indeed could be used to determine if the N-terminal third of the HCMV primase played
a role in primase function or in interacting with other proteins, in particular, other
members of the primase-helicase complex. Unfortunately, the design of the baculovirus
system meant that BPR4 could not be carried forward to express protein since the
recombination between the entry clone and baculovirus linear DNA requires another

118
recombination sequence that is created upon successful recombination between the PCR
fragment and pDONR.
A
GGACGTCATGCAAAAATACTTCTCGCTCGACAACTTTCTACACGATTACGTGGAGACGCATCTACTACGT
| | || |||||| | |
CCGTCCTAGCCTCACTTTCCACGGCGGACCCAGCTTTCTTGTACAAAGTTGGCATTATAAGAAAGCATTG
B AAGCCAACTTTATCGAGACGCGCTCGCTTAACGTGACGCGTTATCGACGCCGCGGTCTCACCGAGGTGCT
| ||||| | | | |||| || | | |
AGCCTCACTTTCCACGGCGGACCCAGCTTTCTTGTACAAAGTTGGCATTATAAGAAAGCATTGCTTATCA
Figure 32 Sequence analysis of BPR4 and BPR8
Comparisons of the recombination site for BPR4 (A) and BPR8 (B) reveals a degree of similarity
between the expected recombination sites (grey) and the random sequence within UL70 (as underlined)
that marked the recombination site for BPR4 and BPR8 with pDONR (purple).
Experiments to express the HCMV primase failed to produce detectable levels of the
107kDa protein. The absence of detectable protein could have been the result of low
translation efficiency caused by contaminants. In particular, the presence of calcium
may reactivate nucleases in the TNT lysate mix that were used to remove endogenous
mRNA and result in degradation of the DNA template (TNT Technical manual). The
unexpected 55kDa band which appeared during expression may indicate that another
peptide was translated from the template due to translation initiating at internal
methionines or that an artifact was created by the methods used for expression.
Increasing the fidelity for the correct initiation methionines can done by optimizing the
Mg2+
or K+ concentration (Hurst, 1996). Antibody detection was done using the S-Tag
system exclusively, which was a unique feature of the pCITE vector. The S-Tag system
is based on the interaction between a 15 amino acid S-Tag peptide and a ribonuclease S-
protein, which differs from current conventional His-Tag antibody detection in that it
does not require a secondary anti-body and hence thought to reduce the amount of
background on the Western Blot.

119
5 Concluding discussion and future directions
In this study we have shown that essential HCMV replication genes UL44, UL57, UL70,
UL102, UL105, and UL84 are highly conserved amongst isolates with variations
occurring outside of known or otherwise predicted functional domains. We have also
shown that the certain components of HCMV primase-helicase complex, UL70 and
UL105, as well as the single-stranded DNA binding protein (UL57) were highly
conserved amongst homologues, affirming the premise that functional domains remain
conserved as species diverged. Then putative functional motifs predicted from protein
motif scanning software were used as a basis for identifying potential functional
domains amongst the homologues under this premise.
We identified two N-linked glycosylation sites and a casein-kinase II phosphorylation
site that were highly conserved amongst 33 homologues of the HCMV helicase
(UL105). In addition, a cAMP-dependent protein kinase phosphorylation site was
identified in motif IV of the six putative helicase motifs previously predicted to exist in
UL105 (Graves-Woodward, 1996), providing some insight into the mechanistic
properties of that domain. In UL70, we identified a highly conserved domain consisting
of aromatic residues preceding a conserved C570 residue that has been implicated in the
structural inhibition of a compound currently being studied for its inhibitory effects on
the HCMV primase (Chen, 2007). In addition, a pair of highly conserved cysteines was
also identified although guesses as to their function, if any, are highly speculative at this
stage of the study. On one side, the domain is reminiscent of a rare PDI motif found in
only a handful of organisms in nature (Woycechowsky, 2003) while on the other, it may
be an important motif for protein structure, interacting with other conserved cysteines,
possibly with C570, or cysteines from the putative zinc finger domain (Chen, 2005), to
stabilize the tertiary structure in a manner that is similar to that of the chemokine class
of proteins (Fernandez, 2002). Similar conserved cysteine residues were previously
predicted to reside in UL57 (Gao, 1988) and were shown here to be highly conserved
across 35 homologues. In particular, a classic disulfide bond motif with the canonical
CxxC sequence was shown to be present in 34 of the 35 homologues studied and may
serve as the starting point for understanding the structural properties for this class of
proteins.

120
While conservation across homologous proteins may act as an indicator of
evolutionarily conserved functional domains, the lack of homology may also
demonstrate the ability of genes to adapt over time to specific roles unique to a
particular species. The DNA polymerase accessory proteins of HSV (UL42) and HCMV
(UL44) are both well characterized and document this distinction well. While
homologous in function and position (Ertl, 1992), UL42 operates as a monomer
(Gottlieb, 1990) and relies on basic interactions (Bridges, 2001; Zuccola, 2000), while
UL44 was shown to form a dimer and rely on hydrophobic interactions (Appleton,
2004). Similarly, UL102 shares no homology with its HSV equivalent, UL8, despite the
primase and helicase components sharing homologies of 27% and 34% respectively to
the HSV equivalents (Chee, 1990) while UL84 has no homolog equivalent in other
herpesviruses, despite being an essential component of HCMV DNA replication (Pari,
1993). Although we cannot use conservation as a means to identify putative domains for
these genes, the putative motifs predicted by the scanning software in this study can be
used as a reference for future studies elucidating protein function.
In this study, three different protein expression techniques were explored and though we
failed to express the HCMV primase, insights can be gained from the methods used in
constructing the vectors and attempts at expression in vitro. In addition, the creation of a
high copy vector for UL70 featuring recombinant arms can be the basis of future
experiments to express the HCMV primase in a BAC system.

121
6 References
Abdel-Haq, N., Chearskul, P., Al-Tatari, H., Asmar, B. (2006) New antiviral agents.
Indian Journal of Pediatrics 73(4), 313-21.
Adamo, J.E., Schroer, J., Shenk, T. (2004) Human cytomegalovirus TRS1 protein is
required for efficient assembly of DNA-containing capsids. Journal of Virology
78(19), 10221-9.
Adjei, A., Armah, H., Narter-Olaga, E. (2006) Seroprevalence of cytomegalovirus
among some voluntary blood donors at the 37 military hospital, Accra, Ghana.
Ghana Medical Journal 40(3), 99-104.
Adler, S.P. (1988) Cytomegalovirus transmission among children in day care, their
mothers and caretakers. The Pediatric Infectious Disease Journal 7(4), 279-85.
Adler, S.P. (1989) Cytomegalovirus and child day care. Evidence for an increased
infection rate among day-care workers. The New England Journal of Medicine
321(19), 1290-6.
Ahlfors, K., Ivarsson, S.A., Harris, S. (1999) Report on a long-term study of maternal
and congenital cytomegalovirus infection in Sweden. Review of prospective
studies available in the literature. Scandinavian Journal of Infectious Diseases
31(5), 443-57.
Alanen, A., Kahala, K., Vahlberg, T., Koskela, P., Vainionpää, R. (2005)
Seroprevalence, incidence of prenatal infections and reliability of maternal
history of varicella zoster virus, cytomegalovirus, herpes simplex virus and
parvovirus B19 infection in South-Western Finland. BJOG: An International
Journal of Obstetrics and Gynaecology 112(1), 50-6.
Altschul, S.F., Madden, T.L., Schäffer, A.A., Zhang, J., Zhang, Z., Miller, W. &
Lipman, D.J. (1997) Gapped BLAST and PSI-BLAST: a new generation of
protein database search programs. Nucleic Acid Research 25, 3389-3402.
Alvisi, G., Jans, D.A., Guo, J., Pinna, L.A., and Ripalti, A. (2005) A protein kinase CK2
site flanking the nuclear targeting signal enhances nuclear transport of human
cytomegalovirus ppUL44. Traffic 6(11), 1002-1013.
Anders, D.G., Gibson, W. (1988) Location, transcript analysis, and partial nucleotide
sequence of the cytomegalovirus gene encoding an early DNA-binding protein
with similarities to ICP8 of herpes simplex virus type. Journal of Virology 62(4),
1364-1372.
Anders, D.G., Kacica, M.A., Pari, G., Punturieri, S.M. (1992) Boundaries and structure
of human cytomegalovirus oriLyt, a complex origin for lytic-phase DNA
replication. Journal of Virology 66(6), 3373-84.
Anders, D.G., Kidd, J.R., Gibson, W. (1987) Immunological characterization of an early
cytomegalovirus single-strand DNA-binding protein with similarities to the
HSV major DNA-binding protein. Virology 161(2), 579-88.
Anders, D.G., McCue, L.A. (1996) The human cytomegalovirus genes and proteins
required for DNA synthesis. Intervirology 39(5-6), 378-88.
Andersson-Ellström, A., Svennerholm, B., Forssman, L. (1995) Prevalence of
antibodies to herpes simplex virus types 1 and 2, Epstein-Barr virus and
cytomegalovirus in teenage girls. Scandinavian Journal of Infectious Diseases
27(4), 315-8.
Appleton, B.A., Brooks, J., Loregian, A., Filman, D.J., Coen, D.M., and Hogle, J.M.
(2006) Crystal structure of the cytomegalovirus DNA polymerase subunit UL44

122
in complex with the C terminus from the catalytic subunit. The Journal of
Biological Chemistry 281(8), 5224-5232.
Appleton, B.A., Loregian, A., Filman, D.J., Coen, D.M., and Hogle J.M. (2004) The
cytomegalovirus DNA polymerase subunit UL44 forms a C clamp-shaped dimer.
Molecular Cell 15, 233-244.
Arezi, B., Kuchta, R.D. (2000) Eukaryotic DNA primase. Trends in Biochemical
Sciences 25(11), 572-6.
Arvin, A. (2007) Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis,
1408 pp. 1st ed, edited by A. Arvin, Mocarski, E., Moore, P.S., Roizman, B.,
Whitley, R., Yamanishi, K., 1. 1 vols. Cambridge University Press, New York.
AuCoin, D.P., Smith, G.B., Meiering, C.D., Mocarski, E.S. (2006) Betaherpesvirus-
conserved cytomegalovirus tegument protein ppUL32 (pp150) controls
cytoplasmic events during virion maturation. Journal of Virology 80(16), 8199-
210.
Baldanti, F., Silini, E., Sarasini, A., Talarico, C.L., Stanat, S.C., Biron, K.K., Furione,
M., Bono, F., Palù, G., Gerna, G. (1995) A three-nucleotide deletion in the
UL97 open reading frame is responsible for the ganciclovir resistance of a
human cytomegalovirus clinical isolate. Journal of Virology 69(2), 796-800.
Baldanti, F., Underwood, M.R., Talarico, C.L., Simoncini, L., Sarasini, A., Biron, K.K.,
Gerna, G. (1998) The Cys607-->Tyr change in the UL97 phosphotransferase
confers ganciclovir resistance to two human cytomegalovirus strains recovered
from two immunocompromised patients. Antimicrobial Agents and
Chemotherapy 42(2), 444-6.
Baldwin, B.R., Zhang, C.O., Keay, S. (2000) Cloning and epitope mapping of a
functional partial fusion receptor for human cytomegalovirus gH. The Journal of
General Virology 81(1), 27-35.
Bale, J.F.J., Zimmerman, B., Dawson, J.D., Souza, I.E., Petheram, S.J., Murph, J.R.
(1999) Cytomegalovirus transmission in child care homes. Archives of
Pediatrics and Adolescent Medicine 153(1), 75-9.
Bar, M., Shannon-Lowe, C., Geballe, A.P. (2001) Differentiation of human
cytomegalovirus genotypes in immunocompromised patients on the basis of
UL4 gene polymorphisms. Journal of Infectious Diseases 183(2), 218-225.
Barnard, E.C., Brown, G., and Stow, N.D. (1997) Deletion mutants of the herpes
simplex virus type 1 UL8 protein effect on DNA synthesis and ability to interact
with and influence the intracellular localization of the UL5 and UL52 proteins.
Virology 237, 97–106.
Barrasa, M.I., Harel, N.Y., Alwine, J.C. (2005) The phosphorylation status of the
serine-rich region of the human cytomegalovirus 86-kilodalton major
immediate-early protein IE2/IEP86 affects temporal viral gene expression.
Journal of Virology 79(3), 1428-37.
Bause, E. (1983) Structural requirements of N-glycosylation of proteins. Studies with
proline peptides as conformational probes. The Biochemical Journal 209(2),
331-6.
Beresford, C.H., Storey, G., Faed, J.M., Milligan, L.M. (1988) Cytomegalovirus
infection and blood transfusion. The New Zealand Medical Journal 101(838), 2-
4.
Bernad, A., Blanco, L., Lázaro, J.M., Martín, G., Salas, M. (1989) A conserved 3'----5'
exonuclease active site in prokaryotic and eukaryotic DNA polymerases. Cell
59(1), 219-28.

123
Biggar, R.J., Andersen, H.K., Ebbesen, P., Melbye, M., Goedert, J.J., Mann, D.L.,
Strong, D.M. (1983) Seminal fluid excretion of cytomegalovirus related to
immunosuppression in homosexual men. British Medical Journal 286(6383),
2010-2.
Biron, K.K. (2006) Antiviral drugs for cytomegalovirus diseases. Antiviral Research
71(2-3), 154-63.
Biron, K.K., Harvey, R.J., Chamberlain, S.C., Good, S.S., Smith, A.A. 3rd, Davis, M.G.,
Talarico, C.L., Miller, W.H., Ferris, R., Dornsife, R.E., Stanat, S.C., Drach, J.C.,
Townsend, L.B., Koszalka, G.W. (2002) Potent and selective inhibition of
human cytomegalovirus replication by 1263W94, a benzimidazole L-riboside
with a unique mode of action. Antimicrobial Agents and Chemotherapy 46(8),
2365-72.
Biswas, N., and Weller, S.K. (1999) A mutation in the C-terminal putative Zn2+ finger
motif of UL52 severely affects the biochemical activities of the HSV-1 helicase-
primase subcomplex. The Journal of Biological Chemistry 274(12), 8068–8076.
Blankenship, C.A., Shenk, T. (2002) Mutant human cytomegalovirus lacking the
immediate-early TRS1 coding region exhibits a late defect. Journal of Virology
76(23), 12290-9.
Boehmer, P.E. (1998) The herpes simplex virus type-1 single-stranded DNA-binding
protein, ICP8, increases the processivity of the UL9 protein DNA helicase. The
Journal of Biological Chemistry 273, 2676–2683.
Boehmer, P.E., Lehman, I.R. (1993) Herpes simplex virus type 1 ICP8: helix-
destabilizing properties. Journal of Virology 67, 711–715.
Boehmer, P.E., Nimonkar, A.V. (2003) Herpes Virus Replication. Life 55, 13-22.
Bogner, E. (2001) Human cytomegalovirus terminase as a target for antiviral
chemotherapy. Reviews in Medical Virology 12(2), 115-27.
Boivin, G., Gilbert, C., Gaudreau, A., Greenfield, I., Sudlow, R., Roberts, N.A. (2001)
Rate of emergence of cytomegalovirus (CMV) mutations in leukocytes of
patients with acquired immunodeficiency syndrome who are receiving
valganciclovir as induction and maintenance therapy for CMV retinitis. The
Journal of Infectious Diseases 184(12), 1598-602.
Bortner, C., Hernandez, T.R., Lehman, I.R., Griffith, J. (1993) Herpes simplex virus 1
single-strand DNA-binding protein (ICP8) will promote homologous pairing and
strand transfer. Journal of Molecular Biology 231, 241-250.
Bridges, K.G., Chow, C.S., and Coen, D.M. (2001) Identification of crucial hydrogen-
bonding residues for the interaction of herpes simplex virus DNA polymerase
subunits via peptide display, mutational, and calorimetric approaches. Journal of
Virology 75, 4990–4998.
Britt, W.J., and Alford, C.A. (1996) Cytomegalovirus. Fields Virology, edited by B.N.
Fields, Knipe, D.M., Howley, P.M. Lippincott-Raven Publishers, Philadelphia.
Burley, S.K., Petsko, G.A. (1985) Aromatic-aromatic interaction: a mechanism of
protein structure stabilization. Science 229(4708), 23-8.
Bush, M., Yager, D.R., Gao, M., Weisshart, K., Marcy, A.I., Coen, D.M., Knipe, D.M.
(1991) Correct intranuclear localization of herpes simplex virus DNA
polymerase requires the viral ICP8 DNA-binding protein. Journal of Virology
65, 1082-1089.
Butcher, S.J., Aitken, J., Mitchell, J., Gowen, B., Dargan, D.J. (1998) Structure of the
human cytomegalovirus B capsid by electron cryomicroscopy and image
reconstruction. Journal of Structural Biology 124(1), 70-6.

124
Calder, J.M., Stow, E.C., Stow, N.D. (1992) On the cellular localization of the
components of the herpes simplex virus type 1 helicase-primase complex and
the viral origin-binding protein. The Journal of General Virology 73(3), 531-8.
Cervia, J.S., Wenz, B., Ortolano, G.A. (2007) Leukocyte reduction's role in the
attenuation of infection risks among transfusion recipients. Clinical Infectious
Diseases 45(8), 1008-13.
Chee, M.S., Bankier, A.T., Beck, S., Bohni, R., Brown, C.M., Cerny, R., Horsnell, T.,
Hutchison, C.A., Kouzarides, T., Mertigneti, J.A., Preddie, E., Satchwell, S.C.,
Tomlinson, P., Weston, K.M., Barrell, B.G. (1990) Analysis of the protein-
coding content of the sequence of human cytomegalovirus strain AD169.
Current Topics in Microbiology and Immunology 154, 125-69.
Chen, D.H., Jiang, H., Lee, M., Liu, F., Zhou, Z.H. (1999) Three-dimensional
visualization of tegument/capsid interactions in the intact human
cytomegalovirus. Virology 260(1), 10-6.
Chen, X., Adrian, J., Cushing, T., DiMaio, D., Liang, L., Mayorga, V., Miao, S.,
Peterson, M.G., Powers, J.P., Spector, F., Stein, C., Wright, M., Xu, D., Ye Q.,
Jaen, J. (2007) SAR studies on a novel series of human cytomegalovirus primase
inhibitors. Bioorganic & Medicinal Chemistry Letters 17(8), 2188-2192.
Chen, Y., Carrington-Lawrence, S.D., Bai, P., Weller, S.K. (2005) Mutations in the
putative zinc-binding motif of UL52 demonstrate a complex interdependence
between the UL5 and UL52 subunits of the human herpes simplex virus type 1
helicase/primase complex. Journal of Virology 79(14), 9088-96.
Chen, Y.M., Knipe, D.M. (1996) A dominant mutant form of the herpes simplex virus
ICP8 protein decreases viral late gene transcription. Virology 221, 281-290.
Child, S.J., Hakki, M., De Niro, K.L., Geballe, A.P. (2004) Evasion of Cellular
Antiviral Responses by Human Cytomegalovirus TRS1 and IRS1. Journal of
Virology 78(1), 197-205.
Chiou, S.H., Yang, Y.P., Lin, J.C., Hsu, C.H., Jhang, H.C., Yang, Y.T., Lee, C.H., Ho,
L.L., Hsu, W.M., Ku, H.H., Chen, S.J., Chen, S.S., Chang, M.D., Wu, C.W.,
Juan, L.J. (2006) The immediate early 2 protein of human cytomegalovirus
(HCMV) mediates the apoptotic control in HCMV retinitis through up-
regulation of the cellular FLICE-inhibitory protein expression. Journal of
Immunology 177(9), 6199-206.
Chou, S. (1999) Antiviral drug resistance in human cytomegalovirus. Transplant
Infectious Disease 1(2), 105-14.
Chou, S., Dennison, K. (1991) Analysis of interstrain variation in cytomegalovirus
glycoprotein B sequences encoding neutralization-related epitopes. Journal of
Infectious Diseases 163, 1229-1234.
Chou, S., Erice, A., Jordan, M.C., Vercellotti, G.M., Michels, K.R., Talarico, C.L.,
Stanat, S.C., Biron, K.K. (1995) Analysis of the UL97 phosphotransferase
coding sequence in clinical cytomegalovirus isolates and identification of
mutations conferring ganciclovir resistance. The Journal of Infectious Diseases
171(3), 576-83.
Chou, S., Lurain, N.S., Thompson, K.D., Miner, R.C., Drew, W.L. (2003) Viral DNA
polymerase mutations associated with drug resistance in human cytomegalovirus.
The Journal of Infectious Diseases 188(1), 32-9.
Chou, S., Marousek, G., Guentzel, S., Follansbee, S.E., Poscher, M.E., Lalezari, J.P.,
Miner, R.C., Drew, W.L. (1997) Evolution of mutations conferring multidrug
resistance during prophylaxis and therapy for cytomegalovirus disease. Journal
of Infectious Diseases 176(3), 786-9.

125
Chou, S., Marousek, G., Parenti, D.M., Gordon, S.M., LaVoy, A.G., Ross, J.G., Miner,
R.C., Drew, W.L. (1998) Mutation in region III of the DNA polymerase gene
conferring foscarnet resistance in cytomegalovirus isolates from 3 subjects
receiving prolonged antiviral therapy. The Journal of Infectious Diseases 178(2),
526-30.
Chou, S., Marousek, G.I., Senters, A.E., Davis, M.G., Biron, K.K. (2004) Mutations in
the human cytomegalovirus UL27 gene that confer resistance to maribavir.
Journal of Virology 78(13), 7124-30.
Chou, S., Marousek, G.I., Van Wechel, L.C., Li, S., Weinberg, A. (2007a) Growth and
drug resistance phenotypes resulting from cytomegalovirus DNA polymerase
region III mutations observed in clinical specimens. Antimicrobial Agents and
Chemotherapy 51(11), 4160-2.
Chou, S., Meichsner, C.L. (2000a) A nine-codon deletion mutation in the
cytomegalovirus UL97 phosphotransferase gene confers resistance to
ganciclovir. Antimicrobial Agents and Chemotherapy 44(1), 183-5.
Chou, S., Miner, R.C., Drew, W.L. (2000) A deletion mutation in region V of the
cytomegalovirus DNA polymerase sequence confers multidrug resistance.
Journal of Infectious Diseases 182(6), 1765-8.
Chou, S., Van Wechel, L.C., Lichy, H.M., Marousek, G.I. (2005) Phenotyping of
cytomegalovirus drug resistance mutations by using recombinant viruses
incorporating a reporter gene. Antimicrobial Agents and Chemotherapy 49(7),
2710-5.
Chou, S., Waldemer, R.H., Senters, A.E., Michels, K.S., Kemble, G.W., Miner, R.C.,
Drew, W.L. (2002) Cytomegalovirus UL97 phosphotransferase mutations that
affect susceptibility to ganciclovir. The Journal of Infectious Diseases 185(2),
162-9.
Chou, S., Wechel, L.C., Marousek, G.I. (2007) Cytomegalovirus UL97 kinase
mutations that confer maribavir resistance. Journal of Infectious Diseases 196(1),
91-4.
Chou, S.W. (2001) Cytomegalovirus drug resistance and clinical implications.
Transplant Infectious Disease 3(2), 20-4.
Cihlar, T., Fuller, M.D., Cherrington, J.M. (1997) Expression of the catalytic subunit
(UL54) and the accessory protein (UL44) of human cytomegalovirus DNA
polymerase in a coupled in vitro transcription/translation system. Protein
Expression and Purification 11(2), 209-18.
Cihlar, T., Fuller, M.D., Cherrington, J.M. (1998) Characterization of drug resistance-
associated mutations in the human cytomegalovirus DNA polymerase gene by
using recombinant mutant viruses generated from overlapping DNA fragments.
Journal of Virology 72(7), 5927-36.
Cihlar, T., Fuller, M.D., Mulato, A.S., Cherrington, J.M. (1998a) A point mutation in
the human cytomegalovirus DNA polymerase gene selected in vitro by cidofovir
confers a slow replication phenotype in cell culture. Virology 248(2), 382-93.
Cocohoba, J.M., McNicholl, I.R. (2002) Valganciclovir: an advance in cytomegalovirus
therapeutics. The Annals of Pharmacotherapy 36(6), 1075-9.
Colberg-Poley, A.M. (1996) Functional roles of immediate early proteins encoded by
the human cytomegalovirus UL36-38, UL115-119, TRS1/IRS1 and US3 loci.
Intervirology 39(5-6), 350-60.
Colletti, K.S., Xu, Y., Cei, S.A., Tarrant, M., Pari, G.S. (2004) Human cytomegalovirus
UL84 oligomerization and heterodimerization domains act as transdominant
inhibitors of oriLyt-dependent DNA replication: evidence that IE2-UL84 and

126
UL84-UL84 interactions are required for lytic DNA replication. Journal of
Virology 78(17), 9203-14.
Colletti, K.S., Xu, Y., Yamboliev, I., Pari, G.S. (2005) Human cytomegalovirus UL84 is
a phosphoprotein that exhibits UTPase activity and is a putative member of the
DExD/H box family of proteins. The Journal of Biological Chemistry 280(12),
11955-60.
Compton, T. (2004) Receptors and immune sensors: the complex entry path of human
cytomegalovirus. Trends in Cell Biology 14(1), 5-8.
Compton, T., Nowlin, D.M., Cooper, N.R. (1993) Initiation of human cytomegalovirus
infection requires initial interaction with cell surface heparan sulfate. Virology
193(2), 834-41.
Craig, J.M., Macauley, J.C., Weller, T.H., Wirth, P. (1957) Isolation of intranuclear
inclusion producing agents from infants with illnesses resembling cytomegalic
inclusion disease. Proceedings of the Society for Experimental Biology and
Medicine 94(1), 4-12.
Craighead, J.E., Kanich, R.E., Almeida, J.D. (1972) Nonviral microbodies with viral
antigenicity produced in cytomegalovirus-infected cells. Journal of Virology
10(4), 766-75.
Craxton, M. (1991) Linear amplification sequencing, a powerful method for sequencing
DNA. Methods 3, 20-24.
Crute, J.J., Lehman, I.R. (1991) Herpes simplex virus-1 helicase-primase: physical and
catalytic properties. The Journal of Biological Chemistry 266, 4484-4488.
Cushing, T.D., Adrian, J., Chen, X., Dimaio, H., Doughan, B., Flygare, J., Liang, L.,
Mayorga, V., Miao, S., Mellon, H., Peterson, M.G., Powers, J.P., Spector, F.,
Stein, C., Wright, M., Xu, D., Ye, Q., Jaen, J. (2006) Discovery of a novel series
of inhibitors of human cytomegalovirus primase. Bioorganic and medicinal
Chemistry Letters 16(18), 4879-83.
Dahle, A.J., Fowler, K.B., Wright, J.D., Boppana, S.B., Britt, W.J., Pass, R.F. (2000)
Longitudinal investigation of hearing disorders in children with congenital
cytomegalovirus. Journal of the American Academy of Audiology 11(5), 283-90.
Dal Monte, P., Pignatelli, S., Rossini, G., Landini, M.P. (2004) Genomic variants
among human cytomegalovirus (HCMV) clinical isolates: the glycoprotein n
(gN) paradigm. Human Immunology 65(5), 387-394.
Dargan, D.J., Jamieson, F.E., MacLean, J., Dolan, A., Addison, C., McGeoch, D.J.
(1997) The published DNA sequence of human cytomegalovirus strain AD169
lacks 929 base pairs affecting genes UL42 and UL43. Journal of Virology
71(12), 9833-6.
Davis-Poynter, N.J., Lynch, D.M., Vally, H., Shellam, G.R., Rawlinson, W.D., Barrell,
B.G., Farrell, H.E. (1997) Identification and characterization of a G protein-
coupled receptor homolog encoded by murine cytomegalovirus. Journal of
Virology 71(2), 1521-1529.
Davison, A.J. (2002) Evolution of the herpesviruses. Veterinary Microbiology 86(1-2),
69-88.
Davison, A.J., Dolan, A., Akter, P., Addison, C., Dargan, D.J., Alcendor, D.J.,
McGeoch, D.J., Hayward, G.S. (2003) The human cytomegalovirus genome
revisited: comparison with the chimpanzee cytomegalovirus genome. The
Journal of General Virology 84(1), 17-28.
Davison, A.J., Stow, N.D. (2005) New genes from old: redeployment of dUTPase by
herpesviruses. Journal of Virology 79(20), 12880-92.

127
de Bruyn Kops, A., Knipe, D. M. (1988) Formation of DNA replication structures in
herpes virus-infected cells requires a viral DNA-binding protein. Cell 55, 857-
868.
de Castro, E., Sigrist, C.J., Gattiker, A., Bulliard, V., Langendijk-Genevaux, P.S.,
Gasteiger, E., Bairoch, A., Hulo, N. (2006) ScanProsite: detection of PROSITE
signature matches and ProRule-associated functional and structural residues in
proteins. Nucleic Acid Research 34(Web server issue), W362-5.
De Clercq, E. (2001) Antiviral drugs: current state of the art. Journal of Clinical
Virology 22(1), 73-89.
de Mello, A.L., Ferreira, E.C., Vilas Boas, L.S., Pannuti, C.S. (1996) Cytomegalovirus
infection in a day-care center in the municipality of São Paulo. Revista do
Instituto de Medicina Tropical de Sao Paulo 38(3), 165-9.
de Ory, F., Ramírez, R., García Comas, L., León, P., Sagües, M.J., Sanz, J.C. (2004) Is
there a change in cytomegalovirus seroepidemiology in Spain? European Journal
of Epidemiology 19(1), 85-9.
Digard, P., Bebrin, W., Weisshart, K., Coen, D.M. (1993) The extreme C-terminus of
herpes simplex virus DNA polymerase is crucial for functional interaction with
processivity factor UL42 and for viral replication. Journal of Virology 67, 398–
406.
Dohner, K., Sodeik, B. (2005) The role of the cytoskeleton during viral infection.
Current Topics in Microbiology and Immunology 285, 67-108.
Dolan, A., Cunningham, C., Hector, R.D., Hassan-Walker, A.F., Lee, L., Addison, C.,
Dargan, D.J., McGeoch, D.J., Gatherer, D., Emery, V.C., Griffiths, P.D., Sinzger,
C., McSharry, B.P., Wilkinson, G.W., Davison, A.J. (2004) Genetic content of
wild-type human cytomegalovirus. Journal of General Virology 85(5), 1301-12.
Dollard, S.C., Grosse, S.D., Ross, D.S. (2007) New estimates of the prevalence of
neurological and sensory sequelae and mortality associated with congenital
cytomegalovirus infection. Reviews in Medical Virology 17(5), 355-63.
Dracheva, S., Koonin, E.V., and Crute, J.J. (1995) Identification of the primase active
site of the herpes simplex virus type 1 helicase-primase. The Journal of
Biological Chemistry 270(23), 14148-14153.
Drew, W.L., Miner, R.C., Marousek, G.I., Chou, S. (2006) Maribavir sensitivity of
cytomegalovirus isolates resistant to ganciclovir, cidofovir or foscarnet. Journal
of Clinical Virology 37(2), 124-7.
Ducancelle, A., Champier, G., Alain, S., Petit, F., Le Pors, M.J., Mazeron, M.C. (2006)
A novel mutation in the UL54 gene of human cytomegalovirus isolates that
confers resistance to foscarnet. Antiviral Therapy 11(4), 537-40.
Dudas, K.C., And Ruyechan, W.T. (1998) Identification of a region of the Herpes
Simplex Virus single-stranded DNA-binding protein involved in cooperative
binding. Journal of Virology 72(1), 257–265.
Dummer, J.S., Hardy, A., Poorsattar, A., Ho, M. (1983) Early infections in kidney, heart,
and liver transplant recipients on cyclosporine. Transplantation 36(3), 259-67.
Dunn, W., Chou, C., Li, H., Hai, R., Patterson, D., Stolc, V., Zhu, H., Liu, F. (2003)
Functional profiling of a human cytomegalovirus genome. Proceedings of the
National Academy of Sciences of the United States of America 100(24), 14223-
8.
Dutch, R.E., Lehman, I.R. (1993) Renaturation of complementary DNA strands by
herpes simplex virus type 1 ICP8. Journal of Virology 67(12), 6945–6949.
Eckle, T., Lang, P., Prix, L., Jahn, G., Klingebiel, T., Handgretinger, R., Selle, B.,
Niethammer, D., Hamprecht, K. (2002) Rapid development of ganciclovir-

128
resistant cytomegalovirus infection in children after allogeneic stem cell
transplantation in the early phase of immune cell recovery. Bone Marrow
Transplantation 30(7), 433-9.
Emery, V.C., Cope, A.V., Bowen, E.F., Gor, D., Griffiths, P.D. (1999) The dynamics of
human cytomegalovirus replication in vivo. The journal of Experimental
Medicine 190(2), 177-82.
Emery, V.C., Griffiths, P.D. (2000) Prediction of cytomegalovirus load and resistance
patterns after antiviral chemotherapy. Proceedings of the National Academy of
Sciences of the United States of America 97(14), 8039-44.
Ertl, P.F., and Powell, K.L. (1992) Physical and functional interaction of human
cytomegalovirus DNA polymerase and its accessory protein (ICP36) expressed
in insect cells. Journal of Virology 66(7), 4126-4133.
Evers, D.L., Komazin, G., Ptak, R.G., Shin, D., Emmer, B.T., Townsend, L.B., Drach,
J.C. (2004) Inhibition of human cytomegalovirus replication by benzimidazole
nucleosides involves three distinct mechanisms. Antimicrobial Agents and
Chemotherapy 48(10), 3918-27.
Faizi-Khan, R., Mori, S., Eizuru, Y., Kumura-Ishii. K., Minamishima, Y. (1998)
Genetic analysis of a ganciclovir-resistant human cytomegalovirus mutant.
Antiviral Research 40(1-2), 95-103.
Falkenberg, M., Elias, P., Lehman, I.R. (1998) The herpes simplex virus type 1
helicase-primase. The Journal of Biological Chemistry 273, 32154–32157.
Feire, A.L., Koss, H., and Compton, T. (2004) Cellular integrins function as entry
receptors for human cytomegalovirus via a highly conserved disintegrin-like
domain. Proceedings of the National Academy of Sciences of the United States
of America 101, 15470–15475.
Felsenstein, J. (1989) PHYLIP -- Phylogeny Inference Package (Version 3.2). Cladistics
5, 164-166.
Fernandez, E.J., Lolis, E. (2002) Structure, function, and inhibition of chemokines.
Annual Review of Phamacology and Toxicology 42, 469-99.
Fishman, J.A., Rubin, R.H. (1998) Infection in organ-transplant recipients. The New
England Journal of Medicine 338(24), 1741-51.
Frick, D.N., Richardson, C.C. (2001) DNA primases. Annual Review of Biochemistry
70, 39-80.
Gandhi, M.K., Khanna, R. (2004) Human cytomegalovirus: clinical aspects, immune
regulation, and emerging treatments. The Lancet Infectious Diseases 4(12), 725-
38.
Gao, M., And Knipe, D.M. (1989) Genetic evidence for multiple nuclear functions of
the Herpes Simplex Virus ICP8 DNA-binding protein. Journal of Virology
63(12), 5258-5267.
Gao, M., Bouchey, J., Curtin, K., Knipe, D.M. (1988) Genetic identification of a portion
of the herpes simplex virus ICP8 protein required for DNA-binding. Virology
163, 319–329.
Gao, M., Knipe, D.M. (1991) Potential role for herpes simplex virus ICP8 DNA
replication protein in stimulation of late gene expression. Journal of Virology 65,
2666–2675.
Gao, M., Knipe, D.M. (1992) Distal protein sequences can affect the function of a
nuclear localization signal. Molecular Cell Biology 12, 1330-9.
Gao, Y., Colletti, K., Pari, G.S. (2007) Identification of Human Cytomegalovirus UL84
viral and cellular encoded binding partners using Proteomics Analysis. Journal
of Virology To be published.

129
Gavel, Y., von Heijne, G. (1990) Sequence differences between glycosylated and non-
glycosylated Asn-X-Thr/Ser acceptor sites: implications for protein engineering.
Protein Engineering 3(5), 433-42.
Gaytant, M.A., Steegers, E.A., Semmekrot, B.A., Merkus, H.M., Galama, J.M. (2002)
Congenital cytomegalovirus infection: review of the epidemiology and outcome.
Obstetrical and Gynecological Survey 57(4), 245-56.
Geary, R.S., Henry, S.P., Grillone, L.R. (2002) Fomivirsen: clinical pharmacology and
potential drug interactions. Clinical Pharmacokinetics 41(4), 255-60.
Gebert, S., Schmolke, S., Sorg, G., Floss, S., Plachter, B., Stamminger, T. (1997) The
UL84 protein of human cytomegalovirus acts as a transdominant inhibitor of
immediate-early-mediated transactivation that is able to prevent viral replication.
Journal of Virology 71(9), 7048-60.
Gibson, W. (1996) Structure and assembly of the virion. Intervirology 39(5-6), 389-400.
Gibson, W. (1999) Cytomegaloviruses (Herpesviridae) | Molecular Biology (Human).
In: A. Granoff, Webster, R.G. (Ed), Encyclopedia of Virology, 2nd Edition ed.,
pp. 351-357. 3 vols. Academic Press.
Gilbert, C., Boivin, G. (2005a) New reporter cell line to evaluate the sequential
emergence of multiple human cytomegalovirus mutations during in vitro drug
exposure. Antimicrobial Agents and Chemotherapy 49(12), 4860-6.
Goldberg, D.E., Wang, H., Azen, S.P., Freeman, W.R. (2003) Long term visual
outcome of patients with cytomegalovirus retinitis treated with highly active
antiretroviral therapy. The British Journal of Ophthalmology 87(7), 853-5.
Goldmacher, V.S., Bartle, L.M., Skaletskaya, A., Dionne, C.A., Kedersha, N.L., Vater,
C.A., Han, J.W., Lutz, R.J., Watanabe, S., Cahir McFarland, E.D., Kieff, E.D.,
Mocarski, E.S., Chittenden, T. (1999) A cytomegalovirus-encoded
mitochondria-localized inhibitor of apoptosis structurally unrelated to Bcl-2.
Proceedings of the National Academy of Sciences of the United States of
America 96(22), 12536-41.
Gorbalenya, A.E., Koonin, E.V., Donchenko, A.P., Blinov, V.M. (1988) A conserved
NTP-motif in putative helicases. Nature 333(6168), 22.
Gorbalenya, A.E., Koonin, E.V., Donchenko, A.P., Blinov, V.M. (1989) Two related
superfamilies of putative helicases involved in replication, recombination, repair
and expression of DNA and RNA genomes. Nucleic Acid Research 17, 4713-
4730.
Gottlieb, J., and Challberg, M.D. (1994) Interaction of herpes simplex virus type 1 DNA
polymerase and the UL42 accessory protein with a model primer template.
Journal of Virology 68(8), 4937-4945.
Gottlieb, J., Marcy, A.I., Coen, D.M., Challberg, M.D. (1990) The herpes simplex virus
type 1 UL42 gene product: a subunit of DNA polymerase that functions to
increase processivity. Journal of Virology 64(12), 5976-87.
Grattan, M.T., Moreno-Cabral, C.E., Starnes, V.A., Oyer, P.E., Stinson, E.B., Shumway,
N.E. (1989) Cytomegalovirus infection is associated with cardiac allograft
rejection and atherosclerosis. JAMA: The Journal of the American Medical
Association 261(24), 3561-6.
Graves-Woodward, K.L., and Weller, S.K. (1996) Replacement of gly815 in helicase
motif V alters the single-stranded DNA-dependent ATPase activity of the herpes
simplex virus type 1 helicase-primase. The Journal of Biological Chemistry
271(23), 13629–13635.
Graves-Woodward, K.L., Gottlieb, J., Challberg, M.D., and Weller, S.K. (1997)
Biochemical analyses of mutations in the HSV-1 helicase-primase that alter ATP

130
hydrolysis, DNA unwinding, and coupling between hydrolysis and unwinding.
The Journal of Biological Chemistry 272(7), 4623–4630.
Group., V.S. (2002) Safety of intravitreous fomivirsen for treatment of cytomegalovirus
retinitis in patients with AIDS. American Journal of Ophthalmology 133(4),
484–98.
Grunewald, K., Desai, P., Winkler, D.C., Heymann, J.B., Belnap, D.M., Baumeister, W.,
Steven, A.C. (2003) Three-dimensional structure of herpes simplex virus from
cryo-electron tomography. Science 302(5649), 1396-8.
Gupte, S.S., Olson, J.W., and Ruyechan, W.T. (1991) The major herpes simplex virus
type-1 DNA-binding protein is a zinc metalloprotein. The Journal of Biological
Chemistry 266(18), 11413-11416.
Hahn, G., Khan, H., Baldanti, F., Koszinowski, U.H., Revello, M.G., Gerna, G. (2002)
The human cytomegalovirus ribonucleotide reductase homolog UL45 is
dispensable for growth in endothelial cells, as determined by a BAC-cloned
clinical isolate of human cytomegalovirus with preserved wild-type
characteristics. Journal of Virology 76(18), 9551-5.
Hahn, G., Revello, M.G., Patrone, M., Percivalle, E., Campanini, G., Sarasini, A.,
Wagner, M., Gallina, A., Milanesi, G., Koszinowski, U., Baldanti, F., Gerna, G.
(2004) Human cytomegalovirus UL131-128 genes are indispensable for virus
growth in endothelial cells and virus transfer to leukocytes. Journal of Virology
78(18), 10023-33.
Hakki, M., Geballe, A.P. (2005) Double-Stranded RNA Binding by Human
Cytomegalovirus pTRS1. Journal of Virology 79(12), 7311-8.
Hamatake, R.K., Bifano, M., Hurlburt, W.W., Tenney, D.J. (1997) A functional
interaction of ICP8, the herpes simplex virus single-stranded DNA-binding
protein, and the helicase-primase complex that is dependent on the presence of
the UL8 subunit. Journal of General Virology 78, 857–865.
Handsfield, H.H., Chandler, S.H., Caine, V.A., Meyers, J.D., Corey, L., Medeiros, E.,
McDougall, J.K. (1985) Cytomegalovirus infection in sex partners: evidence for
sexual transmission. The Journal of Infectious Diseases 151(2), 344-8.
Hansen, S.G., Strelow, L.I., Franchi, D.C., Anders, D.G., Wong, S.W. (2003) Complete
sequence and genomic analysis of rhesus cytomegalovirus. Journal of Virology
77(12), 6620-36.
Hanson, M.N., Preheim, L.C., Chou, S., Talarico, C.L., Biron, K.K., Erice, A. (1995)
Novel mutation in the UL97 gene of a clinical cytomegalovirus strain conferring
resistance to ganciclovir. Antimicrobial Agents and Chemotherapy 39(5), 1204-
5.
Hantz, S., Michel, D., Fillet, A.M., Guigonis, V., Champier, G., Mazeron, M.C.,
Bensman, A., Denis, F., Mertens, T., Dehee, A., Alain, S. (2005) Early selection
of a new UL97 mutant with a severe defect of ganciclovir phosphorylation after
valaciclovir prophylaxis and short-term ganciclovir therapy in a renal transplant
recipient. Antimicrobial Agents and Chemotherapy 49(4), 1580-3.
Hassan-Walker, A.F., Okwuadi, S., Lee, L., Griffiths, P.D., Emery, V.C. (2004)
Sequence variability of the alpha-chemokine UL146 from clinical strains of
human cytomegalovirus. Journal of Medical Virology 74(4), 573-579.
Hassan, J., Connell, J. (2007) Translational mini-review series on infectious disease:
congenital cytomegalovirus infection: 50 years on. Clinical and Experimental
Immunology 149(2), 205-10.
Hayajneh, W.A., Colberg-Poley, A.M., Skaletskaya, A., Bartle, L.M., Lesperance,
M.M., Contopoulos-Ioannidis, D.G., Kedersha, N.L., Goldmacher, V.S. (2001)

131
The sequence and antiapoptotic functional domains of the human
cytomegalovirus UL37 exon 1 immediate early protein are conserved in multiple
primary strains. Virology 279(1), 233-40.
Haynes, R.O. (2002) Integrins: bidirectional, allosteric signaling machines. Cell 110(6),
673-87.
He, X., Lehman, I.R. (2001) An initial ATP-independent step in the unwinding of a
herpes simplex virus type 1 origin of replication by a complex of the viral
origin-binding protein and single-strand DNAbinding protein. Proceedings of
the National Academy of Sciences of the United States of America 98, 3024–
3028.
He, Y.S., Xu, L., Huang, E.S. (1992) Characterization of human cytomegalovirus UL84
early gene and identification of its putative protein product. Journal of Virology
66(2), 1098-108.
Hecker, M., Qiu, D., Marquardt, K., Bein, G., Hackstein, H. (2004) Continuous
cytomegalovirus seroconversion in a large group of healthy blood donors. Vox
Sanguinis 86(1), 41-4.
Heilbronn, R., Jahn, G., Bürkle, A., Freese, U.K., Fleckenstein, B., zur Hausen, H.
(1987) Genomic localization, sequence analysis, and transcription of the putative
human cytomegalovirus DNA polymerase gene. Journal of Virology 61(1), 119-
24.
Hernandez, T.R., Lehman, I.R. (1990) Functional interaction between the herpes
simplex-1 DNA polymerase and UL42 protein. The Journal of Biological
Chemistry 265, 11227–11232.
Heymann, J.B., Cheng, N., Newcomb, W.W., Trus, B.L., Brown, J.C., Steven, A.C.
(2003) Dynamics of herpes simplex virus capsid maturation visualized by time-
lapse cryo-electron microscopy. Nature Structural Biology 10(5), 334-41.
Hitomi, S., Kozuka-Hata, H., Chen, Z., Sugano, S., Yamaguchi, N., Watanabe, S. (1997)
Human cytomegalovirus open reading frame UL11 encodes a highly
polymorphic protein expressed on the infected cell surface. Archives of
Virology 142(7), 1407-27.
Hizel, S., Parker, S., Onde, U. (1999) Seroprevalence of cytomegalovirus infection
among children and females in Ankara, Turkey, 1995. Pediatrics International
41(5), 506-9.
Hjelmesaeth, J., Sagedal, S., Hartmann, A., Rollag, H., Egeland, T., Hagen, M., Nordal,
K.P., Jenssen, T. (2004) Asymptomatic cytomegalovirus infection is associated
with increased risk of new-onset diabetes mellitus and impaired insulin release
after renal transplantation. Diabetologia 47(9), 1550-6.
Ho, E.S., Lin, D.C., Mendel, D.B., Cihlar, T. (2000) Cytotoxicity of antiviral
nucleotides adefovir and cidofovir is induced by the expression of human renal
organic anion transporter 1. Journal of the American Society of Nephrology
11(3), 383-93.
Homa, F.L., Brown, J.C. (1997) Capsid assembly and DNA packaging in herpes
simplex virus. Reviews in Medical Virology 7(2), 107-122.
Hui, E.K., Ralston, K., Judd, A.K., Nayak, D.P. (2003) Conserved cysteine and
histidine residues in the putative zinc finger motif of the influenza A virus M1
protein are not critical for influenza virus replication. Journal of General
Virology 84(11), 3105-13.
Hulo, N., Bairoch, A., Bulliard, V., Cerutti, L., De Castro, E., Langendijk-Genevaux,
P.S., Pagni, M., Sigrist, C.J. (2006) The PROSITE database. Nucleic Acid
Research 34, D227-30.

132
Hurst, R., Maffitt, M., Murray, E., Kappelman, J., Xu, Q., Mendez, J., Butler, B., Ryan
A., and Beckler, G.S. (1996) The TNT® T7 Quick Coupled
Transcription/Translation System. Promega Notes 58, 8-11.
Hwang, C.B., Ruffner, K.L., Coen, D.M. (1992) A point mutation within a distinct
conserved region of the herpes simplex virus DNA polymerase gene confers
drug resistance. Journal of Virology 66(3), 1774-6.
Ijichi, O., Michel, D., Mertens, T., Miyata, K., Eizuru, Y. (2002) GCV resistance due to
the mutation A594P in the cytomegalovirus protein UL97 is partially
reconstituted by a second mutation at D605E. Antiviral Research 53(2), 135-42.
Ilyina, T.V., Gorbalenya, A.E., Koonin, E.V. (1992) Organization and evolution of
bacterial and bacteriophage primase-helicase systems. Journal of Molecular
Evolution 34(4), 351–357.
Iskenderian, A.C., Huang, L., Reilly, A., Stenberg, R.M., Anders, D.G. (1996) Four of
eleven loci required for transient complementation of human cytomegalovirus
DNA replication cooperate to activate expression of replication genes. Journal of
Virology 70(1), 383-92.
Jakob, U., Muse, W., Eser, M., Bardwell, J.C. (1999) Chaperone activity with a redox
switch. Cell 96(3), 341-52.
Jans, D.A., Hübner, S. (1996) Regulation of protein transport to the nucleus: central role
of phosphorylation. Physiological Reviews 76(3), 651-85.
Jarvis, M.A., Jones, T.R., Drummond, D.D., Smith, P.P., Britt, W.J., Nelson, J.A.,
Baldick, C.J. (2004) Phosphorylation of human cytomegalovirus glycoprotein B
(gB) at the acidic cluster casein kinase 2 site (Ser900) is required for localization
of gB to the trans-Golgi network and efficient virus replication. Journal of
Virology.
Jarvis, M.A., Nelson, J.A. (2002) Human cytomegalovirus persistence and latency in
endothelial cells and macrophages. Current Opinion in Microbiology 5(4), 403-7.
Jasanoff, A., Weiss, M.A. (1993) Aromatic-histidine interactions in the zinc finger
motif: structural inequivalence of phenylalanine and tyrosine in the hydrophobic
core. Biochemistry 32(6), 1423-32.
Jiang, X.J., Adler, B., Sampaio, K.L., Digel, M., Jahn, G., Ettischer, N., Stierhof, Y.D.,
Scrivano, L., Koszinowski, U., Mach, M., Sinzger, C. (2008) UL74 of human
cytomegalovirus contributes to virus release by promoting secondary
envelopment of virions. Journal of Virology 82(6), 2802-12.
Kääriäinen, L., Klemola, E., Paloheimo, J. (1966) Rise of cytomegalovirus antibodies in
an infectious-mononucleosis-like syndrome after transfusion. British Medical
Journal 1(5498), 1270-2.
Kangro, H.O., Osman, H.K., Lau, Y.L., Heath, R.B., Yeung, C.Y., Ng, M.H. (1994)
Seroprevalence of antibodies to human herpesviruses in England and Hong
Kong. Journal of Medical Virology 43(1), 91-6.
Kiehl, A., Huang, L., Franchi, D., and Anders, D.G. (2003) Multiple 5' ends of human
cytomegalovirus UL57 transcripts identify a complex, cycloheximide-resistant
promoter region that activates oriLyt. Virology 314, 410–422.
Kiss, P., De Bacquer, D., Sergooris, L., De Meester, M., VanHoorne, M. (2002)
Cytomegalovirus infection: an occupational hazard to kindergarten teachers
working with children aged 2.5-6 years. International Journal of Occupational
and Environmental Health 8(2), 79-86.
Klinedinst, D.K., and Challberg, M.D. (1994) Helicase-primase complex of herpes
simplex virus type 1 a mutation in the UL52 subunit abolishes primase activity.
Journal of Virology 68(6), 3693-3701.

133
Knowles, S.J., Grundy, K., Cahill, I., Cafferkey, M.T., Geary, M. (2005) Low
cytomegalovirus sero-prevalence in Irish pregnant women. Irish Medical Journal
98(7), 210-2.
Kochoyan, M., Keutmann, H.T., Weiss, M.A. (1991) Architectural rules of the zinc-
finger motif: comparative two-dimensional NMR studies of native and
"aromatic-swap" domains define a "weakly polar switch". Proceedings of the
National Academy of Sciences of the United States of America 88(19), 8455-9.
Komazin, G., Ptak, R.G., Emmer, B.T., Townsend, L.B., Drach, J.C. (2003) Resistance
of human cytomegalovirus to the benzimidazole L-ribonucleoside maribavir
maps to UL27. Journal of Virology 77(21), 11499-506.
Komazin, G., Townsend, L.B., Drach, J.C. (2004) Role of a mutation in human
cytomegalovirus gene UL104 in resistance to benzimidazole ribonucleosides.
Journal of Virology 78(2), 710-5.
Kothari, A., Ramachandran, V.G., Gupta, P., Singh, B., Talwar, V. (2002)
Seroprevalence of cytomegalovirus among voluntary blood donors in Delhi,
India. Journal of Health, Population, and Nutrition 20(4), 348-51.
Krishnaswamy, S., Rossmann, M.G. (1990) Structural refinement and analysis of
Mengo virus. Journal of Molecular Biology 211(4), 803-44.
Krosky, P.M., Baek, M-C., Jahng, W.J., Barrera, I., Harvey, R.J., Biron, K.K., Coen,
D.M., and Sethna P.B. (2003) The human cytomegalovirus UL44 protein Is a
substrate for the UL97 protein kinase. Journal of Virology 77(14), 7720–7727.
Krosky, P.M., Baek, M.C., Coen, D.M. (2003a) The human cytomegalovirus UL97
protein kinase, an antiviral drug target, is required at the stage of nuclear egress.
Journal of Virology 77(2), 905-14.
Krosky, P.M., Underwood, M.R., Turk, S.R., Feng, K.W., Jain, R.K., Ptak, R.G.,
Westerman, A.C., Biron, K.K., Townsend, L.B., Drach, J.C. (1998) Resistance
of human cytomegalovirus to benzimidazole ribonucleosides maps to two open
reading frames: UL89 and UL56. Journal of Virology 72(6), 4721-8.
Kubo, T., Rai, S.K., Nakanishi, M., Yamano, T. (1991) Seroepidemiological study of
herpes viruses in Nepal. The Southeast Asian Journal of Tropical Medicine and
Public Health 22(3), 323-5.
Lang, D.J., Kummer, J.F. (1975) Cytomegalovirus in semen: observations in selected
populations. The Journal of Infectious Diseases 132(4), 472-3.
Lee, S.S.-K., Lehman, I.R. (1997) Unwinding of the box I element of a herpes simplex
virus type 1 origin by a complex of the viral origin binding protein, single-strand
DNA binding protein, and singlestranded DNA. Proceedings of the National
Academy of Sciences of the United States of America 94, 2838–2842.
Lemström, K., Koskinen, P., Krogerus, L., Daemen, M., Bruggeman, C., Häyry, P.
(1995) Cytomegalovirus antigen expression, endothelial cell proliferation, and
intimal thickening in rat cardiac allografts after cytomegalovirus infection.
Circulation 92(9), 2594-604.
Limaye, A.P., Corey, L., Koelle, D.M., Davis, C.L., Boeckh, M. (2000) Emergence of
ganciclovir-resistant cytomegalovirus disease among recipients of solid-organ
transplants. Lancet 356(9230), 645-9.
Ling, P.D., Lednicky, J.A., Keitel, W.A., Poston, D.G., White, Z.S., Peng, R., Liu, Z.,
Mehta, S.K., Pierson, D.L., Rooney, C.M., Vilchez, R.A., Smith, E.O., Butel, J.S.
(2003) The dynamics of herpesvirus and polyomavirus reactivation and
shedding in healthy adults: a 14-month longitudinal study. The Journal of
Infectious Diseases 187(10), 1571-80.

134
Lischka, P., Rauh, C., Mueller, R., Stamminger, T. (2006) Human cytomegalovirus
UL84 protein contains two nuclear export signals and shuttles between the
nucleus and the cytoplasm. Journal of Virology 80(20), 10274-80.
Lischka, P., Sorg, G., Kann, M., Winkler, M., Stamminger, T. (2003) A
nonconventional nuclear localization signal within the UL84 protein of human
cytomegalovirus mediates nuclear import via the importin alpha/beta pathway.
Journal of Virology 77(6), 3734-48.
Liu, Z., Wang, E., Taylor, W., Yu, H., Wu, T., Wan, Z., Huang, Y., Ni, Z., Sackett, D.
(1990) Prevalence survey of cytomegalovirus infection in children in Chengdu.
American Journal of Epidemiology 131(1), 143-50.
Loh, L.C., Keeler, V.D., Shanley, J.D. (1999) Sequence requirements for the nuclear
localization of the murine cytomegalovirus M44 gene product pp50. Virology
259, 43–59.
Loh, L.C., Locke, D., Melnychuk, R., and Laferte, S. (2000) The RGD sequence in the
cytomegalovirus DNA polymerase accessory protein can mediate cell adhesion.
Virology 272, 302-314.
Lopper, M., Compton, T. (2004) Coiled-coil domains in glycoproteins B and H are
involved in human cytomegalovirus membrane fusion. Journal of Virology
78(15), 8333-41.
Loregian, A., Appleton, B.A., Hogle, J.M., and Coen, D.M. (2004) Specific residues in
the connector loop of the human cytomegalovirus DNA polymerase accessory
protein UL44 are crucial for interaction with the UL54 catalytic subunit. Journal
of Virology 78(17), 9084–9092.
Loregian, A., Appleton, B.A., Hogle, J.M., and Coen, D.M. (2004a) Residues of human
cytomegalovirus DNA polymerase catalytic subunit UL54 that are necessary and
sufficient for interaction with the accessory protein UL44. Journal of Virology
78(1), 158-167.
Loregian, A., Palu, G. (2005) Disruption of the interactions between the subunits of
herpesvirus DNA polymerases as a novel antiviral strategy. Clinical
Microbiology and Infection 11(6), 437-46.
Loregian, A., Rigatti, R., Murphy, M., Schievano, E., Palu, G., Marsden, H.S. (2003)
Inhibition of human cytomegalovirus DNA polymerase by C-terminal peptides
from the UL54 subunit. Journal of Virology 77(15), 8336-44.
Lorz, K., Hofmann, H., Berndt, A., Tavalai, N., Mueller, R., Schlotzer-Schrehardt, U.,
Stamminger, T. (2006) Deletion of open reading frame UL26 from the human
cytomegalovirus genome results in reduced viral growth, which involves
impaired stability of viral particles. Journal of Virology 80(11), 5423-34.
Lukac, D.M., Alwine, J.C. (1999) Effects of human cytomegalovirus major immediate-
early proteins in controlling the cell cycle and inhibiting apoptosis: studies with
ts13 cells. Journal of Virology 73(4), 2825-31.
Lurain, N.S., Kapell, K.S., Huang, D.D., Short, J.A., Paintsil, J., Winkfield, E., Benedict,
C.A., Ware., C.F., Bremer, J.W. (1999) Human cytomegalovirus UL144 open
reading frame: sequence hypervariability in low-passage clinical isolates.
Journal of Virology 73(12), 10040-10050.
Lurain, N.S., Spafford, L.E., Thompson, K.D. (1994) Mutation in the UL97 open
reading frame of human cytomegalovirus strains resistant to ganciclovir. Journal
of Virology 68(7), 4427-31.
Lurain, N.S., Thompson, K.D., Holmes, E.W., Read, G.S. (1992) Point mutations in the
DNA polymerase gene of human cytomegalovirus that result in resistance to
antiviral agents. Journal of Virology 66(12), 7146-52.

135
Mach, M., Kropff, B., Kryzaniak, M., and Britt, W. (2005) Complex formation by
glycoproteins M and N of human cytomegalovirus: structural and functional
aspects. Journal of Virology 79, 2160–2170.
Makhov, A.M., Boehmer, P.E., Lehman, I.R., Griffith, J.D. (1996) Visualization of the
unwinding of long DNA chains by the herpes simplex virus type 1 UL9 protein
and ICP8. Journal of Molecular Biology 258, 789–799.
Malm, G., Engman, M.L. (2007) Congenital cytomegalovirus infections. Seminars in
Fetal and Neonatal Medicine 12(3), 154-9.
Mansat, A., Mengelle, C., Chalet, M., Boumzebra, A., Mieusset, R., Puel, J., Prouheze,
C., Segondy, M. (1997) Cytomegalovirus detection in cryopreserved semen
samples collected for therapeutic donor insemination. Human Reproduction
12(8), 1663-6.
Mapelli, M., Muhleisen, M., Persico, G., Van Der Zandt, H., and Tucker, P.A. (2000)
The 60-residue C-terminal region of the single-stranded DNA binding protein of
Herpes Simplex Virus type 1 Is required for cooperative DNA binding. Journal
of Virology 74(19), 8812–8822.
Mar, E.C., Patel, P.C., Huang, E.S. (1981) Human cytomegalovirusassociated DNA
polymerase and protein kinase activities. Journal of General Virology 57, 149–
156.
Marfori, J.E., Exner, M.M., Marousek, G.I., Chou, S., Drew, W.L. (2007) Development
of new cytomegalovirus UL97 and DNA polymerase mutations conferring drug
resistance after valganciclovir therapy in allogeneic stem cell recipients. Journal
of Clinical Virology 38(2), 120-5.
Marschall, M., Freitag, M., Suchy, P., Romaker, D., Kupfer, R., Hanke, M.,
Stamminger, T. (2003) The protein kinase pUL97 of human cytomegalovirus
interacts with and phosphorylates the DNA polymerase processivity factor
pUL44. Virology 311(1), 60-71.
Marsden, H.S., McLean, G.W., Barnard, E.C., Francis, G.J., MacEachran, K., Murphy,
M., McVey, G., Cross, A., Abbotts, A.P., Stow, N.D. (1997) The catalytic
subunit of the DNA polymerase of herpes simplex virus type 1 interacts
specifically with the C terminus of the UL8 component of the viral helicase-
primase complex. Journal of Virology 71(9), 6390-7.
Martin, M., Gilbert, C., Covington, E., Boivin, G. (2006) Characterization of human
cytomegalovirus (HCMV) UL97 mutations found in a valganciclovir/oral
ganciclovir prophylactic trial by use of a bacterial artificial chromosome
containing the HCMV genome. Journal of Infectious Diseases 194(5), 579-83.
McGeoch, D.J., Cook, S., Dolan, A., Jamieson, F.E., Telford, E.A.R. (1995) Molecular
Phylogeny and Evolutionary Timescale for the Family of Mammalian
Herpesviruses. Journal of Molecular Biology 247, 443–458.
McGeoch, D.J., Dalrymple, M.A., Dolan, A., McNab, D., Perry, L.J., Taylor, P.,
Challberg, M.D. (1988) Structures of herpes simplex virus type 1 genes required
for replication of virus DNA. Journal of Virology 62(2), 444-53.
McGeoch, D.J., Gatherer, D. (2005) Integrating reptilian herpesviruses into the family
herpesviridae. Journal of Virology 79(2), 725-31.
McGowan, M.P., Hayes, K., Kovacs, G.T., Leydon, J.A. (1983) Prevalence of
cytomegalovirus and herpes simplex virus in human semen. International
Journal of Andrology 6(4), 331-6.
McLean, G.W., Abbotts, A.P., Parry, M.E., Marsden, H.S., Stow, N.D. (1994) The
herpes simplex virus type 1 origin-binding protein interacts specifically with the
viral UL8 protein. The Journal of General Virology 75(10), 2699-706.

136
McMahon, T.P., and Anders, D.G. (2002) Interactions between Human
Cytomegalovirus Helicase-Primase Proteins. Virus Research 86, 39–52.
McSharry, J.J., McDonough, A., Olson, B., Hallenberger, S., Reefschlaeger, J., Bender,
W., Drusano, G.L. (2001) Susceptibilities of human cytomegalovirus clinical
isolates to BAY38-4766, BAY43-9695, and ganciclovir. Antimicrobial Agents
and Chemotherapy 45(10), 2925-7.
Meggio, F., Pinna, L.A. (2003) One-thousand-and-one substrates of protein kinase CK2?
FASEB 17(3), 349-68.
Mehta, S.K., Stowe, R.P., Feiveson, A.H., Tyring, S.K., Pierson, D.L. (2000)
Reactivation and shedding of cytomegalovirus in astronauts during spaceflight.
The Journal of Infectious Diseases 182(6), 1761-4.
Mendelman, L.V., Beauchamp, B.B., Richardson, C.C. (1994) Requirement for a zinc
motif for template recognition by the bacteriophage T7 primase. The EMBO
Journal 13(16), 3909-16.
Mercorelli, B., Sinigalia, E., Loregian, A., Palù, G. (2007) Human cytomegalovirus
DNA replication: antiviral targets and drugs. Reviews in Medical Virology In
Print.
Mettenleiter, T.C. (2004) Budding events in herpesvirus morphogenesis. Virus Research
106(2), 167-80.
Meyer-König, U., Haberland, M., von Laer, D., Haller, O., Hufert, F.T. (1998)
Intragenic variability of human cytomegalovirus glycoprotein B in clinical
strains. The Journal of Infectious Diseases 177(5), 1162-9.
Michael, S.F., Kilfoil, V.J., Schmidt, M.H., Amann, B.T., Berg, J.M. (1992) Metal
binding and folding properties of a minimalist Cys2His2 zinc finger peptide.
Proceedings of the National Academy of Sciences of the United States of
America 89(11), 4796-800.
Michel, D., Milotić, I., Wagner, M., Vaida, B., Holl, J., Ansorge, R., Mertens, T. (2005)
The human cytomegalovirus UL78 gene is highly conserved among clinical
isolates, but is dispensable for replication in fibroblasts and a renal artery organ-
culture system. Journal of General Virology 86(2), 297-306.
Miles, D.J., van der Sande, M., Jeffries, D., Kaye, S., Ismaili, J., Ojuola, O., Sanneh, M.,
Touray, E.S., Waight, P., Rowland-Jones, S., Whittle, H., Marchant, A. (2007)
Cytomegalovirus infection in Gambian infants leads to profound CD8 T-cell
differentiation. Journal of Virology 81(11), 5766-76.
Miller, W.J., McCullough, J., Balfour, H.H. Jr, Haake, R.J., Ramsay, N.K., Goldman,
A., Bowman, R., Kersey, J. (1991) Prevention of cytomegalovirus infection
following bone marrow transplantation: a randomized trial of blood product
screening. Bone Marrow Transplantation 7(3), 227-34.
Mocarski, E.S., Prichard, M.N., Tan, C.S., Brown, J.M. (1997) Reassessing the
organisation of the UL42-UL43 region of the human cytomegalovirus strain
AD169 genome. Virology 239(1), 169-175.
Mocarski, E.S., Tan Courcelle, C. (2001) Cytomegalovirus and Their Replication. In:
D.M. Knipe, Howley, P.M., (Ed), Field's Virology, 4th ed., pp. 2629-2673. Vol.
2. Lippencott Williams and Wilkins, Philadelphia.
Monahan, S.J., Barlam, T.F., Crumpacker, C.S., Parris, D.S. (1993) Two regions of the
herpes simplex virus type 1 UL42 protein are required for its functional
interaction with viral DNA polymerase. Journal of Virology 67, 5922–5931.
Mousavi-Jazi, M., Schloss, L., Drew, W.L., Linde, A., Miner, R.C., Harmenberg, J.,
Wahren, B., Brytting, M. (2001) Variations in the cytomegalovirus DNA

137
polymerase and phosphotransferase genes in relation to foscarnet and
ganciclovir sensitivity. Journal of Clinical Virology 23(1-2), 1-15.
Mousavi-Jazi, M., Schloss, L., Wahren, B., Brytting, M. (2003) Point mutations induced
by foscarnet (PFA) in the human cytomegalovirus DNA polymerase. Journal of
Clinical Virology 26(3), 301-6.
Munro, S.C., Hall, B., Whybin, L.R., Leader, L., Robertson, P., Maine, G.T., Rawlinson,
W.D. (2005) Diagnosis of and screening for cytomegalovirus infection in
pregnant women. Journal of Clinical Microbiology 43(9), 4713-8.
Murphy, E., Yu, D., Grimwood, J., Schmutz, J., Dickson, M., Jarvis, M.A., Hahn, G.,
Nelson, J.A., Myers, R.M., Shenk, T.E. (2003) Coding potential of laboratory
and clinical strains of human cytomegalovirus. Proceedings of the National
Academy of Sciences of the United States of America 100(25), 14976-81.
Natali, A., Valcavi, P., Medici, M.C., Dieci, E., Montali, S., Chezzi, C. (1997)
Cytomegalovirus infection in an Italian population: antibody prevalence, virus
excretion and maternal transmission. The New Microbiologica 20(2), 123-33.
Navarro, D., Paz, P., Tugizov, S., Topp, K., La, Vail. J, Pereira, L. (1993) Glycoprotein
B of human cytomegalovirus promotes virion penetration into cells,
transmission of infection from cell to cell, and fusion of infected cells. Virology
197(1), 143-58.
Neote, K., DiGregorio, D., Mak, J.Y., Horuk, R., Schall, T.J. (1993) Molecular cloning,
functional expression, and signaling characteristics of a C-C chemokine receptor.
Cell 72(3), 415-25.
Nguyen-Huynh, A.T., Schaffer, P.A. (1998) Cellular transcription factors enhance
herpes simplex virus type 1 oriS-dependent DNA replication. Journal of
Virology 72(5), 3635–3645.
Nimonkar, A.V., Boehmer, P.E. (2003) Reconstitution of recombination-dependent
DNA synthesis in herpes simplex virus 1. Proceedings of the National Academy
of Sciences of the United States of America 100(18), 10201-6.
Nishiyama, Y., Maeno, K., Yoshida, S. (1983) Characterization of human
cytomegalovirus-induced DNA polymerase and the associated 3'-to-5'
exonuclease. Virology 124, 221–231.
Nogalski, M.T., Podduturi, J.P., DeMeritt, I.B., Milford, L.E., Yurochko, A.D. (2007)
The human cytomegalovirus virion possesses an activated casein kinase II that
allows for the rapid phosphorylation of the inhibitor of NF-kappaB,
IkappaBalpha. Journal of Virology 81(10), 5305-14.
Ogawa-Goto, K., Tanaka, K., Gibson, W., Moriishi, E., Miura, Y., Kurata, T., Irie, S.,
Sata, T. (2003) Microtubule network facilitates nuclear targeting of human
cytomegalovirus capsid. Journal of Virology 77(15), 8541-7.
Pari, G.S., and Anders, D.G. (1993) Eleven loci encoding trans-acting factors are
required for transient complementation of human cytomegalovirus oriLyt-
dependent DNA replication. Journal of Virology 67(12), 6979-6988.
Pari, G.S., Kacica, M.A., and Anders, D.A. (1993a) Open reading frames UL44,
IRS1/TRS1, and UL36-38 are required for transient complementation of human
cytomegalovirus oriLyt-dependent DNA synthesis. Journal of Virology 67,
2575–2582.
Park, M.Y., Kim, Y.E., Seo, M.R., Lee, J.R., Lee, C.H., Ahn, J.H. (2006) Interactions
among four proteins encoded by the human cytomegalovirus UL112-113 region
regulate their intranuclear targeting and the recruitment of UL44 to
prereplication foci. Journal of Virology 80(6), 2718-27.

138
Pass, R.F. (2005) Congenital cytomegalovirus infection and hearing loss. Herpes: The
Journal of the IHMF 12(2), 50-5.
Pass, R.F., Hutto, C., Lyon, M.D., Cloud, G. (1990) Increased rate of cytomegalovirus
infection among day care center workers. The Pediatric Infectious Disease
Journal 9(7), 465-70.
Pass, R.F., Hutto, S.C., Reynolds, D.W., Polhill, R.B. (1984) Increased frequency of
cytomegalovirus infection in children in group day care. Pediatrics 74(1), 121-6.
Pescovitz, M.D., Rabkin, J., Merion, R.M., Paya, C.V., Pirsch, J., Freeman, R.B.,
O'Grady, J., Robinson, C., To, Z., Wren, K., Banken, L., Buhles, W., Brown, F.
(2000) Valganciclovir results in improved oral absorption of ganciclovir in liver
transplant recipients. Antimicrobial Agents and Chemotherapy 44(10), 2811-5.
Pignatelli, S., Dal Monte, P., Landini, M.P. (2001) gpUL73 (gN) genomic variants of
human cytomegalovirus isolates are clustered into four distinct genotypes.
Journal of General Virology 82(11), 2777-84.
Pilger, B.D., Cui, C., and Coen, D.M. (2004) Identification of a small molecule that
inhibits herpes simplex virus DNA polymerase subunit interactions and viral
replication. Chemical Biology 11, 647–654.
Pinna, L.A. (1990) Casein kinase 2: an 'eminence grise' in cellular regulation?
Biochemica et Biophysica atca 1054(3), 267-84.
Pouteil-Noble, C., Ecochard, R., Landrivon, G., Donia-Maged, A., Tardy, J.C.,
Bosshard, S., Colon, S., Betuel, H., Aymard, M., Touraine, J.L. (1993)
Cytomegalovirus infection--an etiological factor for rejection? A prospective
study in 242 renal transplant patients. Transplantation 55(4), 851-7.
Prabhakar, P., Bailey, A., Smikle, M.F., Ashley, D. (1992) Seroprevalence of
cytomegalovirus infection in a selected population in Jamaica. The West Indian
Medical Journal 41(4), 133-5.
Prichard, M.N., Jairath, S., Penfold, M.E., St Jeor, S., Bohlman, M.C., Pari, G.S. (1998)
Identification of persistent RNA-DNA hybrid structures within the origin of
replication of human cytomegalovirus. Journal of Virology 72(9), 6997-7004.
Prichard, M.N., Penfold, M.E., Duke, G.M., Spaete, R.R., Kemble, G.W. (2001) A
review of genetic differences between limited and extensively passaged human
cytomegalovirus strains. Reviews in Medical Virology 11(3), 191-200.
Prior, J.R., Morroll, D.R., Birks, A.G., Matson P.L. and Lieberman, B.A. (1994) The
screening for cytomegalovirus antibody in semen donors and recipients within a
donor insemination program. Human Reproduction 9(11), 2076-2078.
Pultoo, A., Meetoo, G., Pyndiah, M.N., Khittoo, G. (2001) Seroprevalence of
cytomegalovirus infection in Mauritian volunteer blood donors. Indian Journal
of Medical Sciences 55(2), 73-8.
Ramirez-Aguilar, K.A., Kuchta, R.D. (2004) Mechanism of primer synthesis by the
herpes simplex virus 1 helicase-primase. Biochemistry 43(6), 1754-62.
Ramirez-Aguilar, K.A., Low-Nam, N.A., Kuchta, R.D. (1995) Key Role of Template
Sequence for Primer Synthesis by the Herpes Simplex Virus 1 Helicase-Primase.
Biochemistry 41(49), 14569-79.
Randell, J.C., Coen, D.M. (2004) The herpes simplex virus processivity factor, UL42,
binds DNA as a monomer. Journal of Molecular Biology 335(2), 409-13.
Rasmussen, L., Geissler, A., Winters, M. (2003) Inter- and intragenic variations
complicate the molecular epidemiology of human cytomegalovirus. The Journal
of Infectious Diseases 187(5), 809-19.
Rawlinson, W.D., Barrell, B.G. (1993) Spliced transcripts of human cytomegalovirus.
Journal of Virology 67(9), 5502-13.

139
Rawlinson, W.D., Farrell, H.E., Barrell, B.G. (1996) Analysis of the complete DNA
sequence of murine cytomegalovirus. Journal of Virology 70(12), 8833-49.
Razonable, R.R., Emery, V.C; 11th Annual Meeting of the IHMF (International Herpes
Management Forum). (2004) Management of CMV infection and disease in
transplant patients. 27-29 February 2004. Herpes: The Journal of the IHMF
11(3), 77-86.
Reefschlaeger, J., Bender, W., Hallenberger, S., Weber, O., Eckenberg, P., Goldmann,
S., Haerter, M., Buerger, I., Trappe, J., Herrington, J.A., Haebich, D.,
Ruebsamen-Waigmann, H. (2001) Novel non-nucleoside inhibitors of
cytomegaloviruses (BAY 38-4766): in vitro and in vivo antiviral activity and
mechanism of action. The Journal of Antimicrobial Chemotherapy 48(6), 757-67.
Reeves, M.B., MacAry, P.A., Lehner, P.J., Sissons, J.G., Sinclair, J.H. (2005) Latency,
chromatin remodeling, and reactivation of human cytomegalovirus in the
dendritic cells of healthy carriers. Proceedings of the National Academy of
Sciences of the United States of America 102(11), 4140-5.
Ricart, M.J., Malaise, J., Moreno, A., Crespo, M., Fernández-Cruz, L; Euro-SPK Study
Group. (2005) Cytomegalovirus: occurrence, severity, and effect on graft
survival in simultaneous pancreas-kidney transplantation. Nephrology, Dialysis,
Transplantation 20(Suppl 2:ii25-ii32), ii62.
Ripalti, A., Boccuni, M.C., Campanini, F., Landini, M.P. (1995) Cytomegalovirus-
mediated induction of antisense mRNA expression to UL44 inhibits virus
replication in an astrocytoma cell line: identification of an essential gene.
Journal of Virology 69(4), 2047–2057.
Romanowski, M.J., Shenk, T. (1997) Characterization of the human cytomegalovirus
irs1 and trs1 genes: a second immediate-early transcription unit within irs1
whose product antagonizes transcriptional activation. Journal of Virology 71(2),
1485-96.
Ross, S.A., Boppana, S.B. (2005) Congenital cytomegalovirus infection: outcome and
diagnosis. Seminars in Pediatirc Infectious Diseases 16(1), 44-9.
Ross, S.A., Fowler, K.B., Ashrith, G., Stagno, S., Britt, W.J., Pass, R.F., Boppana, S.B.
(2006) Hearing loss in children with congenital cytomegalovirus infection born
to mothers with preexisting immunity. The Journal of Pediatrics 148(3), 332-6.
Rowe, W.P., Hartley, J.W., Waterman, S., Turner, H.C., Huebner, R.J. (1956)
Cytopathogenic agent resembling human salivary gland virus recovered from
tissue cultures of human adenoids. Proceedings of the Society for Experimental
Biology and Medicine 92(2), 418-24.
Ruebner, B.H., Hirano, T., Slusser, R., Osborn, J., Medearis, D.N. Jr. (1966)
Cytomegalovirus infection. Viral ultrastructure with particular reference to the
relationship of lysosomes to cytoplasmic inclusions. The American Journal of
Pathology 48(6), 971-89.
Salzberger, B., Hartmann, P., Hanses, F., Uyanik, B., Cornely, O.A., Wöhrmann, A.,
Fätkenheuer, G. (2005) Incidence and prognosis of CMV disease in HIV-
infected patients before and after introduction of combination antiretroviral
therapy. Infection 33(5-6), 345-9.
Sarisky, R.T., Hayward, G.S. (1996) Evidence that the UL84 gene product of human
cytomegalovirus is essential for promoting oriLyt-dependent DNA replication
and formation of replication compartments in cotransfection assays. Journal of
Virology 70(11), 7398-413.

140
Schepers, A., Pich, D., Mankertz, J., Hammerschmidt, W. (1993) Cis-acting elements in
the lytic origin of DNA replication of Epstein-Barr virus. Journal of Virology
67(7), 4237–4245.
Scott, G.M., Weinberg, A., Rawlinson, W.D., Chou, S. (2007) Multidrug resistance
conferred by novel DNA polymerase mutations in human cytomegalovirus
isolates. Antimicrobial Agents and Chemotherapy 51(1), 89-94.
Seale, H., MacIntyre, C.R., Gidding, H.F., Backhouse, J.L., Dwyer, D.E., Gilbert, L.
(2006) National Serosurvey of Cytomegalovirus in Australia. Clinical and
Vaccine Immunology 13(11), 1181–1184.
Shen, C.Y., Chang, S.F., Lin, H.J., Ho, H.N., Yeh, T.S., Yang, S.L., Huang, E.S., Wu,
C.W. (1994) Cervical cytomegalovirus infection in prostitutes and in women
attending a sexually transmitted disease clinic. Journal of Medical Virology
43(4), 362-6.
Shen, C.Y., Chang, S.F., Yang, S.L., Wu, C.W., Yang, Y.S., Hong Nerng Ho, Huang,
E.S., Bantel-Schaal, U., Haefelin, D.N., Schieferstein, G. (1994a)
Cytomegalovirus is present in semen from a population of men seeking fertility
evaluation. Journal of Infectious Diseases 169(1), 222-223.
Shen, C.Y., Chang, W.W., Chang, S.F., Chao, M.F., Huang, E.S., Wu, C.W. (1992)
Seroepidemiology of cytomegalovirus infection among children between the
ages of 4 and 12 years in Taiwan. Journal of Medical Virology 37(1), 72-5.
Sherman, G., Gottlieb, J., Challberg, M.D. (1992) The UL8 subunit of the herpes
simplex virus helicase-primase complex is required for efficient primer
utilization. The Journal of Virology 66(8), 4884-92.
Sigrist, C.J., De Castro, E., Langendijk-Genevaux, P.S., Le Saux, V., Bairoch, A., Hulo,
N. (2005) ProRule: a new database containing functional and structural
information on PROSITE profiles. Bioinformatics 21(21), 4060-6.
Smith, C.A., Toogood, H.S., Baker, H.M., Daniel, R.M., Baker, E.N. (1999) Calcium-
mediated thermostability in the subtilisin superfamily: the crystal structure of
Bacillus Ak.1 protease at 1.8 A resolution. Journal of Molecular Biology 294(4),
1027-40.
Smith, J.A., and Pari, G.S. (1995a) Human cytomegalovirus UL102 gene. Journal of
Virology 69(3), 1734–1740.
Smith, J.A., Jairath, S., Cryte, J.J., and Pari, G.S. (1996) Characterization of the human
cytomegalovirus UL105 gene and identification of the putative helicase protein.
Virology 220, 251–255.
Smith, J.A., Pari, G.S. (1995) Expression of human cytomegalovirus UL36 and UL37
genes is required for viral DNA replication. Journal of Virology 69(3), 1925-31.
Smith, M.G. (1956) Propagation in tissue cultures of a cytopathogenic virus from
human salivary gland virus (SGV) disease. Proceedings of the Society for
Experimental Biology and Medicine 92(2), 424-30.
Soderberg-Naucler, C., Streblow, D.N., Fish, K.N., Allan-Yorke, J., Smith, P.P., Nelson,
J.A. (2001) Reactivation of latent human cytomegalovirus in CD14(+)
monocytes is differentiation dependent. Journal of Virology 75(16), 7543-54.
Spaete, R.R., Mocarski, E.S. (1987) Insertion and deletion mutagenesis of the human
cytomegalovirus genome. Proceedings of the National Academy of Sciences of
the United States of America 84(20), 7213-7.
Spector, D.J., Tevethia, M.J. (1994) Protein-protein interactions between human
cytomegalovirus IE2-580aa and pUL84 in lytically infected cells. Journal of
Virology 68(11), 7549-53.

141
Springer, K.L., Chou, S., Li, S., Giller, R.H., Quinones, R., Shira, J.E., Weinberg, A.
(2005) How evolution of mutations conferring drug resistance affects viral
dynamics and clinical outcomes of cytomegalovirus-infected hematopoietic cell
transplant recipients. Journal of Clinical Microbiology 43(1), 208-13.
Stagno, S., Pass, R.F., Cloud, G., Britt, W.J., Henderson, R.E., Walton, P.D., Veren,
D.A., Page, F., Alford, C.A. (1986) Primary cytomegalovirus infection in
pregnancy. Incidence, transmission to fetus, and clinical outcome. JAMA: The
Journal of the American Medical Association 256(14), 1904-8.
Stagno, S., Pass, R.F., Dworsky, M.E., Henderson, R.E., Moore, E.G., Walton, P.D.,
Alford, C.A. (1982) Congenital cytomegalovirus infection: The relative
importance of primary and recurrent maternal infection. The New England
Journal of Medicine 306(16), 945-9.
Staras, S.A., Dollard, S.C., Radford, K.W., Flanders, W.D., Pass, R.F., Cannon, M.J.
(2007) Seroprevalence of cytomegalovirus infection in the United States, 1988-
1994. Clinical Infectious Diseases 43(9), 1143-51.
Steininger, C. (2007) Clinical relevance of cytomegalovirus infection in patients with
disorders of the immune system. Clinical Microbiology and Infection 13(10),
953-63.
Stropes, M.P., Miller, W.E. (2008) Functional analysis of human cytomegalovirus
pUS28 mutants in infected cells. Journal of General Virology 89(1), 97-105.
Sullivan, V., Biron, K.K., Talarico, C., Stanat, S.C., Davis, M., Pozzi, L.M., Coen, D.M.
(1993) A point mutation in the human cytomegalovirus DNA polymerase gene
confers resistance to ganciclovir and phosphonylmethoxyalkyl derivatives.
Antimicrobial Agents and Chemotherapy 37(1), 19-25.
Sullivan, V., Talarico, C.L., Stanat, S.C., Davis, M., Coen, D.M., Biron, K.K. (1992) A
protein kinase homologue controls phosphorylation of ganciclovir in human
cytomegalovirus-infected cells. Nature 358(6382), 162-4.
Takeda, K., Haque, M., Nagoshi, E., Takemoto, M., Shimamoto, T., Yoneda, Y.,
Yamanishi, K. (2000) Characterization of human herpesvirus 7 U27 gene
product and identification of its nuclear localization signal. Virology 272, 394–
401.
Taylor-Wiedeman, J., Sissons, P., Sinclair, J. (1994) Induction of endogenous human
cytomegalovirus gene expression after differentiation of monocytes from healthy
carriers. Journal of Virology 68(3), 1597-604.
Taylor, T.J., Knipe, D.M. (2003) C-terminal region of herpes simplex virus ICP8
protein needed for intranuclear localization. Virology 309(2), 219-31.
Tenney, D.J., Hurlburt, W.W., Micheletti, P.A., Bifano, M., Hamatake, R.K. (1994) The
UL8 component of the herpes simplex virus helicase-primase complex
stimulates primer synthesis by a subassembly of the UL5 and UL52 components.
The Journal of Biological Chemistry 269(7), 5030-5.
Terhune, S., Torigoi, E., Moorman, N., Silva, M., Qian, Z., Shenk, T., Yu, D. (2007)
Human Cytomegalovirus UL38 Protein Blocks Apoptosis. Journal of Virology
In Print.
Thompson, J.D., H.D.G.G.T.J. (1994) CLUSTAL W: improving the sensitivity of
progressive multiple sequence weighting, position-specific gap penalties and
weight matrix choice. Nucleic Acid Research 22, 4673-4680.
Tjiam, K.H., van Heijst, B.Y., Polak-Vogelzang, A.A., Rothbarth, P.H., van Joost, T.,
Stolz, E., Michel, M.F. (1987) Sexually communicable micro-organisms in
human semen samples to be used for artificial insemination by donor.
Genitourinary Medicine 63(2), 116-8.

142
Urwijitaroon, Y., Teawpatanataworn, S., Kitjareontarm, A. (1993) Prevalence of
cytomegalovirus antibody in Thai-northeastern blood donors. The Southeast
Asian Journal of Tropical Medicine and Public Health 24(1), 180-2.
Vink, C., Beuken, E., Bruggeman, C.A. (2000) Complete DNA sequence of the rat
cytomegalovirus genome. Journal of Virology 74(16), 7656-65.
Wagstaff, A.J., Bryson, H.M. (1994) Foscarnet. A reappraisal of its antiviral activity,
pharmacokinetic properties and therapeutic use in immunocompromised patients
with viral infections. Drugs 48(2), 199-226.
Walker, J.E., Saraste, M., Runswick, M.J., and Gay, N.J. (1982) Distantly related
sequences in the alpha- and beta-subunits of ATP synthase, myosin, kinases and
other ATP-requiring enzymes and a common nucleotide binding fold. The
EMBO Journal 1(8), 945-51.
Wang, W., Patterson, C.E., Yang, S., Zhu, H. (2004) Coupling generation of
cytomegalovirus deletion mutants and amplification of viral BAC clones.
Journal of Virological Methods 121(2), 137-43.
Wang, Y.S., Hall, J.D. (1990) Characterization of a major DNA-binding domain in the
herpes simplex virus type 1 DNA-binding protein (ICP8). Journal of Virology
64(5), 2082-2089.
Wathen, M.W. (2002) Non-nucleoside inhibitors of herpesviruses. Reviews in Medical
Virology 12(3), 167-78.
Weber, O., Bender, W., Eckenberg, P., Goldmann, S., Haerter, M., Hallenberger, S.,
Henninger, K., Reefschläger, J., Trappe, J., Witt-Laido, A., Ruebsamen-
Waigmann, H. (2001) Inhibition of murine cytomegalovirus and human
cytomegalovirus by a novel non-nucleosidic compound in vivo. Antiviral
Research 49(3), 179-89.
Weiland, K.L., Oien, N.L., Homa, F., Wathen, M.W. (1994) Functional analysis of
human cytomegalovirus polymerase accessory protein. Virus Research 34(3),
191-206.
Weinberg, A., Jabs, D.A., Chou, S., Martin, B.K., Lurain, N.S., Forman, M.S.,
Crumpacker, C; Cytomegalovirus Retinitis and Viral Resistance Study Group;
Adult AIDS Clinical Trials Group Cytomegalovirus Laboratories. (2003)
Mutations conferring foscarnet resistance in a cohort of patients with acquired
immunodeficiency syndrome and cytomegalovirus retinitis. The Journal of
Infectious Diseases 187(5), 777-84.
Weissman, J.S., Kim, P.S. (1993) Efficient catalysis of disulphide bond rearrangements
by protein disulphide isomerase. Nature 365(6442), 185-8.
White, E.J., Boehmer, P.E. (1999) Photoaffinity labeling of the herpes simplex virus
type-1 single-strand DNA-binding protein (ICP8) with
oligodeoxyribonucleotides. Biochemical and Biophysical Research
Communications 264(2), 493-7.
Wolf, D.G., Smith, I.L., Lee, D.J., Freeman, W.R., Flores-Aguilar, M., Spector, S.A.
(1995) Mutations in human cytomegalovirus UL97 gene confer clinical
resistance to ganciclovir and can be detected directly in patient plasma. The
Journal of Clinical Investigation 95(1), 257-63.
Wong, A., Tan, K.H., Tee, C.S., Yeo, G.S. (2000) Seroprevalence of cytomegalovirus,
toxoplasma and parvovirus in pregnancy. Singapore Medical Journal 41(4), 151-
5.
Wong, S.W., Wahl, A.F., Yuan, P.M., Arai, N., Pearson, B.E., Arai, K., Korn, D.,
Hunkapiller, M.W., Wang, T.S. (1988) Human DNA polymerase alpha gene
expression is cell proliferation dependent and its primary structure is similar to

143
both prokaryotic and eukaryotic replicative DNA polymerases. The EMBO
Journal 7(1), 37-47.
Woycechowsky, K.J., and Raines, R.T. (2003) The CXC Motif: A Functional Mimic of
Protein Disulfide Isomerase. Biochemistry 42(18), 5387-94.
Xu, Y., Cei, S.A., Huete, A.R., Pari, G.S. (2004) Human cytomegalovirus UL84
insertion mutant defective for viral DNA synthesis and growth. Journal of
Virology 78(19), 10360-9.
Xu, Y., Cei, S.A., Rodriguez, Huete, A., Colletti, K.S., Pari, G.S. (2004a) Human
cytomegalovirus DNA replication requires transcriptional activation via an IE2-
and UL84-responsive bidirectional promoter element within oriLyt. Journal of
Virology 78(21), 11664-77.
Xu, Y., Colletti, K.S., Pari, G.S. (2002) Human cytomegalovirus UL84 localizes to the
cell nucleus via a nuclear localization signal and is a component of viral
replication compartments. Journal of Virology 76(17), 8931-8.
Yang, Y.S., Ho, H.N., Chen, H.F., Chen, S.U., Shen, C.Y., Chang, S.F., Huang, E.S.,
Wu, C.W. (1995) Cytomegalovirus infection and viral shedding in the genital
tract of infertile couples. Journal of Medical Virology 45(2), 179-82.
Ye, L.B., Huang, E.S. (1993) In vitro expression of the human cytomegalovirus DNA
polymerase gene: effects of sequence alterations on enzyme activity. Journal of
Virology 67(11), 6339-47.
Yu, D., Silva, M.C., Shenk, T. (2003) Functional map of human cytomegalovirus
AD169 defined by global mutational analysis. Proceedings of the National
Academy of Sciences of the United States of America 100(21), 12396–12401.
Yu, Y., Alwine, J.C. (2002) Human cytomegalovirus major immediate-early proteins
and simian virus 40 large T antigen can inhibit apoptosis through activation of
the phosphatidylinositide 3'-OH kinase pathway and the cellular kinase Akt.
Journal of Virology 76(8), 3731-8.
Yust, I., Fox, Z., Burke, M., Johnson, A., Turner, D., Mocroft, A., Katlama, C.,
Ledergerber, B., Reiss, P., Kirk, O; EuroSIDA. (2004) Retinal and extraocular
cytomegalovirus end-organ disease in HIV-infected patients in Europe: a
EuroSIDA study, 1994-2001. European Journal of Clinical Microbiology and
Infectious Diseases 23(7), 550-9.
Zacny, V.L., Gershburg, E., Davis, M.G., Biron, K.K., Pagano, J.S. (1999) Inhibition of
Epstein-Barr virus replication by a benzimidazole L-riboside: novel antiviral
mechanism of 5, 6-dichloro-2-(isopropylamino)-1-beta-L-ribofuranosyl-1H-
benzimidazole. Journal of Virological Methods 73(9), 7271-7.
Zhang, J., Chung, D.W., Tan, C.K., Downey, K.M., Davie, E.W., So, A.G. (1991)
Primary structure of the catalytic subunit of calf thymus DNA polymerase delta:
sequence similarities with other DNA polymerases. Biochemistry 30(51),
11742-50.
Zhu, H., Shen, Y., Shenk, T. (1995) Human cytomegalovirus IE1 and IE2 proteins
block apoptosis. Journal of Virology 69(12), 7960-70.
Zhu, L., Weller, S.K. (1992) The six conserved helicase motifs of the UL5 gene product,
a component of the herpes simplex virus type 1 helicase-primase, are essential
for its function. Journal of Virology 66(1), 469-79.
Zuccola, H.J., Filman, D.J., Coen, D.M., and Hogle J.M. (2000) The crystal structure of
an unusual processivity factor, herpes simplex virus UL42, bound to the C
terminus of its cognate polymerase. Molecular Cell 5, 267-278.




















